
Submitted:
21 May 2024
Posted:
22 May 2024
You are already at the latest version
Abstract
To elucidate the function of the cold resistance regulatory gene FmNAC1 from Fraxinus mandshurica, this study identified the role that the overexpression of the FmNAC1 gene plays in tobacco growth and cold stress regulation. The cloned FmNAC1 gene from F. mandshurica is 891 bp in length and encodes 296 amino acids. Our subcellular localization analysis confirmed that FmNAC1 is primarily located in the nucleus and functions as a transcription factor. FmNAC1 is responsive to cold and NaCl stress, as well as to the induction of IAA, GA, and ABA hormone signals. To further elucidate its function in cold resistance, four transgenic tobacco lines of FmNAC1 overexpression(FmNAC1-OE)were generated through tissue culture after the agrobacterium-mediated transformation of wild-type (WT) tobacco. These FmNAC1-OE plants exhibited accelerated growth after transplantation. When exposed to low-temperature conditions at -5°C for 24 hours, the rates of wilting and yellowing of the FmNAC1-OE plants were significantly lower than those of the WT tobacco plants. Additionally, the membrane integrity, osmotic regulation, and reactive oxygen species (ROS)-scavenging abilities of the FmNAC1-OE tobacco lines were better than those of the WT plants, indicating the potential of the FmNAC1 gene to improve plant cold resistance. The gene expression results further revealed that the FmNAC1 transcription factor exhibits regulatory interactions with growth-related genes such as IAA and AUX1; cold resistance-related genes such as ICE, DREB, and CBF1; and genes involved in the clearance of reactive oxygen species (ROS), such as CAT and SOD. All of this evidence shows that the FmNAC1 transcription factor from F. mandshurica plays a key role in contributing to the enhancement of growth, cold resistance, and ROS clearance in transgenic tobacco plants.
Keywords:
FmNAC1
; Fraxinus mandshurica
; cold resistance
; transgenic tobacco
1. Introduction
Fraxinus mandshurica Rupr., a deciduous tree native to Northeast China and belonging to Fraxinus Linn, is renowned for its robust growth and exceptional cold resistance [1]. Among the distinctive Fraxinus species in the region, it stands out for its remarkable wood quality, which exhibits excellent dryness and suitability for diverse applications. Due to its versatility, F. mandshurica finds extensive uses across various scenarios, owing to its desirable traits and adaptability [2,3,4,5].
When exposed to prolonged periods of low temperatures, plants undergo a spectrum of physiological, biochemical, and phenotypic changes, which collectively contribute to their cold resistance. Cold environments can impede tree growth to a notable degree, particularly impacting roots and young shoots, thus directly affecting tree productivity [6,7,8]. In response to cold stress, plants exhibit a heightened accumulation of reactive oxygen species within the cell membrane system. While these species pose a threat to plants, enzymes like peroxidase (POD) and superoxide dismutase (SOD) play pivotal roles in their detoxification. Nevertheless, excessive reactive oxygen accumulation can lead to lipid membrane oxidation, resulting in the generation of malondialdehyde (MDA). MDA can interact with membrane proteins, causing protein denaturation, elevated membrane permeability, and increased relative conductivity.
Under the conditions of cold stress, plants augment their resilience by elevating their levels of soluble sugars and soluble proteins [9]. Additionally, numerous genes encoding proteins with cold resistance properties are upregulated, further enhancing their expression [9]. These adaptive mechanisms enable plants to effectively overcome the challenges presented by cold environments, thereby enhancing their chances of survival and growth [11,12,13].
The NAC (NAM, no apical meristem; ATAF1/2, Arabidopsis transcription activation factor; CUC2, cup-shaped cotyledon) family of transcription factors is pivotal in plant development and hormonal signal responses [14,15,16]. Representing one of the largest families of plant-specific transcription factors, NAC-domain proteins have been identified across various plant species including Arabidopsis, rice [17], Populus [18], chestnut [19], Cyclocarya paliurus [20], and Catharanthus roseus [21]. These proteins feature a conserved N-terminal NAC domain, which is responsible for DNA and protein binding, serving as a distinguishing hallmark for their identification. Furthermore, they exhibit a diverse array of C-terminal domains. Unlike a classical helix–turn–helix motif, the NAC domain presents a unique transcription factor fold comprising distorted β-sheets encircled by helical elements [22]. NAC proteins can be categorized into six significant branches: MAM/CUC3, secondary wall NAC (SWN), TIP, SNAC (stress-responsive NAC), ANAC034, and OsNAC4 [23].
The involvement of NAC proteins spans across the transcriptional regulation of diverse plant processes, encompassing the development of shoot apical meristems, floral organs, and lateral roots. Additionally, they play a pivotal role in the plant's stress response, modulating pathways associated with stress tolerance, thus enhancing the plant's resilience to environmental challenges [24,25,26,27,28]. Consequently, the NAC family of transcription factors is revealed to be indispensable in coordinating plant growth, development, and adaptation to fluctuating environmental conditions [29].
The significance of NAC transcription factors in imparting resistance to abiotic stresses has gained increasing recognition, with numerous studies underscoring their importance across various plant species [30,31]. In tomato fruit, the overexpression of MaNAC1 enhances cold resistance, leading to thicker cell walls and increased cellulose content. Similarly, in banana fruit, MaNAC1 acts as a positive regulator in governing cellulose metabolism within the cell wall [32]. Additionally, many NAC transcription factors are reportedly induced by cold stress and play pivotal roles in plant responses to such conditions [32]. The identification of the NAC056 transcription factor, which enhances plant cold tolerance by upregulating genes in the CBF pathway and its association with the CBF1-NIA1 regulatory module, underscores the critical role of NAC056 in balancing plant growth and plant responses to cold stress [34].
The interaction between the growth-hormone-inducible NAC1 protein and the RING structural domain protein SINAT5 mediates the involvement of the NAC1 gene in regulating plant abscisic acid control. This interaction attenuates growth hormone signaling by facilitating the ubiquitination and degradation of NAC1. In corn, a significant correlation was observed between the expression levels of ZmNAC1 and the lateral root density. The overexpression of ZmNAC1 in transgenic Arabidopsis plants resulted in a higher number of lateral roots compared to the wild type, indicating the pivotal role of ZmNAC1 in lateral root development [35,36,37,38].
Specific examples include the banana NAC1 transcription factor, which has been demonstrated to enhance cold tolerance in banana plants [39], and PbeNAC1 in pears, which confers cold tolerance to their respective species [40]. In tomato plants, NAC3 is induced by low temperatures and can enhance the cold tolerance of transgenic plants [41]. Moreover, HuNAC20 and HuNAC25 from Hylocereus undatus exhibited a positive effect, increasing cold tolerance in transgenic Arabidopsis by promoting the expression of stress-responsive genes [41]. It is noteworthy that, while extensive research has been conducted on the NAC1 gene in herbaceous plants, relatively little attention has been paid to this gene in woody plants. It is therefore necessary to explore NAC transcription factors in woody plant species to gain a better understanding of their roles in stress tolerance and their potential applications in enhancing the resilience of these important plant varieties.
Notably, the NAC1 gene in F. mandshurica has been underexplored, necessitating further investigations into its role and function within this specific species. This study addresses the primary challenges encountered in cultivating F. mandshurica, particularly the vulnerability of seedlings to cold damage. F. mandshurica is renowned for its unique flowering and leafing patterns, yet it must endure severe environmental conditions, particularly late-spring cold spells in northern regions, to ensure successful growth and development.
In this study, the FmNAC1 gene, a key regulator of plant growth and stress resistance from F. mandshurica, was identified. The full-length sequence of the FmNAC1 gene was obtained, and significant increases in its expression under cold stress and exogenous hormone treatment were also detected. Moreover, a transgenic validation experiment involving the overexpression of the FmNAC1 gene in tobacco plants was conducted. The results showed that the overexpression of FmNAC1 enhanced the cold tolerance of the transgenic tobacco plants and accelerated their growth. These findings provide valuable insights into the molecular mechanisms underlying the growth and cold stress responses of F. mandshurica and offer potential avenues for the future development of improved F. mandshurica varieties with enhanced cold tolerance and overall performance.
2. Results
2.1. Cloning and Sequence Analysis of FmNAC1 Genes and Prediction of the Protein Tertiary Structure
The gene sequence was screened from the F. mandshurica transcriptome database and designated FmNAC1 [67]. Subsequently, the FmNAC1 gene was successfully amplified via PCR using cDNA extracted from F. mandshurica leaves as a template, following software analysis and primer design (Figure 1A). Through NCBI comparison and analysis, it was determined that the total length of the coding region of the FmNAC1 gene (MG269829.1) is 891 bp, encoding 296 amino acids. The analysis of the NAC1 protein revealed that the NAC1 domain coverage of the FmNAC1 protein was 51.88% (Figure 1B,C).
2.2. Phylogenetic Analysis of the FmNAC1 Gene Reveals Relationships with Homologous NAC1 Sequences in Plants
For the homologous alignment, FmNAC1 and 30 other amino acid sequences were aligned using DANMAN. The results revealed that FmNAC1 exhibited a high level of similarity with homologous genes from other species (Figure 2A). FmNAC1 and the 30 other genes shared a NAC conserved domain (Figure 2A). To analyze the phylogenetic relationships between FmNAC1 and homologous NAC1 sequences in other plants, a phylogenetic tree was constructed using MEGA 5.0 software. The gene sequences of FmNAC1 and the other genes were included in the analysis, which revealed that FmNAC1 was most closely related to OeNAC1 (XP_022848339.1) in Olea europaea var., with a homology of 85.14% (Figure 2B).
2.3. Subcellular Localization of the FmNAC1 Protein
Onion epidermal tissue was cocultured in MS media and subjected to GFP fluorescence observations (Figure 3). Due to the absence of a localization signal in the empty vector, the fluorescence signal of the empty vector was clearly observed in the cell membrane and nuclei of the onion epidermal cells under the microscope. In addition, onion nuclei were stained with DAPI, and blue was used as a positive control. The fluorescence signal of the FmNAC1-GFP fusion protein was exclusively detected in the nucleus of the onion epidermis, indicating the nuclear localization of FmNAC1. These results confirm that FmNAC1 is localized in the nucleus and functions as a transcription factor with transcriptional activation activity.
2.4. Expression Pattern of FmNAC1 in F. mandshurica
The expression pattern of the FmNAC1 gene exhibited varying responses to different treatments (Figure 4). After exposure to salt, the expression of this gene increased at 3 h, 12 h, and 24 h but decreased at 1 h, 6 h, and 48 h. The peak expression level was observed at 12 h, reaching 2.58 times that of the control group. Similarly, in response to the cold treatment, the gene expression increased at 3 h and 6 h and peaked at 12 h, reaching a value 21.16 times that of the control group. Conversely, gene expression decreased at the time points of 1 h, 24 h, and 48 h. Additionally, the FmNAC1 gene exhibited responses to hormone signals such as IAA, GA, and ABA, with distinct upregulation observed at specific time points following treatment with these hormones. These findings highlight the dynamic nature of the FmNAC1 gene and its responsiveness to various environmental stimuli and hormone signals.
