
Submitted:
21 May 2024
Posted:
23 May 2024
You are already at the latest version
Abstract
Candida auris is considered an emerging Candida species that has rapidly spread all over the world. The evidence of its origin and emerging resistance is still unclear. The severe infection by this species causes significant mortality and morbidity among the elderly and immune compressed individuals. Development of drug resistance is the major factor associated with therapeutic failure of existing antifungal agents. Previous studies addressed the antifungal resistance profile and drug discovery for C. auris. However, comprehensive coverage of these information in a single investigation yet to be available. In this review, we have mainly focused on recent development in therapeutic strategy against C. auris. Based on the available information, several differential approaches were discussed, including existing antifungal drugs, chemical compounds, essential oils, natural products, antifungal peptides, immunotherapy, antimicrobial photodynamic therapy, drug repurposing, and drug delivery systems. Among them, chemical medications, natural products, and antifungal peptides are the prime contributors. However, a limited number of resources are available to prove the efficiency of these potential therapies in clinical usage. Hence, we hope that the data gathered in this review can encourage in vivo studies and clinical trials.
Keywords:
Candida auris
; antifungal therapy
; antifungal natural products
; antifungal peptides
; antifungal essential oil
1. Introduction
1.1. Candidiasis, Outbreak, and Epidemiology
Candidiasis is an infection caused by opportunistic pathogens of Candida genus, with manifestations varying from mucocutaneous lesions to life threading bloodstream infections. Candida albicans is the most common Candida species found in various human anatomical sites, including oropharyngeal, esophageal, gastrointestinal, and genital mucosa. Other non albicans species and Candida related species are also simultaneously reported in human body, such as Nakaseomyces glabratus (formerly Candida glabrata), Pichia kudriavzevii (formerly Candida krusei), Candida parapsilosis, Candida tropicalis, etc. Recently, an emerging pathogen named Candida auris has been isolated from different clinical samples like urine, stool, vaginal and rectal swabs. Like other Candida species, patients with comorbidities or weakened immune system, previously exposure to antifungals and subjected to long-stay in healthcare settings, are at risk for C. auris infection [1,2].
C. auris is recognised as an emerging fungal pathogen because of its wide distribution, multidrug-resistant (MDR) behaviour, high transmissibility, strong association with nosocomial infections and high mortality rates. C. auris expresses numerous virulence traits as well as tolerance to common antifungals. This leads to development of therapeutic failure when tried to treat with most common class of antifungals that includes azoles, polyene and echinocandins. In 2009, the first isolate of C. auris was reported from Japanese female patient with ear discharge. In the same year, twelve isolates were obtained from otitis media patients in South Korea. During time frame between 2009 to 2011, twelve isolates of C. auris had identified in India in patients with bloodstream infection. The first outbreak was noted during 2016 and 2017 in Europe and USA respectively. Subsequently, C. auris was reported in more than 47 countries according to Centre for Disease Control (CDC) [3,4], and nowadays it is recognised as a threat to world community by World Health Organisation (WHO). Based on susceptibility profiles, outbreak potential and clinical manifestations, the C. auris isolates were broadly classified into five clades (Figure 1). Recently, an unpublished data indicated a sixth clade of C. auris isolated from the samples collect in Singapore. These uncommon Clade VI isolates were entirely different from all others isolates in relation to antifungal resistance genes, mating type locus, and chromosomal rearrangements. Although C. auris isolates have widely investigated, the real rate of prevalence remains uncertain because of availability of proper dataset [5].
Furthermore, the origin of C. auris has not yet been elucidated; and some authors speculates that global warming may be a possible reason for its spread [6]. C. auris cells remain viable for several month on environmental surfaces and medical equipment and prefer to colonize the skin of patients rather than other mucosal surfaces, leading to high probability of person-to-person transmission [7]. Since they freely live on the biotic and abiotic surfaces and tolerate antifungal and decontamination agents, the eradication of these fungal cells became extremely complicated [8,9,10,11]. All these factors make C. auris a global threat for the immunocompromised patients in healthcare settings.
1.2. Drug Resistance: Molecular Bases
Clinical breakpoints of existing antifungals for C. auris yet to be established; however, CDC has recommended clinical breakpoints established from closely related other Candida species. The general guideline defined to correlate the antifungal resistance of C. auris are ≥32, ≥2, ≥4, ≥2 and ≥4 for fluconazole, Amphotericin B (Amp B), anidulafungin, caspofungin and micafungin respectively (CDC, 2020). Most clinical isolates of C. auris are multidrug resistance or pan-resistant, especially for resistance to fluconazole followed by Amp B and voriconazole[12]. Wide administration of fluconazole for the Candida prophylaxis and the use of azole derivatives in various purposes, such as agriculture area, might be the prime factor for azole resistance. Thus, echinocandins are the only choice for C. auris infection treatment, although some studies already reported echinocandin-resistant strains [13].
Prevalence of drug-resistant C. auris strains were analysed in several studies worldwide. Isolates from India, Pakistan, South Africa, and Venezuela presented 93% of resistance for fluconazole, 35% for Amp B, and 7% for echinocandins [2]. A resistance rate of 90% for fluconazole was observed for isolates from India and United States, whereas all the isolates from UK showed resistance to fluconazole (100%). Amp B was the second most common antifungal related to C. auris resistance, with 20-30% of resistant isolates in India [14], 30% in US [15], 62.5% in Saudi Arabia [16], 23.2% in Kuwait [17] and 33.3% in Oman [18].
Besides prevalence analysis, some studies investigated the molecular bases of drug resistance in C. auris isolates. Different mechanisms of azole resistance have been discussed, including mutation in target genes (ERG11) [19], overexpression of genes encoding drug transporters and increased copy number of the TAC1B gene [20]. Mutation in TAC1B gene was the second most observed mutation in C. auris after ERG11. The mutations like F126L, Y132F, and K143R were frequently noted in ERG11 gene of fluconazole resistant isolates, whereas A640V, A657V, and F862_N866del mutation were observed in TAC1B gene of fluconazole resistant isolates [20,21].
Mechanisms of resistance to echinocandins are mainly associated with FKS1 gene involved in biosynthesis of glucan and maintain the integrity of cell wall. As like C. albicans and C. glabrata, FKS1 is the target of echinocandins and mutations like amino acid substitution and deletion in hot spots of FKS1 results in therapeutic loss [22,23]. The most frequently observed FKS1 mutations are S639F, S639P, and S639Y in hot spot 1; however, less common mutations in hot spot 1 were also reported (F635el, F635L/Y, S639T, D642Y) [24,25,26].
Regarding Amp B, the exact resistance mechanisms were not yet elucidated. The elevated MIC levels observed in Amp B-resistant C. auris isolates were associated with mutation in ERG6 gene [24]. Due to loss of sterol methyltransferase activity, these isolates accumulate more cholestatype sterols [21]. Similarly, in vitro studies evidenced that Amp B-resistant strains bearing nonsense mutations in genes like ERG11 and ERG3 [19].
2. Therapeutic Options
2.1. Current Approved/Considered Antifungal Therapy
The antifungal pipeline for C. auris infections includes different promising antifungal candidates, such as Rezafungin, Ibrexafungerp, Fosmanogepix and T-2307 (Figure 2). Rezafungin is a potential antifungal derived from anidulafungin that exhibited strong in vitro activity against C. auris, including strains resistant to other echinocandins. It boasts a long half-life allowing for once-weekly dosing schedule. In clinical trials, rezafungin has demonstrated a favourable safety profile with minimal side effects. On March 31, 2023, Rezzayo™ (rezafungin for injection) was approved in the US for use in adults with candidemia and invasive candidiasis. Nevertheless, it is still crucial to confirm its efficiency through realtime clinical data. Considering the cost, its more expensive than other antifungals, which could pose an access barrier in most of the clinical setting.
Ibrexafungerp (IBX) was focused by the researchers because of the superior antifungal behaviour. It is a class of (1,3) β-d-glucan synthase inhibitors like echinocandins, a semi-synthetic derivative of enfumafungin. The mode of binding of IBX was not disturbed by FKS mutations, also its mode of binding was different than echinocandins which limited cross-resistance. During the preclinical investigations, it was confirmed its potential as a therapeutic agent for managing highly resistant Candida infections. IBX remined active against fluconazole and echinocandin resistant Candida isolates and multidrug-resistant C. auris. However, IBX expressed concentration-dependent fungicidal activity on different clinically important fungi, including C. auris [28,29,30,31].
Fosmanogepix (known as APX001) is also a promising antifungal agent exhibiting potential to treat C. auris infections. While not yet commercially available, clinical trials provided encouraging results regarding its efficacy and safety against this multidrug-resistant fungus. Fosmanogepix demonstrated strong activity against C. auris strains in laboratory tests, including those resistant to standard antifungals like echinocandins. Phase II trials evaluating fosmanogepix for C. auris infections showed high treatment success rates. A study published in 2022 reported 80% success in clearing Candida from blood cultures with patient survival at the end of treatment. Apart from C. auris, fosmanogepix exhibited a broad spectrum of activity against other Candida species and molds, making it a valuable tool for wider fungal infections. Clinical trials indicated that fosmanogepix was well-tolerated with minimal side effects. Serious adverse events or treatment discontinuations were not reported. Fosmanogepix is available in both intravenous and oral forms, offering flexibility in treatment administration based on patient needs and disease severity [32]. Multiple Phase III clinical trials are currently investigating fosmanogepix for different fungal infections, including invasive candidiasis caused by C. auris. These trials will further evaluate its efficacy and safety on a larger patient population. Fosmanogepix is not yet approved for clinical use in any country. While the promising results from Phase II trials are encouraging, further research and evaluation through Phase III trials are still required [33].
Another novel antifungal agent is T-2307 that belongs to the triterpenoid class. As like other Candida species, C. auris was also sensitive to T-2307. In addition, it exhibited potent in vitro activity against a broad spectrum of fungal pathogens. Its mechanism of action was associated with inhibition of fungal cell wall biosynthesis [34,35,36]. Phase I clinical trial evaluating the safety and tolerability of T-2307 in healthy volunteers was recently completed, showing promising results. Phase II clinical trials to assess its efficacy in patients with fungal infections are planned to begin in 2024. Considering its promising preclinical data and ongoing clinical trials, T-2307 could be approved for clinical use against C. auris infections within the next few years.
2.2. Chemicals as an Emerging Weapon Against C. auris
Whilst some antifungal agents are in clinical trials phase, the scientific community has worked intensely to discover other therapeutic options. Here several new compounds with activity against C. auris are listed (Table 1) and discussed. Toepfer and team investigated the compound clorgyline and its derivatives, which worked as multi-target inhibitors of Cdr1 and Mdr1 efflux pumps of C. albicans and C. glabrata. Especially, clorgyline analogs M19 and M25 expressed high ability to inhibit the efflux pump activity of C. auris [37]. Recent investigations have discovered the active of pyrazole moiety compounds against fluconazole-resistant C. auris isolates. These compounds exhibited a broad spectrum, high potency, high selectivity, low cytotoxicity and anti-drug resistance [38]. Novel tetrazoles featuring isoxazole moiety were also identified as highly selective antifungal agents, displaying outstanding antifungal activity against fluconazole-resistant strains of C. albicans, C. glabrata and C. auris [39]. A novel benzoanilide antifungal (compound A1) showed potent activity against C. auris cells through blocking of virulence biosynthesis and alterations in cell wall by inhibition of glycosylphosphatidylinositol (GPI) and GPI-anchored proteins [40]. Furthermore, a compound named NSC319726 (thiosemicarbazone zinc chelator) exhibited MIC values ranging from 0.125 to 0.25 mg/L for C. auris isolates belonging to five different clades, expressing fungistatic activity in time-kill curves [41].
Lohse and their colleague have aimed to develop antifungal metabolites from a group of FDA (Food and Drug Administration) approved compounds. The selection of these compounds was based on MIC values of <10 µM. Among the hydroxyquinolines tested, clioquinol was more active than others, however the authors were unable to guaranteed their mechanisms of action [42]. In another study, hydroxyquinoline derivative known as nitroxoline, was also active against 35 isolates of C. auris with the MIC range of 0.125 to 1 μg/mL. The resultant MIC values were lesser than the activity of fluconazole and Amp B. At last, nitroxoline was recommended for the treatment of C. auris mediated candiduria. However, in vivo and clinical efficiency remains questionable [43].
Many authors focused on the study of Manogepix, a pyridine-isoxazole-based antifungal agent. Manogepix was active against most clinical isolates of C. auris belonging to South Africa. Over 300 C. auris isolates were studied, including 335 fluconazole resistant, 19 fluconazole and Amp B resistant, 1 Amp B resistant and 2 pan resistant. MIC values of manogepix ranged from 0.002 to 0.063 μg/mL for fluconazole resistant isolates, 0.004 to 0.031 μg/mL for fluconazole and Amp B resistant isolates, and 0.004 μg/mL and 0.008 μg/mL for pan-resistant isolates. The activity of manogepix was more than 3 folds than azoles, 4 folds than echinocandins, and 9 folds than Amp B [44]. Manogepix also showed activity against clinical isolates from the New York Outbreak with MIC values of 0.008 to 0.015 mg/L against pan-resistant isolates [45]. Now, some studies have addressed the efficiency of manogepix with anidulafungin against C. auris [46].
Various studies have reported the biological role of metallic gold or its salt. Among them, gold(I)−phosphine complexes and gold salt auranofin were tested against a panel of 28 fungal strains including Candida spp., Cryptococcus spp., Aspergillus spp., and Fusarium spp. Notably, two (complex 4 and 6) square-planar gold(I) complexes produced a remarkable antifungal activity in most of the tested isolates. In relation to C. auris isolates, complex 4 and 6 resulted in MIC ranges between 3.9 to 7.8 and 1.95 µg/mL respectively. However, auranofin did not produce considerable results (MIC >31.3 µg/mL) in Candida species [47].
