
Submitted:
22 May 2024
Posted:
24 May 2024
You are already at the latest version
Abstract
With a view to understand and resolve the complexity of the food matrix, ohmic technologies alone or in combination were extensively employed. In this sense, the newest developments and advances of proteomics, metabolomics, lipidomics whith their unique benefits could simplify and understand the link between physiological and pathological activities in biology, physiology, pathology, and food science and processing. This review aims to briefly introduce the basis of proteomics, metabolomics, lipidomics, then expansively review their impact on the assessment of meat quality and safety. Here, also, we discuss the application of proteomics, metabolomics, lipidomics for the authentication and adulteration of meat and meat derivatives from different sources, and provides some perspectives regarding the use of emerging techniques such as rapid mass spectrometry (MS) and non-invasive measurements for the analysis of meat quality and safety. This paper summarizes all significant investigation in these matters, and underlines the advances in analytical chemistry technologies and meat science areas. By emphasizing the requirement for additional examinations, this paper attempts a comprehensive knowledge of “foodomics” and their potential to improve their employment in meat science.
Keywords:
lipidomics
; meat quality
; safety and processing
; proteomics
1. Introduction
For many years, the complexity of the food matrix is a matter of concern for the scientific community. As a result foodomic technologies are used widely and advanced methodologies are applied to food science. Foodomics, a high-throughput approach for the exploration of food science, comprise the transcriptomic, genomic, proteomic, and/or metabolomic investigation of foods for compound profile, authenticity, and/or biomarker-detection associated to food quality/safety; food contaminants, toxicity tests; etc. Proteomics, metabolomics, lipidomics and others sub-disciplines viz. epigenomics, interactomics, metallomics, diseasomics, etc. have started to develop, each with their own set of instruments, techniques, reagents and software. In contrast to previous techniques based on hypothesis-driven research, in foodomics technologies the experimental approach is data-driven.
Proteomics focused on characterization of proteins that are expressed in a cell or tissue type [1]. It is a high-throughput tool in elucidating the responses, functions, modifications, the abundance of all proteins and their isoforms as well as the interactions between them [2,3]. It should be noted that through proteomics it is possible to overcome false positive results or limitations that take place in DNA-based and immunoassay techniques. For example the degradation of DNA after exposure of the meat to high temperature or the nutrient loss and excess of pathogens can easily discriminated using mas-spectrometry technologies based on protein analysis [4]. In the bibliography, several techniques are reported in order to obtain the proteome information and its relationship with the specific product characteristics [5] and have been classified as targeted or untargeted. Untargeted proteomic approaches are suitable for early stages of biomarker discovery, while targeted approaches are selected for validation, implementation and detection proteins of interest with high sensitivity [6]. Consequently, the use of proteomics in meat science enable us to deeply explore, understand and predict meat quality through studying and analyzing difficult to detect effects or interactions among a series of complex events [7].
Metabolomics is the study of the metabolite composition of a tissue or biological fluid [8] and focused on small molecules of relative molecular weights <1500Da [9] and have been classified as targeted or untargeted. More specifically, in targeted metabolomics a specific group of metabolites is examined with identification and quantification of many metabolites within the group. According to the bibliography reviewed, targeted metabolomics can be used in order to interpret the behavior of specific group of metabolites in connection with determined conditions and analyze quantitatively pre-selected metabolites [10,11,12] as compared with untargeted metabolomics which can be used in order to detect groups of metabolites, without necessarily identifying nor quantifying a specific metabolites [13,14]. Furthermore, untargeted metabolomics can be further divided into two types: fingerprinting and profiling [15].
Lipidomics was extensively employed for lipid composition analysis and the quality identification of lipids in foods. Lipidomics forms a system-level analysis of lipids on a large scale, and is employed to detect food adulteration and labeling, along with quantification of individual lipid molecular species. Various analytical methods based on lipomics have been used to quantify trace lipid molecules in foods and hence obtain a comprehensive lipid profile [16,17]. For instance, by UPLC-Q-TOF/MS based, Li et al. [16] analyzed lipid composition, and investigated the lipid differences in Xinjiang Bactrian camel meat, hump, beef, and fatty-tails.
A bibliometric analysis on Lens (lens.org) displays that between 2014 and 2024 (May), a total of 1,603 peer-reviewed scholarly works were published on [Metabolomics and meat OR proteomics and meat OR lipidomics and meat] (Figure 1).
Over the last few years, there has been an increasing interest in Foodomics and their use on meat quality and safety, as can be seen from the number of publications on this topic that has increased continuously according to the lens.org (Figure 1). This ever-growing interest could be explained by the increased scientific community attention on foodomics, as well as the rapid technological advances.
The aim of the present review is to cover the resent proteomics, metabolomics as well as lipidomics studies in meat in the areas of meat quality, safety and processing. Our search was carried out in scopus and used the combination of keywords: Metabolomics and meat quality, metabolomics and meat safety, metabolomics and meat. Here, we summarized the latest research advances regarding the application of advanced foodomics in muscle origin and meat processing.
2. An Outline of Proteomics
In recent years, the study of proteomes has been also applied in food technology in order to correlate the quality and safety of foods with health issues and welfare of the public. Microbial metabolism and responses to stress can be described more precisely by using proteomics [2]. Proteomics is an important method because multiple species detection and unknown target screening can be performed by using mass spectrometry technologies, GC-MS and UHPLC-MS. For example, UHPLCMS-MS and LESA-MS method can identify species-specific markers for meat adulteration detection, and heat stable peptide markers in processed meat. In addition, 2DE-MS and OFFGEL-MS technologies successfully determined undeclared species in commercial processed meat products [4] and LC-MS/MS determined the relationship between proteomes and beef exudate with the color and oxidative quality [18]. Additionally, 2-Dimentional Gel Electroforesis together with MS and MALDI-TOF/TOF-MS technologies were used not only to identify specific protein but also to understand proteome modifications as a consequence of growth, development, postmortem metabolism, and tenderness [5,19]. Last but not least, LC-HRMS together with LC-MS/MS MRM methods are considered complementary tools for the detection, identification, and confirmation of species-specific heat-stable peptides in processed meat products [20]. In this line, several MS approaches have developed recently, and along with chromatographic and NMR techniques have become some of the most commonly applied approaches for metabolomic fingerprinting [20]. Conventionally, MS methods are joined with chromatographic separation techniques, such as LC-MS [20].
Briefly, proteomics have been classified as targeted or untargeted and targeted methods can be further classified in four categories (Figure 2). The first category performs targeted data acquisition for the molecule of interest, the second category is based on targeted data analysis acquiring data for all molecular species, the third category is based on peptide ion data and rely on the mass of the entire molecule, while the last category is based on peptide fragments and rely on the fragments of a molecule [21].
Regarding the quantitation using the Mass spectrometry, proteomics can be classified into two broad categories: the label-based method and label-free method. In label-based method, samples are first differentially labeled and pooled, before subjected to LC-MS/MS analysis. As a consequence of using the label-based method, the expected disparities are minimized. On the other hand, in the label-free quantitative proteomics method the samples are labeled, processed and analyzed independently by LC-MS/MS. The quantification is performed by the measurement of the peak area and/or consideration of the number of MS/MS spectra from each peptide using together various software’s due to the large amount of data [22].
2.1. Proteomic Analyses in Meat Quality Control
The general plan of protemics tools on meat quality monitoring is (i) the conduction on animals, treatments or muscles with diverse quality features, (ii) protein extraction, 2-DE, image analysis and statistical investigation, (iii) data assessment and extract of significantly different proteins, and (iv) proteins identification via MS and interpretation.
According to Setyabrata et al. [18], untargeted protein profiling identified 737 proteins in beef exudate using the LC-MS/MS and demonstrated a distinct proteome profile primarily affected by the muscle source and slightly impacted by aging. The forty nine significantly affected proteins have been classified into five groups based on their potential function. Five of the significantly affected proteins were affected by aging, more specifically in the 23rd day postmortem with structural proteins, and 44 proteins were affected by muscle type.
Moreover, through untargeted proteomics the major proteomes in beef exudate were characterized in relation to meat color and oxidative stability of meat. In the beef loin muscle exudate, five proteins were correlated to meat colour stability while no proteins in tenderoin muscles exudate were found to be correlated. The majority of the identified proteins from tenderloin and beef loin muscles revealed a correlation to meat oxidative stability [18].
Proteomic analyses on chicken breast fillets with white striping myopathy were carried out with the use of liquid chromatography-tandem mass spectrometry (LC-MS/MS) [23]. In particular, 148 differentially abundant proteins were identified in the White striping meats compared to normal non-affected meat. Of those, 43 higher and 105 lower abundant proteins were identified in the White striping meat compared with normal non-affected meat. Vimentin, which is known as marker of White striping myopathy as well as Wooden Breast abnormality, was one of the identified up-regulated proteins. Several proteomic analyses have been reporting regarding White striping and Wooden Breast meat in order to understand the protein expression as well as the mechanisms and biochemical pathways behind breast myopathies [24,25,26,27,28].
Similarly, label-free mass spectromic-based proteomics method was used and 127 differential relative abundance proteins were identified in Wooden Breast chicken meat. More specifically, twenty two of them were detected only in chicken breasts with severe wooden breast myopathy. It should be noted that, these proteins involved in redox homeostasis repairing the respective oxidative damage in tissue as well as maintaining the muscle structure and combating the inflammatory process [28]. The label-free proteomic analysis also conducted in order to identify the differences in protein profile between young and older duck breast muscles (60, 300 and 900-day old). To discriminate differences in the meat between young and older ducks, Gu et al. [29] combined metabolomics/proteomics analyses. Three groups were found, 616 differentially expressed proteins were identified and 61 proteins were screened. Among the pathways examined, purine metabolism was uniquely enriched, which regulated flavor improvement. In addition, NME3, RRM2B, AMPD1, and AMPD3 may also be potential targets to distinguish young and older ducks. The results indicated that the oldest meat had a unique biochemical signature providing biomarkers for distinguishing young from older ducks.
Furthermore, the impact on the quality of meat and the proteomic profiling of duck breast muscle by riboflavin supplementation were determined [30]. As revealed by the results, riboflavin supplementation activated mitochondrial aerobic respiration. Therefore, supplementing duck diets with riboflavin improved breast meat quality.
2.2. Proteomic Analyses in Meat Safety
Proteomic analyses have been widely used in the identification of new markers that are suitable to distinguish accidental contamination from intentional adulteration using the LC-MS/MS methods, based on a detection limit of less than 1% (w/w). Generally, they are identified species-specific markers and their resistance to food processing [20].
Moreover, proteomic analyses have been used in order to detect fraud and adulteration of different animal species, issues in production systems, post-mortem processing and storage. [31].
Proteomics with the aim to provide quick information regarding the food proteins can be effectively used for its correlation with food allergies in consumers because of the undeclared compounds. Safety and authenticity issues are successfully addressed [5]. According to Prandi et al. [32], marker peptides were detected in a complex food matrix such as Bolognese sauce samples. Undoubtedly, these processed meat products constitute the most susceptible to adulteration. Prandi’s validated method demonstrated a good specificity (LOD: 0.2-0.8% on finished product) and sensitivity in authentication of eight meat species (duck, rabbit, chicken, turkey, buffalo, equine, deer and sheep).
