
Submitted:
22 May 2024
Posted:
22 May 2024
You are already at the latest version
Abstract
Enhanced electrical activity in detrusor smooth muscle (DSM) cells is a key factor in detrusor overactivity which causes overactive bladder pathological disorders. Transient receptor potential melastatin-4 (TRPM4) channels, calcium-activated cation channels, play a role in regulating DSM electrical activities. These channels likely contribute to depolarizing the DSM cell membrane, leading to bladder overactivity. Our research focuses on understanding TRPM4 channel function in mouse DSM cells using computational modeling. We aimed to create a detailed computational model of the TRPM4 channel based on existing electrophysiological data. We employed a modified Hodgkin-Huxley model with an incorporated TRP-like current to simulate action potential firing in response to current and synaptic stimulus inputs. Validation against experimental data showed close agreement with our simulations. Our model is the first to analyze the TRPM4 channel's role in DSM electrical activity, potentially revealing insights into bladder overactivity. In conclusion, TRPM4 channels are pivotal in regulating human DSM function and TRPM4 channel inhibitors could be promising targets for treating overactive bladder.
Keywords:
urinary incontinence
; TRPM4 ion channel
; action potential
; computational modeling
1. Introduction
People encounter diverse health issues that influence their well-being. Experiencing a non-lethal heart attack can significantly alter one's perspective on life, prompting a heightened awareness of blood pressure, dietary modifications, the adoption of exercise regimens, decreased smoking, and various lifestyle adjustments. The gravity of a health ailment might not always be obvious but can profoundly affect one's quality of life. Many of us may overlook the simplicity of typical urinary bladder function. The primary physiological roles of a healthy urinary bladder are to store urine and facilitate voluntary micturition (action of urinating) [1,2]. However, overactive bladder syndrome (OAB, also known as overactive detrusor function) presents symptoms such as urgency for urination with or without urge incontinence, frequent urination, and nocturia (waking at night to urinate) [3,4,5,6,7,8]. The impact of OAB on the individual's quality of life, healthcare system, and workforce carries considerable economic burdens [9,10,11]. The exact cause of OAB remains largely unknown. The detrusor smooth muscle (DSM) within the bladder wall plays a crucial role in bladder function [12,13]. During urine storage, DSM cells relax, allowing the bladder to hold urine for extended periods without leakage. When urination is necessary, DSM cells contract synchronously with the coordinated action of internal and external urethral sphincters [14,15]. This coordinated activity, orchestrated by the brain and spinal cord, triggers DSM contraction for micturition. In OAB, involuntary contractions of DSM cells can occur during urine storage, leading to leakage [16,17]. Lifestyle changes and pharmacological therapy are among the strategies available to manage OAB and mitigate its complications [18,19]. Pharmacological therapy devoid of side effects is crucial in pathological situations to ensure optimal patient outcomes and minimize treatment-associated risks [20]. It allows for effective management of symptoms while preserving patient safety and quality of life. Treatment options for overactive bladder (OAB) often involve medications like anticholinergics and beta-3 adrenergic agonists, designed to alleviate symptoms by calming the DSM and lessening the urge to urinate frequently [21,22]. Nonetheless, these medications can lead to side effects such as dry mouth, constipation, blurred vision, and cognitive decline with anticholinergics, as well as potential hypertension with beta-3 adrenergic agonists [23,24,25]. Therefore, understanding the DSM electrophysiology becomes essential in exploring new pharmacological targets to mitigate the side effects induced by conventional medications.
Several experimental studies have revealed that DSM cells, found in various species including humans, exhibit enhanced spontaneous phasic contractions, contributing to the development of an overactive bladder [26,27]. The initiation of spontaneous phasic contractions is triggered by the generation of evoked spontaneous action potentials (APs) and the accompanying calcium (Ca2+) dynamics [28,29,30,31]. A thorough biophysical explanation of DSM AP generation is crucial for comprehending spontaneous contractions and for exploring novel pharmacological targets for overactive bladder treatment. The electrical activities in all excitable tissues are regulated by the influx and efflux of charged ions through a network of ion channels situated at the cell membrane. In a resting state, electrically excitable cells maintain a specific resting membrane potential, which varies among different tissue types, including neuronal cells, cardiac cells, and smooth muscle cells. A transient rise in cytoplasmic calcium [Ca2+]i is an important reason behind the DSM cell contraction [32,33]. The influx of extracellular Ca2+ ions occurs mainly via voltage-dependent L-type and T-type Ca2+ channels [34,35,36]. The sarco/endoplasmic reticulum (SR) is the principal Ca2+ store participating in the initial rapid increase in [Ca2+]i by supplying Ca2+ via Ca2+ release mechanism by SR [37,38,39]. T-type Ca2+ channels are activated to depolarize the membrane potential until reaching the threshold potential, which then triggers the opening of L-type Ca2+ channels. Ca2+ influx via the L-type Ca2+ channel is essential for the rising phase of the DSM action potential, whereas various potassium (K+ ) channels mediate the repolarization and after a hyper-polarization period of the action potential, respectively [34,40,41,42]. Of the large family of mammalian K+ channels, the Ca2+ dependent K+ channel, voltage-dependent K+ channel, and delayed rectifier K+ channel are incorporated along with L-type and T-type Ca2+ channels in AP generation [41,42,43,44]. In addition, the Ca2+-dependent K+ channels in DSM cells are categorized into large (BK), intermediate (IK), and small (SK) conductance ion channels [45,46,47,48]. Several research groups have recently conducted an intriguing study revealing a novel regulatory mechanism of the transient receptor potential melastatin 4 (TRPM4) ion channel in modulating DSM cell excitability [49,50]. The TRPM4 channel has been shown to play a crucial role in regulating the resting membrane potential and basal tone of various smooth muscle cells. Through the utilization of the whole-cell voltage clamp method, researchers have successfully recorded TRPM4 channel currents in single smooth muscle cells within urinary bladder tissues from diverse animal subjects [49,51,52,53,54,55,56]. Their findings suggest that TRPM4 could serve as a novel therapeutic target for alleviating symptoms of OAB.
TRPM4 functions as a Ca2+-activated non-selective cation channel, inducing cell membrane depolarization via sodium (Na+) or K+ entry and subsequent activation of L-type Ca2+ channels (Figure 1) [54]. In one study, the application of 9-phenanthrol resulted in hyperpolarization of DSM cell membranes, indicating the first evidence of TRPM4 regulation of human DSM cell resting membrane potential [55]. Moving forward, the researchers conducted in vitro experiments, investigating the impact of TRPM4 on spontaneous and phasic contractions of human DSM isolated strips. Inhibition of TRPM4 channels with 9-phenanthrol significantly reduced spontaneous and phasic contraction amplitude, muscle force integral, contraction duration, contraction frequency, and muscle tone of the DSM, highlighting TRPM4's critical role in human DSM modulation under physiological conditions. The cartoon in Figure 1 explains the possible associated TRPM4 ion channel activation mechanism in DSM cells. Acetylcholine (Ach), a muscarinic agent activates the muscarinic receptors (M3) at the membrane [57,58,59]. Then, phospholipase C (PLC) is activated, leading to the hydrolysis of phosphatidylinositol 4,5-bisphosphate (PIP2) into inositol trisphosphate (IP3) and diacylglycerol (DAG). IP3 then binds to IP3 receptors on the sarcoplasmic reticulum (SR), causing the release of Ca2+ ions from the SR stores into the cytoplasm [53,60]. One portion of the elevated [Ca2+]i activates the TRPM4 ion channel and allows the influx of Na+ ions (or K+ ions) to depolarize the membrane. When the depolarization crosses the threshold potential, the L-type Ca2+ channel, which is a voltage-dependent calcium channel (VDCC) opens and allows an influx of extracellular Ca2+ into the cytoplasm. The cytoplasmic Ca2+ due to release from the SR store and via L-type Ca2+ channel triggers contraction after the generation of the action potential.
Unfortunately, the quantitative description of the modulatory effects of the TRPM4 ion channel on DSM cell electrical properties has not been thoroughly explored due to the highly complex nature of the experimental procedures involved in DSM cell electrophysiology. Unlike cardiac and neuronal electrophysiological studies, obtaining comprehensive electrical recordings from smooth muscle cells is challenging. Consequently, most research focuses on the TRPM4 channel's impact on DSM contraction rather than its effect on passive or active electrical properties, like spike/AP modulation. Understanding the biophysical parameters governing DSM cell electrophysiology is crucial for drug design accuracy. Additionally, there's debate over variations in inward and outward current in DSM electrophysiology. The redundancy of ion channel types regulates cellular functions and maintains physiological balance, ensuring resilience against perturbations [61]. However, investigations of the redundancy and resilience of TRPM4 concerning other inward current ion channels in DSM cells remain unexplored.
Over the past few decades, computational and mathematical modeling techniques have enhanced our comprehension of intricate biological processes. By simulating diverse scenarios and predicting outcomes, these methods offer insights that may be challenging to attain through traditional experimental approaches [62]. The primary goal of computational biophysical modeling is to provide insights into the structure, function, and dynamics of biological systems that may be difficult or impossible to obtain through experimental methods alone. In the realm of smooth muscle electrophysiology, computational simulations have been instrumental in exploring cellular biophysics and modulating cellular electrical activities. These models shed light on ion channel function, encompassing aspects such as conductance, ion selectivity, and channel opening, which hold significance in pathological conditions. Moreover, biophysical modeling directly simulates ion flux through membrane channels, facilitating a deeper understanding of their behavior and their involvement in disease processes. While computational models for various types of smooth muscle cells, such as intestinal [63], uterine [64,65,66], ureter [67,68,69], jejunal [70], vas deferens [71,72,73,74], gastric [75,76], mesenteric [77], small bowel [78], urethra [79,80], and arterial [81] smooth muscle cells, have been developed, models for DSM cells are relatively underdeveloped. The models published on DSM electrophysiology have yet to explore the modulatory properties of TRPM4 ion channels in DSP action potentials [82,83,84,85,86,87,88]. The present in silico model aims to elucidate the biophysical mechanisms underlying the DSM electrophysiology, aiming to investigate the impact of the TRPM4 ion channel on the firing rate of DSM action potentials. Our objectives in developing this model are threefold. Firstly, leveraging experimental data, we will simulate and validate the kinetics of TRPM4 ion channels in response to changes in intracellular Ca2+ concentration. Secondly, we will integrate these ion channels into a single-compartmental biophysical DSM model to simulate APs and explore alterations in AP parameters associated with TRPM4 ion channel activation. Lastly, we seek to derive novel biological insights to corroborate existing hypotheses drawn from various experiments and propose new hypotheses for future research endeavors.
2. Materials and Methods
2.1. Model Adaptation
Biophysically detailed cell modeling for neuronal cells made its debut in 1952 in neuroscience, thanks to the groundbreaking work of Hodgkin and Huxley on the squid giant axon [89]. In the 1960s, Noble pioneered the first models of cardiac cellular activity [90]. Since then, an impressive array of mathematical models for neuronal cells, cardiac muscle, and smooth muscle electrophysiology has emerged. The complexity of these models has steadily increased over the years as more experimental data has become available. Several DSM cell electrophysiology models have been mathematically designed to mimic the behavior of DSM electrical activities [82,83,84,86,87]. These models also serve as a bridge between cellular-level models and tissue-level function, offering a comprehensive understanding of DSM cell electrical activity across different scales. For establishing a single compartmental DSM AP model to integrate the TRPM4 ion channel, we have adapted the model [83] for simulating DSM cell electrical properties. The DSM cell membrane's cylindrical single-cell morphology and passive electrical properties are derived from the experimental data [91]. The length, diameter, membrane resistivity, cytoplasmic resistivity, and membrane capacitance are 200 μm, 6 μm, 138 kΩ.cm2, 183 Ω.cm, and 1 μF/cm2 respectively.
2.2. General Membrane Current Descriptions
The DSM cell is electrically defined by a resistor-capacitor (RC) circuit, where the membrane capacitance Cm is parallel with the variable ion channel conductance gion. All active ion channel conductances gion are associated with respective Nernst potential Eion in series. Figure 2. illustrates the schematic overview of the parallel conductance model for ionic current (Iion), and it shows the flow of ion ‘X+’ under the influence of an electrochemical driving force.