2.5. Acquisition of Transgenic Tobacco Overexpressing FmNAC1
The transgenic tobacco plants overexpressing FmNAC1 demonstrated successful tissue cultures and regeneration (Figure 5). After three days of preculture, the material started to expand and exhibited signs of infection, which were effectively addressed through significant expansion and bacterial removal following three days of coculture in the dark. By day 15, the plates were nearly fully covered, with small buds emerging on the callus at the incision site; they were subsequently transferred to a rooting screening medium for further growth. These findings indicate favorable responses to genetic modification and provide valuable insights into the potential effects of FmNAC1 overexpression on plant growth and development.
2.6. Identification of FmNAC1-OE Transgenic Tobacco Lines
The successful construction of the overexpression vector facilitated its transformation into wild-type (WT) tobacco tissue-culture-generated plants using Agrobacterium-mediated techniques (Figure 6). The resulting transgenic tobacco plants overexpressing the gene of interest were screened for kanamycin resistance, which resulted in the generation of a significant number of robust and resistant plants. Utilizing tissue culture methodologies such as differentiation and rooting, a substantial quantity of transgenic tobacco material was produced. Subsequently, genomic DNA was extracted individually from each WT and transgenic tobacco strain; this was followed by verification using specific primers through PCR amplification. This analysis revealed four transgenic strains exhibiting amplified specific bands consistent with those of the positive plasmid, while the wild-type samples displayed no amplified bands. These results confirm the successful development of transgenic tobacco lines(T0 generation of transgenic tobacco plants)featuring the enhanced expression of the target gene; this constitutes a strong foundation for further exploration of the implications of FmNAC1 overexpression in tobacco. Four transgenic tobacco lines were subjected to real-time-fluorescence quantitative PCR analysis. In this study, when detecting transgenic heterologous overexpression in tobacco, a tobacco gene was used as the internal reference gene instead of F. mandshurica. The quantitative PCR results revealed varying degrees of expression of the FmNAC1 gene across the four strains. Notably, strain 1 exhibited the highest level of heterologous overexpression, at 74.99%. Consequently, we selected the three strains with the highest expression: lines No. 1, 2, and 5(T2 generation of transgenic tobacco plants)were selected for further investigation.
2.7. Growth Phenotype Analysis of FmNAC1-OE Transgenic Tobacco
The FmNAC1-overexpressing transgenic tobacco plants exhibited a faster growth phenotype (Figure 7), with the height of the transgenic plants measuring 11.3 cm, which was 1.61 times greater than that of the WT plants. These results indicated that the overexpression of the FmNAC1 gene accelerated the growth of the tobacco plants.
2.8. Cold Resistance Analysis of FmNAC1-OE Tobacco Plants
We conducted phenotypic observations of transgenic tobacco under low-temperature stress (Figure 8). Tobacco plants that grew robustly, including both transgenic and wild-type plants, were carefully selected for this experiment. The plants were subjected to a 24-hour incubation at -2°C to induce low-temperature stress, after which the response of the plants was assessed. The white arrow indicates the overexpression of the transgenic tobacco, while the other three pots represent the wild-type controls at the same time. After the low-temperature treatment, the conditions of the overexpression lines were similar to those before treatment. In contrast, the leaves of the wild-type tobacco (WT) plants displayed obvious wilting, with main stem yellowing and softening and the branches withering. These results strongly indicate that the FmNAC1 gene has a robust function in conferring cold resistance.
2.9. Analysis of the Physiological Indexes of FmNAC1-OE Transgenic Tobacco Plants under Low-Temperature Stress
Under cold stress conditions, the FmNAC1 transgenic tobacco plants exhibited consistently lower levels of MDA and relative conductivity in their membrane system compared to wild-type (WT) plants. These levels gradually increased and peaked after 24 hours, showcasing reductions of 39.9% and 38.4%, respectively, in comparison to WT tobacco. This observation suggests that the enhanced resistance of FmNAC1 transgenic tobacco to cold stress, coupled with the maintenance of membrane integrity over a prolonged period, contributes to its heightened tolerance to cold stress.
Additionally, the soluble protein and soluble sugar contents in the osmotic system of the FmNAC1 transgenic tobacco plants consistently exceeded those in the WT tobacco plants under cold stress. These levels displayed an increasing trend and reached peak values after 24 hours, with increases of 34.1% and 65.5%, respectively, compared to those of WT tobacco. This suggests that the solute concentration in the osmotic system can increase in FmNAC1 transgenic tobacco plants, decreasing the freezing point and increasing the tolerance of these plants to cold stress.
Moreover, the activities of POD and SOD in the ROS scavenging system of transgenic tobacco consistently surpassed those of WT tobacco under cold stress. These activities exhibited an increasing trend and peaked after 24 hours, showing increases of 275.7% and 3.6%, respectively, compared to those of WT tobacco. This indicates that FmNAC1 transgenic tobacco can endure cold stress and effectively eliminate accumulated ROS, thereby enhancing its tolerance to cold stress.
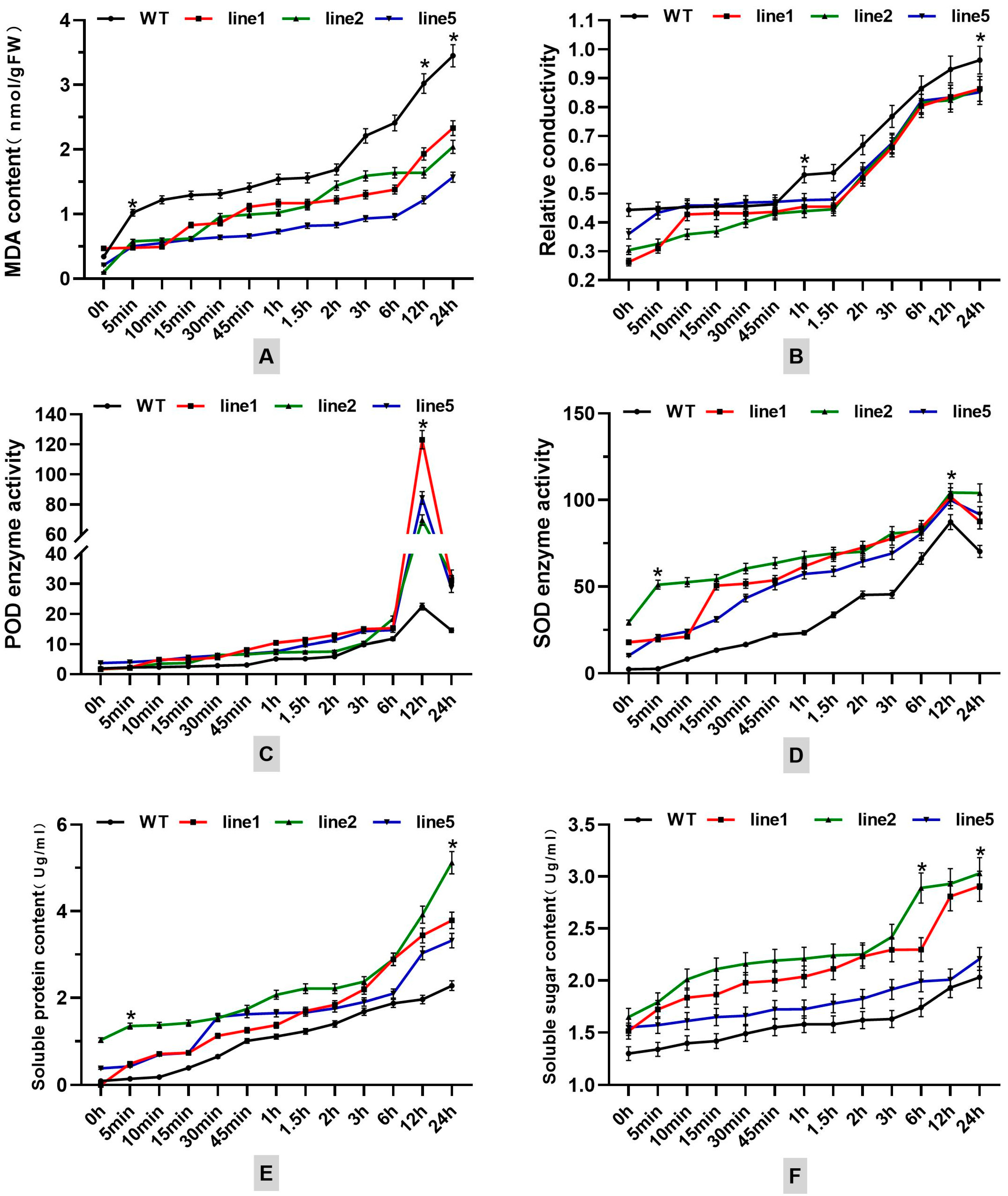
Figure 9.
Analysis of the physiological indexes of transgenic tobacco plants under low-temperature stress: (A) MDA content, (B) relative conductivity, (C) POD activities, (D) SOD activities, (E) soluble protein content, (F) soluble sugar content. The black line represents the wild type, the red line represents transgenic line 1, the green line represents transgenic line 2, and the blue line represents transgenic line 5. * Indicates a significant difference between the control and the transgenic tobacco (p < 0.05).
Figure 9.
Analysis of the physiological indexes of transgenic tobacco plants under low-temperature stress: (A) MDA content, (B) relative conductivity, (C) POD activities, (D) SOD activities, (E) soluble protein content, (F) soluble sugar content. The black line represents the wild type, the red line represents transgenic line 1, the green line represents transgenic line 2, and the blue line represents transgenic line 5. * Indicates a significant difference between the control and the transgenic tobacco (p < 0.05).
2.10. Gene Expression in FmNAC1-OE Transgenic Tobacco Plants
The expression levels of the IAA and AUX1 genes, which are associated with the growth of FmNAC1 transgenic tobacco, were significantly elevated compared to those of the WT, showing increases of 164.8-fold and 19.8-fold, respectively. This indicates a regulatory association between the FmNAC1 transcription factor and the growth-related IAA and AUX1 genes, resulting in the heightened expression of these genes and the promotion of rapid growth in transgenic tobacco.
Under cold stress, the expression of the ICE, DREB and CBF genes, which are linked to cold resistance in FmNAC1 transgenic tobacco, surpassed that in the WT, exhibiting a pattern of an initial increase, a subsequent decrease, and a subsequent increase. Specifically, the expression of ICE peaked after 5 minutes of cold treatment, reaching 54.61 times that of the WT, which decreased to 1.21 times after 30 minutes of cold treatment. Similarly, DREB reached a minor peak after 10 minutes of cold treatment, showing a 19.65-fold increase compared to that of the WT; it reached its highest expression level after 1.5 hours of cold treatment, which was 31.82 times greater than that of the WT. Additionally, the expression of CBF peaked after 10 minutes of cold treatment, exhibiting a 7.79-fold increase compared to that of the WT; it reached its highest level after 24 hours of cold treatment, with a value 10.94 times greater than that of the WT. These findings indicate that the expression of related genes in FmNAC1 transgenic tobacco can respond to cold signals, maintain prolonged high expression levels, and regulate the expression of related genes. This finding suggests that the FmNAC1 transcription factor has a regulatory relationship with the ICE, DREB, and CBF genes associated with cold resistance, resulting in the increased expression of these genes and enhanced cold stress tolerance in transgenic tobacco.
