
Submitted:
18 May 2024
Posted:
21 May 2024
You are already at the latest version
Abstract
Phosphorus (P) is an important and irreplaceable macronutrient. It is central to energy and information storage and exchange in living cell. P is an element with “broken geochemical cycle” since it lacks abundant volatile compounds capable of closing the P cycle. P fertilizers are critical for global food security, but the reserves of minable P are scarce and non-evenly distributed between countries of the world. Accordingly, the risks of global crisis due to limited access to P reserves is expected to be graver than those entailed by competition for fossil hydrocarbons. Paradoxically, despite the scarcity and value of P reserves, its usage is extremely inefficient: current waste rate reaches 80% giving rise to a plethora of unwanted consequences such as eutrophication leading to harmful algal blooms. Microalgal biotechnology comprise a promising solution to tackle this challenge. The proposed review briefly presents the relevant aspects of microalgal P metabolism such as cell P reserve composition and turnover, and the regulation of P uptake kinetics for maximization of P uptake efficiency with the focus on novel knowledge. Multifaceted role of polyPhosphates, the largest cell depot for P, is discussed with emphasis on the P toxicity mediated by short-chain polyPhosphates. Opportunities and hurdles of P bioremoval via P uptake from waste streams with microalgal cultures, either suspended or immobilized, is discussed along with the role of bacterial components of microalgal-bacterial consortia in these processes. Possible avenues of P-rich microalgal biomass such as biofertilizer production or extraction of valuable polyPhosphates and other bioproducts are considered. The review concludes with a comprehensive assessment of the current potential of microalgal biotechnology for ensuring the sustainable usage of phosphorus.
Keywords:
phosphorus
; microalgae
; polyPhosphate(s)
; luxury uptake
; wastewater
; biofertilizer
1. Introduction: Peculiarities of Phosphorus as a Macronutrient
No living cell, including microalgal cell, can exist without phosphorus (P). This nutrient plays a central role in storage and transduction of energy and information therein and serves as an important structural component of the cell. The main organic P pools of the cell include nucleic acids, phospholipids, and diverse low molecular weight phosphometabolites. The main inorganic P pools are dissolved inorganic phosphate, orthophosphate (Pi), and its homopolymer polyPhosphate (PolyP, see section 1.1.4), the main reserve form of P in the cell (the roles of P and its main pools in the cells are briefly discussed in the sections 1.1.1 and 1.1.2). Pi is itself a key metabolite existing in a delicate equilibrium with a broad array of P-metabolites of central metabolism including Calvin cycle, turnover of sugars, adenylates, and nucleotides [1].
Unlike the major biogenic elements such as O, N, or C; P slowly enters the biosphere, mainly during weathering of the base rock releasing Pi into the pedosphere and hydrosphere. There, due to its high reactivity, a significant part of P gets bound into poorly soluble compounds [2]. Therefore, P is characterized by a low bioavailability despite its high abundance (P is 11th most abundant element in Earth crust [3], see also section 1.1). Some particulate Pi-containing material is deposited on ocean and lake bottoms, where it can be recycled at an extremely slow rate. The net result of this, is a very slow turnover of P which is even called “an element with a broken geochemical cycle” [4] and living organism most of the time experience a shortage of P and therefore became adapted to it in the process of evolution [5].
Recently, humankind exerted a profound impact on P cycling in the biosphere. In the pre-industrial epoch, the production of food depended mostly on and was limited by the availability of organic waste as a source of P. With intensification of crop production after the green revolution the demand of P fertilizer soared, and extensive mining of rock phosphates for fertilizer production created two major concerns [6,7,8]: dwindling of P resources and wasteful P usage. Recently estimated rock phosphate global consumption exceeds 160 million tons or ca 21 million tons of P [9]. These “concentrates” or, in other words, economically extractable source of P, may be exhausted in the nearest future, although what is ‘economically extractable’ will certainly change with increasing demand for P and technological development. This will lead to an unprecedented crisis in agriculture complicated by the extremely uneven distribution of the minable rock phosphate between different countries similarly to the distribution of oil and other fossil hydrocarbons [4,8]. Nevertheless, there is a crucial difference between fossil fuel and P: while the former can be replaced with alternative carbon and energy sources, P is irreplaceable. Although new deposits may be found, solving the broken P cycle is still of critical importance for global agriculture.
Furthermore, the usage of P fertilizers is woefully inefficient: in many cases less than 20% of P applied with the fertilizers is taken up by crop plants [10]. These losses along with those in the supply chain of agricultural products are leaching from agricultural soils to terrestrial and aquatic ecosystems. The abrupt increase of Pi availability promotes algal and cyanobacterial blooms [11] with subsequent depletion of O2 in the environment and suffocation of the hydrobionts [12]. Frequently the eutrophication gets exacerbated by proliferation of toxic cyanobacteria and dinoflagellates whose toxins increase the damage to the ecosystems [13].
While the society becomes increasingly aware of the non-renewable and finite nature of P resources (see section 1.1), the paramount challenge of establishing a sustainable anthropogenic phosphate cycle did not change in the last decade [6]. The need to act is urgent and to cope with P shortage several action priorities have been recently suggested including: recovery of phosphate from anthropogenic point sources, precision fertilizer placement technology, engineering of plants for minimizing their P requirements and maximizing phosphate uptake [7,14,15]. The evidence from the fields of microalgal physiology, ecology, and biotechnology strongly supports the idea that at least some of these priorities are attainable with the use of microalgal cultures.
Under ample P availability conditions, microalgal cells take up much more P than it is necessary for the next cell division judging from the Redfield ratio 106 : 16 : 1 (C:N:P) [16]. Previous estimation of global potential of even non-optimized P uptake capacity of cultivated microalgae [7] (calculated on a rather optimistic assumption of 40 g (DW) ∙ m–2 ∙ day–1 and a modest cell dry weight P percentage of 1.8%) suggests that a production area of approximately 26,600 km2 (a little more than, e.g., the area of Israel) would be required to sequester the 7 Mt of P that is annually lost worldwide with animal manure. In reality, the required area could even be less since microalgal P uptake and accumulation capacity can be boosted at least to 7% of Cell DW [14,15]. Therefore, the goal of increasing the sustainability of using P with help of microalgal biotechnologies seems to be, in principle, feasible. Moreover, the potential of microalgal biotechnology for P upcycling is vigorously discussed in the recent literature (the number of publications retrieved by Scopus query “microalgae phosphorus removal” increased more than three times over last 10 years, see also [17,18]). At the same time, there are limitations and concerns (which are often overlooked) that should be realized and circumvented before the practical implementation of the microalgae-based technologies for P biocapture will became widely available. Towards this end, we attempted to summarize a broad spectrum of opinions from somewhat pessimistic to strongly optimistic about the potential of microalgal biotechnology for achieving the sustainable usage of P, particularly in agricultural systems, and in frame of the development of innovative bio-economies.
1.1. Global Perspective on P: A Nutrient That is “Abundant but Scarce” and Mostly Wasted
Although P is abundant in the Earth crust (1200 mg kg–1, 11th most abundant element) [3], it is dispersed over wide areas. In the soil, its abundance is lower (900 mg kg–1 [29]). Moreover, it does not exist as free element due to its high reactivity, it is therefore frequently bound to chemical compounds making it scarcely available for uptake by phototrophs. The geochemical cycle of P is very different from those of other biogenic elements since there is no cycling through the atmosphere, so P was called “an element with broken geochemical cycle” [4]. In contrast to situation with other macronutrients such as nitrogen and carbon, most ecosystems depend entirely on the aqueous transfer of P [2]. Therefore, availability of P is the factor limiting productivity of primary producers such as phytoplankton in most aquatic ecosystems. In turns microalgae acquired diverse mechanisms to cope with P scarcity during their evolution (section 2.1).
Likewise, P often becomes the limiting nutrient for terrestrial plant productivity, especially in agroecosystems. Thus, from the time of the Green Revolution, crop size in agriculture (and hence food security) depends on intensive fertilizing. Consequently, P fertilizers which are made from rock phosphate is of utmost importance. However, the minable P resources are finite and non-renewable. The estimates of rock phosphate resources are highly controversial, most of them concur on the time span from several decades to several centuries. Moreover, economically extractable deposits of rock phosphate are extremely unevenly distributed between different countries: Morocco controls more than two thirds of the reserves estimated to be commercially exploitable, while China, Algeria and Syria possess their shares in the single-digit percentages [30]. Fears of running out of P reserves and thereby jeopardizing global food security are continuously resurface, so geopolitics come into play, and the main supply nations could drive up prices, and the consequences of this can be graver than those of competition for fossil hydrocarbons.
More than 90% of the P mined as rock phosphate is currently used in the food supply chain. Paradoxically, the processes in this supply chain are woefully inefficient: more than 80% of the extracted P ends up in waste and is lost in the environment, mainly in the hydrosphere [31]. A part of it gets bound into poorly soluble chemical compounds , hence it becomes poorly bioavailable and the remaining can cause its own set of problems such as widespread water quality issues mediated by excessive microalgae growth. Ironically, these microalgae could also provide a mean to recover and recycle P as they can store this precious nutrient.
1.2. P Acquisition and Storage in the Cell
1.2.1. Cell P quota and P uptake Capacity
The capacity of microalgal cell for taking up and accumulating P (cell P quota) is defined by the difference between the lowest and the highest P content [32]. The lowest P content (also called the minimal cell P quota) is typical of microalgal cells whose P reserves are depleted to the largest possible extent. That is, the cell division is slowed down or arrested but the cell sustained no irreversible damage so that its division resumes upon replenishment of P [5,33]. Accordingly, the largest cell P content or largest cell P quota is recorded when all the intracellular P pools (see below) are filled, and no further P uptake is possible in any form. This does not include the P adsorbed on the surface structures of the cell which might be significant; in certain microalgal species such as Micractinium simplicissimum, it can even surpass the cell P quota (see e.g., [34]). Cell P quota can be modulated by the mode of cultivation and the operating parameters [35,36,37,38]. Maximal values of P content of biomass (and per cell) were achieved for Nannochloropsis salina in continuous steady-state mode of cultivation under highest applied light intensity [39]. In practice, both cell P quota and cell P adsorption capacity are important determinants of the ability of the microalgal culture to capture P from waste streams.
The absolute P content of microalgal biomass under conditions of scarce availability but sufficient for the progression of cell cycle normally obeys the aforementioned Redfield ratio constituting, on an average, 1–2 % of the cell dry weight e.g., of natural phytoplankton [40]. Interestingly, the proportion of P in biomass of terrestrial plants is much lower due to a large carbon content [31], especially in mechanical and protective tissues and, likely, due to the inability to accumulate large quantities of polyPs. The microalgae cultivated under ample conditions e.g., in wastewater, frequently reach 3–4% P in their cell dry weight (CDW) [41,42]. Transient P shortage triggers a complex of mechanisms dramatically increasing microalgal cell capacity of P acquisition, so it exceeds “normal” metabolic demand. This phenomenon called “luxury uptake” (for more detail, see 1.2.3) pushes microalgal P content to 7 % of CDW and higher [14,15,33,43,44,45]. For example, P-hyperaccumulating microalgae were isolated from the Revolving Algal Biofilm (RAB) system used for P recovery and reported to accumulate up to 14% PolyP of CDW [46] which seem to be at the top of realistic P content values. The discrepancy between studies in estimating total P content and, particularly, PolyP-P content can be related to the different methodologies used for P metabolites quantification, as other phosphorous molecules (nucleic acids) may interfere with the accurate determination of polyPs [47].
1.1.2. Phosphorus Pools in the Cell
The typical intracellular Pi concentrations in the cell are in the range of 5–10 µM [48,49]. Pi is present almost in all cell compartments where it gets involved in a plethora of metabolic and regulatory reactions. Although the intracellular Pi concentrations can be estimated as modest, the turnover of intracellular Pi is significant [50]. Intracellular Pi concentration is maintained, independent of external Pi concentration, by the system of Pi homeostasis. This system includes the mechanisms of Pi uptake and the main cell P storage depot in form of polyP (discussed in sections 1.1.3 and 1.1.4) and several pools of phosphometabolites as further described below.
In the cell, P exists within different groups of molecules and/or cell compartments designated as P pools (Figure 1). One of the largest P pools is comprised by nucleic acids serving for storage and transduction of genetic information and developmental signals in living systems. Ribosomal RNA comprises the largest and most flexible P pool whereas DNA-P pool is quite stable even during P shortage periods. There are also exceptions when chloroplast rRNA increase, and chloroplast DNA decrease, during P shortage [51]. This P pool can play an important role in the rapidly growing cultures used for sequestration of P from waste streams.
A relatively small but functionally important P pool is comprised by phosphoproteins involved in intracellular signaling and regulation i.e., when the proteins are phosphorylated and dephosphorylated by assorted protein kinases.
Phosphate is a component of a polar group of phospholipids, an important class of membrane lipids. Phospholipids, such as phosphatidylglycerol, occur in plastidial membranes and play an important role in photosynthesis. In higher plants, it is estimated that phosphate is present in less than half of the envelope membrane lipids within chloroplasts, and less than 15% of thylakoid membrane lipids (see [52] and references therein). These estimations can differ in microalgae due to their dynamic nature, complex evolution of membrane systems and exposure to rapidly changing environment. Other common phospholipids can be present in both the plastidial and extraplastidial lipids, as a component of cellular membranes (ER, mitochondria, plasma membrane). Although the content of phospholipids is relatively low compared to other glycerolipids, they are considered as an internal P resource important for P sparing under conditions of P scarcity. Additionally, microalgal cells use lipid remodeling strategies to maintain cellular homeostasis under conditions of variable P and environment. Under conditions of P shortage, P-containing lipids can be replaced with the non-phosphorous structural lipids, galactolipids, S-containing plastidial sulfoquinovosyldiacylglycerol or N-containing betaine lipids [53]; implementation of this replacement is genotypically and phenotypically variable [54].