Rapid parallel reaction monitoring (PRM) method was developed using high resolution Orbitrap MS in order to detect pork meat. When the most sensitive peptide was selected, the LOD in mixed meat can attain 0.5%. The RSD values between detected and designated pork levels (1%, 5% and 50%) were 4–15%. A total of 125 peptides were identified; however only 5 peptide markers were selected derived from myosin-1 and myosin-4, which could be used for authenticating pork meat in the mixture of sheep, beef and chicken at. Nevertheless, the respective peptide markers were heat sensitive, as a result this method can only be applied for authentication of raw meat samples rather than cooked [33].
Notwithstanding the above method, Naveena et al. [34], developed another accurate OFFGEL electrophoresis together with label-free mass spectromic-based proteomics approach, which could be used in both raw and cooked round meat mixes containing cattle, water buffalo, sheep and goat meat. In this investigation, species-specific peptides derived from myosin light chain-1 and 2 were recognized for authenticating buffalo meat spiked at a minimum 0.5% level in sheep meat with high confidence. Relative quantification of buffalo meat mixed with sheep meat was developed by UPLC-QTOF and PLGS search engine to substantiate the confidence level of the data. In the DNA-based method, PCR amplification of mitochondrial D loop gene using species specific primers found 226 bp and 126 bp product amplicons for buffalo and cattle meat, respectively. These authors reported that this method was effective in perceiving a minimum of 0.5 -1.0% when buffalo meat was spiked with cattle meat in raw and cooked meat combinations. Same trends were reported by Naveena et al., [35] who developed a 2DE and OFFGEL-based proteomic for authenticating raw and cooked water buffalo, sheep and goat meat, and their blends. In-gel and OFFGEL-based proteomic approaches are effectual in validating meat mixes spiked at minimum 1.0% and 0.1% levels, respectively, in triple meat mix for both raw and cooked samples.
2.1. Proteomic Analyses in Meat Processing
Identification of proteins on chicken breast after various thermal treatments was conducted using a label-free proteomics strategy. According to Yang et al. [36], a total of 638 proteins were identified, 84 of them were differentially abundant proteins after steaming process, 89 after boiling, 50 after roasting and 43 after microwaving between processed pale, soft, exudative and normal chicken breast muscles. Through the statistical analysis, 20 proteins with significant contributions to color and texture were screened. Regarding the texture in processed meat samples, the deterioration is associated with the changes in myofibrillar and connective tissue structural proteins as well as sarcoplasmic proteins relevant to heat induced oxidation. Possible indicators for the stability or variation of colour of the above chicken breast muscles (pale, soft, exudative and normal) could be the collagen, tropomyosin, myoglobin, as well as hemoglobin.
According to Wang et al. [37], 20 heat-stable peptides were identified by LC-MS in meat from cooked pork, chicken, duck, beef, and sheep and confirmed by NMR. Furthermore, 24 peptides were identified for the raw meats. Study showed that processing such as grilling and boiling, did not affect the structure of peptides and as a result a species identification can be obtained. Moreover 26 heat stable peptides have been identified for chicken, 1 for turkey and 1 for rabbit [20].

Table 1.
Proteomics for quality control of meat products.
| Meat substrate | Extraction method | Protein identification methodology | Data analysis | Results | Reference | |
|---|---|---|---|---|---|---|
| Meat exudate | Kim et al method, Mohallem & Aryal et al method | LC-MS/MS | ANOVA, PCA, HCA, KEGG | 737 proteins were detected. 222 affected by muscles, aging or their interaction. The samples clustered based on muscle type | [18] | |
| Small-tailed Han sheep, Simmental cattle, Sanyuan hybrid pig, Pekin duck, broiler chicken |
Sarah et al method | UPLC-TripleTOF-MS, NMR | Analyst 1.6.2 software | 53 biomarkers were identified in total. 20 heat-stable peptides were identified for cooked meat and 24 peptides for the raw meats. | [37] | |
| Chicken breast fillets | Kong et al & Kuttappan et al method withmodifications | Orbitrap Lumos, tandem mass tag (TMT) analysis, LC-MS/MS | t-test, IP analysis | 148 differentially abundant proteins were identified in the White striping meats compared with normal non-affected meat. | [23] | |
| Chicken | Montowska & Fornal et al mehod | LC-HRMS LC-MS/MS MRM |
26 heat-stable peptides | [20] | ||
| Normal and Woody Broiler Breast Muscles |
Zhang et al method | 2DE, LC−MS/MS | SAS 9.4 General Linear Model, Fisher’s test, | 20 differentially abundant proteins were identified among at 0 min, 15 min, 4 h, and 24 h postmortem time points in either normal broiler or woody broiler breasts muscles. |
[27] | |
| Normal and Wooden breast chicken meat | Zhu et al method | SDS-PAGE, Q-Exactive Plus MS, coupled to a Dionex Ultimate 3000 RSLCnano |
t-test, Bonferroni, ANOVA, Tukey’s Test, XLSTAT |
127 differential relative abundance proteins, 22 of them detected only in Wooden breast meat and 2 in N breast. | [28] | |
| Duck Breast muscle | UHPLC, Orbitrap, LC-MS/MS |
UniProt-GOA, KEGG, Fisher’s test, one-way ANOVA, GraphPadPrism8.0software | 616 differentially expressed proteins were identified. 61 proteins were screened | [29] | ||
| Pale, soft, exudative and normal chicken breasts (pectoralis major muscle) |
Yang et al method | Q-Exactive HF-X MS/MS, HPLC-MS/MS | UniProt-gallus, MaxQuant 1.6.1.0., Fisher’s test, ANOVA, PCA, PLS-DA | Total 638 proteins were identified, 84, 89, 50 and 43 differentially abundant proteins were identified in steaming, boiling, roasting and microwaving respectively | [36] | |
| Duck breast muscle |
Tang et al mathod | iTRAQ | ANONA, Student’s t-test KEGG |
Total 1641 proteins were identified, 23 selected differentially expressed proteins were involved in the energy metabolism |
[30] | |
| Bolognese sauce | UHPLC/ESI-MS/MS, μHPLC-LTQ-OrbiTRAP | Peaks Studio, SRM |
Good specificity (LOD: 0.2-0.8% on finished product) and sensitivity in authentication of duck, rabbit, chicken, turkey, buffalo, equine, deer and sheep. | [32] | ||
| Shitou and Wuzong geese | UHPLC- MS/MS, 4D-DIA, | ANOVA, PCA, KEGG, |
Total 63.436 peptides were identified, which covered 5.183 proteins. 163 differentially expressed proteins were identified, in the comparison between the leg muscles of Shitou goose and Wuzhong goose. Metabolic pathway, played major role in determining the quality differences in two breeds. |
[38] |
3. An Outline of Metabolomics
Considering that metabolites are able to give information of the biochemical activity of an organism as intermediates or end products of enzymatic reactions, it is consequential to use of metabolomic technologies. As mentioned above metabolomics focused on small molecules of relative molecular weights <1500Da [9] in a biological system and became known by the usage of high-resolution analytical technologies such as nuclear magnetic resonance spectrometry (NMR) as well as mass spectrometry (MS). MS can identify relevant metabolites by comparing database information, for example Fiehn library, Golm Metaabolome Database and Wiley database [39] while NMR can provide structural information about metabolites [40]. Moreover Liquid Chromatography Mass Spectrometry (LC-MS/MS) is widely used because of its high sensitivity and wide detection range and respective analyses were performed using a UHPLC system with Q Exactive HF-X mass spectrometer under the control of the Xcalibur acquisition software [41]. Generally, target analysis is commonly performed with UPLC-ESI-MS/MS, LC-MS/MS and NMR [9,42,43] while in untargeted analysis is commonly used UHPLC-QTOF-MS/MS and LC-Orbitrap-MS/MS [9,42,44] as referred to in Figure 3.
It should be noted that targeted and untargeted metabolomics are highly complementary and both of ohmic methods used to identify metabolites that change in abundance between two or more conditions [42].
3.1. Metabolomic Analyses in Meat Quality Control
In meat quality studies, metabolomics could offer a sign for drawing maps of the metabolic network in postmortem muscle aging and flavor development during cooking. Despite the impact on meat quality, the metabolic changes have not easily predicted, because coordinated metabolism in live muscle is no longer maintained due to lack of energy supply and arrest of de novo gene expression after animal death. Postmortem metabolic maps could result in an outcome of the metabolic factors responsible for meat quality traits and thereby contribute to the exploration of biomarkers in quality monitoring, processing, and authentication of meat products [47]. In the last decade extensive research has been conducted to determine and evaluate the volatile organic compounds in meat products such as aldehydes, ketones, hydrocarbons, alcohols as well as sulfur containing compounds [47] and identified the most important compounds in sausages by the SPME-GC–MS technique [48,49,50].
Flavor substances formation is directly related to metabolic mechanisms and LC-MS/MS is widely used in food flavor metabolomic research. According to recent study [41] metabolomics was used to characterize key aroma substances and their water-soluble markers affecting chicken meat. Metabolomics analyses showed the presence of L-glutamine, a key metabolite of nonanal, hexanal, heptanal, octanal and 1-octen-3-ol affecting and improving chicken flavor through the Maillard reaction.
Moreover physiological and metabolic differences in meat quality between two local breeds of Tibetan sheep was investigated by analyzing differential metabolites through untargeted metabolomic analysis. White Tibetan sheep was clearly distinguished from Black Tibetan sheep as 49 differential metabolites were successfully identified including carbohydrates, amino acids and derivatives, fatty acids and derivatives and other organic compounds [9].
Through metabolomic profiling more information regarding quality in beef muscles can be reported using the exudate as analytical sample and indicator due to the presence of sarcoplasmic proteins, amino acids, sugars, lipids and enzymes [51]. According to Setyabrata et al. [18], metabolomic analysis of meat exudate used aging periods for clustering and clustering of the metabolites by muscle types (Beef loin muscles and tenderloin muscles) was revealed by PCA analysis of exudate metabolites.
Another metabolomic study, according to Zhang et al. [52], revealed significant differences between the meat extract from Liancheng white duck breast meat and Cherry Valley duck meat. The differentiate metabolites were categorized in 28 classes. From these, 4 were the main including carbohydrates, amino acids, fatty acids and eikosanoids. The results revealed that breed has a great impact on the meat quality along with metabolomic profiling of duck meat and correlated with meat quality metabolites may function as markers for the for the above breeds. Similarly, metabolomic profile was employed for the determination of the metabolites of volatile compounds in slow-growing Liancheng white duck and fast-growing Cherry Valley duck meat. Targeted metabolimics showed a lower carnitine content in Liancheng white duck meat. This might promote lipid deposition in order for production of additional octanal and nonanal. Conversely, the higher carnitine content showed in Cherry Valley duck meat may be attributed to the lower lipid oxidation products. In addition the results showed that, the sweet and meaty aroma in slow-growing Liancheng ducks derived from higher sugar and amino acid contents [53].
In recent study by Weng et al. [43], the effect of the age (70, 120 and 300days old) on the nutritive profile of goose meat was investigated. In detail, using widely targeted metabolomics analysis, a total of 776 metabolites were detected in goose meat. In particular, carnitine, anserine as well as nicotinamide riboside increased with the increasing age and played key role in greater nutritional value. Conversely, hypoxanthine, 2-methylsuccinic acid and glutaric acid decreased with the increasing age. In short, Weng et al. [43] concluded that the older geese meat (300 days old) was more nutritious and healthy to human.