Furthermore, under cold stress, the expression of the CAT and SOD genes involved in the clearance of reactive oxygen species (ROS) in the transgenic tobacco plants was greater than that in the WT plants, displaying a pattern of an initial increase, a subsequent decrease, and a subsequent increase. The expression of both genes peaked after 10 minutes of treatment, showing increases of 23.65-fold and 6.04-fold compared to those in the WT, respectively. These results indicate that FmNAC1 transgenic tobacco can promptly respond to cold signals, maintain prolonged high expression levels, and regulate the expression of ROS-clearance-related genes. This finding suggests that the FmNAC1 transcription factor has a regulatory relationship with the CAT and SOD genes associated with ROS clearance, resulting in the increased expression of these genes, the enhanced ability of transgenic tobacco to scavenge reactive oxygen species, and its increased tolerance to cold stress.
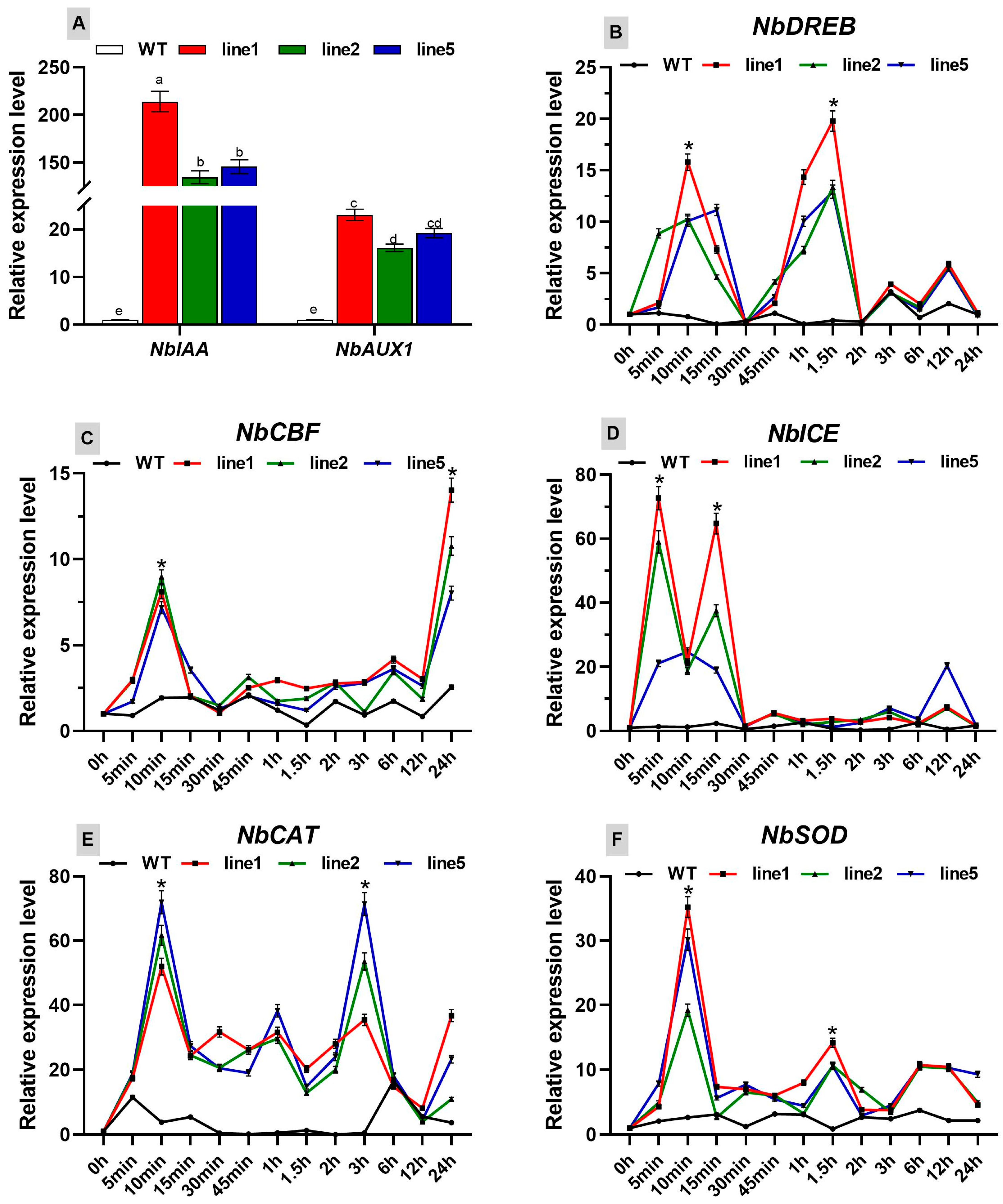
Figure 10.
Gene expression levels in FmNAC1-OE transgenic tobacco plants under cold stress: (A) NbIAA gene; NbAUX1 gene; (B) NbDREB gene; (C) NbCBF gene; (D) NbICE gene; (E) NbCAT gene; and (F) NbSOD gene. The black line represents the wild type, the red line represents transgenic line 1, the green line represents transgenic line 2, and the blue line represents transgenic line 5. * Indicates a significant difference between the control and the transgenic tobacco (p < 0.05).
Figure 10.
Gene expression levels in FmNAC1-OE transgenic tobacco plants under cold stress: (A) NbIAA gene; NbAUX1 gene; (B) NbDREB gene; (C) NbCBF gene; (D) NbICE gene; (E) NbCAT gene; and (F) NbSOD gene. The black line represents the wild type, the red line represents transgenic line 1, the green line represents transgenic line 2, and the blue line represents transgenic line 5. * Indicates a significant difference between the control and the transgenic tobacco (p < 0.05).
3. Discussion
Transcription factors in the NAC family are well known to play a pivotal role in plant development and stress response [44]. Introducing or augmenting key NAC transcription factors such as NAC1 in plants has the potential to enhance wood quality, boost yields, and fortify plant resistance to diverse stressors. Our study aimed to elucidate the molecular mechanism underlying FmNAC1's contribution to stress resilience in F. mandshurica, a species of significant economic and ecological value.
Upon exposure to NaCl, cold stress, and hormonal signals such as IAA, GA, and ABA, the expression of FmNAC1 in F. mandshurica initially increased before subsequently decreasing. This pattern suggests that FmNAC1 can respond to a variety of stressors including cold, salinity, and hormonal fluctuations, indicating its crucial role in both stress adaptation and growth regulation in F. mandshurica. When exposed to stress, this species adjusts its hormonal balance to adapt to external environmental conditions. Notably, the expression of FmNAC1 was significantly more responsive to IAA than to GA or ABA. This heightened responsiveness to IAA may be due to the impact of cold stress on NAC1 gene expression, potentially enhancing cold resistance. The involvement of FmNAC1 in plant growth, particularly in response to IAA hormonal changes, appears to be pivotal, highlighting its integral role in the developmental processes and stress responses of F. mandshurica.
After the transgenic tobacco was transplanted, measurements were taken every 2 days for a total of 11 days to determine the plant height; this resulted in a final height of 11.3 cm, marking a 1.61-fold increase compared to their wild-type counterparts. These findings highlight the role of the overexpression of the FmNAC1 gene in accelerating tobacco plant growth. Upon exposure to low temperatures (-5℃ for 24 hours), wild-type tobacco (WT) exhibited visible wilting, leaf yellowing, stem softening, and branch withering. Conversely, transgenic lines overexpressing FmNAC1 showcased significantly enhanced growth compared to the WT, maintaining robustness with thick stems and green leaves. These observations underscore the robust cold resistance conferred by the FmNAC1 gene. Furthermore, the heightened growth and structural integrity observed in transgenic tobacco plants bolster their resilience to environmental fluctuations. By stimulating root growth, the FmNAC1 gene facilitates nutrient absorption, thereby promoting rapid growth and augmenting cold tolerance [34].
In assessing the membrane damage caused by low temperatures, relative conductivity and MDA content serve as reliable indicators. In our investigation, we consistently observed a lower MDA content and lower relative conductivity in the membrane system of FmNAC1 transgenic tobacco compared to regular tobacco when subjected to cold stress. Even after 24 hours of cold treatment, the transgenic plants overexpressing FmNAC1 maintained a lower MDA content and lower relative conductivity compared to the control plants. With prolonged cold treatment, the MDA content gradually increased, indicating exacerbated cell damage. Notably, all samples exhibited a gradual rise in MDA content before 12 hours of cold treatment, which can potentially be attributed to elevated catalase and superoxide dismutase activity. However, despite the heightened activities of these enzymes after 24 hours of cold treatment, MDA accumulation persisted due to the extensive cellular damage induced by prolonged cold stress [46,47,48,49,50].
Under cold stress, FmNAC1 transgenic tobacco consistently exhibited elevated levels of soluble protein, soluble sugar, peroxidase (POD), and superoxide dismutase (SOD) in the osmotic system compared to conventional tobacco. The modification of osmoregulatory substances, such as soluble sugars and soluble proteins, enhances plants' ability to withstand extreme temperatures [51,52,53]. Soluble sugars play a pivotal role in plant responses to cold stress, with an increased content being crucial in mitigating damage from low temperatures. Throughout our study, prolonged exposure to cold treatment led to increased levels of soluble sugars and soluble proteins. This rise in content could effectively enhance the cold resistance of transgenic plants overexpressing FmNAC1. The enhanced cold stress tolerance observed in transgenic FmNAC1 tobacco suggests the gene's potential to augment nutrient accumulation, promoting robust growth and influencing the plants' ability to scavenge active oxygen, thus bolstering cold resistance.
GmNAC20 functions by controlling the expression of stress-tolerance-enhancing COR genes, DREBIA/CBF3, and DREB1C/CBF2. It directly interacts with the promoters of DREB1A/CBF3 and DREB1C/CBF2, leading to increased CBF3 transcription and decreased CBF2 transcription [54]. In bananas, MaNAC1 is regulated by MaICE1, and this protein interacts with MaCBF1 to govern cold tolerance in banana fruit [55]. Furthermore, the PbeNAC1 protein can enhance the transcription level of stress-related genes by interacting with PbeDREB1 and PbeDREB2A, thereby boosting the tolerance of transgenic tobacco [56]. These findings underscore the involvement of NAC TEs in downstream ICE1 and upstream CBF regulation.
Similarly, in F. mandshurica, FmNAC1 has been shown to have a regulatory relationship with genes associated with growth (IAA and AUX1), cold resistance (ICE, DREB, and CBF1), and ROS clearance (CAT and SOD) in transgenic tobacco under cold treatment [57,58,59,60]. It is suggested that FmNAC1 genes directly regulate CBF and DREB genes downstream, thereby promoting rapid growth, cold resistance, and ROS clearance in transgenic tobacco. NAC1 is characterized as a regulator of fast plant growth and cold resistance in F. mandshurica, potentially serving as a pivotal factor in balancing these two biological traits [61]. However, the specific mechanism by which NAC1 maintains this balance remains a primary focus of future research [62,63].