The calculation of the individual ionic current follows the principles of Ohm's law, traditionally expressed through Equation 1.
In equation (1), and ENerrnst represents the maximum channel conductance and Nernst potential specific to the ion channel under consideration. The variable parameters m and h correspond to dimensionless activation and inactivation gating variables, respectively, which are dependent on time, voltage, and Ca2+ concentration. To facilitate equation fitting, an additional pair of dimensionless parameters denoted as 'x' and 'y' are introduced. determined through a system of first-order differential equations, following the classical Hodgkin-Huxley (HH) formulation mechanisms [89]
For example, equation 2 computes the instantaneous value of the activation variable “m” in our DSM cell model.
In this equation, m∞ represents the steady-state value, and τm denotes the time constant, both of which are functions of voltage and/or ionic concentrations.
In this context, the relationship between the state parameter and the membrane potential (Vm) for ion channels is elucidated through the Boltzmann equation.
Where V1/2 is the half activation potential and S is the slope factor.
The large conductance calcium-activated potassium (BK) channel kinetics have been elucidated through a multi-state Markov model (MM) [92], which offers a detailed representation of the channel's sensitivity to Ca2+, enhancing precision. The MM model enhances the accuracy of the description of a multi-agent activated ion channel by incorporating multiple closed and open states to represent its internal conditions. This model comprises closed states denoted as C0, C1, C2, C3, and C4, and five corresponding open states denoted as O0, O1, O2, O3, and O4. Among these, the open state O4 facilitates the passage of K+ ions via BK channels driven by the instantaneous electrochemical driving force (EDF). The BK current (IBK) is determined using the following equation.
where is the maximum conductance and O is the summation of O1, O2, O3, and O4.
Common rate equations:
Kon=335,Kcoff=26,Kooff=26, O=O1+O2+O3+O4
Rate equations for voltage-dependent transitions:
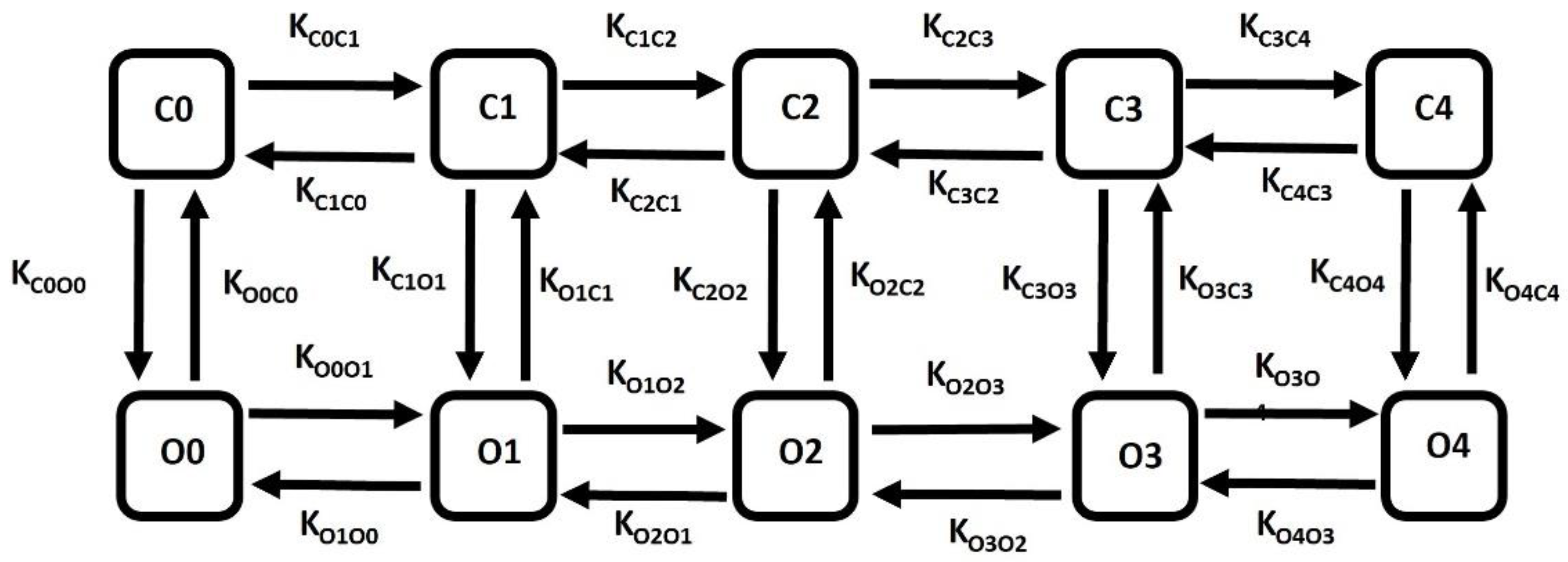
KC0O0 = 0.03162 * a, KC1O1 = 0.000969 * a, KC2O2 = 0.0000381 * a,
KC3O3 = 0.000881 * a, KC4O4 = 0.054324 * a, KO0C0 = 328.1084 * b,
KO1C1 = 154. 1736 * b, KO2C2 = 33.6594 * b, KO3C3= 0.097312 *b,KO4C4=0.000406*b*cai
KC3O3 = 0.000881 * a, KC4O4 = 0.054324 * a, KO0C0 = 328.1084 * b,
KO1C1 = 154. 1736 * b, KO2C2 = 33.6594 * b, KO3C3= 0.097312 *b,KO4C4=0.000406*b*cai
Figure 3.
Schematic diagram of 10-state Markov model for BK channel. A detailed explanation is provided in the text.
Figure 3.
Schematic diagram of 10-state Markov model for BK channel. A detailed explanation is provided in the text.
State equations for calcium (cai) dependent transitions:
KC0C1= 3 * Kon * cai, KC1C2= 4* Kon * cai, KC2C3= 3 * Kon * cai, KC3C4= Kon * cai
KC4C3= 3 * Kcoff * cai, KC3C2= 4 * Kcoff * cai, KC2C1= 3 * Kcoff * cai, KC1C0= Kcoff * cai
KO0O1= 3 * Kon * cai, KO1O2= 4 * Kon * cai, KO2O3= 3 * Kon * cai, KO3O4= Kon * cai
KO4O3= 3 * Kooff * cai, KO3O2= 4 * Kooff * cai, KO2O1= 3 * Kooff * cai, KO1O0 = Kooff * cai
KC4C3= 3 * Kcoff * cai, KC3C2= 4 * Kcoff * cai, KC2C1= 3 * Kcoff * cai, KC1C0= Kcoff * cai
KO0O1= 3 * Kon * cai, KO1O2= 4 * Kon * cai, KO2O3= 3 * Kon * cai, KO3O4= Kon * cai
KO4O3= 3 * Kooff * cai, KO3O2= 4 * Kooff * cai, KO2O1= 3 * Kooff * cai, KO1O0 = Kooff * cai
In equations 5, 6, and 7, the parameter values are defined to obtain the most accurate model of the BK ion channel, aiming to simulate the current-voltage relationship curve and BK current under different membrane potentials and internal Ca2+ concentration.
2.3. TRPM 4 Channel with Ca2+ Sensing Mechanism
Here the mathematical interpretation of TRPM4 ion channel is based on conventional Hodgkin-Huxley formalism. In this model, Hodgkin-Huxley formalism is adapted for INa ionic currents in equation (8):
where is maximum conductance, Erev is Nernst potential for sodium ion, m is intracellular calcium concentration-dependent activation variable and x is the power for gating variable.
The first-order differential equation is used in equation (9) to describe the time and calcium-dependent nature of gating variable m.
Where m∞ and τm are defined as steady-state value and time constant of gating variable ‘m’ respectively.
Here the activation steady state is expressed by the Hills equation.
Where Cah is half inhibition intracellular calcium ion centration, Cai is free calcium concentration and n is the Hill coefficient. All modeling parameters are borrowed from literature based on experimental studies. Cah and n are fitted to match with experimental result.
2.4. Whole DSM Cell Model and Simulation
Figure 4 shows the list of active ion channels incorporated in our DSM cell. In our model, two types of voltage-gated Ca2+ channels are T-type (ICaT) and L-type (ICaL) Ca2+ channels. Voltage-gated K+ channels are inward rectifying channels (IKIR), delayed rectifier channels (IKDR), and fast (IKv1) channels. Ca2+-activated K+ channels are large conductance (IBKCa), intermediate conductance (IIKCa), and small conductance (ISKCa) channels. The TRPM4 channel (ITRPM4) and leakage channels (Il) are also added in this single compartmental DSM cell model.
The synaptic input defined by the alpha function [93] or a brief square pulse of injected current (variable duration and magnitude) serves as the external stimulus to generate action potentials in the whole-cell model. Subsequently, the conductance is adjusted to explore the modified response in action potentials and resting membrane potential. Simulations were conducted with a consistent time step of 0.04 ms using the Euler Method on a PC equipped with an Intel Core i7 CPU operating at 3.80 GHz and a dual-core setup. The NEURON simulation environment was utilized for model creation, renowned for its accurate portrayal of excitable cells [94]. While NEURON primarily employs implicit integration techniques like backward Euler and a variant of Crank-Nicolson for stability, it also supports Euler's method due to its efficiency and minimal memory usage [95]. We have also adapted NEURON ‘s multiple run fitter optimization algorithms to optimize our running fitness for action potential generation. This process usually entails scrutinizing different aspects of our running performance and leveraging the gathered data to refine our training routine, aiming for improved outcomes.
Following model creation, stability, and consistency were evaluated by adjusting the maximum conductance (gmax) of ionic conductances within +/- 30% of the default value. Results demonstrated stable action potentials, exhibiting anticipated responses to changes in conductance; for instance, increased gmax led to higher AP peak amplitudes while preserving the AP's normal characteristics.
Goodness-of-fit (GoF) involves assessing how well the observed data aligns with the data predicted by a model, typically through a fit statistic or measure of deviation, such as residuals, Chi-square, or deviance. The standard error of the regression (S) or root mean squared error (RMSE) serves as a measure of GoF in our analyses of simulated action potentials, chosen due to the inadequacy of R2 for nonlinear comparisons [96]. S is computed using the following formula:
In the equation 11, Yexpt represents the experimental value, YSim denotes the corresponding simulated fit value, N stands for the number of parameters utilized in the fitting equation, and M represents the total number of data points. A smaller S value indicates lower average errors and indicates a better fit. We established a threshold for a satisfactory model as 5% of the difference between the maximum and minimum values observed in the experimental data. Any S value below this threshold is deemed indicative of a good fit.
The model code will be accessible on GitHub and the repository (https://modeldb.science, accessed on 05/05/2024) for open-source code sharing.
3. Results
The TRPM4 channel's current behavior is modeled and simulated based on equations 8, 9, and 10, following the principles of the Hodgkin-Huxley (HH) formalism, which incorporates activation and inactivation parameters. The equilibrium potential of TRPM4, denoted as ENa, is set at -40mV. Activation parameters are computed across varying intracellular Ca2+ concentrations (Cai), with a baseline Cai concentration of 0.1 mM during rest. In Figure 5, we present the steady-state activation curves for the TRPM4 channel, comparing our model's simulated curve (depicted by the red solid line) with experimental data (shown as filled triangles) sourced from Demion et al. (2007) [97]. The steady-state activation curve is unit less and ranges between 0 and 1. The X-axis is the logarithmic value of the Cai value. The comparison demonstrates a close alignment between our simulated curve and the experimental data. Notably, higher concentrations of Cai correspond to increased steady-state activation values, indicating an augmented TRPM4 current.
Subsequently, we integrated the TRPM4 channel into a single-compartmental DSM model, as outlined by [83]. Earlier sections of this paper have detailed the inclusion of various ion channels within the DSM model. Before introducing the TRPM4 channel, we meticulously evaluated the stability, robustness, and flexibility of the DSM cell model, adhering to the methodologies outlined in the methods section. Minor adjustments were made to certain parameters (particularly with ion channel conductances and time constant values) to ensure the stability of the resting membrane potential. The resting intracellular calcium concentration is maintained at 150 nM. Table 1 provides a comprehensive list of all ion channels incorporated in the model, along with their respective maximum conductances necessary for maintaining the stable resting membrane potential.