Ge et al. [54], using HPLC-QTRAP-MS based metabolomic approach found that the age was affecting the metabolites of Beijing You chicken, a Chinese local breed with superior meat quality [55]. More specifically, the metabolites in the breast muscle were significantly changed during their aging (56, 98 and 120days old). A total of 544 metabolites were detected, of which 60 differential metabolites were detected between 56-98 days old as well as 55 differential metabolites between 98-120 days old chicken. L-carnitine, L-methionine and 3-hydroxybutyrate increased with the increasing age. L-methionine is responsible for the flavor of food, as it produces dimethyl sulfide after heating and equally important is L-carnitine as its promotes growth and improves antioxidant capacity.
Furthermore, during the aging of Wuding-chickens significant changes in metabolic profile were also demonstrated [56]. Using 1H NMR-based metabolomic approach found that metabolic compositional profile of lactate, creatine, IMP, glucose, carnosine, anserine, taurine and glutamine was significantly different in 230days old chicken meat compared to the younger ones.
3.2. Metabolomic Analyses in Meat Safety Control
In order to reduce the incidence of food borne diseases caused by micro-organisms, WHO encourages the use of the irradiation process only if the irradiation dose is up to 10kGy. Even if irradiation is reported as a safe and effective method to extent the self-life of the meat and protect its hygienic quality, in the European Union only a few products which are irradiated in some Member States in substantial amounts are proposed. Especially for meat products, only Chicken meat, poultry, poultry preparations, mechanical recovered chicken meat and chicken offal are proposed for irradiation in Netherlands and France [57].
Metabolomic analyses are expected to be a tool to monitor the impact of irradiation and absolutely it could be useful food inspection tool. Qualitative untargeted metabolomics using HPLC-HRMS–Q-Orbitrap analysis was used by Panseri et al. [46] to evaluate the changes in metabolome profiling of irradiated meat, such as chicken, turkey, mixed ground meat for sausages and connecting to food safety issues relating to metabolome alteration. 402 metabolites were detected, and all 3 matrices exhibited a specific metabolome profile that was not prejudiced by the application of irradiation intensities. The three meat groups exhibited the following similarities (i) free amino acids pool stayed insensible by irradiation, (ii) taurine seemed as the most important differentiator for all three meat groups, (iii) the reduction of the glutathione level and (iv) an increase of adenosine nucleotide degradation. More specifically, changes in amino acids, monosaccharides, nucleoyides and free fatty acid profile as well as potentially presence of any oxidative products that could be formed due to irradiation. According to this study, the irradiation did not cause changes in main food ingredients such as free amino acids pool. Metabolomic analysis did not determine any relevant negative impact of irradiation in meat (only alteration in few metabolic pathways) and as a result the original quality of the meat is maintained.
Additionally the effect of electron beam irradiation treatment in spicy yak jerky, a typical snack meat product and local specialty in China, is investigated. According to this study, low irradiation dose did not result in obvious changes of the protein biological value of the spicy yak jerky, but high dose irradiation (9 kGy) decreased protein nutrition value of spicy yak jerky. More specifically, when the spicy yak jerky was irradiated with 0, 2, 5, and 7 kGy, no significant change was observed in amino acid values, while when it was irradiated with 9kGy a significant decrease of total amino acids was observed [58].
3.3. Metabolomic Analyses in Meat Processing
According to Trithavisup et al. [44], metabolomic profile between cooked Wooden Breast chicken meat and chicken breast without Wooden Breast abnormality was compared in order to identify the differences of metabolic compositions of pepsin-hydrolyzed samples. Untargeted metabolomic analysis did not show any toxic metabolites in both of samples, no apparent compounds that could cause acute adverse health effects due to consumption of cooked Wooden Breast chicken meat. Metabolomic changes between chicken breast with or without abnormality also remained in the cooked product. More specifically, 322 differential metabolites were identified between the cooked samples. Metabolites associated with taurine, hypotaurine metabolism, phenylalanine, tyrosine, tryptophan biosynthesis, D-glutamine and D-glutamate metabolism were most affected because of the Wooden Breast abnormality. Moreover, amino acids together with short peptides as well as carboxamides were also identified.
3.4. Metabolomic Analyses in Meat Authenticity
According to Wang et al. [41], the basic characteristic aroma substances of chicken are nonanal, octanal and dimethyl tetrasulfide, whereas breed-specific aroma compounds found only in native Chinese chickens contain hexanal, 1-octen-3-ol, (E)-2-nonenal, heptanal, and (E,E)-2.4-decadienal. Through this study, using non-targeted metabolome, metabolic differences between white-feathered boilers and native Chinese breeds were identified. More specifically, 821 metabolites detected and divided into 16 substance classes of which the largest (314 metabolites) belonged to amino acids and their metabolites class followed by organic acids and their derivatives (102 metabolites).
Based on Ma et al. [9], identification of local breeding varieties of Tiberian sheep (White Tibetan sheep and the Black Tibetan sheep in Qinghai) was investigated. The results showed that Black Tibetan sheep was superior to the White Tibetan sheep. In targeted metabolomic profiling significant differences in terms of amino acid composition as well as amino acid content observed. Total amino acid and non-essential amino acid for the Black Tibetan sheep were higher measured up with the White Tibetan sheep while no differences were found in the saturated and monosaturated fatty acids.
Using NMR-based metabolomics fingerprinting, differential metabolites were identified. The major metabolites were succinate and various amino acids including isoleucine, leucine, methionine, tyrosine, and valine from beef extracts originating from Australia, Korea, New Zealand, and United States, making them usable biomarkers in order to distinguish the geographical origin of beef [59].
According to Weng et al. [53], differential metabolites identified between two indigenous poultry breeds, the Liancheng white duck and Cherry Valley duck meat. In particular carnitines, were among the top up-regulated metabolites in Cherry Valley duck meat compared to Liancheng white duck meat. Moreover according to Zhou et al. [60], the metabolome data were different between the Liancheng white duck and Mianyang Shelduck duck. In particular, L-Arginine, L-Ornithine, and L-lysine, were found in considerably higher concentrations in Liancheng white duck meat than in Mianyang Shelduck meat.
Furthermore widely targeted metabolomics together with statistical analysis was performed in order to identify potential biomarkers for authentication of older geese meat [43]. Carnitine, anserine as well as nicotinamide riboside could be considered as good biomarkers in order to guarantee the authenticity.
3.5. Metabolomic Analyses in Meat and Impact on Human Health
Untargeted 1H NMR has been used to identify the association between metabolites metabolite-inflammation and diet-inflammation. According to Wood et al. [61] higher intake of processed red meat is associated with lower levels of two anti-inflammatory amino acids. Similarly, red meat intake and cardiovascular disease risk involves higher levers of inflammation according to the same author in another research [62]. Untargeted 1H NMR metabolomic analysis shows that glutamine, anti-inflammatory metabolite, associated with red meat intake when controlling for body mass index. Meanwhile, glutamine was also associated with lower C-reactive protein levels.
Table 2.
Metabolomics for quality control of meat products.
| Meat substrate | Extraction method | Metabolite identification methodology | Data analysis | Results | Reference |
|---|---|---|---|---|---|
| Beijing You chicken | HPLC-QTRAP-MS | SPSS 22.0, one way ANOVA and Ducan΄s, PCA, OPLS-DA | 544 metabolites identified into 32 categories. L-carnitine, L-methionine and 3-hydroxybutyrate increased with the increasing age. | [54] | |
| Cooked Wooden Breast chicken and chicken breast without Wooden Breast abnormality | Solid Phase Extraction | LC-MS/MS, Orbitrap HF MS | Students t-test | Total 1155 metabolites were identified. 322 differential metabolites were identified between the cooked samples. taurine, hypotaurine metabolism, phenylalanine, tyrosine, tryptophan biosynthesis, D-glutamine and D-glutamate metabolism were most affected because of the Wooden Breast abnormality | [44] |
| Chicken, turkey, mixed ground meat for sausages | HPLC-HRMS–Q-Orbitrap | Hierarchical Clustering Analysis for BWC and VP, one way ANOVA with Tukey post-hoc test, multivariate paired t-test. | Irradiation did not cause changes in main food ingredients such as free amino acids pool, only alteration in few metabolic pathways | [46] | |
| Goose meat | Chen et al. method | UPLC-ESI-MS/MS | OPLS-DA, K-means cluster, KEGG | 776 metabolites were detected into 16 classes. Carnitine, anserine, nicotinamide riboside increased with the increasing age. Conversely, hypoxanthine, 2-methylsuccinic acid and glutaric acid decreased with the increasing age. | [43] |
| Red meat | 1H NMR | Bonferroni Correction | Glutamine, anti-inflammatory metabolite, associated with red meat intake when controlling for body mass index and lower CRP levels. | [62] | |
| Liancheng white duck breast meat and Cherry Valley duck meat | UHPLC-QTOF-MS | SPSS 17.0, one way ANOVA and Mann-Whitney test , PCA, OPLS-DA | Significant differences between the two breeds. 28 differentiate metabolites were classified. From these, 4 were the main including carbohydrates, amino acids, fatty acids and eikosanoids | [52] | |
| Meat exudate | Bligh & Dyer et al method | UPLC-MS | ANOVA, PCA, HCA, KEGG | 518 metabolites were detected. 159 affected by muscles, aging or their interavction. The samples clustered based on aging periods | [18] |
| White and Black Tibetan sheep | UPLC- QTOF-MS, NMR for targeted, UHPLC-QTOF-MS/MS for untargeted | SPSS 20.0, PCC | Black Tibetan sheep was superior to the White Tibetan sheep. 49 differential metabolites were identified including carbohydrates, amino acids and derivatives, fatty acids and derivatives and other organic compounds | [9] | |
| Chicken | UHPLC- Orbitrap MS | PCA, OPLS-DA | 821 metabolites detected and divided into 16 classes. The amino acids and their metabolites class was the largest (314 metabolites) followed by organic acids and their derivatives (102 metabolites). | [41] | |
4. An Outline of Lipidomics
Lipidomics and quality
Lipidomics is the sector responsible for the identification of key lipid biomarkers in metabolic regulation. This is achieved mainly by comparison of lipid metabolic changes under different conditions. The final aim is the revealing of the mechanisms of lipid action [63].
Differences in lipid metabolizing capacity arise from different salt substitutions affecting microorganisms’ activity associated with lipolytic enzymes. Hence, the overall flavor might be affected by lipid changes in meat products [64]. Moreover, there might be an accumulation of certain fat-soluble volatile flavor compounds in lipid molecules leading to a slow release over time [65].
Free fatty acids are the end of product of metabolism of fats in raw meat by microorganisms and this can lead to improvement of the quality of the product. Rosinic acid, α -linolenic acid, palmitoleic acid, and 13 s -hydroxyoctadecadienoic acid are associated with Psychrobacter, a potential flavor producer [66].
Free fatty acids (FFA), glycerolipids (GL), glycerophospholipids (GP), sphingolipids (SL), sterol lipids, prenol lipids, saccharolipids and polyketides are considered the main lipid categories of meat [67]. Triacylglycerols (TAGs) and phospholipids (PLs) constitute two of the most common and abundant categories of lipids rich in the contents of ω3 polyunsaturated fatty acids (PUFA). These play health and nutritional roles in the body. Lipidomics is the analytical strategy for investigation of comprehensive studies of lipids in various food matrices. Fingerprinting and dynamic alterations related to lipids understanding, continue to be unexplored and ambiguous [68,69]. Lipidomics based on HPLC-HRMS can be employed for characterization of lipids in pork [70], duck [71] and lamb [72].
Lipid metabolism changes and their compositions for discrimination of lipid species have been detected by Mass spectrometry (MS)-based lipidomics [73].
iKnife and rapid evaporative ionization mass spectrometry (REIMS)-based lipidomics have been employed by Song et al. [74] for identification of salmon and rainbow trout in real time [75]. In this direction, lipid composition changes of hams arising from different origins have been studied extensively [72].