The activity of new promising antifungal compounds has also investigated against C. auris in biofilm stage. Among several phenylthiazole small molecules, compound 1 emerged as the most potent antifungal, inhibiting the growth of C. albicans and C. auris strains at concentrations ranging from 0.25 to 2µg/mL. This compound reduced 50% of biofilm produced by C. auris with similar activity to Amp B [48]. A class of molecule ceragenins were also found to inhibit both planktonic and biofilm form of C. auris. Promisingly, they led to significant reductions of fungal infections in ex vivo mucosal tissues [49].
2.3. Essential Oils Are the Potential Sources of Novel Antifungal Skeletons
Essential oils are the fatty acids moieties derived from bioactive plants. These chemical substances alone or in combination with other drugs produce significant biological activities among the different targets. Thus, essential oils have been extensively studied as antimicrobial agents against bacteria and fungus, including C. auris strains.
For instance, Parker et al. demonstrated the effectiveness of selected essential oils against C. auris and found the superior activity for the oils from Cinnamon leaf, clove bud, lemongrass and basil. The effective eradication of C. auris occurred with MIC values ranged from 0.01% to 1.0%. The same study reported the interactions between conventional antifungal drugs and essential oils. Clove bud oil synergistically interacted with fluconazole and flucytosine to combat C. auris [50].
Essential oil extracted from the seeds of Withania somnifera was also tested against C. auris, producing IC50 at 5.96 mg/mL and fungistatic mechanism confirmed by killing assay. Its mechanism of action was associated with disturb in the membrane integrity of C. auris cells, evidenced through ergosterol binding and sorbitol protection assays. However, seed oil was inactive against mature biofilm formed by C. auris [51].
In a more detailed study, Di Vito et al. tested 15 essential oils against 10 clinical strains of C. auris. The results indicated that Cinnamomum zeylanicum essential oil was most effective against C. auris (MIC; 0.06% v/v) in synergy with antifungal drug fluconazole. Further they verified that cinnamaldehyde was the sole reason for the antifungal activity [52]. In the same year, another group of researchers identified the antifungal potency of Cinnamomum cassia essential oil [53], this oil also rich in chemical constitute like cinnamaldehyde [54]. The level of cinnamaldehyde in plants may vary depending upon the species; however, C. cassia and C. zeylanicum has 85.3% and 90.5% cinnamaldehyde respectively [55].
2.4. Natural Products against Candida auris
The world of medicine is increasingly turning to nature's bounty for solutions to modern health challenges. In the realm of fungal infections, a fascinating arsenal of weapons lies hidden within plants, microbes, and even marine organisms. These diverse and potent molecules, meticulously crafted by living organisms, offer a promising alternative to traditional antifungal drugs. With the rising tide of fungal resistance to existing therapies, natural products present a glimmer of hope in the fight against these tenacious pathogens. A list of natural products with anti-candidal activity are listed in Table 2 and Figure 3.
Among the compounds from plants, great focus has been given to Carvacrol, a phenolic monoterpenoid found in essential oils of oregano, thyme, pepperwort, wild bergamot, and other plants. Due to its broad spectrum of biological responses [56]. Carvacrol was active against C. auris by modulating the expression level and action of certain antioxidant enzymes [57]. As like carvacrol, geraniol is another monoterpene alcohol found in geranium oil as a major component. Recently, Fatima and her colleague utilized geraniol against C. auris. Geraniol displayed fungicidal activity and inhibition effect on metabolically active biofilm of C. auris. Subsequently, geraniol improved the survival rate of C. elegans infected by C. auris [58].
Penta-O-galloyl-β-D-glucose (PGG), a bioactive product of many plants (firstly isolated from the leaves of S. terebinthifolia) was also investigated against C. auris. Chemically, PPG is hydrolysable tannin reported with plenty of biological activities such as antibacterial, anticancer, and antiviral activities. PPG demonstrated anticandidal activity with the MIC ranges of 1-8µg/mL against drug-resistant C. auris [59]. Kim and Eom explored the antifungal and anti-biofilm properties of 6-shogaol against C. auris. Shogaols are pungent constituents of ginger similar in chemical structure to gingerol [60]. 6-shogaol demonstrated effectiveness in inhibiting the growth of C. auris at the concentration range of 16-32µg/mL, further it showed promise activity in preventing the formation of biofilms and controlled the secreted aspartyl proteinase activity [61].
In relation to natural compounds from microbial sources, Rubiginosin C obtained from the stromata of the ascomycetes Hypoxylon rubiginosum and Hypoxylon texense, effectively inhibited the formation of biofilms of C. auris and C. albicans [62]. In the last years, many studies focused on the compound Enfumafungin, a triterpene glycoside found in the culture supernatant of Hormonema carpetanum. This compound was acted as a probe to produce biologically active antifungal Ibrexafungerp. Enfumafungin analogues were isolated; they are enfumafungin B and C. Both compounds were effective against clinically relevant C. auris with the MIC of 64 µg/mL. Further molecular docking studies confirmed that these compounds binded in transmembrane region of FKS1 of β-(1,3)-D-glucansynthase [63].
In another study, Persephacin isolated from the endophytic fungus Sphaceloma sp showed activity against a wide spectrum of fungal species (C. albicans, C. glabrata, C. parapsilosis, C. krusei, C. kefyr, C. tropicalis and C. auris). Persephacin resulted in the MIC values around 2.5 µg/mL, which was equal to the activity expressed by Amp B [64]. A group of new linear lipopeptides (Myropeptin C−E and Myropeptin A1) isolated from saprotrophic filamentous fungus Myrothecium inundatum, also produced good inhibition in C. auris cells. In vitro hemolysis, cell viability, and ionophore assays indicated that these compounds target mitochondrial and cellular membranes, inducing cell depolarization and cell death [65]. Subsequentely, Hakuhybotrol along with six known cladobotric acids F, E, H, A, pyrenulic acid A and F2928-1 were isolated from culture broth of Hypomyces pseudocorticiicola FKA-73. Most of these compounds showed promising antifungal activity against azole resistant and sensitive strains of C. auris. In particular cladobotric acids F and E were able to cause a high inhibition in the fungal growth [66].
Although most antifungal compounds from microbial sources were isolated from fungus, some studies investigated compounds from bacterial cultures. For example, culture extract of Lactobacillus paracasei 28.4 inhibited several C. auris strains, acting against planktonic cells, biofilms, and persister cells. Further experiments confirmed that supplementation derived from L. paracasei 28.4 protected G. mellonella from C. auris infection [67].
Finally, compounds from marine sources have also been investigated against C. auris. Turbinmicin, a potent lead obtained from the marine reservoir (Turbinmicin-producing bacterium Micromonospora sp. WMMC-415.), displayed antifungal properties in several in vitro and in vivo experiments. Turbinmicin expressed fungal-specific mode of action, targeting Sec14 of the vesicular trafficking pathway, a unique target yet to be investigated [68]. A subsequent investigation showed that Turbinmicin exhibited a MIC value of 0.125 mg/mL and an inhibitory action on mature biofilm of C. auris [69].
2.5. Peptide-Based Strategies for Eradicating C. auris
Antimicrobial peptides (AMPs) are another alternative group of components reported with superior biological property. They are an active form of smaller segment of protein produced by various organisms that includes plants, insects, human and other small animals. They have its own function when it presents within the host; therefore, they expressed some unexceptional behaviour towards medically important pathogens like bacteria, fungi, and virus. On the other hand, antimicrobial resistance mechanism of AMPs has not yet been elucidated. Wider investigations identified some direct or indirect mechanisms of AMPs against pathogens, with capacity to reduce the virulence traits. As per the information available [70], there are more than 3940 AMPs reported until now that include 3146 natural peptides, 190 predicted and 314 synthetic AMPs. In more specific, HsAFP1 (Heuchera sanguinea); NaD1 (Nicotiana alata flowers); Psd1 (Pisum sativum seeds); Psoriasin, CGA-N46, β-Defensin-1 to 4, Histatin-5 from Homo sapiens; Gomesin, Heliomicin, Jelleine I to IV, Lasioglossin I to III from insects and arachnids; and NFAP2 from filamentous fungi Neosartorya fischeri were reported with anticandidal activity [71]. The list of AMPs with its significant biological role and its origin was presented in Table 3.
Among the microorganisms, both fungi and bacteria produce potent antimicrobial peptides. The fungus Neosartorya fischeri produces two different peptides; they are NFAP and NFAP2 with 57 and 52 amino acids length, respectively. Cystine residues present in these structures have remarkable importance, since they significantly improve the stability of the peptides at high temperature [85,86]. They were identified as potent molecules against fluconazole (FLC)-resistant C. albicans [87] and C. auris [88]. Notoriously, NFAP2 interacted with most of the azoles and echinocandins, producing significant FICI values [88]. The bacteria Bacillus subtilis produces a lipopeptide (AF4) that showed broad spectrum of antifungal activity on more than 110 fungal isolates. Recently, AF4 at 8 mg/L was found to kill most of C. auris cells. Mode of killing was associated with severe cellular membrane disruption and elevated generation of Reactive Oxygen Species (ROS) [76].
Human body is also a potent source of antifungal peptides, so far many peptides with different biological functions were reported from the human body. Peptides like human β-defensin-3 [72], human cathelicidin peptides LL-37 [73] and salivary histatin-5 [74] were recently reported with antifungal activity on C. auris (Table 3). Human β-defensin-3 and cathelicidin peptides LL-37 produced 100% and 70% of synergy with fluconazole [72,73]. Anticandidal activity of human histatin-5 was also documented against other non albicans species like C. glabrata, C. parapsilosis, C. tropicalis, C. krusei, C. guilliermondii and C. tropicalis [89]. In addition, some studies highlighted the ex vivo effects of histatin-5 on mouse models, by reducing the fungal burden in both oral and vaginal infections models [90,91].
Plant defensins are potent molecules that protect plants from infections by effectively combating microbes without harming host cells. Certain plant-derived peptides, like HsAFP1 [92], NaD1[93], Psd1 [94] and D-lp1 [75] exhibited strong activity against C. auris. Among them, D-lp1completely inhibited biofilm formation and virulence of C. auris [75]. As like plant AMPs, other living organisms were found to be a source of antifungal molecules. Crotamine (from south American rattlesnake) acted on multidrug-resistant C. auris without harming cells [77]. Pom-1 and Pom-2 peptides from Pomacea poeyana (Cuban freshwater snail) inhibited bacteria like Pseudomonas aeruginosa and fungi including C. auris [78,95]. Scorpion venom peptides ToAP1 and ToAP2 showed promising antifungal effects against C. auris, alone and associated with other antimicrobial drugs [79] (Table 3).
2.6. Antifungal Immune Therapy against C. auris
Apart from AMPs, there are some other proteinous molecules that arouse interest as therapeutic strategy towards C. auris due to its immunological properties. These group of molecules can be derived from immune system of humans or certain animals. Amongst them, complement receptor 3-related protein (CR3-RP) is one of the key surface antigens expressed during the biofilm formation of Candida species. Previous investigation identified the presence of CR3-RP moieties on the surface of C. auris. Upon in vitro exposure to prepared anti-CR3-RP, C. auris cells failed to form biofilm, confirming the ability of anti-CR3-RP for eradicating C. auris biofilms [96]. Similarly, Singh et al. utilized anti-Hyr1p monoclonal antibody (mAb) to control the C. auris infection. The anti-Hyr1p mAb prevented the biofilm formation and enhanced opsonophagocytic killing of C. auris by macrophages. In vivo studies showed that anti-Hyr1p mAb protected 55% of mice from the systemic infection causes by C. auris [97]. Other than these, NDV-3A (a vaccine based on the N-terminus of Als3 protein formulated with alum) also showed effects against C. auris, blocking the formation of biofilms and encouraging the macrophage-mediated killing of C. auris [98].
A new humanized antibody H5K1 was recently identified and found to be active against C. auris. H5K1 expressed significant results when tested alone or in combination with Caspofungin and Amp B [99]. Recent findings suggested that H5K1 specifically binds to β-1,3-glucans derived from C. auris, causing perturbation and remodeling of the fungal cell wall and facilitating the loss of cellular membrane integrity [100]. In support to this investigation, other Candida cell-surface-specific mAbs were investigated in mouse model of C. auris invasive infection. For example, the specific monoclonal antibody C3.1, that targets β-1,2-mannotriose (β-Man3) of C. auris, was able to improve the survival of animals and reduce the fungal burden in vital organs. In the same study, other peptide-specific mAbs such as 6H1 and 9F2 were reported with targeting two hyphal specific protein 1 (Hwp1) and phosphoglycerate kinase 1 (Pgk1), respectively. It’s also showed the same outcome as like C3.1 in comparison with control group. All together 6H1+9F2 cocktail enhanced the therapeutic outcome than monotherapy. Therefore, all the three antibodies reported here might be an alternative to treat C. auris mediated infections [101].
Intravenous immunoglobulins (IVIG) have been considered an alternative therapeutic strategy to treat the patient who present primary antibody deficiencies. They are the therapeutic product of normal human IgG [102,103,104]. Xin et al. recently demonstrated the role of IVIG in prevention and control of C. auris and C. albicans mediated disseminated candidiasis in animal models. Treatment with IVIG prolonged the survival and reduced the fungal burden in organs of treated animals. In combination, IVIG enhanced the therapeutic index of Amp B in comparison with monotherapy [105].