Incorporating all ion channel models, including TRPM4, our primary aim was to uphold a physiological resting membrane potential (RMP) of -52 mV. The model's robustness was confirmed by maintaining the RMP at -52 mV for 2000 ms, as illustrated in Figure 6. However, at the outset (0 ms), the model experienced a brief period of instability due to the behavior of all ion channels, resulting in slight fluctuations in membrane potential (Figure 6A). To address this, the time scale (x-axis) in Figure 6B begins from 500 ms onwards, excluding these initial fluctuations.
We administered current stimuli with varying amplitudes lasting 10 ms to investigate evoked depolarization, action potential (AP) generation and threshold estimation. No spikes occurred until the stimulus reached 0.56 mA, at which point the AP was triggered (Figure 7A). Analysis of the AP (depicted by the red solid line) and depolarization (represented by the red dashed line) facilitated the prediction of the threshold required to initiate the AP, estimated at -38.56 mV. Subsequently, we introduced synaptic input with different amplitudes to study evoked depolarization, AP generation, and threshold prediction. No spikes were observed with a stimulus of 0.0078 µS, while an AP was elicited with a stimulus of 0.0079 µS. Analysis of the simulated AP (depicted by the red solid line) and depolarization (shown as the red dashed line) predicted the threshold for AP initiation at -38.42 mV. Experimental data on AP in mouse DSM cells under synaptic stimulus, as reported by [83], were compared with our model-simulated AP. The extracted data from this experimental AP (illustrated by the blue dashed line) exhibited a close match with our model's output, corroborating the accuracy of our model.
Our subsequent investigation aimed to explore the modulatory impact of the TRPM4 channel on the electrical characteristics of the DSM cell. It is important to highlight one of the most intriguing aspects of DSM cell action potential generation, namely, the variability in action potential shapes. Unlike neuronal and cardiac cells, which typically exhibit uniform action potentials for a given cell type, isolated DSM cells can produce various types of action potentials. These variations stem from differences in action potential duration, the shape of depolarizing and repolarizing phases, after-depolarization potential, and after-hyperpolarizations. Figure 8 depicts one type of DSM action potential following a 10% and 20% increase in the maximum conductance of the TRPM4 ion channel. The action potential represented by the red solid line arises after a 10% increase in the maximum conductance of the TRPM4 ion channel. Notably, the resting membrane potential becomes more positive following action potential generation. However, this generated action potential aligns with physiological expectations. In contrast, the action potential depicted by the black solid line emerges after a 20% increase in the maximum conductance of the TRPM4 ion channel. Although an action potential is generated, this particular type of action potential does not match physiological norms. Elevated conductance of the TRPM4 channel induces model instability.
To conduct a more comprehensive quantitative examination, we performed sensitivity analysis on the TRPM4 conductance concerning action potential parameters. Figure 9 illustrates the alterations in action potential parameters, specifically the resting membrane potential (red solid line) and action potential duration (dashed doted red line), in response to variations in the maximum conductance of the TRPM4 channel. All parameters are normalized for enhanced clarity. It's noteworthy that each action potential parameter fluctuates within a range of up to +20% and -30%, indicating the stability of the action potential across the range of TRPM4 channel conductance.
The prevalence of ion channels in excitable cells (such as neurons, cardiac cells, and muscle cells) prompts fundamental inquiries into how the unique intrinsic characteristics of individual cells arise from their specific complement of channels. Across all excitable cells, many ion channels exhibit overlapping voltage and time-dependent traits. We propose that these shared properties contribute to the robustness of physiological function. Despite displaying consistent and similar behaviors, individual excitable cells of the same type demonstrate variability in ion channel conductance densities [61,98]. This complexity complicates the direct function assignment to any particular conductance and is associated with the diverse responses of similar cells to disturbances, deletions, and pharmacological treatments. Several experimental investigations have shown that the activation of the T-type Ca2+ channel depolarizes the DSM cell to the threshold of the L-type Ca2+ channel, for which the cell generates AP [99]. To examine our hypothesis concerning a mutation in the T-type Ca2+ channel (whereby the maximum conductance of the T-type Ca2+ channel is set to zero), we adjusted the conductance of the TRPM4 channel to maintain the electrophysiological behavior of the DSM cell. As illustrated in Figure 10, when the T-type Ca2+ channel is muted (depicted by the red dashed line), the DSM cell fails to generate any action potential. Further adjustments to the parameters of the TRPM4 channel render the DSM cell incapable of generating action potentials. However, through fine-tuning the parameters of the TRPM4, L-type Ca2+ channel, and inward rectifying channel, the DSM cell is able to generate action potentials without the presence of the T-type Ca2+ channel (depicted by the red solid line). Initially, the model exhibited stochastic behaviors, but subsequent adjustments to other ion channel parameters enabled the generation of action potentials. This finding strongly supports the variability and resilience of our computational model. It should be noted that other ion channel blockers were applied in the DSM models, and their effects on AP were shown in [83].
4. Discussion
OAB syndrome has a profound impact on people's lives and poses significant financial challenges. This condition, characterized by sudden and uncontrollable urges to urinate, often results in frequent bathroom visits, disrupted sleep, and limitations in daily activities. The anxiety and embarrassment surrounding potential accidents can lead to social isolation and reduced productivity. Additionally, managing OAB requires ongoing medical expenses, such as doctor visits, medications, and possibly surgeries, placing strain on both personal finances and healthcare systems. Despite its widespread occurrence and consequences, much remains unknown about the underlying causes and effective treatments for OAB. Fundamental research plays a crucial role in unraveling the complexities of this condition, including understanding the neural pathways involved in bladder control, identifying early detection biomarkers, and developing targeted therapies with minimal side effects. DSM contraction is closely linked to OAB syndrome [16,17], as heightened activity in these muscles can lead to involuntary and frequent bladder contractions, resulting in the characteristic urgency and urge incontinence experienced by individuals with OAB. A thorough exploration of the underlying biophysical mechanisms behind DSM electrophysiology is crucial for identifying new pharmacological targets with minimal side effects. As a mechanical event, the contraction is initiated by intracellular calcium elevation and the generation of action potential. Ion channels play a crucial role across the DSM cell membrane in modulating the underlying electrical properties, including the action potential. Hence, accurately quantifying the individual ion channel's contribution to modulating the action potential will aid in exploring more effective drugs for overactive bladder treatment. In recent years, multiple experimental findings have indicated the presence of the TRPM4 ion channel in DSM cells and its role in modulating DSM contraction [49,50]. Due to the highly complex nature of experimental smooth muscle electrophysiology setups, the quantified contribution of the TRPM4 channel to DSM cell membrane potential and action potential has not yet been studied.
Computational and in silico approaches have revolutionized the field of electrophysiology by providing valuable insights into the behavior of ion channels at a fundamental level. These methods simulate the complex interactions of ion channels with high precision, allowing researchers to explore diverse scenarios and predict ion channel behavior under different conditions. By complementing experimental studies, computational models enhance our understanding of ion channel function, kinetics, and pharmacology. Moreover, they offer a platform for hypothesis testing and guide experimental design, ultimately accelerating the discovery of novel targets and therapies for various physiological and pathological conditions. While numerous computational models have been published regarding DSM cell electrophysiology, none of them have incorporated the TRPM4 channel for investigation [82,83,84,86,87]. Computational modeling is an ongoing process, wherein established models are continuously refined with new experimental findings to yield more fruitful research outcomes. To the best of our knowledge, the model presented is the first biophysically constrained DSM cell model to investigate the contribution of the TRPM4 channel to DSM cell excitability.
Our first objective in developing this model was to employ differential equations, Hodgkin-Huxley formalisms, and parameters to accurately replicate the internal kinetics of the TRPM4 channel. We have depicted the simulated steady-state activation curve across various calcium concentrations (Figure 5). Validation against experimental data reinforces the heightened accuracy of our TRPM4 ion channel model, confirming the successful achievement of our first objective. Our second objective was to integrate the TRPM4 channel into a DSM cell model and examine its modulating effects on cellular excitability. Before integrating the new TRPM4 channel, we validated the previous model to ensure its robustness and reproducibility. Once we confirmed the proper functioning of the model, we proceeded with the integration of the TRPM4 channel. However, the addition of a foreign element to an established model often results in stochastic aberrant behavior. Similarly, our model exhibited instability after the incorporation of the TRPM4 channel. To address this, we carefully adjusted the maximum conductances of ion channels to stabilize the model and replicate previously simulated outputs. We validated the simulated synaptic potentials and action potentials by comparing them with experimental data, confirming our model's ability to replicate experimental findings [83] (Figure 7). Subsequently, we adjusted the maximum conductance of the TRPM4 channel within physiological ranges to assess its impact on the resting membrane potential, action potential threshold, and peak amplitude. Activation of the TRPM4 channel leads to a shift in the resting membrane potential towards a more positive state, reduces the threshold potential for action potential initiation, and increases the peak amplitude of the action potential. This suggests that overexpression of TRPM4 channels across the DSM cell membrane could induce heightened spontaneous contractions, a primary symptom of overactive bladder [49,50]. Therefore, controlled doses of TRPM4 channel blockers may mitigate this pathological condition. Conversely, in the absence of overexpressed channels, their inherent activity may still lead to overactivity due to spontaneous intracellular calcium elevation. Experimental studies have demonstrated transient increases in intracellular calcium, such as calcium sparks, puffs, and waves [100], which could activate TRPM4 channels locally, exacerbating overactive bladder symptoms. The term "localization" distinguishes TRPM4 channel activation from calcium-dependent potassium channel activation, which hyperpolarizes the membrane, reducing cellular excitability. To further validate our model, we conducted a sensitivity analysis, providing additional quantitative insights into how the TRPM4 channel influences cellular electrical properties. As a primary goal of any computational physiology model is to generate new hypotheses, our third objective was to propose novel insights regarding the TRPM4 channel in detrusor muscle biophysics. In the introduction section, we briefly discussed the redundancy of ion channel types to ensure resilience against perturbations. Building on this concept, we inactivated the T-type calcium channel by reducing its maximum conductance to zero, then simulated all electrical properties by adjusting conductances and other parameters for the remaining ion channels. This revealed that the TRPM4 channel could compensate for the absence of the T-type calcium channel. Our hypothesis suggests that the TRPM4 channel may serve as a standby ion channel to maintain bladder physiology in cases of dysfunction or mutation in the T-type calcium channel.
Computational models inherently have several limitations. Firstly, our current model investigated the modulating effects of TRPM4 channels on spike-type action potentials in the DSM model. However, DSM cells also generate pacemaking-type action potentials, which are not simulated in this model. To date, no biophysically detailed DSM model has been developed to simulate pacemaking action potentials. Additionally, most of the experimental research papers highlighted the spike-type action potentials in single isolated DSM cells [82,83]. Although the action potential was validated against experimental data from mouse urinary bladder, our ion channel parameters are derived from other animals (mice and guinea pigs) and organs due to a lack of specific data [82,83]. Additionally, while actual smooth muscle morphology may not perfectly align with a cylindrical shape, our model is designed based on this simplified geometry. Furthermore, DSM cells function as a syncytium, where multiple cells are interconnected via gap junctions, enabling electrical signal propagation. However, our model is limited to a single-compartment isolated cell. Future model expansions could involve investigating the effects of TRPM4 channel modulation in multicompartmental tissues with complex calcium dynamics.
Author Contributions
C.M conceived the idea of draft, wrote manuscript. R.T also help in writing, editing and formatting the final draft. C.M also responded to reviewers’ comments.
Funding
This research received no external funding.
Institutional Review Board Statement
“Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Chitaranjan Mahapatra is thankful to UCSF, San Francisco for providing space and other required facilities for the completion of this manuscript. RK is thankful to UC Berkeley for providing facilities and space for the completion of the manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- de Groat, William C., and N. A. O. K. I. Yoshimura. "Anatomy and physiology of the lower urinary tract." Handbook of clinical neurology 130 (2015): 61-108. [CrossRef]
- Tyagi, Shachi, Pradeep Tyagi, Naoki Yoshimura, and Michael B. Chancellor. "Physiology of micturition." In Textbook of Female Urology and Urogynecology-Two-Volume Set, pp. 232-246. CRC Press, 2017.