Raw pork meat has been discriminated by lipidomic fingerprints [70] and the mechanism of lipid fragmentation in refrigerated Tan sheep has been well documented [76]. Many previous studies have been reported on ham lipids, focusing on the famous dry-cured pork ham produced in the Mediterranean region and China [77,78,79,80,81]. There is a significant variation in lipid composition among different raw materials [72].
Chen et al. [63] used non-targeted lipidomics, correlation analysis and KEGG pathway analysis and investigated microbial and lipid metabolism changes during fermentation of restructured duck ham of different salt substitutions.
Different techniques have been employed in the classification of hams based on the lipid profile. For example gas chromatography with flame ionization detector (GC-FID) of the lipidic fraction after transmethylation in acid medium [82].
Lipid analysis of different types of food has been accomplished by Proton nuclear magnetic resonance (1H-NMR) spectroscopy [83,84,85], and analysis of the lipid profile of Iberian dry-cured hams using high field 1H-NMR spectroscopy [86,87].
Figure 4 depicts an MS-based lipidomics approach, which can be divided into: lipid extraction, lipid identification by either Gas chromatography (GC)/Liquid chromatography (LC)-MS, and data processing with lipid classification using a chemometric approach by PCA, PLS-DA or other bioinformatics data analysis.
5. Discussion
Meat and meat products are complex food matrix with several challenging difficulties. Their biochemical mechanism and composition are affected during breed, processing and storage. In addition to this, consumer needs for quality food with potential health effects as well as challenges in food production industry regarding distribution and several health and quality risks lead to the need for improving the regulatory system and analytical methods used.
Without a doubt foodomic technologies are able to resolve the quality, safety, processing and authenticity issues. Numerus researches have been conducting using proteomics, metabolomics, and lipidomics technologies in meat and meat products. Based on the reviewed data, the implementation of such advanced methodologies together with statistical approaches can predict the critical issues relating to quality, adulteration, processing and authenticity.
First and foremost, it should be noted that using omic technologies many problems can be overcome, like false positive results or limitations that take place in DNA-based and immunoassay techniques [4]. Taking into consideration that proteomics have been classified as untargeted or targeted, in order to detect biomarkers at the beginning of the research or proteins of interest for validation and implementation respectively with high sensitivity [6]. In addition to this, metabolomics have been also classified as targeted or untargeted in order to interpret the behavior of specific group of metabolites in connection with determined conditions and analyze quantitatively pre-selected metabolites or detect groups of metabolites respectively without necessarily identifying nor quantifying a specific metabolites [10,11,12,13,14]. So it is clear enough, that foodomic technologies in meat science enable us to deeply explore, understand and predict meat quality.
In proteomics different mass spectrometry technologies like GC-MS, UHPLC-MS, LESA-MS, 2DE-MS, MALDI-TOF/TOF-MS and OFFGEL-MS have been successfully used in order to determine not only specific protein or undeclared species in meat products and relationship between proteomes and meat quality, but also to understand proteome modifications as a consequence of meat aging [4,5,18,19]. Last but not least, LC-HRMS together with LC-MS/MS MRM methods are used for the detection, identification, and confirmation of species-specific heat-stable peptides in processed meat products, giving us an excellent tool for distinguishing accidental contamination from intentional adulteration using [20].
In metabolomics of high-resolution analytical technologies like NMR as well as MS have been successfully used in order to identify relevant metabolites by comparing database information, while NMR can provide structural information about metabolites [39,40]. In particular, LC-MS/MS, UPLC-ESI-MS/MS, UHPLC-QTOF-MS/MS, LC-Orbitrap-MS/MS methods are used because of its high sensitivity and wide detection range [9,41,42,43,44].
Despite the extensive research, further research is needed in order to explore new aspects in foodomic methodologies, possible application of them in industry and providing valuable insights in meat safety and quality.
6. Conclusions
Though traditional methods for evaluating intrinsic meat quality and safety characteristics comprising instrumental texture and color studies, pH and sensory traits delivering evidence on meat quality, these latter are not appropriate to estimating the biochemical mechanisms, linking meat quality modifications, or expecting meat quality in the upcoming. Unquestionably, proteomic, metabolomic and lipidomic methodologies or a combination of them are considered complementary tools. In this sense, these attractive measurement tools are progressively popular for researchers intending to clarify quality differences of meat products caused by many features from farm to fork.
While the certain reliability of discussed tools, proteomic, metabolomic and lipidomic fronts encounters in practical applications, mostly in the scope of data investigation, and chemometrics to assist a comprehensive valuation of experimental data and a profounder understanding of how internal metabolic pathways or biological processes change in meat and meat derivatives. Accordingly, to expand the metabolites range and enlarge data quality with the grouping of diverse analytical approaches and platforms, it is indispensable to improve the precision and the sensitivity of instruments. As an illustration, the new technology as IMS (Ion mobility separation) was able to grant a novel dimension for chromatography and MS, permitting the track of quality traits through meat processing over an in situ automatic sampling. Additionally, potent statistical program tools need to be developed to manage and control huge amounts of experimental data for efficiently measuring the security of new or traditional processing technologies in meat systems. Similarly, metabolite libraries of MS/NMR software and web servers require being frequently amplified by totaling more metabolites, forming and recording more reliable standards, mining data from the literature, and explaining structures of new metabolites.
Author Contributions
All authors have read and agreed to the published version of the manuscript.”.
Funding
“This research received no external funding”.
Data Availability Statement
Ν/A.
Acknowledgments
Ν/A.
Conflicts of Interest
“The authors declare no conflicts of interest.”
References
- Bendixen, Em. The use of proteomics in meat science. Meat Sci. 2005, 71, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.; Pius, B. A.; Sun, D.; Yang, K.; Shan, K.; Li, C. Overview of omics applications in elucidating the underlying mechanisms of biochemical and biological factors associated with meat safety and nutrition. J. Proteom. 2023, 276, 104840. [Google Scholar] [CrossRef] [PubMed]
- Tyers, M.; Mann, M. From genomics to proteomics. Nature. 2003, 422, 193–197. [Google Scholar] [CrossRef]
- Li, Y.C.; Liu, S.Y.; Meng, F.B.; Liu, D.Y.; Zhang, Y.; Wang, W.; Zhang, J.M. Comparative review and the recent progress in detection technologies of meat product adulteration. Compr. Rev. Food Sci. Food Saf. 2020, 1–41. [Google Scholar] [CrossRef]
- Agregan, R.; Pateiro, M.; Kumar, M.; Franco, D.; Capanoglu, E.; Dhama, K.; Lorenzo, J.M. The potential of proteomics in the study of processed meat products. J. Proteom. 2023, 270, 104744. [Google Scholar] [CrossRef]
- Sobsey, C.A.; Ibrahim, S.; Richard, V.R.; Gaspar, V.; Mitsa, G.; Lacasse, V.; Zahedi, R.P.; Batist, G.; Borchers, C.H. Targeted and Untargeted Proteomics Approaches in Biomarker Development. Proteomics. 2020, 20, 1900029. [Google Scholar] [CrossRef] [PubMed]
- Purslow, P.; Gagaoua, M.; Warner, R.D. Insights on meat quality from combining traditional studies and proteomics. Meat sci. 2021, 174, 108423. [Google Scholar] [CrossRef]
- Alonso, A.; Marsal, S.; Julia, A. Analytical methods in untargeted metabolomics: state of the art in 2015. Front. bioeng. biotechnol, 2015, 3, article 23. [Google Scholar] [CrossRef]
- Ma, Y.; Han, L.; Zhang, S.; Zhang, X.; Hou, S.; Gui, L.; Sun, S.; Yuan, Z.; Wang, Z.; Yanget, B. Insight into the differences of meat quality between Qinghai white Tibetan sheep and black Tibetan sheep from the perspective of metabolomics and rumen microbiota. Food Chem.: X. 2023, 19, 100843. [Google Scholar] [CrossRef]
- Ramautar, R.; Demirci, A.; Jong, G. J.D. Capillary electrophoresis in metabolomics. Trends Anal. Chem. 2006, 25, 5–455. [Google Scholar] [CrossRef]
- Cevallos-Cevallos, J.M.; Reyes-De-Corcuera, J.I.; Etxeberria, E.; Danyluk, M.D. Rodrick, G.E. Metabolomic analysis in food science: a review. Trends Food Sci. Technol. 2009, 20, 557–566. [Google Scholar] [CrossRef]
- Maier, T.V.; Schmitt-Kopplin, P. Capillary Electrophoresis in Metabolomics. In Capillary Electrophoresis, Schmitt-Kopplin, P., Eds,; Humana Press: 2016; Methods in Molecular Biology, volume 1483, 437-470. https://10. 1007. [Google Scholar]
- Cevallos-Cevallos, J.; Rouseff, R.; Reyes-De-Corcuera, J. Untargeted metabolite analysis of healthy and Huanglongbing-infected orange leaves by CE-DAD. J. Electrophor. 2009, 30, 1240–1248, https://101002/elps200800594. [Google Scholar] [CrossRef] [PubMed]
- Monton, M.R.N.; Soga, T. Metabolome analysis by capillary electrophoresis-mass spectrometry. J. Chromatogr. A. 2007, 1168, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Utpott, M.; Rodrigues, E.; Oliveira Rios, A.; Mercali, G.D.; Flôres, S.H. Metabolomics: An analytical technique for food processing evaluation. Food Chem, 2022; 366, 130685. [Google Scholar] [CrossRef]
- Li, Q.; Yang, L.; Li, R.; Chen, G.; Dong, J.; Wu, L.; Fu, Y.; Yang, J. Lipid analysis of meat from Bactrian camel (Camelus bacterianus), beef, and tails of fat-tailed sheep usingUPLC-Q-TOF/MS based lipidomics. Front. Nutr. 2023, 10, 1053116. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Z.; Ran, J.; Yang, C.; Lin, Z.; Liu, Y. LC/MS-based lipidomics to characterize breed-specific and tissue-specific lipid composition of chickenmeat and abdominal fat. LWT-Food Sci. Technol. 2022, 163, 113611. [Google Scholar] [CrossRef]