Immuno-informatics-based approaches offer an alternative and burgeoning avenue for designing suitable vaccine candidates against fungal infections. Recently, subtractive proteomics approaches were employed to design vaccines against C. auris. Khan et al. adopted this method and generated multi-epitope vaccine candidates, which elicited immune responses against C. auris infection by inducing various immune factors such as IgM, IgG, IL-6, and Interferon-α [106]. Similarly, Gupta and colleagues developed a vaccine based on novel CD4+ epitopes through genome-wide scanning and a reverse vaccinology approach. This vaccine had a reduced chance to become ineffective because rapidly evolving genes of C. auris were eliminated from the epitope selection process [107].
2.7. Photosensitizers Based Antimicrobial Photodynamic Therapy (APDT)
APDT is a promising approach as an adjuvant therapy for fungal infections. As advantage, APDT simultaneously targets different biomolecules of pathogens. Thus, it doesn’t have any specific mechanism of action which restrict to development of cross resistance. Therefore, APDT is considered an alternative therapeutic strategy against multiple clinically important organisms that includes C. auris. APDT involves the association of a photosensitizer with irradiation by a light source, that results in generation of ROS, a prime factor responsible for effectiveness of APDT.
Phenothiazinium based photosensitizers are promising agent proven to be inactivated the C. auris cells. Methylene blue, toluidine blue, new methylene blue, and the pentacyclic derivative S137 were assessed as photosensitizers for APDT on C. auris (CDC B11903). Their efficacy was evaluated using MIC and the G. mellonella insect model. Based on the findings, the pentacyclic derivative S137 was identified as a potent treatment for C. auris [108]. Previously, it had demonstrated strong inhibition of C. albicans [109]. In another study researchers utilised red, green, and blue visible lights alone and in combination with photosensitizers (new methylene blue, toluidine blue O and rose bengal) against C. auris. The results showed that blue light alone disturbed the mature biofilm, but it was significantly improved when the photosensitizer was combined. On the other hand, red or green light alone had no effect on Candida biofilm. The biofilms were disturbed only in combination of light and photosensitizers [110].
Recently, Silva and their coworkers assessed the impact of methylene blue and 1,9-dimethyl methylene blue in addition with red LED on C. auris. At 3 μM regardless of the light dose, 1,9-dimethyl methylene blue reduced the metabolic activity of Candida cells. Furthermore, it promoted high level of ROS, lipid peroxidation and mitochondrial membrane damage. In contrast methylene blue was active only in concentration of 100 μM when exposed to highest dose of light. Further, studies evidenced that 1,9-dimethyl methylene blue was capable of inhibiting biofilm formation and mature biofilm formed by C. auris [111]. Earlier studies by Stefanek and his team confirmed the positive effect of methylene blue with a red laser on the biofilm of C. auris. They observed a maximum of 90% biofilm inhibition after 300 seconds of irradiation compared to the growth control. Interestingly, in the presence of 0.25 mM methylene blue, the expression of both the MDR1 and CDR1 genes was affected [112].
2.8. Repurposing of Drugs with Antifungal Properties
Drug repositioning or repurposing is a process of utilizing commercially available drug for treating diseases outside the scope of its original indication. Drug repurposing is considered an important approach to manage emerging diseases caused by bacteria, fungi and virus [113,114]. The availability of various information rather than therapeutic indexes is a valuable point to consider in the repurposing process. Drug repurposing reduce the time and cost of new drug development since it has previous data on toxicity profiles and preclinical parameters [115]. Therefore, repurposing of drug is recognized as an alternative to combat antifungal drug resistance, and several commercial non-antifungal drugs with activity against C. auris has been reported (Figure 4). Among them, Sertraline (an antidepressant agent comes under serotonin reuptake inhibitors) showed active against three different isolates of C. auris, possessing efficient antifungal activity by supressing the action of yeast to hyphae conversion and biofilm formation [116].
Synergistic drug interactions have also been investigated to increase the success of drug repurposing [117]). Pitavastatin, a cholesterol-lowering drug, proved to be a potent azole chemosensitizer. It reduced Candida biofilm formation and lowered MIC ranges when combined with fluconazole against C. auris [118]. Aprepitant, an antiemetic drug, showed ability to disrupt metal ion homeostasis in C. auris, synergizing with azoles to reduce MIC by up to eight-fold and inhibit biofilm formation by 95±0.13% [119]. Miltefosine, an antiparasitic drug licensed for leishmaniasis, demonstrated potential against C. auris and other Candida strains, especially in combination with other antifungal drugs [120,121]. Colistin, an antibiotic used for multidrug-resistant Gram-negative infections like pneumonia, showed synergistic effects when combined with caspofungin, with FICI values ranging from 0.08 to 0.14. However, combining colistin with micafungin yields indifferent results, with FICI values ranging from 0.51 to 1.01 [122]. Synergistic combinations between HIV protease inhibitors and azoles were also found to be active against drug-resistant C. auris. Lopinavir combined with itraconazole achieved potent effects, increasing the survival rate of C. auris-infected C. elegans by up to 90% and reducing fungal burden by 88.5% [123]. Additionally, lopinavir and ritonavir interacted synergistically with itraconazole, effectively combating disseminated candidiasis in a rat model [124]. Atazanavir resensitizes C. auris to azoles by inhibiting efflux pumps, glucose transport, and ATP synthesis [125]. Moreover, the combination of saquinavir and itraconazole significantly reduced fungal burden in murine models, with an 88% decrease in colony-forming units compared to itraconazole alone [126].
Recently, some studies validated the synergistic potential of azoles in combination with Chlorhexidine, used as skin antiseptic and mouthwash due to its broad-spectrum antibacterial effects. It was reported that chlorhexidine can bind to cellular membrane phospholipids, causing changes in osmotic pressure and cell lysis [127]. When combined with fluconazole, chlorhexidine significantly reduced the viability of both planktonic and biofilm forms of C. auris [128]. These results suggest the combined use of chlorhexidine and azoles to control the C. auris infections in cutaneous and mucosal surfaces.
2.9. Nanotechnology Mediated Antifungal Therapy
Metallic nanoparticles have been investigated as antimicrobial agent against a large number of microorganisms. The detailed investigations and its potent antifungal property of different nanoparticles was presented in this review. In the last decades, researchers have extensively worked to develop different metallic nanoparticles like Ag, Zn, and Au targeted to combat pathogenic microorganisms.
In relation to C. auris, most studies focused on silver-based nanoparticles. Humberto and colleagues verified that silver nanoparticles effectively limited the biofilm development at 0.48 ppm, suggesting their use for controlling C. auris in healthcare settings [129]. Another study investigated silver nanoparticles against multidrug-resistant C. auris, showing strong antifungal properties with <0.5 μg/mL on planktonic cells and MIC <2 μg/mL on preformed biofilm [130]. Consistent findings also revealed a significant reduction in viable C. auris cells, in both planktonic and biofilm form, upon treatment with silver nanoparticles [131].
More recently, several functionalised silver nanoparticles have been produced by green synthesis using plants compounds as metal ions reducing agents. Polyphenol-capped metallic silver nanoparticles, such as those derived from Cynara cardunculus extract, exhibited an antifungal effect on C. auris by inducing mitochondrial toxicity and DNA fragmentation at 50 µg/mL [132]. Trimetallic (Ag-Cu-Co) nanoparticles, synthesized with compounds from Salvia officinalis, also showed potent antifungal properties, inducing apoptosis and G2/M phase cell cycle arrest in C. auris, with MIC values ranging from 0.39–0.78 μg/mL and minimum fungicidal concentration ranging from 0.78–1.56 μg/mL [133].
Besides silver nanoparticles, various other metal nanoparticles had proved action against C. auris. For example, bismuth nanoparticles have shown promising effects in combating multidrug-resistant C. auris, exhibiting anticandidal activity with MIC ranging from 1 to 4 µg/mL, and disrupting both cells and biofilms of C. auris [134]. Caspofungin loaded zinc oxide nanoparticles have demonstrated antifungal activity against caspofungin-resistant C. auris. Interestingly, caspofungin-ZnO nanoparticles did not develop acquired or cross resistance in C. auris [135].
2.10. Liposomal Technology for Efficient Drug Delivery
Liposomal technology represents a promising avenue for antifungal therapy, using different approaches to prepare liposomal vehicles. The key-point among these methods is the careful selection of lipid moieties for encapsulating a specific drug. This selection influences the surface charge of the liposome, enabling tailored delivery of potent molecules. The advantages of liposomal technology are manifold, including improved bioavailability, reduced toxicity and targeted delivery. For instance, liposomal formulations of amphotericin B offer enhanced solubility and reduced nephrotoxicity compared to conventional amphotericin B, confirming the potential of liposomal technology in optimizing therapeutic outcomes.
De Alteriis et al. examined the role of liposomal technology to improve the antifungal effects of essential oil from Lavandula angustifolia, reaching antibiofilm activity of persister-derived biofilm of C. auris [136]. Similarly, Lippia sidoides essential oil was loaded in lipid carriers, and improvements in the MIC values were observed on C. auris [137]. In line with earlier investigations, Jaromin et al. utilized liposomal formulation to improve and modulate the surface characteristics of PQA-Az-13, which is the combination of indazole, pyrrolidine, and arylpiperazine scaffolds substituted with a trifluoromethyl moiety. Here, addition of liposome displayed a mean size of 76.4 nm, a positive charge of +45.0 mV with excellent stability, and no toxicity to normal human dermal fibroblasts. PQA-Az-13 showed MIC between 0.67 and 1.25 µg/mL against C. auris and demonstrated promissing results in in-vitro biofilms and ex vivo skin colonization models [138].
3. Conclusion and Future Perspectives
The treatment of C. auris infections faces multiple challenges that are associated with multidrug resistance, global spread, limited therapeutic options, biofilm formation, diagnostic failure, lack of standardized treatment guidelines, underreporting, and surveillance issues. The future of antifungal development is promising with a range of new strategies addressed to the control of C. auris and other emerging fungi. Unlike bacteria, fungi are eukaryotes and share its genetic traits with human genome. Thus, the identification of fungal-specific pathways are required to provide selective targets to fungi, reducing the risk of side effects in humans and minimizing toxicity.
For this, researchers have been working to design compounds with new modes of action, exploring the chemical structures of existing antifungal drugs or investigating new molecules from natural sources, such as plants, microorganisms, insects and other small animals. Another approach is the selection of specific drug carrier such as metallic and liposomal nanoparticles, that can be designed to deliver antifungal drugs directly to C. auris cells, minimizing side effects and improving its efficacy. Face to the advances in the light-based technologies, the antimicrobial photodynamic therapy raises as a promising therapy that simultaneously acts on multiple cell targets in different pathogens. This mechanism of action makes APDT an important adjuvant therapy to control skin and mucosal fungal infections.
Additionally, some researchers are focusing on the drug repurposing strategies. Adapting drugs already approved for other conditions can be a faster and cheaper way to bring new antifungals to market, especially against emerging or resistant fungal threats. While still in early stages, research on fungal vaccines is ongoing, focusing on stimulating the immune system to recognize and fight fungal infections. Tailoring antifungal treatment based on individual patient characteristics and the specific fungal strain involved in the infection can improve outcomes and reduce the risk of resistance.
Finally, combining different therapies can be an effective approach to reach multiple cellular targets and to obtain synergistic interactions, increasing the antifungal efficacy and preventing the emergence of resistant C. auris strains. Although several approaches discussed here had showed promising antifungal activity against C. auris, few researchers extended their results to animal models and clinical trials. Overall, hope that the data gathered in this review can provides support and insights into the advances of new treatments for C. auris infections.
Acknowledgments
The present work was carried out with the support of the Coordination for the Improvement of Higher Education Personnel - Brazil (CAPES) - 88887.890668/2023-00. We are grateful to the Brazilian National Council for Scientific and Technological Development (CNPq 310265/2022-3) and office of Naval Research Global (N62909-20-1-2034).