- Abrams, Paul. "Describing bladder storage function: overactive bladder syndrome and detrusor overactivity." Urology 62, no. 5 (2003): 28-37. [CrossRef]
- Wein, Alan J., and Raymond R. Rackley. "Overactive bladder: a better understanding of pathophysiology, diagnosis, and management." The Journal of urology 175, no. 3 (2006): S5-S10. [CrossRef]
- Drake, Marcus J. "The Overactive Bladder." In Textbook of Female Urology and Urogynecology-Two-Volume Set, pp. 584-593. CRC Press, 2017.
- Chapple, Christopher R., and Altaf Mangera. "Urgency incontinence and overactive bladder." Oxford Textbook of Urological Surgery (2017): 282.
- Dwyer, Peter L., and Anna Rosamilia. "Evaluation and diagnosis of the overactive bladder." Clinical obstetrics and gynecology 45, no. 1 (2002): 193-204. [CrossRef]
- Brown, Jeanette S., William F. McGhan, and Sudhansu Chokroverty. "Comorbidities associated with overactive bladder." Am J Manag Care 6, no. 11 Suppl (2000): S574-9.
- Palma-Zamora, Isaac D., and Humphrey O. Atiemo. "Understanding the economic impact of neurogenic lower urinary tract dysfunction." Urologic Clinics 44, no. 3 (2017): 333-343. [CrossRef]
- Lin, Kuan-Yin, Ka-Chun Siu, and Kuan-Han Lin. "Impact of lower urinary tract symptoms on work productivity in female workers: A systematic review and meta-analysis." Neurourology and Urodynamics 37, no. 8 (2018): 2323-2334. [CrossRef]
- Pierce, Heather, Lin Perry, Pauline Chiarelli, and Robyn Gallagher. "A systematic review of prevalence and impact of symptoms of pelvic floor dysfunction in identified workforce groups." Journal of Advanced nursing 72, no. 8 (2016): 1718-1734. [CrossRef]
- Chakrabarty, Basu, J. Crook, Marcus Drake, Niall Gilliland, Dev Gulur, Darryl Kitney, Aditya Manjunath, Pavlo Somov, and Bahareh Vahabi. "The Urinary Tract: Form and Function." Lower Urinary Tract Symptoms in Adults: A Clinical Approach (2020): 1-17.
- Kitney, Darryl Graham. Bladder spontaneous activity: influence of mild heating and inert injectables. University of Surrey (United Kingdom), 2016.
- Bortolini, Maria Augusta T., Andreisa PM Bilhar, and Rodrigo A. Castro. "Neural control of lower urinary tract and targets for pharmacological therapy." International urogynecology journal 25 (2014): 1453-1462. [CrossRef]
- Afrashteh, Behnaz. "Identification of lower urinary tract dysfunction after spinal cord injury at lumbosacral level by developing a clinical relevance rat model." PhD diss., University of Salzburg, 2023.
- Asl, Seyed Omid Komari Zadeh. Analysis and modeling of the roles of actin-myosin interactions in bladder smooth muscle biomechanics. Virginia Commonwealth University, 2014.
- Guntu, Vinay Sandeep Kumar. "Biophysical modeling to reverse engineer two mammalian neural circuits lower urinar Y tract and hippocampus." PhD diss., University of Missouri--Columbia, 2020.
- Wyman, J. F., K. L. Burgio, and D. K. Newman. "Practical aspects of lifestyle modifications and behavioural interventions in the treatment of overactive bladder and urgency urinary incontinence." International journal of clinical practice 63, no. 8 (2009): 1177-1191. [CrossRef]
- Willis-Gray, Marcella G., Alexis A. Dieter, and Elizabeth J. Geller. "Evaluation and management of overactive bladder: strategies for optimizing care." Research and reports in urology (2016): 113-122. [CrossRef]
- Smith, Louise E., Rebecca K. Webster, and G. James Rubin. "A systematic review of factors associated with side-effect expectations from medical interventions." Health Expectations 23, no. 4 (2020): 731-758. [CrossRef]
- Mostafaei, Hadi, Shahrokh F. Shariat, Hanieh Salehi-Pourmehr, Florian Janisch, Keiichiro Mori, Fahad Quhal, and Sakineh Hajebrahimi. "The clinical pharmacology of the medical treatment for overactive bladder in adults." Expert review of clinical pharmacology 13, no. 7 (2020): 707-720. [CrossRef]
- Hutchinson, Alexander, Alexander Nesbitt, Andre Joshi, Adrian Clubb, and Marlon Perera. "Overactive bladder syndrome: management and treatment options." Australian journal of general practice 49, no. 9 (2020): 593-598. [CrossRef]
- Fogaing, Cora, Abubakr H. Mossa, and Lysanne Campeau. "Are beta 3 adrenergic agonists now the preferred pharmacologic management of overactive bladder?." Current urology reports 21 (2020): 1-12. [CrossRef]
- Duong, Vi, Aya Iwamoto, Jon Pennycuff, Bela Kudish, and Cheryl Iglesia. "A systematic review of neurocognitive dysfunction with overactive bladder medications." International Urogynecology Journal 32, no. 10 (2021): 2693-2702. [CrossRef]
- Goodridge, Sophia Delpe, and Leslie M. Rickey. "Medical Therapy with Antimuscarinics and ß3-Agonists." In Female Urinary Incontinence, pp. 147-164. Cham: Springer International Publishing, 2022.
- Drake, Marcus J., Christopher H. Fry, Hikaru Hashitani, Ruth Kirschner-Hermanns, Mohammad S. Rahnama'i, John E. Speich, Hikaru Tomoe, Anthony J. Kanai, and Karen D. McCloskey. "What are the origins and relevance of spontaneous bladder contractions? ICI-RS 2017." Neurourology and urodynamics 37, no. S4 (2018): S13-S19. [CrossRef]
- Mitsui, Retsu, Ken Lee, Aoi Uchiyama, Shunta Hayakawa, Fumio Kinoshita, Shunichi Kajioka, Masatoshi Eto, and Hikaru Hashitani. "Contractile elements and their sympathetic regulations in the pig urinary bladder: a species and regional comparative study." Cell and tissue research 379 (2020): 373-387. [CrossRef]
- Burdyga, T. V., and Susan Wray. "The relationship between the action potential, intracellular calcium and force in intact phasic, guinea-pig uretic smooth muscle." The Journal of Physiology 520, no. 3 (1999): 867-883. [CrossRef]
- Ter Keurs, Henk EDJ, and Penelope A. Boyden. "Calcium and arrhythmogenesis." Physiological reviews 87, no. 2 (2007): 457-506. [CrossRef]
- Bolton, T. B., D. V. Gordienko, V. Pucovský, S. Parsons, and O. Povstyan. "Calcium release events in excitation–contraction coupling in smooth muscle." In Role Of The Sarcoplasmic Reticulum In Smooth Muscle: Novartis Foundation Symposium 246, vol. 246, pp. 154-173. Chichester, UK: John Wiley & Sons, Ltd, 2002.
- Sanders, Kenton M. "Spontaneous electrical activity and rhythmicity in gastrointestinal smooth muscles." Smooth muscle spontaneous activity: Physiological and pathological modulation (2019): 3-46. [CrossRef]
- Boopathi, Ettickan, Cristiano Gomes, Stephen A. Zderic, Bruce Malkowicz, Ranjita Chakrabarti, Darshan P. Patel, Alan J. Wein, and Samuel Chacko. "Mechanical stretch upregulates proteins involved in Ca2+ sensitization in urinary bladder smooth muscle hypertrophy." American Journal of Physiology-Cell Physiology 307, no. 6 (2014): C542-C553. [CrossRef]
- Poley, Rainer N., Christopher R. Dosier, John E. Speich, Amy S. Miner, and Paul H. Ratz. "Stimulated calcium entry and constitutive RhoA kinase activity cause stretch-induced detrusor contraction." European journal of pharmacology 599, no. 1-3 (2008): 137-145. [CrossRef]
- Kajioka, Shunichi, Shinsuke Nakayama, Gordon McMurray, Kihachiro Abe, and Alison F. Brading. "Ca2+ channel properties in smooth muscle cells of the urinary bladder from pig and human." European journal of pharmacology 443, no. 1-3 (2002): 19-29. [CrossRef]
- Wu, C., G. Sui, and C. H. Fry. "The role of the L-type Ca2+ channel in refilling functional intracellular Ca2+ stores in guinea-pig detrusor smooth muscle." The Journal of Physiology 538, no. 2 (2002): 357-369. [CrossRef]
- Petkov, Georgi V. "14 Role of Ion Channels in Urinary Bladder Smooth Muscle Function." Signal Transduction and Smooth Muscle (2018): 281. [CrossRef]
- Wray, Susan, and Theodor Burdyga. "Sarcoplasmic reticulum function in smooth muscle." Physiological reviews 90, no. 1 (2010): 113-178. [CrossRef]
- Wu, C., and C. H. Fry. "Na+/Ca2+ exchange and its role in intracellular Ca2+ regulation in guinea pig detrusor smooth muscle." American Journal of Physiology-Cell Physiology 280, no. 5 (2001): C1090-C1096. [CrossRef]
- Herrera, Gerald M., and Mark T. Nelson. "Sarcoplasmic reticulum and membrane currents." In Role Of The Sarcoplasmic Reticulum In Smooth Muscle: Novartis Foundation Symposium 246, vol. 246, pp. 189-207. Chichester, UK: John Wiley & Sons, Ltd, 2002.
- Provence, Aaron. "Kv7 Channels Of The Urinary Bladder Smooth Muscle: Functional Roles And Therapeutic Potential." (2018).
- Takagi, Hiroaki, and Hikaru Hashitani. "Effects of K+ channel openers on spontaneous action potentials in detrusor smooth muscle of the guinea-pig urinary bladder." European journal of pharmacology 789 (2016): 179-186. [CrossRef]
- Hayase, Masa, Hikaru Hashitani, Kenjiro Kohri, and Hikaru Suzuki. "Role of K+ channels in regulating spontaneous activity in detrusor smooth muscle in situ in the mouse bladder." The Journal of urology 181, no. 5 (2009): 2355-2365. [CrossRef]
- Xin, Wenkuan, Qiuping Cheng, Rupal P. Soder, and Georgi V. Petkov. "Inhibition of phosphodiesterases relaxes detrusor smooth muscle via activation of the large-conductance voltage-and Ca2+-activated K+ channel." American Journal of Physiology-Cell Physiology 302, no. 9 (2012): C1361-C1370. [CrossRef]
- Hristov, Kiril L., Shankar P. Parajuli, Rupal P. Soder, Qiuping Cheng, Eric S. Rovner, and Georgi V. Petkov. "Suppression of human detrusor smooth muscle excitability and contractility via pharmacological activation of large conductance Ca2+-activated K+ channels." American Journal of Physiology-Cell Physiology 302, no. 11 (2012): C1632-C1641. [CrossRef]
- Petkov, Georgi V. "Role of potassium ion channels in detrusor smooth muscle function and dysfunction." Nature Reviews Urology 9, no. 1 (2012): 30-40. [CrossRef]
- Hristov, Kiril L., Amy C. Smith, Shankar P. Parajuli, John Malysz, and Georgi V. Petkov. "Large-conductance voltage-and Ca2+-activated K+ channel regulation by protein kinase C in guinea pig urinary bladder smooth muscle." American Journal of Physiology-Cell Physiology 306, no. 5 (2014): C460-C470. [CrossRef]
- Parajuli, Shankar P., and Georgi V. Petkov. "Activation of muscarinic M3 receptors inhibits large-conductance voltage-and Ca2+-activated K+ channels in rat urinary bladder smooth muscle cells." American Journal of Physiology-Cell Physiology 305, no. 2 (2013): C207-C214. [CrossRef]
- Jenkins, David Paul. Modulators of Small and Intermediate Conductance Calcium-Activated Potassium Channel: Mechanism of Action and In Vivo Effects. University of California, Davis, 2012.