Another large P pool deeply involved in energy storage and transduction are nucleoside triphosphates comprising one of the energy “currencies” of the cell, with ATP as a ubiquitous energy storage form. Overall, a considerable part of intracellular P is also incorporated into diverse P-metabolites such as phosphorylated sugars. The main P storage of the microalgal cell is represented by vacuolar polyP which is a dynamic P depot (discussed in 1.2.4) whereas in P-sufficient plants, 85 to 95% of P reserves are found as vacuolar Pi [55].
1.2.3. Phosphorus Uptake
While different genes are involved in P acquisition and metabolism within different taxa of phototrophs, the general responses to P shortage are similar with the up-regulation of genes responsible for P acquisition, transport, and storage. As mentioned above, Pi homeostasis in the cytoplasm is maintained in the lower mM range by Pi transport system comprised by intricately regulated mechanisms translocating Pi across the tonoplast (dis)engaged in response to changes in availability of external P and other environmental conditions. Normally, uptake of Pi takes place against a steep gradient: its concentration in the cell is higher than in the cell surroundings (Table 1), so Pi uptake is carried out mostly by active mechanisms. At the same time, the uptake capacity of this system is limited (for most microalgal species, Ki for Pi uptake is below 4 µM [5]). Loading Pi into vacuole across the tonoplast also requires ATP [56] and contributes to Pi homeostasis of the cell, especially during luxury P uptake. It is believed that photophosphorylation is the main source of ATP energy for the active Pi uptake in phototrophs, although under severe Pi depletion cyanobacteria can leverage their internal energy sources for Pi acquisition [57]. It was also hypothesized that in marine cyanobacteria, Pi uptake is likely limited by the surface of their outer membrane and not by the cell energy reserves [57].
Two main Pi uptake systems of microalgae are represented by two subsystems (Figure 2). One is the high affinity transporters operating when Pi is scarce (normally in natural environments, see Table 1 and Tables S1-S3 for the genes/enzymes involved). These are among the oldest and highly conserved proteins, more detail on their structure, operation, and evolution can be found elsewhere [55]. Mechanistically, the Pi translocation through the plasma membrane is a co-transport process driven by protons generated by a cytoplasmic membrane H+-ATPase. The high-affinity transporters are easily saturated by Pi; in other words, their Michaelis–Menten constant, Km (the concentration of substrate that allows the reaction to proceed at one-half its maximum rate) is low varying from 0.1–0.3 μM (as was determined for C. reinhardtii). As a result of the deployment of the high-affinity Pi transporters, Vmax of Pi transport increases 10–20 times [58]. Important characteristics of the high-affinity P transporters are their rapid and specific induction upon the onset of Pi shortage and repression after re-supplementation of Pi(within ca. 24 h [58]).
Another subsystem is the low affinity Pi transporters which function constitutively (i.e., largely independently of the external Pi availability). While the low-affinity Pi transporters cannot acquire Pi when it is present in low concentrations (e.g., PTC1, PTA1), they are not so readily saturated by Pi when it is ample in the cell surroundings (i.e., they have a higher Km about 10 μM). The low-affinity Pi transporters are therefore responsible for the translocation of the bulk of Pi (ca. 80%) taken up by the cell under ample P conditions, whereas in P-starved cells nearly all Pi uptake takes place via the high-affinity system [59].
An important role of the periplasm in Pi acquisition by marine cyanobacteria has been revealed recently [57]: they acquire Pi by maintaining a periplasmic concentration below environmental levels; the Pi accumulated in the extracellular buffer can be removed hypo-osmotically by ATP-powered transport. Interestingly, the proton motive force (PMF) is not required for the Pi retention in the periplasm by loading of Pi into the periplasm across the outer membrane is PMF-dependent and can be augmented by the energy coming from photosynthesis. As a result, marine cyanobacteria can uptake Pi even at its very low external concentrations, making these organisms good candidates for polishing of wastewater with low Pi levels.
The Pi transporter proteins of terrestrial plants or fungi (yeast) are relatively well known whereas these from microalgae are much less studied; current knowledge of the Pi transporter proteins originates mostly from a handful of model organisms such as Chlamydomonas reinhardtii or Arabidopsis thaliana. Thus, 25 putative genes—homologs of the higher plant PHT family encoding Pi transporters in C. reinhardtii divided into four subfamilies of CrPTA (H/Pi cotransporter), CrPTB (Na/Pi symporter), CrPHT3, and CrPHT4 whose expression is putatively regulated by CrPSR1 [60].
Overall, higher plants and fungi (yeast) harbor genes involved in Pi uptake and its regulation—homologs of the corresponding genes are present in microalgae suggesting similarity of their regulation mechanisms.
1.1.4. PolyPhosphate Turnover and Its Regulation
The maintenance of Pi homeostasis in the cytosolic and other cell compartments is critical for maintenance of normal cell metabolism which can be easily disturbed by excess of Pi. After a sudden increase of the external Pi concentration (e.g., because of P fertilizer leaching), it may exceed a thermodynamic threshold, so the energy available to the cell will become sufficient for massive Pi uptake. The Pi taken up in excess of the current metabolic demand is stored in the form of polyP, the relatively metabolically inert and osmotically safe storage form of P [61].
Several organic and inorganic molecules can be classified as polyP(s) with inorganic linear polyP being the main storage molecule. Inorganic polyP is homopolymer of orthophosphoric acid residues varying in chain length. It has been discovered in all kingdoms of life including oxygenic phototrophs. In addition to its function of main P depot of the microalgal cell, it is claimed to serve as a primordial source of energy (stored in phosphoanhydride bonds linking the Pi residues) that may have been used by biological systems prior to the evolutionary advent of ATP, hence it is often referred to as “molecular fossil” [62,63]. Synthesis and accumulation of polyP is related with numerous cellular functions, apart from P storage: it is involved in maintaining adenylate and metal cation homeostasis, counter-ion for cation sequestration, protein activity modulation, and stress acclimation [63,64]. At the same time, in certain conditions uncontrolled synthesis of short-chain polyP can likely mediate P toxicity (section 2.3).
Numerous studies on PolyP turnover in the cells of microalgae suggest that polyP, as other intracellular reserves, are accumulated (i) when bioavailable P is ample in the cell surroundings and (ii) when the metabolic demand of P is lower than its influx into the cell. Accordingly, polyP accumulation typically occurs in the microalgal cells at early and advances stationary phase when cell division slows down whereas in the cells of rapidly dividing exponential cultures polyP are scarce [15,45].
The biosynthesis of polyP is energy intensive; it consumes energy-rich substrates such as ATP or inositol phosphates. While the bulk of energy demand for biosynthesis of polyP in photoautotrophic cell is satisfied by photosynthesis, the energy for polyP assembly can be partly supplied by respiration or, under anaerobic conditions, by fermentation. Although the latter source is much less efficient in driving polyP biosynthesis [45].
In microlagal cells, polyP reserves are mainly stored in specialized vacuoles called acidocalcisomes [65,66]. According to the current understanding, the bulk of polyP in microalgal cell is synthesized by a complex molecular machinery comprised by several subunits—proteins from the VTC (vacuolar transport chaperone) family [67]. The structure and function of the VTC complex are most studied in baker’s yeast (Saccharomyces cereviseae) where it is a sophisticated protein complex assembled from the polyP polymerase VTC4 and location-specific combinations of the accessory VTC1, VTC2, VTC3, and VTC5 subunits [68,69]. In yeast, VTC was found to be activated by binding of inositol pyrophosphate with participation of (presumably) Pi-sensing SPX domain [67], a domain discovered in many other genes involved in Pimetabolism. Genomes of many microalgal species harbor genes encoding VTC complex subunits and recent in silico evidence showed that VTC4 proteins are structurally highly conserved [70] suggesting that the VTC complex is broadly distributed in microalgae and commonly involved in polyP synthesis in their cells [14,15]. The structure and mechanism of VTC operation in microalgae remains underexplored [42,70,71].
In cyanobacteria, polyP are synthesized by polyPhosphate kinase 1 (PPK1 which is evolutionary unrelated to VTC of eukaryotes [63]) degraded by polyPhosphate kinase 2 (PPK2), an enzyme with reversible activity, and the exopolyPhosphatase (PPX) cleaving the PolyP units. While homologs of PPK2 and cytosolic PPX as well as other enzymes with documented PolyP hydrolase activity were not found in in the proteome of C. reinhardtii [70,72], a diadenosine and diphosphoinositol polyPhosphate phosphohydrolase (DIPP) enzyme was suggested to be involved in PolyP degradation in C. reinhardtii [72]. Notably, variation of ppk/ppx genes expression might be a very flexible mechanism of acclimation of cyanobacteria to diurnal variation of Pi availability [73], so this mechanism can be of certain importance for biosequestration of Pi e.g., from waste streams with varying Pi abundance.
Regulation of the polyP biosynthesis in microalgae experiencing sudden fluctuations of Pi availability appears to be very dynamic. Thus, the cells of Synechocystis can be “primed” for luxury uptake of Pi and storage even by a short period of P deficiency (PolyP accumulation took place in first 1–3 min after the replenishment of Pi). In eukaryotic microalgae these processes generally take more time but follow the same pattern (see e.g., [15]). The accumulation of polyP can be triggered by other limitations (e.g., by sulfur deficiency [74]) slowing down cell division rate and the corresponding Pi expenditure. The polyP content of cyanobacterial biomass can also be enhanced by slowing down its degradation under ample P conditions by knocking out the phoU gene, a negative regulator of Pi-responsive genes in bacteria sensitive to the external Pi level [75]. This approach can be beneficial from the standpoint of technology since it would allow to retain the high polyP content in the grown biomass which normally tends to decline upon resumption of cell division after the transient PolyP accumulation following Pi refeeding of the culture.
Importantly, in eukaryotic algae, shortage of bioavailable P readily up-regulates the synthesis of the components of VTC compleх rendering the cell capable of rapid polyP accumulation whenever Pi becomes available. Proteomics studies revealed that the turnover time of the proteins involved in polyP biosynthesis is on the scale of hours since in C. reinhardtii, PolyP biosynthesis continues when the genes encoding VTC subunits 1 and 4 (Cre12.g510250 and Cre09.g402775) are already downregulated [71] suggesting the involvement of post-translational regulation in the control of polyP biosynthesis by the VTC complex, at least in C. reinhardtii. However, the similarity of response of many species from the transition to P depletion to P repletion suggest similar regulation in eukaryotic microalgae [71].
It should be also noted that many proteins involved in polyP turnover harbor SPX domains thought to be responsible for sensing of Pi levels in the cell. Thus, PolyP synthesis is thought to be stimulated by binding inositol pyrophosphate to the SPX domain of the VTC4 subunit as was revealed in VTC4 of C. reinhardtii [63,71,72] as well as Chlorella vulgaris, Desmodesmus armatus and Gonium pectorale [70].
The evidence on the effects of polyP on resource allocation and culture productivity are controversial. On one hand, there is a consensus that polyP biosynthesis is energy intensive. Indeed, knockout of ppk gene encoding polyP-kinase increased the productivity of Synechocystis sp. under favorable conditions [76]. On the other hand, in certain eukaryotic microalgae, growth was not negatively affected by P assimilation and polyP synthesis [70].
Overall, cells and cell populations of cyanobacteria and microalgae are heterogeneous in their ability to accumulate and metabolize polyP to adapt to fluctuating Pi availability [77]. They may adopt different strategies encompassing either slow growth with a high amount of stored polyP or fast growth and cell division at the expense of the stored polyP reserve [77].
2. Between Scylla and Charybdis: P-Starvation and P-Toxicity
In nature, microalgae experience mostly oligotrophic conditions. It means that most of the time microalgae are facing scarce and fluctuating availability of key mineral nutrients including phosphorus. This is the case for the habitats with very low P availability such as the oligotrophic central oceanic gyres [78], and very old soils found e.g., in Western Australia and South Africa [2]. As a result, microalgae are naturally equipped to cope with P scarcity by multiple mechanisms they developed during their evolution. Evolutionary adaptation to P shortages has left a deep marks on the genomic landscape of oxygenic phototrophs manifested e.g., by diverse regulation mechanisms affecting a wide range of genes triggered by a decline in P availability [13,79]. Briefly, these mechanisms enable the microalgae to get as much of the bioavailable P as possible and to do it as quickly as possible. The intracellular P is then neutralized during storage and can safely be stored for future P shortage.
By contrast, the mechanisms allowing to cope with extremely high nutrient spikes that may be harmful (see section 2.3) are apparently lacking in microalgae [14,33]. Abrupt increase of the nutrient availability either due to the natural reasons (upwelling, runoff after storms, etc.) or anthropogenic (wastewater discharge, fertilizer run-off from the fields, etc.) reasons leads to harmful algal blooms (HABs), poisoning and eutrophication.
