
Submitted:
23 May 2024
Posted:
24 May 2024
You are already at the latest version
Abstract
The increasing emergence of multidrug resistant (MDR) pathogens causes difficult-to-treat infections, with long-term hospitalizations and high incidence of death, thus representing a global public health problem. To manage MDR bacteria bugs, new antimicrobial strategies are necessary, and their introduction in practice is a daily challenge for scientists in the field. An extensively studied approach to treat MDR infections consists in inducing high levels of reactive oxygen species (ROS) by several methods. Although further clinical investigations are mandatory on the possible toxic effects of ROS to mammalian cells, clinical evaluations are extremely promising, and their topical use to treat infected wounds and ulcers, also in presence of biofilm, is already clinically approved. Biochar (BC) is a carbonaceous material obtained by pyrolysis of different vegetable and animal biomass feedstocks, at 200−1000 °C in limited presence of O2. Recently, it has been demonstrated that BC capability of removing organic and inorganic xenobiotics is mainly due to the presence of persistent free radicals (PFRs), which can activate oxygen, H2O2 or persulfate in the presence or not of transition metals by electron transfer thus generating ROS, which in turn degrade pollutants by advanced oxidation processes (AOPs). In this context, the antibacterial effects of BC containing PFRs have been demonstrated by some authors against Escherichia coli and Staphylococcus aureus, thus giving birth to our idea of the possible use of BC-derived PFRs as a novel method capable of inducing ROS generation for antimicrobial oxidative therapy. Here, the general aspects concerning ROS physiological and pathological production and regulation, and the mechanism by which they could exert antimicrobial effects have been reviewed. The methods currently adopted to induce ROS production for antimicrobial oxidative therapy have been discussed. Finally, for the first time, BC-related PFRs have been proposed as a new source of ROS for antimicrobial therapy via AOPs.
Keywords:
multidrug resistant pathogens
; antimicrobial oxidative therapy
; reactive oxygen species (ROS)
; biochar (BC)
; biochar-derived permanent free radicals (PFRs)
; advanced oxidation processes (AOPs)
1. Introduction
The incessant and rapid increase of multi drug resistant (MDR) pathogens causes the emergence of difficult-to-treat infections, with long-term hospitalizations, high-costs, and a frightening incidence of death. In the United States, more than 2.8 million antibiotic-resistant infections occur each year, resulting in 35,000 deaths [1]. Resistant bacteria are becoming an uncontrollable worldwide hazard to both humans and animals [2,3]. Management of the antimicrobial resistance through the available antibiotic is a global public health problem and represents a daily challenge for experts in the field. To limit the global antibiotics resistance crisis, the main solution would be to reduce the volume of the unscrupulous use of antibiotics both in medicine, agriculture and in the environment [4], as well as to perform an efficient infection control strategy to prevent the spread out of contagions. Anyway, the development of novel antimicrobial drugs remains urgent and mandatory [1]. Although several new agents based on existing classes of antibiotics are being developed, there has been little advancement on the exclusive discovery of novel agents [4].
Moreover, the capability of several bacterial and fungal species to form biofilms is an added alarming mechanism through which pathogens develop a very complex form of resistance. Biofilm producing pathogens represent a significant problem in many clinical settings, since biofilm further increases their tolerance towards conventionally prescribed antimicrobials [5,6]. The high-dosage use of antibiotics in biofilm conditions to treat chronic wounds, burns, chronic respiratory diseases, cystic fibrosis and recurrent cystitis leads to intense selective pressure, which paradoxically drives to further antibacterial resistance [4].
On this alarming scenario, the need for the development of optional per se effective therapeutic approaches, or of alternative treatments which can improve the antimicrobial efficacy of existing drugs also towards biofilms, is imperative [7]. This second strategy would allow to reduce the amounts of antibiotics to be used thus limiting the emergence of resistance in pathogens. Entirely novel antimicrobial instruments characterized by a unique mechanism of action are represented by reactive oxygen species (ROS) induced by different methods [4].
ROS have demonstrated in vitro and in vivo a significant antimicrobial action against a wide spectrum of Gram-positive and Gram-negative organisms, including MDR isolates and biofilm-producing pathogens [8]. The use of ROS could represent a new therapeutic approach for topical use on skin, mucosal membranes, or internal tissue that may be colonized with microbial inhabitants and biofilms [9].
Treatments involving ROS as antimicrobial agents are already available for topical application and are clinically approved to treat infected wounds and are being developed for clinical use also in other settings [10].
As mentioned above, ROS is the well-known acronym used in several sectors including medicine, to indicate reactive oxygen species, including radical and not radical oxygen-containing atoms and molecules, such as superoxide anion (O2−), singlet oxygen (1O2), peroxide (O2−2), hydrogen peroxide (H2O2), hydroxyl radicals (OH•), and hydroxyl anions (OH−), that are constantly being formed as byproducts of the physiologic aerobic metabolism of cells [11]. Additionally, ROS can react with NO produced by cells from intracellular L-arginine via action of epithelial nitrogen oxide synthetase (NOS), neuronal NOS, and inducible NOS, forming other reactive species such as NO• and ONOO−, while NO• in combination with O2, provides ONOO•. These molecules are referred to as reactive nitrogen species (RNS) [11].
In normal conditions, ROS and RNS generation is kept under control by the antioxidant defenses and repair systems of cell [12]. On the contrary, when overproduced, the detoxification systems of cell fail to maintain ROS and RNS physiological levels, which accumulate, thus causing the onset of oxidative stress (OS) and inflammation. Irreversible damage to DNA, lipids, and proteins occurs, thus promoting aging, age-related diseases, and several degenerative human disorders [13].
Collectively, OS is a cascade of events that frequently triggers and accompanies molecular/cellular pathogenic events. It is responsible for several human disorders, including carcinogenesis [14,15], atherosclerosis, cardiovascular, and neurodegenerative diseases [16,17].
As occurring in humans, also in pathogens, OS builds up when prooxidants overpower antioxidants. Therefore, ROS get accumulated in the microorganism’s cell, thus exceeding the cell’s capacity to readily detoxify them [18].
As examples, the host immune response, as well as several antimicrobials counteract infections by inducing ROS accumulation. While the interaction between the host and pathogens causes exogenous OS in bacteria, the intracellular redox reactions, antibiotics, and not controlled aerobic respiration contribute to endogenous OS [19]. ROS cause multiple damages to the bacterial cells, including double stranded breaks in DNA by oxidizing dCTP and dGTP pools, which results in the misincorporation of bases into DNA. Additionally, ROS induce lipids peroxidation, and proteins carbonatization [20], thus exerting a very rapid bactericidal activity. It has been reported that ROS were able to cause 3 log CFU reduction in 30 min and total eradication in 2 hours when used against Staphylococcus aureus [21]. Several conventional and alternative antibiotics including metal nanoparticles and natural molecules exert their antimicrobial properties by inducing ROS hyperaccumulation in pathogens [22]. Other methods to exert ROS-mediated antimicrobial effects, include photodynamic therapy (PDT), honey reactive oxygen (HRO) therapy, and hyperbaric oxygen treatment (HBOT).
On these considerations, ROS could really represent an effective option for eradicating MDR pathogens. While procedures to induce ROS in microbial cells, such as HBOT and PDT are traditional, the use of nanomaterials and engineered medical honey are rather novel promising methods. Nevertheless, nanotoxicology is a field still not clearly defined, which lacks sufficient and unequivocable epidemiologic data, information, and regulation. Furthermore, such methods may induce ROS formation also in host cells. To make possible an enlargement of the clinical use of ROS to counteract MDR pathogens and related biofilm, the development of other delivery strategies for increasing the selectivity of ROS for microbial pathogens over the host tissue is necessary. In this regard, based on our recent studies on biochar (BC) [23,24], we profit of this review to propose a possible innovative method to be studied to produce ROS by a natural low-cost source. Biochar (BC) is a carbonaceous material obtained by pyrolysis of different vegetable and animal biomass feedstocks and waste, at 200−1000 °C in the limited presence of absence of oxygen. Due to its strong adsorption capacity, BC can exert a plethora of beneficial effect, including the removal of environmental pollutants and xenobiotics, thus preventing their uptake in plants, animals, and humans [25,26,27,28]. In microbiology, BC has demonstrated to be helpful in limiting the antimicrobial resistance by degrading/removing residual antibiotics in soil and waters [24]. The so called, environmentally persistent free radicals (EPFRs) are known to exist in significant concentration in atmospheric particulate matter (PM) and are primarily emitted from combustion and thermal processing of organic materials. While their existence in combustion has been known for over half-a-century, only recently their presence in environmental media and their healthy and/or hazardous effects have been researched [29]. Nowadays, it has been demonstrated that also BC can contain persistent free radicals (PFRs) as well, bound to the external or internal surfaces of its solid particles [29]. PFRs are reactive species due to unpaired electrons, that can persist up to several months, in contrast to traditional transient radicals [24]. Studies reported that PFRs are the main responsible for the BC capacity to degrade organic pollutants through the generation of active oxygen species (ROS) and sulfate radicals [23]. It was reported that the generated ROS, including radical (•OH, •O2−, •O2H, SO4•-) and non-radical species (1O2) successfully degrade several organic pollutants, hormones and eDNA, by advanced oxidation processes (AOPs). Interesting, PFRs-mediated ROS showed antibacterial effects against Escherichia coli and S. aureus [30,31,32], thus supporting the idea of the possible use of BC-derived PFRs as a novel method to induce ROS generation for antimicrobial oxidative therapy. In the following sections, all that was introduced in this section will be more in deep reviewed and discussed.
2. Reactive Oxygen Species (ROS) and Oxidative Stress (OS)
2.1. Physiological and Pathological Origin of ROS
Figure 1 schematizes the main processes by which ROS can form in cells and the detrimental effects they can determine to health [11].
Table 1 reports the possible enzymatic and not enzymatic endogenous sources of ROS, as well as those exogenous. The main reactive species of both oxygen and nitrogen which can be consequently produced have been also included.
Based on the recently acquired knowledge and literature reports [33,34,35,36], respect to the Table reported by us in 2020 [11], environmental persistent free radicals (EPFRs) and biochar-related persistent free radicals (BC-PFRs) have been included among the exogenous sources of ROS in Table 1. Upon their formation according to reported processes and mechanisms the can induce ROS formation by reacting with atmospheric or water dissolved O2, as well as with H2O2 and/or persulfate [24]. Briefly, phagocytic cells (neutrophils, monocytes, or macrophages) use NOX for one-electron reduction of molecular oxygen to the radical superoxide anion (O2•−), during cellular respiration. O2•− is considered the primary ROS, and when reacts with other molecules through enzymatic or non-enzymatic metals-catalyzed processes, generates secondary ROS.
O2•− is mainly transformed by superoxide dismutase (SOD) into the hydrogen peroxide (H2O2), from which the highly reactive ROS hydroxyl ion (•OH) and radical HOO•, are formed through the Fenton or Haber–Weiss reactions in presence of transition metals. O2•− is also produced from the irradiation of molecular oxygen with UV rays, photolysis of water, and by exposure of O2 to organic radicals formed in aerobic cells such as NAD•, FpH•, semiquinone radicals, cation radical pyridinium, or by hemoproteins.
While the radical O2•− does not react directly with lipids, polypeptides, sugars, or nucleic acids, •OH and HOO• react especially with phospholipids in cell membranes and proteins, thus causing oxidative damage, DNA damage, telomere reduction, ageing and apoptosis [11].
Furthermore, H2O2 can be converted by MPO to hypochlorous acid, particularly hazardous for cellular proteins [37].
Additionally, ROS can react with NO produced from intracellular L-arginine by cells as a defense mechanism, using three different kinds of NOS, such as epithelial NOS, neuronal NOS, and inducible NOS, thus forming reactive nitrogen species (RNS) such as NO• and ONOO−. Finally, NO• in combination with O2, can provide ONOO•, which induces lipid peroxidation in lipoproteins [11,38,39,40].
Table 2 summarizes the specific source of the most representative radical and not radical reactive species produced in biological aerobic systems and their physiological function.
Whatever their origin, ROS and RNS cause indifferently detrimental oxidative modifications of cellular macromolecules such as carbohydrates, lipids, proteins, DNA, and RNA, thus determining the production of molecules, which are considered markers of OS (Table 3).
To counteract the detrimental effects reported in Table 3, cells have developed several repair systems able to restore or eliminate lipids, proteins, and DNA damaged by the action of ROS and RNS. Particularly, cytosolic and mitochondrial enzymes which include polymerases, glycosylases, and nucleases, repair the damaged DNA, while proteinases, proteases, and peptidases make part of the proteolytic enzymes, remove damaged proteins. In addition, biological systems have developed both physiological and biochemical mechanisms to limit free radicals’ production and reactive species toxicity. At physiological level, the microvascular system exerts the function of maintaining the levels of O2 in the tissues, while at biochemical level a protective activity is exerted both by endogenous (enzymatic and non-enzymatic) and exogenous molecules, as reported in Table 4.
2.2. Pathogens Responses to OS
As reported previously in the introduction, ROS cause multiple damages to the bacterial cells [22]. Anyway, bacteria can evade OS by several defenses including detoxifying methods using enzymes like catalase, alkyl hydroperoxide reductase, thioredoxin, and superoxide dismutase (SOD). Additionally, they use pigments like carotenoids; metal homeostasis, and repairing devices including DNA restoration, general stress response, and SOS response [18,46]. All these mechanisms are regulated by gene networks [46]. E. coli reacts to OS mainly producing SOD and catalase, which convert O2•− to H2O2 (SOD) and in turn H2O2 in H2O and O2 (catalase). While mammalian cells possess two types of SOD, E. coli owns three isoforms of SOD characterized by different metal cores. Particularly, sodA contains Mn, sodB includes Fe, and sodC comprises both Cu and Zn. E. coli has also two types of catalases, namely hydro-peroxidase I and hydro-peroxidase II [47]. Also, E. coli has several major regulators activated during OS such as OxyR, SoxRS, OhrR,and RpoS. OxyR and SoxR control the catalases and SODs transcription in relation to the O2•− and H2O2 concentration. They undergo conformation changes when oxidized in the presence of hydrogen peroxide and superoxide radicals, respectively, and subsequently control the expression of cognate genes [48]. In contrast, the RpoS regulon is induced by an increase in RpoS levels. It is a specialized sigma factor which governs the expressions of gene that lead to general stress resistance of cells [49]. These genes may be involved in eliminating oxidative agents, in repairing systems of affected biomolecules and in maintaining normal cellular physiologic circumstances. Despite the enormous genomic diversity of bacteria, OS response regulators present in E. coli are functionally conserved in a wide range of bacterial groups. Bacteria have developed complex, adapted gene regulatory responses to OS, probably due to the high level of ROS produced endogenously through their basic metabolism. Additionally, several bacterial pathogens prevent the increase of ROS by directly inhibiting the synthesis of NADPH oxidase [50]. Iron homeostasis and remodeling of metabolism are two other methods by which bacteria lessen the damage caused by ROS. Bacteria can remodel their metabolism via upregulation of the glycoxylate shunt, thus reducing endogenous ROS formation, or by redirecting the metabolism toward the pentose phosphate pathway and augmenting the production of NADH which refills the level of antioxidants. Ketoacids such as pyruvate and α-ketoglutarate can decarboxylate in the presence of ROS, thus originating toxic molecules and diminishing damage caused by ROS [51]. Iron, which is involved also in ROS generation by Fenton reactions, is crucial for the growth and survival of bacteria and paradoxically iron acquisition by siderophore action is pivotal to counteract OS [52]. Siderophores are compounds that bacteria produce when intracellular iron concentration is low to facilitate its uptake [53]. Two siderophores namely Staphyloferrin A and B have found to enhance the resistance to OS in S. aureus, while E. coli produces an enterobactin siderophore to alleviate damage by OS [51,52]. It was demonstrated that OS in turn regulates the bacterial siderophore production [53]. When E. coli was exposed to H2O2 and paraquat, the expression of enterobactin increased also in presence of high concentration of iron, which reduced the sensitivity of the isolate to both H2O2 and paraquat [53]. Similarly, when methicillin resistant S. aureus (MRSA) was exposed to the antimicrobial surface coating AGXX®, siderophore biosynthesis genes were highly upregulated. Some bacterial species produce biofilm as a highly specialized and organized form of resistance in which bacteria cooperate and stay protected by a self-produced biomass [54]. Persisters are bacterial cells in a dormant state with low metabolic activity existing in biofilm that showcase high antibiotic tolerance, can recolonize post-therapy [55], are less sensitive to ROS and demonstrated increased expression of efflux pumps. Efflux pumps major expression is another mechanism to react to OS which allows bacteria to pump out the ROS-damaged proteins [56].
3. Antimicrobial Oxidative Therapies: Available Methods to Induce ROS Formation
As previously reported in the Introduction, the induction of high levels of reactive oxygen species (ROS) by several procedures thus causing OS detrimental for bacteria cells, has been extensively studied for inhibit several species of Gram-positive and Gram-negative bacteria, viruses, and fungi. The use of ROS represents a new therapeutic approach for topical use on skin, mucosal membranes, or internal tissue that may be colonized with microbial inhabitants and biofilms [57]. It was found that the antibacterial effect of several conventional and alternative antibiotics, metal nanoparticles and natural molecules is based also on their capability of inducing ROS hyperaccumulation in pathogens [22]. Therefore, other methods have been developed and are clinically applied or in clinical trials to produce ROS finalized to antimicrobial oxidative therapy. They include the photodynamic therapy (PDT), the honey reactive oxygen (HRO) therapy, and hyperbaric oxygen treatments (HBOT) [57].
3.1. ROS Formation Induced by Conventional, Alternative and Natural Antimicrobials
Table 5 reports information concerning conventional and alternative antimicrobials including some antibiotics, nanoparticles, and natural compounds, which exert their effects by generating ROS.
Clinically approved antibiotics such as erythromycin, by protein synthesis inhibition, and rifampicin, by inhibiting RNA synthesis, are effective on Rhodococcus equi, while vancomycin is antibacterial against R. equi, Mycobacterium tuberculosis and S. aureus, by inhibiting cell wall synthesis inhibition. Moreover norfloxacin, by inhibiting DNA gyrase, is effective against R. equi, S. aureus and E. coli. Clofazimine, by DNA replication inhibition, ethambutol and isoniazid, by cell wall synthesis inhibition, are antibacterial against M. tuberculosis. Finally, quinones, by different cellular targets, are active on Enterococcus sp., Streptococcus sp., Staphylococcus sp. and Moraxela catarrhalis [64]. Anyway, studies observed that antibiotics functioning with a primary mode of action not correlated with OS, interfering with some bacterial cell targets, as above reported, were found to cause bacterial damage also generating ROS [71,72]. As shown in Figure 2, the interaction of antibiotics with bacteria cell targets can cause both ROS hyperproduction and cell damage which in turn cause additional ROS hyperproduction, while the improved concentration of ROS cause OS in bacteria lethally amplifying cellular damage.
Particularly, some antibiotics generate ROS through overstimulation of electrons via the tricarboxylic acid cycle and the release of iron from the iron–sulfur clusters, thus activating the Fenton chemistry. Nitrofurantoin and Polymyxin B are two commonly used ROS-mediated antibiotics [22]. Nitrofurantoin, used to treat urinary tract infections by E. coli [18], acts by a NADH-dependent reduction, producing nitroaromatic anion radicals. The autooxidation of these anion radicals in presence of O2, produces O2−, by which ROS generate, thus causing OS and toxicity on bacteria [18]. Polymyxin B (PMB) makes part of the family of antimicrobial peptides and is active mainly on Gram-negative bacteria such as A. baumannii, P. aeruginosa, and carbapenemase-producing Enterobacteriaceae [18,73]. Due to its neurotoxic and nephrotoxic properties, PMB is advised to be used only as a last resort antibiotic [18]. Sampson et al. demonstrated that PMB, in addition to be a membrane disruptor [74], induced cell death in Gram-negative bacteria by the accumulation of OH•[58]. Anyway, Arriaga-Alba and co-workers were the first authors to report oxidative stress induction as a part of the mechanism of action of nalidixic acid and norfloxacin in Salmonella typhimurium. [59]. Antibiotics were shown to upregulate many oxidative stress genes in P. aeruginosa[22]. Wang and Zhao found out that norfloxacin was more lethal in E. coli deficient in catalase gene katG, than in its isogenic mutants, thus confirming a potential pathway linking hydroxyl radicals to the antibiotic lethality [60]. Also, ampicillin and kanamycin showed increased lethality to an alkyl hydroperoxide reductase ahpC E. coli mutant, lacking the defense system to contrast hydroxyl radicals. These studies evidenced increased superoxide levels in the bacterium, which were the source of H2O2 which in turn generated the highly toxic hydroxyl radical, responsible for the improved lethality of antibiotics[60]. Hong et al demonstrated that E. coli exposed to lethal oxidative stressors caused by antibiotics including nalidixic acid, trimethoprim, ampicillin, and aminoglycosides, did not die only during the actual treatment but also post-treatment, after the removal of the initial stressor, due to post-stress ROS-mediated toxicity[61,62]. Anyway, the connection between antibiotics action and ROS are not still clearly demonstrated. Since it was demonstrated that antibiotics work also under anoxic conditions, some reports oppose that the generation of ROS contributes to their lethality, which is instead influenced by the bacterial metabolism, iron homeostasis and iron-sulfur proteins [75,76,77]. Although contradictory studies exist concerning the possibly ROS influence on antibiotic lethality, it was established that numerous alternative antimicrobials work by inducing ROS-mediated OS in bacteria. Unfortunately, some ROS-producing antibiotics, such as aminoglycosides, fluoroquinolones, and β-lactam antibiotics may induce host cellular damage in specific tissues such as the renal cortex or tendons by generating OS, anyway manageable by specific antioxidant molecules [64].
3.1.2. ROS Formation Induced by Alternative Antimicrobials
Several novel alternative antimicrobials are under development whose primary mode of action seems to be through the generation of ROS causing OS in bacteria. Generally, these antimicrobials often target the redox defenses, such as the thiol-dependent enzyme thioredoxin reductase (TrxR) in bacteria[63]. Examples of ROS-mediated antimicrobials include Ebselen, nanoparticles, nanozymes, and AGXX®.
Ebselen
Ebselen is an organo-selenium-based antioxidant drug endowed with anti-inflammatory, antioxidant, and cryoprotective effects. It acts by inhibiting TrxR in bacteria lacking glutathione, thus triggering OS, thus being lethal to these pathogens. Ebselen was recently shown to efficiently inhibit in vitro the growth of MDR S. aureus, to improve wound healing in rats, and to reduce the bacterial load in S. aureus skin lesions in rats [63]. Ebselen has been reported to inhibit M. tuberculosis. Important, Ebselen could also be combined with other ROS-stimulating compounds that block the antioxidant defenses of bacteria such as silver nanoparticles (NPs) [64].
Nanoparticles (NPs)
Mainly due to their small size (<100 nm) nanoparticles (NPs) can cause hyperproduction of ROS which can cause carbonylation of proteins, peroxidation of lipids, DNA/RNA breakage, and membrane structure destruction, thus damaging cells [78]. Among NPs, those made of silver, such as silver oxide NPs (AgNPs), titanium dioxide, silicon, copper oxide, zinc oxide, gold, calcium oxide, and magnesium oxide NPs have been ported to have antibacterial effects against both Gram-positive and Gram-negative pathogens [57]. Mesoporous silica NPs (MSNPs) containing a maleamato ligand (MSNPs-maleamic) and others containing also copper (III) coordinator ions (MSNPs-maleamic-Cu) synthesized by Diaz-Garcia et al, demonstrated antibacterial activity towards E. coli and S. aureus by OS induction [65]. The minimum inhibitory concentration values (MICs) of MSNPs-maleamic and MSNPs-maleamic-Cu established that both preparations performed better against E. coli than on S. aureus and that MSNPs-maleamic (MIC = 62.5 µg/mL) was more effective than MSNPs-maleamic-Cu (MIC = 125 µg/mL) against E. coli [65]. Both preparations caused a significant increase of ROS in both species (30-50% more than in control) and NPs that caused the major increase displayed lower MICs (MSNPs-maleamic, 50% in S. aureus, and 40% in E. coli), thus confirming that ROS and OS generation contribute to the antibacterial mechanism of action of MSN-maleamic and MSN-maleamic-Cu [65].