- Hristov, Kiril L., Amy C. Smith, Shankar P. Parajuli, John Malysz, Eric S. Rovner, and Georgi V. Petkov. "Novel regulatory mechanism in human urinary bladder: central role of transient receptor potential melastatin 4 channels in detrusor smooth muscle function." American Journal of Physiology-Cell Physiology 310, no. 7 (2016): C600-C611. [CrossRef]
- Hamilton, Kirk L. "New life in overactive bladder. Focus on “Novel regulatory mechanism in human urinary bladder: central role of transient receptor potential melastatin 4 channels in detrusor smooth muscle function”." American Journal of Physiology-Cell Physiology 310, no. 7 (2016): C597-C599. [CrossRef]
- Smith, Amy C., Kiril L. Hristov, Qiuping Cheng, Wenkuan Xin, Shankar P. Parajuli, Scott Earley, John Malysz, and Georgi V. Petkov. "Novel role for the transient potential receptor melastatin 4 channel in guinea pig detrusor smooth muscle physiology." American Journal of Physiology-Cell Physiology 304, no. 5 (2013): C467-C477. [CrossRef]
- Smith, Amy C., Shankar P. Parajuli, Kiril L. Hristov, Qiuping Cheng, Rupal P. Soder, Serge AY Afeli, Scott Earley, Wenkuan Xin, John Malysz, and Georgi V. Petkov. "TRPM4 channel: a new player in urinary bladder smooth muscle function in rats." American Journal of Physiology-Renal Physiology 304, no. 7 (2013): F918-F929. [CrossRef]
- Provence, Aaron, Eric S. Rovner, and Georgi V. Petkov. "Regulation of transient receptor potential melastatin 4 channel by sarcoplasmic reticulum inositol trisphosphate receptors: Role in human detrusor smooth muscle function." Channels 11, no. 5 (2017): 459-466. [CrossRef]
- Launay, Pierre, Andrea Fleig, Anne-Laure Perraud, Andrew M. Scharenberg, Reinhold Penner, and Jean-Pierre Kinet. "TRPM4 is a Ca2+-activated nonselective cation channel mediating cell membrane depolarization." Cell 109, no. 3 (2002): 397-407. [CrossRef]
- Parajuli, Shankar P., Kiril L. Hristov, Michelle N. Sullivan, Wenkuan Xin, Amy C. Smith, Scott Earley, John Malysz, and Georgi V. Petkov. "Control of urinary bladder smooth muscle excitability by the TRPM4 channel modulator 9-phenanthrol." Channels 7, no. 6 (2013): 537-540. [CrossRef]
- Malysz, John, and Georgi V. Petkov. "Urinary bladder smooth muscle ion channels: expression, function, and regulation in health and disease." American Journal of Physiology-Renal Physiology 319, no. 2 (2020): F257-F283. [CrossRef]
- Hegde, Sharath S., and Richard M. Eglen. "Muscarinic receptor subtypes modulating smooth muscle contractility in the urinary bladder." Life sciences 64, no. 6-7 (1999): 419-428. [CrossRef]
- Ehlert, Frederick J. "Contractile role of M2 and M3 muscarinic receptors in gastrointestinal, airway and urinary bladder smooth muscle." Life sciences 74, no. 2-3 (2003): 355-366. [CrossRef]
- Andersson, Karl-Erik. "Potential benefits of muscarinic M3 receptor selectivity." European Urology Supplements 1, no. 4 (2002): 23-28. [CrossRef]
- Nausch, Bernhard, Thomas J. Heppner, and Mark T. Nelson. "Nerve-released acetylcholine contracts urinary bladder smooth muscle by inducing action potentials independently of IP3-mediated calcium release." American Journal of Physiology-Regulatory, Integrative and Comparative Physiology 299, no. 3 (2010): R878-R888. [CrossRef]
- Goaillard, Jean-Marc, and Eve Marder. "Ion channel degeneracy, variability, and covariation in neuron and circuit resilience." Annual review of neuroscience 44 (2021): 335-357. [CrossRef]
- Walpole, Joseph, Jason A. Papin, and Shayn M. Peirce. "Multiscale computational models of complex biological systems." Annual review of biomedical engineering 15 (2013): 137-154. [CrossRef]
- Aliev RR, Richards W, Wikswo JP. A simple nonlinear model of electrical activity in the intestine. Journal of theoretical biology. 2000 May 7;204(1):21-8. [CrossRef]
- Bursztyn L, Eytan O, Jaffa AJ, Elad D. Mathematical model of excitation-contraction in a uterine smooth muscle cell. American Journal of Physiology-Cell Physiology. 2007 May 1;292(5):C1816-29. [CrossRef]
- Tong WC, Choi CY, Karche S, Holden AV, Zhang H, Taggart MJ. A computational model of the ionic currents, Ca2+ dynamics and action potentials underlying contraction of isolated uterine smooth muscle. PloS one. 2011 Apr 29;6(4):e18685. [CrossRef]
- Rihana S, Terrien J, Germain G, Marque C. Mathematical modeling of electrical activity of uterine muscle cells. Medical and Biological Engineering and Computing. 2009 Jun 1;47(6):665-75. [CrossRef]
- Mahapatra, C. (2023). # 2696 IN SILICO ELECTROPHYSIOLOGICAL STUDY REVEALS TAMSULOSIN MEDIATES URETER SMOOTH MUSCLE CONTRACTION BY INCREASING POTASSIUM CURRENT. Nephrology Dialysis Transplantation, 38(Supplement_1), gfad063c_2696. [CrossRef]
- Mahapatra, C., & Manchanda, R. (2019, July). Computational studies on ureter smooth muscle: modeling ion channels and their role in generating electrical activity. In Proceedings of the 2019 Summer Simulation Conference (pp. 1-6).
- Mahapatra, C., & Pradhan, A. (2022). FC025: Physiological Role of KV Channel in Ureter Smooth Muscle Cell Investigated Quantitatively by Electrophysiological Modeling. Nephrology Dialysis Transplantation, 37(Supplement_3), gfac100-001. [CrossRef]
- Poh YC, Corrias A, Cheng N, Buist ML. A quantitative model of human jejunal smooth muscle cell electrophysiology. PloS one. 2012 Aug 17;7(8): e42385. [CrossRef]
- Mahapatra, C., & Manchanda, R. (2022). In-Silico Investigation of Castration on Vas Deferens Smooth Muscle Electrophysiology. The FASEB Journal, 36. [CrossRef]
- Mahapatra, C., & Manchanda, R. (2019). Modeling Vas Deferens Smooth Muscle Electrophysiology: Role of Ion Channels in Generating Electrical Activity. In Soft Computing for Problem Solving: SocProS 2017, Volume 2 (pp. 655-663). Springer Singapore.
- Mahapatra, C. (2021). MODULATING EFFECTS OF CASTRATION ON VAS DEFERENS SMOOTH MUSCLE ELECTRICAL ACTIVITIES: INSIGHTS FROM A QUANTITATIVE STUDY. Fertility and Sterility, 116(3), e349. [CrossRef]
- Mahapatra, C., & Manchanda, R. (2020) Modeling VAS Deferens Smooth Muscle Electrophysiology: Role of Ion Channels in Generating Electrical Activity. Biophysical Journal, 118(3), 259a-260a. [CrossRef]
- Corrias A, Buist ML. A quantitative model of gastric smooth muscle cellular activation. Annals of biomedical engineering. 2007 Sep 1;35(9):1595-607. [CrossRef]
- Corrias A, Buist ML. Quantitative cellular description of gastric slow wave activity. American Journal of Physiology-Gastrointestinal and Liver Physiology. 2008 Apr 1;294(4): G989-95. [CrossRef]
- Kapela A, Bezerianos A, Tsoukias NM. A mathematical model of Ca2+ dynamics in rat mesenteric smooth muscle cell: agonist and NO stimulation. Journal of theoretical biology. 2008 Jul 21;253(2):238-60. [CrossRef]
- Miftakhov RN, Abdusheva GR, Wingate DL. Model predictions of myoelectrical activity of the small bowel. Biological cybernetics. 1996 Jan 7;74(2):167-79. [CrossRef]
- Mahapatra, C. (2021). Computational Study of Action Potential Generation in Urethral Smooth Muscle Cell. In Computational Advances in Bio and Medical Sciences: 10th International Conference, ICCABS 2020, Virtual Event, December 10-12, 2020, Revised Selected Papers 10 (pp. 26-32). Springer International Publishing.
- Mahapatra, C., Adam, S., & Gupta, A. (2021, December). Computational modeling of electrophysiological properties in urethral smooth muscle cell. In JOURNAL OF COMPUTATIONAL NEUROSCIENCE (Vol. 49, No. SUPPL 1, pp. S81-S83). VAN GODEWIJCKSTRAAT 30, 3311 GZ DORDRECHT, NETHERLANDS: SPRINGER.
- Cha CY, Earm KH, Youm JB, Baek EB, Kim SJ, Earm YE. Electrophysiological modelling of pulmonary artery smooth muscle cells in the rabbits—special consideration to the generation of hypoxic pulmonary vasoconstriction. Progress in biophysics and molecular biology. 2008 Apr 30;96(1):399-420. [CrossRef]
- Mahapatra, C., & Samuilik, I. (2024). A Mathematical Model of Spontaneous Action Potential Based on Stochastics Synaptic Noise Dynamics in Non-Neural Cells. Mathematics, 12(8), 1149. [CrossRef]
- Mahapatra, C., Brain, K. L., & Manchanda, R. (2018). A biophysically constrained computational model of the action potential of mouse urinary bladder smooth muscle. PloS one, 13(7), e0200712. [CrossRef]
- Mahapatra, C., Dave, V., & Manchanda, R. (2017). A Mathematical Modeling of Voltage-gated Calcium ion channel-based Calcium Transient Response in UrinaryBladder Smooth Muscle Cell. International Journal of Pure and Applied Mathematics, 117(9), 71-75. [CrossRef]
- Mahapatra, C., & Manchanda, R. (2017). Simulation of In Vitro-Like Electrical Activities in Urinary Bladder Smooth Muscle Cells. Journal of Biomimetics, Biomaterials and Biomedical Engineering, 33, 45-51. [CrossRef]
- Mahapatra, C., & Manchanda, R. (2019). Modulating Properties of Hyperpolarization-Activated Cation Current in Urinary Bladder Smooth Muscle Excitability: A Simulation Study. In Recent Findings in Intelligent Computing Techniques: Proceedings of the 5th ICACNI 2017, Volume 1 (pp. 261-266). Springer Singapore.
- Mahapatra, C., Brain, K. L., & Manchanda, R. (2018, October). Computational study of Hodgkin-Huxley type calcium-dependent potassium current in urinary bladder over activity. In 2018 IEEE 8th international conference on computational advances in bio and medical sciences (ICCABS) (pp. 1-4). IEEE.
- Dave, V., Mahapatra, C., & Manchanda, R. (2015, August). A mathematical model of the calcium transient in urinary bladder smooth muscle cells. In 2015 37th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC) (pp. 5359-5362). IEEE.
- Hodgkin, A.L.; Huxley, A.F. A quantitative description of membrane current and its application to conduction and excitation in nerve. J. Physiol. 1952, 117, 500. [CrossRef]
- Noble, Denis. "A modification of the Hodgkin—Huxley equations applicable to Purkinje fibre action and pacemaker potentials." The Journal of physiology 160, no. 2 (1962): 317. [CrossRef]
- Fry, C. H., M. Cooklin, J. Birns, and A. R. Mundy. "Measurement of intercellular electrical coupling in guinea-pig detrusor smooth muscle." The Journal of urology 161, no. 2 (1999): 660-664.
- Geng, Yanyan, and Karl L. Magleby. "Single-channel kinetics of BK (Slo1) channels." Frontiers in physiology 5 (2015): 129016. [CrossRef]
- Heeger, David. "Synaptic Input." (2000).