References
- Frías-De-león, M.G.; Hernández-Castro, R.; Vite-Garín, T.; Arenas, R.; Bonifaz, A.; Castañón-Olivares, L.; Acosta-Altamirano, G.; Martínez-Herrera, E. Antifungal Resistance in Candida Auris: Molecular Determinants. Antibiotics 2020, 9, 568. [Google Scholar] [CrossRef] [PubMed]
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous Emergence of Multidrug-Resistant Candida Auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clin Infect Dis 2017, 64, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.K.; Yasir, M.; Willcox, M. Candida Auris: An Emerging Antimicrobial-Resistant Organism with the Highest Level of Concern. Lancet Microbe 2023, 4, e482–e483. [Google Scholar] [CrossRef]
- Sanyaolu, A.; Okorie, C.; Marinkovic, A.; Abbasi, A.F.; Prakash, S.; Mangat, J.; Hosein, Z.; Haider, N.; Chan, J. Candida Auris: An Overview of the Emerging Drug-Resistant Fungal Infection. Infect Chemother 2022, 54, 236–246. [Google Scholar] [CrossRef]
- Suphavilai, C.; Ko, K.K.K.; Lim, K.M.; Tan, M.G.; Boonsimma, P.; Chu, J.J.K.; Goh, S.S.; Rajandran, P.; Lee, L.C.; Tan, K.Y.; et al. Discovery of the Sixth Candida Auris Clade in Singapore. medRxiv 2023. [Google Scholar] [CrossRef]
- Adams, E.; Quinn, M.; Tsay, S.; Poirot, E.; Chaturvedi, S.; Southwick, K.; Greenko, J.; Fernandez, R.; Kallen, A.; Vallabhaneni, S.; et al. Candida Auris in Healthcare Facilities, New York, USA, 2013-2017. Emerg Infect Dis 2018, 24, 1816–1824. [Google Scholar] [CrossRef] [PubMed]
- Abdolrasouli, A.; Armstrong-James, D.; Ryan, L.; Schelenz, S. In Vitro Efficacy of Disinfectants Utilised for Skin Decolonisation and Environmental Decontamination during a Hospital Outbreak with Candida Auris. Mycoses 2017, 60, 758–763. [Google Scholar] [CrossRef] [PubMed]
- Welsh, R.M.; Bentz, M.L.; Shams, A.; Houston, H.; Lyons, A.; Rose, L.J.; Litvintseva, A.P. Survival, Persistence, and Isolation of the Emerging Multidrug-Resistant Pathogenic Yeast Candida Auris on a Plastic Health Care Surface. J Clin Microbiol 2017, 55, 2996–3005. [Google Scholar] [CrossRef] [PubMed]
- Sherry, L.; Ramage, G.; Kean, R.; Borman, A.; Johnson, E.M.; Richardson, M.D.; Rautemaa-Richardson, R. Biofilm-Forming Capability of Highly Virulent, Multidrug-Resistant Candida Auris. Emerg Infect Dis 2017, 23, 328–331. [Google Scholar] [CrossRef]
- Ledwoch, K.; Maillard, J.Y. Candida Auris Dry Surface Biofilm (DSB) for Disinfectant Efficacy Testing. Materials 2018, 12, 18. [Google Scholar] [CrossRef]
- Ruiz-Gaitán, A.; Martínez, H.; Moret, A.M.; Calabuig, E.; Tasias, M.; Alastruey-Izquierdo, A.; Zaragoza, Ó.; Mollar, J.; Frasquet, J.; Salavert-Lletí, M.; et al. Detection and Treatment of Candida Auris in an Outbreak Situation: Risk Factors for Developing Colonization and Candidemia by This New Species in Critically Ill Patients. Expert Rev Anti Infect Ther 2019, 17, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Lone, S.A.; Ahmad, A. Candida Auris-the Growing Menace to Global Health. Mycoses 2019, 62, 620–637. [Google Scholar] [CrossRef] [PubMed]
- Ademe, M.; Girma, F. Candida Auris: From Multidrug Resistance to Pan-Resistant Strains. Infect Drug Resist 2020, 13, 1287–1294. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, A.; Sood, P. On the Emergence, Spread and Resistance of Candida Auris: Host, Pathogen and Environmental Tipping Points. J Med Microbiol 2021, 70. [Google Scholar] [CrossRef] [PubMed]
- Lyman, M.; Forsberg, K.; Sexton, D.J.; Chow, N.A.; Lockhart, S.R.; Jackson, B.R.; Chiller, T. Worsening Spread of Candida Auris in the United States, 2019 to 2021. Ann Intern Med 2023, 176, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Almaghrabi, R.S.; Albalawi, R.; Mutabagani, M.; Atienza, E.; Aljumaah, S.; Gade, L.; Forsberg, K.; Litvintseva, A.; Althawadi, S. Molecular Characterisation and Clinical Outcomes of Candida Auris Infection: Single-Centre Experience in Saudi Arabia. Mycoses 2020, 63, 452–460. [Google Scholar] [CrossRef] [PubMed]
- Khan, Z.; Ahmad, S.; Al-Sweih, N.; Joseph, L.; Alfouzan, W.; Asadzadeh, M. Increasing Prevalence, Molecular Characterization and Antifungal Drug Susceptibility of Serial Candida Auris Isolates in Kuwait. PLoS ONE 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Al Maani, A.; Paul, H.; Al-Rashdi, A.; Al Wahaibi, A.; Al-Jardani, A.; Al Abri, A.M.A.; Al Balushi, M.A.H.; Al Abri, S.; Al Reesi, M.; Al Maqbali, A.; et al. Ongoing Challenges with Healthcare-Associated Candida Auris Outbreaks in Oman. J Fungi 2019, 5, 101. [Google Scholar] [CrossRef] [PubMed]
- Whaley, S.G.; Berkow, E.L.; Rybak, J.M.; Nishimoto, A.T.; Barker, K.S.; Rogers, P.D. Azole Antifungal Resistance in Candida Albicans and Emerging Non- Albicans Candida Species. Front Microbiol, 2017; 7. [Google Scholar] [CrossRef]
- Rybak, J.M.; Muñoz, J.F.; Barker, K.S.; Parker, J.E.; Esquivel, B.D.; Berkow, E.L.; Lockhart, S.R.; Gade, L.; Palmer, G.E.; White, T.C.; et al. Mutations in TAC1B: A Novel Genetic Determinant of Clinical Fluconazole Resistance in Candida Auris. mBio 2020, 11. [Google Scholar] [CrossRef]
- Rybak, J.M.; Barker, K.S.; Muñoz, J.F.; Parker, J.E.; Ahmad, S.; Mokaddas, E.; Abdullah, A.; Elhagracy, R.S.; Kelly, S.L.; Cuomo, C.A.; et al. In Vivo Emergence of High-Level Resistance during Treatment Reveals the First Identified Mechanism of Amphotericin B Resistance in Candida Auris. Clin Microbiol Infect 2022, 28, 838–843. [Google Scholar] [CrossRef]
- Biagi, M.J.; Wiederhold, N.P.; Gibas, C.; Wickes, B.L.; Lozano, V.; Bleasdale, S.C.; Danziger, L. Development of High-Level Echinocandin Resistance in a Patient With Recurrent Candida Auris Candidemia Secondary to Chronic Candiduria. Open Forum Infect Dis 2019, 6. [Google Scholar] [CrossRef]
- Kordalewska, M.; Lee, A.; Park, S.; Berrio, I.; Chowdhary, A.; Zhao, Y.; Perlin, D.S. Understanding Echinocandin Resistance in the Emerging Pathogen Candida Auris. Antimicrob Agents Chemother 2018, 62. [Google Scholar] [CrossRef]
- Rybak, J.M.; Cuomo, C.A.; David Rogers, P. The Molecular and Genetic Basis of Antifungal Resistance in the Emerging Fungal Pathogen Candida Auris. Curr Opin Microbiol 2022, 70. [Google Scholar] [CrossRef]
- Berkow, E.L.; Lockhart, S.R. Activity of CD101, a Long-Acting Echinocandin, against Clinical Isolates of Candida Auris. Diagn Microbiol Infect Dis 2018, 90, 196–197. [Google Scholar] [CrossRef]
- Rhodes, J.; Abdolrasouli, A.; Farrer, R.A.; Cuomo, C.A.; Aanensen, D.M.; Armstrong-James, D.; Fisher, M.C.; Schelenz, S. Genomic Epidemiology of the UK Outbreak of the Emerging Human Fungal Pathogen Candida Auris. Emerg Microbes Infect 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Pan, J.; Gu, L.; Wang, W.; Wei, B.; Zhang, H.; Chen, J.; Wang, H. Review of Treatment Options for a Multidrug-Resistant Fungus: Candida Auris. Med Mycol 2024, 62. [Google Scholar] [CrossRef] [PubMed]
- Quindós, G.; Miranda-Cadena, K.; San-Millán, R.; Borroto-Esoda, K.; Cantón, E.; Linares-Sicilia, M.J.; Hamprecht, A.; Montesinos, I.; Tortorano, A.M.; Prigitano, A.; et al. In Vitro Antifungal Activity of Ibrexafungerp (SCY-078) Against Contemporary Blood Isolates From Medically Relevant Species of Candida: A European Study. Front Cell Infect Microbiol 2022, 12. [Google Scholar] [CrossRef]
- Wiederhold, N.P.; Najvar, L.K.; Olivo, M.; Morris, K.N.; Patterson, H.P.; Catano, G.; Patterson, T.F. Ibrexafungerp Demonstrates In Vitro Activity against Fluconazole-Resistant Candida Auris and In Vivo Efficacy with Delayed Initiation of Therapy in an Experimental Model of Invasive Candidiasis. Antimicrob Agents Chemother 2021, 65. [Google Scholar] [CrossRef]
- Larkin, E.; Hager, C.; Chandra, J.; Mukherjee, P.K.; Retuerto, M.; Salem, I.; Long, L.; Isham, N.; Kovanda, L.; Borroto-Esoda, K.; et al. The Emerging Pathogen Candida Auris: Growth Phenotype, Virulence Factors, Activity of Antifungals, and Effect of SCY-078, a Novel Glucan Synthesis Inhibitor, on Growth Morphology and Biofilm Formation. Antimicrob Agents Chemother 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Berkow, E.L.; Angulo, D.; Lockhart, S.R. In Vitro Activity of a Novel Glucan Synthase Inhibitor, SCY-078, against Clinical Isolates of Candida Auris. Antimicrob Agents Chemother 2017, 61. [Google Scholar] [CrossRef]
- Hodges, M.R.; Ople, E.; Wedel, P.; Shaw, K.J.; Jakate, A.; Kramer, W.G.; van Marle, S.; van Hoogdalem, E.J.; Tawadrous, M. Safety and Pharmacokinetics of Intravenous and Oral Fosmanogepix, a First-in-Class Antifungal Agent, in Healthy Volunteers. Antimicrob Agents Chemother 2023, 67. [Google Scholar] [CrossRef] [PubMed]
- Pappas, P.G.; Vazquez, J.A.; Oren, I.; Rahav, G.; Aoun, M.; Bulpa, P.; Ben-Ami, R.; Ferrer, R.; Mccarty, T.; Thompson, G.R.; et al. Clinical Safety and Efficacy of Novel Antifungal, Fosmanogepix, for the Treatment of Candidaemia: Results from a Phase 2 Trial. J Antimicrob Chemother 2023, 78, 2471–2480. [Google Scholar] [CrossRef] [PubMed]
- Vahedi-Shahandashti, R.; Lass-Flörl, C. Novel Antifungal Agents and Their Activity against Aspergillus Species. J Fungi 2020, 6, 213. [Google Scholar] [CrossRef] [PubMed]
- Wiederhold, N.P. Review of T-2307, an Investigational Agent That Causes Collapse of Fungal Mitochondrial Membrane Potential. J Fungi 2021, 7, 130. [Google Scholar] [CrossRef] [PubMed]
- Santana, D.J.; Anku, J.A.E.; Zhao, G.; Zarnowski, R.; Johnson, C.J.; Hautau, H.; Visser, N.D.; Ibrahim, A.S.; Andes, D.; Nett, J.E.; et al. A Candida Auris-Specific Adhesin, Scf1, Governs Surface Association, Colonization, and Virulence. Science 2023, 381, 1461–1467. [Google Scholar] [CrossRef] [PubMed]
- Toepfer, S.; Lackner, M.; Keniya, M.V.; Zenz, L.M.; Friemert, M.; Bracher, F.; Monk, B.C. Clorgyline Analogs Synergize with Azoles against Drug Efflux in Candida Auris. J Fungi 2023, 9, 663. [Google Scholar] [CrossRef] [PubMed]
- Chi, X.; Zhang, H.; Wu, H.; Li, X.; Li, L.; Jiang, Y.; Ni, T. Discovery of Novel Tetrazoles Featuring a Pyrazole Moiety as Potent and Highly Selective Antifungal Agents. ACS Omega 2023, 8, 17103–17115. [Google Scholar] [CrossRef]
- Ni, T.; Chi, X.; Xie, F.; Li, L.; Wu, H.; Hao, Y.; Wang, X.; Zhang, D.; Jiang, Y. Design, Synthesis, and Evaluation of Novel Tetrazoles Featuring Isoxazole Moiety as Highly Selective Antifungal Agents. Eur J Med Chem 2023, 246. [Google Scholar] [CrossRef] [PubMed]