Nanozymes (NZs)
The major drawback of ROS-based antibacterial therapy consists in the low selectivity of ROS, that are detrimental also for human cells. Recently, nanomaterials possessing enzyme-like characteristics and referred to as nanozymes (NZs), have been reported to produce surface-bound ROS which were selective in killing bacterial cells over mammalian ones [66]. Particularly, ROS bound on silver and palladium bimetallic alloy (AgPd0.38) efficiently killed antibiotic resistant bacteria including S. aureus, Bacillus subtilis, E. coli and P. aeruginosa (MBC = 4-16 µg/mL) without developing drug resistance and inhibited biofilm formation [66].
AGXX®
AGXX® is a silver and ruthenium-based antimicrobial surface coating, that can be coated or deposited on various carriers such as cellulose, plastics, ceramics, or metals. AGXX® demonstrated low levels of toxicity to mammalian cells [79], and no tendency to develop resistance, due to its multiple mode of action [67]. ROS are produced catalytically by AGXX® which demonstrated to inhibit the growth of Enterococcus faecalis, and MRSA, as well as to prevent biofilm formation in MRSA [54,68]. In both species, AGXX® affected oxidative stress defenses such as superoxide dismutase (SodA), catalase (KatA), alkyl hydroperoxide reductase (AhpCF), thioredoxin/thioredoxin reductase (Trx/TrxR), disulfide reductase (MerA), and oxidized bacillithiol (BSH) [69]. AGXX® induced OS, general stress, heat shock (expression of Clp proteases), and copper stress [54,68]. In MRSA, it affected the iron homeostasis and upregulated several siderophore biosynthesis (sbn) genes [54,69], while in S. aureus, increased protein-thiol-oxidations, protein aggregations, and a BSH redox state were observed[67,69]. The mechanism of action of AGXX® was assessed in S. aureus observing two inter-connected redox cycles by which AGXX® simultaneously exerts ROS-mediated (superoxide anion, hydrogen peroxide, and highly toxic HO•) antimicrobial effect and achieves self-renewal.
3.1.3. ROS Formation Induced by Natural Compounds
In addition to synthetic compounds, also natural compounds can exert ROS-mediated antimicrobial effects. It is the case of allicin and honey, being the latter the main ingredient of a clinically approved gel formulation for topical administration. Honey is used in the honey antimicrobial therapy, that has been here discussed in a dedicated section. There are many other secondary metabolites produced by plants that may elicit oxidative stress in bacteria, such as catechins, ferulic acid, and their derivatives [64]. The combination of other ROS- and RNS-generating antimicrobials with these compounds may lead to the development of promising therapeutic strategies against different intracellular bacterial pathogens [64]. Particularly, allicin is a constituent of garlic, which works as a thiol-reactive compound; decreasing the levels of low molecular weight thiols, which should function as a defense against ROS. Their increasing causes OS to bacterial cells, thus inhibiting their growth [70].
3.2. ROS Formation Induced by Antibacterial Photodynamic Therapy
By a study on the toxicity of small concentrations of acridine red on Paramecium spp. it was recognized that the observed toxicity was dependent on the time of the day and the amount of daylight [80]. Later, von Tappeiner with Albert Jesionek applied clinically this approach, to treat skin carcinomas and coined the term “photodynamic phenomenon” [81,82,83] .Thus, anticancer photodynamic therapy was born. In those years, the successful photodynamic inactivation of bacteria was also described [80]. Anyway, while anti-cancer PDT is clinically applied from 25 years, at least in the treatment of actinic keratosis or basal cell carcinoma, its application as antimicrobial option has only more recently been rediscovered, to manage the emergence of the first drug-resistant infections in the healthcare sector during the early 1990s [84,85].
3.2.1. Basic Principles of Photodynamic Therapy
PDT is based on the combination of three elements including a not toxic compound referred to as photosensitizer (PS), light in a spectral range appropriate for exciting the PS (typically from the visible to near infrared (NIR) spectrum) and molecular oxygen [85]. The mechanism of PDT is described by the Jablonski diagram reproduced in Figure 3.
Briefly, upon absorption of a photon (A), the PS moves from its ground singlet state (S0) to an excited singlet state (Sn). In this status, PS can lose energy, thus returning to S0 by emitting fluorescence (F) or heat (H) via internal conversion. On the contrary, it can pass to a longer-living excited triplet state PS (T1), through an inter-system crossing (ISC) process. Following, it can either return to S0 state by phosphorescence emission (P) or generating reactive oxygen species (ROS), by two mechanisms [86]. When a type I mechanism is followed, electrons are transferred to surrounding substrates thus forming of superoxide radical anions (O2−•), that undergo dismutation into hydrogen peroxide (H2O2) from which the highly reactive hydroxyl radical (HO•) derives via Fenton-like reactions. Differently, in type II mechanism, energy and not charge is transferred directly to the ground state molecular oxygen (3O2), leading to emergence of singlet oxygen (1O2), which is nothing else than energized molecular oxygen [87]. At this point, PS is returned in its S0 status, ready to begin another cycle with production of additional ROS. Interesting, one PS molecule can generate thousands of molecules of 1O2 before being destroyed. The singlet oxygen quantum yield describes the amount of type II mechanism [87]. During antibacterial PDT, type I and type II reactions can occur simultaneously, and the ratio between these processes depends on the type of PS utilized, its chemical structure and the specific microenvironment in which the PDT is implemented. The interplay between type I and type II mechanisms is a critical factor to consider for an optimized treatment and understanding its underlying photochemical processes [88,89,90]. As already reported, ROS are detrimental for bacteria by targeting several vital microbial molecules, such as proteins, lipids, and nucleic acids, thus determining bacterial death. An additional type III reaction has been recently incorporated into the familiar categories of type I and II mechanisms. In this novel process, free radicals of inorganic compounds are generated, regardless of the presence of oxygen, which would participate in the photoinactivation of microorganisms [91].
3.2.2. Photosensitizers Used in Clinical Trials
The most common PSs clinically used in anticancer PDT have been extensively studied also for their antibacterial properties and effectiveness in the treatment of various bacterial infections. Many published studies have determined that phenothiazinium PSs such as MB and TB are effective on planktonic bacteria. Furthermore, some studies also tested the efficacy of phenothiazinium against biofilm structures [92]. Recently, new derivatives such as dimethyl methylene blue (derived from MB) and EtNBS (N-ethylpropylsulfonamido) have been studied. These dyes possess a high cationic charge that makes them more effective against bacterial cells [92]. Rose Bengal is an anionic synthetic xanthene dye that has shown promise in several clinical trials for PDT, particularly in the treatment of localized bacterial infections. Other synthetic anionic xanthene dyes derived from Fluorescein are Eosin Y and Erythrosine (ERY). All these dyes have an absorption peak in the green wavelength range (480–550 nm). The attachment and uptake of anionic PSs by the bacterial cells are lower than cationic ones [92]. Amino-levulinic acid (ALA) is a prodrug that can be converted to protoporphyrin IX (PpIX), clinically used (in the form of the hydrochloride salt) in combination with blue light illumination for the treatment of minimally to moderately thick actinic keratosis of the face or scalp. It has demonstrated to be effective also in the treatment of bacterial infections, including periodontal infections. Additionally, a variety of evidence proved that 5-aminolevulinic acid-based photodynamic therapy (ALA-PDT) is clinically effective in the management of Acne vulgaris and is recommended as an alternative treatment modality for severe acne [119]. Concerning chlorins, mainly cationic derivatives of chlorin-e6 are used for PDT. Photodithazine® is a commercially available chlorin-e6 derivative with two positive charges. Chlorine e6 (Ce6) and various phthalocyanines have strong antibacterial properties and have been used in preclinical studies and in some early clinical trials for antibacterial PDT. Curcumin is a natural compound found in Curcuma longa, and its cationic derivatives have also been investigated as possible PSs. Collectively, the use of specific PSs in clinical trials strongly vary depending on the target bacterial infection and the research objectives of each study [92,120,121].
3.2.3. Antibacterial PDT vs. Antibiotics
Unfortunately, as reported in the following Table 7, even if significant are the advantages of using antibacterial PDT (APDT) compared to antibiotics, APDT has certain limitations that need careful consideration.

Table 8.
Advantages and limitations of APDT compared to conventional antibiotics.
| Advantages | Limitations | Discussion on limitations |
|---|---|---|
| ↑ BS action than antibiotics including bacteria, protozoa, fungi |
Light limited penetration capabilities |
Possible problems in reaching deep seated infections by scattering phenomena [122] Bacterial colonies located beneath the skin’s surface or within organs could be difficult to reach * [123] |
| ↓ Adverse effects and damage to the host tissue | Potential lack of target specificity |
↑ Selectivity and efficiency of PSs toward bacteria and ↓ toxicity on mammalian cells can be achieved using proper vectors or by co-administration, conjugation or incorporation with polycationic materials, bacterial-targeting peptides, polymers, antibiotics, or antibodies [124,125,126,127] |
| Bactericidal effects independent of antibiotic resistance pattern | Risk of antibiotic inactivation |
When in combination with certain antibiotics APDT can inactivate the antibiotics [123] |
| No resistance following multiple sessions of therapy | Potential side effects | Emergence of skin sensitivity, redness, and pain at the treatment site [128] |
* In this case the correct choice of the wavelength and PS is pivotal; PS = Photosensitizer; BS = broad spectrum; ↑ means increase, more; ↓ means decrease/reduction, less, minor.
Despite promising results have been observed in some diseases, the clinical translation of PDT for bacterial infections has progressed slower than for cancer treatment and leisurelier than anticipated. Therefore, its widespread adoption in medical practice remains limited. Among limitations, the modest capability of light to penetrate skin, tissues and organs hampers the application of APDT for systemic infection and reduce its effectiveness in the topical treatments. The penetration of light depends on optical properties of the tissue and the wavelength of used light. There exists a heterogeneity between tissues and even within a tissue. These inhomogeneity sites (e.g. nuclei, membranes, etc.) cause light scattering, reflecting, transmitting or absorption [122]. Light within the 620–850 nm spectrum range achieves the optimum tissue penetration and PDT applications [122]. Mainly concerning potential side effects, understanding, and effectively managing them is critical for optimizing patient outcomes. To fully harness the potential of antibacterial PDT as a valuable therapeutic strategy in combating bacterial infections, it is imperative to address these drawbacks through ongoing research efforts and comprehensive approaches [120,129]. Successful outcome of antibacterial PDT depends on the selected PS. To enhance PS selectivity over host tissue, while maintaining nonselective activity against microbial species represents a pivotal challenge. The optimal PS should target both Gram-positive and Gram-negative species, accounting for their differential treatment responses. Several very recent studies have described the latest advancements in the field over the past years, emphasizing new PS systems relevant for antibacterial PDT’s methodologies [80,130,131,132], which have recently reviewed by Sébastien Clément & Jean-Yves Winum [123].
3.2.4. Light Sources
The main light sources employed to activate PSs have been included in Table 7.
Table 7.
Main light sources employed to activate PSs.
| Light source | LASER | Argon |
Monochromatic, coherent, and collimated light High irradiance Couplable into optical fiber bundles Expansive, cumbersome |
Refs. | |
| Diode | [133] | ||||
| Neodymium doped | Yttrium Aluminum Garnet lasers |
||||
| Light-emitting diodes (LEDs) | Deliver a slightly wider emission spectrum than LASER Low costs No monochromatic, no coherent |
[133] | |||
| Gas-discharge lamps | Quartz-tungsten-halogen lamps Xenon-discharge lamps Sodium lamps |
They can be spectrally filtered to match any PS No efficiently couplable into optical fiber bundles Cause more heating as compared to LASERs and LEDs |
[134] | ||
| Daylight | Broad-spectral range from UV to IR region Free of cost Can illuminate a very large area with high uniformity Variable in irradiance, radiant exposure is poorly controlled |
[133,135] | |||
LASER = Light Amplification by Stimulated Emission of Radiation.
In addition to the light sources reported in Table 7, “non-coherent” or “non-thermal” light sources without giving details of the actual light source used have been reported. Additionally, there are also occasional reports of other light sources such as endoscopy systems, photopolymerisers and supra-luminous diodes (SLD) [133]. All in all, for the irradiation of a given PS, parameters such as radiant exposure, light irradiance, power output, spectral emission, intensity of the respective light source, as well as the mode of light-delivery (via optical fiber or directly) are more important than the type of the light source itself.
3.2.5. Clinical Trials
Due to the multi-faceted nature of the antimicrobial photodynamic process and its diverse targets, the emergence of resistance in microbes becomes highly improbable. Consequently, the anti-bacterial PDT should be considered promising as a potent and innovative approach in combating bacterial infections [136]. In the last years, the volume of research focused on PDT as therapeutic device to inhibit a wide range of microorganisms, including bacteria is remarkably increased. Anyway, the antibacterial PDT has yet to attain the capability to tackle systemic infections. On the contrary, it has demonstrated high potential for addressing localized infections sustained by MDR bacteria including hospital-acquired pathogens like E. faecium, S. aureus, K. pneumoniae, A. baumannii, P. aeruginosa, and Enterobacter spp., which together constitute the group ESKAPE [137]. Additionally, in vitro and in vivo studies have demonstrated the capability of PDT of eradicating or significantly reducing biofilms, thus finding applications in dental diseases, skin infections and orthopedic implants treatment [138,139]. The ongoing advancement of antibacterial PDT systems is also marked by the continuous refinement of the strategies, prominently through synergistic combinations of diverse chemicals. Antibacterial PDT has been applied in about 40 clinical trials for treatment of dermatological disorders and oral infections. Clinical trials on antibacterial PDT have focused on evaluating its safety and efficacy in treating specific bacterial infections, especially those that are challenging to manage with traditional antibiotics [85]. Some of the key findings from these trials include those obtained in dermatological disorders such as acne vulgaris, and bacterial skin conditions where antibacterial PDT studies have shown positive outcomes in reducing the severity of acne lesions [140,141,142,143]. Some studies have also explored applications in treating infected wounds, especially those associated with MDR bacteria. The results suggest that antibacterial PDT can be beneficial in promoting wound healing and controlling bacterial growth [144,145]. In the field of periodontal diseases, clinical trials have shown promising results for treating chronic periodontitis, a common bacterial infection that affects the gums and supporting structures of the teeth [146,147,148,149]. Other clinical trials studies have also examined the use of antibacterial PDT in managing bacterial infections related to medical implants, such as catheters and prosthetic devices [150,151].
3.3. ROS Formation Induced by Antibacterial Honey Therapy
Honey was used in traditional medicine mainly to treat wounds due to its antimicrobial effects and healing properties. The bactericidal efficacy of honey was reported more than a century ago by Van Ketel [152], whose findings prompted extensive research on honey over the next decades. With the advent of modern medicine, the interest in honey and its medical use decreased [152]. Nowadays, honey is undergoing a revival in its consideration for antimicrobial and wound healing applications, due to the rising global antibiotic resistance, which makes the development of novel alternative therapies to combat infections necessary. Several factors can contribute to its effective antimicrobial activity, which can strongly vary depending on different microbial strains, including the geographical and botanical source, its harvesting, processing, and storage conditions. It has been demonstrated that a range of both Gram-positive and Gram-negative bacteria, including MDR strains and biofilms, fungi and viruses can be inhibited by honey. Furthermore, susceptibility to antibiotics can be restored when used synergistically with honey. Table 8 reports the antibacterial effects of honeys from different geographical sources and the related target bacteria.
Table 8.
Antimicrobial effect of honey from different geographical locations and target pathogens.
| Country of Origin | Honey Sample | Organisms | Ref. |
|---|---|---|---|
| Australia | |||
| New Zealand | Manuka | S. aureus, P. aeruginosa | [153] |
| New Zealand | Manuka |
S. aureus, MRSA, MSSA Coagulase-negative S. epidermidis K. pneumonia, ESBL E. coli |
[154] |
| Australia | Leptospermum based honey | S. aureus | [155] |
| North America | |||
| Canada | Canadian honey | E. coli, Bacillus subtilis | [156] |
| Cuba | Christmas vine, Morning glory Black mangrove Linen vine, Singing bean |
S. aureus, P. aeruginosa E. coli, B. subtilis |
[157] |
| South America | |||
| Chile | Ulmo honey | MRSA, E. coli, P. aeruginosa | [158] |
| Argentina | Algarrobo, citrus and multifloral honey |
S. aureus, E. faecalis, E. coli Morganella morganii P. aeruginosa |
[159] |
| Europe | |||
| Scotland | Blossom, heather, Highland, Portobello Orchard |
Acinetobactor calcoaceticus S. aureus, P. aeruginosa, E. coli |
[160] |
| Northwest Spain | Rubus honey |
S. aureus, S. epidermidis Micrococcus luteus, E. faecalis B. cereus, Proteus mirabilis, E. coli P. aeruginosa Salmonella typhimurium |
[161] |
| Denmark | Heather, raspberry, rapeseed, hawthorn White clover | S. aureus, P. aeruginosa, E. coli | [162] |
| Slovakia | Honeydew honey | P. aeruginosa, S. aureus | [163] |
| Asia | |||
| China | Buckwheat honey | S. aureus, P. aeruginosa | [153] |
| Saudi Arabia | Sider honey |
S. aureus, Streptococcus pyogenes Corynebacteria pseudotuberculosis K. pneumonia, P. aeruginosa E. coli |
[164] |
| Africa | |||
| Algeria | Astragalus, wall-rocket, eucalyptus Legume, peach, juniper, buckthorn multifloral |
Clostridium perfringens, S. aureus, E. coli, B. subtili. | [165] |
| Nigeria | Wildflower and bitter leaf honey |
S. typhimurium Shigella dysenteries, E. coli B. cereus, S. aureus |
[166] |
| Egypt | Cotton, blackseed, orange, eucalyptus Sider, clover honey |
E. coli, S. aureus Streptococcus mutans, P. mirabilis P. aeruginosa K. pneumoniae |
[167] |
| Egypt | Acacia, citrus, clover, coriander, cotto palm honey |
S. aureus, S. pyogenes C. pseudotuberculosis K. pneumonia, P. aeruginosa, E. coli |
[164] |
ESBL =extended spectrum β-lactamase.
3.3.1. Medical Application of Honey
Although the knowledge of the antibacterial compounds involved in the antibacterial effects of honey remains still incomplete, the information on honey has remarkably expanded in the recent years. Otherwise, despite the variability of the antibacterial activity of honey limits its extensive applicability in medicine, several honeys have been approved for clinical application [168]. Currently, honey is mainly used as a topical antibacterial agent in applications on wounds. Its high viscosity grants an effective hydrated and protective barrier between the wound site and external environment. Honey has been used to treat wounds such as burns, trauma, and chronic wounds, where the complex wound healing process could be interrupted by infection or specific disease states (e.g., diabetes), thus limiting the development of irreversible chronic wounds, recurrent infections, amputation/limb salvage, and life-threatening conditions [152]. For mild to moderate superficial and partial thickness burns, honey was more effective than conventional treatment for reduction of microbial colonization and improved wound healing [169,170]. In a study, the application of honey on tunneled cuffed hemodialysis catheters, resulted in a comparable bacteremia-free period compared with that obtained with mupirocin treatment [171]. Honey has also been widely explored as a tissue-regenerative agent. In this case, it has been applied directly on wound or in combination with traditional wound dressings, which allow honey remaining in direct contact with the wound bed, thus providing a persistent and long-term release of antimicrobial agents to contribute to all stages of wound healing. Furthermore, the presence of reactive oxygen species (ROS) such as H2O2 has been shown to promote wound healing by promoting cellular repair processes and tissue regeneration [10,172]. Anyway, some limitations exist, such as being absorbed by the dressing, poor penetration into the wound site, and short-term antimicrobial action. To address these issues, tissue engineering approaches have been developed, such as its formulation in electrospun fibres and hydrogels [173,174,175,176,177]. Collectively, the use of honey, honey-derived, and honey-inspired products in tissue engineering applications combined with other biomaterials, may enable its use in a variety of other clinical situations outside wound care, where the combination of antimicrobial properties and tissue regeneration is desirable.
The Case of Surgihoney Reactive Oxygen (SHRO)
The first therapeutic agent based on the oxidative activity of ROS was a pharmaceutical honey gel for treating wounds, referred to as Surgihoney Reactive Oxygen (SHRO). Particularly, SHRO is an engineered sterilized honey created to act as a preventive antimicrobial agent for soft tissue infections. SHRO deriving from natural organic honey from different origins, is capable to provide a constant level of ROS over a prolonged period of time, when topically applied to a wound. Subsequently, ROS induce OS due to •OH production and inhibit the essential metabolic procedure for bacterial growth [178]. As more in detail discussed in the subsequent sections the antimicrobial activities of SHRO are mainly due to the generation of H2O2[179]. Other formulated honey prototypes (PT1 and PT2) were designed to furthers increase the generation of H2O2. Subsequently, honey ROS-based antibiotic agents differently formulated, such as sprays, nebulizers and infusions, that employ this mechanism are being developed and may be particularly useful for delivery ROS to other clinical sites. Nowadays, there are many types of therapeutic honey in the market (e.g., Paterson’s curse, Rosemary, Manuka, Thyme, Revamil, Rewa Rewa, heather honey, Khadi, Kraft honey, Multifloral, and Medihoney) [180,181]. Table 9 summarizes a clinical register using SHRO in various complex and severe infections [4].
With the rising age of the population in many countries, and the global epidemic of obesity and type 2 diabetes [4], the disease of chronic soft tissue lesions is becoming enormous. Most chronic breaks in the skin often become colonized with bacteria [182], that regardless their pathogenicity play an essential role in slowing tissue healing, establishing biofilm and resulting in wound slough and an unpleasant odor [182]. As reported in Table 9, in these cases where often available antibiotics are no longer functioning, the effects of SHRO on bioburden and biofilm [178] can be of great help. In fact, the early use of ROS in such lesions can control bioburden and biofilm, thus sparing conventional antibiotic use, and supporting infection control [178,183,184]. In clinical studies, ROS therapy via SHRO has demonstrated satisfactory safety and tolerability, and clinical- and cost effective in practice [183,185]. Most outstandingly, SHRO has demonstrated impressive capacity to clean up bacterial bioburden and biofilm in chronic wounds, being also active on MDR bacteria such as P. aeruginosa, MRSA and vancomycin-resistant enterococci (VRE) present in an ischemic ulcer [4].
Medical Grade Honey for Clinical Applications
Medical-grade honey (MGH) intended for clinical application must be sterilized by gamma irradiation to destroy potentially present bacterial spores, including those of Bacillus spp. and Clostridium botulinum, which could cause wound botulism or gangrene [186]. Several types of honey, including MGH, have recently been re-introduced into modern medicine. There is no clear definition of MGH, but according to a study by Hermann et al., MGH should satisfy the criteria reported in Figure 4 [187].
Manuka and Revamil® are the major medical-grade honeys currently approved for clinical application. Manuka honey is produced from the manuka bush (Leptospermum scoparium) native of New Zealand and Australia. The raw honey used as a source for Revamil®, is instead produced by a standardized process in greenhouses. Since Revamil® honey is registered as a medical device for applications in wound healing and not as an antimicrobial agent, the antimicrobial activity is not specified for individual batches of this honey. However, in a quantitative liquid bactericidal assay, both Revamil® and manuka honeys demonstrated potent bactericidal activity [188,189], being manuka honey the most performant against S. aureus, B. subtilis, and P. aeruginosa. On the contrary, these honeys had identical bactericidal activity against E. coli. The following Table 10 reports the approved and already commercially available wound healing products based on honey.