- Hines, Michael L., and Nicholas T. Carnevale. "The NEURON simulation environment." Neural computation 9, no. 6 (1997): 1179-1209. [CrossRef]
- Carnevale, Ted. "Neuron simulation environment." Scholarpedia 2, no. 6 (2007): 1378. [CrossRef]
- Spiess, Andrej-Nikolai, and Natalie Neumeyer. "An evaluation of R 2 as an inadequate measure for nonlinear models in pharmacological and biochemical research: a Monte Carlo approach." BMC pharmacology 10 (2010): 1-11. [CrossRef]
- Demion, Marie, Patrick Bois, Pierre Launay, and Romain Guinamard. "TRPM4, a Ca2+-activated nonselective cation channel in mouse sino-atrial node cells." Cardiovascular research 73, no. 3 (2007): 531-538. [CrossRef]
- Marom, Shimon, and Eve Marder. "A biophysical perspective on the resilience of neuronal excitability across timescales." Nature Reviews Neuroscience 24, no. 10 (2023): 640-652. [CrossRef]
- Chow, K-Y., C. Wu, G. P. Sui, and C. H. Fry. "Role of the T-type Ca2+ current on the contractile performance of guinea pig detrusor smooth muscle." Neurourology and Urodynamics: Official Journal of the International Continence Society 22, no. 1 (2003): 77-82. [CrossRef]
- Jaggar, Jonathan H., Valerie A. Porter, W. Jonathan Lederer, and Mark T. Nelson. "Calcium sparks in smooth muscle." American Journal of Physiology-Cell Physiology 278, no. 2 (2000): C235-C256. [CrossRef]
Figure 1.
Schematic illustration of the proposed physiological role for TRPM4 channels in DSM cells. According to the postulated mechanism, the TRPM4 channels via sarcoplasmic reticulum Ca2+-dependent activation participate in a positive feedback loop to maximize DSM contractility by providing Na+-depolarizing conductance.
Figure 1.
Schematic illustration of the proposed physiological role for TRPM4 channels in DSM cells. According to the postulated mechanism, the TRPM4 channels via sarcoplasmic reticulum Ca2+-dependent activation participate in a positive feedback loop to maximize DSM contractility by providing Na+-depolarizing conductance.
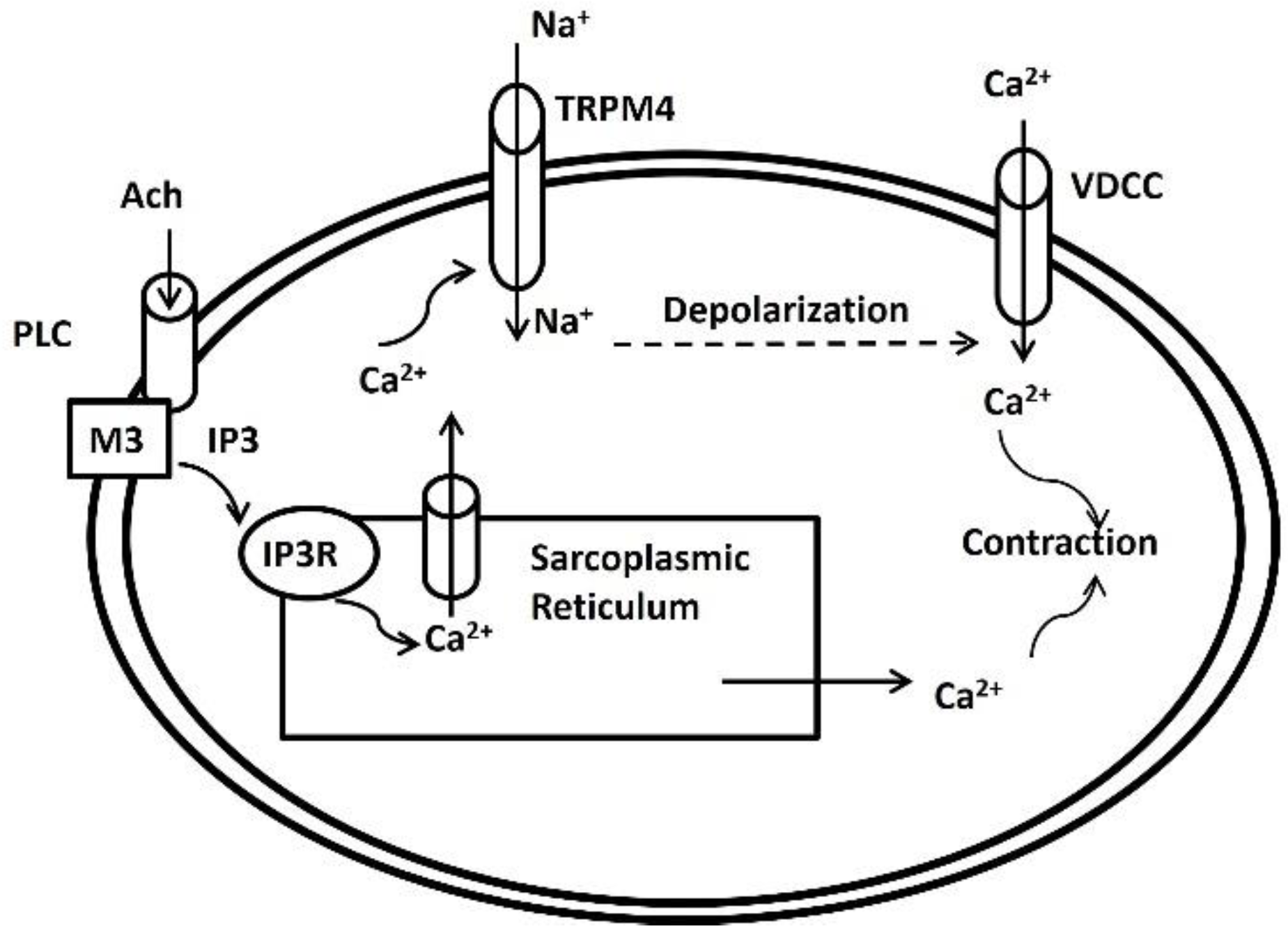
Figure 2.
Schematic overview of parallel conductance model for ionic current. Further elucidation is provided in the subsequent paragraph.
Figure 2.
Schematic overview of parallel conductance model for ionic current. Further elucidation is provided in the subsequent paragraph.
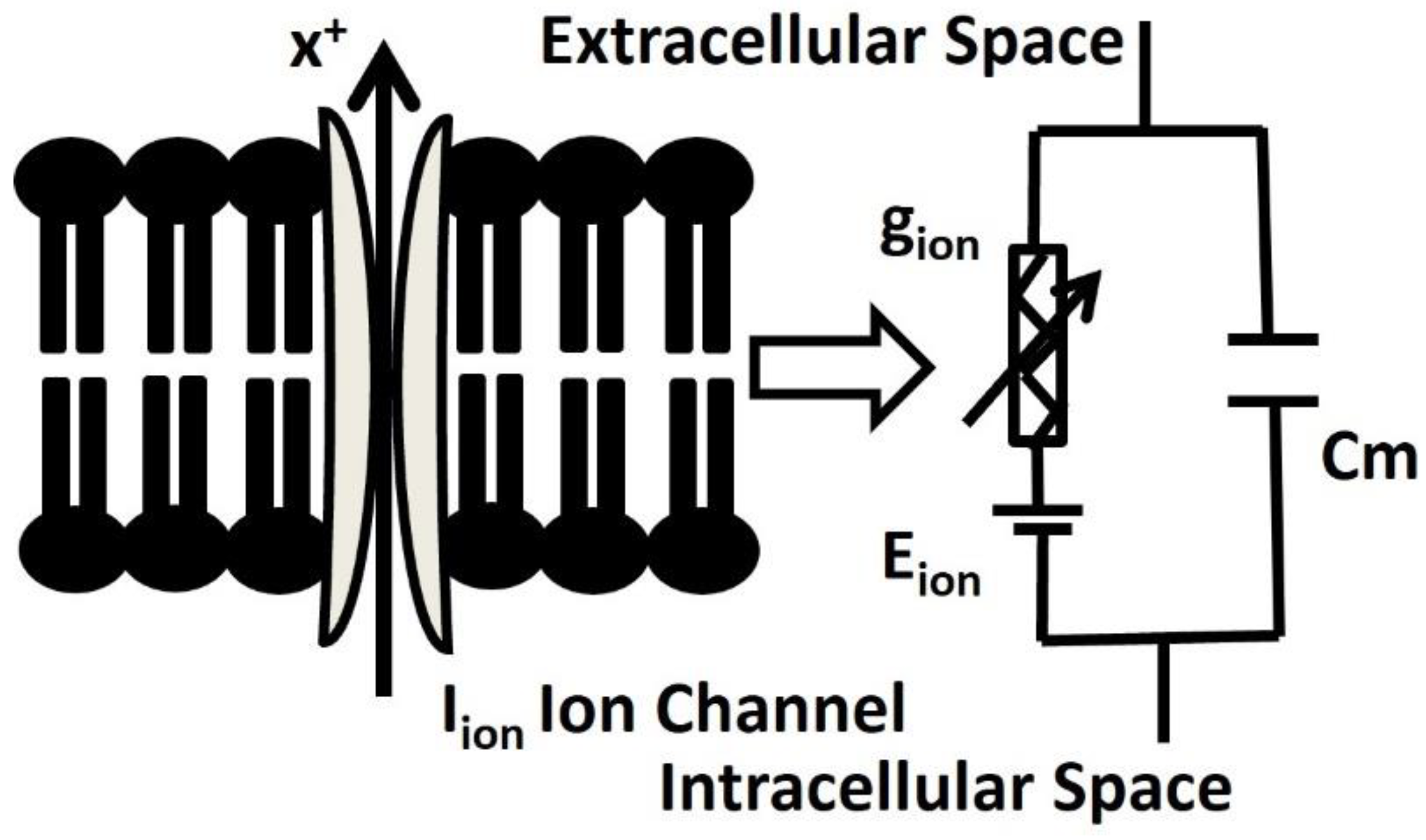
Figure 4.
Schematic diagram illustrating all ionic components within a DSM cell. The accompanying paragraph provides descriptions for each component.
Figure 4.
Schematic diagram illustrating all ionic components within a DSM cell. The accompanying paragraph provides descriptions for each component.
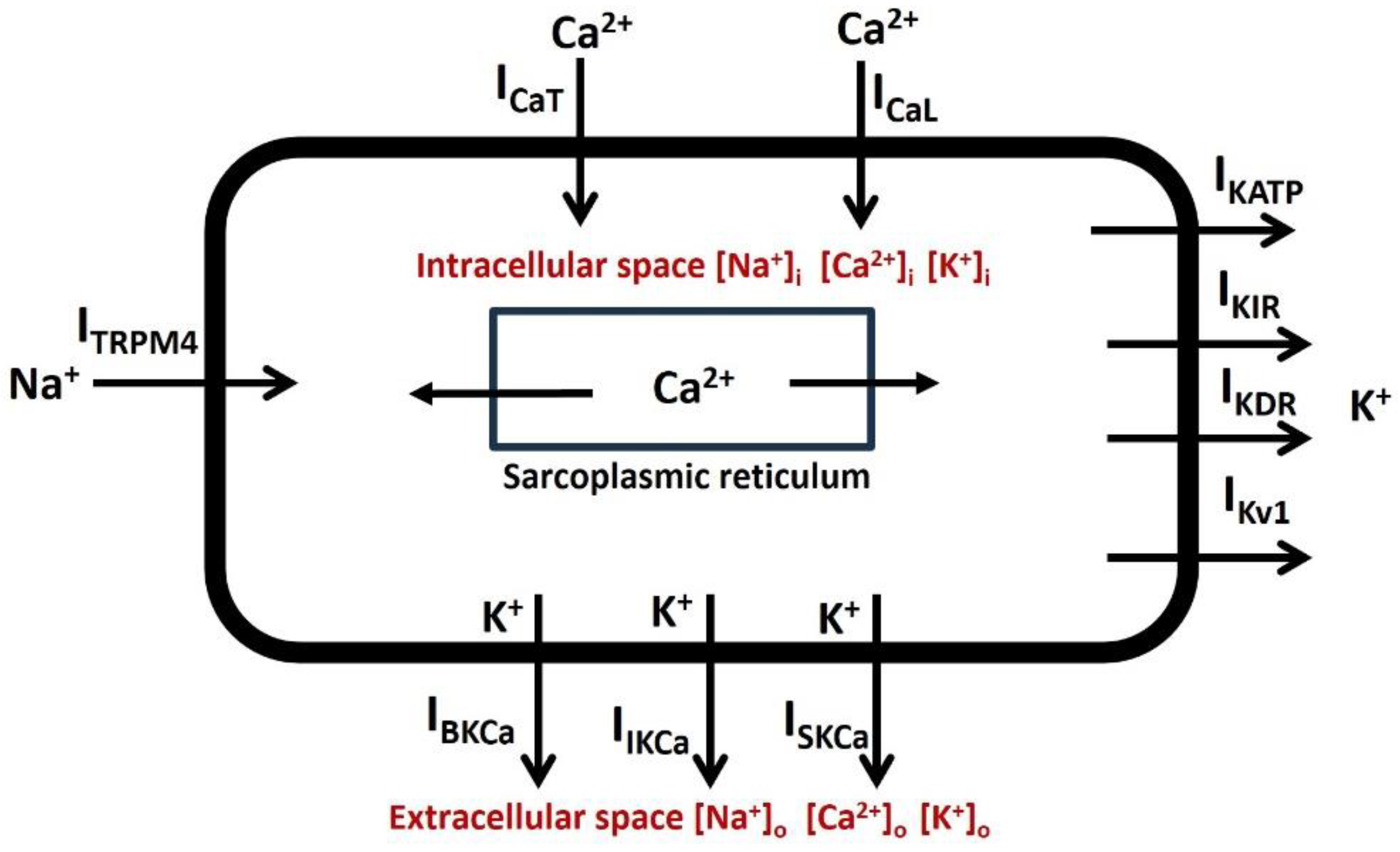
Figure 5.