4. Conclusion
The aim of this study was to confirm the function of FmNAC1 from F. mandshurica in plant cold stress resistance and growth regulation. The transcription factor FmNAC1 in F. mandshurica was identified, and its expression was responsive to cold stress and various exogenous hormones. The overexpression of the FmNAC1 gene not only promoted growth but also enhanced cold resistance in tobacco plants. FmNAC1 overexpression enhanced growth and cold stress tolerance by modulating osmoregulatory substances and the expression of key genes, such as growth-related genes (IAA and AUX1), cold resistance-related genes (ICE, DREB, and CBF), and ROS-clearance-related genes (CAT and SOD) in transgenic tobacco plants. These findings provide valuable insights into the molecular mechanisms underlying the growth and cold stress responses of the valuable timber tree species F. mandshurica.
5. Materials and Methods
5.1. Plant Material and Growth Conditions
Nicotiana tabacum and F. mandshurica seeds from the Northeast Forestry University experimental forest farm were used. The seeds were surface sterilized and grown at 25 °C under long-day conditions (16h light/8h dark) on a standard field.
5.2. Cloning and Identification of the FmNAC1 Gene
A MiniBEST Plant RNA Extraction Kit (Takara Bio, Inc., Japan) was used for the total RNA extraction. The synthesized cDNA was created using the PrimeScript First Strand cDNA Synthesis Kit (Takara Bio Inc., Shiga, Japan). Then, a PCR was performed to amplify the F. mandshurica cDNA (template) for 40 cycles using 2×Phanta Flash Master Mix (Dye Plus; Vazyme, Nanjing, China) and FmNAC1 primers (Table 1). The PCR program was as follows: predenaturation at 94°C for 5 min; denaturation at 94°C for 30s; annealing at 53°C for 45s; extension at 72°C for 30s [64]. After electrophoresis on 2% agarose gel, the PCR products were purified using the Zymoclea Gel DNA Recovery Kit (ZYMO), cloned into the pCE2 TA/Blunt-Zero Vector (Vazyme), and transformed into Escherichia coli DH5a cells [66]. The sequence of the FmNAC1 gene was submitted to GenBank with the accession number MG269829.1.
5.3. Experimental Approach for FmNAC1 Protein Subcellular Localization
The Thermo ScientificTM Spel single enzyme-cut PBI121-GFP plasmid was incubated at 37°C for 15min and then heat-inactivated at 80°C for 5min. The linearized vector obtained from single-enzyme digestion was designed for the homologous recombination of the target gene according to the ClonExpress principle. Primers for the target gene were designed as follows: PBNAC1-GFP-F (5’-tacccgggtcgactgactagtATGAGTAGTTTGAGCATGATAGAGGC-3’) and PBNAC1-GFP-R (5’- gcccttgctcaccatactagtTCAATAATGGTTCCACTGCAATCC-3’). The insert fragment was obtained via high-fidelity enzyme PCR using the subcloned vector of the target gene as a template. Homologous recombination was performed using ClonExpress/Mut Express enzymes (Vazyme, Nanjing, China), and the resulting construct was transformed into Escherichia coli DH5a cells (Figure 11A).
A transient expression vector for PBI121-FmNAC1-GFP was constructed and transformed into Agrobacterium GV3103 cells. Five to six layers of onion scale were soaked in 75% ethanol for 10 min and sterile water was added 3–5 times; then, l cm2 of onion epidermis was cut with a sterile scalpel, spread in hypertonic medium, and cultured for 24 h. At OD = l, the cells were infected for 20 min, washed several times, and placed in coculture medium for 24–36 h.
5.4. Abiotic Stresses and Hormone Signal Treatments
Thirty-day-old uniformly growing manchurian seedlings were subjected to hormone and abiotic stress treatments. For the low-temperature stress treatment, manchurian seedlings were transferred to liquid MS medium and placed in a 4℃ low-temperature culture chamber [68]. For the salt stress, seedlings were transferred to MS liquid medium containing 200 mmol/L NaCl. For the hormone treatments, seedlings were transferred to MS liquid medium containing 100 mmol/L ABA, 100 pmol/L GA3, and 100 pmol/L IAA. For the control group, untreated seedlings were transferred to MS liquid medium and cultured at 25℃ under long-day photoperiod conditions (16h light/8h dark). All materials were treated for 48 hours, and samples of the treated manchurian seedlings were taken at 1 h, 3 h, 6 h, 12 h, 24 h, and 48 h before being held in liquid nitrogen and then stored in a -80℃ freezer for later use. Each treatment group was set up with 5 biological replicates.
5.5. Agrobacterium-Mediated Genetic Transformation, Identification, and Propagation of Nicotiana
The Thermo ScientificTM XbaI and KpnI double-enzyme-cut vector pROKII-GUS plasmid was incubated at 37°C for 15 min and then heat-inactivated at 80°C for 5 min. The linearized vector obtained from the double-enzyme digestion was designed for the homologous recombination of the target gene according to the ClonExpress principle. Primers for the target gene were designed as follows: pRNAC1-GUS-F (5’- gagaacacgggggactctagaATGAGTAGTTTGAGCATGATAGAGGC-3’) and pRNAC1-GUS-R (5’- catggtcaagagtccggtaccTCAATAATGGTTCCACTGCAATCC-3’). The insert fragment was obtained via a high-fidelity enzyme PCR, using the subcloned vector of the target gene as a template. Homologous recombination was performed using ClonExpress/Mut Express enzymes (Vazyme, Nanjing, China), and the resulting construct was transformed into Escherichia coli DH5a cells (Figure 11B). A transient expression vector for pROKII-FmNAC1-GUS was constructed and transformed into Agrobacterium GV3103 cells.
Native tobacco seeds were disinfected on an ultraclean workbench, soaked in 30% 84 disinfection dilution solution for 30 min, and washed 3–5 times in sterile water. Afterwards, the seeds were disinfected with 75% alcohol for 30 s, and the triangular bottle was constantly shaken. The plants were quickly transferred to 0.1% mercury solution and shaken for 6–8 min, sterile water was removed many times, and the water on the seed surface was left to dry on sterilized filter paper. The bacteria were vaccinated in MS media and then cultured once after 28 days of sterile culture. The native tobacco plants were transformed via the leaf disc method as follows: high-quality tender green tobacco leaves were picked, the vein trunk was removed, the plants were cut into 1 cm2 pieces on MS media, and the plants were precultured for 1–2 days. The engineered bacteria were inoculated into sterile Petri dishes, the leaves were soaked for 5 min, and then the leaf surface liquid was removed. The plants were transferred to a coculture media and grown in the dark for 2 days. After sterilization once a day for 5 d, the leaf explants were washed 3–5 times with 500 mg/L cephalosporin, dried for surface water, and transferred to sterile media (MS+0.05 mg/L NAA+0.5 mg/L 6-BA + 500 mg/L cephalosporin + 25 mg/L Kana, pH=6). Leaf explants were placed in differentiation screening media (MS+0.05 mg/L NAA+0.5 mg/L 6-BA + 500 mg/L cephalosporin + 50 mg/L Kana, pH=6) to select the resistant buds. Days 7 to 14 were selected for bud production so that the buds could fully contact the selection medium. The buds that were able to grow normally were resistant buds. When the resistant buds grew to approximately 3 cm, they were removed and placed in differentiation screening media. The resistant shoots were cultured for 7 d in rooting screening media (MS+0.25 mg/L NAA + 500 mg/L cephalosporin + 50 mg/L Kana, pH=6) for further transgenic testing of the positive transformed shoots [64].&#;
Identification and propagation of transgenic tobacco: DNA was extracted from tobacco leaves using the cetyltrimethylammonium bromide (CTAB) method [65]. DNA was extracted from positive tobacco plants selected on the resistance culture medium and was then amplified and identified using primers for cloning the FmNAC1 gene and PCR conditions, with the recombinant plasmid as positive control. Part of the material was subsequently transplanted to a 1:1 mixture of chat soil/vermiculite matrix and cultured at a constant temperature and humidity.
5.6. Determination of Physiological Traits&#;&#;
POD activity was assayed using the photochemical guaiacol method. SOD activity was assayed using the photochemical NBT method. The concentration of MDA was assayed using the thiobarbituric acid method [2]. The soluble sugar content was determined using the anthrone colorimetric method [44]. The soluble protein content was determined using the Coomassie Brilliant Blue G-250 method [43].
5.7. Analysis of Low-Temperature Tolerance in Transgenic Tobacco Plants
The plants in the transgenic tobacco group were subjected to a -2°C low-temperature treatment for 0 min, 5 min, 10 min, 15 min, 30 min, 45 min, 1 h, 1.5 h, 2 h, 3 h, 6 h, 12 h, and 24 h. The tobacco status was observed, and the plants were precooled in liquid nitrogen in a -80°C refrigerator. The physiological and biochemical indicators were subsequently measured, and qRT-PCR was used to detect changes in the expression of genes related to low-temperature-resistance-related phenotypes at the molecular level of the FmNAC1 protein.
5.8. Analysis of Gene Expression
The quantified RNA was reverse transcribed into cDNA using the PrimeScript™ RT reagent kit with a gDNA Eraser (Perfect Real-Time) (Takara Bio Inc., Shiga, Japan). Quantitative RT-PCR (qRTPCR) was conducted in a 7500 Real-Time PCR system (Applied Biosystems, Forster City, CA, USA) using the Takara SYBR® Premix Ex Taq™ II (Perfect Real Time) (Takara Bio, Inc., Japan). All reactions were performed in triplicate to ensure technical and biological reproducibility, and the relative abundance of the transcripts was calculated using 7500 Software v 2.0.6 (Applied Biosystems, Forster City, CA, USA) using the comparative 2−△△CT method. The qRT-PCR primer pairs are shown in Table 2. Tubulin was used as an internal control to determine the expression levels of the target genes.
5.9. Statistical Analysis of Data
Data processing and statistics: all physiological and biochemical index (MDA, antioxidant enzyme activity, qPCR, phytohormone content) data are presented as the mean ± the standard deviation (n = 3). The effects of the cold treatment, the biotype, and the interactions between the two factors were determined via an analysis of variance (ANOVA) followed by Duncan’s multiple range test (with a probability level of 0.05 indicating statistical significance) using Excel and SPSS 20.0, Inc. (Chicago, IL, USA). The figures were drawn using Prism 8.0.
Author Contributions
Y.Z, L.H., and Y.C. conceived the study and designed the experiments. C.Z.Y.C. and L.H. performed most of these experiments, and the other authors provided their help in the treatment and sample collection. Y.C. and G.P. analyzed the data and generated the figures. Y.W., Y.C., L.H., Y.Z.and B.L. were involved in writing and revising the manuscript. All authors approved the final manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Key R&D Program of China (2021YFD2200303) and the Postdoctoral Fund of Heilongjiang Province (LBH-Z21095). These funders played a role in the design of the study and data collection and analysis, and in the writing of the manuscript.
Availability of Data and Materials
The datasets generated and analyzed during the course of the current study are available from the National Center for Biotechnology Information repository. The sequence of FmNAC1 used in this study can be accessed at accession number FmNAC1 (https://www.ncbi.nlm.nih.gov/nucleotide/MG269829.1?report=genbank&log$=nuclalign&blast_rank=1&RID=HDXDRHRJ013)).
Acknowledgments
Not applicable.
Ethical Approval and Consent to Participate
The experimental materials were obtained from complete plants cultured in the greenhouse of Northeast Forestry University.
Consent for Publication
Not applicable.