The antimicrobial activity of the majority of honeys is mainly due to its capability to generate high levels of hydrogen peroxide (H2O2) [10,21,179,212,213,214,215] by the oxidative action the enzyme glucose oxidase (GOx) on glucose. Secreted into the nectar by bees during the preparation of honey, GOx oxidizes glucose to gluconic acid thus producing H2O2 [10,215,216,217,218,219]. The enzyme presents no activity in raw honey, due to a lack of free water. Therefore, to initiate the peroxide-dependent antimicrobial mechanism, the honey needs to be diluted. Other important antimicrobial features responsible for the non-peroxide activity of honey include low water content (osmotic effect), low pH (acidic environment), phenolic compounds, bee defensin-1 (Def-1), and methylglyoxal (MGO) (in Leptospermum-derived manuka honey). Table 11 summarizes the factors that confer honey its antibacterial effects.
Briefly, sucrose captured by bee from flowers is broken down via diastase and invertase enzymes into glucose and fructose. Glucose oxidase (Gluox) secreted by the bee’s hypopharyngeal glands, in presence of O2 and sufficient H2O oxidizes glucose forming gluconolactone/gluconic acid, that make honey acidic and H2O2 [223]. H2O2 is the most responsible of honey’s antimicrobial activity, by killing pathogens through DNA damage, and being destructive towards several cellular targets [223]. Interesting, the antimicrobial effect of hydrogen peroxide in honey increases upon dilution, enabling the glucose oxidase enzyme to bind to glucose more readily, thus resulting in a continuous production of hydrogen peroxide [215]. Moreover, honey, predominantly due to gluconolactone/gluconic acid formation, is acidic with an average pH of 3.91 (ranges between 3.4 to 6.1), which makes it powerful against microbial strains, preventing their growth. Bee Def-1 is an antibacterial peptide originating in the bee’s hypopharyngeal gland and identified in bee hemolymph (the bee blood system) [224]. Within the bee it acts as an innate immune response, exhibiting activity against fungi, yeast, protozoa and both Gram-positive and Gram-negative bacteria [189]. Importantly, bee Def-1 is mainly effective against Gram-positive bacteria, most notably B. subtilis, S. aureus and Paenibacillus larvae, while it has limited effectiveness against MDR organisms [168]. Although the full mechanism of action for bee Def-1 has not been elucidated, defensin proteins from other species have been shown to create pores within the bacterial cell membrane, resulting in cell death [225]. Bee Def-1 demonstrated to play an important role in wound healing, through stimulation of MMP-9 secretions from keratinocytes [226]. It interferes with bacterial adhesion to surfaces, or in the early biofilm stage by inhibiting the growth of attached cells, and by altering the production of extracellular polymeric substances (EPS). MGO is generated in honey during storage by the non-enzymatic conversion of dihydroxyacetone, a saccharide found in high concentrations in the nectar of Leptospermum flowers [220]. The antimicrobial activity of MGO is attributed to alterations in bacterial fimbriae and flagella, which prohibit the bacterium’s adherence and motility. Honey is a super-saturated solution of sugars. The strong interaction between these sugars with water molecules prevents the abundance of free water molecules (low water activity) available for microbes to grow [221]. Finally, the combination of different phenols acts as an enhancer of honey’s antimicrobial efficacy. Produced as plant secondary metabolites, these bioactive compounds are transferred from the plant to the honey by bee and have been identified as the major responsible for the health-promoting properties of honey [222]. In alkaline conditions (pH 7.0–8.0), polyphenols can display pro-oxidative properties, inhibiting microbial growth by accelerating hydroxyl radical formation and oxidative strand breakage in DNA. They could also support the production of considerable amounts of H2O2 via a non-enzymatic pathway.
3.4. ROS Formation Induced by Antibacterial Hyperbaric Oxygen Therapy
Differently from antibacterial honey therapy, which is a topical treatment, antibacterial hyperbaric oxygen therapy (HBOT), making part of hyperbaric medicine, is a systemic method to treat soft tissue infections [57]. Particularly, in a typical HBOT treatment, the patient (mono-place) or more than one patient (multi-place) inhale 100% O2 for a specified time in a pressurized chamber [227]. From the lung, the inhaled oxygen is delivered to the whole body, where induces the formation of ROS, which overwhelm the antioxidant defenses of the facultative anaerobic bacteria and aerobic bacteria causing lipids peroxidation and membranes disruption, DNA injury, protein disfunction and death. A pressure greater than 1.4 atmosphere absolute (ATA) is necessary to have an effective antibacterial effect against some facultative anaerobic bacteria and aerobic bacteria [227]. In addition to be directly bactericidal via ROS formation, it has been reported that HBOT enhances the antimicrobial effects of the immune system, such as those of leukocytes in hypoxic wounds (Figure 5) [228].
Moreover, HBOT potentiates the antibacterial effects of some antibiotics such as imipenem and tobramycin (Figure 6).
Hyperoxia (98% O2 at 2.8 absolute ATA—approximately 284.6 kPa) has been shown to enhance the effect of nitrofurantoin, sulfamethoxazole, trimethoprim, gentamicin, and tobramycin in E. coli strains (serotype 018 and ATCC 25922) [230]. Two works by Kolpen et al report that the combination therapy of ciprofloxacin and HBOT may be potentially beneficial for the eradication of infections caused by the biofilm forming P. aeruginosa [231,232]. Anyway, HBOT did not show any additive or synergistic effect with other antimicrobial agents, such as distamycin and rifampicin [57]. Very recently, it has been reported that wild strains of Pseudomonads, Burkholderias, and Stenotrophomonads stopped growing under hyperbaric conditions at a pressure of 2.8 ATA of 100% oxygen [230]. HBOT is used as a primary or alternative method for the treatment of infections such as diabetic foot infection, surgical site infections, gas gangrenes, osteomyelitis, and necrotizing compartments [229]. In addition to the bactericidal effects, HBOT suppresses the production of clostridia alpha toxin in gas gangrene diseases. HBOT has demonstrated also anti-inflammatory effects, that may play a significant role in decreasing tissue damage and infection expansion. While generally safe, HBOT may have side effects such as ear discomfort, sinus pain, and temporary nearsightedness. Other contraindications include untreated pneumothorax (collapsed lung), certain medications, and claustrophobia. Although patients treated by HBOT need carful pre-examination and monitoring, when safety standards are strictly tracked, HBOT can be considered a suitable procedure to treat severe infections sustained by MDR bacteria, with an acceptable rate of complication.
3.4.1. Clinical Application of HBOT in Infections
Several studies have evidenced that HBOT, either alone or as an accessory treatment, can be a valuable therapeutic option to cure patients with several diseases, difficult to treat infections included (Table 12).
Basically, HBOT strongly improves the levels of O2 concentration in blood and the oxygen pressure both in blood and tissues (2000 mmHg and 500 mmHg, respectively), determining hyperoxia conditions, which provide beneficial effects in patients suffering from several diseases as reported in Table 12. HBOT was used to adjust immunology, to maintain the durability of an allograft [253]. HBOT has demonstrated beneficial effects on vascular endothelium, thus promoting angiogenesis, and to induce partial high tensions of O2 in circulating plasma, thus stimulating O2 dependent collagen matrix formation, which is an essential phase in wound healing [254]. On the other hand, HBOT effects on sepsis, urinary tract infections and meningitis are not well known, so far. Unequivocally. the most frequent clinical application of HBOT remains for several skin soft tissue infection and osteomyelitis infections, which are associated with hypoxia, caused by anaerobic infections due antibiotic resistant bacteria [255,256]. Currently, HBOT is considered both alone and in combination with different antibacterial treatment a relevant option for solving several cute or chronic diseases [249,251,257,258]. Although animal studies have shown the inhibitory effect of HBOT on inflammation and apoptosis after cerebral ischemia [259,260], clinical trials on humans have not shown any significant benefit. Anyway, it has been indicated that HBOT can improve some neuropsychological and inflammatory outcomes, especially in stroke patients, within the first few hours [261,262]. Furthermore, studies on animals have shown that HBOT is associated with reduced blood–brain barrier breakdown, reduced cerebral edema, improved cerebral oxygenation, decreased intracranial pressure, reduced oxidative burden, reduced metabolic derangement, and increased neural regeneration [260,261,263]. The following Table 13 contains an overview of some clinical studies investigating the application of HBOT for different infections, while Table 14 collects the most relevant studies concerning specifically, the clinical application of HBOT in Surgical Site Infections (SSI) categorized based on the type of SSI or surgery. Sternal wound infections following cardiac surgery, SSIs following neuromodulation or neuro-muscular surgery, and SSIs following the male-to-female gender affirmation surgery (urogenital surgery) have been included.
4. Possible Novel Methods to Induce ROS Formation: Our Proposal
4.1. Environmental Persistent Free Radicals (EPFRs)
Environmentally persistent free radicals (EPFRs) are defined as long-living organic free radicals stabilized on or inside particles [294]. EPFRs are persistent because of the protection by the particles containing them and the presence of transition metals, thus having lifetimes exceptionally longer (from days to years) than other free radicals. These surface-bound radicals are found in contaminated soil, tar balls, and cigarette smoke, and primarily form during thermal processes such as pyrolysis and combustion of organic materials, waste incineration and photoactivation [295]. In fact, the byproducts from these processes, such as phenols and aromatic polycyclic hydrocarbons provide a breeding ground for EPFR generation. In fact, EPFRs are also found in the matrix of ultrafine and airborne fine particulate matter (PM), which is emitted into the environment by both natural and anthropogenic processes, including coal combustion emissions (16.8%), vehicular emission (32.1%), industrial processes (11.7%), dust storm (27.2%), and nitrates (3.4%). PM is a mixture of organic, inorganic species, solid and liquid components of metals which have the tendency to form radicals with or without the sunlight photoactivation [295,296]. The environmental factors affecting the EPFRs’ formation, lifetime and abundance such as precursors, temperature, light irradiation, presence of metals, temperature, pH, humidity, thermal processing time, and oxygen [297]. EPFRs are categorized as no decay, low decay, and fast decay radicals. The no decay EFFRs are unpaired electrons of delocalized over aromatic bonds entrapped inside PM. Phenoxyl radicals are fast decay EPFRs, while semiquinone radicals are subjected to slow decay [298]. EPFRs can produce reactive oxygen species, including hydroxyl radicals, which induce oxidative stress in living organisms, posing adverse environmental and human health effects. The atmospheric oxygen is the only sink for stable free radicals converting them into not particles and/or metal stabilized molecular species thus decaying [295]. The EPFRs decay in atmosphere depends on reaction of EPFRs with molecular oxygen. By reaction with atmospheric oxygen, they generate high levels of reactive oxygen species (ROS), via electron transfer, such as hydroxyl radicals and superoxide anion radicals, thereby inducing cellular oxidative stress [23,24]. For this reason and their possible redox recycling, EPFRs are emerging as environmental pollutants with a ROS-dependent significant toxicity to organisms, including humans, plants, animals, and microorganisms. Anyway, recent research has also explored their potential for degrading organic environmental contaminants. By activating hydrogen peroxide or persulfate EPFRs produce ROS species such as superoxide, singlet oxygen, or OH−, thus inducing the degradation of environmental organic pollutants [295]. Collectively, although EPFRs are long-lived environmental pollutants, harmful to environment and living beings, their capacity of activating ROS generation, if properly controlled, can make them a wide range tool to environment remediation.
4.2. Biochar-Derived Persistent Free Radicals (PFRs)
Biochar (BC) is a carbonaceous material obtained by the pyrolysis of different vegetable and animal biomass feedstocks at 200–1000 ◦C in limited presence or in absence of O2. BC has demonstrated a broad prospective use in the treatment of environmental pollutants, in soil amendment. It has been used in photocatalytic and photothermal systems, for photothermal conversion, to construct electrical and thermal device, as well as 3D solar vapor-generation devices for water desalination [24]. All these potentials are due to its high surface area and rich pore structure, which determines great physical absorptivity [23]. Additionally, they also depend on the chemical characteristics of BC, which in turn depend on the type of biomass used to produce BC, on the original biomass chemical composition, and pyrolysis conditions [299,300]. Whereas, starting from the year 2014 (Figure 7), the presence of persistent free radicals (PFRs) in BC deriving from lignocellulosic biomasses, like the radicals previously detected in combustion-generated particulate matter (PMs), sediments, and contaminated soils, known as environmental persistent free radicals (EPFRs), have been reported.
As EPFRs, such reactive species can remain stable for months or years and play a crucial role in the capacity of BC to degrade different types of xenobiotics and pollutant by oxidative reactions via ROS formation. Unlike other free radicals, including ROS, PFRs are resonance-stabilized since they are bound to the external or internal surface of solid particles of BC [24]. If BC is conserved under vacuum, lifetime of PFRs could be infinite (no decay radicals), while when air-exposed they react with molecular oxygen in the air, decay over time, thus producing ROS. Similarly, in aqueous systems, PFRs act as transition metals like Fe2+, forming ROS, as well [301,302,303,304]. PFRs are categorized into three classes, i.e., oxygen-centered PFRs (OCPFRs), carbon-centered PFRs (CCPFRs), and oxygenated carbon-centered radicals (CCPFRs-O). The possible presence of PFRs onto a BC, their type and concentrations are significantly affected by pyrolysis conditions, biomass types, the elemental composition of pristine biomass, and the presence of external transition metals as detailed in Table 15.
4.1.1. Proposed Mechanisms for PFRs Formation During Biomass Pyrolysis
The actual mechanism by which PFRs form during pyrolysis remains not fully clarified. However, transition metals capable of electron transferring and substituted aromatics molecules present in the lignin component of pristine biomass have been recognized to be essential for PFRs formation. Anyway, high concentration of PFRs have been detected also in products obtained by the pyrolysis of non-aromatic cellulose in absence of transition metals [308]. Collectively, PFRs can form by different pathways including or not the presence of transition metals, and once formed PFRs, could be either only surface-stabilized or surface-stabilized in metal-radical complexes [310]. Scheme 1 (concerning lignin) and Scheme 2 (concerning cellulose and emicellulose) report the possible chemical paths by which PFRs may form.
4.1.2. Possible Activities of PFRs and Our Proposal
PFRs formed in BC during combustion of lignocellulosic biomasses either in the presence or absence of external transition metals could promote several beneficial reactions, such as PFRs-mediated remediation and degradation of organic and inorganic pollutants by different actions and mechanisms, including oxidative and reductive processes. PFRs on BC can activate hydrogen peroxide (H2O2) or oxygen (O2), as well as persulfate (S2O82−) to produce different radical and not radical oxygenated species (ROS) capable of efficiently degrade organic contaminants by oxidative mechanism, as ROS generated by the previously reported antimicrobial therapies are bactericidal to pathogens inducing OS via ROS stimulation. Therefore, we thought that ROS induction using BC-derived PFRs, whose type, concentration and reactivity can be tunable varing pyrolysis conditions could be a novel method to form ROS for a possibly more selective BC-based antibacterial oxidative therapy. In this regard, in a recent review on BC-derived PFRs, a random selection of the main experimental works regarding the applications of PFRs found in BCs conveyed in the last five years (2019-2023) has been reported. Among the reported PFRs applications, three regarded their use as antibacterial agents (Table 16), thus supporting our idea.
5. Conclusions
Upon the colonization of the host cell during infection, the maintenance of redox homeostasis (RH) is a key process for bacterial survival and for escaping the oxidative stress physiologically generated by macrophages to opposing the development of the infection. Pathogens succeed in strictly controlling RH by a mechanism based on different redoxins and low molecular weight-thiol molecules. Therapeutic strategies based on the capacity of different compounds or methods to cause ROS and RNS hyper-generation during phagocytosis to unbalance bacterial redox defences and stop host cell colonization have a great potential to solve the increasing problem of antibiotic-resistant infections. ROS have demonstrated to be effective in inhibiting clinically important microbial pathogens, by lipid peroxidation thus damaging their membranes, by harming DNA and impairing protein functions. It has been observed that certain traditional antibiotics and alternative antimicrobials, including nanomaterials, as well as their combination, induce ROS as a secondary or main mechanism of antibacterial effect. Additionally, on the worrying scenario of an increasing global emergence of difficult-to-treat infections due to bacterial resistance to available antibiotics, old procedures that inhibit microbial growth by forming ROS, such as HBOT and medical honey, have been reinvigorated. Together with the more recent PDT, they are in fact currently successfully employed for the treatment or prevention of soft tissue infections and chronic ulcerations. The development of resistance to these methods has not been reported, but unfortunately, since ROS-mediated OS is destructive also on eukaryotes, their clinical application to treat systemic infections is at present impracticable. Further studies, aimed at identifying novel delivery techniques for using ROS with superior selectivity for microbial pathogens is required. As our contribution to this challenge, we have now proposed BC-associated PFRs as a promising novel low-cost and eco-friendly method for ROS generation to be studied for the oxidative inhibition of MDR pathogens and as a potential treatment for a wide range of infections. A greater knowledge concerning the proper pyrolysis conditions employed to obtain the type of PFRs more suitable for this purpose and in optimized concentration could make this ROS-delivering method more selective for bacterial pathogens.