Simulated steady-state activation curve concerning log (Cai) Solid line represents the result from simulation, where solid filled triangle shows the adapted experimental data from Demion et al, 2007 [].
Figure 5.
Simulated steady-state activation curve concerning log (Cai) Solid line represents the result from simulation, where solid filled triangle shows the adapted experimental data from Demion et al, 2007 [].
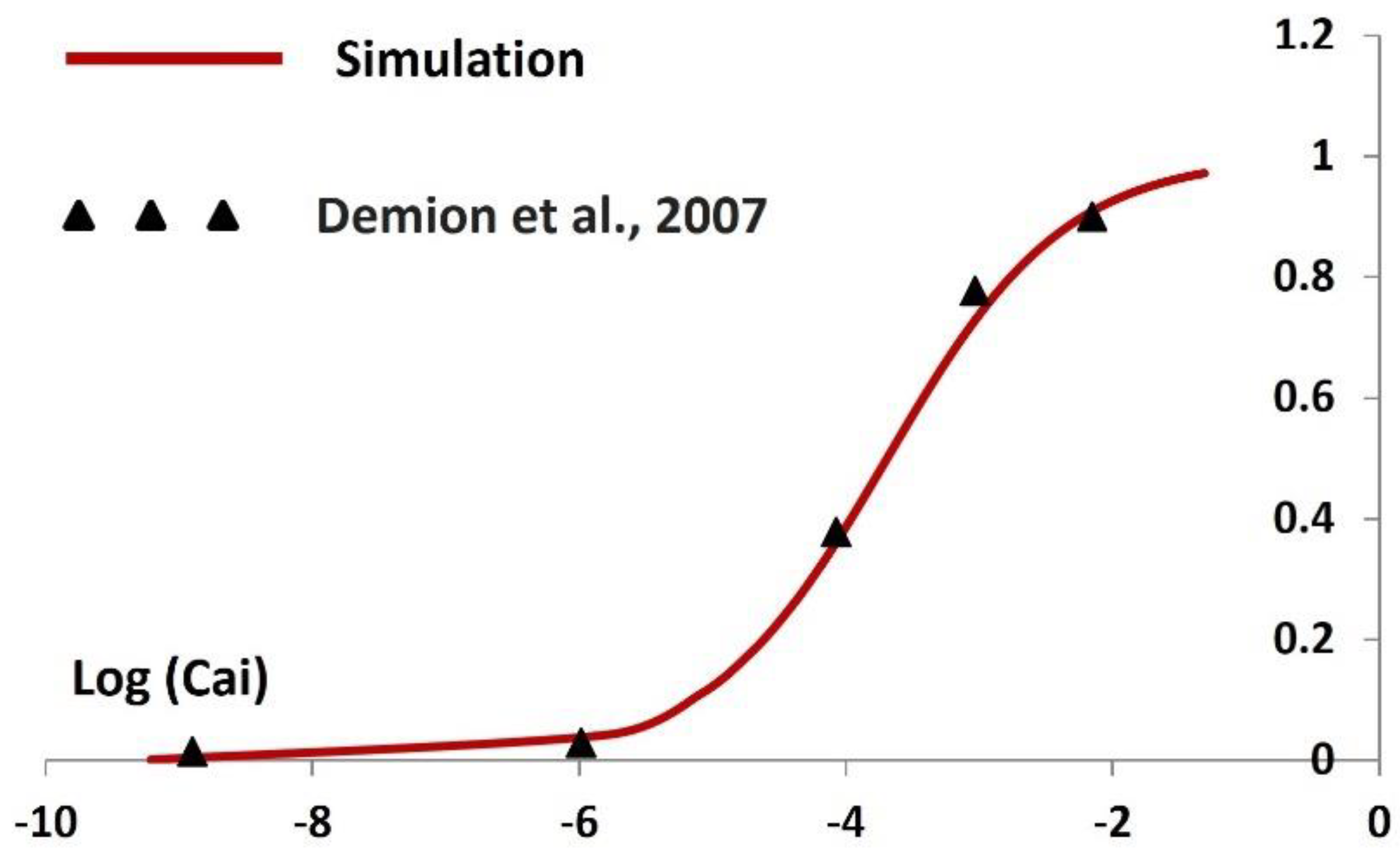
Figure 6.
DSM model shows initial fluctuation (A) and constant resting membrane potential maintained at– 52 mV (B).
Figure 6.
DSM model shows initial fluctuation (A) and constant resting membrane potential maintained at– 52 mV (B).
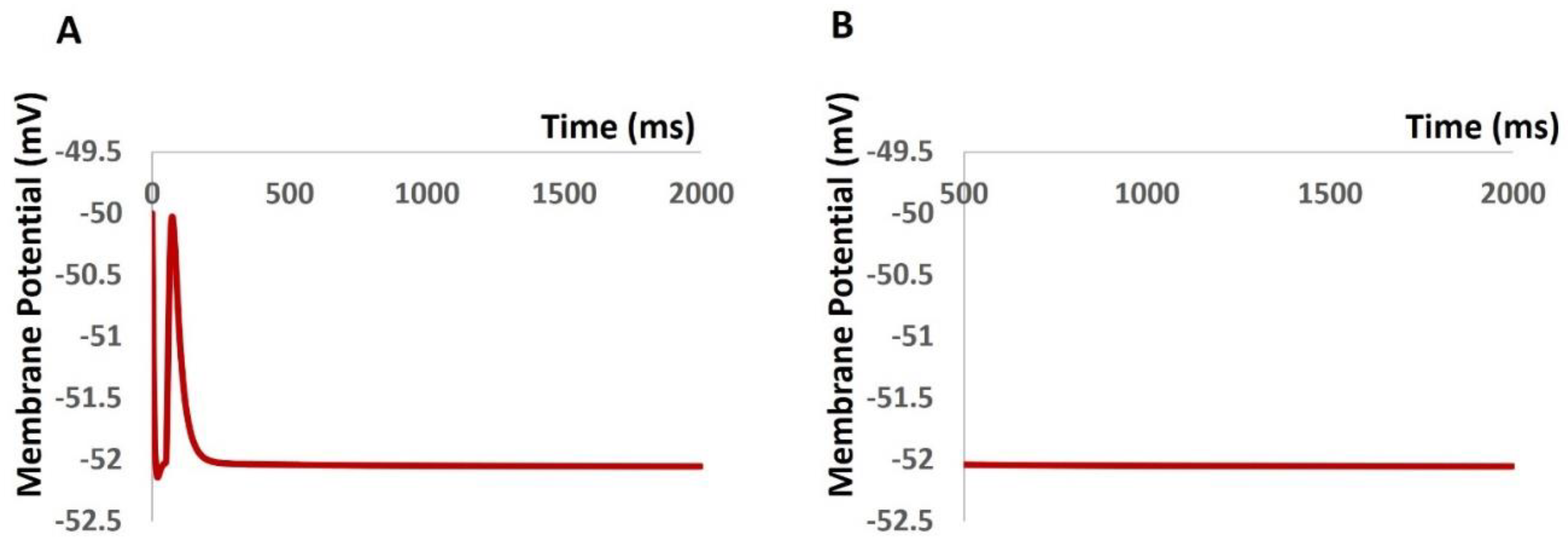
Figure 7.
(A)The model generated AP (solid red line) and depolarization (dashed red line) with the current stimulus. (B)The model generated AP (solid red line), experimental AP (solid blue line), and simulated depolarization (dashed red line) with synaptic input stimulus.
Figure 7.
(A)The model generated AP (solid red line) and depolarization (dashed red line) with the current stimulus. (B)The model generated AP (solid red line), experimental AP (solid blue line), and simulated depolarization (dashed red line) with synaptic input stimulus.
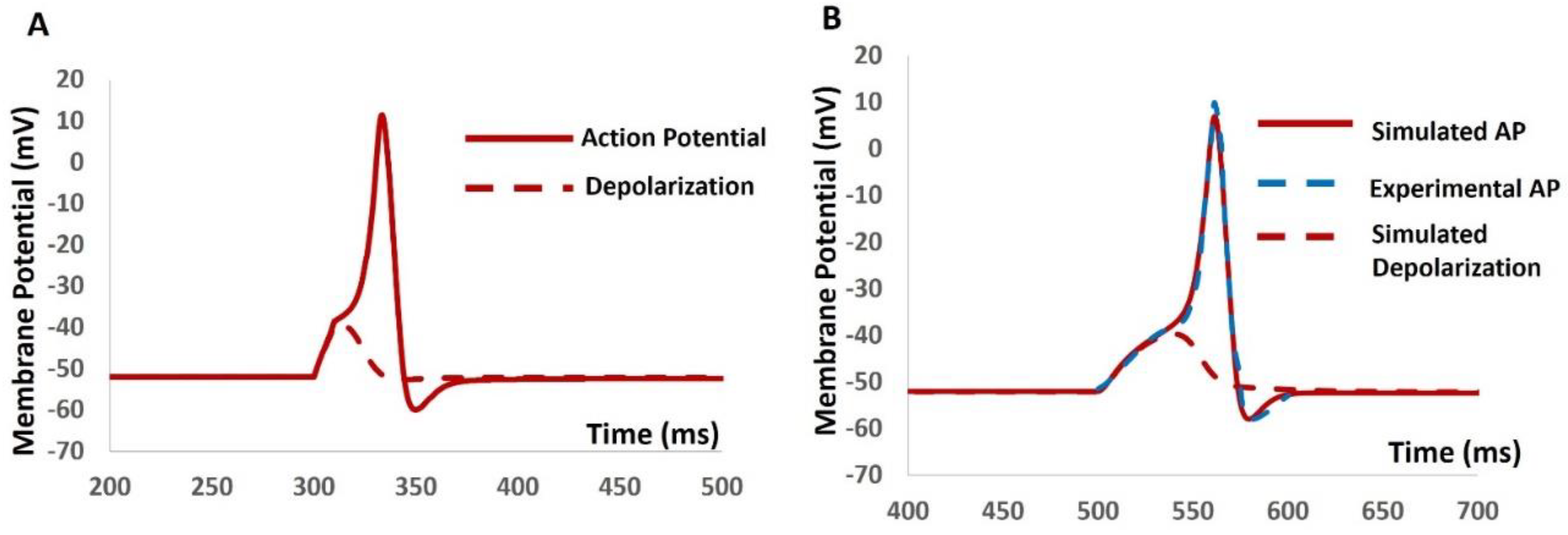
Figure 8.
DSM action potential after a 10% (red solid line) and 20% (black solid line) increase in the maximum conductance of the TRPM4 ion channel.
Figure 8.
DSM action potential after a 10% (red solid line) and 20% (black solid line) increase in the maximum conductance of the TRPM4 ion channel.