- Setyabrata, D.; Ma, D.; Xie, S.; Thimmapuram, J.; Cooper, B.R.; Aryal, U.K.; Kim, Y.H.B. Proteomics and metabolomics profiling of meat exudate to determine the impact of postmortem aging on oxidative stability of beef muscles. Food Chem. 2023, 18, 100660. [Google Scholar] [CrossRef] [PubMed]
- Mora, L.; Gallego, M.; Toldra, F. New approaches based on comparative proteomics for the assessment of food quality. Curr. Opin. Food Sci. 2018, 22, 22–27. [Google Scholar] [CrossRef]
- Stachniuk, A.; Sumara, M.; Montowska, E. Fornal, Liquid chromatography–mass spectrometry bottom-up proteomic methods in animal species analysis of processed meat for food authentication and the detection of adulterations. Mass Spectrom. Rev. 2019, 40, 3–30. [Google Scholar] [CrossRef] [PubMed]
- Borras, E.; Sabido, E. What is targeted proteomics? A concise revision of targeted acquisition and targeted data analysis in mass spectrometry, Proteomics, 2017, 17, 1700180. [Google Scholar] [CrossRef]
- Anand, S.; Samuel, M.; Ang, C.S.; Keerthikumar, S.; Mathivanan, S. ; Label-Based and Label-Free Strategies for Protein Quantitation. In Proteome Bioinformatics, Keerthikumar, S., Mathivanan, S., Eds.; Humana Press, 2017; Methods in Molecular Biology, volume 1549, pp. 31–43. [CrossRef]
- Kong, B.; Owens, C.; Bottje, W.; Shakeri, M.; Choi, J.; Zhuang, H.; Bowker, B. Proteomic analyses on chicken breast meat with white striping myopathy. Poult. Sci. 2024, 103, 103682. [Google Scholar] [CrossRef]
- Kuttappan,V. A.; Bottje, W.; Ramnathan, R.; Hartson, S.D.; Coon, C.N.; Kong, B.W.; Owens, C.M.; Vazquez-Anon, M.; Hargis, B.M. Proteomic analysis reveals changes in carbohydrate and protein metabolism associated with broiler breast myopathy. Poult. Sci. 2017, 96, 2992–2999. [Google Scholar] [CrossRef] [PubMed]
- Cai, K.; Shao, W.; Chen, X.; Campbell, Y.L.; Nair, M.N.; Suman, S.P.; Beach, C.M.; Guyton, M.C.; Schilling, M.W. Meat quality traits and proteome profile of woody broiler breast (pectoralis major) meat. Poult. Sci. 2018, 97, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Antonelo, D.; Hendrix, J.; To, V.; Campbell, Y.; Staden, M.V.; Li, S.; Suman, S.P.; Zhai, W.; Chen, J.; Zhu, H.; Schilling, M.W. Proteomic characterization of normal and woody breast meat from broilers of five genetic strains. MMB, 2020, 4, 1. [Google Scholar] [CrossRef]
- Zhang, X.; Zhai, W.; Li, S.; Suman, S.P.; Chen, J.; Zhu, H.; Antonelo, D.S.; Schilling, M.W. Early postmortem proteome changes in normal and woody broiler breast muscles. J. Agric. Food Chem. 2020, 68, 39–11000. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, L.M.; Rocha, T.C.; Delgado, J.; Diaz-Velasco, S.; Madruga, M.S.; Estevez, M. Deciphering the underlying mechanisms of the oxidative perturbations and impaired meat quality in Wooden breast myopathy by label-free quantitative MS-based proteomics. Food Chem. 2023, 423, 136314. [Google Scholar] [CrossRef] [PubMed]
- Gu, T.; Duan, M.; Chen, L.; Tian, Y.; Xu, W.; Zeng, T.; Lu, L. ; Proteomic-metabolomic combination analysis reveals novel biomarkers of meat quality that differ between young and older ducks. Poultry Sci. 2024, 103, 103530. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Zhang, B.; Liu, D.; Gao, K.; Dai, Y.; Liang, S.; Cai, W.; Li, Z.; Guo, Z.; Hu, J.; Zhou, Z.; Xie, M.; Hou, S. Dietary riboflavin supplementation improves meat quality, antioxidant capacity, fatty acid composition, lipidomic, volatilomic, and proteomic profiles of breast muscle in Pekin ducks. Food Chem.: X, 2023, 19, 100799. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Pavan, E.; Ross, A.B.; Deb-choudhury, S.; Dixit, Y.; Mungure, T.E.; Realini, C.E.; Cao, M.; Farouk, M.M. Molecular insights into quality and authentication of sheep meat from proteomics and metabolomics. J. Proteom. 2023, 276, 104836. [Google Scholar] [CrossRef]
- Prandi, B.; Varania, M.; Faccinic, A.; Lambertinid, F.; Sumand, M.; Leporatid, A.; Tedeschia, T.; Sforza, S. Species specific marker peptides for meat authenticity assessment: A multispecies quantitative approach applied to Bolognese sauce. Food Control, 2019, 97, 15–24. [Google Scholar] [CrossRef]
- Pan, X.D.; Chen, J.; Chen, Q.; Huang, B.F.; Han, J.L. Authentication of pork in meat mixtures using PRM mass spectrometry of myosin peptides. RSC Adv. 2018, 8, 11157–11162. [Google Scholar] [CrossRef]
- Naveena, B.M.; Jagadeesh, D.S.; Jagadeesh, B.A.; Madhava, R.T.; Kamuni, V.; Vaithiyanathan, S.; Kulkarni, V.V.; Rapole, S. OFFGEL electrophoresis and tandem mass spectrometry approach compared with DNA-based PCR method for authentication of meat species from raw and cooked ground meat mixtures containing cattle meat, water buffalo meat and sheep meat. Food Chem. 2017, 233, 311–320. [Google Scholar] [CrossRef]
- Naveena, B.M.; Jagadeesh, D.S.; Kamuni, V.; Muthukumar, M.; Kulkarni, V.V.; Kiran, M.; Rapole, S. In-gel and OFFGEL-based proteomic approach for authentication of meat species from minced meat and meat products. J Sci. Food Agric. 2017, 98, issue 3, 1188–1196. [Google Scholar] [CrossRef]
- Yang, T.; Yang, Y.; Zhang, P.; Li, W.; Ge, Q.; Yu, H.; Wu, M.; Xing, L.; Qian, Z.; Gao, F.; Liu, R. Quantitative proteomics analysis on the meat quality of processed pale, soft, and exudative (PSE)-like broiler pectoralis major by different heating methods. Food Chem. 2023, 426, 136602. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.J.; Zhou, G.Y.; Ren, H.W.; Xu, Y.; Yang, Y.; Guo, L.H.; Liu, N. Peptide biomarkers identified by LC–MS in processed meats of five animal species. J. Food Compos. Anal. 2018, 73, 47–54. [Google Scholar] [CrossRef]
- Chen, G.; Qi, L.; Zhang, S.; Peng, H.; Lin, Z.; Zhang, X.; Nie, Q.; Luo, W. Metabolomic, lipidomic, and proteomic profiles provide insights on meat quality differences between Shitou and Wuzong geese. Food Chem. 2024, 438, 137967. [Google Scholar] [CrossRef]
- Putri, S. P.; Ikram, M.; Sato, A.; Dahlan, H.A.; Rahmawati, D.; Ohto, Y.; Fukusaki, E. Application of gas chromatography-mass spectrometry-based metabolomics in food science and technology. J. Biosci. Bioeng. 2022, 133, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Zhang, L.; Zheng, X.; Huang, Q.; Farag, M.A.; Zhu, R.; Zhao, C. Emerging applications of metabolomics in food science and future trends. Food Chem.: X, 2022, 16, 100500. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, L.; Liu, X.; Wang, Y.; Wang, Y.; Zhao, W.; Zhao, G.; Cui, H.; Wen, J. Identification of characteristic aroma compounds in chicken meat and their metabolic mechanisms using gas chromatography–olfactometry, odor activity values, and metabolomics. Food Res. Int. 2024, 175, 113782. [Google Scholar] [CrossRef]
- Li, S.; Tian, Y.; Jiang, P.; Lin, Y.; Liu, X.; Yang, H. Recent advances in the application of metabolomics for food safety control and food quality analyses. Crit. Rev. Food Sci. Nutr. 2020, 61, 1448–1469. [Google Scholar] [CrossRef]
- Weng, K.; Huo, W.; Song, L.; Cao, Z.; Zhang, Y.; Zhang, Y.; Chen, G.; Xu, Q. Effect of marketable age on nutritive profile of goose meat based on widely targeted metabolomics. LWT - Food Sci. Technol. 2022, 170, 114071. [Google Scholar] [CrossRef]
- Trithavisup, T.; Krobthonget, S.; Yingchutrakul, Y.; Sanpinit, P.; Malila, Y. Impact of Wooden Breast myopathy on in vitro protein digestibility, metabolomic profile, and cell cytotoxicity of cooked chicken breast meat. Poultry Sci. 2024, 103, 103261. [Google Scholar] [CrossRef]
- Wu, M.; Zuo, S.; Maiorano, G.; Kosobucki, P.; Stadnicka, K. How to employ metabolomic analysis to research on functions of prebiotics and probiotics in poultry gut health? Front. Microbiol. 2022, 13, 1040434. [Google Scholar] [CrossRef] [PubMed]
- Panseri, S.; Arioli, F.; Pavlovic, R.; Di Cesare, F.; Nobile, M.; Mosconit, G.; Villa, R.; Chiesa, L.M.; Bonerba, E. Impact of irradiation on metabolomics profile of ground meat and its implications toward food safety. LWT - Food Sci. Technol. 2022, 161, 113305. [Google Scholar] [CrossRef]
- Feng, Y.; Cai, Y.; Fu, X.; Zheng, L.; Xiao, Z.; Zhao, M. Comparison of aromaactive compounds in broiler broth and native chicken broth by aroma extract dilution analysis (AEDA), odor activity value (OAV) and omission experiment. Food Chem. 2018, 265, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Sidira, M.; Kandylis, P.; Kanellaki, M.; Kourkoutas, Y. Effect of immobilized Lactobacillus casei on the evolution of flavor compounds in probiotic dry-fermented sausages during ripening. Meat Sci. 2015, 100, 41–51. [Google Scholar] [CrossRef]
- Sidira, M.; Kandylis, P.; Kanellaki, M.; Kourkoutas, Y. Effect of immobilized Lactobacillus casei on volatile compounds of heat treated probiotic dry-fermented sausages. Food Chem. 2015, 178, 201–207. [Google Scholar] [CrossRef]
- Sidira, M.; Kandylis, P.; Kanellaki, M.; Kourkoutas, Y. Effect of curing salts and probiotic cultures on the evolution of flavor compounds in dry-fermented sausages during ripening. Food Chem. 2016, 201, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Bowker, B.C.; Eastridge, J.S.; Solomon, M.B. Measurement of Muscle Exudate Protein Composition as an Indicator of Beef Tenderness. J. Food Sci. 2014, 79, C1392–C1297. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, L.; Xin, Q.; Zhu, Z.; Miao, Z.; Zheng, N. Metabolomic characterization of Liancheng white and Cherry Valley duck breast meat and their relation to meat quality. Poultry Sci. 2023, 102, 103020. [Google Scholar] [CrossRef]
- Weng, K.; Song, L.; Bao, Q.; Cao, Z.; Zhang, Y.; Zhang, Y.; Chen, G.; Xu, Q. Comparative Characterization of Key Volatile Compounds in Slow- and Fast-Growing Duck Raw Meat Based on Widely Targeted Metabolomics. Foods. 2022, 11, 3975. [Google Scholar] [CrossRef]
- Ge, Y.; Gai, K.; Li, Z.; Chen, Y.; Wang, L.; Qi, X.; Xing, K.; Wang, X.; Xiao, L.; Ni, H.; Guo, Y.; Chen, L.; Sheng, X. HPLC-QTRAP-MS-based metabolomics approach investigates the formation mechanisms of meat quality and flavor of Beijing You chicken. Food Chem.: X, 2023, 17, 100550. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.P.; Cui, H.X.; Liu, R.R.; Zheng, M.Q.; Chen, J.L.; Wen, J. Comparison of breast muscle meat quality in 2 broiler breeds. Poultry Sci. 2011, 90, 2355–2359. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Ge, C.; Zhou, G.; Zhang, W.; Liao, G. 1H NMR-based metabolic characterization of Chinese Wuding chicken meat. Food Chem. 2019, 274, 574–582. [Google Scholar] [CrossRef] [PubMed]
- European Commission, Communication from the Commission to the European Parliament and the Council on Foods and food ingredients authorized for treatment with ionizing radiation in the Community, Brussels, 2001, Document 52001DC0472, https://eur-lex.europa.eu/legal-content/EN/TXT/? 5200.