Author Contributions
The authors (S.A., G.C.S., A.M.S. and G.Z.) contributed equally to this work.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Saliba, R.; Zahar, J.-R.; Dabar, G.; Riachy, M.; Karam-Sarkis, D.; Husni, R. Limiting the Spread of Multidrug-Resistant Bacteria in Low-to-Middle-Income Countries: One Size Does Not Fit All. Pathogens 2023, 12, 144. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Courvalin, P.; Dantas, G.; Davies, J.; Eisenstein, B.; Huovinen, P.; Jacoby, G.A.; Kishony, R.; Kreiswirth, B.N.; Kutter, E.; et al. Tackling Antibiotic Resistance. Nat Rev Microbiol 2011, 9, 894–896. [Google Scholar] [CrossRef] [PubMed]
- Franco, B.E.; Altagracia Martínez, M.; Sánchez Rodríguez, M.A.; Wertheimer, A.I. The Determinants of the Antibiotic Resistance Process. Infect Drug Resist 2009, 2, 1–11. [Google Scholar] [PubMed]
- Dryden, M. Reactive Oxygen Species: A Novel Antimicrobial. Int J Antimicrob Agents 2018, 51, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.C.; Martinez, L.; Kirsner, R. The Diabetic Foot: The Importance of Biofilms and Wound Bed Preparation. Curr Diab Rep 2006, 6, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Caviglia, D. Prevention and Eradication of Biofilm by Dendrimers: A Possibility Still Little Explored. Pharmaceutics 2022, 14, 2016. [Google Scholar] [CrossRef]
- Dryden, M.; Cooke, J.; Salib, R.; Holding, R.; Pender, S.L.F.; Brooks, J. Hot Topics in Reactive Oxygen Therapy: Antimicrobial and Immunological Mechanisms, Safety and Clinical Applications. J Glob Antimicrob Resist 2017, 8, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Dryden, M. Reactive Oxygen Therapy: A Novel Therapy in Soft Tissue Infection. Curr Opin Infect Dis 2017, 30, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Dryden, M.S.; Cooke, J.; Salib, R.J.; Holding, R.E.; Biggs, T.; Salamat, A.A.; Allan, R.N.; Newby, R.S.; Halstead, F.; Oppenheim, B.; et al. Reactive Oxygen: A Novel Antimicrobial Mechanism for Targeting Biofilm-Associated Infection. J Glob Antimicrob Resist 2017, 8, 186–191. [Google Scholar] [CrossRef]
- Dunnill, C.; Patton, T.; Brennan, J.; Barrett, J.; Dryden, M.; Cooke, J.; Leaper, D.; Georgopoulos, N.T. Reactive Oxygen Species (ROS) and Wound Healing: The Functional Role of ROS and Emerging ROS-modulating Technologies for Augmentation of the Healing Process. Int Wound J 2017, 14, 89–96. [Google Scholar] [CrossRef]
- Alfei, S.; Marengo, B.; Zuccari, G. Oxidative Stress, Antioxidant Capabilities, and Bioavailability: Ellagic Acid or Urolithins? Antioxidants 2020, 9, 707. [Google Scholar] [CrossRef] [PubMed]
- Genestra, M. Oxyl Radicals, Redox-Sensitive Signalling Cascades and Antioxidants. Cell Signal 2007, 19, 1807–1819. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, K.; Khurana, S.; Tai, T. Oxidative Stress in Aging-Matters of the Heart and Mind. Int J Mol Sci 2013, 14, 17897–17925. [Google Scholar] [CrossRef] [PubMed]
- Marengo, B.; Nitti, M.; Furfaro, A.L.; Colla, R.; Ciucis, C. De; Marinari, U.M.; Pronzato, M.A.; Traverso, N.; Domenicotti, C. Redox Homeostasis and Cellular Antioxidant Systems: Crucial Players in Cancer Growth and Therapy. Oxid Med Cell Longev 2016, 2016, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Marengo, B.; Raffaghello, L.; Pistoia, V.; Cottalasso, D.; Pronzato, M.A.; Marinari, U.M.; Domenicotti, C. Reactive Oxygen Species: Biological Stimuli of Neuroblastoma Cell Response. Cancer Lett 2005, 228, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, T.; N. Setzer, W.; Fazel Nabavi, S.; Erdogan Orhan, I.; Braidy, N.; Sobarzo-Sanchez, E.; Mohammad Nabavi, S. Insights Into Effects of Ellagic Acid on the Nervous System: A Mini Review. Curr Pharm Des 2016, 22, 1350–1360. [Google Scholar] [CrossRef]
- Kaneto, H.; Katakami, N.; Matsuhisa, M.; Matsuoka, T. Role of Reactive Oxygen Species in the Progression of Type 2 Diabetes and Atherosclerosis. Mediators Inflamm 2010, 2010, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Park, C.; Jang, H.-J.; Kim, B.; Bae, H.-W.; Chung, I.-Y.; Kim, E.S.; Cho, Y.-H. Antibacterial Strategies Inspired by the Oxidative Stress and Response Networks. Journal of Microbiology 2019, 57, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Gaupp, R.; Ledala, N.; Somerville, G.A. Staphylococcal Response to Oxidative Stress. Front Cell Infect Microbiol 2012, 2. [Google Scholar] [CrossRef]
- Hong, Y.; Zeng, J.; Wang, X.; Drlica, K.; Zhao, X. Post-Stress Bacterial Cell Death Mediated by Reactive Oxygen Species. Proceedings of the National Academy of Sciences 2019, 116, 10064–10071. [Google Scholar] [CrossRef]
- Dryden, M.; Lockyer, G.; Saeed, K.; Cooke, J. Engineered Honey: In Vitro Antimicrobial Activity of a Novel Topical Wound Care Treatment. J Glob Antimicrob Resist 2014, 2, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Vaishampayan, A.; Grohmann, E. Antimicrobials Functioning through ROS-Mediated Mechanisms: Current Insights. Microorganisms 2021, 10, 61. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Pandoli, O.G. Bamboo-Based Biochar: A Still Too Little-Studied Black Gold and Its Current Applications. J Xenobiot 2024, 14, 416–451. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Pandoli, O.G. Biochar-Derived Persistent Free Radicals: A Plethora of Environmental Applications in a Light and Shadows Scenario. Toxics 2024, 12, 245. [Google Scholar] [CrossRef] [PubMed]
- Beesley, L.; Moreno-Jiménez, E.; Gomez-Eyles, J.L.; Harris, E.; Robinson, B.; Sizmur, T. A Review of Biochars’ Potential Role in the Remediation, Revegetation and Restoration of Contaminated Soils. Environmental Pollution 2011, 159, 3269–3282. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, S.; Verheijen, F.G.A.; van der Velde, M.; Bastos, A.C. A Quantitative Review of the Effects of Biochar Application to Soils on Crop Productivity Using Meta-Analysis. Agric Ecosyst Environ 2011, 144, 175–187. [Google Scholar] [CrossRef]
- Kinney, T.J.; Masiello, C.A.; Dugan, B.; Hockaday, W.C.; Dean, M.R.; Zygourakis, K.; Barnes, R.T. Hydrologic Properties of Biochars Produced at Different Temperatures. Biomass Bioenergy 2012, 41, 34–43. [Google Scholar] [CrossRef]
- Qin, Y.; Li, G.; Gao, Y.; Zhang, L.; Ok, Y.S.; An, T. Persistent Free Radicals in Carbon-Based Materials on Transformation of Refractory Organic Contaminants (ROCs) in Water: A Critical Review. Water Res 2018, 137, 130–143. [Google Scholar] [CrossRef]
- Vejerano, E.P.; Rao, G.; Khachatryan, L.; Cormier, S.A.; Lomnicki, S. Environmentally Persistent Free Radicals: Insights on a New Class of Pollutants. Environ Sci Technol 2018, 52, 2468–2481. [Google Scholar] [CrossRef]
- Chen, Y.; Duan, X.; Zhang, C.; Wang, S.; Ren, N.; Ho, S.-H. Graphitic Biochar Catalysts from Anaerobic Digestion Sludge for Nonradical Degradation of Micropollutants and Disinfection. Chemical Engineering Journal 2020, 384, 123244. [Google Scholar] [CrossRef]
- Wang, T.; Zheng, J.; Cai, J.; Liu, Q.; Zhang, X. Visible-Light-Driven Photocatalytic Degradation of Dye and Antibiotics by Activated Biochar Composited with K+ Doped g-C3N4: Effects, Mechanisms, Actual Wastewater Treatment and Disinfection. Science of The Total Environment 2022, 839, 155955. [Google Scholar] [CrossRef]
- Shi, J.; Wang, J.; Liang, L.; Xu, Z.; Chen, Y.; Chen, S.; Xu, M.; Wang, X.; Wang, S. Carbothermal Synthesis of Biochar-Supported Metallic Silver for Enhanced Photocatalytic Removal of Methylene Blue and Antimicrobial Efficacy. J Hazard Mater 2021, 401, 123382. [Google Scholar] [CrossRef] [PubMed]
- Arshad, U.; Altaf, M.T.; Liaqat, W.; Ali, M.; Shah, M.N.; Jabran, M.; Ali, M.A. Biochar: Black Gold for Sustainable Agriculture and Fortification Against Plant Pathogens—A Review. Gesunde Pflanzen 2023. [Google Scholar] [CrossRef]
- Liu, X.; Chen, Z.; Lu, S.; Shi, X.; Qu, F.; Cheng, D.; Wei, W.; Shon, H.K.; Ni, B.-J. Persistent Free Radicals on Biochar for Its Catalytic Capability: A Review. Water Res 2024, 250, 120999. [Google Scholar] [CrossRef]
- Fang, G.; Liu, C.; Gao, J.; Dionysiou, D.D.; Zhou, D. Manipulation of Persistent Free Radicals in Biochar To Activate Persulfate for Contaminant Degradation. Environ Sci Technol 2015, 49, 5645–5653. [Google Scholar] [CrossRef]
- Odinga, E.S.; Waigi, M.G.; Gudda, F.O.; Wang, J.; Yang, B.; Hu, X.; Li, S.; Gao, Y. Occurrence, Formation, Environmental Fate and Risks of Environmentally Persistent Free Radicals in Biochars. Environ Int 2020, 134, 105172. [Google Scholar] [CrossRef]
- Andrés, C.M.C.; Pérez de la Lastra, J.M.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. Hypochlorous Acid Chemistry in Mammalian Cells-Influence on Infection and Role in Various Pathologies. Int J Mol Sci 2022, 23. [Google Scholar] [CrossRef]
- Adams, L.; Franco, M.C.; Estevez, A.G. Reactive Nitrogen Species in Cellular Signaling. Exp Biol Med 2015, 240, 711–717. [Google Scholar] [CrossRef]
- Salisbury, D.; Bronas, U. Reactive Oxygen and Nitrogen Species. Nurs Res 2015, 64, 53–66. [Google Scholar] [CrossRef] [PubMed]
- Genestra, M. Oxyl Radicals, Redox-Sensitive Signalling Cascades and Antioxidants. Cell Signal 2007, 19, 1807–1819. [Google Scholar] [CrossRef]
- Frijhoff, J.; Winyard, P.G.; Zarkovic, N.; Davies, S.S.; Stocker, R.; Cheng, D.; Knight, A.R.; Taylor, E.L.; Oettrich, J.; Ruskovska, T.; et al. Clinical Relevance of Biomarkers of Oxidative Stress. Antioxid Redox Signal 2015, 23, 1144–1170. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, E. Role of Protein Carbonylation in Skeletal Muscle Mass Loss Associated with Chronic Conditions. Proteomes 2016, 4, 18. [Google Scholar] [CrossRef] [PubMed]
- Trpkovic, A.; Resanovic, I.; Stanimirovic, J.; Radak, D.; Mousa, S.A.; Cenic-Milosevic, D.; Jevremovic, D.; Isenovic, E.R. Oxidized Low-Density Lipoprotein as a Biomarker of Cardiovascular Diseases. Crit Rev Clin Lab Sci 2015, 52, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Reynaert, N.L.; Gopal, P.; Rutten, E.P.A.; Wouters, E.F.M.; Schalkwijk, C.G. Advanced Glycation End Products and Their Receptor in Age-Related, Non-Communicable Chronic Inflammatory Diseases; Overview of Clinical Evidence and Potential Contributions to Disease. Int J Biochem Cell Biol 2016, 81, 403–418. [Google Scholar] [CrossRef] [PubMed]
- Jacob, K.D.; Noren Hooten, N.; Trzeciak, A.R.; Evans, M.K. Markers of Oxidant Stress That Are Clinically Relevant in Aging and Age-Related Disease. Mech Ageing Dev 2013, 134, 139–157. [Google Scholar] [CrossRef] [PubMed]
- Gaupp, R.; Ledala, N.; Somerville, G.A. Staphylococcal Response to Oxidative Stress. Front Cell Infect Microbiol 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Doukyu, N.; Taguchi, K. Involvement of Catalase and Superoxide Dismutase in Hydrophobic Organic Solvent Tolerance of Escherichia Coli. AMB Express 2021, 11, 97. [Google Scholar] [CrossRef]
- Chiang, S.M.; Schellhorn, H.E. Regulators of Oxidative Stress Response Genes in Escherichia Coli and Their Functional Conservation in Bacteria. Arch Biochem Biophys 2012, 525, 161–169. [Google Scholar] [CrossRef]
- Battesti, A.; Majdalani, N.; Gottesman, S. The RpoS-Mediated General Stress Response in Escherichia Coli. Annu Rev Microbiol 2011, 65, 189–213. [Google Scholar] [CrossRef]
- Nguyen, G.T.; Green, E.R.; Mecsas, J. Neutrophils to the ROScue: Mechanisms of NADPH Oxidase Activation and Bacterial Resistance. Front Cell Infect Microbiol 2017, 7. [Google Scholar] [CrossRef]
- Li, H.; Zhou, X.; Huang, Y.; Liao, B.; Cheng, L.; Ren, B. Reactive Oxygen Species in Pathogen Clearance: The Killing Mechanisms, the Adaption Response, and the Side Effects. Front Microbiol 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhu, L.; Pan, D.; Li, S.; Xiao, H.; Zhang, Z.; Shen, X.; Wang, Y.; Long, M. Siderophore-Mediated Iron Acquisition Enhances Resistance to Oxidative and Aromatic Compound Stress in Cupriavidus Necator JMP134. Appl Environ Microbiol 2019, 85. [Google Scholar] [CrossRef] [PubMed]
- Peralta, D.R.; Adler, C.; Corbalán, N.S.; Paz García, E.C.; Pomares, M.F.; Vincent, P.A. Enterobactin as Part of the Oxidative Stress Response Repertoire. PLoS One 2016, 11, e0157799. [Google Scholar] [CrossRef] [PubMed]
- Vaishampayan, A.; de Jong, A.; Wight, D.J.; Kok, J.; Grohmann, E. A Novel Antimicrobial Coating Represses Biofilm and Virulence-Related Genes in Methicillin-Resistant Staphylococcus Aureus. Front Microbiol 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; El Meouche, I.; Dunlop, M.J. Bacterial Persistence Induced by Salicylate via Reactive Oxygen Species. Sci Rep 2017, 7, 43839. [Google Scholar] [CrossRef] [PubMed]
- Grant, S.S.; Hung, D.T. Persistent Bacterial Infections, Antibiotic Tolerance, and the Oxidative Stress Response. Virulence 2013, 4, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Memar, M.Y.; Ghotaslou, R.; Samiei, M.; Adibkia, K. Antimicrobial Use of Reactive Oxygen Therapy: Current Insights. Infect Drug Resist 2018, 11, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Sampson, T.R.; Liu, X.; Schroeder, M.R.; Kraft, C.S.; Burd, E.M.; Weiss, D.S. Rapid Killing of Acinetobacter Baumannii by Polymyxins Is Mediated by a Hydroxyl Radical Death Pathway. Antimicrob Agents Chemother 2012, 56, 5642–5649. [Google Scholar] [CrossRef]
- Arriaga-Alba, M.; Rivera-Sánchez, R.; Parra-Cervantes, G.; Barro-Moreno, F.; Flores-Paz, R.; Garcı́a-Jiménez, E. Antimutagenesis of β-Carotene to Mutations Induced by Quinolone on Salmonella Typhimurium. Arch Med Res 2000, 31, 156–161. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, X. Contribution of Oxidative Damage to Antimicrobial Lethality. Antimicrob Agents Chemother 2009, 53, 1395–1402. [Google Scholar] [CrossRef]
- Rasouly, A.; Nudler, E. Reactive Oxygen Species as the Long Arm of Bactericidal Antibiotics. Proceedings of the National Academy of Sciences 2019, 116, 9696–9698. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Zeng, J.; Wang, X.; Drlica, K.; Zhao, X. Post-Stress Bacterial Cell Death Mediated by Reactive Oxygen Species. Proceedings of the National Academy of Sciences 2019, 116, 10064–10071. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Zhou, J.; Wang, P.; Li, T.; Zhao, Y.; Ren, X.; Lu, J.; Wang, J.; Holmgren, A.; Zou, L. Topical Therapeutic Efficacy of Ebselen Against Multidrug-Resistant Staphylococcus Aureus LT-1 Targeting Thioredoxin Reductase. Front Microbiol 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Mourenza, Á.; Gil, J.A.; Mateos, L.M.; Letek, M. Oxidative Stress-Generating Antimicrobials, a Novel Strategy to Overcome Antibacterial Resistance. Antioxidants 2020, 9, 361. [Google Scholar] [CrossRef] [PubMed]
- Díaz-García, D.; Ardiles, P.; Prashar, S.; Rodríguez-Diéguez, A.; Páez, P.; Gómez-Ruiz, S. Preparation and Study of the Antibacterial Applications and Oxidative Stress Induction of Copper Maleamate-Functionalized Mesoporous Silica Nanoparticles. Pharmaceutics 2019, 11, 30. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Shao, T.; Yu, Y.; Xiong, Y.; Yang, L. Surface-Bound Reactive Oxygen Species Generating Nanozymes for Selective Antibacterial Action. Nat Commun 2021, 12, 745. [Google Scholar] [CrossRef] [PubMed]
- Linzner, N.; Antelmann, H. The Antimicrobial Activity of the AGXX® Surface Coating Requires a Small Particle Size to Efficiently Kill Staphylococcus Aureus. Front Microbiol 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Clauss-Lendzian, E.; Vaishampayan, A.; de Jong, A.; Landau, U.; Meyer, C.; Kok, J.; Grohmann, E. Stress Response of a Clinical Enterococcus Faecalis Isolate Subjected to a Novel Antimicrobial Surface Coating. Microbiol Res 2018, 207, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Loi, V. Van; Busche, T.; Preuß, T.; Kalinowski, J.; Bernhardt, J.; Antelmann, H. The AGXX® Antimicrobial Coating Causes a Thiol-Specific Oxidative Stress Response and Protein S-Bacillithiolation in Staphylococcus Aureus. Front Microbiol 2018, 9. [Google Scholar] [CrossRef]
- Loi, V. Van; Huyen, N.T.T.; Busche, T.; Tung, Q.N.; Gruhlke, M.C.H.; Kalinowski, J.; Bernhardt, J.; Slusarenko, A.J.; Antelmann, H. Staphylococcus Aureus Responds to Allicin by Global S-Thioallylation – Role of the Brx/BSH/YpdA Pathway and the Disulfide Reductase MerA to Overcome Allicin Stress. Free Radic Biol Med 2019, 139, 55–69. [Google Scholar] [CrossRef]
- Paulander, W.; Wang, Y.; Folkesson, A.; Charbon, G.; Løbner-Olesen, A.; Ingmer, H. Bactericidal Antibiotics Increase Hydroxyphenyl Fluorescein Signal by Altering Cell Morphology. PLoS One 2014, 9, e92231. [Google Scholar] [CrossRef] [PubMed]
- Kohanski, M.A.; Dwyer, D.J.; Hayete, B.; Lawrence, C.A.; Collins, J.J. A Common Mechanism of Cellular Death Induced by Bactericidal Antibiotics. Cell 2007, 130, 797–810. [Google Scholar] [CrossRef] [PubMed]
- Chua, N.G.; Zhou, Y.P.; Tan, T.T.; Lingegowda, P.B.; Lee, W.; Lim, T.P.; Teo, J.; Cai, Y.; Kwa, A.L. Polymyxin B with Dual Carbapenem Combination Therapy against Carbapenemase-Producing Klebsiella Pneumoniae. Journal of Infection 2015, 70, 309–311. [Google Scholar] [CrossRef] [PubMed]
- Alfei, S.; Schito, A.M. Positively Charged Polymers as Promising Devices against Multidrug Resistant Gram-Negative Bacteria: A Review. Polymers (Basel) 2020, 12, 1195. [Google Scholar] [CrossRef] [PubMed]
- Ezraty, B.; Vergnes, A.; Banzhaf, M.; Duverger, Y.; Huguenot, A.; Brochado, A.R.; Su, S.-Y.; Espinosa, L.; Loiseau, L.; Py, B.; et al. Fe-S Cluster Biosynthesis Controls Uptake of Aminoglycosides in a ROS-Less Death Pathway. Science (1979) 2013, 340, 1583–1587. [Google Scholar] [CrossRef] [PubMed]
- Keren, I.; Wu, Y.; Inocencio, J.; Mulcahy, L.R.; Lewis, K. Killing by Bactericidal Antibiotics Does Not Depend on Reactive Oxygen Species. Science (1979) 2013, 339, 1213–1216. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Imlay, J.A. Cell Death from Antibiotics Without the Involvement of Reactive Oxygen Species. Science (1979) 2013, 339, 1210–1213. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Li, Q.; Wang, J.; Yu, Y.; Wang, Y.; Zhou, Q.; Li, P. Reactive Oxygen Species-Related Nanoparticle Toxicity in the Biomedical Field. Nanoscale Res Lett 2020, 15, 115. [Google Scholar] [CrossRef] [PubMed]
- Linzner, N.; Antelmann, H. The Antimicrobial Activity of the AGXX® Surface Coating Requires a Small Particle Size to Efficiently Kill Staphylococcus Aureus. Front Microbiol 2021, 12. [Google Scholar] [CrossRef]
- Cieplik, F.; Deng, D.; Crielaard, W.; Buchalla, W.; Hellwig, E.; Al-Ahmad, A.; Maisch, T. Antimicrobial Photodynamic Therapy – What We Know and What We Don’t. Crit Rev Microbiol 2018, 44, 571–589. [Google Scholar] [CrossRef]
- Mitton, D.; Ackroyd, R. A Brief Overview of Photodynamic Therapy in Europe. Photodiagnosis Photodyn Ther 2008, 5, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Dolmans, D.E.J.G.J.; Fukumura, D.; Jain, R.K. Photodynamic Therapy for Cancer. Nat Rev Cancer 2003, 3, 380–387. [Google Scholar] [CrossRef] [PubMed]
- Ackroyd, R.; Kelty, C.; Brown, N.; Reed, M. The History of Photodetection and Photodynamic Therapy. Photochem Photobiol 2001, 74, 656–669. [Google Scholar] [CrossRef] [PubMed]
- Wainwright, M. Dyes, Flies, and Sunny Skies: Photodynamic Therapy and Neglected Tropical Diseases. Coloration Technology 2017, 133, 3–14. [Google Scholar] [CrossRef]
- Wainwright, M.; Maisch, T.; Nonell, S.; Plaetzer, K.; Almeida, A.; Tegos, G.P.; Hamblin, M.R. Photoantimicrobials—Are We Afraid of the Light? Lancet Infect Dis 2017, 17, e49–e55. [Google Scholar] [CrossRef] [PubMed]
- Correia, J.H.; Rodrigues, J.A.; Pimenta, S.; Dong, T.; Yang, Z. Photodynamic Therapy Review: Principles, Photosensitizers, Applications, and Future Directions. Pharmaceutics 2021, 13, 1332. [Google Scholar] [CrossRef] [PubMed]
- Lutkus, L. V.; Rickenbach, S.S.; McCormick, T.M. Singlet Oxygen Quantum Yields Determined by Oxygen Consumption. J Photochem Photobiol A Chem 2019, 378, 131–135. [Google Scholar] [CrossRef]
- Liu, Y.; Qin, R.; Zaat, S.A.J.; Breukink, E.; Heger, M. Antibacterial Photodynamic Therapy: Overview of a Promising Approach to Fight Antibiotic-Resistant Bacterial Infections. J Clin Transl Res 2015, 1, 140–167. [Google Scholar]
- Wainwright, M. Photoantimicrobials and PACT: What’s in an Abbreviation? Photochemical & Photobiological Sciences 2019, 18, 12–14. [Google Scholar] [CrossRef]
- Yan, E.; Kwek, G.; Qing, N.S.; Lingesh, S.; Xing, B. Antimicrobial Photodynamic Therapy for the Remote Eradication of Bacteria. Chempluschem 2023, 88. [Google Scholar] [CrossRef]
- Hamblin, M.R.; Abrahamse, H. Oxygen-Independent Antimicrobial Photoinactivation: Type III Photochemical Mechanism? Antibiotics 2020, 9, 53. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, J.; Rahban, D.; Aghamiri, S.; Teymouri, A.; Bahador, A. Photosensitizers in Antibacterial Photodynamic Therapy: An Overview. Laser Ther 2018, 27, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Youf, R.; Müller, M.; Balasini, A.; Thétiot, F.; Müller, M.; Hascoët, A.; Jonas, U.; Schönherr, H.; Lemercier, G.; Montier, T.; et al. Antimicrobial Photodynamic Therapy: Latest Developments with a Focus on Combinatory Strategies. Pharmaceutics 2021, 13, 1995. [Google Scholar] [CrossRef] [PubMed]
- Fontana, C.R.; Abernethy, A.D.; Som, S.; Ruggiero, K.; Doucette, S.; Marcantonio, R.C.; Boussios, C.I.; Kent, R.; Goodson, J.M.; Tanner, A.C.R.; et al. The Antibacterial Effect of Photodynamic Therapy in Dental Plaque-derived Biofilms. J Periodontal Res 2009, 44, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Zanin, I.C.J.; Lobo, M.M.; Rodrigues, L.K.A.; Pimenta, L.A.F.; Höfling, J.F.; Gonçalves, R.B. Photosensitization of in Vitro Biofilms by Toluidine Blue O Combined with a Light-emitting Diode. Eur J Oral Sci 2006, 114, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Fekrazad, R.; Zare, H.; Vand, S.M.S. Photodynamic Therapy Effect on Cell Growth Inhibition Induced by Radachlorin and Toluidine Blue O on Staphylococcus Aureus and Escherichia Coli: An in Vitro Study. Photodiagnosis Photodyn Ther 2016, 15, 213–217. [Google Scholar] [CrossRef] [PubMed]
- Voos, A.C.; Kranz, S.; Tonndorf-Martini, S.; Voelpel, A.; Sigusch, H.; Staudte, H.; Albrecht, V.; Sigusch, B.W. Photodynamic Antimicrobial Effect of Safranine O on an Ex Vivo Periodontal Biofilm. Lasers Surg Med 2014, 46, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Collins, T.L.; Markus, E.A.; Hassett, D.J.; Robinson, J.B. The Effect of a Cationic Porphyrin on Pseudomonas Aeruginosa Biofilms. Curr Microbiol 2010, 61, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Di Poto, A.; Sbarra, M.S.; Provenza, G.; Visai, L.; Speziale, P. The Effect of Photodynamic Treatment Combined with Antibiotic Action or Host Defence Mechanisms on Staphylococcus Aureus Biofilms. Biomaterials 2009, 30, 3158–3166. [Google Scholar] [CrossRef]