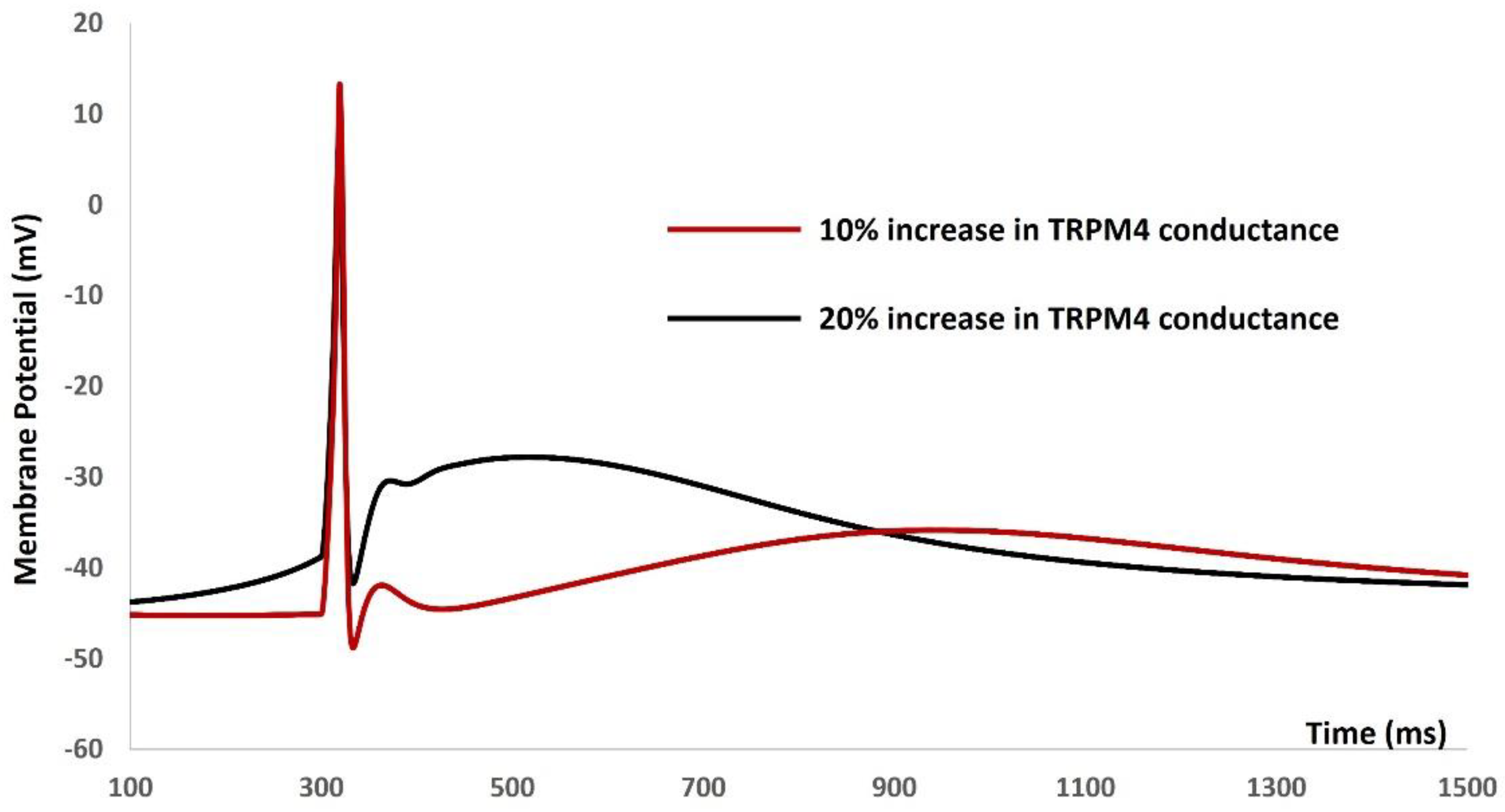
Figure 9.
Sensitivity analysis of the TRPM4 channel conductance for DSM resting membrane potential and action potential duration.
Figure 9.
Sensitivity analysis of the TRPM4 channel conductance for DSM resting membrane potential and action potential duration.
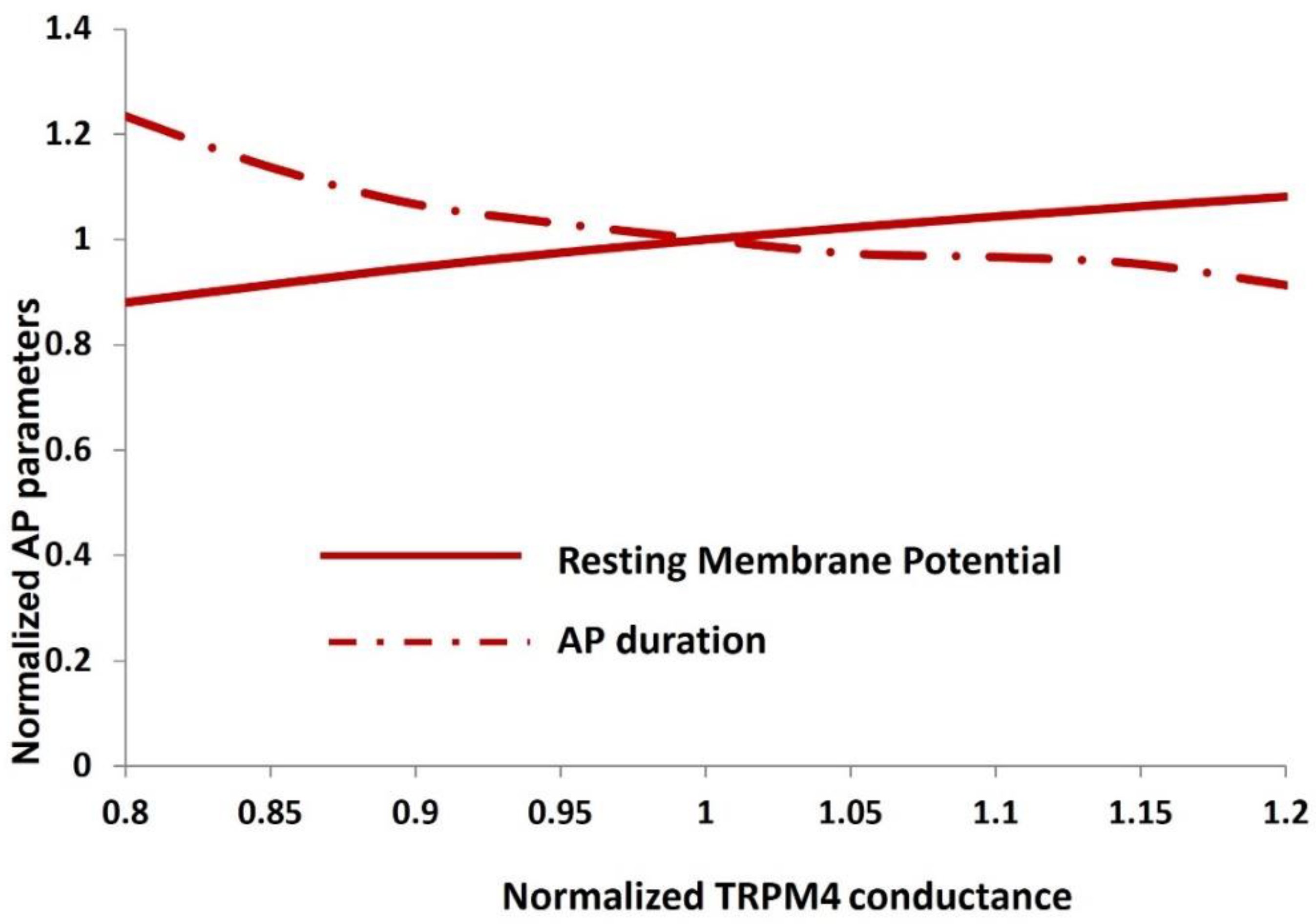
Figure 10.
DSM cell generates action potential (red solid line) with mutation of T-type Ca2+ channel. The dashed solid line depicts the absence of action potential.
Figure 10.
DSM cell generates action potential (red solid line) with mutation of T-type Ca2+ channel. The dashed solid line depicts the absence of action potential.
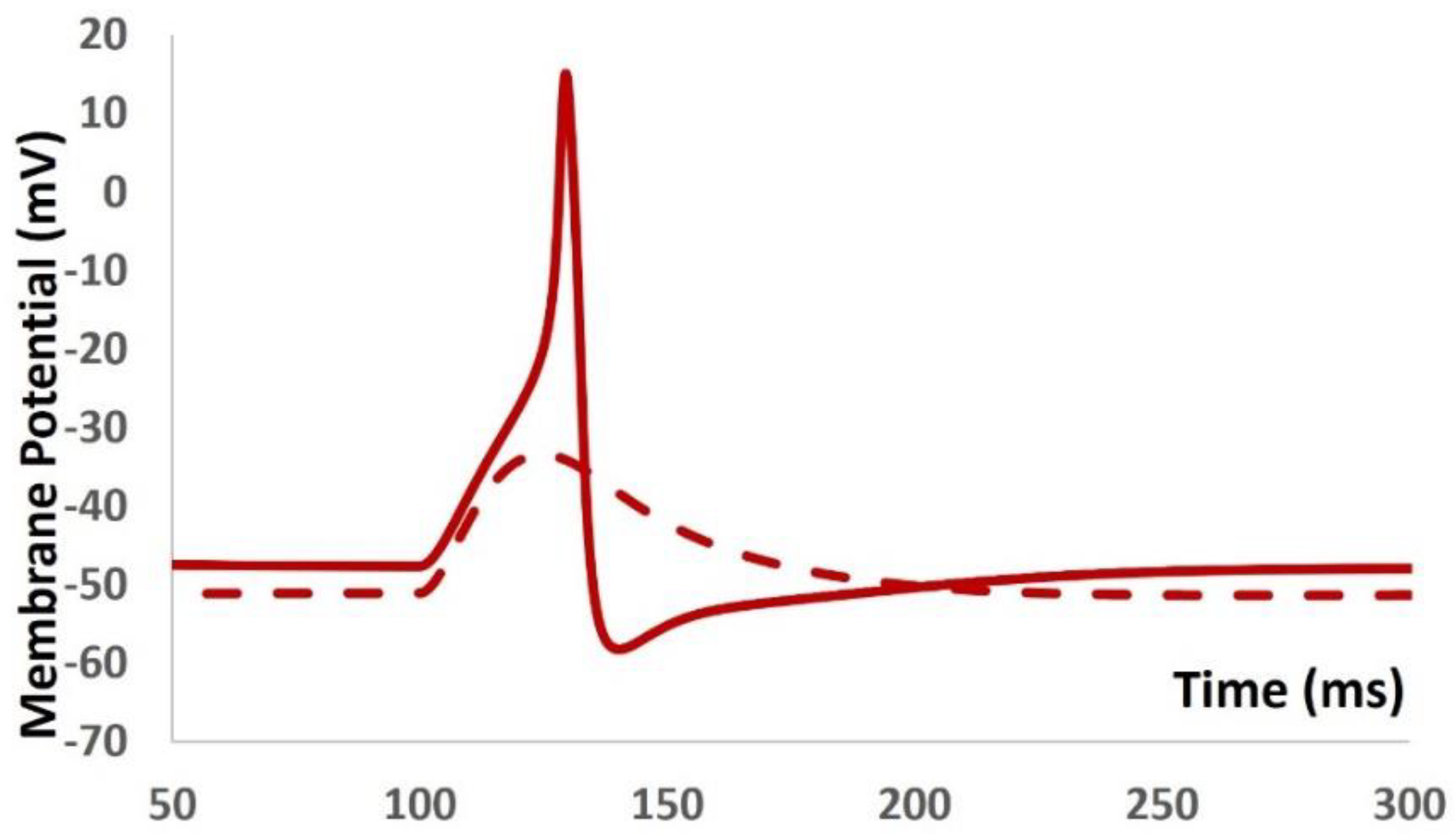

Table 1.
Ion channel maximum conductance.
| Ion Channel | Conductance (S/cm2) |
| T- type Ca2+ channel | 0.0002 |
| L- type Ca2+ channel | 0.0003 |
| Voltage-gated K+ channel –Kv1 | 0.0006 |
| Voltage-gated K+ channel-KDR | 0.0009 |
| Calcium-dependent K+ channel (BK) | 0.0008 |
| Calcium-dependent K+ channel (IK) | 0.0007 |
| Calcium-dependent K+ channel (SK) | 0.0001 |
| ATP-dependent K+ channel | 0.0001 |
| Inward-rectifying channel | 0.0001 |
| TRPM4 Channel | 0.0002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