Situations where fluctuating availability of P occur can be problematic in microalgal biotechnology. Indeed, mineral nutrient composition of many waste stream types is not balanced in terms of N:P ratio leading to incomplete removal of either nitrogen or P (section 3.2.2), other challenges may arise during treatment of wastewater with very high content of P e.g., those from phosphate mines and/or rock phosphate processing plants.
2.1. Phosphorus Starvation
The phenomenon of P starvation in microalgae is relatively well studied. This is because a significant research effort was directed to revealing the dynamics of phytoplankton in the aquatic ecosystems which is believed to be modulated, to a large extent, by limited P availability [13,55]. Most of the studied microalgal species are capable to accumulate ample reserves of P (section 1.3) sufficient to support the cell division for several generations (up to 20 [80]) in the absence of external bioavailable P. Besides as further described below, the cellular plasticity to P shortage allow the cell to survive for many days. Therefore, deprivation of P is considered to be a relatively mild stress in comparison with e.g., nitrogen deprivation [81]. Deprivation of P leads to activation of several acclimatory mechanisms which can be divided into two types: (i) the mechanisms increasing the efficiency of P acquisition by the cell and (ii) the mechanisms of mobilization of intracellular P reserves and P sparing mechanisms such as membrane lipid remodeling.
The first is the changeover of the Pi transporters in the cell membrane: the high-affinity Pi transporters genes which (section 1.2.3) are up-regulated, lead to an increase in high-affinity Pi transporters, replacing the low-affinity transporters. As a result, the cell increases it capacity to fetch Pi at its low external concentrations, though at a cost of addition expenditure of energy. Then, the expression and secretion of the extracellular enzymes such purple phosphatase(s) capable of liberation of Pi from DOP is increased (section 2.2.) so the capability of the cell to acquire P from its surroundings is enhanced.
The second is salvaging P from cell components such as expendable types of nucleic acids including ribosomal RNA and other surplus RNA molecules. The total rRNA declines during P shortage; it can drop to the level of the P content within DNA as was demonstrated for the marine picoplanktonic cyanobacterium Prochlorococcus marina [82]. The pools of Pi in different cell compartments as well as P-metabolites such sugar phosphates are also being gradually depleted.
Another critical mechanism enabling microalgal cell metabolic plasticity in response to limiting P is membrane lipid remodeling. In higher plants, the well-studied response to P-limitation constitutes the replacement of phospholipids with non-phosphorous galactolipids, namely the extraplastidial phospholipids with a bilayer forming galactolipid DGDG, and in the plastid, the replacement of the acidic phosphatidylglycerol with SQDG to sustain photosynthetic activity [83]. This strategy is also used by microalgae and an increased ratio of SQDG to PG was reported as a biomarker of P-limited oceanic waters [84]. The replacement of degrading phospholipids with certain galactolipids, betaine lipids and sulfolipids was also shown in the microalga C. reinhardtii and Nannochlorpsis oceanica [53,71].
In P-replete cells, non-phosphorous glycoglycerolipids include the predominant lipid classes in a photosynthetic cell, such as galactolipids, monogalactosyldiacylglycerol (MGDG), digalactosyldiacylglycerol (DGDG), and sulfoquinovosyldiacylglycerol (SQDG). Phosphorous-containing membrane lipids include phosphatidylglycerol (mainly present in the plastids) and several classes of phospholipids, involved in the key reactions of membrane and storage lipid biosynthesis. Importantly, many eukaryotic microalgae contain an additional class of membrane lipids – the non-phosphorous betaine lipids. This group of membrane lipids was deemed to be lost in higher plants during evolution along with the establishment of sessile lifestyle. Betaine lipids are present in many groups of microalgae and represented by three main types. Diacylglyceryl-N,N,N-trimethylhomoserine (DGTS), abundant in green microalgae, has structural similarity to a phospholipid phosphatidylcholine, PC. Its content generally increases along with galactolipid DGDG under conditions of P scarcity, while the content of phospholipids decreases. Recent biophysical studies suggested that bilayers formed by DGTS has some beneficial features in terms of thickness and water repulsion [85]. The capacity to swiftly remodel membrane lipids in response to P availably may have important consequences for P luxury uptake and P retaining in the cells. This direction warrants further studies. To note, P-starvation responses are regulated in the cell at different levels, including the transcriptome level, and regulation of many transcriptional factors and lipid metabolism genes. Overall, survival and growth of microalgae in P-deprivation or limitation needed for obtaining low cellular P quota are tightly related to lipid metabolism.
Lipid remodeling has also been shown to be a key component of cellular autophagy during P shortage. Autophagic degradation of cellular components supports cell homeostasis under nutrient starvation. Autophagosome formation relies on membrane modeling and re-modeling events, from the nucleation of the phagophore to its expansion and closure [86]. Autophagy may play a role in response to P deprivation, which is associated with a decrease in energy-rich molecules such as ATP, and a range of phosphorylated metabolites [87,88]. Phospholipids are required for the construction of autophagosomes and the operation of autophagy flux at several levels. For instance, for lipidation of the hallmark autophagy protein ATG8. The lipids associated with the autophagosome has started to be revealed in plants. While a new player in plant autophagosome formation appeared to be a negatively charged phosphatidylinositol-4-phosphate (PtdIns4P) [86], the lipid(s) involved in microalgal ATG8 lipidation is currently unknown. Because autophageous vacuoles decline with the duration of P starvation, we hypothesize that prolonged P deficiency depletes phospholipids required for the autophagy machinery and thus cells are unable to completely degrade polyP reserves.
Critically, the responses to P shortage are complex and cell have been shown to “prioritize” certain strategies for P homeostasis. For instance, P. shikokuense prioritizes the preservation of RNA and polyP for crucial metabolic processes while sparing and recycling phospholipids with non-phospholipids. Concurrently, autophagy is triggered in P. shikokuense under phosphorus deficiency, thereby decreasing energy exhaustion and potentially conserving and releasing phosphorus resources for more vital metabolic pathways.
However, the microalgal cells can retain some polyP reserves even after long P starvation (30+ days), the reasons for this remain enigmatic [89]. A plausible hypothesis explaining the loss of access to polyP reserves is failure of autophagic mechanisms and due to e.g., lack of energy resulting from overall metabolic quiescence under stress. On the other hand, polyPs are molecules with many metabolic functions (as mentioned earlier in the review), so it might be expectable that not all the polyP is consumed. This evidence that the regulation/sensing of P levels (as well as difference between species) needs to be elucidated.
Eventually, when all P resources, both internal and external, are exhausted, the cell division stops, and the typical rearrangements of nutrient-starvation stress in microalgae take place. Briefly, these are reduction of photosynthetic apparatus (decline in photosynthetic pigments such as chlorophylls, phycobilins, and primary carotenoids and dismantling of the thylakoid membranes) to avoid excessive light capture and hence the photooxidative damage when light and dark reactions of photosynthesis are off-balance [90]. Another hallmark of changes in the cell induced by nutrient starvation including P starvation is the accumulation of carbon reserves, mostly in form of starch and neutral lipids. Accordingly, the nutrient limitation/deprivation is frequently used to control the biochemical composition of industrially cultivated microalgae. Although, as it was noted above, the P starvation stress is milder than e.g., nitrogen starvation [91] and hence is less manageable.
As a net result of acclimation to P shortage, microalgal cells gain the capability for rapid uptake of Pi in large qualities. On one hand, this capability is thought to be advantageous: the species who captures P from the environment faster than other can thereby starve its competitors [92]. On the other hand, there is a danger of Pi intoxication mediated by short-chain polyPhosphates (see section 2.3.).
2.2. Mobilization of External DOP
Like higher plants and heterotrophic microorganisms, most microalgae [89,93,94,95] possess the ability to express external phosphatases under P shortage conditions facilitating the mobilization of P from the dissolved organic molecules in the cell surroundings (commonly termed dissolved organic phosphorus, DOP). In the model microalga C. reinhardtii, several extracellular and cell-wall associated phosphatases were discovered, including those constitutionally expressed and the alkaline and neutral phosphatases induced by limited P conditions, PHOX [79].
Omics studies revealed diverse phosphatases many of which may be responsive to environmental P levels in many other microalgal species such as coccolithophore Emiliania huxleyi; dinoflagellates Karenia brevis, Amphidinium carterae, and Alexandrium catenella (reviewed in [13]). At the same time, reservations should be made regarding relevance of these findings since it is relatively easy to identify a putative phosphatase gene, and even show that transcripts from the gene respond to the environmental P status, but it is more difficult to reveal the cellular location, kinetic features and, ultimately, the actual function of a putative phosphatase in the P metabolism of the certain microalga [96].
Clearly, the ability to mobilize external DOP is very important for treatment of organic-rich waste streams like those from diverse food industries. So, the strains with a large repertoire of efficient external phosphatase should be prospected for this application. The ability of microalgae to grow and store P from organic sources during biotechnological processes has not been investigated. Considering that they possess the machinery for it, this should be investigated in the future as it was done in yeast [97].
2.3. Phosphate Toxicity and Resilience to Elevated Pi Concentrarions
Literature survey indicates that the studies of microalgae under conditions where P is in large excess of the cell demand are much scarcer than studies in P starvation. Accordingly, the phenomenon of Pi toxicity remains largely underexplored though it can decline the robustness of microalgal cultures in P-eutrophicated systems. Therefore, understanding its mechanisms is important for the development of microalgal treatment of waste streams with high P load e.g., those from phosphate mining sites or fertilizer production plants. It is important to realize that deteriorative effects of excess Pi on microalgae are beyond the mere osmotic stress. Recently, reports appeared containing the phenomenological description of inhibitory or toxic effects of high external Pi e.g., in Chlorella vulgaris [98]. In these reports, the Pi toxicity was putatively associated with the formation of ample fine-grained polyP inclusions in the microalgal cells incubated at high external Pi.
Current reports on P metabolism in microalgae do not suggest the existence of specific pathways for detoxication of Pi when it presents in high concentrations. Extracellular structures like cell wall with high Pi adsorption capacity can contribute to the elevated Pi resilience. Another determinant of elevated Pi resilience is the ability to throttle its influx into the cell by down-regulation or gating of Pi transporters.
To prevent the disturbance of the metabolism by Piwhich has been taken up already, it should be converted into “safe”, less osmotically and metabolically active form of polyP and isolated in a storage cell compartment such as the vacuole. However, biosynthesis of polyP requires a large investment of energy, mainly in form of ATP which may not be readily available, especially under stressful conditions limiting the metabolic resources of the cell.
In view of the phenotypical hallmarks of high Pi toxicity, it was hypothesized that: At high Pi influx, synthesis of many polyP chains can be initiated simultaneously, but due to limited availability of energy it never completes resulting likely in the formation of abundant short-chain polyP. As short-chain polyP can interfere with protein folding and possibly other processes in the cell, their accumulation can result in slowdown of the cell division and other signs of inhibition observed under large excess of Pi [99,100].
This hypothesis was corroborated by the appearance of diffuse signal attributable to polyP on the EDX spectra taken from the cytoplasm of the cells exposed to a high Pi level in the absence of visible PolyP formation [98]. Moreover, the failure of high Pi tolerance in microalgae was observed upon abrupt Pi re-feeding of the microalgal cultures pre-starved of P and hence expected to be metabolically quiescent; notably, this was the case even upon addition of Pi in concentrations well below the toxic level [98]. An abrupt transition from P shortage to ample P conditions leads to a dramatic increase in Pi influx into the cell (since 10-fold increase in the Vmax of the Pi transport to the cells). Interestingly, the symptoms of P toxicity were also developed in higher plants upon an abrupt transition from P-depleted to P-replete conditions [101]. This needs to be taken into consideration for the development of bio-processes for P-upcycling.
3. Biotechnological Implications
3.1. Microalgae: The Curse of Eutrophication and the Boon of Biosequestration
As noted above, in nature, Pi slowly enters aquatic ecosystems, remains in the water column for a long time (from a few to thousands of years, and ends up sedimented on the ocean and lake bottoms (see [2,78] and references therein). There, it is recycled at an extremely slow rate subject to the action of many environmental factors including pH, redox potential, and salinity [78]. Anthropogenic intervention dramatically changed the distribution of bioavailable Pi in the environment. Thus, according to recent estimations, human activity mobilizes as much phosphate each year from rock phosphate deposits and other sources as is mobilized by ‘natural’ processes from base rocks in soil genesis [102]. One of the reasons for this is the massive application of P fertilizers. Much of the Pi applied with the fertilizers to the soil becomes unavailable to plants because it either gets immobilized (changes chemically) even though it stays in the soil or is washed away from the root zone [2]. Unfortunately, there are no known technologies for preventing such diffuse P losses in the environment. Although microalgae cause deleterious HABs under eutrophic conditions, they are increasingly claimed as an efficient vehicle for recovering P from point sources such as sewage and waste from animal farming. Microalgae are believed to be capable of closing the P loop by converting the P-rich microalgal biomass into biofertilizer, supplements to animal feed, and other valuable bioproducts. Suitability of microalgae for this role stems from their adaptations to fluctuating P availability in the environment acquired during their long evolution under such conditions (see below).