- Zhao, L.; Zhang, Y.; Pan, Z.; Venkitasamy, C.; Zhang, L.; Xiong, W.; Guo, S.; Xia, H.; Liu, W. Effect of electron beam irradiation on quality and protein nutrition values of spicy yak jerky. Lebensm. Wiss. Technol. 2018, 87, 1–7. [Google Scholar] [CrossRef]
- Jung, Y.; Lee, J.; Kwon, J.; Lee, K.S.; Ryu, D.H.; Hwang, G.S. Discrimination of the geographical origin of beef by 1H NMR-based metabolomics. J. Agric. Food Chem. 2010, 58, 10458–10466. [Google Scholar] [CrossRef]
- Zhou, H.; Yang, Y.; Wang, L.; Ye, S.; Liu, J.; Gong, P.; Qian, Y.; Zeng, H.; Chen, X. Integrated multi-omic data reveal the potential molecular mechanisms of the nutrition and flavor in Liancheng white duck meat. Front Genet, 2022, 13, 939585. [Google Scholar] [CrossRef] [PubMed]
- Wood, A.; Senn, M.; Gadgil, M.; Graca, G.; Allison, M.; Tzoulaki, I.; Greenland, P.; Ebbels, T.; Elliott, P.; Goodarzi, M.; Tracy, R.; Rotter, J. Herrington, D. Untargeted Metabolomic Analysis Reveals Inverse Associations Between Red Meat Intake and Two Anti- Inflammatory Amino Acids. Curr. Dev. Nutr. 2023, 7, 101326. [Google Scholar] [CrossRef]
- Wood, A.C.; Graca, G.; Gadgil, M.; Senn, M.K.; Allison, M.A.; Tzoulaki, I.; Greenland, P.; Ebbels, T. , Elliott, P.; Goodarzi, M.O.; Tracy, R,; Rotter, J.I.; Herrington D. Untargeted metabolomic analysis investigating links between unprocessed red meat intake and markers of inflammation. Am. J. Clin. Nutr. 2023, 118, 989–999. [Google Scholar] [CrossRef]
- Chen, C.; Shi, Z.; Fan, X.; Du, L.; Zhou, C.; Pan, D. Combined application of high-throughput sequencing and LC-MS-based lipidomics in the evaluation of microorganisms and lipidomics of restructured ham of different salted substitution. Food Res Int. 2023, 174, 113596. [Google Scholar] [CrossRef]
- Chen, J.N.; Zhang, Y.Y.; Huang, X.; Wang, H.P.; Dong, X.; Zhu, B. Analysis of Lipid Molecule Profiling and Conversion Pathway in Mandarin Fish (Siniperca chuatsi) during Fermentation via Untargeted Lipidomics. J. Agric. Food Chem. 2023, 71, 8673–8684. [Google Scholar] [CrossRef]
- Guo, X.; Shi, D.; Liu, C.; Huang, Y.; Wang, Q.; Wang, J. UPLC-MS-MS-based lipidomics for the evaluation of changes in lipids during dry-cured mutton ham processing. Food Chem. 2022, 377, 131977. [Google Scholar] [CrossRef] [PubMed]
- Mu, Y.; Su, W.; Mu, Y.; Jiang, L. Combined Application of High-Throughput Sequencing and Metabolomics Reveals Metabolically Active Microorganisms During Panxian Ham Processing. Front Microbiol. 2019, 10, 3012. [Google Scholar] [CrossRef] [PubMed]
- Fahy, E.; Cotter, D.; Sud, M.; Subramaniam, S. Lipid classification, structures and tools. Biochim Biophys Acta, 2011, 1811, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Sun, T. , Wang, X., Cong, P., Xu, J., & Xue, C. Mass spectrometry-based lipidomics in food science and nutritional health: A comprehensive review. Comprehensive Reviews in Food Science and Food Safety 2020, 19, 2530–2558. [Google Scholar]
- Bourlieu, C. , Cheillan, D., Blot, M., Daira, P., Trauchessec, M., Ruet, S., & Leconte, N. J. Polar lipid composition of bioactive dairy co-products buttermilk and butterserums: Emphasis on sphingolipid and ceramide isoforms. Food Chemistry, 2017; 240, 67–74. [Google Scholar]
- Mi, S.I.; Shang, K.E.; Li, X.; Zhang, C.H.; Liu, J.Q.; Huang, D.Q. Characterization and discrimination of selected China’s domestic pork using an LC–MS-based lipidomics approach. Food Control, 2019, 100, 305–314. [Google Scholar] [CrossRef]
- Li, C.; Li, X.; Huang, Q.; Zhuo, Y.; Xu, B.; Wang, Z. Changes in the phospholipid molecular species in water-boiled salted duck during processing based on shotgun lipidomics. Food Res. Int. 2020, 132, 109064. [Google Scholar] [CrossRef]
- Li, J.; Tang, C.; Zhao, Q.; Yang, Y.; Li, F.; Qin, Y.; Zhang, J. Integrated lipidomics and targeted metabolomics analyses reveal changes in flavor precursors in psoas major muscle of castrated lambs. Food Chem. 2020, 333, 127451. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Cao, J.; Bao, Y.; Sun, Z.; Liu, Z.; Li, C. Characterization of lipid profiling in three parts (muscle, head and viscera) of Tilapia (oreochromis niloticus) using lipidomics with UPLC-ESI-Q-TOF-MS. Food Chem. 2021, 347, 129057–129066. [Google Scholar] [CrossRef] [PubMed]
- Song, G.; Zhang, M.; Zhang, Y.; Wang, H.; Li, S.; Dai, Z.; Shen, Q. In situ method for real-time discriminating salmon and rainbow trout without sample preparation using iKnife and rapid evaporative ionization mass spectrometry-based lipidomics. J. Agric. Food Chem. 2019, 67, 4679–4688. [Google Scholar] [CrossRef]
- Song, G.; Wang, Q.; Zhang, M.; Yang, H.; Xie, H.; Zhao, Q. Real-time in situscreening of omega-7 phospholipids in marine biological resources using an iKnife-rapid-evaporative-lonization-mass-spectrometry-based lipidomics phenotype. J. Agric. Food Chem. 2021, 69, 9004–9011. [Google Scholar] [CrossRef]
- Jia, W.; Shi, Q.; Shi, L. Effect of irradiation treatment on the lipid composition and nutritional quality of goat meat. Food Chem, 2021, 351, 129295. [Google Scholar] [CrossRef] [PubMed]
- Benet, I. ; Dolors, Guardia, M. ; Ibanez, C.; Sola, J.; Arnau, J.; Roura, E. Analysis of SPME or SBSE extracted volatile compounds from cooked cured pork ham differing in intramuscular fat profiles. LWT-Food Sci. Technol. 2015, 60, 393–399. [Google Scholar] [CrossRef]
- Marusic, N.; Vidacek, S.; Janci, T.; Petrak, T.; Medic, H. Determination of volatile compounds and quality parameters of traditional Istrian dry-cured ham. Meat Sci. 2014, 96, 1409–1416. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Guo, H.; Wu, T.; Tao, N.; Wang, X.; Zhong, J. Effect of three types of thermal processing methods on the lipidomics profile of tilapia fillets by UPLC-Q-Extractive Orbitrap mass spectrometry. Food Chem. 2019, 298, 125029. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Li, X.; Huang, A. A metabolomics-based approach investigates volatile flavor formation and characteristic compounds of the Dahe black pig dry-cured ham. Meat Sci. 2019, 158, 107904. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Guo, M.; Wang, Q.; Dong, J.; Lu, S.; Lyu, B.; Ma, X. Antioxidant activities of peptides derived from mutton ham, Xuanwei ham and Jinhua ham. Food Res. Int., 2021, 142, 110195. [Google Scholar] [CrossRef]
- Petron, M.J.; Muriel, E.; Timon, M.L.; Martin, L.; Antequera, T. Fatty acids and triacylglycerols profiles from different types of Iberian dry-cured hams. Meat Sci. 2004, 68, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Alexandri, E.; Ahmed, R.; Siddiqui, H.; Choudhary, M.; Tsiafoulis, C.; Gerothanassis, I. High resolution NMR spectroscopy as a structural and analytical tool for unsaturated lipids in solution. Molecules, 2017, 22, 1663–1735. [Google Scholar] [CrossRef]
- Lankhorst, P. P.; Chang, A.N. The application of NMR in compositional and quantitative analysis of oils and lipids. In Modern magnetic resonance, Webb, G.A., Eds.; Springer, 2017; pp. 1–22. [CrossRef]
- Li, J.; Vosegaard, T.; Guo, Z. Applications of nuclear magnetic resonance in lipid analyses: An emerging powerful tool for lipidomics studies. Prog. Lipid Res. 2017, 68, 37–56. [Google Scholar] [CrossRef]
- Pajuelo, A.; Sánchez, S.; Pérez-Palacios, T.; Caballero, D.; Díaz, J. , Antequera, T.; Marcos, C.F. 1H NMR to analyse the lipid profile in the glyceride fraction of different categories of Iberian dry-cured hams. Food Chem. 2022, 383, 132371–132380. [Google Scholar] [CrossRef]
- Pajuelo, A.; Ramiro, J. L.; Fernández-Marcos, M. L.; Sánchez, S.; Pérez-Palacios, T.; Antequera, T.; Marcos, C.F. Lipidomic analysis and classification of Iberian dry-cured hams with low field NMR. Food Frontiers. 2023, 4, 1810–1818. [Google Scholar] [CrossRef]
- Harlina, P. W.; Maritha, V.; Geng, F.; Subroto, E.; Yuliana,T. ; Shahzad, R.; Sun J. Lipidomics: a comprehensive review in navigating the functional quality of animal and fish products. Int. J. Food Prop., 2023, 26, 2–3115. [Google Scholar] [CrossRef]
- Jin, R.; Li, L.; Feng, J.; Dai, Z.; Huang, Y.W.; Shen, Q. Zwitterionic hydrophilic interaction solid-phase extraction and multi-dimensional mass spectrometry for shotgun lipidomic study of Hypophthalmichthys nobilis. Food Chem. 2017, 216, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Wang, Y.; Gong, L.; Guo, R.; Dong, W.; Cheung, H.Y. Shotgun lipidomics strategy for fast analysis of phospholipids in fisheries waste and its potential in species differentiation. J. Agric. Food Chem., 2012, 60, 9384–9393. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Chen, K.; Li, S.; Wang, Y.; Shen, Q. Lipidomics differentiation of soft-shelled turtle strains using hydrophilic interaction liquid chromatography and mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2019, 1112, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Han, X. Accurate quantification of lipid species by electrospray ionization mass spectrometry – Meet a key challenge in lipidomics. Metabolites, 2011, 1, 21–40. [Google Scholar] [CrossRef]
- Yamamoto, S.; Kato, S.; Senoo, N.; Miyoshi, N.; Morita, A.; Miura, S. Differences in phosphatidylcholine profiles and identification of characteristic phosphatidylcholine molecules in meat animal species and meat cut locations. Biosci. Biotechnol. Biochem. 2021, 85, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Xie, D.; Wang, D.; Xu, W.; Zhang, C.; Li, P.; Sun, C. Lipidomic Profile Changes of Yellow-Feathered Chicken Meat during Thermal Processing Based on UPLC-ESI-MS Approach. Food Chem. 2023, 399, 133977. [Google Scholar] [CrossRef] [PubMed]
- Chiesa, L. M.; Di Cesare, F.; Mosconi, G.; Pavlovic, R.; Campaniello, M.; Tomaiuolo, M.; Mangiacotti, M.; Chiaravalle, E.; Panseri, S. Lipidomics Profile of Irradiated Ground Meat to Support Food Safety. Food Chem. 2022, 375, 131700. [Google Scholar] [CrossRef]
- Feng, X.; Li, J.; Zhang, L.; Rao, Z.; Feng, S.; Wang, Y.; Liu, H.; Meng, Q. Integrated Lipidomic Metabolomics Analysis Revealing the Effects of Frozen Storage Duration on Pork Lipids. Metabolites. 2022, 12, 977. [Google Scholar] [CrossRef]
- Daley, C. A.; Abbott, A.; Doyle, P. S.; Nader, G. A.; Larson, S. A. Review of Fatty Acid Profiles and Antioxidant Content in Grass-Fed and Grain-Fed Beef. Nutr. J. 2010, 9, 1–12. [Google Scholar] [CrossRef]
- Lv, J.; Ma, J.; Liu, Y.; Li, P.; Wang, D.; Geng, Z.; Xu. W. Lipidomics analysis of Sanhuang chicken during cold storage reveals possible molecular mechanism of lipid changes. Food Chem. 2023, 417, 135914. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Wu, X.; Zhang, R.; Shi, L. 2022. UHPLC-Q-Orbitrap-based lipidomics reveals molecular mechanism of lipid changes during preservatives treatment of Hengshan goat meat sausages. Food Chem. 2022, 369, 130948. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Cui, H.; Hu, Y.; Li, J.; Yue, S.; Tang, C.; Zhao, Q.; Yu, Y.; Li, H.; Qin, Y. Yang, Y.; Zang, J. Comparative characterization of Taihe silky chicken and cobb chicken using LC/MS-based lipidomics and GC/MS-based volatilomics. Lwt, 2022, 163, 113554. [Google Scholar] [CrossRef]
- Li, C.; Al-Dalali, S.; Zhou, H.; Wang, Z.; Xu, B. 2021. Influence of mixture of spices on phospholipid molecules during water-boiled salted duck processing based on shotgun lipidomics. Food Res. Int. 1106; 51. [Google Scholar] [CrossRef]
- Li, M.; Zhu, M.; Chai, W.; Wang, Y.; Fan, D.; Lv, M.; Jiang, X.; Liu, Y.; Wei, Q.; Wang, C. Determination of lipid profiles of Dezhou donkey meat using an LC-MS-based lipidomics method. J. Food Sci., 2021, 86, 4511–4521. [Google Scholar] [CrossRef] [PubMed]
- Ulmer, C.Z.; Jones, C.M.; Yost, R.A.; Garrett, T.J.; Bowden, J.A. Optimization of Folch, Bligh-Dyer, and Matyash sample-toextraction solvent ratios for human plasma-based lipidomics studies. Anal. Chim. Acta. 2018, 1037, 351–357. [Google Scholar] [CrossRef]
- Jia, W.; Guo, A.; Bian, W.; Zhang, R.; Wang, X.; Shi, L. Integrative deep learning framework predicts lipidomics-based investigation of preservatives on meat nutritional biomarkers and metabolic pathways, Crit. Rev. Food Sci. Nutr. 2023, 1–15. [Google Scholar] [CrossRef]
Figure 1.