Competing Interests
The authors declare no conflicts of interest.
Author Details
National Key Laboratory of Tree Genetics and Breeding, Northeast Forestry University, Harbin 150040, People’s Republic of China; Heilongjiang Touyan Innovation Team Program (Tree Genetics and Breeding Innovation), Northeast Forestry University, Harbin, China; College of Life Sciences, Northeast Forestry University, Harbin, China.
Abbreviations
IAA: 3-Indoleacetic acid; AUX1: auxin resistant 1; DREB: dehydration-responsive element-binding protein; CBF1: C-repeat/DRE binding factor 1; CAT: catalase; SOD: superoxide dismutase; GA: gibberellin; ABA: abscisic acid; MDA: malondialdehyde; SOD: superoxide dismutase; POD: peroxidase.
References
- Zhao, X.; Zhang, X.; Liu, Z.; et al. Comparing the Effects of N and P Deficiency on Physiology and Growth for Fast- and Slow-Growing Provenances of Fraxinus mandshurica [J]. Forests, 2021, 12, 1760. [Google Scholar] [CrossRef]
- Cao, Y.; He, L.; Song, F.; et al. Physiological and Gene Expression Response of Interspecific Hybrids of Fraxinus mandshurica × Fraxinus americana to MJ or SNP under Drought [J]. Forests, 2023, 14, 1277. [Google Scholar] [CrossRef]
- Hu, L.J.; Uchiyama, K.; Shen, H.L.; et al. Nuclear DNA microsatellites reveal genetic variation but a lack of phylogeographical structure in an endangered species, Fraxinus mandshurica, across north‒east China [J]. Annals of Botany, 2008, 102, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.M.; Preece, J.E.; Shen, H.L. Somatic embryogenesis in immature cotyledons of Manchurian ash (Fraxinus mandshurica Rupr. ) [J]. Plant Cell, Tissue and Organ Culture (PCTOC), 2012, 108, 485–492. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, W.W.; Li, N.; et al. Photosynthesis and growth responses of Fraxinus mandshurica Rupr. seedlings to a gradient of simulated nitrogen deposition [J]. Annals of Forest Science, 2018, 75, 1. [Google Scholar]
- Chinnusamy, V.; Zhu, J.; Zhu, J.K. Cold stress regulation of gene expression in plants [J]. Trends in plant science, 2007, 12, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: an overview [J]. Archives of biochemistry and biophysics, 2005, 444, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Thomashow, M.F. Plant cold acclimation: freezing tolerance genes and regulatory mechanisms [J]. Annual review of plant biology, 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.T.; Wang, W.; Mao, B.; et al. Cold stress tolerance in rice: physiological changes, molecular mechanism, and future prospects. [J]. 2018, 40, 171–185. [Google Scholar]
- Jung, J.H.; Domijan, M.; Klose, C.; et al. Phytochromes function as thermosensors in Arabidopsis [J]. Science, 2016, 354, 886–889. [Google Scholar] [CrossRef]
- Legris, M.; Klose, C.; Burgie, E.S.; et al. Phytochrome B integrates light and temperature signals in Arabidopsis [J]. Science, 2016, 354, 897–900. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Kodama, Y. Refinements to light sources used to analyze the chloroplast cold-avoidance response over the past century [J]. Plant Signaling & Behavior, 2018, 13, 9206–211. [Google Scholar]
- Cook, D.; Fowler, S.; Fiehn, O.; et al. A prominent role for the CBF cold response pathway in configuring the low-temperature metabolome of Arabidopsis [J]. Proceedings of the National Academy of Sciences, 2004, 101, 15243–15248. [Google Scholar] [CrossRef] [PubMed]
- Hannah, M.A.; Heyer, A.G.; Hincha, D.K. A global survey of gene regulation during cold acclimation in Arabidopsis thaliana [J]. PLoS genetics, 2005, 1, e26. [Google Scholar] [CrossRef]
- Maruyama, K.; Takeda, M.; Kidokoro, S.; et al. Metabolic pathways involved in cold acclimation identified by integrated analysis of metabolites and transcripts regulated by DREB1A and DREB2A [J]. Plant physiology, 2009, 150, 1972–1980. [Google Scholar] [CrossRef]
- Nakashima, K.; Takasaki, H.; Mizoi, J.; et al. NAC transcription factors in plant abiotic stress responses [J]. Biochimica et Biophysica Acta (BBA)-Gene Regulatory Mechanisms, 2012, 1819, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P.; et al. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana [J]. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef]
- Hu, R.; Qi, G.; Kong, Y.; et al. Comprehensive analysis of NAC domain transcription factor gene family in Populus trichocarpa [J]. BMC plant biology, 2010, 10, 1–23. [Google Scholar] [CrossRef]
- Cao, F.; Guo, C.; Wang, X.; Wang, X.; Yu, L.; Zhang, H.; Zhang, J. Genome-wide identification, evolution, and expression analysis of the NAC gene family in chestnut (Castanea mollissima) [J]. Front. Genet. 2024, 15, 1337578. [Google Scholar] [CrossRef]
- Yang, Z.; An, Y.; Ye, Q.; Zhang, N.; Liu, X.; He, F.; Zeng, Y.; Tang, M.; Yang, Z.; Li, K. Genome-Wide Identification and Expression Analysis of Salt-Tolerance-Associated NAC Family Genes in Cyclocarya paliurus [J]. Forests 2024, 15, 479. [Google Scholar] [CrossRef]
- Ahmed, J.; Sajjad, Y.; Gatasheh, M.K.; Ibrahim, K.E.; Huzafa, M.; Khan, S.A.; Situ, C.; Abbasi, A.M.; Hassan, A. Genome-wide identification of NAC transcription factors and regulation of monoterpenoid indole alkaloid biosynthesis in Catharanthus roseus [J]. Front. Plant Sci. 2023, 14, 1286584. [Google Scholar] [CrossRef] [PubMed]
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; et al. NAC proteins: regulation and role in stress tolerance [J]. Trends in plant science, 2012, 17, 369–381. [Google Scholar] [CrossRef]
- Nakashima, K.; Takasaki, H.; Mizoi, J.; et al. NAC transcription factors in plant abiotic stress responses [J]. Biochimica et Biophysica Acta (BBA) - Gene Regulatory Mechanisms 2012, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Nuruzzaman, M.; Sharoni, A.M.; Kikuchi, S. Roles of NAC transcription factors in the regulation of biotic and abiotic stress responses in plants [J]. Frontiers in microbiology, 2013, 4, 248. [Google Scholar] [CrossRef] [PubMed]
- Uauy, C.; Distelfeld, A.; Fahima, T.; et al. A NAC gene regulating senescence improves grain protein, zinc, and iron content in wheat [J]. Science, 2006, 314, 1298–1301. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.S.P.; Nakashima, K.; Sakuma, Y.; et al. Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress 1 promoter [J]. The Plant Cell, 2004, 16, 2481–2498. [Google Scholar] [CrossRef] [PubMed]
- Bielefeld, E.C.; Kopke, R.D.; Jackson, R.L.; et al. Noise protection with N-acetyl-l-cysteine (NAC) using a variety of noise exposures, NAC doses, and routes of administration [J]. Acta oto-laryngologica, 2007, 127, 914–919. [Google Scholar] [CrossRef] [PubMed]
- Ernst, H.A.; Nina Olsen, A.; Skriver, K.; et al. Structure of the conserved domain of ANAC, a member of the NAC family of transcription factors [J]. EMBO reports, 2004, 5, 297–303. [Google Scholar] [CrossRef]
- Fang, Y.; You, J.; Xie, K.; et al. Systematic sequence analysis and identification of tissue-specific or stress-responsive genes of NAC transcription factor family in rice [J]. Molecular Genetics and Genomics, 2008, 280, 547–563. [Google Scholar] [CrossRef]
- Shao, H.; Wang, H.; Tang, X. NAC transcription factors in plant multiple abiotic stress responses: progress and prospects [J]. Frontiers in plant science, 2015, 6, 902. [Google Scholar] [CrossRef]
- Jensen, M.K.; Kjaersgaard, T.; Nielsen, M.M.; et al. The Arabidopsis thaliana NAC transcription factor family: structure–function relationships and determinants of ANAC019 stress signaling [J]. Biochemical Journal, 2010, 426, 183–196. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.; Qin, W.; Zhou, Z.; et al. Banana MaNAC1 activates secondary cell wall cellulose biosynthesis to enhance chilling resistance in fruit [J]. Plant Biotechnology Journal 2024, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, N.; Wen, D.; Yu, J.; Hong, J.; Wu, M.; Cheng, L.; Meng, S. Identification of Stress Responsive NAC Genes in Casuarina equisetifolia L.and Its Expression Analysis under Abiotic Stresses [J]. Agronomy, 2024, 14, 535. [Google Scholar]
- Xu, P.; Ma, W.; Feng, H.; Cai, W. NAC056 transcription factor confers freezing tolerance by positively regulating the expression of CBFs and NIA1 in Arabidopsis, PLANT COMMUNICATIONS, 2024.
- Bu, Q.; Jiang, H.; Li, C.B.; et al. Role of the Arabidopsis thaliana NAC transcription factors ANAC019 and ANAC055 in regulating jasmonic acid-signaled defense responses [J]. Cell research, 2008, 18, 756–767. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Richardson, E.A.; Ye, Z.H. Two NAC domain transcription factors, SND1 and NST1, function redundantly in regulation of secondary wall synthesis in fibers of Arabidopsis [J]. Planta, 2007, 225, 1603–1611. [Google Scholar] [CrossRef]
- Wang, X.; Basnayake, B.M.V.S.; Zhang, H.; et al. The Arabidopsis ATAF1, a NAC transcription factor, is a negative regulator of defense responses against necrotrophic fungal and bacterial pathogens [J]. Molecular Plant‒Microbe Interactions, 2009, 22, 1227–1238. [Google Scholar] [CrossRef] [PubMed]
- Jensen, M.K.; Hagedorn, P.H.; De Torres-Zabala, M.; et al. Transcriptional regulation by an NAC (NAM–ATAF1, 2–CUC2) transcription factor attenuates ABA signaling for efficient basal defense toward Blumeria graminis f. sp. hordei in Arabidopsis [J]. The Plant Journal, 2008, 56, 867–880. [Google Scholar] [CrossRef] [PubMed]
- Shan, W.; Kuang, J.F.; Lu, W.J.; Chen, J.Y. Banana fruit NAC transcription factor MaNAC1 is a direct target of MaICE1 and involved in cold stress through interacting with MaCBF1 [J]. Plant Cell Environ. 2014, 37, 2116–2127. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, Z.; Wu, Y.; Mu, M.; Jiang, J.; Nie, W.; Zhao, S.; Cui, G.; Yin, X. Genome-wide identification and characterization of NAC transcription factor family members in Trifolium pratense and expression analysis under lead stress [J]. BMC Genom. 2024, 25, 128. [Google Scholar] [CrossRef]
- Tao, W.; Xue, M.M.; Ying, C.; Cui, C.W.; Zhen, X.X.; Zixi, L.; Li, H.G.; Wen, N.Z. SlNAC3 suppresses cold tolerance in tomatoes by enhancing ethylene biosynthesis [J]. Plant, Cell & Environment, 2024.May (1).