3.2. Microalgae-Mediated Biocapture of P
The findings reviewed above suggest that microalgae per se are very capable organisms regarding the uptake of different forms of P and using microalgal biomass for biocapture of P from waste streams is arguably promising. Although there are established practices of P bioremoval from waste streams with heterotrophic bacteria such as Enhanced Biological Phosphorus Removal (EBPR) to manage eutrophication risks [103], these are still technologically complex and expensive and hence unaffordable by small communities and companies. At the same time, the use of microalgae for biocapture of P is advantageous because of (i) more efficient wastewater treatment due to photogenerated oxygen [104]; (ii) generation of biomass convertible to valuable products such as biofertilizer (section 3.3); (iii) Simplicity with low capital and operational costs (low energy requirements), consequently having a lower environmental footprint that other alternatives [17].
Still, practical implementation of this approach can be problematic without a deep understanding of the relationships between culture operational parameters (cell density, growth phase), cultivation conditions (light, carbon and other nutrient supply, temperature, pH, mixing, presence of toxicants etc.) and the kinetics of P uptake translating into the amount of P that can be sustainably removed from the medium per unit of time. Of special importance, e.g., for wastewater polishing, is the completeness of P removal. Finally, the opportunities of the utilization of P-rich biomass obtained during P biosequestration are of crucial importance.
From the standpoint of technology, it is important to realize how large is the potential of microalgal culture for P removal under specific conditions and constraints. There are many reports in the literature presenting specific figures on P uptake under diverse experimental setups and a broad range of wastewater compositions (for a recent summary, see Table 1 in [17]), however it is more difficult to find a summary relating the culture conditions and the efficiency of P bioremoval, above all under full-scale conditions. In view of this, we attempt to generalize on the reported data to infer general prerequisites for efficient bioremoval of P from waste streams using microalgae.
3.2.1. Cultivation Conditions and P Nutrition History of the Culture
The natural acclimations of microalgae to fluctuations of Pi in the environment including the dramatic increase in P uptake capacity and overplus response (section 2) can be exploited to achieve the highest Pi uptake rates and P contents of the resulting biomass. Moreover, nearly complete removal of P from the medium can be achieved induced by P pre-starving of the culture [38].
Knowledge of the kinetics of Pi uptake and conversion into polyP is important to obtain microalgal biomass enriched with PolyP which represents a valuable biofertilizer. Thus, the cyanobacterium Synechocystis sp. PCC6803 accumulated polyP after 3 min. of the exposure to ample Pi conditions whereas 1 h later polyP content started to decline [77]. Similarly, the transient increase of polyP in P-starving Chlorella vulgaris IPPAS C-1 cells was observed 4 h after a Pi spike [15]. This capability of luxury Pi uptake renders microalgae particularly suitable for dealing with the spikes of Pi content in waste streams.
Importantly, the polyP accumulation depends on the energy coming (in form of ATP) from photosynthesis and it is sensitive to extreme temperatures [45,77]. Therefore, the adequate supply of light energy to the phototrophic cultures is important to ensure efficient polyP accumulation. However, these requirements may be difficult to fulfill as the wastewaters are turbid. Since atmospheric CO2 levels are limiting for microalgal growth, enrichment of microalgal cultures with CO2 augments their growth thereby boosting their P uptake capacity [105] although this requirement may be less strict in organic-rich wastewater and/or microalgae capable of mixotrophic growth. A decline of pH by injection of CO2 also increases the Pi bioavailability for microalgae though care should be taken to keep the pH within the range suitable for the microalgal culture.
The P-sufficient vigorously dividing cultures continue to consume P at a steady rate, and the sustained bioremoval of Pi is currently the mainstream approach in environmental applications [17,106,107]. Although the absolute amount of P removed by such cultures can be significant, complete P removal from the medium of dense P-sufficient cultures is difficult to achieve because of the release of P into the medium e.g., from dead cells. This is a common reason of the notoriously low efficiency of dense microalgal cultures at removal of low amounts of P (typical completeness of P removal is around 90% at initial P load of 4–30 mg L–1 [17]).
To achieve both goals of polyP-rich biomass production and removal of P from treated wastewater, a biphasic approach has been suggested when the bulk of P is removed by P-sufficient microalgal culture and the polishing of the effluent is carried out by a slightly P-starved culture [14,38]. Based on the fundamental metabolic knowledge discussed in Section 1.2.3 and 1.2.4., such systems would indeed trigger polyP synthesis and P uptake via up-regulation of P transporters and VTC proteins.
3.2.2. Algal-Bacterial Communities and P acquizition
Microalgae and bacteria are already forming a successful synergy in Waste Stabilization Ponds (WSPs), microalgae providing the oxygen needed for bacteria to degrade organic compounds. To the best of our knowledge, the extent of this synergy in relation to P is unknown. Bacteria can play a key role in P availability in soil, rendering P bioavailable through pH changes, however, P bioavailability should not be an issue during wastewater treatment. The environmental conditions in algae-based wastewater treatment systems are such that most of the P storage would be performed by the algae. However, many factors including the availability of the dissolved organic carbon can affect the efficiency of microalgae and cyanobacteria by modulating their relationships with heterotrophic bacteria and recycling of P released after cell death as was recently established for Microcystis [108]. A promising approach to using of the microalgal-bacterial co-cultures taking P from wastewater as biofertilizers is comprised by their immobilization on biodegradable carriers [109].
3.2.3. Phosphorus Load and Nutrient Balancing
Generally, the capability of microalgal culture to take P is determined by its cell P quota and cell division rate which can be limited by stresses like other nutrient shortage, cell shading or extreme temperatures. This translates into the requirements of optimal nutrient load and composition within the wastewater treated with the microalgal culture. Bioprocess for P biocapture with microalgae should be developed taking into account the available cell P quota and the amount of P to be removed: as mentioned above, over-saturated cells with their P quota full will not take up Pi even if it is available, and the conditions are favorable. Furthermore, the rate of the nutrient loading on the culture should match its growth rate and cell P quota size, otherwise the efficiency of P bioremoval will decline. Thus, nearly complete Pi removal can be easily achieved at P load around 5–10 mg L–1 but increasing P load above ca. 70–100 mg L–1 results in decline of the removal efficiency to 80–90% [17] (however, there can be more than one reason for such a behavior, see below).
Unfortunately, most of the reports on P removal of microalgae seldom include determination of cell P quota size (i.e., “benchmarking” of the cultures regarding their Pi uptake capacity). Moreover, current summaries of these reports lack indication of actual culture density (cell number and/or biomass content), pre-cultivation conditions, and its dynamics during the experiments. Consequently, it is difficult to infer the actual efficiency of the microalgal cultures at P bioremoval and it is even more difficult to compare results obtained in different studies with the same algal species and wastewater types.
Nutrient balancing is also important since P uptake of microalgal cultures experiencing a shortage of nitrogen, sulfur or other essential element shortage will be low as well. Still, the relatively low amounts of Pi which are taken up under conditions limiting cell division rate can be converted into polyP more efficiently [66,74]. In this situation, the accumulation of polyP per cell might increase [56] but the overall culture productivity will decline. On the other hand, P shortage seems to be less impactful for bioremoval of nitrogen [110].
Waste streams are frequently characterized by imbalance of the key nutrient (P, nitrogen, organic and inorganic carbon) composition differing considerably from their proportions of the Redfield ratio [16]. Particularly, nitrogen (more often) or P can be in excess, so to achieve a complete bioremoval of the nutrients the limiting one should be supplemented. An alternative approach comprises the selection of microalgal species/strains with smaller N/P ratio in the biomass matching that of the wastewater more closely [107].
Unforeseen problems may arise during treatment of wastewater contaminated by hazardous micropollutants and/or heavy metals. Although ample Pi can be, in principle, beneficial for the contaminant resilience of algal-bacterial communities in treatment facilities, toxic effects of the pollutants can deteriorate their P recovery capacity [111].
3.2.4. PolyP and Stress Resilience in the Context of Waste Stream Phycoremediation
As noted above, polyP due to its polyanionic nature functions as a counter-ion for metal cation sequestration, mainly in the vacuole. Normally, this function is carried out for physiologically relevant cations such as Na+, K+, Ca2+, or Mg2+ [66]. At the same time, polyP were shown to participate in detoxication of heavy metal cations, both required for essential cell functions (Mn2+, Zn2+, Fe3+, Cu2+) and toxic metals uninvolved in the cell metabolism (Cd2+, Hg2+, Pb2+) and even radionuclides such as cesium and uranium. Overall, the reports summarized in [63] are consistent with the existence of the relationship between polyP accumulation and tolerance to high external concentrations of heavy metals based likely on the ability of polyP to bind metal ions and thereby detoxify them. This relationship can be important for bioremediation of waste streams containing, apart from P, high amounts of heavy metals as is the case for rock phosphate mine leachate.
PolyP has also been suggested to participate in the mitigation of excessively alkaline pH harmful for photosynthesis and ATP generation which might arise in microalgal cells incubated in the presence of a high concentration of ammonium [112]. This capacity is especially relevant for bioremediation of certain wastewater types like dairy wastewater and aquaculture without biofilters rich in ammonia. There also indications of the polyP participation of the resilience to osmotic stress in microalgae by maintenance of ATP levels [113,114].
3.3. Microalgal Biomass Is an Efficient and Environtally Friendly Biofertilizer
The waste streams rich in macronutrients including P and organics cannot be applied for irrigation or fertilization directly because of the presence of hazardous micropollutants (e.g., antibiotics) and other substances endangering the soil as well as public health. This is reflected by the current legislative ban for application of the untreated wastewater on the fields. On the other hand, novel German regulation on P recovery from wastewater treatment plants effective from 2029 prescribes the recycling of 50% of P from sludge and 80% of P from ash [6].
Over the past decade, a consensus built up regarding the feasibility of closing the P loop by captured upcycling from waste streams with microalgal cultures [7]. One way to achieve this is to restrict the use of the conventional P fertilizers by increasingly substituting them by biofertilizer from P-rich microalgal biomass. This approach looks attractive above all when alternatives such as increasing P use efficiency either by purely conventional and/or genome modification-assisted breeding are challenging.
The effects of biofertilizers from microalgal biomass have been systematically studied for around two decades, although the studies dedicated to P are markedly scarcer that those focused e.g., on nitrogen fertilization (see [115,116,117] and references therein). It became clear in general that this type of biofertilizer has distinct advantages over traditional chemical fertilizers [18]. First of all, it is the rate of the release P available for the uptake by plants which, in case of microalgal biomass, turned to be close to the rate of P uptake by crop plants [118]. This makes the microalgal biomass a natural analog of the expensive man-made controlled-release fertilizers. Another important advantage is on the side of agrotechnology: microalgal biomass can be applied on the soil surface without tillering into the soil resulting in a considerable savings of fuel, labor, and reduced impact on the soil.
As fertilizers are intended to support soil fertility and increase the crop harvest size (see e.g., Table 2 in [18]), it is important to ensure that microalgal biomass satisfies these requirements. Indeed, recent experiments with nutrient-poor soil substrates showed that (i) the P from the microalgal biomass is nearly quantitatively transferred to the soil and then recovered by crop plants increasing available P in the soil 2–5 times [18] and (ii) the growth of the plants fertilized with microalgae was commensurate to that of the plants fertilized by conventional P fertilizers [118]. Fertilization with microalgal biomass is also expected to replenish the soil pool of bioavailable microelements such as Zn, Mn, Cu, which is depleted by intensive plant cultivation thereby preventing further limitations of these elements. Despite a considerable number of optimistic reports, the capability of the microalgal biomass as a substitute of the conventional P fertilizer depends on many factors such as crop type, soil, agricultural practices, and climatic conditions etc., so it must be evaluated on a case-by-case basis [119].
Additional beneficial effects on crops frequently arise due to hormone-like substances synthesized and released by the microalgae and, likely, their bacterial symbionts [117]. Strictly speaking, these effects cannot be termed as biofertilizer (since they are not related to the bulk inflow of the macronutrients and bioactive molecules to the soil). Application of microalgal biomass also increases the bioavailability of the P initially present in the soil [116] and improves its cycling [117]; however, this effect is of secondary importance in the context of this review, further details can be found elsewhere [117].
As reviewed in [117], the exogenous polysaccharides (EPS) produced by cyanobacteria and specific microalgae improve soil physical properties in agricultural settings, stabilize soil and form additional pores. The effect related with waterholding capacity was especially evident in experiments with inoculating low-organic carbon soils. In view of these findings, microalgal biomass applied as P biofertilizer is expected to exert beneficial effect(s) on the soil and crop condition. Indeed, no adverse changes either in the soil microbiome or in its activity were found following the fertilization with microalgal biomass [120].
Finally, fertilizers should not interfere with soil microbial activity and other parameters commonly designated as “soil health”. Microalgal biomass application was found to be beneficial for soil health by increasing the organic carbon content of the soil without measurable enhancement of greenhouse emission except for CO2 in certain cases [116,121]. Still, the data on the greenhouse gas emission from the soil fertilized by microalgal biomass remain controversial [18,122] as well as the comparative estimations of the integral environmental impact by the conventional (e.g., triple superphosphate) and microalgal biofertilizer production [123].
Although the evidence supporting the suitability of microalgal biomass for the production of the green P-rich biofertilizer, biostimulants, biocontrol agents, and soil conditioners is growing, there is a need for further research in this direction. Thus, the rate of microalgal biomass supplementation to the conventional P fertilizers such as triple superphosphate to reduce the application rate of the latter requires precise optimization to avoid adverse effects of biomass on the dissolvability of the superphosphate [106,124].
Another concern is the requirement of CO2 enrichment of the cultures to achieve realistic growth rate, but the demand for the inorganic carbon can be satisfied by nearby point CO2 sources such as flue gases from for e.g., thermal powerplants.