- Tu, J.; Zhu, T.; Wang, Q.; Yang, W.; Huang, Y.; Xu, D.; Liu, N.; Sheng, C. Discovery of a New Chemical Scaffold for the Treatment of Superbug Candida Auris Infections. Emerg Microbes Infect 2023, 12. [Google Scholar] [CrossRef]
- Li, J.; Coste, A.T.; Bachmann, D.; Sanglard, D.; Lamoth, F. Assessment of the In Vitro and In Vivo Antifungal Activity of NSC319726 against Candida Auris. Microbiol Spectr 2021, 9. [Google Scholar] [CrossRef]
- Lohse, M.B.; Laurie, M.T.; Levan, S.; Ziv, N.; Ennis, C.L.; Nobile, C.J.; DeRisi, J.; Johnson, A.D. Broad Susceptibility of Candida Auris Strains to 8-Hydroxyquinolines and Mechanisms of Resistance. mBio 2023, 14, e0137623. [Google Scholar] [CrossRef]
- Fuchs, F.; Hof, H.; Hofmann, S.; Kurzai, O.; Meis, J.F.; Hamprecht, A. Antifungal Activity of Nitroxoline against Candida Auris Isolates. Clin Microbiol Infect 2021, 27, e7–e1697. [Google Scholar] [CrossRef] [PubMed]
- Maphanga, T.G.; Mpembe, R.S.; Naicker, S.D.; Govender, N.P. In Vitro Antifungal Activity of Manogepix and Other Antifungal Agents against South African Candida Auris Isolates from Bloodstream Infections. Microbiol Spectr 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.C.; Kilburn, S.; Kapoor, M.; Chaturvedi, S.; Shaw, K.J.; Chaturvedi, V. In Vitro Activity of Manogepix against Multidrug-Resistant and Panresistant Candida Auris from the New York Outbreak. Antimicrob Agents Chemother 2020, 64. [Google Scholar] [CrossRef] [PubMed]
- John, L.L.H.; Thomson, D.D.; Bicanic, T.; Hoenigl, M.; Brown, A.J.P.; Harrison, T.S.; Bignell, E.M. Heightened Efficacy of Anidulafungin When Used in Combination with Manogepix or 5-Flucytosine against Candida Auris In Vitro. Antimicrob Agents Chemother 2023, 67. [Google Scholar] [CrossRef] [PubMed]
- Dennis, E.K.; Kim, J.H.; Parkin, S.; Awuah, S.G.; Garneau-Tsodikova, S. Distorted Gold(I)-Phosphine Complexes as Antifungal Agents. J Med Chem 2020, 63, 2455–2469. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, H.; Eldesouky, H.E.; Hazbun, T.; Mayhoub, A.S.; Seleem, M.N. Identification of a Phenylthiazole Small Molecule with Dual Antifungal and Antibiofilm Activity Against Candida Albicans and Candida Auris. Sci Rep 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, M.M.; Rovig, J.; Holden, B.S.; Taylor, M.F.; Weber, S.; Wilson, J.; Hilton, B.; Zaugg, A.L.; Ellis, S.W.; Yost, C.D.; et al. Ceragenins Are Active against Drug-Resistant Candida Auris Clinical Isolates in Planktonic and Biofilm Forms. J Antimicrob Chemother 2018, 73, 1537–1545. [Google Scholar] [CrossRef]
- Parker, R.A.; Gabriel, K.T.; Graham, K.D.; Butts, B.K.; Cornelison, C.T. Antifungal Activity of Select Essential Oils against Candida Auris and Their Interactions with Antifungal Drugs. Pathogens 2022, 11, 821. [Google Scholar] [CrossRef]
- Balkrishna, A.; Kharayat, B.; Rastogi, S.; Kabdwal, M.; Haldar, S.; Varshney, A. Withania Somnifera Seed Oil Exhibits Antibiofilm Properties against Drug-Resistant Candida Auris Clinical Isolate through Modulation in Cell Permeability. J Appl Microbiol 2023, 134. [Google Scholar] [CrossRef]
- Di Vito, M.; Garzoli, S.; Rosato, R.; Mariotti, M.; Gervasoni, J.; Santucci, L.; Ovidi, E.; Cacaci, M.; Lombarini, G.; Torelli, R.; et al. A New Potential Resource in the Fight against Candida Auris: The Cinnamomum Zeylanicum Essential Oil in Synergy with Antifungal Drug. Microbiol Spectr 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Rosato, R.; Napoli, E.; Granata, G.; Di Vito, M.; Garzoli, S.; Geraci, C.; Rizzo, S.; Torelli, R.; Sanguinetti, M.; Bugli, F. Study of the Chemical Profile and Anti-Fungal Activity against Candida Auris of Cinnamomum Cassia Essential Oil and of Its Nano-Formulations Based on Polycaprolactone. Plants 2023, 12, 358. [Google Scholar] [CrossRef] [PubMed]
- Shreaz, S.; Wani, W.A.; Behbehani, J.M.; Raja, V.; Irshad, M.; Karched, M.; Ali, I.; Siddiqi, W.A.; Hun, L.T. Cinnamaldehyde and Its Derivatives, a Novel Class of Antifungal Agents. Fitoterapia 2016, 112, 116–131. [Google Scholar] [CrossRef] [PubMed]
- Doyle, A.A.; Stephens, J.C. A Review of Cinnamaldehyde and Its Derivatives as Antibacterial Agents. Fitoterapia 2019, 139. [Google Scholar] [CrossRef] [PubMed]
- Sharifi-Rad, M.; Varoni, E.M.; Iriti, M.; Martorell, M.; Setzer, W.N.; del Mar Contreras, M.; Salehi, B.; Soltani-Nejad, A.; Rajabi, S.; Tajbakhsh, M.; et al. Carvacrol and Human Health: A Comprehensive Review. Phytother Res 2018, 32, 1675–1687. [Google Scholar] [CrossRef] [PubMed]
- Ismail, M.; Srivastava, V.; Marimani, M.; Ahmad, A. Carvacrol Modulates the Expression and Activity of Antioxidant Enzymes in Candida Auris. Res Microbiol 2022, 173. [Google Scholar] [CrossRef] [PubMed]
- Fatima, T.; Fatima, Z.; Hameed, S. Abrogation of Efflux Pump Activity, Biofilm Formation, and Immune Escape by Candidacidal Geraniol in Emerging Superbug, Candida Auris. Int Microbiol 2023, 26, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Marquez, L.; Lee, Y.; Duncan, D.; Whitesell, L.; Cowen, L.E.; Quave, C. Potent Antifungal Activity of Penta-O-Galloyl-β-d-Glucose against Drug-Resistant Candida Albicans, Candida Auris, and Other Non-Albicans Candida Species. ACS Infect Dis 2023, 9, 1685–1694. [Google Scholar] [CrossRef] [PubMed]
- Ok, S.; Jeong, W.S. Optimization of Extraction Conditions for the 6-Shogaol-Rich Extract from Ginger (Zingiber Officinale Roscoe). Prev Nutr Food Sci 2012, 17, 166–171. [Google Scholar] [CrossRef]
- Kim, H.R.; Eom, Y.B. Antifungal and Anti-Biofilm Effects of 6-Shogaol against Candida Auris. J Appl Microbiol 2021, 130, 1142–1153. [Google Scholar] [CrossRef]
- Zeng, H.; Stadler, M.; Abraham, W.R.; Müsken, M.; Schrey, H. Inhibitory Effects of the Fungal Pigment Rubiginosin C on Hyphal and Biofilm Formation in Candida Albicans and Candida Auris. J Fungi (Basel) 2023, 9, 726. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Wu, W.; Liu, Y.; Chen, S.; Li, H.; Yang, X.; Zhu, X.; Chen, X.; Yan, L.; Chu, Z.; et al. Natural Enfumafungin Analogues from Hormonema Carpetanum and Their Antifungal Activities. J Nat Prod 2023, 86, 2407–2413. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Haldar, S.; King, J.B.; Mattes, A.O.; Srivastava, S.; Wendt, K.L.; You, J.; Cunningham, C.; Cichewicz, R.H. Persephacin Is a Broad-Spectrum Antifungal Aureobasidin Metabolite That Overcomes Intrinsic Resistance in Aspergillus Fumigatus. J Nat Prod 2023, 86, 1980–1993. [Google Scholar] [CrossRef] [PubMed]
- Jagels, A.; Adpressa, D.A.; Kaweesa, E.N.; McCauley, M.; Philmus, B.; Strother, J.A.; Loesgen, S. Metabolomics-Guided Discovery, Isolation, Structure Elucidation, and Bioactivity of Myropeptins C-E from Myrothecium Inundatum. J Nat Prod 2023, 86, 1723–1735. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Takahashi, S.; Ito, S.; Tokiwa, T.; Noguchi, Y.; Azami, H.; Kojima, H.; Higo, M.; Ban, S.; Nagai, K.; et al. Hakuhybotrol, a Polyketide Produced by Hypomyces Pseudocorticiicola, Characterized with the Assistance of 3D ED/MicroED. Org Biomol Chem 2023, 21, 2320–2330. [Google Scholar] [CrossRef] [PubMed]
- Rossoni, R.D.; de Barros, P.P.; Mendonça, I. do C.; Medina, R.P.; Silva, D.H.S.; Fuchs, B.B.; Junqueira, J.C.; Mylonakis, E. The Postbiotic Activity of Lactobacillus Paracasei 28.4 Against Candida Auris. Front Cell Infect Microbiol 2020, 10, 397. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhao, M.; Braun, D.R.; Ericksen, S.S.; Piotrowski, J.S.; Nelson, J.; Peng, J.; Ananiev, G.E.; Chanana, S.; Barns, K.; et al. A Marine Microbiome Antifungal Targets Urgent-Threat Drug-Resistant Fungi. Science 2020, 370, 974–978. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Zhang, F.; Zarnowski, R.; Barns, K.; Jones, R.; Fossen, J.; Sanchez, H.; Rajski, S.R.; Audhya, A.; Bugni, T.S.; et al. Turbinmicin Inhibits Candida Biofilm Growth by Disrupting Fungal Vesicle-Mediated Trafficking. J Clin Invest 2021, 131. [Google Scholar] [CrossRef] [PubMed]
- Antimicrobial Peptide Database https://aps.unmc.edu/ (accessed May 8, 2024).
- Perez-Rodriguez, A.; Eraso, E.; Quindós, G.; Mateo, E. Antimicrobial Peptides with Anti- Candida Activity. Int J Mol Sci 2022, 23, 9264. [Google Scholar] [CrossRef] [PubMed]
- Shaban, S.; Patel, M.; Ahmad, A. Fungicidal Activity of Human Antimicrobial Peptides and Their Synergistic Interaction with Common Antifungals against Multidrug-Resistant Candida Auris. Int Microbiol 2023, 26, 165–177. [Google Scholar] [CrossRef]
- Rather, I.A.; Sabir, J.S.M.; Asseri, A.H.; Ali, S. Antifungal Activity of Human Cathelicidin LL-37, a Membrane Disrupting Peptide, by Triggering Oxidative Stress and Cell Cycle Arrest in Candida Auris. J Fungi 2022, 8, 204. [Google Scholar] [CrossRef] [PubMed]
- Pathirana, R.U.; Friedman, J.; Norris, H.L.; Salvatori, O.; McCall, A.D.; Kay, J.; Edgerton, M. Fluconazole-Resistant Candida Auris Is Susceptible to Salivary Histatin 5 Killing and to Intrinsic Host Defenses. Antimicrob Agents Chemother 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Kamli, M.R.; Sabir, J.S.M.; Malik, M.A.; Ahmad, A. Characterization of Defensin-like Protein 1 for Its Anti-Biofilm and Anti-Virulence Properties for the Development of Novel Antifungal Drug against Candida Auris. J Fungi 2022, 8, 1298. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, S.; Madduri, M.; Rudramurthy, S.M.; Roy, U. Functional Characterization of a Bacillus-Derived Novel Broad-Spectrum Antifungal Lipopeptide Variant against Candida Tropicalis and Candida Auris and Unravelling Its Mode of Action. Microbiol Spectr 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Dal Mas, C.; Rossato, L.; Shimizu, T.; Oliveira, E.B.; Da Silva Junior, P.I.; Meis, J.F.; Hayashi, M.A.F.; Colombo, A.L. Effects of the Natural Peptide Crotamine from a South American Rattlesnake on Candida Auris, an Emergent Multidrug Antifungal Resistant Human Pathogen. Biomolecules 2019, 9, 205. [Google Scholar] [CrossRef]
- Raber, H.F.; Sejfijaj, J.; Kissmann, A.K.; Wittgens, A.; Gonzalez-Garcia, M.; Alba, A.; Vázquez, A.A.; Vicente, F.E.M.; Erviti, J.P.; Kubiczek, D.; et al. Antimicrobial Peptides Pom-1 and Pom-2 from Pomacea Poeyana Are Active against Candidaauris, C. Parapsilosis and C. Albicans Biofilms. Pathogens 2021, 10. [Google Scholar] [CrossRef]
- Pinheiro, T.K.S.; Nicola, A.M.; Bentes, D.B.; Zhang, S.X.; Felipe, M.S.S.; Silva-Pereira, I.; Albuquerque, P. The Antimicrobial Peptide ToAP2 Is Synergic with Caspofungin and Amphotericin B against Candida Auris; 2023.