- Cieplik, F.; Späth, A.; Regensburger, J.; Gollmer, A.; Tabenski, L.; Hiller, K.-A.; Bäumler, W.; Maisch, T.; Schmalz, G. Photodynamic Biofilm Inactivation by SAPYR—An Exclusive Singlet Oxygen Photosensitizer. Free Radic Biol Med 2013, 65, 477–487. [Google Scholar] [CrossRef]
- Malá, Z.; Žárská, L.; Bajgar, R.; Bogdanová, K.; Kolář, M.; Panáček, A.; Binder, S.; Kolářová, H. The Application of Antimicrobial Photodynamic Inactivation on Methicillin-Resistant S. Aureus and ESBL-Producing K. Pneumoniae Using Porphyrin Photosensitizer in Combination with Silver Nanoparticles. Photodiagnosis Photodyn Ther 2021, 33, 102140. [Google Scholar] [CrossRef] [PubMed]
- Board-Davies, E.L.; Rhys-Williams, W.; Hynes, D.; Williams, D.; Farnell, D.J.J.; Love, W. Antibacterial and Antibiofilm Potency of XF Drugs, Impact of Photodynamic Activation and Synergy With Antibiotics. Front Cell Infect Microbiol 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Karygianni, L.; Ruf, S.; Follo, M.; Hellwig, E.; Bucher, M.; Anderson, A.C.; Vach, K.; Al-Ahmad, A. Novel Broad-Spectrum Antimicrobial Photoinactivation of In Situ Oral Biofilms by Visible Light plus Water-Filtered Infrared A. Appl Environ Microbiol 2014, 80, 7324–7336. [Google Scholar] [CrossRef] [PubMed]
- Mesquita, M.Q.; Menezes, J.C.J.M.D.S.; Neves, M.G.P.M.S.; Tomé, A.C.; Cavaleiro, J.A.S.; Cunha, Â.; Almeida, A.; Hackbarth, S.; Röder, B.; Faustino, M.A.F. Photodynamic Inactivation of Bioluminescent Escherichia Coli by Neutral and Cationic Pyrrolidine-Fused Chlorins and Isobacteriochlorins. Bioorg Med Chem Lett 2014, 24, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Souza, B.M.N.; Pinto, J.G.; Pereira, A.H.C.; Miñán, A.G.; Ferreira-Strixino, J. Efficiency of Antimicrobial Photodynamic Therapy with Photodithazine® on MSSA and MRSA Strains. Antibiotics (Basel) 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Bertoloni, G.; Rossi, F.; Valduga, G.; Jori, G.; Ali, H.; van Lier, J.E. Photosensitizing Activity of Water- and Lipid-Soluble Phthalocyanines on Prokaryotic and Eukaryotic Microbial Cells. Microbios 1992, 71, 33–46. [Google Scholar] [PubMed]
- Fan, B.; Peng, W.; Zhang, Y.; Liu, P.; Shen, J. ROS Conversion Promotes the Bactericidal Efficiency of Eosin Y Based Photodynamic Therapy. Biomater Sci 2023, 11, 4930–4937. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.L.L.; Sobral, A.P.T.; Gallo, J.M.A.S.; Gimenez, T.; Ferri, E.P.; Ianello, S.; Motta, P. de B.; Motta, L.J.; Horliana, A.C.R.T.; Santos, E.M.; et al. Antimicrobial Photodynamic Therapy with Erythrosine and Blue Light on Dental Biofilm Bacteria: Study Protocol for Randomised Clinical Trial. BMJ Open 2023, 13, e075084. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.; Kishen, A. Polycationic Chitosan-Conjugated Photosensitizer for Antibacterial Photodynamic Therapy †. Photochem Photobiol 2012, 88, 577–583. [Google Scholar] [CrossRef]
- Spesia, M.B.; Milanesio, M.E.; Durantini, E.N. Synthesis, Properties and Photodynamic Inactivation of Escherichia Coli by Novel Cationic Fullerene C60 Derivatives. Eur J Med Chem 2008, 43, 853–861. [Google Scholar] [CrossRef]
- Wang, M.; Huang, L.; Sharma, S.K.; Jeon, S.; Thota, S.; Sperandio, F.F.; Nayka, S.; Chang, J.; Hamblin, M.R.; Chiang, L.Y. Synthesis and Photodynamic Effect of New Highly Photostable Decacationically Armed [60]- and [70]Fullerene Decaiodide Monoadducts To Target Pathogenic Bacteria and Cancer Cells. J Med Chem 2012, 55, 4274–4285. [Google Scholar] [CrossRef]
- Cieplik, F.; Pummer, A.; Regensburger, J.; Hiller, K.-A.; Späth, A.; Tabenski, L.; Buchalla, W.; Maisch, T. The Impact of Absorbed Photons on Antimicrobial Photodynamic Efficacy. Front Microbiol 2015, 6. [Google Scholar] [CrossRef]
- Maisch, T.; Eichner, A.; Späth, A.; Gollmer, A.; König, B.; Regensburger, J.; Bäumler, W. Fast and Effective Photodynamic Inactivation of Multiresistant Bacteria by Cationic Riboflavin Derivatives. PLoS One 2014, 9, e111792. [Google Scholar] [CrossRef] [PubMed]
- Najafi, S.; Khayamzadeh, M.; Paknejad, M.; Poursepanj, G.; Kharazi Fard, M.J.; Bahador, A. An In Vitro Comparison of Antimicrobial Effects of Curcumin-Based Photodynamic Therapy and Chlorhexidine, on Aggregatibacter Actinomycetemcomitans. J Lasers Med Sci 2016, 7, 21–25. [Google Scholar] [CrossRef]
- Dahl, T.A.; McGowan, W.M.; Shand, M.A.; Srinivasan, V.S. Photokilling of Bacteria by the Natural Dye Curcumin. Arch Microbiol 1989, 151, 183–185. [Google Scholar] [CrossRef]
- García, I.; Ballesta, S.; Gilaberte, Y.; Rezusta, A.; Pascual, Á. Antimicrobial Photodynamic Activity of Hypericin against Methicillin-Susceptible and Resistant Staphylococcus Aureus Biofilms. Future Microbiol 2015, 10, 347–356. [Google Scholar] [CrossRef]
- Yow, C.M.N.; Tang, H.M.; Chu, E.S.M.; Huang, Z. Hypericin-mediated Photodynamic Antimicrobial Effect on Clinically Isolated Pathogens †. Photochem Photobiol 2012, 88, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, K.; Ozawa, T.; Awazu, K.; Ito, N.; Honda, N.; Matsumoto, S.; Tsuruta, D. Photodynamic Therapy Using Systemic Administration of 5-Aminolevulinic Acid and a 410-Nm Wavelength Light-Emitting Diode for Methicillin-Resistant Staphylococcus Aureus-Infected Ulcers in Mice. PLoS One 2014, 9, e105173. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, B.; Zhang, L.; Liu, X.; Shi, L.; Kang, X.; Lei, X.; Chen, K.; Chen, Z.; Li, C.; et al. Clinical Practice Guidelines for 5-Aminolevulinic Acid Photodynamic Therapy for Acne Vulgaris in China. Photodiagnosis Photodyn Ther 2023, 41, 103261. [Google Scholar] [CrossRef]
- Polat, E.; Kang, K. Natural Photosensitizers in Antimicrobial Photodynamic Therapy. Biomedicines 2021, 9, 584. [Google Scholar] [CrossRef]
- Abrahamse, H.; Hamblin, M.R. New Photosensitizers for Photodynamic Therapy. Biochemical Journal 2016, 473, 347–364. [Google Scholar] [CrossRef] [PubMed]
- Gunaydin, G.; Gedik, M.E.; Ayan, S. Photodynamic Therapy-Current Limitations and Novel Approaches. Front Chem 2021, 9, 691697. [Google Scholar] [CrossRef] [PubMed]
- Clément, S.; Winum, J.-Y. Photodynamic Therapy Alone or in Combination to Counteract Bacterial Infections. Expert Opin Ther Pat 2024, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Embleton, M.L. Selective Lethal Photosensitization of Methicillin-Resistant Staphylococcus Aureus Using an IgG-Tin (IV) Chlorin E6 Conjugate. Journal of Antimicrobial Chemotherapy 2002, 50, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Dosselli, R.; Gobbo, M.; Bolognini, E.; Campestrini, S.; Reddi, E. Porphyrin−Apidaecin Conjugate as a New Broad Spectrum Antibacterial Agent. ACS Med Chem Lett 2010, 1, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Sperandio, F.; Huang, Y.-Y.; Hamblin, M. Antimicrobial Photodynamic Therapy to Kill Gram-Negative Bacteria. Recent Pat Antiinfect Drug Discov 2013, 8, 108–120. [Google Scholar] [CrossRef] [PubMed]
- Rice, D.R.; Gan, H.; Smith, B.D. Bacterial Imaging and Photodynamic Inactivation Using Zinc(Ii)-Dipicolylamine BODIPY Conjugates. Photochemical & Photobiological Sciences 2015, 14, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Soto-Moreno, A.; Montero-Vilchez, T.; Diaz-Calvillo, P.; Molina-Leyva, A.; Arias-Santiago, S. The Impact of Photodynamic Therapy on Skin Homeostasis in Patients with Actinic Keratosis: A Prospective Observational Study. Skin Res Technol 2023, 29, e13493. [Google Scholar] [CrossRef]
- Rapacka-Zdończyk, A.; Woźniak, A.; Michalska, K.; Pierański, M.; Ogonowska, P.; Grinholc, M.; Nakonieczna, J. Factors Determining the Susceptibility of Bacteria to Antibacterial Photodynamic Inactivation. Front Med (Lausanne) 2021, 8. [Google Scholar] [CrossRef]
- Klausen, M.; Ucuncu, M.; Bradley, M. Design of Photosensitizing Agents for Targeted Antimicrobial Photodynamic Therapy. Molecules 2020, 25, 5239. [Google Scholar] [CrossRef]
- Zhou, W.; Jiang, X.; Zhen, X. Development of Organic Photosensitizers for Antimicrobial Photodynamic Therapy. Biomater Sci 2023, 11, 5108–5128. [Google Scholar] [CrossRef] [PubMed]
- Badran, Z.; Rahman, B.; De Bonfils, P.; Nun, P.; Coeffard, V.; Verron, E. Antibacterial Nanophotosensitizers in Photodynamic Therapy: An Update. Drug Discov Today 2023, 28, 103493. [Google Scholar] [CrossRef] [PubMed]
- Piksa, M.; Lian, C.; Samuel, I.C.; Pawlik, K.J.; Samuel, I.D.W.; Matczyszyn, K. The Role of the Light Source in Antimicrobial Photodynamic Therapy. Chem Soc Rev 2023, 52, 1697–1722. [Google Scholar] [CrossRef] [PubMed]
- Usui, Y. DETERMINATION OF QUANTUM YIELD OF SINGLET OXYGEN FORMATION BY PHOTOSENSITIZATION. Chem Lett 1973, 2, 743–744. [Google Scholar] [CrossRef]
- Wiegell, S.R.; Skødt, V.; Wulf, H.C. Daylight-mediated Photodynamic Therapy of Basal Cell Carcinomas – an Explorative Study. Journal of the European Academy of Dermatology and Venereology 2014, 28, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Al-Mutairi, R.; Tovmasyan, A.; Batinic-Haberle, I.; Benov, L. Sublethal Photodynamic Treatment Does Not Lead to Development of Resistance. Front Microbiol 2018, 9. [Google Scholar] [CrossRef]
- Nakonieczna, J.; Wozniak, A.; Pieranski, M.; Rapacka-Zdonczyk, A.; Ogonowska, P.; Grinholc, M. Photoinactivation of ESKAPE Pathogens: Overview of Novel Therapeutic Strategy. Future Med Chem 2019, 11, 443–461. [Google Scholar] [CrossRef] [PubMed]
- Songca, S.P.; Adjei, Y. Applications of Antimicrobial Photodynamic Therapy against Bacterial Biofilms. Int J Mol Sci 2022, 23, 3209. [Google Scholar] [CrossRef]
- Ribeiro, M.; Gomes, I.B.; Saavedra, M.J.; Simões, M. Photodynamic Therapy and Combinatory Treatments for the Control of Biofilm-Associated Infections. Lett Appl Microbiol 2022, 75, 548–564. [Google Scholar] [CrossRef]
- Barolet, D.; Boucher, A. Radiant near Infrared Light Emitting Diode Exposure as Skin Preparation to Enhance Photodynamic Therapy Inflammatory Type Acne Treatment Outcome. Lasers Surg Med 2010, 42, 171–178. [Google Scholar] [CrossRef]
- Sakamoto, F.H.; Torezan, L.; Anderson, R.R. Photodynamic Therapy for Acne Vulgaris: A Critical Review from Basics to Clinical Practice. J Am Acad Dermatol 2010, 63, 195–211. [Google Scholar] [CrossRef] [PubMed]
- Boen, M.; Brownell, J.; Patel, P.; Tsoukas, M.M. The Role of Photodynamic Therapy in Acne: An Evidence-Based Review. Am J Clin Dermatol 2017, 18, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Wojewoda, K.; Gillstedt, M.; Tovi, J.; Salah, L.; Wennberg Larkö, A.-M.; Sjöholm, A.; Sandberg, C. Optimizing Treatment of Acne with Photodynamic Therapy (PDT) to Achieve Long-Term Remission and Reduce Side Effects. A Prospective Randomized Controlled Trial. J Photochem Photobiol B 2021, 223, 112299. [Google Scholar] [CrossRef] [PubMed]
- Ning, X.; He, G.; Zeng, W.; Xia, Y. The Photosensitizer-Based Therapies Enhance the Repairing of Skin Wounds. Front Med (Lausanne) 2022, 9. [Google Scholar] [CrossRef]
- Brown, S. Clinical Antimicrobial Photodynamic Therapy: Phase II Studies in Chronic Wounds. Journal of the National Comprehensive Cancer Network 2012, 10, S–80. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Laguna, V.; Gilaberte, Y.; Millán-Lou, M.I.; Agut, M.; Nonell, S.; Rezusta, A.; Hamblin, M.R. A Combination of Photodynamic Therapy and Antimicrobial Compounds to Treat Skin and Mucosal Infections: A Systematic Review. Photochemical & Photobiological Sciences 2019, 18, 1020–1029. [Google Scholar] [CrossRef] [PubMed]
- Prażmo, E.; Mielczarek, A.; Kwaśny, M.; Łapiński, M. Photodynamic Therapy As a Promising Method Used in the Treatment of Oral Diseases. Advances in Clinical and Experimental Medicine 2016, 25, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Dragana, R.; Jelena, M.; Jovan, M.; Biljana, N.; Dejan, M. Antibacterial Efficiency of Adjuvant Photodynamic Therapy and High-Power Diode Laser in the Treatment of Young Permanent Teeth with Chronic Periapical Periodontitis. A Prospective Clinical Study. Photodiagnosis Photodyn Ther 2023, 41, 103129. [Google Scholar] [CrossRef] [PubMed]
- Vohra, F.; Akram, Z.; Safii, S.H.; Vaithilingam, R.D.; Ghanem, A.; Sergis, K.; Javed, F. Role of Antimicrobial Photodynamic Therapy in the Treatment of Aggressive Periodontitis: A Systematic Review. Photodiagnosis Photodyn Ther 2016, 13, 139–147. [Google Scholar] [CrossRef]
- Bechara Andere, N.M.R.; dos Santos, N.C.C.; Araujo, C.F.; Mathias, I.F.; Rossato, A.; de Marco, A.C.; Santamaria, M.; Jardini, M.A.N.; Santamaria, M.P. Evaluation of the Local Effect of Nonsurgical Periodontal Treatment with and without Systemic Antibiotic and Photodynamic Therapy in Generalized Aggressive Periodontitis. A Randomized Clinical Trial. Photodiagnosis Photodyn Ther 2018, 24, 115–120. [Google Scholar] [CrossRef]
- Ohba, S.; Sato, M.; Noda, S.; Yamamoto, H.; Egahira, K.; Asahina, I. Assessment of Safety and Efficacy of Antimicrobial Photodynamic Therapy for Peri-Implant Disease. Photodiagnosis Photodyn Ther 2020, 31, 101936. [Google Scholar] [CrossRef]
- Yupanqui Mieles, J.; Vyas, C.; Aslan, E.; Humphreys, G.; Diver, C.; Bartolo, P. Honey: An Advanced Antimicrobial and Wound Healing Biomaterial for Tissue Engineering Applications. Pharmaceutics 2022, 14, 1663. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Liu, R.; Lu, Q.; Hao, P.; Xu, A.; Zhang, J.; Tan, J. Biochemical Properties, Antibacterial and Cellular Antioxidant Activities of Buckwheat Honey in Comparison to Manuka Honey. Food Chem 2018, 252, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Girma, A.; Seo, W.; She, R.C. Antibacterial Activity of Varying UMF-Graded Manuka Honeys. PLoS One 2019, 14, e0224495. [Google Scholar] [CrossRef] [PubMed]
- Cokcetin, N.; W.S.; B.S.; C.D.; B.P.; H.L. Cokcetin, N.; Williams, S.; Blair, S.; Carter, D.; Brooks, P.; Harry, L.Active Australian Leptospermum Honey: New Sources and Their Bioactivity; Agrifutures Australia:Canberra, Australia,.; 2019;
- Brudzynski, K. Effect of Hydrogen Peroxide on Antibacterial Activities of Canadian Honeys. Can J Microbiol 2006, 52, 1228–1237. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Suarez, J.M.; Tulipani, S.; Díaz, D.; Estevez, Y.; Romandini, S.; Giampieri, F.; Damiani, E.; Astolfi, P.; Bompadre, S.; Battino, M. Antioxidant and Antimicrobial Capacity of Several Monofloral Cuban Honeys and Their Correlation with Color, Polyphenol Content and Other Chemical Compounds. Food and Chemical Toxicology 2010, 48, 2490–2499. [Google Scholar] [CrossRef]
- Sherlock, O.; Dolan, A.; Athman, R.; Power, A.; Gethin, G.; Cowman, S.; Humphreys, H. Comparison of the Antimicrobial Activity of Ulmo Honey from Chile and Manuka Honey against Methicillin-Resistant Staphylococcus Aureus, Escherichia Coli and Pseudomonas Aeruginosa. BMC Complement Altern Med 2010, 10, 47. [Google Scholar] [CrossRef] [PubMed]
- Isla, M.I.; Craig, A.; Ordoñez, R.; Zampini, C.; Sayago, J.; Bedascarrasbure, E.; Alvarez, A.; Salomón, V.; Maldonado, L. Physico Chemical and Bioactive Properties of Honeys from Northwestern Argentina. LWT - Food Science and Technology 2011, 44, 1922–1930. [Google Scholar] [CrossRef]
- Fyfe, L.; Okoro, P.; Paterson, E.; Coyle, S.; McDougall, G.J. Compositional Analysis of Scottish Honeys with Antimicrobial Activity against Antibiotic-Resistant Bacteria Reveals Novel Antimicrobial Components. LWT - Food Science and Technology 2017, 79, 52–59. [Google Scholar] [CrossRef]
- Escuredo, O.; Silva, L.R.; Valentão, P.; Seijo, M.C.; Andrade, P.B. Assessing Rubus Honey Value: Pollen and Phenolic Compounds Content and Antibacterial Capacity. Food Chem 2012, 130, 671–678. [Google Scholar] [CrossRef]
- Matzen, R.D.; Zinck Leth-Espensen, J.; Jansson, T.; Nielsen, D.S.; Lund, M.N.; Matzen, S. The Antibacterial Effect In Vitro of Honey Derived from Various Danish Flora. Dermatol Res Pract 2018, 2018, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bucekova, M.; Jardekova, L.; Juricova, V.; Bugarova, V.; Di Marco, G.; Gismondi, A.; Leonardi, D.; Farkasovska, J.; Godocikova, J.; Laho, M.; et al. Antibacterial Activity of Different Blossom Honeys: New Findings. Molecules 2019, 24, 1573. [Google Scholar] [CrossRef]
- Ahmed, G. Hegazi Antimicrobial Activity of Different Egyptian Honeys as Comparison of Saudi Arabia Honey. Res J Microbiol 2011, 6, 488–495. [Google Scholar]
- Laallam, H.; Boughediri, L.; Bissati, S.; Menasria, T.; Mouzaoui, M.S.; Hadjadj, S.; Hammoudi, R.; Chenchouni, H. Modeling the Synergistic Antibacterial Effects of Honey Characteristics of Different Botanical Origins from the Sahara Desert of Algeria. Front Microbiol 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- John-Isa, J.F.; Adebolu, T.T.; Oyetayo, V.O. Antibacterial Effects of Honey in Nigeria on Selected Diarrhoeagenic Bacteria. South Asian Journal of Research in Microbiology 2019, 1–11. [Google Scholar] [CrossRef]
- ElBorai, A. Antibacterial and Antioxidant Activities of Different Varieties of Locally Produced Egyptian Honey. Egyptian Journal of Botany 2018, 0, 0–0. [Google Scholar] [CrossRef]
- Kwakman, P.H.S.; Zaat, S.A.J. Antibacterial Components of Honey. IUBMB Life 2012, 64, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Moore, O.A.; Smith, L.A.; Campbell, F.; Seers, K.; McQuay, H.J.; Moore, R.A. Systematic Review of the Use of Honey as a Wound Dressing. BMC Complement Altern Med 2001, 1, 2. [Google Scholar] [CrossRef] [PubMed]
- Jull, A.B.; Rodgers, A.; Walker, N. Honey as a Topical Treatment for Wounds. In Cochrane Database of Systematic Reviews; Jull, A.B., Ed.; John Wiley & Sons, Ltd: Chichester, UK, 2008. [Google Scholar]
- Johnson, D.W.; van Eps, C.; Mudge, D.W.; Wiggins, K.J.; Armstrong, K.; Hawley, C.M.; Campbell, S.B.; Isbel, N.M.; Nimmo, G.R.; Gibbs, H. Randomized, Controlled Trial of Topical Exit-Site Application of Honey (Medihoney) versus Mupirocin for the Prevention of Catheter-Associated Infections in Hemodialysis Patients. Journal of the American Society of Nephrology 2005, 16, 1456–1462. [Google Scholar] [CrossRef]
- Love, N.R.; Chen, Y.; Ishibashi, S.; Kritsiligkou, P.; Lea, R.; Koh, Y.; Gallop, J.L.; Dorey, K.; Amaya, E. Amputation-Induced Reactive Oxygen Species Are Required for Successful Xenopus Tadpole Tail Regeneration. Nat Cell Biol 2013, 15, 222–228. [Google Scholar] [CrossRef]
- Hixon, K.R.; Klein, R.C.; Eberlin, C.T.; Linder, H.R.; Ona, W.J.; Gonzalez, H.; Sell, S.A. A Critical Review and Perspective of Honey in Tissue Engineering and Clinical Wound Healing. Adv Wound Care (New Rochelle) 2019, 8, 403–415. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Li, W.; Zhang, F.; Liu, Z.; Zanjanizadeh Ezazi, N.; Liu, D.; Santos, H.A. Electrospun Fibrous Architectures for Drug Delivery, Tissue Engineering and Cancer Therapy. Adv Funct Mater 2019, 29. [Google Scholar] [CrossRef]
- Li, J.; Mooney, D.J. Designing Hydrogels for Controlled Drug Delivery. Nat Rev Mater 2016, 1, 16071. [Google Scholar] [CrossRef] [PubMed]
- Rossi, M.; Marrazzo, P. The Potential of Honeybee Products for Biomaterial Applications. Biomimetics 2021, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Minden-Birkenmaier, B.; Bowlin, G. Honey-Based Templates in Wound Healing and Tissue Engineering. Bioengineering 2018, 5, 46. [Google Scholar] [CrossRef] [PubMed]
- Halstead, F.D.; Webber, M.A.; Rauf, M.; Burt, R.; Dryden, M.; Oppenheim, B.A. In Vitro Activity of an Engineered Honey, Medical-Grade Honeys, and Antimicrobial Wound Dressings against Biofilm-Producing Clinical Bacterial Isolates. J Wound Care 2016, 25, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Cooke, J.; Dryden, M.; Patton, T.; Brennan, J.; Barrett, J. The Antimicrobial Activity of Prototype Modified Honeys That Generate Reactive Oxygen Species (ROS) Hydrogen Peroxide. BMC Res Notes 2015, 8, 20. [Google Scholar] [CrossRef]
- Wilkinson, J.M.; Cavanagh, H.M.A. Antibacterial Activity of 13 Honeys Against Escherichia Coli and Pseudomonas Aeruginosa. J Med Food 2005, 8, 100–103. [Google Scholar] [CrossRef]
- Mullai, V.; Menon, T. Bactericidal Activity of Different Types of Honey against Clinical and Environmental Isolates of Pseudomonas Aeruginosa. The Journal of Alternative and Complementary Medicine 2007, 13, 439–442. [Google Scholar] [CrossRef]
- Percival, S.L.; Hill, K.E.; Williams, D.W.; Hooper, S.J.; Thomas, D.W.; Costerton, J.W. A Review of the Scientific Evidence for Biofilms in Wounds. Wound Repair and Regeneration 2012, 20, 647–657. [Google Scholar] [CrossRef]
- Dryden, M.; Dickinson, A.; Brooks, J.; Hudgell, L.; Saeed, K.; Cutting, K.F. A Multi-Centre Clinical Evaluation of Reactive Oxygen Topical Wound Gel in 114 Wounds. J Wound Care 2016, 25, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Dryden, M.; Milward, G.; Saeed, K. Infection Prevention in Wounds with Surgihoney. Journal of Hospital Infection 2014, 88, 121–122. [Google Scholar] [CrossRef] [PubMed]
- Dryden, M.; Tawse, C.; Adams, J.; Howard, A.; Saeed, K.; Cooke, J. The Use of Surgihoney to Prevent or Eradicate Bacterial Colonisation in Dressing Oncology Long Vascular Lines. J Wound Care 2014, 23, 338–341. [Google Scholar] [CrossRef] [PubMed]
- Nair, H.K.R.; Tatavilis, N.; Pospíšilová, I.; Kučerová, J.; Cremers, N.A.J. Medical-Grade Honey Kills Antibiotic-Resistant Bacteria and Prevents Amputation in Diabetics with Infected Ulcers: A Prospective Case Series. Antibiotics 2020, 9, 529. [Google Scholar] [CrossRef] [PubMed]
- Hermanns, R.; Mateescu, C.; Thrasyvoulou, A.; Tananaki, C.; Wagener, F.A.D.T.G.; Cremers, N.A.J. Defining the Standards for Medical Grade Honey. J Apic Res 2020, 59, 125–135. [Google Scholar] [CrossRef]
- Kwakman, P.H.S.; Velde, A.A. te; Boer, L.; Speijer, D.; Christina Vandenbroucke-Grauls, M.J.; Zaat, S.A.J. How Honey Kills Bacteria. The FASEB Journal 2010, 24, 2576–2582. [Google Scholar] [CrossRef] [PubMed]
- Kwakman, P.H.S.; te Velde, A.A.; de Boer, L.; Vandenbroucke-Grauls, C.M.J.E.; Zaat, S.A.J. Two Major Medicinal Honeys Have Different Mechanisms of Bactericidal Activity. PLoS One 2011, 6, e17709. [Google Scholar] [CrossRef] [PubMed]
- Advancis Medical Activon® Manuka Honey. Available online: Https://Uk.Advancismedical.Com/Products/Activon-Manuka-Honey(accessed on 5 May 2024).