Critically, the technologies for economically viable large-scale growing of the P-rich microalgal biomass, its preservation and transportation are not yet established. Another possible limitation stems from the economic viability of and high energy consumption of microalgal-based biofertilizers . While currently high andestimated at 6.51 kWh per 1 g of recycled P, the use of photovoltaic solar energy during bioprocessing [18,122] will increase competitiveness with conventional fertilizers in the near future.
The production of biofertilizer from the wastewater-grown microalgal biomass requires strict safety control to exclude the contamination of the biomass and, subsequently, soil by hazardous micropollutants. The latter include heavy metal ions, microplastics, drugs, and nanoparticles which became increasingly widespread in the waste streams all over the world and are readily taken up by microalgae [125,126,127].
Admittedly, the development and application of microalgae as P biofertilizer needs further research which will be fueled by the expansion of their global biofertilizer market forecasted for the coming years (to USD 3.1 billion by the end of 2024 [128]).
3.4. PolyP as a Valuable Commodity
PolyP, due to their unique properties and relatively low cost of synthesis, nontoxicity, and biodegradability, are widely used in various industry (see above and [6,129]).
In addition to the functions of polyP within the cell (section 1.1.4), polyP also appears to be involved in symbiotic and parasitic associations. In higher non-photosynthetic eukaryotes, polyP levels were proposed to moderate host-pathogen interaction [130], play a role in cancer cell proliferation, apoptosis, exert a procoagulant and proinflammatory effects and disrupt TOR signaling (see [63,131] and references therein). PolyP also participate in bone tissue development, they are promising candidates in therapy for bone and blood diseases [131]. Accordingly, polyP are considered for application in developing novel bone substitute materials, carriers for prolonged action pharmaceuticals, and donor of P for enzymatic syntheses of biologically active compounds [132].
PolyPs are widely used as reagents in water treatment, fertilizers, flame retardants and food additives due to unique properties, nontoxicity and biodegradability (reviewed in [131]). Thus, polyPs are considered a less dangerous alternative to other water softeners and anticorrosion agents [131]. PolyPs are also widely used in the food industry as a food, specifically meat, preservative and water retaining agent. Still, taking into account the important role of PolyP in cell metabolism, bone tissue development and blood coagulation, it is also necessary to control polyP amounts in food.
Importantly, while the ability of microalgae to store P as polyP granules has been repeatedly demonstrated for decades, the exact chemistry of the granules still remains unclear. The central paradigm is that the polyP granules are formed from linear orthophosphoric acid units linked by phosphoanhydride bonds surrounded by counter-ions [70] and possibly other (organic) compounds [15]. While several reports showed that microalgal cell at different cellular ages stored different amount of polyP in different intracellular locations, the knowledge about different structures of polyPs (e.g., short vs long chain polyP etc) according to cell age or ecology is currently elusive. Because different types of polyP have specific chemical characteristics and consequently different biotechnological applications, polyPs characterization should be an area of future investigation in the field of microalgal P up-cycling.
4. Conclusions and Outlook
Sustainable usage of P, an irreplaceable macronutrient, is essential to solve the pressing problem of ensure food security of the increasing population while preserving the fragile ecosystem services and biodiversity. This problem is exacerbated by global climatic instability, exhaustion of water and fossil energy supply as well as by geopolitical complications. Recently announced stricter P discharge regulations are expected to intensify the cooperation of wastewater treatment plants and industrial partners, also in the field of bioremediation and circular bioeconomy. According to [6], a favored strategy in Germany is mono-combustion of the sludge and purification of the Pi from the residual ash which can be, in principle, accomplished with microalgal cultures.
This review clearly shows that microalgae have a formidable potential for valorizing P-rich waste to increase the sustainability of the usage of this indispensable macronutrient. The scientific community have made a significant progress in understanding the interactions in the P-microalgae-plant-environment system. Such advances have improved our understanding of the mechanisms of P acquisition, allocation, and regulation thereof at the molecular, cellular, organism, and community levels. Still, current knowledge of P metabolism and its regulation in microalgae is insufficient for the development of a robust, economically viable technology harnessing microalgae for efficient capture waste P and turning into valuable products. Particularly promising are the biofertilizers from P-rich microalgal biomass helping to re-route P from the anthropogenic output to agricultural ecosystems. Thus, the evidence on the interplay between native soil microbiome and the microbiome of microalgal culture introduced to the soil with microalgal biomass remains scarce and often limited to phenomenological descriptions.
P biofertilizers from microalgal biomass will especially be beneficial and hence most efficient when applied to marginal soils and the soils low on organic carbon content. Further effort is required to identify the strains combining a high P biocapture efficiency with resilience to wastewater components and fast growth capability. Native and artificial consortia formed by diverse microbes around microalgae deserve close attention in this regard for their synergistic beneficial effects on the soil and crop plants. More field tests including different crops, soil types, and agronomic practices needed to evaluate agronomic efficiency of the microalgae-based P biofertilizers.
A basic problem is finding the most economically efficient cultivation system and cutting down on the microalgal biomass production costs. Most likely it will be an outdoor growing system leveraging local waste nutrient sources (waste streams and concentrated CO2 e.g., from flue gas). To make sure that the developed solutions for P recycling with microalgae is economically feasible, a detailed techno-economic analysis will be required along with the assessment of greenhouse emissions and secondary environmental impacts. Further steps in this direction can be made by bioprospecting of microalgal strains more efficient at Pi uptake and PolyP accumulation [46].
The biotechnological achievements should complement other essential advances such as crop varieties bred for the P use efficiency, precision P fertilizer applications, and other measures needed to cut down the P loss to groundwater, inland water systems and the ocean with a corresponding decrease in eutrophication. Further study of polyP biochemistry and cell biology is important to expand their applications in medicine, environmental protection, and agriculture. The problem of research standardization and the need of strain benchmarking are highly relevant to the progress in this field.
A critical breakthrough in sustainable usage of P must be achieved to slow down our linear Pi use—from mining of phosphate rock, to human Pi-applications and finally to P sedimentation in the ocean (as highlighted by Blank [6] and Raven [133]), before the available phosphate reserves will be exhausted. This is projected to take place within two centuries or even earlier. Prudent use of the currently available and novel knowledge and technologies inspires us with hope to significantly delay – if not avoid – a looming P crisis, and to mitigate environmental and geopolitical problems associated with sustainable P use.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, A.S.; writing—original draft preparation, A.S and I.K..; writing—review and editing, A.S and M.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Russian Science Foundation, grant 23-44-00006.
Data Availability Statement
N/A.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Lauer, M.J.; Blevins, D.G.; Sierzputowska-Gracz, H. 31P-nuclear magnetic resonance determination of phosphate compartmentation in leaves of reproductive soybeans (Glycine max L.) as affected by phosphate nutrition. Plant Physiology 1989, 89, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Lambers, H.; Plaxton, W.C. Phosphorus: Back to the Roots. In Annual Plant Reviews Volume 48, John Wiley & Sons, Inc.: 2015; pp. 1-22. [CrossRef]
- Tiessen, H. Phosphorus in the global environment. In The Ecophysiology of Plant-Phosphorus Interactions, White, P.J., Hammond, J.P., Eds. Springer Netherlands: Dordrecht, 2008; pp. 1-7. [CrossRef]
- Bennett, E.; Elser, J. A broken biogeochemical cycle. Nature 2011, 478, 29–31. [Google Scholar]
- Cembella, A.D.; Antia, N.J.; Harrison, P.J. The utilization of inorganic and organic phosphorous compounds as nutrients by eukaryotic microalgae: A multidisciplinary perspective: Part I. Critical reviews in microbiology 1982, 10, 317–391. [Google Scholar] [CrossRef] [PubMed]
- Blank, L.M. (Poly)phosphate biotechnology: Envisaged contributions to a sustainable P future. Microb Biotechnol 2023, 16, 1616–1622. [Google Scholar] [CrossRef] [PubMed]
- Solovchenko, A.; Verschoor, A.M.; Jablonowski, N.D.; Nedbal, L. Phosphorus from wastewater to crops: An alternative path involving microalgae. Biotechnology advances 2016, 34, 550–564. [Google Scholar] [CrossRef] [PubMed]
- Cordell, D.; White, S. Life’s Bottleneck: Implications of Global Phosphorus Scarcity and Pathways for a Sustainable Food System. Annual Review of Environment and Resources 2014, 39. [Google Scholar]
- Fixen, P.E.; Johnston, A.M. World fertilizer nutrient reserves: a view to the future. Journal of the Science of Food and Agriculture 2012, 92, 1001–1005. [Google Scholar] [CrossRef]
- Simpson, R.J.; Oberson, A.; Culvenor, R.A.; Ryan, M.H.; Veneklaas, E.J.; Lambers, H.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A. Strategies and agronomic interventions to improve the phosphorus-use efficiency of farming systems. Plant and Soil 2011, 349, 89–120. [Google Scholar] [CrossRef]
- Hallegraeff, G.; Anderson, D.; Cembella, A. Manual on harmful marine microalgae; UNESCO Publising:, 2003; p. 770. [Google Scholar]
- Smith, V.H.; Schindler, D.W. Eutrophication science: where do we go from here? Trends in ecology & evolution 2009, 24, 201–207. [Google Scholar]
- Grossman, A.R.; Aksoy, M. Algae in a phosphorus-limited landscape. In Annual Plant Reviews, Phosphorus Metabolism in Plants, Plaxton, W., Lambers, H., Eds.; Wiley-Blackwell: 2015; Vol. 48, pp. 337–374.