- Hu, X.; Xie, F.; Liang, W.; Liang, Y.; Zhang, Z.; Zhao, J.; Hu, G.; Qin, Y. HuNAC20 and HuNAC25, Two Novel NAC Genes from Pitaya, Confer Cold Tolerance in Transgenic Arabidopsis [J]. Int. J. Mol. Sci. 2022, 23, 2189. [Google Scholar] [CrossRef]
- Bradrord, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding [J]. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hao, Y.-Q.; Lu, G.-Q.; Wang, L.-H.; Wang, C.-L.; Guo, H.-M.; Li, Y.-F.; Cheng, H.-M. Overexpression of AmDUF1517 enhanced tolerance to salinity, drought, and cold stress in transgenic cotton [J]. Integr. Agric. 2018, 17, 2204–2214. [Google Scholar] [CrossRef]
- Liu, S.; Guan, Y.; Weng, Y.; et al. Genome-wide identification of the NAC gene family and its functional analysis in Liriodendron [J]. BMC Plant Biol. 2023, 23, 415. [Google Scholar] [CrossRef]
- Huang, X.; Shi, H.; Hu, Z.; et al. ABA is involved in regulation of cold stress response in bermudagrass [J]. Frontiers in plant science, 2017, 8, 1613. [Google Scholar] [CrossRef]
- He, Z.; Zhan, Y.; Zeng, F.; et al. Drought physiology and gene expression characteristics of Fraxinus interspecific hybrids [J]. Plant growth regulation, 2016, 78, 179–193. [Google Scholar] [CrossRef]
- Bouslama, M.; Schapaugh, W.T., Jr. Stress tolerance in soybeans. I. Evaluation of three screening techniques for heat and drought tolerance 1 [J]. Crop science, 1984, 24, 933–937. [Google Scholar]
- Ni, Z.; Kim, E.D.; Ha, M.; et al. Altered circadian rhythms regulate growth vigor in hybrids and allopolyploids [J]. Nature, 2009, 457, 327–331. [Google Scholar] [CrossRef]
- Shi, Y.; Gong, Z. One SNP in COLD1 determines cold tolerance during rice domestication [J]. J Genet Genomics, 2015, 42, 133–134. [Google Scholar] [CrossRef] [PubMed]
- Yue, C.; Cao, H.L.; Wang, L.; et al. Effects of cold acclimation on sugar metabolism and sugar-related gene expression in tea plant during the winter season [J]. Plant molecular biology, 2015, 88, 591–608. [Google Scholar] [CrossRef]
- Heidarvand, L.; Maali-Amiri, R. Physio-biochemical and proteome analysis of chickpea in early phases of cold stress [J]. Journal of plant physiology, 2013, 170, 459–469. [Google Scholar] [CrossRef]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.O.N.; et al. ROS and redox signaling in the response of plants to abiotic stress [J]. Plant, cell & environment, 2012, 35, 259–270. [Google Scholar]
- Yarra, R.; Wei, W. The NAC-type transcription factor GmNAC20 improves cold, salinity tolerance, and lateral root formation in transgenic rice plants [J]. Functional & Integrative Genomics, 2021, 21, 473–487. [Google Scholar] [CrossRef] [PubMed]
- Shan, W.; Kuang, J.; Lu, W.; et al. Banana fruit NAC transcription factor MaNAC1 is a direct target of MaICE1 and involved in cold stress through interacting with MaCBF1 [J]. Plant, Cell & Environment 2014, 2116–2127. [Google Scholar]
- Jin, C.; Li, K.Q.; Xu, X.Y.; et al. A Novel NAC Transcription Factor, PbeNAC1, of Pyrus betulifolia Confers Cold and Drought Tolerance via Interacting with PbeDREBs and Activating the Expression of Stress-Responsive Genes [J]. Frontiers in Plant Science 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Achard, P.; Gong, F.; Cheminant, S.; et al. The cold-inducible CBF1 factor–dependent signaling pathway modulates the accumulation of the growth-repressing DELLA proteins via its effect on gibberellin metabolism [J]. The Plant Cell, 2008, 20, 2117–2129. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.J.; Wei, W.; Song, Q.X.; et al. Soybean NAC transcription factors promote abiotic stress tolerance and lateral root formation in transgenic plants [J]. Plant Journal for Cell & Molecular Biology, 2011, 68, 302–313. [Google Scholar]
- He, Z.; Zhan, Y.; Zeng, F.; et al. Drought physiology and gene expression characteristics of Fraxinus interspecific hybrids [J]. Plant growth regulation, 2016, 78, 179–193. [Google Scholar] [CrossRef]
- Bouslama, M.; Schapaugh, W.T., Jr. Stress tolerance in soybeans. I. Evaluation of three screening techniques for heat and drought tolerance 1 [J]. Crop science, 1984, 24, 933–937. [Google Scholar]
- Ni, Z.; Kim, E.D.; Ha, M.; et al. Altered circadian rhythms regulate growth vigor in hybrids and allopolyploids [J]. Nature, 2009, 457, 327–331. [Google Scholar] [CrossRef]
- Shi, Y.; Gong, Z. One SNP in COLD1 determines cold tolerance during rice domestication [J]. J Genet Genomics, 2015, 42, 133–134. [Google Scholar] [CrossRef]
- Diao, P.; Chen, C.; Zhang, Y.; et al. The role of NAC transcription factor in plant cold response [J]. Plant Signaling & Behavior, 2020, 15, 1785668. [Google Scholar]
- Gao, S.; Li, C.; Chen, X.; et al. Basic helix-loop-helix transcription factor PxbHLH02 enhances drought tolerance in Populus (Populus simonii× P. nigra) [J]. Tree Physiology, 2023, 43, 185–202. [Google Scholar] [CrossRef]
- CLARKE, J.D. Cetyltrimethyl Ammonium Bromide (CTAB) DNA Miniprep for Plant DNA Isolation [J]. Cold Spring Harbor Protocols 2009, pdb.prot5177. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Xue, W.; Li, X.; et al. Mitotically heritable epigenetic modifications of CmMYB6 control anthocyanin biosynthesis in Chrysanthemum. [J]. New Phytologist 2022, 1075–1088. [Google Scholar] [CrossRef] [PubMed]
- Nan S L, Ya G Z, YU, et al. Characteristics and Expression Analysis of FmTCP15 under Abiotic Stresses and Hormones and Interact with DELLA Protein in Fraxinus mandshurica Rupr. Forests 2019, 10, 343.
- Kahrizi, D.; Ghaheri, M.; Yari, Z.; et al. Investigation of different concentrations of MS media effects on gene expression and steviol glycosides accumulation in Stevia rebaudiana Bertoni [J]. Cellular and Molecular Biology, 2018, 64, 23–27. [Google Scholar] [CrossRef]
Figure 1.
Sequence analysis of FmNAC1 genes and prediction of the protein tertiary structure. (A) The FmNAC1 gene and its encoded amino acid sequence. (B) The tertiary structure of the FmNAC1 protein. (C) The tertiary structure of the conserved domain of the Oryza sativa L. stress-responsive NAC1 protein. (D) The different regions, identified with different colors, on the amino acid sequences of FmNAC1and OsNAC1.
Figure 1.
Sequence analysis of FmNAC1 genes and prediction of the protein tertiary structure. (A) The FmNAC1 gene and its encoded amino acid sequence. (B) The tertiary structure of the FmNAC1 protein. (C) The tertiary structure of the conserved domain of the Oryza sativa L. stress-responsive NAC1 protein. (D) The different regions, identified with different colors, on the amino acid sequences of FmNAC1and OsNAC1.
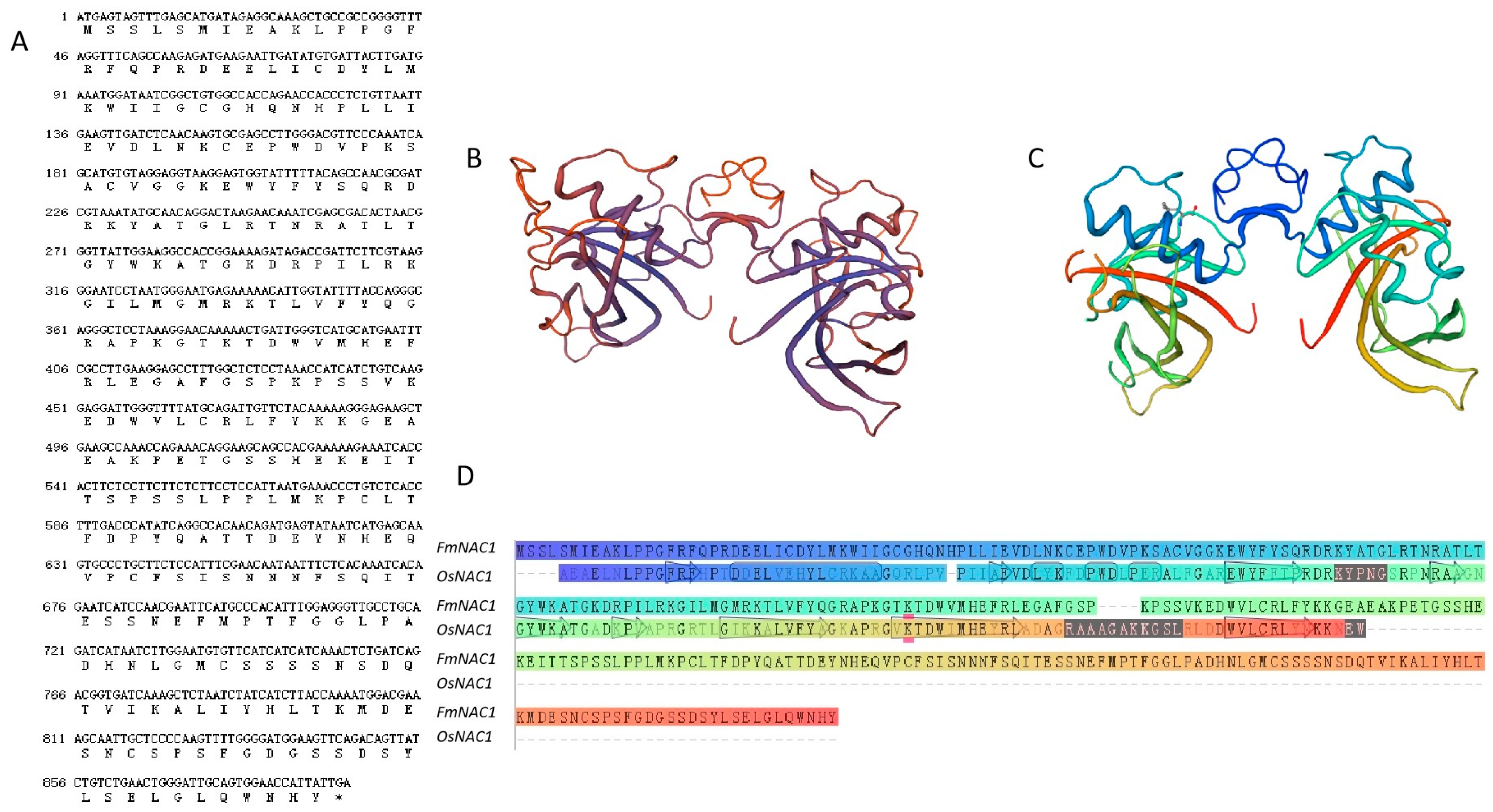
Figure 2.