Number of publications on the foodomics and meat quality/safety between 2014 and 2024 grouped by (A) year of publication and (B) publication subject (Data was obtained from lens.org).
Figure 1.
Number of publications on the foodomics and meat quality/safety between 2014 and 2024 grouped by (A) year of publication and (B) publication subject (Data was obtained from lens.org).
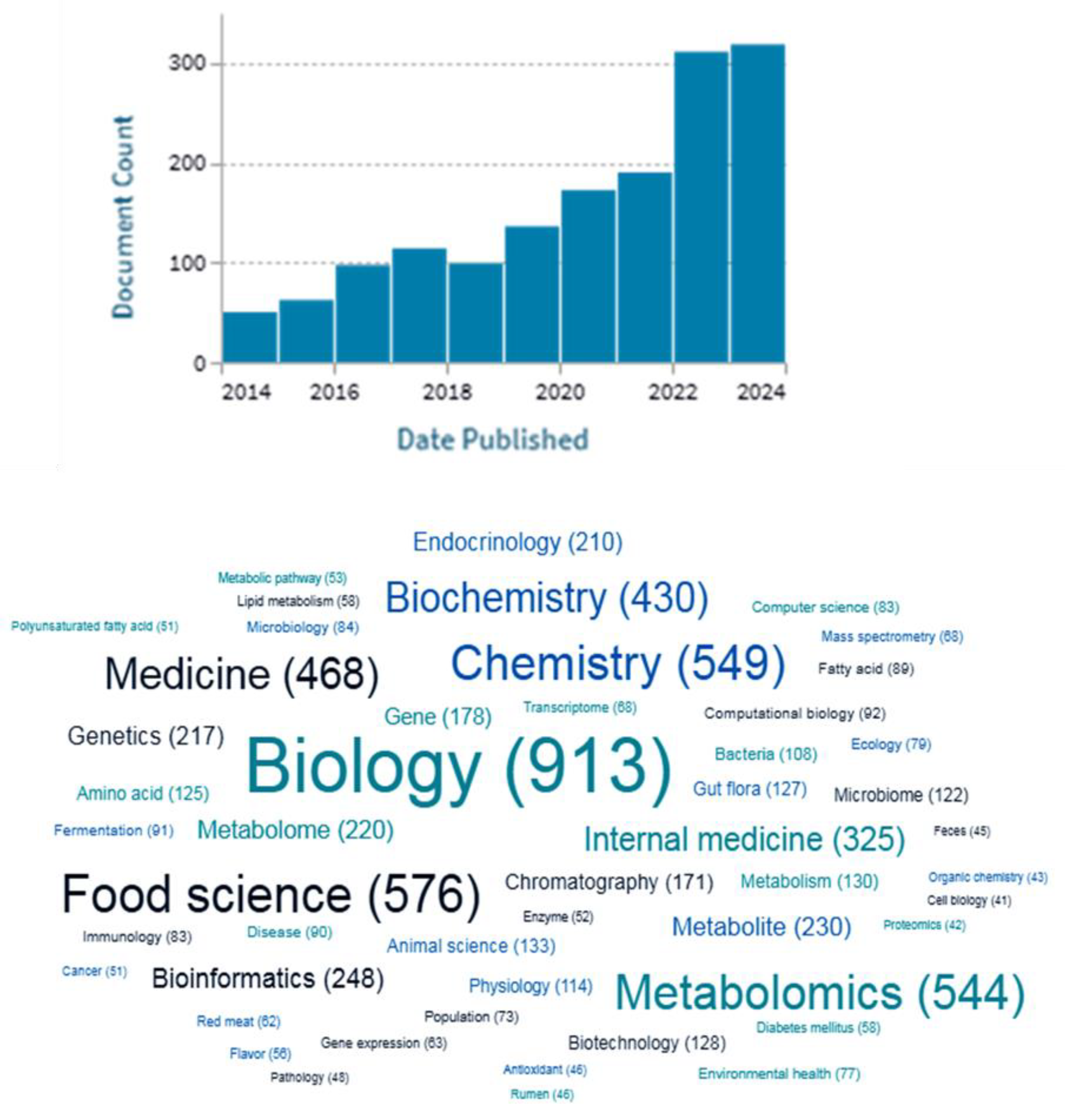
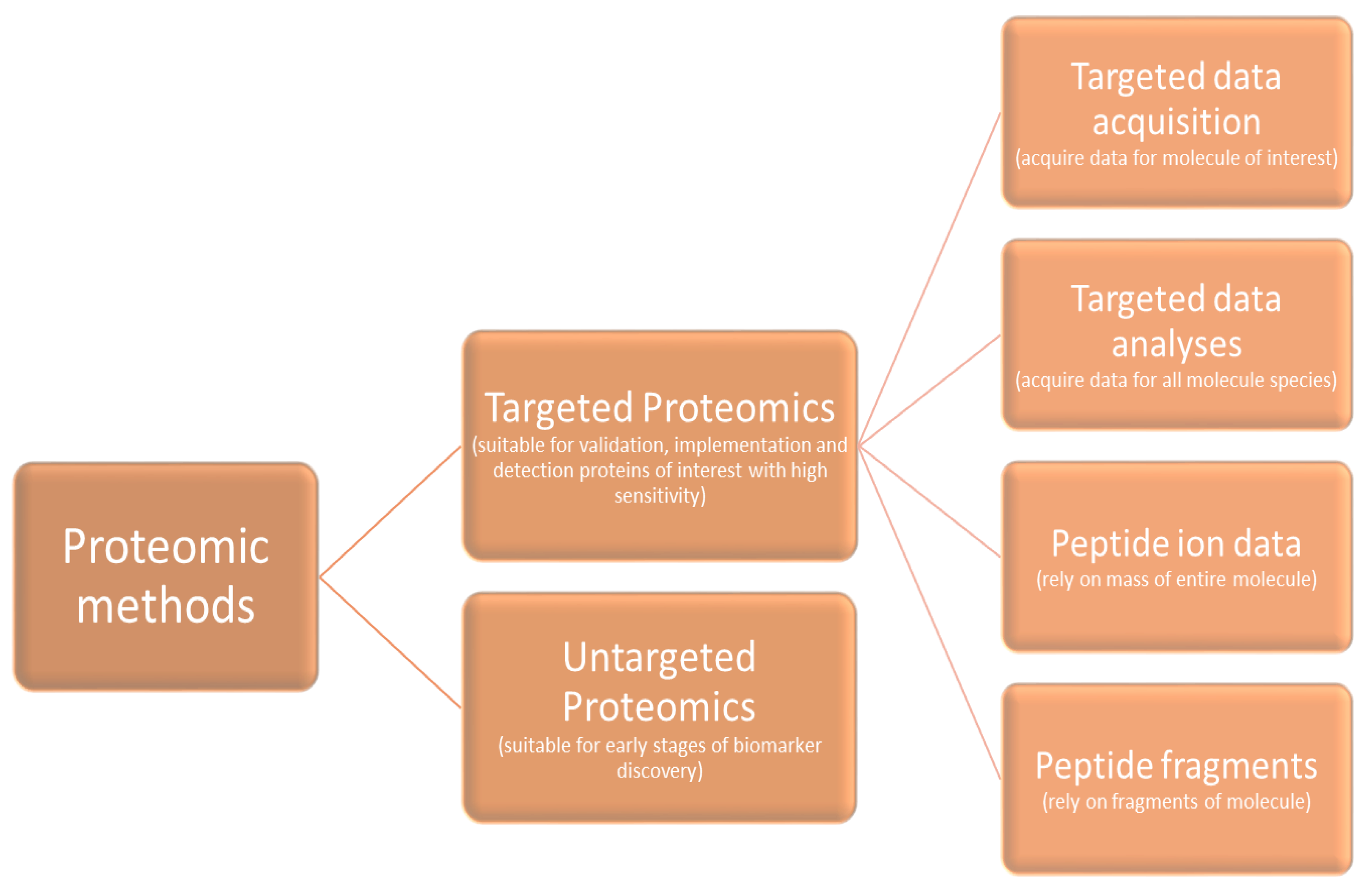
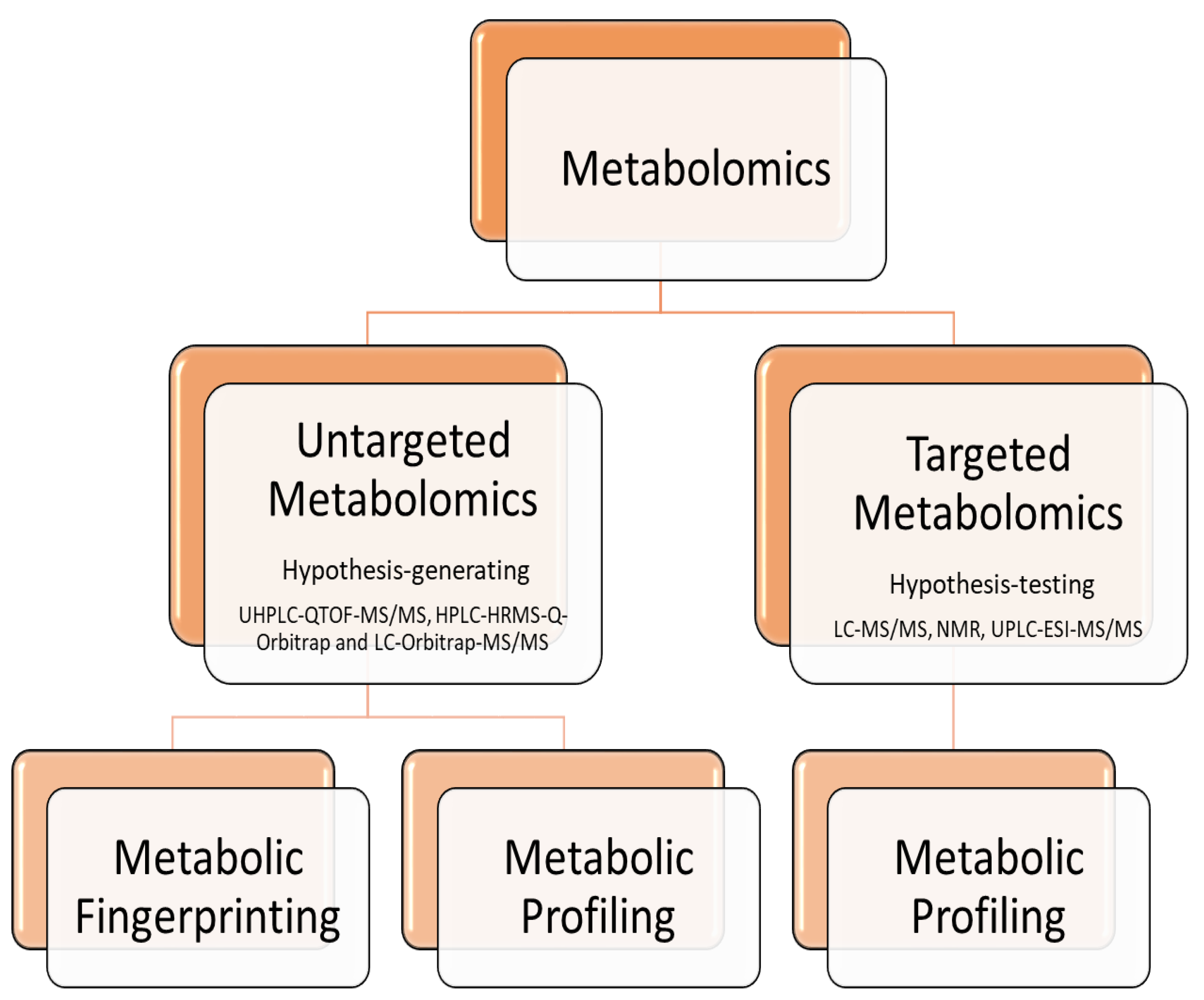
Figure 4.
Lipidomic Approach for determination of the Functional Quality of meat Products (Adapted from Harlina et al. [88]).
Figure 4.
Lipidomic Approach for determination of the Functional Quality of meat Products (Adapted from Harlina et al. [88]).
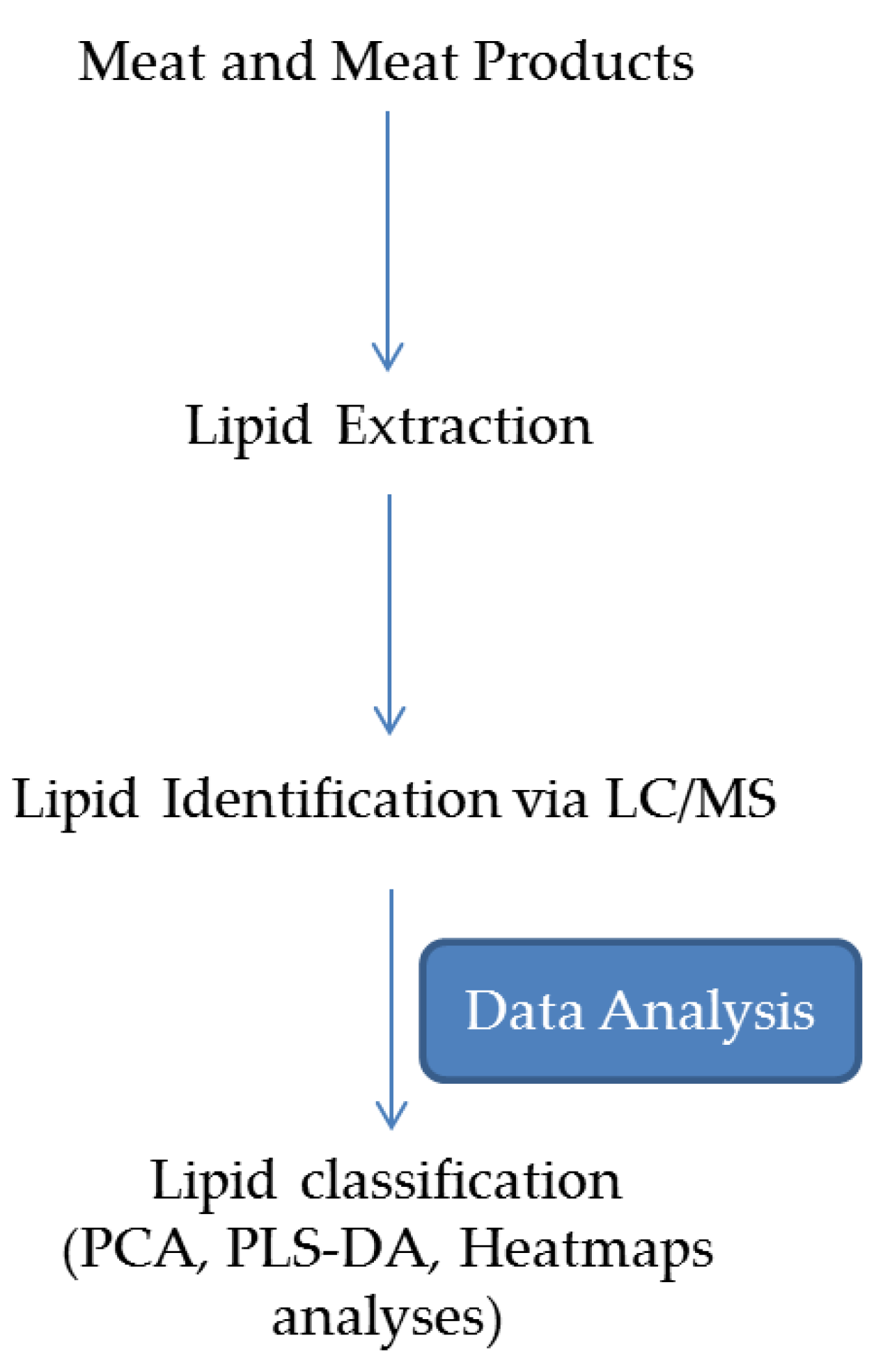
Table 3.
Lipidomics for quality control of meat products (Adapted from Jia et al. [104]; Harlina et al. [88]).
| Meat substrate | Extraction method | Lipid identification methodology | Data analysis | Results | Reference |
|---|---|---|---|---|---|
| Chicken meat | Folch et al. method | UPLC-ESI-MS | PCA, PLS-DA, OPLS-DA | Significant phospholipids’ decrease, lysophospholipid increase | [94] |
| Chicken, turkey and mixed ground meat for sausage preparation | Bligh and Dyer method | GC analysis of fatty acid methyl esters, HPLC Q-Exactive Orbitrap high resolution mass spectrometry for lipidomics analysis | PCA, Volcano plot | Identification of 345 lipids categorized into 14 subclasses. Identification of oxidized glycerophosphoethanolamines and oxidized glycerophosphoserines in irradiated turkey meat | [95] |
| Pork | Folch et al. method (from Ulmer et al. 103) | Ultra-Performance Liquid Chromatography coupled with triple-quadrupole mass spectrometry | PCA and OPLS-DA analysis | ether-linked phosphatidylethanolamine and phosphatidylcholine containing more than one unsaturated bond were greatly influenced by frozen storage |
[96] |
| Grass-fed and grain-fed beef | - | - | - | Variations in the fatty acid composition between grass-fed and grain-fed beef. Grass-based diets have been shown to enhance total conjugated linoleic acid (CLA) (C18:2) isomers, trans vaccenic acid (TVA) (C18:1 t11), a precursor to CLA, and omega-3 (n-3) FAs |