- Solovchenko, A.E.; Ismagulova, T.T.; Lukyanov, A.A.; Vasilieva, S.G.; Konyukhov, I.V.; Pogosyan, S.I.; Lobakova, E.S.; Gorelova, O.A. Luxury phosphorus uptake in microalgae. Journal of Applied Phycology 2019, 31, 2755–2770. [Google Scholar] [CrossRef]
- Solovchenko, A.; Khozin-Goldberg, I.; Selyakh, I.; Semenova, L.; Ismagulova, T.; Lukyanov, A.; Mamedov, I.; Vinogradova, E.; Karpova, O.; Konyukhov, I.; et al. Phosphorus starvation and luxury uptake in green microalgae revisited. Algal Research 2019, 43, 101651. [Google Scholar] [CrossRef]
- Redfield, A.C. The biological control of chemical factors in the environment. American scientist 1958, 46, 230A–221. [Google Scholar]
- Yu, D.; Yan, L.; Shi, J.; Liu, Y.; Zhang, A.; Wang, Y.; Zhang, Y.; Xie, T. Phosphorus Removal and Recovery During Microalgae-Based Wastewater Treatment: A Mini-review. International Journal of Environmental Research 2024, 18, 34. [Google Scholar] [CrossRef]
- Cao, T.N.-D.; Mukhtar, H.; Le, L.-T.; Tran, D.P.-H.; Ngo, M.T.T.; Nguyen, T.-B.; Bui, X.-T. Roles of microalgae-based biofertilizer in sustainability of green agriculture and food-water-energy security nexus. Science of The Total Environment 2023, 870, 161927. [Google Scholar] [CrossRef]
- Crimp, A.; Brown, N.; Shilton, A. Microalgal luxury uptake of phosphorus in waste stabilization ponds – frequency of occurrence and high performing genera. Water Science and Technology 2017, 78, 165–173. [Google Scholar] [CrossRef] [PubMed]
- You, K.; Ge, F.; Wu, X.; Song, K.; Yang, Z.; Zhang, Q.; Liu, Y.; Ruan, R.; Zheng, H. Nutrients recovery from piggery wastewater and starch wastewater via microalgae-bacteria consortia. Algal Research 2021, 60, 102551. [Google Scholar] [CrossRef]
- Wang, S.; Li, N.; Yuan, Q.; Liang, D.; Chang, J.; Wang, X.; Ren, N. Vivianite recovery from high concentration phosphorus wastewater with mine drainage as iron sources. Science of The Total Environment 2023, 858, 160098. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Dyhrman, S.T.; Lomas, M.W.; Poulton, N.J.; Van Mooy, B.A.S. Accumulation and enhanced cycling of polyphosphate by Sargasso Sea plankton in response to low phosphorus. Proceedings of the National Academy of Sciences 2014, 111, 8089–8094. [Google Scholar] [CrossRef]
- Diaz, J.M.; Björkman, K.M.; Haley, S.T.; Ingall, E.D.; Karl, D.M.; Longo, A.F.; Dyhrman, S.T. Polyphosphate dynamics at Station ALOHA, North Pacific subtropical gyre. Limnology and Oceanography 2016, 61, 227–239. [Google Scholar] [CrossRef]
- Bolier, G.; de Koningh, M.C.J.; Schmale, J.C.; Donze, M. Differential luxury phosphate response of planktonic algae to phosphorus removal. Hydrobiologia 1992, 243, 113–118. [Google Scholar] [CrossRef]
- Li, J.; Plouchart, D.; Zastepa, A.; Dittrich, M. Picoplankton accumulate and recycle polyphosphate to support high primary productivity in coastal Lake Ontario. Scientific Reports 2019, 9, 19563. [Google Scholar] [CrossRef] [PubMed]
- Weihrauch, C.; Opp, C. Ecologically relevant phosphorus pools in soils and their dynamics: The story so far. Geoderma 2018, 325, 183–194. [Google Scholar] [CrossRef]
- Fang, Z.; Shao, C.; Meng, Y.; Wu, P.; Chen, M. Phosphate signaling in Arabidopsis and Oryza sativa. Plant Science 2009, 176, 170–180. [Google Scholar] [CrossRef]
- Hentrich, S.; Hebeler, M.; Grimme, L.H.; Leibfritz, D.; Mayer, A. P-31 NMR saturation transfer experiments in Chlamydomonas reinhardtii : evidence for the NMR visibility of chloroplastidic Pi. European Biophysics Journal 1993, 22, 31–39. [Google Scholar] [CrossRef]
- Filippelli, G.M. The global phosphorus cycle: past, present, and future. Elements 2008, 4, 89–95. [Google Scholar] [CrossRef]
- Gross, M. Where is all the phosphorus? Current Biology 2017, 27, R1141–R1144. [Google Scholar] [CrossRef]
- Smil, V. Phosphorus in the environment: natural flows and human interferences. Annual review of energy and the environment 2000, 25, 53–88. [Google Scholar] [CrossRef]
- Shen, A.; Gao, S.; Jiang, J.; Hu, Q.; Wang, H.; Yuan, S. Oscillations of algal cell quota: Considering two-stage phosphate uptake kinetics. J Theor Biol 2024, 581, 111739. [Google Scholar] [CrossRef] [PubMed]
- Solovchenko, A.; Gorelova, O.; Karpova, O.; Selyakh, I.; Semenova, L.; Chivkunova, O.; Baulina, O.; Vinogradova, E.; Pugacheva, T.; Scherbakov, P. Phosphorus feast and famine in cyanobacteria: is luxury uptake of the nutrient just a consequence of acclimation to its shortage? Cells 2020, 9, 1933. [Google Scholar] [CrossRef]
- Lobakova, E.S.; Selyakh, I.O.; Semenova, L.R.; Scherbakov, P.N.; Fedorenko, T.A.; Chekanov, K.A.; Chivkunova, O.B.; Baulina, O.I.; Vasilieva, S.G.; Solovchenko, A.E. Hints for understanding microalgal phosphate-resilience from Micractinium simplicissimum IPPAS C-2056 (Trebouxiophyceae) isolated from a phosphorus-polluted site. Journal of Applied Phycology 2022, 34, 2409–2422. [Google Scholar] [CrossRef]
- Brown, N.; Sells, M.; Jayamaha, N.; Shilton, A. Predicting phosphorus accumulation and proposing conditions needed for an algal-based phosphorus uptake process. Environmental Technology 2018, 132, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Sells, M.D.; Brown, N.; Shilton, A.N. Determining variables that influence the phosphorus content of waste stabilization pond algae. Water Research 2018, 132, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Powell, N.; Shilton, A.N.; Pratt, S.; Chisti, Y. Factors influencing luxury uptake of phosphorus by microalgae in waste stabilization ponds. Environmental Science & Technology 2008, 42, 5958–5962. [Google Scholar]
- Lavrinovičs, A.; Mežule, L.; Juhna, T. Microalgae starvation for enhanced phosphorus uptake from municipal wastewater. Algal Research 2020, 52, 102090. [Google Scholar] [CrossRef]
- Sforza, E.; Calvaruso, C.; La Rocca, N.; Bertucco, A. Luxury uptake of phosphorus in Nannochloropsis salina: Effect of P concentration and light on P uptake in batch and continuous cultures. Biochemical Engineering Journal 2018, 134, 69–79. [Google Scholar] [CrossRef]
- Grobbelaar, J.U. Inorganic Algal Nutrition. In Handbook of Microalgal Culture: Applied Phycology and Biotechnology, 2 ed.; Richmond, A., Hu, Q., Eds.; John WIley and Sons: Chichester, UK, 2013; pp. 123–133. [Google Scholar]
- Brown, N.; Shilton, A. Luxury uptake of phosphorus by microalgae in waste stabilisation ponds: current understanding and future direction. Reviews in Environmental Science and Bio/Technology 2014, 13, 321–328. [Google Scholar] [CrossRef]
- Plouviez, M.; Bolot, P.; Shilton, A.; Guieysse, B. Phosphorus uptake and accumulation in Chlamydomonas reinhardtii: Influence of biomass concentration, phosphate concentration, phosphorus depletion time, and light supply. Algal Research 2023, 71. [Google Scholar] [CrossRef]
- Jensen, T.E.; Sicko, L.M. Phosphate metabolism in blue-green algae. I. Fine structure of the” polyphosphate overplus” phenomenon in Plectonema boryanum. Canadian journal of microbiology 1974, 20, 1235–1239. [Google Scholar] [CrossRef] [PubMed]
- Grillo, J.F.; Gibson, J. Regulation of phosphate accumulation in the unicellular cyanobacterium Synechococcus. Journal of bacteriology 1979, 140, 508–517. [Google Scholar] [CrossRef]
- Aitchison, P.; Butt, V. The relation between the synthesis of inorganic polyphosphate and phosphate uptake by Chlorella vulgaris. Journal of Experimental Botany 1973, 24, 497–510. [Google Scholar] [CrossRef]
- Schaedig, E.; Cantrell, M.; Urban, C.; Zhao, X.; Greene, D.; Dancer, J.; Gross, M.; Sebesta, J.; Chou, K.J.; Grabowy, J.; et al. Isolation of phosphorus-hyperaccumulating microalgae from revolving algal biofilm (RAB) wastewater treatment systems. Front Microbiol 2023, 14, 1219318. [Google Scholar] [CrossRef] [PubMed]
- Christ, J.J.; Willbold, S.; Blank, L.M. Methods for the Analysis of Polyphosphate in the Life Sciences. Analytical Chemistry 2020, 92, 4167–4176. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.R.; Kornberg, A. Inorganic polyphosphate in the origin and survival of species. Proceedings of the National Academy of Sciences of the United States of America 2004, 101, 16085–16087. [Google Scholar] [CrossRef] [PubMed]
- Rasala, B.; Mayfield, S. Photosynthetic biomanufacturing in green algae; production of recombinant proteins for industrial, nutritional, and medical uses. Photosynthesis Research 2015, 123, 227–239. [Google Scholar] [CrossRef]
- Blank, L.M. The cell and P: from cellular function to biotechnological application. Current Opinion in Biotechnology 2012, 23, 846–851. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Wang, X.; Tong, Y.; Chen, X.; Liao, H. Bioengineering and management for efficient phosphorus utilization in crops and pastures. Current Opinion in Biotechnology 2012, 23, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Cook, R.; Lupette, J.; Benning, C. The Role of Chloroplast Membrane Lipid Metabolism in Plant Environmental Responses. Cells 2021, 10, 706. [Google Scholar] [CrossRef] [PubMed]
- Mühlroth, A.; Winge, P.; El Assimi, A.; Jouhet, J.; Maréchal, E.; Hohmann-Marriott, M.F.; Vadstein, O.; Bones, A.M. Mechanisms of Phosphorus Acquisition and Lipid Class Remodeling under P Limitation in a Marine Microalga. Plant Physiology 2017, 175, 1543–1559. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A. Interactions between nitrogen and phosphorus metabolism. Annual Plant Reviews Volume 48: Phosphorus Metabolism in Plants 2015, 48, 187–214. [Google Scholar]
- Raghothama, K. Phosphate acquisition. Annual review of plant biology 1999, 50, 665–693. [Google Scholar] [CrossRef]
- Rao, N.N.; Gómez-García, M.R.; Kornberg, A. Inorganic polyphosphate: essential for growth and survival. Annual review of biochemistry 2009, 78, 605–647. [Google Scholar] [CrossRef] [PubMed]
- Kamennaya, N.A.; Geraki, K.; Scanlan, D.J.; Zubkov, M.V. Accumulation of ambient phosphate into the periplasm of marine bacteria is proton motive force dependent. Nat Commun 2020, 11, 2642. [Google Scholar] [CrossRef] [PubMed]
- Grossman, A.; Takahashi, H. Macronutrient utilization by photosynthetic eukaryotes and the fabric of interactions. Annual Review of Plant Biology 2001, 52, 163–210. [Google Scholar] [CrossRef] [PubMed]
- Shimogawara, K.; Wykoff, D.D.; Usuda, H.; Grossman, A.R. Chlamydomonas reinhardtii Mutants Abnormal in Their Responses to Phosphorus Deprivation. Plant Physiology 1999, 120, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xiao, L.; Yang, H.; Chen, G.; Zeng, H.; Zhao, H.; Zhu, Y. Genome-Wide Identification, Expression Profiling, and Evolution of Phosphate Transporter Gene Family in Green Algae. Front Genet 2020, 11, 590947. [Google Scholar] [CrossRef] [PubMed]
- Falkner, G.; Falkner, R. The complex regulation of the phosphate uptake system of cyanobacteria. In Bioenergetic Processes of Cyanobacteria; Springer: 2011; pp. 109–130.
- Achbergerová, L.; Nahálka, J. Polyphosphate-an ancient energy source and active metabolic regulator. Microb Cell Fact 2011, 10, 14170–14175. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Luque, E.; Bhaya, D.; Grossman, A.R. Polyphosphate: A Multifunctional Metabolite in Cyanobacteria and Algae. Frontiers in Plant Science 2020, 11. [Google Scholar] [CrossRef]
- Kulaev, I.; Vagabov, I.; Kulakovskaya, T. The Biochemistry of Inorganic Polyphosphates, 2 ed.; John Wiley & Sons, Ltd.: Chichester, England, 2004. [Google Scholar]
- Ruiz, F.A.; Marchesini, N.; Seufferheld, M.; Govindjee, *!!! REPLACE !!!*; Docampo, R. The Polyphosphate Bodies of Chlamydomonas reinhardtii Possess a Proton-pumping Pyrophosphatase and Are Similar to Acidocalcisomes. Journal of Biological Chemistry 2001, 276, 46196–46203. [Google Scholar] [CrossRef]
- Goodenough, U.; Heiss, A.A.; Roth, R.; Rusch, J.; Lee, J.-H. Acidocalcisomes: Ultrastructure, Biogenesis, and Distribution in Microbial Eukaryotes. Protist 2019, 170, 287–313. [Google Scholar] [CrossRef]
- Guan, Z.; Chen, J.; Liu, R.; Chen, Y.; Xing, Q.; Du, Z.; Cheng, M.; Hu, J.; Zhang, W.; Mei, W.; et al. The cytoplasmic synthesis and coupled membrane translocation of eukaryotic polyphosphate by signal-activated VTC complex. Nat Commun 2023, 14, 718. [Google Scholar] [CrossRef]
- Desfougeres, Y.; Gerasimaite, R.U.; Jessen, H.J.; Mayer, A. Vtc5, a Novel Subunit of the Vacuolar Transporter Chaperone Complex, Regulates Polyphosphate Synthesis and Phosphate Homeostasis in Yeast. J Biol Chem 2016, 291, 22262–22275. [Google Scholar] [CrossRef] [PubMed]
- Müller, O.; Neumann, H.; Bayer, M.J.; Mayer, A. Role of the Vtc proteins in V-ATPase stability and membrane trafficking. Journal of Cell Science 2003, 116, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Cliff, A.; Guieysse, B.; Brown, N.; Lockhart, P.; Dubreucq, E.; Plouviez, M. Polyphosphate synthesis is an evolutionarily ancient phosphorus storage strategy in microalgae. Algal Research 2023. [Google Scholar] [CrossRef]
- Plouviez, M.; Abyadeh, M.; Hasan, M.; Mirzaei, M.; Paulo, J.A.; Guieysse, B. The proteome of Chlamydomonas reinhardtii during phosphorus depletion and repletion. Algal Research 2023, 71. [Google Scholar] [CrossRef]
- Plouviez, M.; Fernandez, E.; Grossman, A.R.; Sanz-Luque, E.; Sells, M.; Wheeler, D.; Guieysse, B. Responses of Chlamydomonas reinhardtii during the transition from P-deficient to P-sufficient growth (the P-overplus response): The roles of the vacuolar transport chaperones and polyphosphate synthesis. J Phycol 2021. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Garcia, M.R.; Fazeli, F.; Grote, A.; Grossman, A.R.; Bhaya, D.J.J.o.b. Role of polyphosphate in thermophilic Synechococcus sp. from microbial mats. 2013, 195, 3309–3319. [Google Scholar]
- Ota, S.; Yoshihara, M.; Yamazaki, T.; Takeshita, T.; Hirata, A.; Konomi, M.; Oshima, K.; Hattori, M.; Bisova, K.; Zachleder, V.; et al. Deciphering the relationship among phosphate dynamics, electron-dense body and lipid accumulation in the green alga Parachlorella kessleri. Scienific Reports 2016, 6, 25731. [Google Scholar] [CrossRef] [PubMed]
- Ryu, H.B.; Kang, M.J.; Choi, K.M.; Yang, I.K.; Hong, S.J.; Lee, C.G. Inhibition of Polyphosphate Degradation in Synechocystis sp. PCC6803 through Inactivation of the phoU Gene. J Microbiol Biotechnol 2024, 34, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Sebesta, J.; Cantrell, M.; Schaedig, E.; Hou, H.J.M.; Pastore, C.; Chou, K.J.; Xiong, W.; Guarnieri, M.T.; Yu, J. Polyphosphate kinase deletion increases laboratory productivity in cyanobacteria. Front Plant Sci 2024, 15, 1342496. [Google Scholar] [CrossRef]
- Voronkov, A.; Sinetova, M. Polyphosphate accumulation dynamics in a population of Synechocystis sp. PCC 6803 cells under phosphate overplus. Protoplasma, 2019. [Google Scholar] [CrossRef]
- Paytan, A.; McLaughlin, K. The oceanic phosphorus cycle. Chemical Reviews 2007, 107, 563–576. [Google Scholar] [CrossRef]
- Moseley, J.L.; Chang, C.-W.; Grossman, A.R.J.E.c. Genome-based approaches to understanding phosphorus deprivation responses and PSR1 control in Chlamydomonas reinhardtii. 2006, 5, 26–44.