Multiple sequence alignment and phylogenetic relationships of the FmNAC1 protein. (A) Homolog alignment of the FmNAC1 protein. The red box represents the NAC conserved domain. (B) Construction of the FmNAC1 phylogenetic tree. The phylogenetic relationships of the FmNAC1 proteins were investigated by obtaining the NAC1 protein sequences from 30 plant species from the NCBI database (http://www.ncbi.nlm.nih.gov/) using the BLASTP method. Sequence alignment was performed using the ClustalW method, and a phylogenetic tree was constructed using the neighbor-joining method with MEGA 5.0 software. In the constructed phylogenetic tree, FmNAC1 is represented by a red triangle. (Different colors indicate different clades, with distinct phyla marked on the outside). Abrus precatorius (XP_027343628.1), Beta vulgaris subsp. (XP_010666181.1), Cajanus cajan (XP_020223961.1), Camellia sinensis (XP_028081260.1), Carya illinoinensis (XP_042951768.1), Cicer arietinum (XP_004488843.1), Coffea arabica (XP_027061236.1), Fraxinus mandshurica (AXO66323.1), Glycine soja (XP_028244150.1), Herrania umbratica (XP_021288005.1), Hevea brasiliensis (XP_021644348.1), Jatropha curcas (XP_012072721.1), Juglans microcarpa x Juglans regia (XP_040989265.1), Juglans regia (XP_018838950.1), Lupinus angustifolius (XP_019443758.1), Manihot esculenta (XP_021606973.1), Medicago truncatula (XP_003595973.1), Nelumbo nucifera (XP_010258675.1), Nicotiana tomentosiformis (XP_009605814.1), Olea europaea var. (XP_022848339.1), Phaseolus vulgaris (XP_007149262.1), Populus alba (XP_034910133.1), Populus euphratica (XP_011012784.1), Populus trichocarpa (XP_002310688.1), Prunus avium (XP_021832498.1), Pyrus x bretschneideri (XP_009336675.1), Quercus lobata (XP_030961789.1), Ricinus communis (XP_002529954.1), Sesamum indicum (XP_011095676.1), Theobroma cacao (XP_007022298.1), Vigna angularis (XP_017423259.1).
Figure 2.
Multiple sequence alignment and phylogenetic relationships of the FmNAC1 protein. (A) Homolog alignment of the FmNAC1 protein. The red box represents the NAC conserved domain. (B) Construction of the FmNAC1 phylogenetic tree. The phylogenetic relationships of the FmNAC1 proteins were investigated by obtaining the NAC1 protein sequences from 30 plant species from the NCBI database (http://www.ncbi.nlm.nih.gov/) using the BLASTP method. Sequence alignment was performed using the ClustalW method, and a phylogenetic tree was constructed using the neighbor-joining method with MEGA 5.0 software. In the constructed phylogenetic tree, FmNAC1 is represented by a red triangle. (Different colors indicate different clades, with distinct phyla marked on the outside). Abrus precatorius (XP_027343628.1), Beta vulgaris subsp. (XP_010666181.1), Cajanus cajan (XP_020223961.1), Camellia sinensis (XP_028081260.1), Carya illinoinensis (XP_042951768.1), Cicer arietinum (XP_004488843.1), Coffea arabica (XP_027061236.1), Fraxinus mandshurica (AXO66323.1), Glycine soja (XP_028244150.1), Herrania umbratica (XP_021288005.1), Hevea brasiliensis (XP_021644348.1), Jatropha curcas (XP_012072721.1), Juglans microcarpa x Juglans regia (XP_040989265.1), Juglans regia (XP_018838950.1), Lupinus angustifolius (XP_019443758.1), Manihot esculenta (XP_021606973.1), Medicago truncatula (XP_003595973.1), Nelumbo nucifera (XP_010258675.1), Nicotiana tomentosiformis (XP_009605814.1), Olea europaea var. (XP_022848339.1), Phaseolus vulgaris (XP_007149262.1), Populus alba (XP_034910133.1), Populus euphratica (XP_011012784.1), Populus trichocarpa (XP_002310688.1), Prunus avium (XP_021832498.1), Pyrus x bretschneideri (XP_009336675.1), Quercus lobata (XP_030961789.1), Ricinus communis (XP_002529954.1), Sesamum indicum (XP_011095676.1), Theobroma cacao (XP_007022298.1), Vigna angularis (XP_017423259.1).
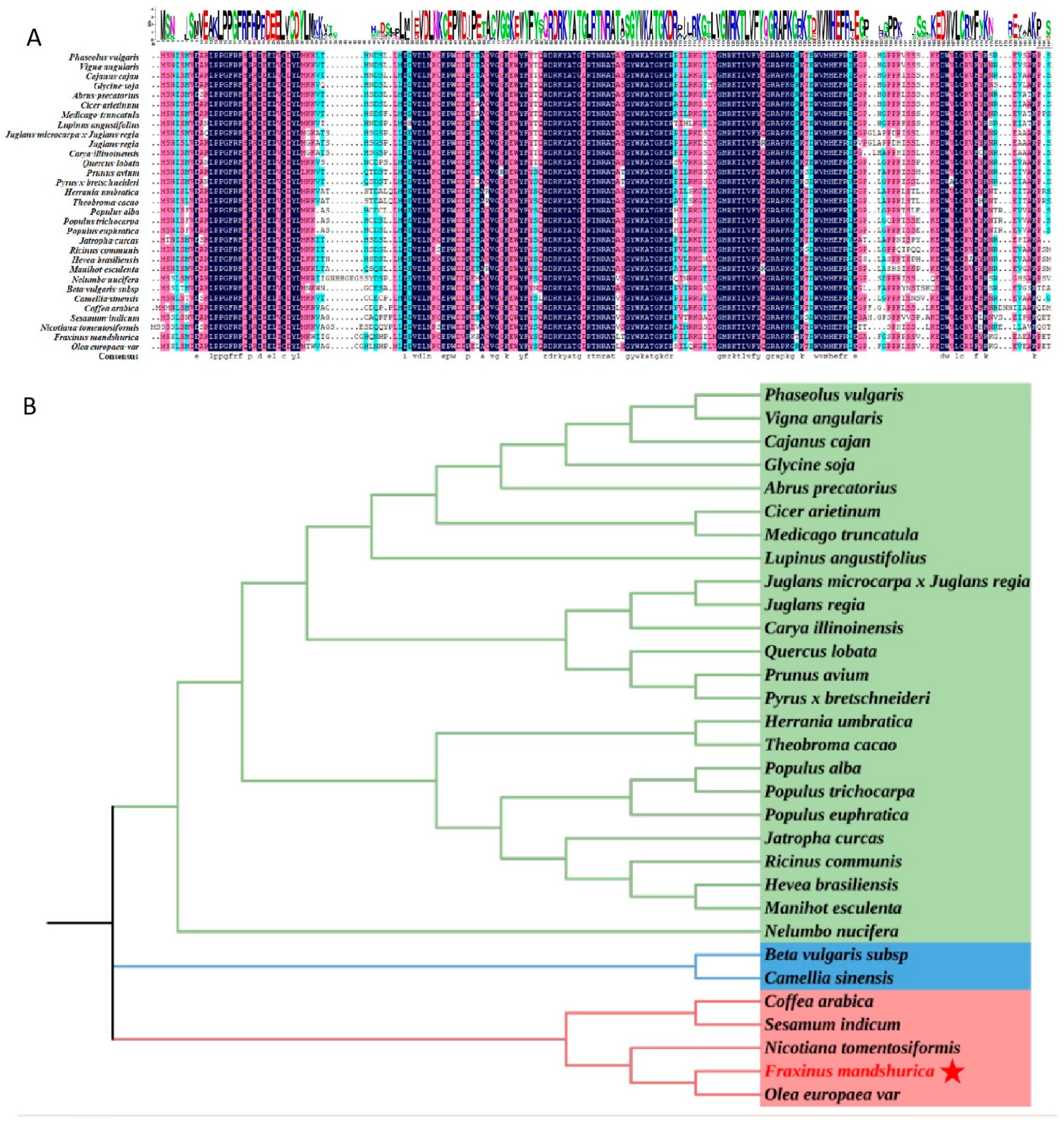
Figure 3.
Subcellular localization of FmNAC1. The photographs were taken under dark-field illumination for green fluorescence localization (GFP), bright-field illumination to examine the cell morphology (bright field), and merged-field illumination (merged). The bar represents 10 μm. The red arrows indicate that green fluorescence occurred in the nucleus.
Figure 3.
Subcellular localization of FmNAC1. The photographs were taken under dark-field illumination for green fluorescence localization (GFP), bright-field illumination to examine the cell morphology (bright field), and merged-field illumination (merged). The bar represents 10 μm. The red arrows indicate that green fluorescence occurred in the nucleus.

Figure 4.
The expression of the FmNAC1 gene fluctuated with the duration of the biological stress. Note: The horizontal axes represent the cold, salt, ABA, GA, and IAA treatments, and the vertical axes represent 1 hour, 3 hours, 6 hours, 12 hours, 24 hours, and 48 hours of treatment.
Figure 4.
The expression of the FmNAC1 gene fluctuated with the duration of the biological stress. Note: The horizontal axes represent the cold, salt, ABA, GA, and IAA treatments, and the vertical axes represent 1 hour, 3 hours, 6 hours, 12 hours, 24 hours, and 48 hours of treatment.