[97] |
| Dry-cured muton ham | lipid extraction buffer (MTBE: Methanol = 3:1, v/v) | lipid metabolomics based on UPLC-MS-MS. | PCA and OPLS-DA | Most abundant lipids were glycerolipids (GL) followed by glycerophospholipids. Quality of mutton ham changed during the P3 fermenting stage |
[65] |
| Chicken breast | Soxhlet extraction | Ultra-high performance liquid chromatography coupled with mass spectrometry (UHPLC-MS)- | Volcano plot analysis | Triacylglycerol (TAG), phosphatidylcholine (PC) and phosphatidylethanolamine (PE) significantly decreased | [98] |
| Hengshan goat meat sausages | LC-ESI –MS (Q-Orbitrap | lipid variables related to glycerophospholipid and sphingolipid metabolism | [99] | ||
| Chicken | Soxhlet extractio | UPLC-Q-Exactive Orbitrap/MS | PCA, PLS-DA, PCA of E-tongue | significant differences between Cobb chicken and Taihe silky chicken lipids at the taxonomic and molecular levels | [100] |
| Duck | Phospholipid extraction according to previous methodology | DI -ESI –MS (Q-Trap) | PCA, PLS-DA | The spices had a significant effect on individual phospholipid molecules during processing | [101] |
| Donkey meat | FAME by GC, Muscle lipids were extracted with CHCl3:CH3OH (2:1, v/v | LC –MS (Triple TOF) |
OPLS-DA, heatmap analysis | 1143 lipids belonging to 14 subclasses were identified in donkey meat, of which 73 lipids (23 upregulated and 50 downregulated) including glycerolipids (GLs), glycerophospholipids (GPs) and sphingolipids (SPs) | [102] |
| Camel meat | lipid fraction was extracted with MTBE | UPLC-Q-TOF/MS | PCA, OPLS-DA, volcano plot | 342 lipid species were detected, 192, 64, and 79 distinguishing lipids were found in the groups camel hump compared to camel meat, camel meat compared to beef, and camel hump compared to fatty-tails, respectively | [16] |
| Goat meat irradiated | dual-phase extraction with methanol and MTBE | UHPLC–Q-Orbitrap | PCA, PLS-DA | 12 subclasses of 174 lipids were identified with significant differences ( | [76] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