- Falkner, R.; Falkner, G. Distinct adaptivity during phosphate uptake by the cyanobacterium Anabaena variabilis reflects information processing about preceding phosphate supply. Journal of trace and microprobe techniques 2003, 21, 363–375. [Google Scholar] [CrossRef]
- Vítová, M.; Bišová, K.; Kawano, S.; Zachleder, V. Accumulation of energy reserves in algae: From cell cycles to biotechnological applications. Biotechnology Advances 2015, 33, 1204–1218. [Google Scholar] [CrossRef] [PubMed]
- Bertilsson, S.; Berglund, O.; Karl, D.M.; Chisholm, S.W. Elemental composition of marine Prochlorococcus and Synechococcus: Implications for the ecological stoichiometry of the sea. Limnology and oceanography 2003, 48, 1721–1731. [Google Scholar] [CrossRef]
- Abida, H.; Dolch, L.-J.; Meï, C.; Villanova, V.; Conte, M.; Block, M.A.; Finazzi, G.; Bastien, O.; Tirichine, L.; Bowler, C.; et al. Membrane Glycerolipid Remodeling Triggered by Nitrogen and Phosphorus Starvation in Phaeodactylum tricornutum. Plant Physiology 2014, 167, 118–136. [Google Scholar] [CrossRef]
- Canavate, J.P.; Armada, I.; Hachero-Cruzado, I. Aspects of phosphorus physiology associated with phosphate-induced polar lipid remodelling in marine microalgae. Journal of Plant Physiology 2017, 214, 28–38. [Google Scholar] [CrossRef]
- Bolik, S.; Schlaich, A.; Mukhina, T.; Amato, A.; Bastien, O.; Schneck, E.; Demé, B.; Jouhet, J. Lipid bilayer properties potentially contributed to the evolutionary disappearance of betaine lipids in seed plants. BMC biology 2023, 21, 275. [Google Scholar] [CrossRef]
- Gomez, R.E.; Castets, J.; Lupette, J.; Chambaud, C.; Joubès, J.; Bernard, A. Phosphatidylinositol-4-phosphate joins the dance of plant autophagosome formation. Autophagy 2023, 19, 1609–1610. [Google Scholar] [CrossRef]
- Kokabi, K.; Gorelova, O.; Ismagulova, T.; Itkin, M.; Malitsky, S.; Boussiba, S.; Solovchenko, A.; Khozin-Goldberg, I. Metabolomic foundation for differential responses of lipid metabolism to nitrogen and phosphorus deprivation in an arachidonic acid-producing green microalga. Plant Science 2019, 283, 95–115. [Google Scholar] [CrossRef] [PubMed]
- Shemi, A.; Schatz, D.; Fredricks, H.F.; Van Mooy, B.A.; Porat, Z.; Vardi, A. Phosphorus starvation induces membrane remodeling and recycling in Emiliania huxleyi. New Phytologist 2016, 211, 886–898. [Google Scholar] [CrossRef]
- Martin, P.; Dyhrman, S.T.; Lomas, M.W.; Poulton, N.J.; Van Mooy, B.A. Accumulation and enhanced cycling of polyphosphate by Sargasso Sea plankton in response to low phosphorus. Proc Natl Acad Sci U S A 2014, 111, 8089–8094. [Google Scholar] [CrossRef]
- Gorelova, O.; Baulina, O.; Ismagulova, T.; Kokabi, K.; Lobakova, E.; Selyakh, I.; Semenova, L.; Chivkunova, O.; Karpova, O.; Scherbakov, P. Stress-induced changes in the ultrastructure of the photosynthetic apparatus of green microalgae. Protoplasma 2019, 256, 261–277. [Google Scholar] [CrossRef] [PubMed]
- Brányiková, I.; Maršálková, B.; Doucha, J.; Brányik, T.; Bišová, K.; Zachleder, V.; Vítová, M. Microalgae—novel highly efficient starch producers. Biotechnology and bioengineering 2011, 108, 766–776. [Google Scholar] [CrossRef] [PubMed]
- de Mazancourt, C.; Schwartz, M.W. Starve a competitor: evolution of luxury consumption as a competitive strategy. Theoretical Ecology 2010, 5, 37–49. [Google Scholar] [CrossRef]
- Tetu, S.G.; Brahamsha, B.; Johnson, D.A.; Tai, V.; Phillippy, K.; Palenik, B.; Paulsen, I.T. Microarray analysis of phosphate regulation in the marine cyanobacterium Synechococcus sp. WH8102. The ISME Journal 2009, 3, 835–849. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Martin, M.A.; Mateo, P.; Leganes, F.; Fernandez-Pinas, F. Novel cyanobacterial bioreporters of phosphorus bioavailability based on alkaline phosphatase and phosphate transporter genes of Anabaena sp. PCC 7120. Anal Bioanal Chem 2011, 400, 3573–3584. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Zhang, H.; Huang, B.; Lin, S. Alkaline phosphatase gene sequence and transcriptional regulation by phosphate limitation in Amphidinium carterae (Dinophyceae) 1. Journal of phycology 2011, 47, 1110–1120. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Litaker, R.W.; Sunda, W.G. Phosphorus physiological ecology and molecular mechanisms in marine phytoplankton. Journal of Phycology 2016, 52, 10–36. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, K.R.; Fees, J.; Christ, J.J.; Hofmann, I.; Block, C.; Herzberg, D.; Bröring, S.; Reckels, B.; Visscher, C.; Blank, L.M.; et al. Biotechnological production of food-grade polyphosphate from deoiled seeds and bran. EFB Bioeconomy Journal 2023, 3, 100048. [Google Scholar] [CrossRef]
- Lobakova, E.; Gorelova, O.; Selyakh, I.; Semenova, L.; Scherbakov, P.; Vasilieva, S.; Zaytsev, P.; Shibzukhova, K.; Chivkunova, O.; Baulina, O. Failure of Micractinium simplicissimum Phosphate Resilience upon Abrupt Re-Feeding of Its Phosphorus-Starved Cultures. International Journal of Molecular Sciences 2023, 24, 8484. [Google Scholar] [CrossRef]
- Li, Q.; Fu, L.; Wang, Y.; Zhou, D.; Rittmann, B.E. Excessive phosphorus caused inhibition and cell damage during heterotrophic growth of Chlorella regularis. Bioresource Technology 2018, 268, 266–270. [Google Scholar] [CrossRef]
- Fu, L.; Li, Q.; Yan, G.; Zhou, D.; Crittenden, J.C. Hormesis effects of phosphorus on the viability of Chlorella regularis cells under nitrogen limitation. Biotechnol Biofuels 2019, 12, 121. [Google Scholar] [CrossRef] [PubMed]
- Cogliatti, D.H.; Clarkson, D.T. Physiological changes in, and phosphate uptake by potato plants during development of, and recovery from phosphate deficiency. Physiologia Plantarum 1983, 58, 287–294. [Google Scholar] [CrossRef]
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S.J.N. Agricultural sustainability and intensive production practices. 2002, 418, 671.
- Martín, H.G.; Ivanova, N.; Kunin, V.; Warnecke, F.; Barry, K.W.; McHardy, A.C.; Yeates, C.; He, S.; Salamov, A.A.; Szeto, E. Metagenomic analysis of two enhanced biological phosphorus removal (EBPR) sludge communities. Nature biotechnology 2006, 24, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, R.; Guieysse, B. Algal-bacterial processes for the treatment of hazardous contaminants: A review. Water research 2006, 40, 2799–2815. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.L.; Rodrigues, C.M.; Pires, J.C.M.; Simões, M. The effect of increasing CO2 concentrations on its capture, biomass production and wastewater bioremediation by microalgae and cyanobacteria. Algal Research 2016, 14, 127–136. [Google Scholar] [CrossRef]
- de Siqueira Castro, J.; Calijuri, M.L.; Mattiello, E.M.; Ribeiro, V.J.; Assemany, P.P. Algal biomass from wastewater: soil phosphorus bioavailability and plants productivity. Science of the total environment 2020, 711, 135088. [Google Scholar] [CrossRef]
- Kandasamy, S.; Narayanan, M.; Raja, R.; Devarayan, K.; Kavitha, R. The current state of algae in wastewater treatment and energy conversion: a critical review. Current Opinion in Environmental Science & Health 2023, 100469. [Google Scholar]
- Li, T.; Xu, L.; Li, W.; Wang, C.; Gin, K.Y.; Chai, X.; Wu, B. Dissolved organic carbon spurs bacterial-algal competition and phosphorus-paucity adaptation: Boosting Microcystis’ phosphorus uptake capacity. Water Res 2024, 255, 121465. [Google Scholar] [CrossRef] [PubMed]
- Odibo, A.; Janpum, C.; Pombubpa, N.; Monshupanee, T.; Incharoensakdi, A.; Ur Rehman, Z.; In-Na, P. Microalgal-bacterial immobilized co-culture as living biofilters for nutrient recovery from synthetic wastewater and their potential as biofertilizers. Bioresour Technol 2024, 398, 130509. [Google Scholar] [CrossRef]
- Jakhwal, P.; Daneshvar, E.; Skalska, K.; Matsakas, L.; Patel, A.; Park, Y.; Bhatnagar, A. Nutrient removal and biomass production of marine microalgae cultured in recirculating aquaculture systems (RAS) water with low phosphate concentration. J Environ Manage 2024, 358, 120859. [Google Scholar] [CrossRef]
- Tang, C.C.; Hu, Y.R.; Zhang, M.; Chen, S.L.; He, Z.W.; Li, Z.H.; Tian, Y.; Wang, X.C. Role of phosphate in microalgal-bacterial symbiosis system treating wastewater containing heavy metals. Environ Pollut 2024, 123951. [Google Scholar] [CrossRef]
- Pick, U.; Bental, M.; Chitlaru, E.; Weiss, M. Polyphosphate-hydrolysis-a protective mechanism against alkaline stress? FEBS Letters 1990, 274, 15–18. [Google Scholar]
- Bental, M.; Pick, U.; AVRON, M.; Degani, H. The role of intracellular orthophosphate in triggering osmoregulation in the alga Dunaliella salina. European journal of biochemistry 1990, 188, 117–122. [Google Scholar] [CrossRef]
- Leitão, J.M.; Lorenz, B.; Bachinski, N.; Wilhelm, C.; Müller, W.E.; Schröder, H.C. Osmotic-stress-induced synthesis and degradation of inorganic polyphosphates in the alga Phaeodactylumtricornutum. Marine Ecology Progress Series 1995, 121, 279–288. [Google Scholar] [CrossRef]
- Miranda, A.M.; Hernandez-Tenorio, F.; Villalta, F.; Vargas, G.J.; Sáez, A.A. Advances in the Development of Biofertilizers and Biostimulants from Microalgae. Biology 2024, 13, 199. [Google Scholar] [CrossRef]
- Ammar, E.E.; Aioub, A.A.A.; Elesawy, A.E.; Karkour, A.M.; Mouhamed, M.S.; Amer, A.A.; El-Shershaby, N.A. Algae as Bio-fertilizers: Between current situation and future prospective. Saudi Journal of Biological Sciences 2022, 29, 3083–3096. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, A.L.; Weyers, S.L.; Goemann, H.M.; Peyton, B.M.; Gardner, R.D. Microalgae, soil and plants: A critical review of microalgae as renewable resources for agriculture. Algal Research 2021, 54, 102200. [Google Scholar] [CrossRef]
- Schreiber, C.; Schiedung, H.; Harrison, L.; Briese, C.; Ackermann, B.; Kant, J.; Schrey, S.D.; Hofmann, D.; Singh, D.; Ebenhöh, O.; et al. Evaluating potential of green alga Chlorella vulgaris to accumulate phosphorus and to fertilize nutrient-poor soil substrates for crop plants. Journal of Applied Phycology 2018, 30, 2827–2836. [Google Scholar] [CrossRef]
- Alvarez-Gonzalez, A.; Uggetti, E.; Serrano, L.; Gorchs, G.; Ferrer, I.; Diez-Montero, R. Can microalgae grown in wastewater reduce the use of inorganic fertilizers? J Environ Manage 2022, 323, 116224. [Google Scholar] [CrossRef]
- Kublanovskaya, A.; Khapchaeva, S.; Zotov, V.; Zaytsev, P.; Lobakova, E.; Solovchenko, A. The effect of the microalga Chlorella vulgaris Ippas C-1 biomass application on yield, biological activity, and the microbiome of the soil during bean growing. Moscow University Biological Sciences Bulletin 2019, 74, 227–234. [Google Scholar] [CrossRef]
- Solovchenko, A.; Zaitsev, P.; Zotov, V. Phosphorus biofertilizer from microalgae. Biofertilizers 2021, 57–68. [Google Scholar]
- de Souza, M.H.B.; Calijuri, M.L.; Assemany, P.P.; Castro, J.d.S.; de Oliveira, A.C.M. Soil application of microalgae for nitrogen recovery: A life-cycle approach. Journal of Cleaner Production 2019, 211, 342–349. [Google Scholar] [CrossRef]
- Castro, J.d.S.; Calijuri, M.L.; Ferreira, J.; Assemany, P.P.; Ribeiro, V.J. Microalgae based biofertilizer: A life cycle approach. Science of The Total Environment 2020, 724, 138138. [Google Scholar] [CrossRef]
- Sharma, G.K.; Khan, S.A.; Shrivastava, M.; Bhattacharyya, R.; Sharma, A.; Gupta, D.K.; Kishore, P.; Gupta, N. Circular economy fertilization: Phycoremediated algal biomass as biofertilizers for sustainable crop production. Journal of Environmental Management 2021, 287, 112295. [Google Scholar] [CrossRef]
- Cao, M.; Wang, F.; Zhou, B.; Chen, H.; Yuan, R.; Ma, S.; Geng, H.; Li, J.; Lv, W.; Wang, Y.; et al. Nanoparticles and antibiotics stress proliferated antibiotic resistance genes in microalgae-bacteria symbiotic systems. J Hazard Mater 2023, 443, 130201. [Google Scholar] [CrossRef]
- Eheneden, I.; Wang, R.; Zhao, J. Antibiotic removal by microalgae-bacteria consortium: Metabolic pathways and microbial responses. Sci Total Environ 2023, 891, 164489. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Jin, P.; Xie, S.; Beardall, J.; Feng, Y.; Guo, C.; Ma, Z.; Gao, G. Micro- and nanoplastics interact with conventional pollutants on microalgae: Synthesis through meta-analysis. Environ Pollut 2023. [Google Scholar] [CrossRef]
- Behera, B.; Venkata Supraja, K.; Paramasivan, B. Integrated microalgal biorefinery for the production and application of biostimulants in circular bioeconomy. Bioresource Technology 2021, 339, 125588. [Google Scholar] [CrossRef]
- Demling, P.; Baier, M.; Deitert, A.; Fees, J.; Blank, L.M. Biotechnological polyphosphate as an opportunity to contribute to the circularization of the phosphate economy. Current Opinion in Biotechnology 2024, 87, 103107. [Google Scholar] [CrossRef] [PubMed]
- Bowlin, M.Q.; Gray, M.J. Inorganic polyphosphate in host and microbe biology. Trends Microbiol 2021, 29, 1013–1023. [Google Scholar] [CrossRef]
- Kulakovskaya, T.V.; Vagabov, V.M.; Kulaev, I.S. Inorganic polyphosphate in industry, agriculture and medicine: Modern state and outlook. Process Biochemistry 2012, 47, 1–10. [Google Scholar] [CrossRef]
- Feng, G.; Dong, S.; Huang, M.; Zeng, M.; Liu, Z.; Zhao, Y.; Wu, H. Biogenic Polyphosphate Nanoparticles from a Marine Cyanobacterium Synechococcus sp. PCC 7002: Production, Characterization, and Anti-Inflammatory Properties In Vitro. Mar Drugs 2018, 16. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A. Phosphorus and the future. In The Ecophysiology of Plant-Phosphorus Interactions. Springer, Dordrecht, The Netherlands, White, P., Hammond, J., Eds. Springer: Dordrecht, 2008; pp. 271-283. [CrossRef]
Figure 1.