- dos Reis, T.F.; de Castro, P.A.; Bastos, R.W.; Pinzan, C.F.; Souza, P.F.N.; Ackloo, S.; Hossain, M.A.; Drewry, D.H.; Alkhazraji, S.; Ibrahim, A.S.; et al. A Host Defense Peptide Mimetic, Brilacidin, Potentiates Caspofungin Antifungal Activity against Human Pathogenic Fungi. Nat Commun 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Bugli, F.; Massaro, F.; Buonocore, F.; Saraceni, P.R.; Borocci, S.; Ceccacci, F.; Bombelli, C.; Di Vito, M.; Marchitiello, R.; Mariotti, M.; et al. Design and Characterization of Myristoylated and Non-Myristoylated Peptides Effective against Candida Spp. Clinical Isolates. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Szerencsés, B.; Gácser, A.; Endre, G.; Domonkos, I.; Tiricz, H.; Vágvölgyi, C.; Szolomajer, J.; Howan, D.H.O.; Tóth, G.K.; Pfeiffer, I.; et al. Symbiotic NCR Peptide Fragments Affect the Viability, Morphology and Biofilm Formation of Candida Species. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Vicente, F.E.M.; González-Garcia, M.; Diaz Pico, E.; Moreno-Castillo, E.; Garay, H.E.; Rosi, P.E.; Jimenez, A.M.; Campos-Delgado, J.A.; Rivera, D.G.; Chinea, G.; et al. Design of a Helical-Stabilized, Cyclic, and Nontoxic Analogue of the Peptide Cm-P5 with Improved Antifungal Activity. ACS Omega 2019, 4, 19081–19095. [Google Scholar] [CrossRef]
- Basso, V.; Garcia, A.; Tran, D.Q.; Schaal, J.B.; Tran, P.; Ngole, D.; Aqeel, Y.; Tongaonkar, P.; Ouellette, A.J.; Selsteda, M.E. Fungicidal Potency and Mechanisms of θ-Defensins against Multidrug-Resistant Candida Species. Antimicrob Agents Chemother 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Tóth, L.; Kele, Z.; Borics, A.; Nagy, L.G.; Váradi, G.; Virágh, M.; Takó, M.; Vágvölgyi, C.; Galgóczy, L. NFAP2, a Novel Cysteine-Rich Anti-Yeast Protein from Neosartorya Fischeri NRRL 181: Isolation and Characterization. AMB Express 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Kovács, L.; Virágh, M.; Takó, M.; Papp, T.; Vágvölgyi, C.; Galgóczy, L. Isolation and Characterization of Neosartorya Fischeri Antifungal Protein (NFAP). Peptides (N.Y.) 2011, 32, 1724–1731. [Google Scholar] [CrossRef] [PubMed]
- Kovács, R.; Holzknecht, J.; Hargitai, Z.; Papp, C.; Farkas, A.; Borics, A.; Tóth, L.; Váradi, G.; Tóth, G.K.; Kovács, I.; et al. In Vivo Applicability of Neosartorya Fischeri Antifungal Protein 2 (NFAP2) in Treatment of Vulvovaginal Candidiasis. Antimicrob Agents Chemother 2019, 63. [Google Scholar] [CrossRef] [PubMed]
- Kovács, R.; Nagy, F.; Tóth, Z.; Forgács, L.; Tóth, L.; Váradi, G.; Tóth, G.K.; Vadászi, K.; Borman, A.M.; Majoros, L.; et al. The Neosartorya Fischeri Antifungal Protein 2 (NFAP2): A New Potential Weapon against Multidrug-Resistant Candida Auris Biofilms. Int J Mol Sci 2021, 22, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Nikawa, H.; Jin, C.; Fukushima, H.; Makihira, S.; Hamada, T. Antifungal Activity of Histatin-5 against Non-Albicans Candida Species. Oral Microbiol Immunol 2001, 16, 250–252. [Google Scholar] [CrossRef]
- Peters, B.M.; Zhu, J.; Fidel, P.L.; Scheper, M.A.; Hackett, W.; El Shaye, S.; Jabra-Rizk, M.A. Protection of the Oral Mucosa by Salivary Histatin-5 against Candida Albicans in an Ex Vivo Murine Model of Oral Infection. FEMS Yeast Res 2010, 10, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.; Liu, S.; Wang, H.; Su, H.; Liu, Z. Efficacy of Histatin5 in a Murine Model of Vulvovaginal Candidiasis Caused by Candida Albicans. Pathog Dis 2017, 75. [Google Scholar] [CrossRef]
- Aerts, A.M.; Bammens, L.; Govaert, G.; Carmona-Gutierrez, D.; Madeo, F.; Cammue, B.P.A.; Thevissen, K. The Antifungal Plant Defensin HsAFP1 from Heuchera Sanguinea Induces Apoptosis in Candida Albicans. Front Microbiol, 2011; 2. [Google Scholar] [CrossRef]
- Hayes, B.M.E.; Bleackley, M.R.; Wiltshire, J.L.; Anderson, M.A.; Traven, A.; Van Der Weerden, N.L. Identification and Mechanism of Action of the Plant Defensin NaD1 as a New Member of the Antifungal Drug Arsenal against Candida Albicans. Antimicrob Agents Chemother 2013, 57, 3667–3675. [Google Scholar] [CrossRef]
- Gonçalves, S.; Silva, P.M.; Felício, M.R.; de Medeiros, L.N.; Kurtenbach, E.; Santos, N.C. Ps D1 Effects on Candida Albicans Planktonic Cells and Biofilms. Front Cell Infect Microbiol, 2017; 7. [Google Scholar] [CrossRef]
- García, M.G.; Rodríguez, A.; Alba, A.; Vázquez, A.A.; Vicente, F.E.M.; Pérez-Erviti, J.; Spellerberg, B.; Stenger, S.; Grieshober, M.; Conzelmann, C.; et al. New Antibacterial Peptides from the Freshwater Mollusk Pomacea Poeyana (Pilsbry, 1927). Biomolecules 2020, 10, 1–17. [Google Scholar] [CrossRef]
- Dekkerová, J.; Lopez-Ribot, J.L.; Bujdáková, H. Activity of Anti-CR3-RP Polyclonal Antibody against Biofilms Formed by Candida Auris, a Multidrug-Resistant Emerging Fungal Pathogen. Eur J Clin Microbiol Infect Dis 2019, 38, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Barbarino, A.; Youssef, E.G.; Coleman, D.; Gebremariam, T.; Ibrahim, A.S. Protective Efficacy of Anti-Hyr1p Monoclonal Antibody against Systemic Candidiasis Due to Multi-Drug-Resistant Candida Auris. J Fungi (Basel) 2023, 9. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Uppuluri, P.; Mamouei, Z.; Alqarihi, A.; Elhassan, H.; French, S.; Lockhart, S.R.; Chiller, T.; Edwards, J.E.; Ibrahim, A.S. The NDV-3A Vaccine Protects Mice from Multidrug Resistant Candida Auris Infection. PLoS Pathog 2019, 15. [Google Scholar] [CrossRef] [PubMed]
- Di Mambro, T.; Vanzolini, T.; Bruscolini, P.; Perez-Gaviro, S.; Marra, E.; Roscilli, G.; Bianchi, M.; Fraternale, A.; Schiavano, G.F.; Canonico, B.; et al. A New Humanized Antibody Is Effective against Pathogenic Fungi in Vitro. Sci Rep 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Vanzolini, T.; Di Mambro, T.; Magnani, M.; Menotta, M. AFM Evaluation of a Humanized Recombinant Antibody Affecting C. Auris Cell Wall and Stability. RSC Adv 2023, 13, 6130–6142. [Google Scholar] [CrossRef] [PubMed]
- Rosario-colon, J.; Eberle, K.; Adams, A.; Courville, E.; Xin, H. Candida Cell-Surface-Specific Monoclonal Antibodies Protect Mice against Candida Auris Invasive Infection. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Ben-Nathan, D.; Lustig, S.; Tam, G.; Robinzon, S.; Segal, S.; Rager-Zisman, B. Prophylactic and Therapeutic Efficacy of Human Intravenous Immunoglobulin in Treating West Nile Virus Infection in Mice. J Infect Dis 2003, 188, 5–12. [Google Scholar] [CrossRef] [PubMed]
- Bayry, J.; Lacroix-Desmazes, S.; Kazatchkine, M.D.; Kaveri, S.V. Intravenous Immunoglobulin for Infectious Diseases: Back to the Pre-Antibiotic and Passive Prophylaxis Era? Trends Pharmacol Sci 2004, 25, 306–310. [Google Scholar] [CrossRef] [PubMed]
- Diep, B.A.; Le, V.T.M.; Badiou, C.; Le, H.N.; Pinheiro, M.G.; Duong, A.H.; Wang, X.; Dip, E.C.; Aguiar-Alves, F.; Basuino, L.; et al. IVIG-Mediated Protection against Necrotizing Pneumonia Caused by MRSA. Sci Transl Med 2016, 8. [Google Scholar] [CrossRef]
- Xin, H.; Rosario-Colon, J.A.; Eberle, K. Novel Intravenous Immunoglobulin Therapy for the Prevention and Treatment of Candida Auris and Candida Albicans Disseminated Candidiasis. mSphere 2023, 8. [Google Scholar] [CrossRef]
- Khan, T.; Suleman, M.; Ali, S.S.; Sarwar, M.F.; Ali, I.; Ali, L.; Khan, A.; Rokhan, B.; Wang, Y.; Zhao, R.; et al. Subtractive Proteomics Assisted Therapeutic Targets Mining and Designing Ensemble Vaccine against Candida Auris for Immune Response Induction. Comput Biol Med 2022, 145. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Osmanoglu, Ö.; Minocha, R.; Bandi, S.R.; Bencurova, E.; Srivastava, M.; Dandekar, T. Genome-Wide Scan for Potential CD4+ T-Cell Vaccine Candidates in Candida Auris by Exploiting Reverse Vaccinology and Evolutionary Information. Front Med (Lausanne) 2022, 9. [Google Scholar] [CrossRef]
- Grizante Barião, P.H.; Tonani, L.; Brancini, G.T.P.; Nascimento, E.; Braga, G.Ú.L.; Wainwright, M.; von Zeska Kress, M.R. In Vitro and in Vivo Photodynamic Efficacies of Novel and Conventional Phenothiazinium Photosensitizers against Multidrug-Resistant Candida Auris. Photochem Photobiol Sci 2022, 21, 1807–1818. [Google Scholar] [CrossRef]
- Rodrigues, G.B.; Dias-Baruffi, M.; Holman, N.; Wainwright, M.; Braga, G.U.L. In Vitro Photodynamic Inactivation of Candida Species and Mouse Fibroblasts with Phenothiazinium Photosensitisers and Red Light. Photodiagnosis Photodyn Ther 2013, 10, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Bapat, P.S.; Nobile, C.J. Photodynamic Therapy Is Effective Against Candida Auris Biofilms. Front Cell Infect Microbiol 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Silva, A.R.; Cabral, F.V.; Silva, C.R.; Silva, D.F.T.; Freitas, A.Z.; Fontes, A.; Ribeiro, M.S. New Insights in Phenothiazinium-Mediated Photodynamic Inactivation of Candida Auris. J Fungi (Basel) 2023, 9. [Google Scholar] [CrossRef]
- Štefánek, M.; Černáková, L.; Dekkerová, J.; Bujdáková, H. Photodynamic Inactivation Effectively Eradicates Candida Auris Biofilm despite Its Interference with the Upregulation of CDR1 and MDR1 Efflux Genes. J Fungi (Basel) 2022, 8. [Google Scholar] [CrossRef]
- Capoci, I.R.G.; Faria, D.R.; Sakita, K.M.; Rodrigues-Vendramini, F.A.V.; Bonfim-Mendonça, P. de S.; Becker, T.C.A.; Kioshima, É.S.; Svidzinski, T.I.E.; Maigret, B. Repurposing Approach Identifies New Treatment Options for Invasive Fungal Disease. Bioorg Chem 2019, 84, 87–97. [Google Scholar] [CrossRef]
- Kim, J.H.; Cheng, L.W.; Chan, K.L.; Tam, C.C.; Mahoney, N.; Friedman, M.; Shilman, M.M.; Land, K.M. Antifungal Drug Repurposing. Antibiotics (Basel) 2020, 9, 1–29. [Google Scholar] [CrossRef]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug Repurposing: Progress, Challenges and Recommendations. Nat Rev Drug Discov 2019, 18, 41–58. [Google Scholar] [CrossRef]
- Gowri, M.; Jayashree, B.; Jeyakanthan, J.; Girija, E.K. Sertraline as a Promising Antifungal Agent: Inhibition of Growth and Biofilm of Candida Auris with Special Focus on the Mechanism of Action in Vitro. J Appl Microbiol 2020, 128, 426–437. [Google Scholar] [CrossRef]
- Talevi, A.; Bellera, C.L. Challenges and Opportunities with Drug Repurposing: Finding Strategies to Find Alternative Uses of Therapeutics. Expert Opin Drug Discov 2020, 15, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Eldesouky, H.E.; Salama, E.A.; Li, X.; Hazbun, T.R.; Mayhoub, A.S.; Seleem, M.N. Repurposing Approach Identifies Pitavastatin as a Potent Azole Chemosensitizing Agent Effective against Azole-Resistant Candida Species. Sci Rep 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Eldesouky, H.E.; Lanman, N.A.; Hazbun, T.R.; Seleem, M.N. Aprepitant, an Antiemetic Agent, Interferes with Metal Ion Homeostasis of Candida Auris and Displays Potent Synergistic Interactions with Azole Drugs. Virulence 2020, 11, 1466–1481. [Google Scholar] [CrossRef] [PubMed]
- Barreto, T.L.; Rossato, L.; de Freitas, A.L.D.; Meis, J.F.; Lopes, L.B.; Colombo, A.L.; Ishida, K. Miltefosine as an Alternative Strategy in the Treatment of the Emerging Fungus Candida Auris. Int J Antimicrob Agents 2020, 56. [Google Scholar] [CrossRef] [PubMed]
- Aghaei Gharehbolagh, S.; Izadi, A.; Talebi, M.; Sadeghi, F.; Zarrinnia, A.; Zarei, F.; Darmiani, K.; Borman, A.M.; Mahmoudi, S. New Weapons to Fight a New Enemy: A Systematic Review of Drug Combinations against the Drug-Resistant Fungus Candida Auris. Mycoses 2021, 64, 1308–1316. [Google Scholar] [CrossRef] [PubMed]
- Bidaud, A.L.; Djenontin, E.; Botterel, F.; Chowdhary, A.; Dannaoui, E. Colistin Interacts Synergistically with Echinocandins against Candida Auris. Int J Antimicrob Agents 2020, 55. [Google Scholar] [CrossRef] [PubMed]
- Eldesouky, H.E.; Salama, E.A.; Lanman, N.A.; Hazbun, T.R.; Seleem, M.N. Potent Synergistic Interactions between Lopinavir and Azole Antifungal Drugs against Emerging Multidrug-Resistant Candida Auris. Antimicrob Agents Chemother 2020, 65. [Google Scholar] [CrossRef]
- Salama, E.A.; Eldesouky, H.E.; Elgammal, Y.; Abutaleb, N.S.; Seleem, M.N. Lopinavir and Ritonavir Act Synergistically with Azoles against Candida Auris in Vitro and in a Mouse Model of Disseminated Candidiasis. Int J Antimicrob Agents 2023, 62. [Google Scholar] [CrossRef]
- Elgammal, Y.; Salama, E.A.; Seleem, M.N. Atazanavir Resensitizes Candida Auris to Azoles. Antimicrob Agents Chemother 2023, 67. [Google Scholar] [CrossRef]
- Elgammal, Y.; Salama, E.A.; Seleem, M.N. Saquinavir Potentiates Itraconazole’s Antifungal Activity against Multidrug-Resistant Candida Auris in Vitro Andin Vivo. Med Mycol 2023, 61. [Google Scholar] [CrossRef] [PubMed]
- Giacobbe, D.R.; Magnasco, L.; Sepulcri, C.; Mikulska, M.; Koehler, P.; Cornely, O.A.; Bassetti, M. Recent Advances and Future Perspectives in the Pharmacological Treatment of Candida Auris Infections. Expert Rev Clin Pharmacol 2021, 14, 1205–1220. [Google Scholar] [CrossRef] [PubMed]
- Hao, W.; Wang, Y.; Xi, Y.; Yang, Z.; Zhang, H.; Ge, X. Activity of Chlorhexidine Acetate in Combination with Fluconazole against Suspensions and Biofilms of Candida Auris. J Infect Chemother 2022, 28, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Lara, H.H.; Ixtepan-Turrent, L.; Jose Yacaman, M.; Lopez-Ribot, J. Inhibition of Candida Auris Biofilm Formation on Medical and Environmental Surfaces by Silver Nanoparticles. ACS Appl Mater Interfaces 2020, 12, 21183–21191. [Google Scholar] [CrossRef]
- Vazquez-Munoz, R.; Lopez, F.D.; Lopez-Ribot, J.L. Silver Nanoantibiotics Display Strong Antifungal Activity Against the Emergent Multidrug-Resistant Yeast Candida Auris Under Both Planktonic and Biofilm Growing Conditions. Front Microbiol 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- AlJindan, R.; AlEraky, D.M. Silver Nanoparticles: A Promising Antifungal Agent against the Growth and Biofilm Formation of the Emergent Candida Auris. J Fungi (Basel) 2022, 8. [Google Scholar] [CrossRef] [PubMed]
- Malik, M.A.; Batterjee, M.G.; Kamli, M.R.; Alzahrani, K.A.; Danish, E.Y.; Nabi, A. Polyphenol-Capped Biogenic Synthesis of Noble Metallic Silver Nanoparticles for Antifungal Activity against Candida Auris. J Fungi (Basel) 2022, 8. [Google Scholar] [CrossRef] [PubMed]
- Kamli, M.R.; Srivastava, V.; Hajrah, N.H.; Sabir, J.S.M.; Hakeem, K.R.; Ahmad, A.; Malik, M.A. Facile Bio-Fabrication of Ag-Cu-Co Trimetallic Nanoparticles and Its Fungicidal Activity against Candida Auris. J Fungi (Basel) 2021, 7, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Munoz, R.; Lopez, F.D.; Lopez-Ribot, J.L. Bismuth Nanoantibiotics Display Anticandidal Activity and Disrupt the Biofilm and Cell Morphology of the Emergent Pathogenic Yeast Candida Auris. Antibiotics (Basel) 2020, 9, 1–15. [Google Scholar] [CrossRef]
- Fayed, B.; Jayakumar, M.N.; Soliman, S.S.M. Caspofungin-Resistance in Candida Auris Is Cell Wall-Dependent Phenotype and Potential Prevention by Zinc Oxide Nanoparticles. Med Mycol 2021, 59, 1243–1256. [Google Scholar] [CrossRef]
- de Alteriis, E.; Maione, A.; Falanga, A.; Bellavita, R.; Galdiero, S.; Albarano, L.; Salvatore, M.M.; Galdiero, E.; Guida, M. Activity of Free and Liposome-Encapsulated Essential Oil from Lavandula Angustifolia against Persister-Derived Biofilm of Candida Auris. Antibiotics (Basel) 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Baldim, I.; Paziani, M.H.; Grizante Barião, P.H.; von Zeska Kress, M.R.; Oliveira, W.P. Nanostructured Lipid Carriers Loaded with Lippia Sidoides Essential Oil as a Strategy to Combat the Multidrug-Resistant Candida Auris. Pharmaceutics 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Jaromin, A.; Zarnowski, R.; Markowski, A.; Zagórska, A.; Johnson, C.J.; Etezadi, H.; Kihara, S.; Mota-Santiago, P.; Nett, J.E.; Boyd, B.J.; et al. Liposomal Formulation of a New Antifungal Hybrid Compound Provides Protection against Candida Auris in the Ex Vivo Skin Colonization Model. Antimicrob Agents Chemother 2024, 68. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
An updated illustration of distribution pattern of C. auris in different geographical location around the world. The data presented here are based on the information retained from recent publication A colour scheme was used to represent the year of C. auris reported. Over 40 countries reported positive for C. auris between 2009 and 2022. By 2022, five clades were reported worldwide; however, recently unpublished data reported the existence of a sixth clade, suspected to have originated in Singapore [5].