- Rafter, L.; Reynolds, T.; Collier, M.; Rafter, M.; West, M. A Clinical Evaluation of Algivon® Plus Manuka Honey Dressings for Chronic Wounds. Wounds UK ; 2017; Vol. 13, pp. 132–140;
- Edwards, J. A Prospective Evaluation of the Use of Honey Dressings to Manage Burn Wounds. Wounds UK ; 2013; Vol. 9, pp. 102–106;
- Cryer, S. The Use of Manuka Honey to Facilitate the Autoamputation of Fingertip Necrosis. Wounds UK ; 2016; Vol. 12, pp. 66–70;
- Welland Medical Aurum with Manuka Honey. Available online: Https://Wellandmedical.Com/Brand_type/Aurum/(accessed on 6 May 2024).
- Martin-Skurr, K. Case Study: Pyoderma Gangrenosum and the Effects of Manuka Honey. Available online: Https://Wellandmedical.Com/Wp-Content/Uploads/2019/02/Case-Study_NZ-Pyoderma-Gangrenosum-Manuka-Honey.Pdf(accessed on 6 May 2024).
- L-Mesitran. Products. Available online: Https://L-Mesitran.Com/Eu/En/Products(accessed on 6 May 2024).
- Boggust, A. Evaluation of L-Mesitran® Dressings in the Treatment of Minor Burns and Scalds at a Paediatric Emergency Department. Wounds UK 2013, 9, 114–117. [Google Scholar]
- Stephen-Haynes, J.; Callaghan, R. Properties of Honey: Its Mode of Action and Clinical Outcomes. Wounds UK 2011, 7, 50–57. [Google Scholar]
- Callaghan, R. Treating Difficult-to-Debride Wounds Using a Manuka Honey Dressing: A Case Study Evaluation. Wounds UK 2014, 10, 104–109. [Google Scholar]
- Bradbury, S.; Callaghan, R.; Ivins, N. ManukaDress Made Easy. Wounds UK 2014, 10, 1–6. [Google Scholar]
- Derma Science Europe Antibacterial Dressing Medihoney®: Antibacterial Medical Honey. Available online: Https://Media.Supplychain.Nhs.Uk/Media/Documents/ELY302/Marketing/48251_ELY302_3.Pdf.Pdf(accessed on 6 May 2024).
- Robson, V.; Dodd, S.; Thomas, S. Standardized Antibacterial Honey (MedihoneyTM) with Standard Therapy in Wound Care: Randomized Clinical Trial. J Adv Nurs 2009, 65, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.W.; van Eps, C.; Mudge, D.W.; Wiggins, K.J.; Armstrong, K.; Hawley, C.M.; Campbell, S.B.; Isbel, N.M.; Nimmo, G.R.; Gibbs, H. Randomized, Controlled Trial of Topical Exit-Site Application of Honey (Medihoney) versus Mupirocin for the Prevention of Catheter-Associated Infections in Hemodialysis Patients. Journal of the American Society of Nephrology 2005, 16, 1456–1462. [Google Scholar] [CrossRef] [PubMed]
- Kestrel Health Information MEDIHONEY® Calcium Alginate Dressing. Available online: Https://Www.Woundsource.Com/Product/Medihoney-Calcium-Alginate-Dressing(accessed on 6 May 2024).
- Jull, A.; Walker, N.; Parag, V.; Molan, P.; Rodgers, A. Randomized Clinical Trial of Honey-Impregnated Dressings for Venous Leg Ulcers. British Journal of Surgery 2008, 95, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Derma Sciences Europe Antibacterial Dressing Medihoney®: Barrier Cream. Available online: Https://Media.Supplychain.Nhs.Uk/Media/Documents/ELY289/Marketing/48248_ELY289.Pdf.Pdf(accessed on 6 May 2024).
- Nijhuis, W.; Houwing, R.; Van der Zwet, W.; Jansman, F. A Randomised Trial of Honey Barrier Cream versus Zinc Oxide Ointment. British Journal of Nursing 2012, 21, S10–S13. [Google Scholar] [CrossRef]
- Comvita Antibacterial Wound GelTM 25g. Available online: Https://Www.Comvita.Co.Uk/Product/Antibacterial-Wound-Gel%E2%84%A2-25g/6011(accessed on 6 May 2024).
- Robson, V.; Yorke, J.; Sen, R.A.; Lowe, D.; Rogers, S.N. Randomised Controlled Feasibility Trial on the Use of Medical Grade Honey Following Microvascular Free Tissue Transfer to Reduce the Incidence of Wound Infection. British Journal of Oral and Maxillofacial Surgery 2012, 50, 321–327. [Google Scholar] [CrossRef] [PubMed]
- Matoke Holdings Surgihoney, RO. Available Online:. Available online: Https://Www.Surgihoneyro.Com/(accessed on 6 May 2024).
- Dryden, M.; Goddard, C.; Madadi, A.; Heard, M.; Saeed, K.; Cooke, J. Using Antimicrobial Surgihoney to Prevent Caesarean Wound Infection. Br J Midwifery 2014, 22, 111–115. [Google Scholar] [CrossRef]
- Molan, P. Why Honey Is Effective as a Medicine. Bee World 2001, 82, 22–40. [Google Scholar] [CrossRef]
- Molan, P.C. The Antibacterial Activity of Honey. Bee World 1992, 73, 5–28. [Google Scholar] [CrossRef]
- Mandal, M.D.; Mandal, S. Honey: Its Medicinal Property and Antibacterial Activity. Asian Pac J Trop Biomed 2011, 1, 154–160. [Google Scholar] [CrossRef]
- Nolan, V.C.; Harrison, J.; Cox, J.A.G. Dissecting the Antimicrobial Composition of Honey. Antibiotics 2019, 8, 251. [Google Scholar] [CrossRef] [PubMed]
- Bucekova, M.; Valachova, I.; Kohutova, L.; Prochazka, E.; Klaudiny, J.; Majtan, J. Honeybee Glucose Oxidase—Its Expression in Honeybee Workers and Comparative Analyses of Its Content and H2O2-Mediated Antibacterial Activity in Natural Honeys. Naturwissenschaften 2014, 101, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Cebrero, G.; Sanhueza, O.; Pezoa, M.; Báez, M.E.; Martínez, J.; Báez, M.; Fuentes, E. Relationship among the Minor Constituents, Antibacterial Activity and Geographical Origin of Honey: A Multifactor Perspective. Food Chem 2020, 315, 126296. [Google Scholar] [CrossRef]
- Wong, C.M.; Wong, K.H.; Chen, X.D. Glucose Oxidase: Natural Occurrence, Function, Properties and Industrial Applications. Appl Microbiol Biotechnol 2008, 78, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Kuś, P.M.; Szweda, P.; Jerković, I.; Tuberoso, C.I.G. Activity of Polish Unifloral Honeys against Pathogenic Bacteria and Its Correlation with Colour, Phenolic Content, Antioxidant Capacity and Other Parameters. Lett Appl Microbiol 2016, 62, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Johnston, M.; McBride, M.; Dahiya, D.; Owusu-Apenten, R.; Singh Nigam, P. Antibacterial Activity of Manuka Honey and Its Components: An Overview. AIMS Microbiol 2018, 4, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Albaridi, N.A. Antibacterial Potency of Honey. Int J Microbiol 2019, 2019, 1–10. [Google Scholar] [CrossRef]
- Güneş, M.E.; Şahin, S.; Demir, C.; Borum, E.; Tosunoğlu, A. Determination of Phenolic Compounds Profile in Chestnut and Floral Honeys and Their Antioxidant and Antimicrobial Activities. J Food Biochem 2017, 41, e12345. [Google Scholar] [CrossRef]
- Ball, D.W. The Chemical Composition of Honey. J Chem Educ 2007, 84, 1643. [Google Scholar] [CrossRef]
- McLoone, P.; Warnock, M.; Fyfe, L. Honey: A Realistic Antimicrobial for Disorders of the Skin. Journal of Microbiology, Immunology and Infection 2016, 49, 161–167. [Google Scholar] [CrossRef]
- Ganz, T. Defensins: Antimicrobial Peptides of Innate Immunity. Nat Rev Immunol 2003, 3, 710–720. [Google Scholar] [CrossRef] [PubMed]
- Bucekova, M.; Sojka, M.; Valachova, I.; Martinotti, S.; Ranzato, E.; Szep, Z.; Majtan, V.; Klaudiny, J.; Majtan, J. Bee-Derived Antibacterial Peptide, Defensin-1, Promotes Wound Re-Epithelialisation in Vitro and in Vivo. Sci Rep 2017, 7, 7340. [Google Scholar] [CrossRef]
- Memar, M.Y.; Yekani, M.; Alizadeh, N.; Baghi, H.B. Hyperbaric Oxygen Therapy: Antimicrobial Mechanisms and Clinical Application for Infections. Biomedicine & Pharmacotherapy 2019, 109, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Çimşit, M.; Uzun, G.; Yıldız, Ş. Hyperbaric Oxygen Therapy as an Anti-Infective Agent. Expert Rev Anti Infect Ther 2009, 7, 1015–1026. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Fu, D.; Yan, L.; Xie, L. The Role of Hyperbaric Oxygen Therapy in the Treatment of Surgical Site Infections: A Narrative Review. Medicina (B Aires) 2023, 59, 762. [Google Scholar] [CrossRef] [PubMed]
- Chmelař, D.; Rozložník, M.; Hájek, M.; Pospíšilová, N.; Kuzma, J. Effect of Hyperbaric Oxygen on the Growth and Susceptibility of Facultatively Anaerobic Bacteria and Bacteria with Oxidative Metabolism to Selected Antibiotics. Folia Microbiol (Praha) 2024, 69, 101–108. [Google Scholar] [CrossRef]
- Kolpen, M.; Mousavi, N.; Sams, T.; Bjarnsholt, T.; Ciofu, O.; Moser, C.; Kühl, M.; Høiby, N.; Jensen, P.Ø. Reinforcement of the Bactericidal Effect of Ciprofloxacin on Pseudomonas Aeruginosa Biofilm by Hyperbaric Oxygen Treatment. Int J Antimicrob Agents 2016, 47, 163–167. [Google Scholar] [CrossRef]
- Kolpen, M.; Lerche, C.J.; Kragh, K.N.; Sams, T.; Koren, K.; Jensen, A.S.; Line, L.; Bjarnsholt, T.; Ciofu, O.; Moser, C.; et al. Hyperbaric Oxygen Sensitizes Anoxic Pseudomonas Aeruginosa Biofilm to Ciprofloxacin. Antimicrob Agents Chemother 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Weaver, L.K. Hyperbaric Oxygen Therapy for Carbon Monoxide Poisoning. Undersea Hyperb Med 2014, 41, 339–354. [Google Scholar]
- Ortega, M.A.; Fraile-Martinez, O.; García-Montero, C.; Callejón-Peláez, E.; Sáez, M.A.; Álvarez-Mon, M.A.; García-Honduvilla, N.; Monserrat, J.; Álvarez-Mon, M.; Bujan, J.; et al. A General Overview on the Hyperbaric Oxygen Therapy: Applications, Mechanisms and Translational Opportunities. Medicina (B Aires) 2021, 57, 864. [Google Scholar] [CrossRef]
- Harnanik, T.; Soeroso, J.; Suryokusumo, M.G.; Juliandhy, T. Effects of Hyperbaric Oxygen on T Helper 17/Regulatory T Polarization in Antigen and Collagen-Induced Arthritis: Hypoxia-Inducible Factor-1α as a Target. Oman Med J 2020, 35, e90–e90. [Google Scholar] [CrossRef] [PubMed]
- Baiula, M.; Greco, R.; Ferrazzano, L.; Caligiana, A.; Hoxha, K.; Bandini, D.; Longobardi, P.; Spampinato, S.; Tolomelli, A. Integrin-Mediated Adhesive Properties of Neutrophils Are Reduced by Hyperbaric Oxygen Therapy in Patients with Chronic Non-Healing Wound. PLoS One 2020, 15, e0237746. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yi, H.; Kato, M.; Suzuki, H.; Kobayashi, S.; Takahashi, H.; Nakashima, I. Differential Sensitivities to Hyperbaric Oxygen of Lymphocyte Subpopulations of Normal and Autoimmune Mice. Immunol Lett 1997, 59, 79–84. [Google Scholar] [CrossRef] [PubMed]
- SAITO, K.; TANAKA, Y.; OTA, T.; ETO, S.; YAMASHITA, U. Suppressive Effect of Hyperbaric Oxygenation on Immune Responses of Normal and Autoimmune Mice. Clin Exp Immunol 2008, 86, 322–327. [Google Scholar] [CrossRef] [PubMed]
- Schottlender, N.; Gottfried, I.; Ashery, U. Hyperbaric Oxygen Treatment: Effects on Mitochondrial Function and Oxidative Stress. Biomolecules 2021, 11, 1827. [Google Scholar] [CrossRef] [PubMed]
- Růžička, J.; Dejmek, J.; Bolek, L.; Beneš, J.; Kuncová, J. Hyperbaric Oxygen Influences Chronic Wound Healing - a Cellular Level Review. Physiol Res 2021, 70, S261–S273. [Google Scholar] [CrossRef] [PubMed]
- Baethge, C.; Goldbeck-Wood, S.; Mertens, S. SANRA—a Scale for the Quality Assessment of Narrative Review Articles. Res Integr Peer Rev 2019, 4, 5. [Google Scholar] [CrossRef] [PubMed]
- Lerche, C.J.; Schwartz, F.; Pries-Heje, M.M.; Fosbøl, E.L.; Iversen, K.; Jensen, P.Ø.; Høiby, N.; Hyldegaard, O.; Bundgaard, H.; Moser, C. Potential Advances of Adjunctive Hyperbaric Oxygen Therapy in Infective Endocarditis. Front Cell Infect Microbiol 2022, 12. [Google Scholar] [CrossRef]
- Löndahl, M.; Boulton, A.J.M. Hyperbaric Oxygen Therapy in Diabetic Foot Ulceration: Useless or Useful? A Battle. Diabetes Metab Res Rev 2020, 36. [Google Scholar] [CrossRef]
- Brouwer, R.J.; Lalieu, R.C.; Hoencamp, R.; van Hulst, R.A.; Ubbink, D.T. A Systematic Review and Meta-Analysis of Hyperbaric Oxygen Therapy for Diabetic Foot Ulcers with Arterial Insufficiency. J Vasc Surg 2020, 71, 682–692. [Google Scholar] [CrossRef]
- Lalieu, R.C.; Brouwer, R.J.; Ubbink, D.T.; Hoencamp, R.; Bol Raap, R.; van Hulst, R.A. Hyperbaric Oxygen Therapy for Nonischemic Diabetic Ulcers: A Systematic Review. Wound Repair and Regeneration 2020, 28, 266–275. [Google Scholar] [CrossRef] [PubMed]
- Lerche, C.J.; Christophersen, L.J.; Kolpen, M.; Nielsen, P.R.; Trøstrup, H.; Thomsen, K.; Hyldegaard, O.; Bundgaard, H.; Jensen, P.Ø.; Høiby, N.; et al. Hyperbaric Oxygen Therapy Augments Tobramycin Efficacy in Experimental Staphylococcus Aureus Endocarditis. Int J Antimicrob Agents 2017, 50, 406–412. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-E.; Ko, J.-Y.; Fong, C.-Y.; Juhn, R.-J. Treatment of Diabetic Foot Infection with Hyperbaric Oxygen Therapy. Foot and Ankle Surgery 2010, 16, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.J.; Chen, T.X.; Wald, K.J.; Tooley, A.A.; Lisman, R.D.; Chiu, E.S. Hyperbaric Oxygen Therapy in Ophthalmic Practice: An Expert Opinion. Expert Rev Ophthalmol 2020, 15, 119–126. [Google Scholar] [CrossRef]
- Oguz, H.; Sobaci, G. The Use of Hyperbaric Oxygen Therapy in Ophthalmology. Surv Ophthalmol 2008, 53, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, F.; Khalatbary, A. A Review on the Neuroprotective Effects of Hyperbaric Oxygen Therapy. Med Gas Res 2021, 11, 72. [Google Scholar] [CrossRef] [PubMed]
- Moen, I.; Stuhr, L.E.B. Hyperbaric Oxygen Therapy and Cancer—a Review. Target Oncol 2012, 7, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Borab, Z.; Mirmanesh, M.D.; Gantz, M.; Cusano, A.; Pu, L.L.Q. Systematic Review of Hyperbaric Oxygen Therapy for the Treatment of Radiation-Induced Skin Necrosis. Journal of Plastic, Reconstructive & Aesthetic Surgery 2017, 70, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Song, Z.; Zhou, Y.; Pan, S.; Wang, F.; Guo, Z.; Jiang, M.; Wang, G.; Kong, R.; Sun, B. The Apoptosis of Peripheral Blood Lymphocytes Promoted by Hyperbaric Oxygen Treatment Contributes to Attenuate the Severity of Early Stage Acute Pancreatitis in Rats. Apoptosis 2014, 19, 58–75. [Google Scholar] [CrossRef]
- Godman, C.A.; Chheda, K.P.; Hightower, L.E.; Perdrizet, G.; Shin, D.-G.; Giardina, C. Hyperbaric Oxygen Induces a Cytoprotective and Angiogenic Response in Human Microvascular Endothelial Cells. Cell Stress Chaperones 2010, 15, 431–442. [Google Scholar] [CrossRef]
- Fielden, M.P.; Martinovic, E.; Ells, A.L. Hyperbaric Oxygen Therapy in the Treatment of Orbital Gas Gangrene. Journal of American Association for Pediatric Ophthalmology and Strabismus 2002, 6, 252–254. [Google Scholar] [CrossRef] [PubMed]
- Bumah, V.V.; Whelan, H.T.; Masson-Meyers, D.S.; Quirk, B.; Buchmann, E.; Enwemeka, C.S. The Bactericidal Effect of 470-Nm Light and Hyperbaric Oxygen on Methicillin-Resistant Staphylococcus Aureus (MRSA). Lasers Med Sci 2015, 30, 1153–1159. [Google Scholar] [CrossRef] [PubMed]
- Mogami, H.; Hayakawa, T.; Kanai, N.; Kuroda, R.; Yamada, R.; Ikeda, T.; Katsurada, K.; Sugimoto, T. Clinical Application of Hyperbaric Oxygenation in the Treatment of Acute Cerebral Damage. J Neurosurg 1969, 31, 636–643. [Google Scholar] [CrossRef]
- Edwards, M.L. Hyperbaric Oxygen Therapy. Part 2: Application in Disease. Journal of Veterinary Emergency and Critical Care 2010, 20, 289–297. [Google Scholar] [CrossRef]
- Wang, R.-Y.; Yang, Y.-R.; Chang, H.-C. The SDF1-CXCR4 Axis Is Involved in the Hyperbaric Oxygen Therapy-Mediated Neuronal Cells Migration in Transient Brain Ischemic Rats. Int J Mol Sci 2022, 23, 1780. [Google Scholar] [CrossRef] [PubMed]