A schematic representation of main P pools in microalgal cells including Pi present in all cell compartments, structural phospholipids of cell membranes, phosphoproteins and phosphometabolites, nucleic acids, and the main storage form of P in the cells, polyP. For more detail on P uptake, see section 1.2.3.
Figure 1.
A schematic representation of main P pools in microalgal cells including Pi present in all cell compartments, structural phospholipids of cell membranes, phosphoproteins and phosphometabolites, nucleic acids, and the main storage form of P in the cells, polyP. For more detail on P uptake, see section 1.2.3.
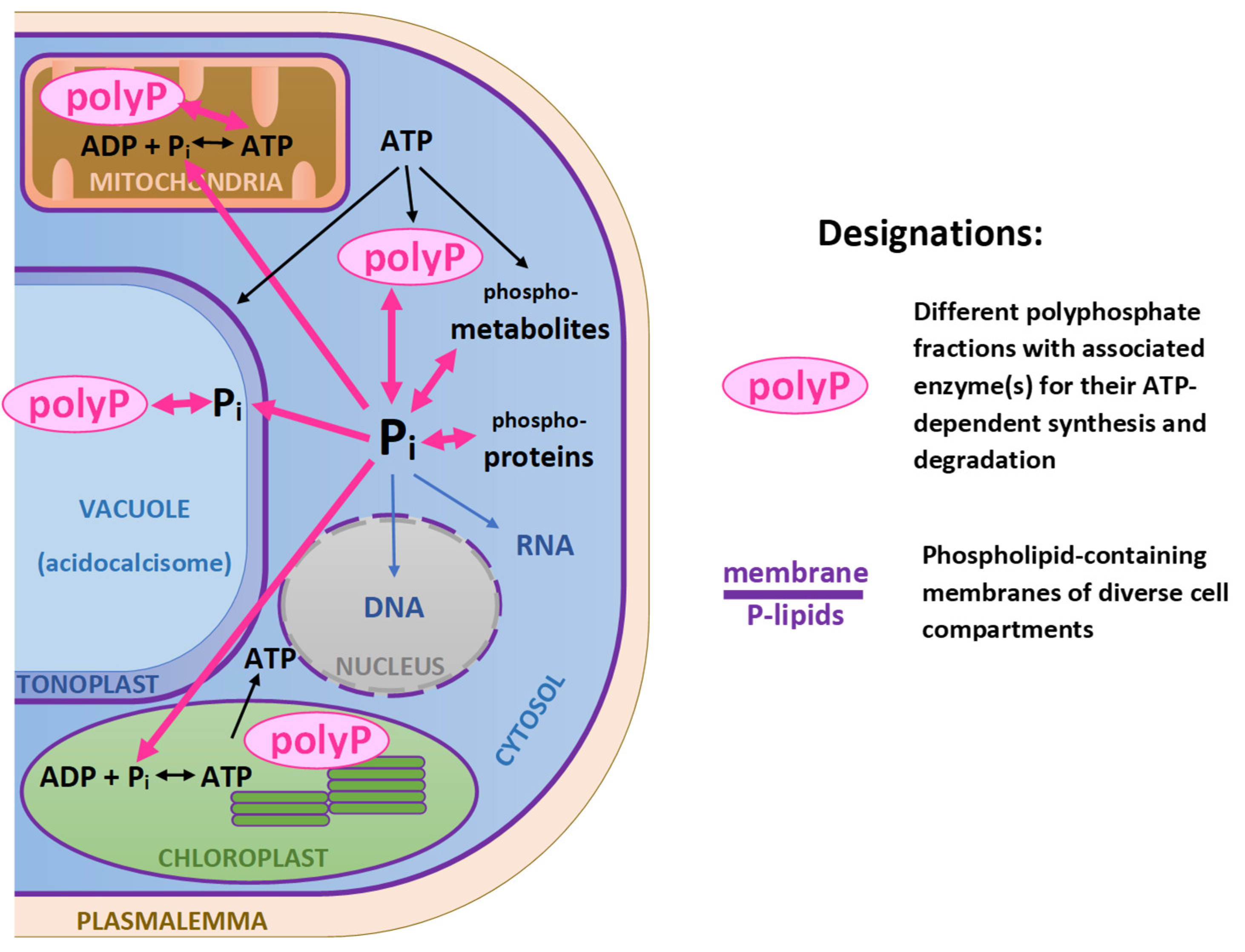
Figure 2.
Microalgae including cyanobacteria acquire P with a sophisticated system of Pi transporters operating with a high efficiency across a broad range of external Pi concentrations and environmental conditions. For details, see text and Appendix B.
Figure 2.
Microalgae including cyanobacteria acquire P with a sophisticated system of Pi transporters operating with a high efficiency across a broad range of external Pi concentrations and environmental conditions. For details, see text and Appendix B.
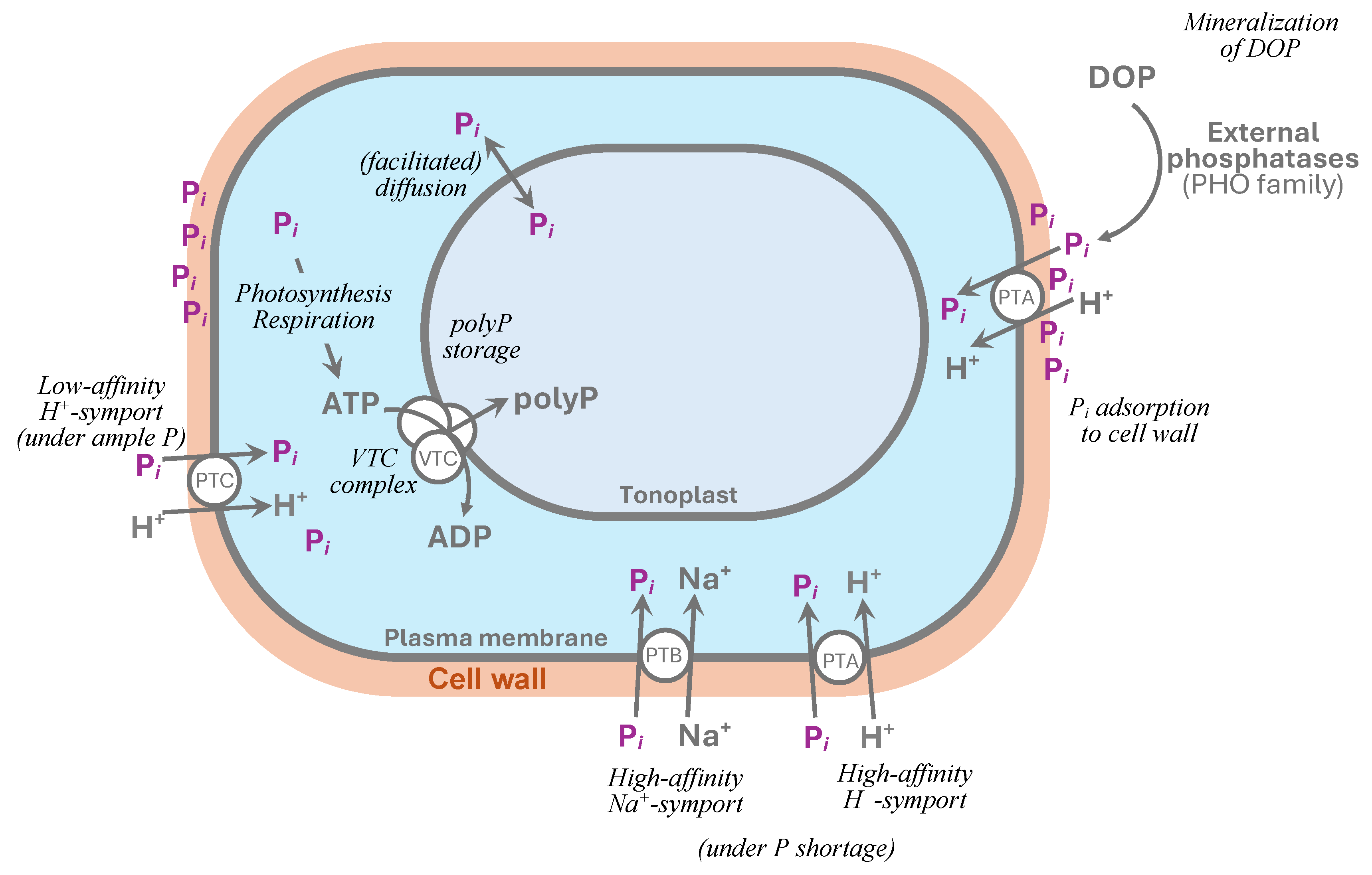

Table 1.
Ranges of P and/or Pi concentrations in pristine natural and anthropogenically impacted habitats and typical organisms.
Table 1.
Ranges of P and/or Pi concentrations in pristine natural and anthropogenically impacted habitats and typical organisms.
| Habitat or source | P or Pi range | Ref. |
|---|---|---|
| Wastewater | 3–330 mg L–1 (3 µM–3 mM) Pi | [19] |
| Domestic: 0.5 – 8.6 mg-P L-1 (0.02-0.3 mM) | [20] | |
| Industrial e.g., mine drainage: 186 – 558 mg-P L-1 (6 – 18 mM) | [21] | |
| Deep aphotic ocean | Soluble reactive P: 0.8 – 5.4 µg-P L-1 (0.025 – 0.175 µM) | [22,23] |
| River | Total P 0.04-0.4 mg-P L–1 (0.001-0.01 mM ) | [24] |
| Lakes | Soluble reactive P: 0.01-0.85 mg-P–1 (0.3 µM – 0.3 mM)) | [25] |
| Unfertilized soil | 12.4 – 341 µg-P L-1 (0.4––11 μM)* | [2] |
| Fertilized soil | 0.1–5.0 mg-P L-1 (0.003-0.16 mM) | [26] |
| Intracellular concentration | Crop plants: 155 – 620 mg-P L–1 (5–20 µM) | [27] |
| Microalgae: C. reinhardtii: 152 ± 37.0 µg free Pi (4.9±1.2 µmol mostly in the chloroplast) | [28] |
* Bioavailable for plants from the soil solution.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