Figure 1.
An updated illustration of distribution pattern of C. auris in different geographical location around the world. The data presented here are based on the information retained from recent publication A colour scheme was used to represent the year of C. auris reported. Over 40 countries reported positive for C. auris between 2009 and 2022. By 2022, five clades were reported worldwide; however, recently unpublished data reported the existence of a sixth clade, suspected to have originated in Singapore [5].
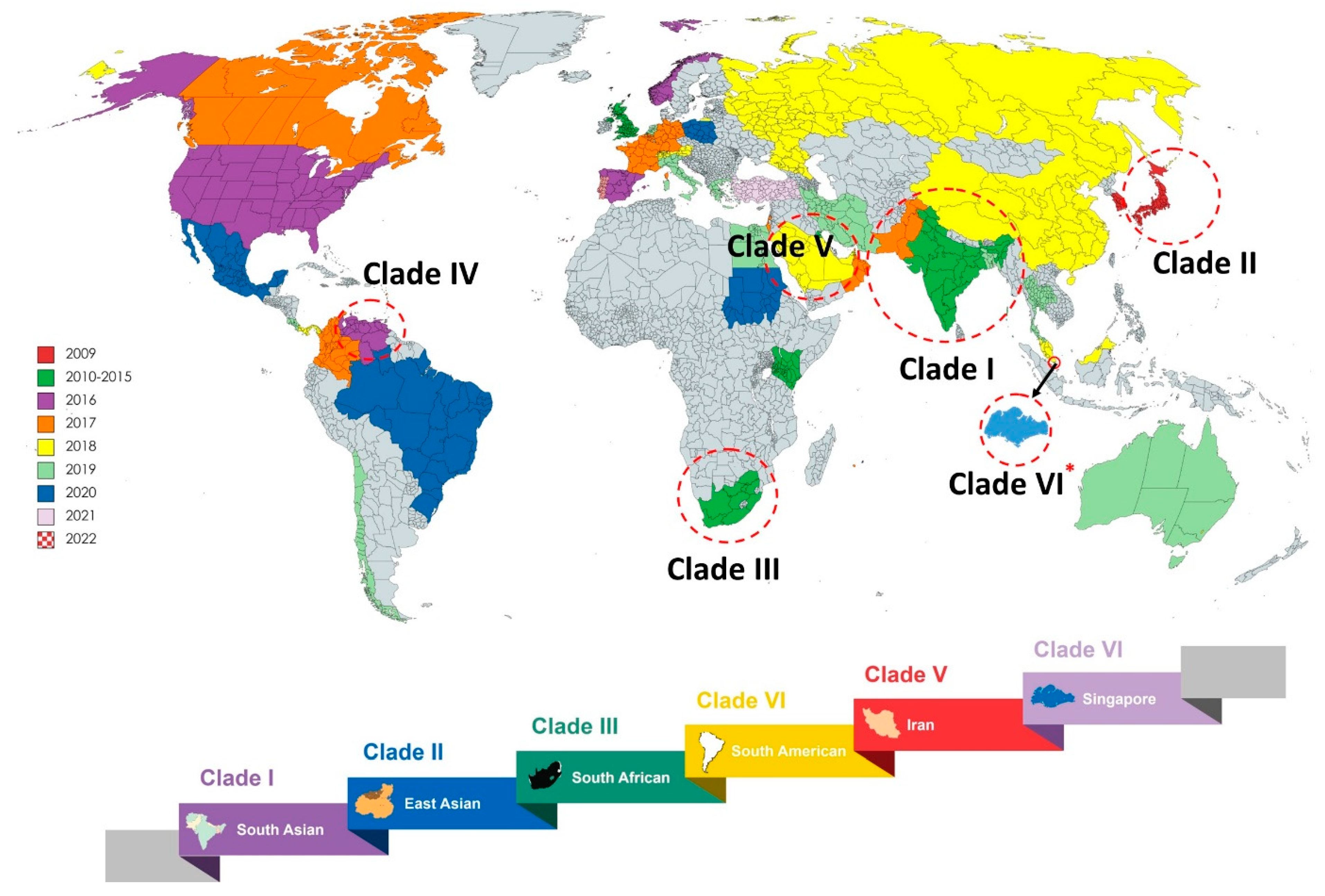
Figure 2.
Current treatment options for C. auris medicated prophylaxis. As of December 2023, only one molecule was approved by US-FDA to manage C. auris and other Candida medicated infections. Rezafungin is a modified version of anidulafungin, the occurrence of structural modification aimed to reduce the hepatotoxicity of the molecules while its efficiency was retained. The data presented here is based on the recent information provided by Wang et al., 2024 [27].
Figure 2.
Current treatment options for C. auris medicated prophylaxis. As of December 2023, only one molecule was approved by US-FDA to manage C. auris and other Candida medicated infections. Rezafungin is a modified version of anidulafungin, the occurrence of structural modification aimed to reduce the hepatotoxicity of the molecules while its efficiency was retained. The data presented here is based on the recent information provided by Wang et al., 2024 [27].
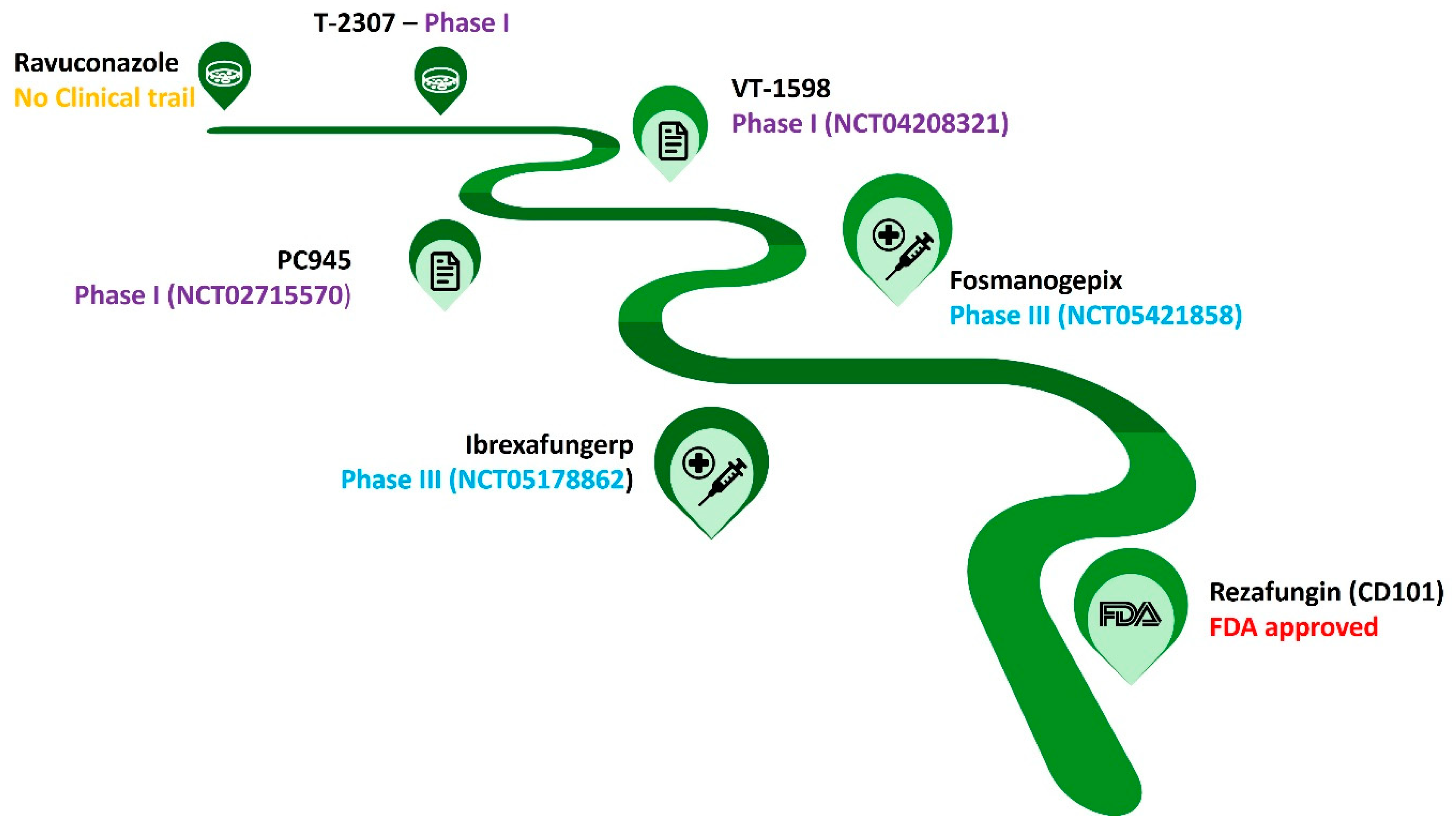
Figure 3.
Scheme of potential molecules from different sources of natural products against C. auris.
Figure 3.
Scheme of potential molecules from different sources of natural products against C. auris.