- Helms, A.K.; Whelan, H.T.; Torbey, M.T. Hyperbaric Oxygen Therapy of Cerebral Ischemia. Cerebrovascular Diseases 2005, 20, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Matchett, G.A.; Martin, R.D.; Zhang, J.H. Hyperbaric Oxygen Therapy and Cerebral Ischemia: Neuroprotective Mechanisms. Neurol Res 2009, 31, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Cevolani, D.; Di Donato, F.; Santarella, L.; Bertossi, S.; Cellerini, M. Functional MRI (FMRI) Evaluation of Hyperbaric Oxygen Therapy (HBOT) Efficacy in Chronic Cerebral Stroke: A Small Retrospective Consecutive Case Series. Int J Environ Res Public Health 2020, 18, 190. [Google Scholar] [CrossRef] [PubMed]
- Thiankhaw, K.; Chattipakorn, N.; Chattipakorn, S.C. The Effects of Hyperbaric Oxygen Therapy on the Brain with Middle Cerebral Artery Occlusion. J Cell Physiol 2021, 236, 1677–1694. [Google Scholar] [CrossRef]
- Chiang, I.-H.; Chen, S.-G.; Huang, K.-L.; Chou, Y.-C.; Dai, N.-T.; Peng, C.-K. Adjunctive Hyperbaric Oxygen Therapy in Severe Burns: Experience in Taiwan Formosa Water Park Dust Explosion Disaster. Burns 2017, 43, 852–857. [Google Scholar] [CrossRef]
- M. Misiuga; J. Glik; M. Kawecki; I. Dziurzyńska; M. Ples; W. Łabuś; M. Nowak The Effect of Hyperbaric Oxygen Therapy on Burn Wounds Covered With Skin Allografts. JOTSRR 2016, 1, 17–27. [Google Scholar]
- Bartek, J.; Jakola, A.S.; Skyrman, S.; Förander, P.; Alpkvist, P.; Schechtmann, G.; Glimåker, M.; Larsson, A.; Lind, F.; Mathiesen, T. Hyperbaric Oxygen Therapy in Spontaneous Brain Abscess Patients: A Population-Based Comparative Cohort Study. Acta Neurochir (Wien) 2016, 158, 1259–1267. [Google Scholar] [CrossRef] [PubMed]
- Inanmaz, M.E.; Kose, K.C.; Isik, C.; Atmaca, H.; Basar, H. Can Hyperbaric Oxygen Be Used to Prevent Deep Infections in Neuro-Muscular Scoliosis Surgery? BMC Surg 2014, 14, 85. [Google Scholar] [CrossRef] [PubMed]
- Barili, F.; Polvani, G.; Topkara, V.K.; Dainese, L.; Cheema, F.H.; Roberto, M.; Naliato, M.; Parolari, A.; Alamanni, F.; Biglioli, P. Role of Hyperbaric Oxygen Therapy in the Treatment of Postoperative Organ/Space Sternal Surgical Site Infections. World J Surg 2007, 31, 1702–1706. [Google Scholar] [CrossRef]
- Larsson, A.; Uusijärvi, J.; Lind, F.; Gustavsson, B.; Saraste, H. Hyperbaric Oxygen in the Treatment of Postoperative Infections in Paediatric Patients with Neuromuscular Spine Deformity. European Spine Journal 2011, 20, 2217–2222. [Google Scholar] [CrossRef]
- George, M.E.; Rueth, N.M.; Skarda, D.E.; Chipman, J.G.; Quickel, R.R.; Beilman, G.J. Hyperbaric Oxygen Does Not Improve Outcome in Patients with Necrotizing Soft Tissue Infection. Surg Infect (Larchmt) 2009, 10, 21–28. [Google Scholar] [CrossRef]
- Wilkinson, D. Hyperbaric Oxygen Treatment and Survival From Necrotizing Soft Tissue Infection. Archives of Surgery 2004, 139, 1339. [Google Scholar] [CrossRef] [PubMed]
- Massey, P.R.; Sakran, J. V.; Mills, A.M.; Sarani, B.; Aufhauser, D.D.; Sims, C.A.; Pascual, J.L.; Kelz, R.R.; Holena, D.N. Hyperbaric Oxygen Therapy in Necrotizing Soft Tissue Infections. Journal of Surgical Research 2012, 177, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Shupak, A.; Oren, S.; Goldenberg, I.; Barzilai, A.; Moskuna, R.; Bursztein, S. Necrotizing Fasciitis: An Indication for Hyperbaric Oxygenation Therapy? Surgery 1995, 118, 873–878. [Google Scholar] [CrossRef]
- Duzgun, A.P.; Satır, H.Z.; Ozozan, O.; Saylam, B.; Kulah, B.; Coskun, F. Effect of Hyperbaric Oxygen Therapy on Healing of Diabetic Foot Ulcers. The Journal of Foot and Ankle Surgery 2008, 47, 515–519. [Google Scholar] [CrossRef]
- Löndahl, M.; Katzman, P.; Nilsson, A.; Hammarlund, C. Hyperbaric Oxygen Therapy Facilitates Healing of Chronic Foot Ulcers in Patients With Diabetes. Diabetes Care 2010, 33, 998–1003. [Google Scholar] [CrossRef] [PubMed]
- Faglia, E.; Favales, F.; Aldeghi, A.; Calia, P.; Quarantiello, A.; Oriani, G.; Michael, M.; Campagnoli, P.; Morabito, A. Adjunctive Systemic Hyperbaric Oxygen Therapy in Treatment of Severe Prevalently Ischemic Diabetic Foot Ulcer: A Randomized Study. Diabetes Care 1996, 19, 1338–1343. [Google Scholar] [CrossRef] [PubMed]
- Kessler, L.; Bilbault, P.; Ortéga, F.; Grasso, C.; Passemard, R.; Stephan, D.; Pinget, M.; Schneider, F. Hyperbaric Oxygenation Accelerates the Healing Rate of Nonischemic Chronic Diabetic Foot Ulcers. Diabetes Care 2003, 26, 2378–2382. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Li, P.; Shi, Z.; Hou, T.; Chen, X.; Du, J. A Prospective, Randomized, Controlled Study of Hyperbaric Oxygen Therapy: Effects on Healing and Oxidative Stress of Ulcer Tissue in Patients with a Diabetic Foot Ulcer. Ostomy Wound Manage 2013, 59, 18–24. [Google Scholar] [PubMed]
- Kalani, M.; Jörneskog, G.; Naderi, N.; Lind, F.; Brismar, K. Hyperbaric Oxygen (HBO) Therapy in Treatment of Diabetic Foot Ulcers. J Diabetes Complications 2002, 16, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Abidia, A.; Laden, G.; Kuhan, G.; Johnson, B.F.; Wilkinson, A.R.; Renwick, P.M.; Masson, E.A.; McCollum, P.T. The Role of Hyperbaric Oxygen Therapy in Ischaemic Diabetic Lower Extremity Ulcers: A Double-Blind Randomised-Controlled Trial. European Journal of Vascular and Endovascular Surgery 2003, 25, 513–518. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, R.; Severson, M.A.; Traynelis, V.C. Role of Hyperbaric Oxygen Therapy in the Treatment of Bacterial Spinal Osteomyelitis. J Neurosurg Spine 2009, 10, 16–20. [Google Scholar] [CrossRef]
- Chen, C.-E.; Shih, S.-T.; Fu, T.-H.; Wang, J.-W.; Wang, C.-J. Hyperbaric Oxygen Therapy in the Treatment of Chronic Refractory Osteomyelitis: A Preliminary Report. Chang Gung Med J 2003, 26, 114–121. [Google Scholar]
- Delasotta, L.A.; Hanflik, A.; Bicking, G.; Mannella, W.J. Hyperbaric Oxygen for Osteomyelitis in a Compromised Host. Open Orthop J 2013, 7, 114–117. [Google Scholar] [CrossRef]
- Yu, W.-K.; Chen, Y.-W.; Shie, H.-G.; Lien, T.-C.; Kao, H.-K.; Wang, J.-H. Hyperbaric Oxygen Therapy as an Adjunctive Treatment for Sternal Infection and Osteomyelitis after Sternotomy and Cardiothoracic Surgery. J Cardiothorac Surg 2011, 6, 141. [Google Scholar] [CrossRef]
- Petzold, T.; Feindt, P.R.; Carl, U.M.; Gams, E. Hyperbaric Oxygen Therapy in Deep Sternal Wound Infection After Heart Transplantation. Chest 1999, 115, 1455–1458. [Google Scholar] [CrossRef] [PubMed]
- Siondalski, P.; Keita, L.; Sićko, Z.; Zelechowski, P.; Jaworski, Ł.; Rogowski, J. [Surgical Treatment and Adjunct Hyperbaric Therapy to Improve Healing of Wound Infection Complications after Sterno-Mediastinitis]. Pneumonol Alergol Pol 2003, 71, 12–16. [Google Scholar] [PubMed]
- Sun, I.; Lee, S.; Chiu, C.; Lin, S.; Lai, C. Hyperbaric Oxygen Therapy with Topical Negative Pressure: An Alternative Treatment for the Refractory Sternal Wound Infection. J Card Surg 2008, 23, 677–680. [Google Scholar] [CrossRef] [PubMed]
- Larsson, A.; Uusijärvi, J.; Lind, F.; Gustavsson, B.; Saraste, H. Hyperbaric Oxygen in the Treatment of Postoperative Infections in Paediatric Patients with Neuromuscular Spine Deformity. European Spine Journal 2011, 20, 2217–2222. [Google Scholar] [CrossRef] [PubMed]
- Egito, J.G.T. do; Abboud, C.S.; Oliveira, A.P.V. de; Máximo, C.A.G.; Montenegro, C.M.; Amato, V.L.; Bammann, R.; Farsky, P.S. Evolução Clínica de Pacientes Com Mediastinite Pós-Cirurgia de Revascularização Miocárdica Submetidos à Oxigenoterapia Hiperbárica Como Terapia Adjuvante. Einstein (São Paulo) 2013, 11, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Litwinowicz, R.; Bryndza, M.; Chrapusta, A.; Kobielska, E.; Kapelak, B.; Grudzień, G. Hyperbaric Oxygen Therapy as Additional Treatment in Deep Sternal Wound Infections – a Single Center’s Experience. Polish Journal of Cardio-Thoracic Surgery 2016, 3, 198–202. [Google Scholar] [CrossRef]
- Bartek Jr., J.; Skyrman, S.; Nekludov, M.; Mathiesen, T.; Lind, F.; Schechtmann, G. Hyperbaric Oxygen Therapy as Adjuvant Treatment for Hardware-Related Infections in Neuromodulation. Stereotact Funct Neurosurg 2018, 96, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Copeland, H.; Newcombe, J.; Yamin, F.; Bhajri, K.; Mille, V.A.; Hasaniya, N.; Bailey, L.; Razzouk, A.J. Role of Negative Pressure Wound Care and Hyperbaric Oxygen Therapy for Sternal Wound Infections After Pediatric Cardiac Surgery. World J Pediatr Congenit Heart Surg 2018, 9, 440–445. [Google Scholar] [CrossRef]
- Stizzo, M.; Manfredi, C.; Spirito, L.; Sciorio, C.; Romero Otero, J.; Martinez Salamanca, J.I.; Crocetto, F.; Verze, P.; Imbimbo, C.; Fusco, F.; et al. Hyperbaric Oxygen Therapy as Adjuvant Treatment for Surgical Site Infections after Male-to-female Gender Affirmation Surgery: A 10-year Experience. Andrology 2022, 10, 1310–1316. [Google Scholar] [CrossRef]
- Pan, B.; Li, H.; Lang, D.; Xing, B. Environmentally Persistent Free Radicals: Occurrence, Formation Mechanisms and Implications. Environmental Pollution 2019, 248, 320–331. [Google Scholar] [CrossRef]
- Vinayak, A.; Mudgal, G.; Singh, G.B. Environment Persistent Free Radicals: Long-Lived Particles. In; 2021; pp. 1–19.
- Saravia, J.; Lee, G.I.; Lomnicki, S.; Dellinger, B.; Cormier, S.A. Particulate Matter Containing Environmentally Persistent Free Radicals and Adverse Infant Respiratory Health Effects: A Review. J Biochem Mol Toxicol 2013, 27, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Yao, D.; Qian, Y.; Zhong, S.; Zhang, L.; Xue, G.; Jia, H. Factors Controlling the Formation of Persistent Free Radicals in Hydrochar during Hydrothermal Conversion of Rice Straw. Environ Chem Lett 2018, 16, 1463–1468. [Google Scholar] [CrossRef]
- Xu, Y.; Yang, L.; Wang, X.; Zheng, M.; Li, C.; Zhang, A.; Fu, J.; Yang, Y.; Qin, L.; Liu, X.; et al. Risk Evaluation of Environmentally Persistent Free Radicals in Airborne Particulate Matter and Influence of Atmospheric Factors. Ecotoxicol Environ Saf 2020, 196, 110571. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Sun, P.; Faye, M.C.A.S.; Zhang, Y. Characterization of Biochar Derived from Rice Husks and Its Potential in Chlorobenzene Degradation. Carbon N Y 2018, 130, 730–740. [Google Scholar] [CrossRef]
- Kumbhar, D.; Palliyarayil, A.; Reghu, D.; Shrungar, D.; Umapathy, S.; Sil, S. Rapid Discrimination of Porous Bio-Carbon Derived from Nitrogen Rich Biomass Using Raman Spectroscopy and Artificial Intelligence Methods. Carbon N Y 2021, 178, 792–802. [Google Scholar] [CrossRef]
- Wu, C.; Fu, L.; Li, H.; Liu, X.; Wan, C. Using Biochar to Strengthen the Removal of Antibiotic Resistance Genes: Performance and Mechanism. Science of The Total Environment 2022, 816, 151554. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Qin, F.; Zhang, C.; Huang, D.; Tang, L.; Yan, M.; Wang, W.; Song, B.; Qin, D.; Zhou, Y.; et al. Effects of Heterogeneous Metals on the Generation of Persistent Free Radicals as Critical Redox Sites in Iron-Containing Biochar for Persulfate Activation. ACS ES&T Water 2023, 3, 298–310. [Google Scholar] [CrossRef]
- Zhou, B.; Liu, Q.; Shi, L.; Liu, Z. Electron Spin Resonance Studies of Coals and Coal Conversion Processes: A Review. Fuel Processing Technology 2019, 188, 212–227. [Google Scholar] [CrossRef]
- Ruan, X.; Sun, Y.; Du, W.; Tang, Y.; Liu, Q.; Zhang, Z.; Doherty, W.; Frost, R.L.; Qian, G.; Tsang, D.C.W. Formation, Characteristics, and Applications of Environmentally Persistent Free Radicals in Biochars: A Review. Bioresour Technol 2019, 281, 457–468. [Google Scholar] [CrossRef]
- Jothirani, R.; Kumar, P.S.; Saravanan, A.; Narayan, A.S.; Dutta, A. Ultrasonic Modified Corn Pith for the Sequestration of Dye from Aqueous Solution. Journal of Industrial and Engineering Chemistry 2016, 39, 162–175. [Google Scholar] [CrossRef]
- S. , S.; P., S.K.; A., S.; P., S.R.; C., R. Computation of Adsorption Parameters for the Removal of Dye from Wastewater by Microwave Assisted Sawdust: Theoretical and Experimental Analysis. Environ Toxicol Pharmacol 2017, 50, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, A.; Kumar, P.S.; Renita, A.A. Hybrid Synthesis of Novel Material through Acid Modification Followed Ultrasonication to Improve Adsorption Capacity for Zinc Removal. J Clean Prod 2018, 172, 92–105. [Google Scholar] [CrossRef]
- Luo, Z.; Yao, B.; Yang, X.; Wang, L.; Xu, Z.; Yan, X.; Tian, L.; Zhou, H.; Zhou, Y. Novel Insights into the Adsorption of Organic Contaminants by Biochar: A Review. Chemosphere 2022, 287, 132113. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Tang, L.; Jiang, H.; Mao, Y.; Song, W.; Chen, H. Prediction of Heavy Metals Adsorption by Hydrochars and Identification of Critical Factors Using Machine Learning Algorithms. Bioresour Technol 2023, 383, 129223. [Google Scholar] [CrossRef]
- Chauhan, S.; Shafi, T.; Dubey, B.K.; Chowdhury, S. Biochar-Mediated Removal of Pharmaceutical Compounds from Aqueous Matrices via Adsorption. Waste Dispos Sustain Energy 2023, 5, 37–62. [Google Scholar] [CrossRef]
Figure 1.
Schematic pathways of reactive oxygen species (ROS) production and their main effects on biological systems. Nrf2 = erythroid nuclear transcription factor-2; NF-kB = transcription factor involved in cellular responses to stimuli such as stress, cytokines, free radicals, heavy metals, ultraviolet irradiation, oxidized low density lipoproteins (LDL) etc. Reproduced from our article [11].
Figure 1.
Schematic pathways of reactive oxygen species (ROS) production and their main effects on biological systems. Nrf2 = erythroid nuclear transcription factor-2; NF-kB = transcription factor involved in cellular responses to stimuli such as stress, cytokines, free radicals, heavy metals, ultraviolet irradiation, oxidized low density lipoproteins (LDL) etc. Reproduced from our article [11].
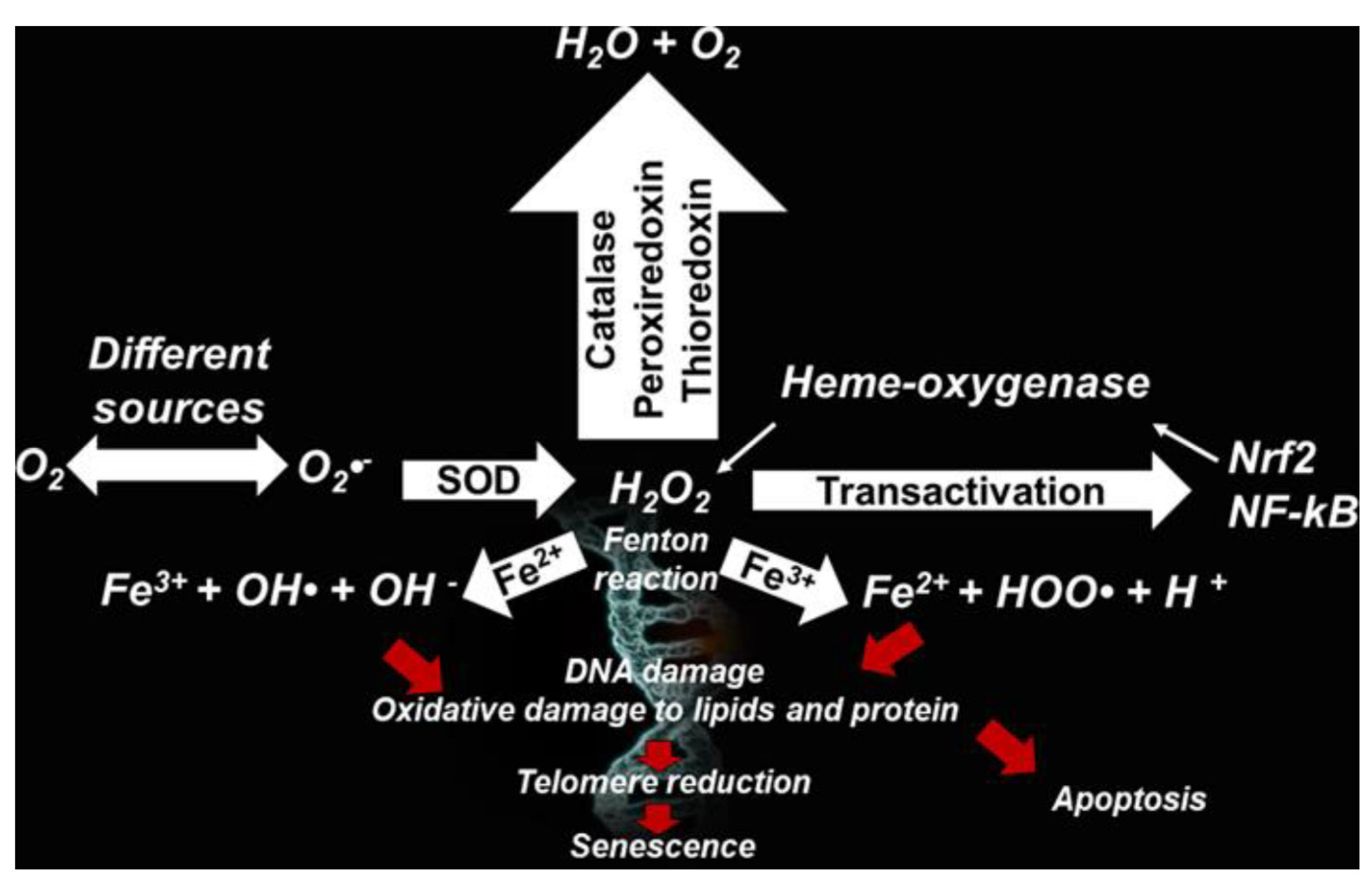
Figure 2.
ROS induction by antibiotics.
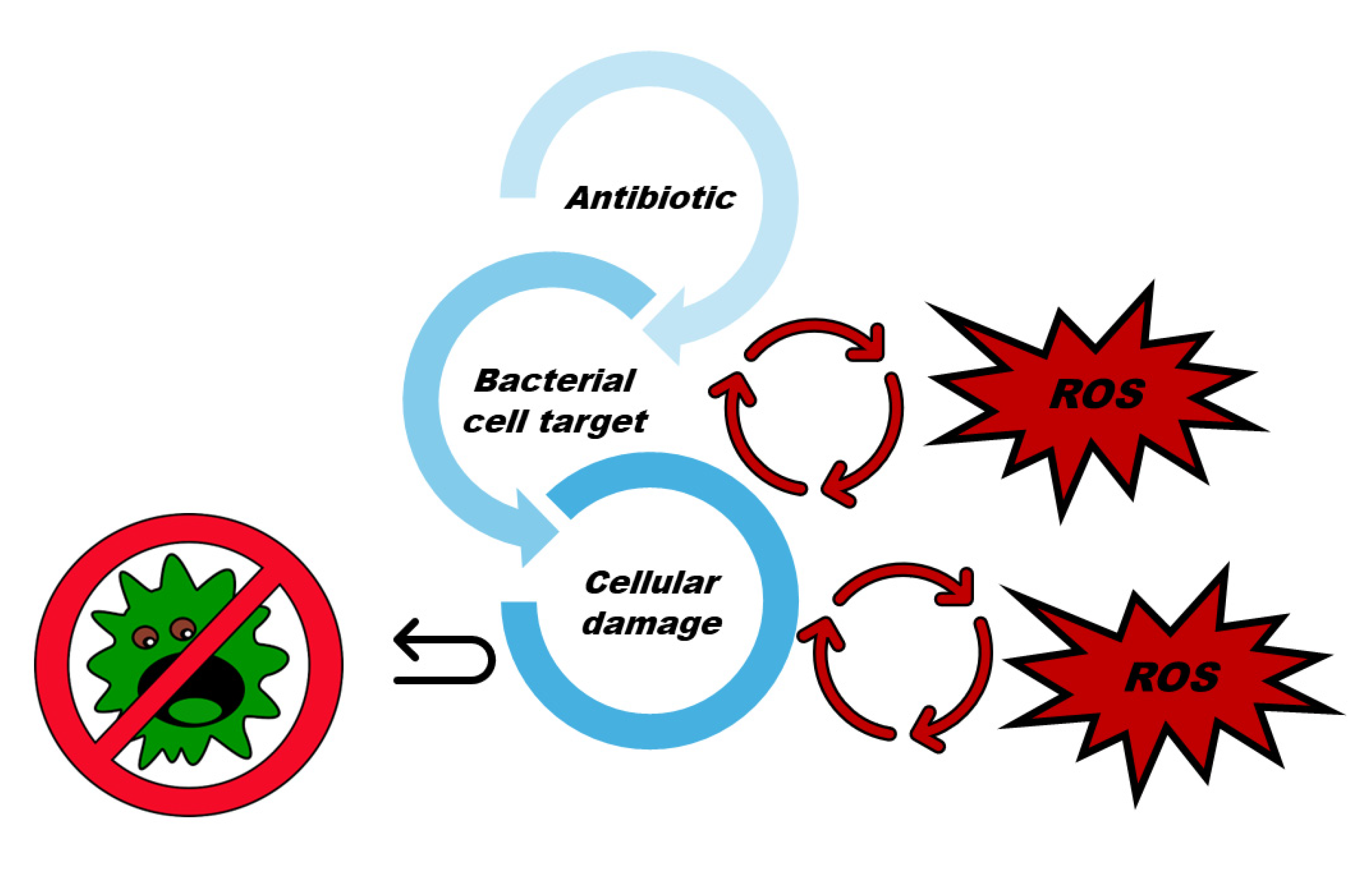
Figure 3.
Jablonski diagram showing the photochemical and photophysical mechanisms of antimicrobial photodynamic therapy (PDT). S0: ground singlet state of the PS molecule; Sn: excited singlet state of the PS molecule; T1: triplet excited state of the PS molecule; A: absorption of light; F: fluorescence emission; H: heat generation (internal conversion); ISC: inter-system crossing; P: phosphorescence emission; 3O2: ground state oxygen; 1O2: singlet oxygen; O2−•: superoxide anion; HO•: hydroxyl radical; H2O2: hydrogen peroxide. The image is an adaptation from an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/Licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium[80].
Figure 3.
Jablonski diagram showing the photochemical and photophysical mechanisms of antimicrobial photodynamic therapy (PDT). S0: ground singlet state of the PS molecule; Sn: excited singlet state of the PS molecule; T1: triplet excited state of the PS molecule; A: absorption of light; F: fluorescence emission; H: heat generation (internal conversion); ISC: inter-system crossing; P: phosphorescence emission; 3O2: ground state oxygen; 1O2: singlet oxygen; O2−•: superoxide anion; HO•: hydroxyl radical; H2O2: hydrogen peroxide. The image is an adaptation from an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/Licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium[80].