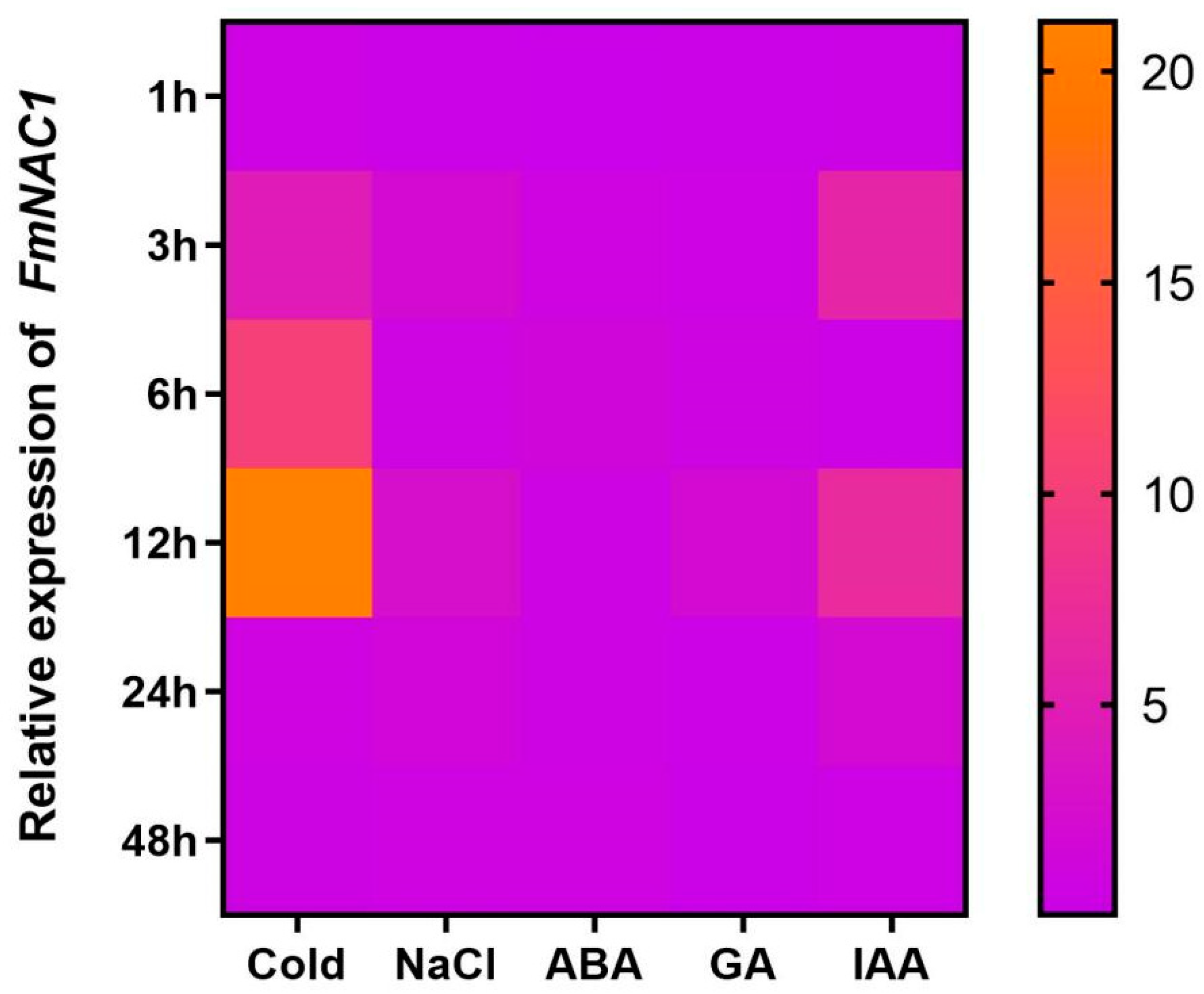
Figure 5.
Transgenic tobacco from FmNAC1-OE plants was obtained. Note: the above figure represents the growth status of transgenic tobacco plants cultured for 1 day, 7 days, 10 days, 15 days, 28 days, 45 days, 70 days, and 45 days after successive generations; the plants were finally transplanted into the greenhouse.
Figure 5.
Transgenic tobacco from FmNAC1-OE plants was obtained. Note: the above figure represents the growth status of transgenic tobacco plants cultured for 1 day, 7 days, 10 days, 15 days, 28 days, 45 days, 70 days, and 45 days after successive generations; the plants were finally transplanted into the greenhouse.

Figure 6.
Verification of the transgenic tobacco strain overexpressing FmNAC1. Note: (A) The columns are arranged as follows: the first column represents the marker, the second column represents the plasmid, the third column represents the water control, columns four to ten represent tobacco lines overexpressing FmNAC1, and the eleventh column represents the wild-type group. (B) The gene expression of FmNAC1 in transgenic tobacco. The first column represents the wild-type group, followed by columns for samples 1, 2, 5, and 7.
Figure 6.
Verification of the transgenic tobacco strain overexpressing FmNAC1. Note: (A) The columns are arranged as follows: the first column represents the marker, the second column represents the plasmid, the third column represents the water control, columns four to ten represent tobacco lines overexpressing FmNAC1, and the eleventh column represents the wild-type group. (B) The gene expression of FmNAC1 in transgenic tobacco. The first column represents the wild-type group, followed by columns for samples 1, 2, 5, and 7.
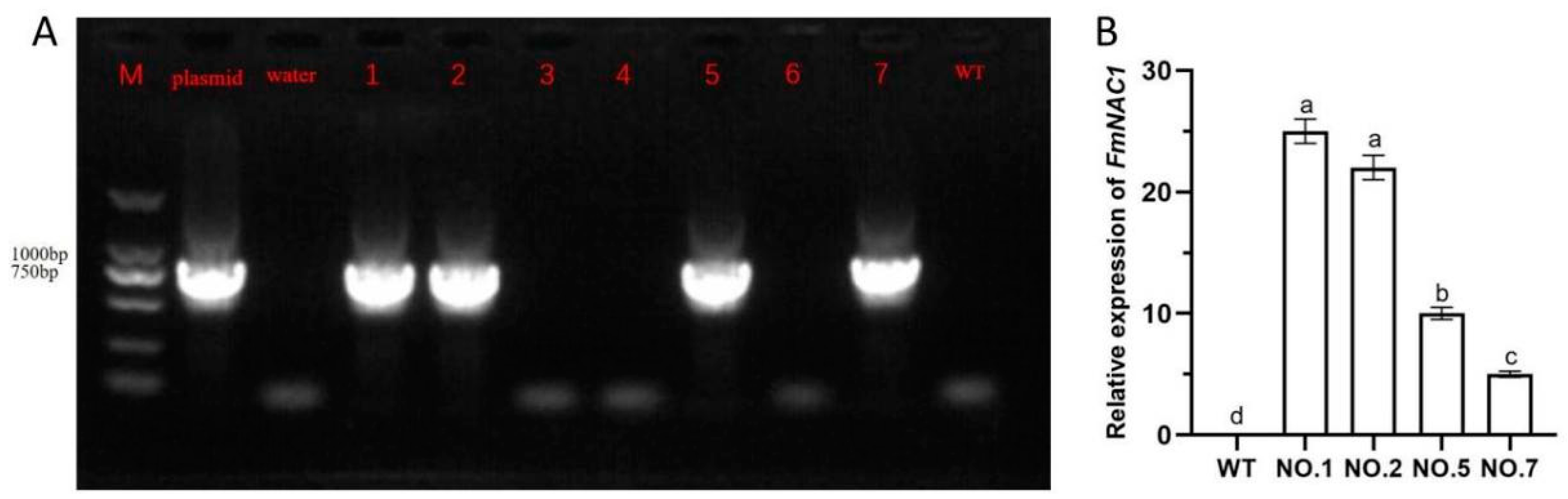
Figure 7.
Changes in the height of the FmNAC1 transgenic tobacco plants after transplanting. (A) Experimental plants, with a white arrow indicating the overexpression of transgenic tobacco, while the other three pots represent wild-type controls from the same time period. The height of the transgenic tobacco plants was measured one day after transplantation. (B) The x-axis represents time, the y-axis represents height, the black line represents the wild type, the red line represents transgenic line 1, the green line represents transgenic line 2, and the blue line represents transgenic line 5. * Indicates a significant difference between the control and transgenic tobacco (p < 0.05).
Figure 7.
Changes in the height of the FmNAC1 transgenic tobacco plants after transplanting. (A) Experimental plants, with a white arrow indicating the overexpression of transgenic tobacco, while the other three pots represent wild-type controls from the same time period. The height of the transgenic tobacco plants was measured one day after transplantation. (B) The x-axis represents time, the y-axis represents height, the black line represents the wild type, the red line represents transgenic line 1, the green line represents transgenic line 2, and the blue line represents transgenic line 5. * Indicates a significant difference between the control and transgenic tobacco (p < 0.05).

Figure 8.
Phenotypic changes in the FmNAC1 transgenic tobacco gene under cold stress. Note: from left to right, transgenic line 1, transgenic line 2, transgenic line 5, and the wild type.
Figure 8.
Phenotypic changes in the FmNAC1 transgenic tobacco gene under cold stress. Note: from left to right, transgenic line 1, transgenic line 2, transgenic line 5, and the wild type.

Figure 11.
The vector map of the FmNAC1 gene overexpression construct. (A) Vector map of the PBI121-FmNAC1-GFP overexpression construct. (B) Vector map of the pROKII-FmNAC1-GUS overexpression construct.
Figure 11.
The vector map of the FmNAC1 gene overexpression construct. (A) Vector map of the PBI121-FmNAC1-GFP overexpression construct. (B) Vector map of the pROKII-FmNAC1-GUS overexpression construct.

Table 1.
The primers corresponding to the coding region of the cloned FmNAC1 gene.
| Gene | GenBank accession numbers | Primer name | Primer sequences | Melting temperatures |
|---|---|---|---|---|
| FmNAC1 | MG269829.1 | FmNAC1-F | 5'-TCTATCACCTCCGCCATC -3' | 53.1℃ |
| FmNAC1-R | 5'-TCAATAATGGTTCCACTGC-3' | 52.3℃ |
Table 2.
Sequences of the primers used for the qRT-PCR.
| Gene | GenBank accession numbers | Primer Name | Primer Sequence | Melting temperatures |
|---|---|---|---|---|
| NbIAA | XM_016592822.1 | q NbIAA -F | 5' GCCTCCTTCCACATTCCC 3' | 57℃ |
| q NbIAA -R | 5' AGTCCACGTTTCGCACCA 3' | 57.4℃ | ||
| NbAUX1 | NM_001326023.1 | q NbAUX1-F | 5' AGGCTACTGCCCTACCAC 3' | 52℃ |
| qNbAUX1-R | 5' CAGCAACTGTATTCCCACA 3' | 51.8℃ | ||
| NbICE | XM_016610394.1 | q NbICE-F | 5' CCCATCATTCCATTTGCT 3' | 53.3℃ |
| q NbICE-R | 5' CCATTATCCCACTTCCTCT 3' | 51.1℃ | ||
| NbDREB | NM_001325812.1 | q NbDREB-F | 5' GGCTTGGCACTTTCCCTT 3' | 57.3℃ |
| qNbDREB-R | 5' AATAGCGCCTCCTCATCC 3' | 54.3℃ | ||
| NbCBF | NM_001325227.1 | q NbCBF-F | 5' GGGGAATAAGGAGGAGAA 3' | 51℃ |
| q NbCBF-R | 5' CTGAAGTCGGGATGGGTA 3' | 53.2℃ | ||
| NbCAT | NM_001325412.1 | q NbCAT-F | 5' AATCATAGCCACGCCACC 3' | 56.2℃ |
| q NbCAT-R | 5' CGAATAGTAAAGACCAGGGA 3' | 52.2℃ | ||
| NbSOD | XM_016581519.1 | q NbSOD -F | 5' GATCTGGGAAGAGGTGGA 3' | 51.7℃ |
| q NbSOD -R | 5' CAGCCCTAATGATAAACTGA 3' | 50.5℃ | ||
| NbTU | XM_016640346.1 | qNbTU-F | 5'ATGTTGTTAGGAAGGAGGCT3' | 53.2℃ |
| qNbTU-R | 5' ATCATGCGGTCAGGGTAT 3' | 52.6℃ | ||
| FmNAC1 | MG269829.1 | qFmNAC1-F | 5' AATGGATAATCGGCTGTG 3' | 50.9℃ |
| qFmNAC1-R | 5' CCTTACGAAGAATCGGTCTA 3' | 52.4℃ | ||
| FmTU | qPT-PCR | qFmTU-F | 5' AGGACGCTGCCAACAACTTT 3' | 59.8℃ |
| qFmTU-R | 5' TTGAGGGGAAGGGTAAATAGTG 3' | 58.0℃ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