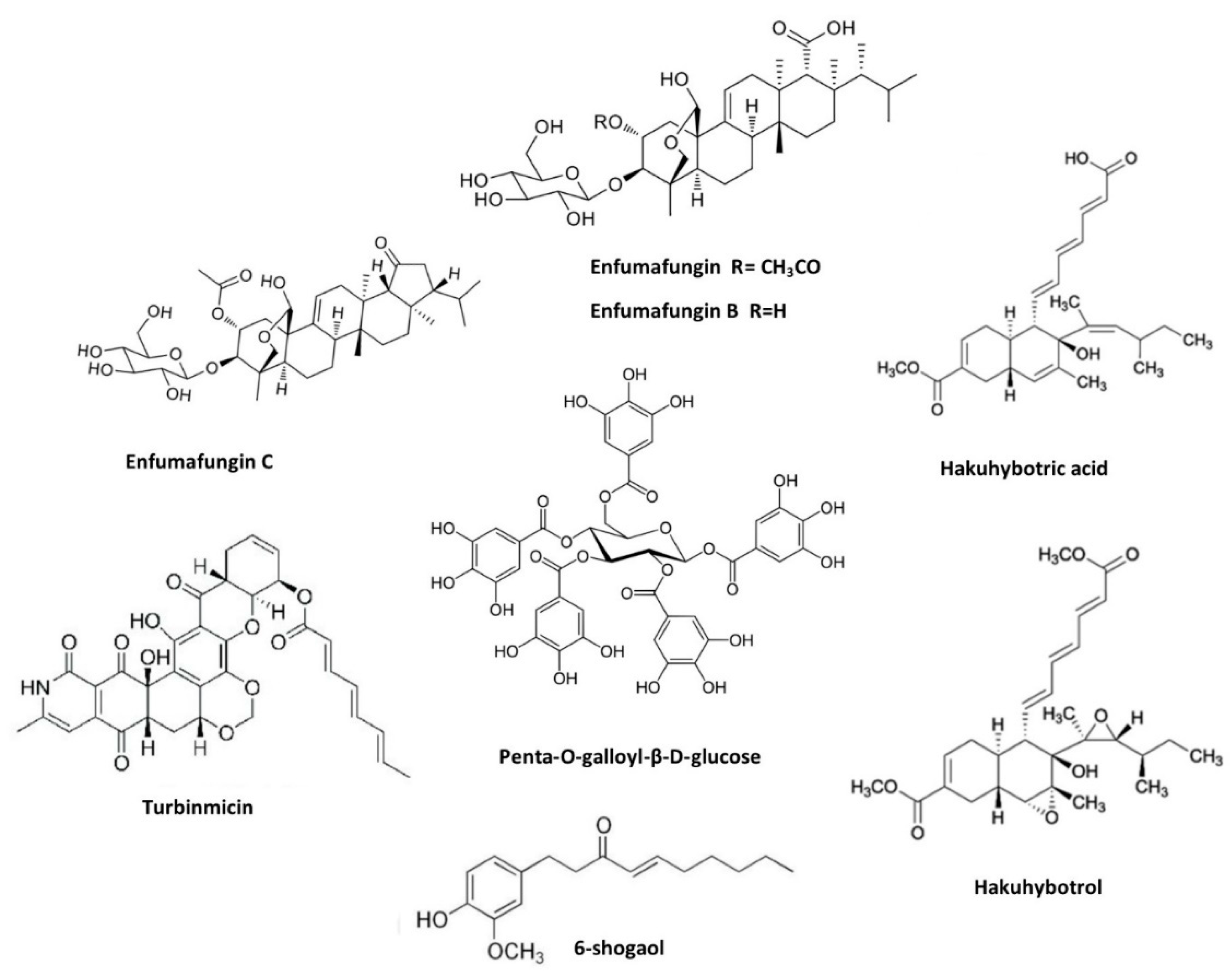
Figure 4.
The list of commercial drug molecules used to manage other disease conditions showing antifungal action against C. auris.
Figure 4.
The list of commercial drug molecules used to manage other disease conditions showing antifungal action against C. auris.
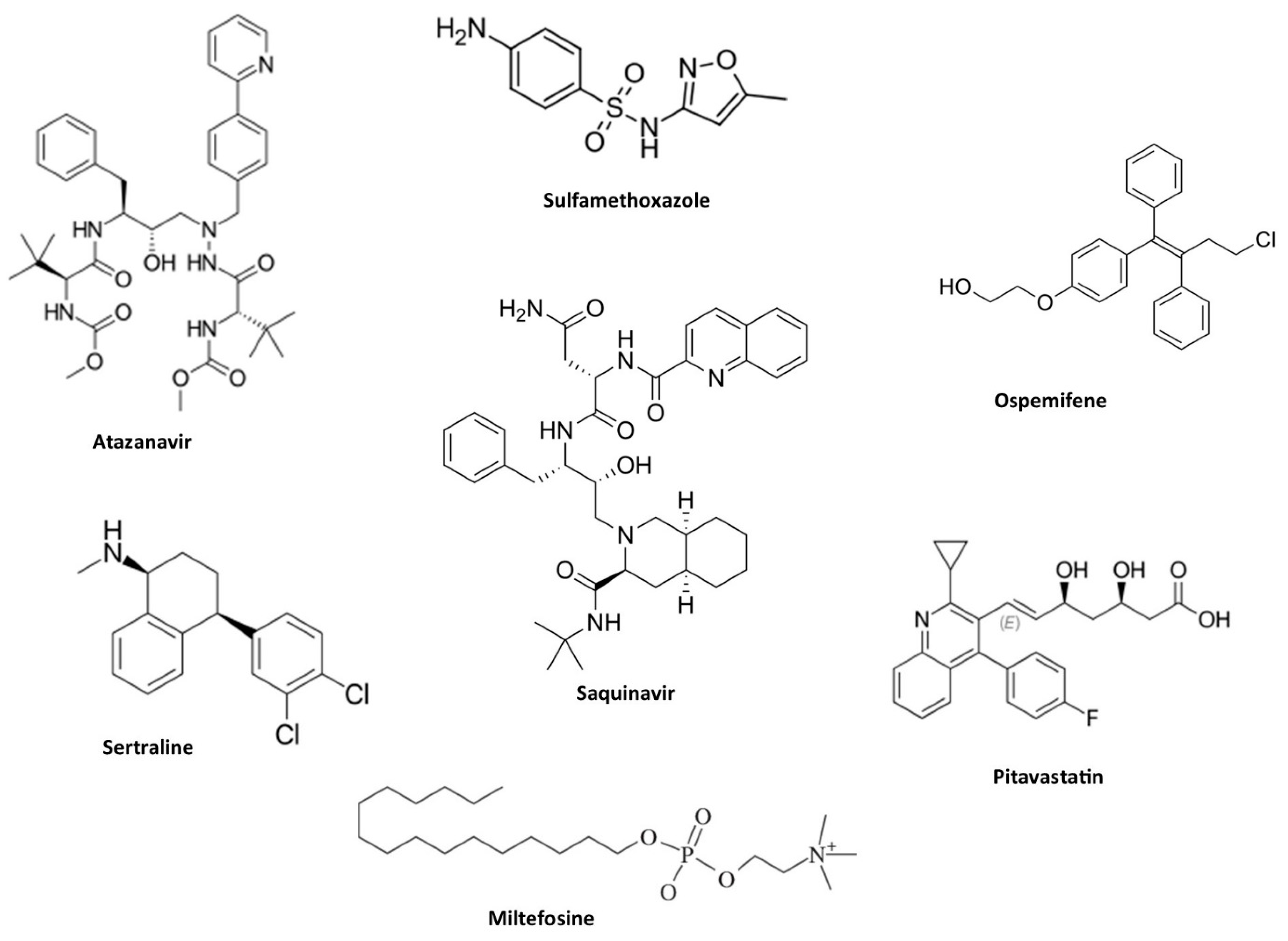

Table 1.
Minimal Inhibitory Concentration (MIC) of different classes of chemically derived compounds against C. auris.
Table 1.
Minimal Inhibitory Concentration (MIC) of different classes of chemically derived compounds against C. auris.
| Group | Compound | MIC/MIC range (µg/ml) | Reference |
|---|---|---|---|
| Clorgyline analogs |
M19 | 86.7 - 98.3 µM | Toepfer et al., 2023 [37] |
| M25 | 137 - 228 µM | ||
| Novel Tetrazoles Featuring a Pyrazole Moiety |
8, 11, 15, 24, 25 | < 0.0625 – 4, 0.125 – 8, < 0.0625 – 2, 0.5 – 64, < 0.0625 - 16 | Chi et al., 2023 [38] |
| Novel tetrazoles featuring isoxazole moiety | 10d, 10h, 13r, 13u | 0.008, < 0.008, 0.0313, 0.0313 | Ni et al., 2023 [39] |
| Benzoanilide group | Compound A1 | 0.5 - 2.0 | Tu et al., 2023 [40] |
| Piperidine based 1,2,3-triazolylacetamide derivatives | pta1, pta2, pta3, pta4, pta5, pta6 | 0.48 - 0.97, 0.24-0.48, 0.12-0.24, >250, >250, >250 | Srivastava et al., 2020 [41] |
| Pyrrolidine-based 1,2,3-triazole | P1 – P10 | 0.97 - 62.5 | Wani et al., 2023 [42] |
| Nitroxoline | 0.125 to 1 | Fuchs et al., 2021[43] | |
| NSC319726 is a thiosemicarbazone | 0.125 to 0.25 | Li et al., 2021 [44] | |
| Manogepix |
FluR | 0.002 to 0.063 | Maphanga et al., 2022 [45] |
| Flu and Amp BR | 0.004 to 0.031 | ||
| PanR | 0.004 μg/mL and 0.008 | ||
| Manogepix | PanR - New York | 0.008 to 0.015 | Zhu et al., 2020 [46] |
| Gold(I)−Phosphine Complex 4 | 3.9 to 7.8 | Dennis et al., 2019 [47] |
|
| Gold(I)−Phosphine Complex 6 | 1.95 | ||
| Phenylthiazole - Compound 1 | 0.25–2 | Mohammad et al., 2019[48] | |
| Ceragenins |
CSA-44, CSA-131, CSA-142, CSA-144 | 0.5 – 1, 0.5 – 1, 2 to 8, 0.5 - 2 | Hashemi et al., 2018 [49] |
FluR – Fluconazole resistant; Amp BR – Amphotericin B resistant; PanR – Resistant to more than two antifungals.
Table 2.
Antifungal natural products against C. auris .
| Species | Molecule | MIC range (µg/ml) | Ref |
|---|---|---|---|
| Turbinmicin-producing bacterium Micromonospora sp. WMMC-415 | Turbinmicin | 0.125-0.50 | Zhang et al., 2020 [56] |
| Hypoxylon rubiginosum and Hypoxylon texense | Penta-O-galloyl-β-D-Glucose | 1-8 | Marquez et al., 2023 [57] |
|
Hormonema carpetanum |
Enfumafungin | 64 | Cheng et al., 2023 [58] |
| Enfumafungin B | > 64 | ||
| Enfumafungin C | > 64 | ||
| Sphaceloma sp from the leaf of Poplar sp | Persephacin | 2.5 | Du et al., 2023 [59] |
|
Myrothecium inundatum |
Myropeptin C | 16 | Jagels et al., 2023 [60] |
| Myropeptin D | 16 | ||
| Myropeptin E | 16 | ||
| Myropeptin A1 | 4 | ||
|
Hypomyces pseudocorticiicola FKA-73 |
Hakuhybotrol | >128 | Watanabe et al., 2023[61] |
| Cladobotric acids F | >128 | ||
| Pyrenulic acid A | 16 | ||
| F2928-1 | 2 | ||
| Cladobotric acids E | 2 to 4 | ||
| Cladobotric acids H | 16 to 32 | ||
| Cladobotric acids A | 4 to 8 | ||
| Lactobacillus paracasei 28.4 | Culture extract | 3.75 to 7.5 mg/mL | Rossoni et al., 2020 [62] |
| Carvacrol | 125-500 | Ismail et al., 2022 [63] | |
| Geraniol | 225 | Fatima et al., 2023 [64] | |
| Nikkomycin Z | 0.125 to >64 | Bentz et al., 2021[65] | |
| 6-shogaol | 16-32 | Kim & Eom, 2021) [66] |
Table 3.
Antifungal peptides against C. auris derived from different classes of organisms.
| S.no | Origin | Name | MIC (µg/ml) | Ref |
|---|---|---|---|---|
| 1 | Homo sapiens | Human β-defensin-3 | 3.125 to 12.5 | Shaban et al., 2023[72] |
| 2 | Homo sapiens | Cathelicidin peptides LL-37 | 25-100 | Rather et al., 2022 [73] |
| 3 | Homo sapiens | Histatin-5 | 7.5 µM | Pathirana et al., 2018[74] |
| 4 | Barley plant | Defensin-like Protein 1 (D-lp1) | 0.047–0.78 mg/mL | Kamli et al., 2022 [75] |
| 5 | Bacillus subtilis | AF4 | 8 | Ramesh et al., 2023[76] |
| 6 | American rattlesnake (Crotalus durissus terrificus) | Crotamine | 40–80 µM which is equal to 0.2 - 0.4 mg/ml | Dal Mas et al., 2019 [77] |
| 7 | Pomacea poeyana - freshwater snail | Pom-1, Pom-2 | 8.5, 8.4 | Raber et al., 2021[78] |
| 8 | Scorpion venom | ToAP1, ToAP2 | >100 µM, 50 - >100 µM | Pinheiro et al., 2023[79] |
| 9 | Brilacidin | Semi synthetic | 80 | Dos Reis et al., 2023 [80] |
| 10 | Chemically prepared, Myristoylated and Non-Myristoylated Peptides | Pep-A, Myr-A, Pep-B, Myr-B, Pep-C, Myr-C | >256, >256, >256, 16-32, >256, 16-64 | Bugli et al., 2022 [81] |
| 11 | Symbiotic NCR Peptide Fragments |
NCR169C 17–38 | 6.25 µM | Szerencsés et al., 2021[82] |
| NCR169C 17–38 ox | 12.5 µM | |||
| 12 | Analogue of the Peptide Cm-p5 | |||
| Monomers | Cm-p5, Cyclic, Hcy | 11, 27, Not active | Vicente et al., 2019[83] |
|
| Dimer | Dimer 1 (parallel) | 30 | ||
| Dimer 2 (anti-parallel) | 31 | |||
| 13 | Rhesus macaque θ-defensin (RTD) | RTD – 1, RTD - 2 | 6.25, 6.25 | Basso et al., 2018[84] |
| Olive baboon θ-defensins (BTD) | BTD – 2, BTD – 4, BTD - 8 | 3.125, > 25, 3.12 - 6.25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