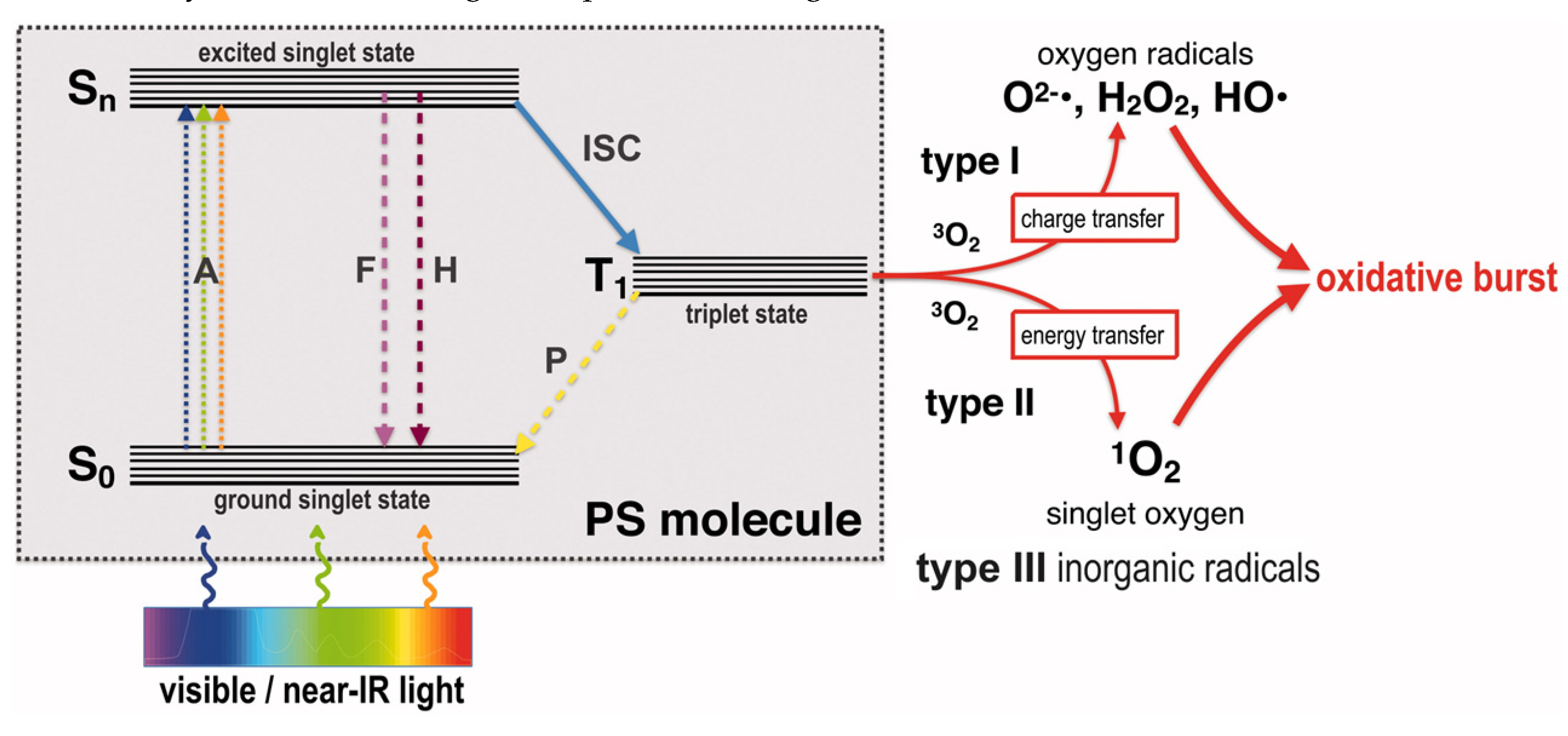
Figure 4.
Characteristic and criteria that a medical grade honey (MGH) should fulfil according to Hermann et al [187].
Figure 4.
Characteristic and criteria that a medical grade honey (MGH) should fulfil according to Hermann et al [187].

Figure 5.
HBOT enhances the immune system’s antimicrobial effects: Increased O2 levels during HBOT have a variety of biological effects, including suppression of proinflammatory mediators, transitory reduction in the CD4:CD8 T cell ratio, and stimulation of lymphocyte and neutrophil death through caspase-3-, caspase-7-, and caspase-9-dependent mechanisms. In general, these effects can boost the antibacterial processes of the immune system and infection recovery. Abbreviations: ROS, reactive oxygen species; IL, interleukin; INF, interferon; TNF, tumor necrosis factor; CAS, caspase; NO, nitric oxide. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/) [229].
Figure 5.
HBOT enhances the immune system’s antimicrobial effects: Increased O2 levels during HBOT have a variety of biological effects, including suppression of proinflammatory mediators, transitory reduction in the CD4:CD8 T cell ratio, and stimulation of lymphocyte and neutrophil death through caspase-3-, caspase-7-, and caspase-9-dependent mechanisms. In general, these effects can boost the antibacterial processes of the immune system and infection recovery. Abbreviations: ROS, reactive oxygen species; IL, interleukin; INF, interferon; TNF, tumor necrosis factor; CAS, caspase; NO, nitric oxide. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/) [229].
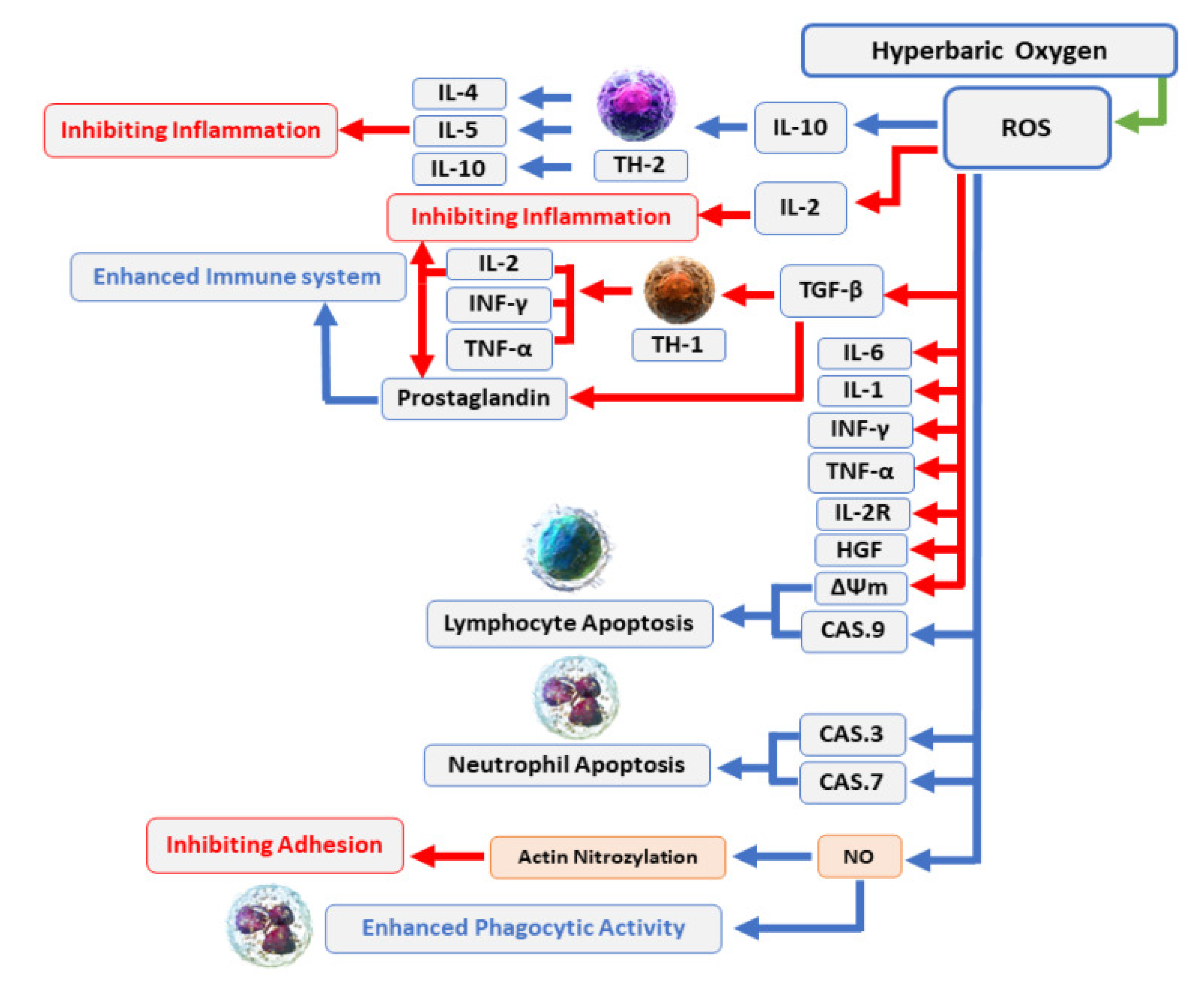
Figure 6.
Events caused by hyperbaric oxygen therapy by which its antibacterial effects derive.
Figure 7.
Number of publications on BCs-derived PFRs from 2014 according to Scopus dataset (reviews and chapters in books included). The survey used the following keywords: persistent AND free AND radicals AND biochar [24].
Figure 7.
Number of publications on BCs-derived PFRs from 2014 according to Scopus dataset (reviews and chapters in books included). The survey used the following keywords: persistent AND free AND radicals AND biochar [24].
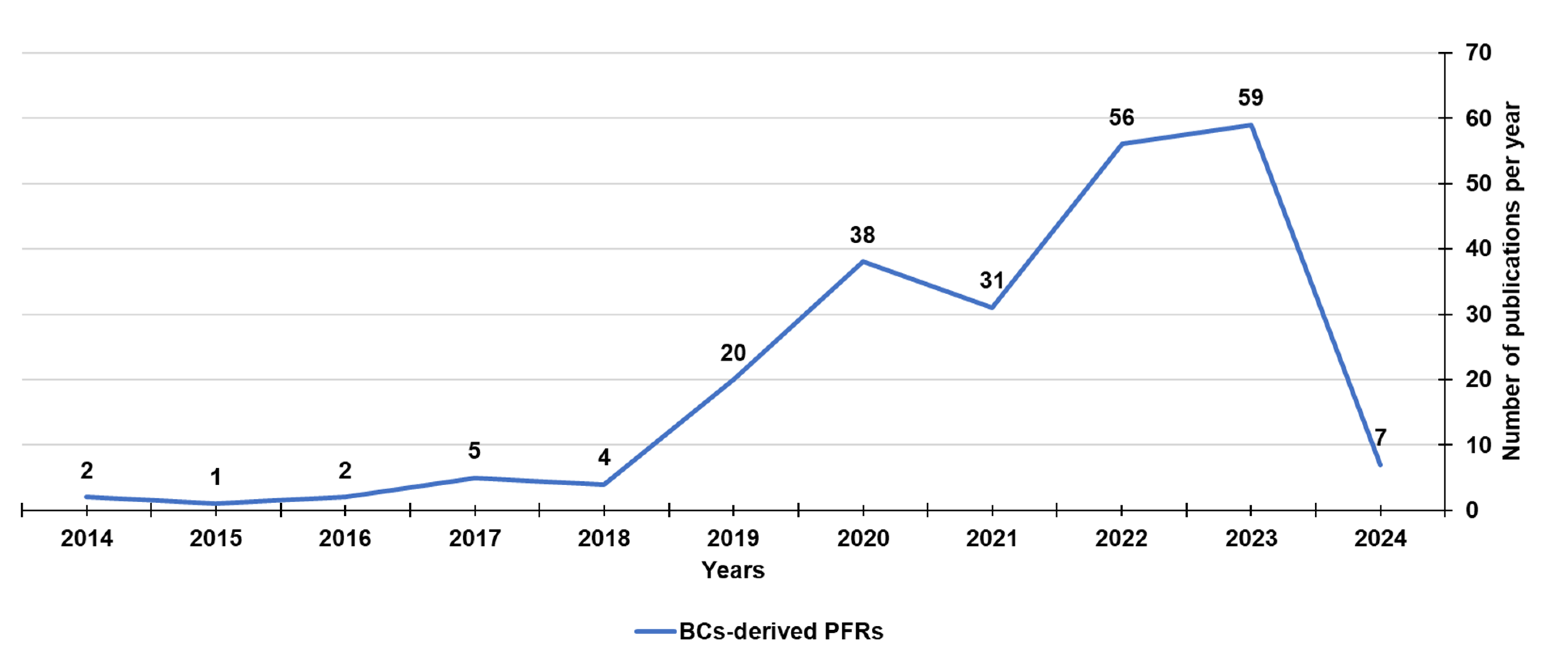
Scheme 1.
Possible mechanisms leading to the formation of BC-bounded PFRs from lignin. The orange sphere represents biomass, while the black sphere represents BC whose hypothetic structures depending on pyrolysis condition have been shown at the bottom of the Scheme [24].
Scheme 1.
Possible mechanisms leading to the formation of BC-bounded PFRs from lignin. The orange sphere represents biomass, while the black sphere represents BC whose hypothetic structures depending on pyrolysis condition have been shown at the bottom of the Scheme [24].
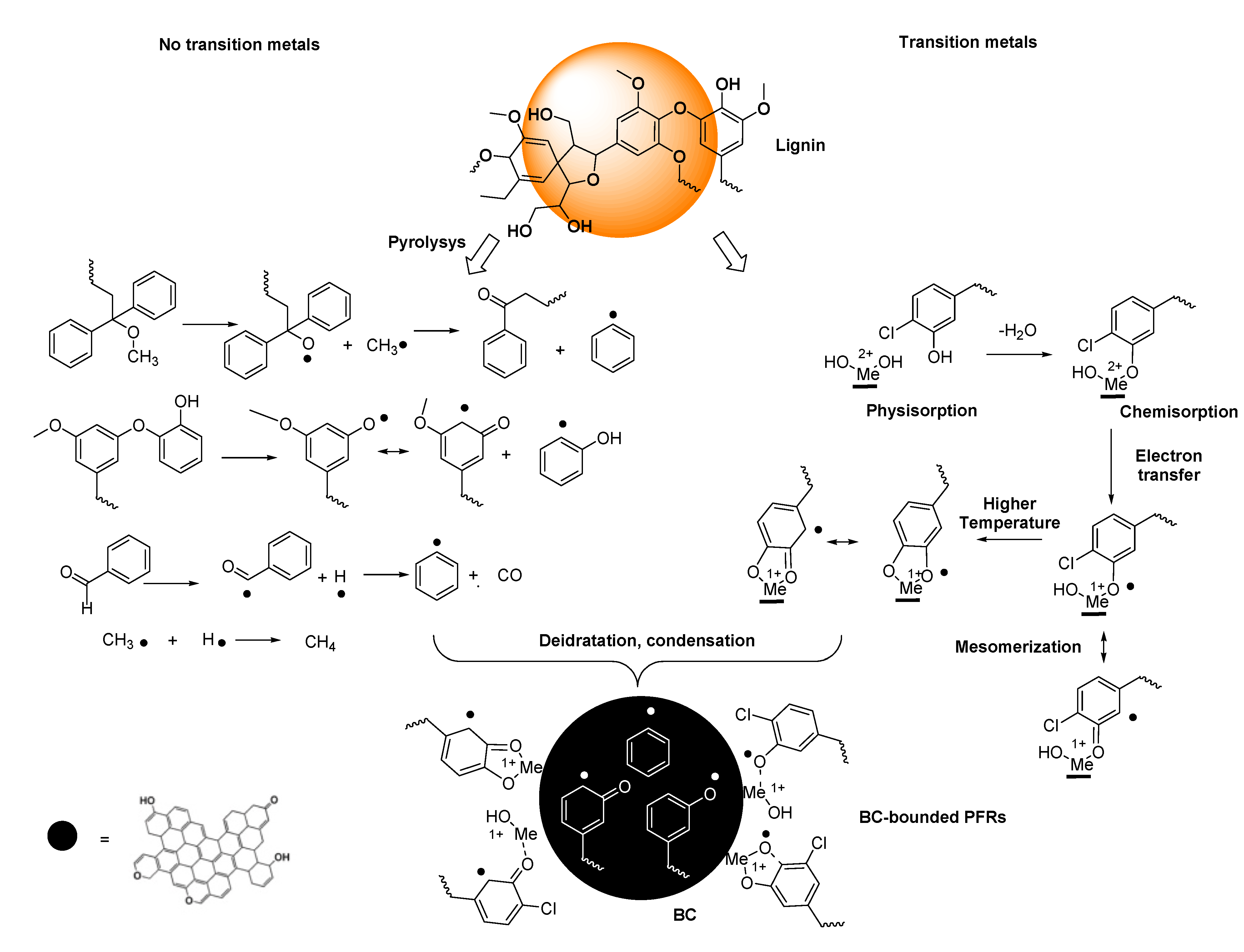
Scheme 2.
Possible mechanisms leading to the formation of BC-bounded graphitic PFRs from cellulose (left side) and emicellulose (right side). The orange sphere represents biomass, while the black sphere represents BC whose hypothetic structures depending on pyrolysis condition have been shown at the bottom of the Scheme [24].
Scheme 2.
Possible mechanisms leading to the formation of BC-bounded graphitic PFRs from cellulose (left side) and emicellulose (right side). The orange sphere represents biomass, while the black sphere represents BC whose hypothetic structures depending on pyrolysis condition have been shown at the bottom of the Scheme [24].
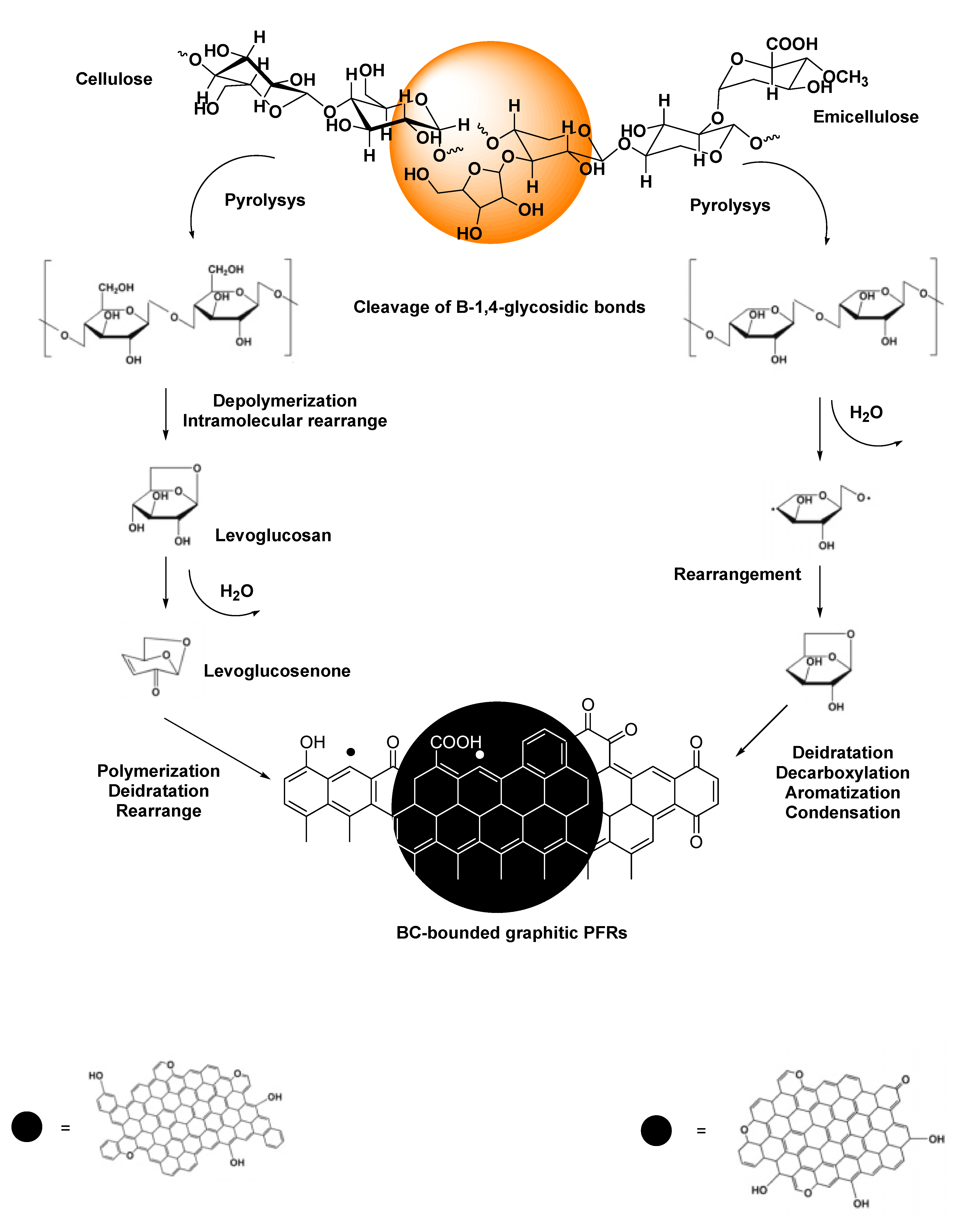
Table 1.
Endogenous and exogenous sources of ROS and the main reactive species of both oxygen and nitrogen (RNS) which can be consequently produced.
Table 1.
Endogenous and exogenous sources of ROS and the main reactive species of both oxygen and nitrogen (RNS) which can be consequently produced.
| Endogenous Sources | Exogenous Sources | Reactive Species | |
|---|---|---|---|
| Enzymatic | Non-Enzymatic | ||
| NOX MPO Cytochrome P450 Lipoxygenase Angiotensin II Xanthene oxidase Cyclooxygenase FpH• |
Mitochondria Respiratory chain Glucose auto-oxidation NAD• Semiquinone radicals Radical pyridinium Hemoproteins |
Air Water pollution Tobacco Alcohol Heavy/transition metals Drugs Industrial solvents Cooking Radiation EPFRs BC-PFRs |
O2•− H2O2 •OH •OOH ONOO• NO2• NO• ONOOCO2− NO2+ ONOOH N2O3 ONOO− ONOOCO2− CO3•− |
NADPH = nicotinamide adenine dinucleotide phosphate; MPO = myeloperoxidase; NOX = NADPH oxidase; NAD = nicotinammide adenina dinucleotide; Fp = flavoprotein enzymes; EPFRs = environmental persistent free radicals; BC-PFRs = biochar-related persistent free radicals.
Table 2.
The most representative radicals and not radical reactive species produced in biological aerobic systems with sources and functions.
Table 2.
The most representative radicals and not radical reactive species produced in biological aerobic systems with sources and functions.
| Reactive Specie | Source | Function |
|---|---|---|
| O2•− | Enzymatic process Autoxidation reactions Non-enzymatic electron transfer reactions |
Reduces iron complexes such as cytochrome C Oxidizes ascorbic acid and α-tocopherol |
| HOO• | Protonation of O2•− | Initiates fatty acid peroxidation |
| HO• | By H2O2 via Fenton reaction and HWR | Reacts with organic and inorganic molecules * |
| NO• |
L-arginine (substrate) NADPH (electron source) Nitric oxide-synthase |
Intracellular second messenger Stimulates GC and PK Causes smooth muscle relaxation in blood vessels |
| NO2• | Protonation of ONOO− Homolytic fragmentation of ONOOCO2− |
Acts on the antioxidant mechanism ↓ Ascorbate and α-tocopherol in plasma |
| ONOO• | Reaction of O2 with NO• | Oxidizes and nitrates methionine and L-tyrosine Oxidizes DNA to form nitroguanine |
| CO3•− | (SOD)-Cu2+ By reaction of •OH reacts with HCO3- |
Oxidizes proteins and nucleic acids |
| ONOOCO2− | Reaction of ONOO− with CO2 | Promotes nitration of oxyhaemoglobin’s tyrosine of the via free radicals |
HWR = Haber Weiss recombination; * DNA, proteins, lipids, carbohydrates; GC = guanylate cyclase; PK = protein kinases.
Table 3.
Oxidative modification of cellular macromolecules: reactions involved and produced markers of OS.
Table 3.
Oxidative modification of cellular macromolecules: reactions involved and produced markers of OS.
| Cellular Macromolecules | Reactions | OS biomarkers | Ref |
|---|---|---|---|
| Proteins | RNS with L-tyrosine | NT | [41] |
| Fenton reaction of ROS with L-lysine, L-arginine L-proline, L-threonine |
PC | [42] | |
| Proteins/lipids | Michael-addition of aldehydic lipid oxidation products to L-lysine, L-cysteine, L-histidine | PC | [42] |
| Proteins/lipids | Complex oxidative process | Ox-LDL | [43] |
| Proteins/carbohydrates | Glyco-oxidation between L-lysine amino groups and L-arginine carbonyl groups linked to carbohydrates |
AGEs | [44] |
| Lipids | •OH and HOO• mediated lipid peroxidation of poly-unsaturated fatty acids * | 4-HNE, MDA, F2-IsoPs | [41] |
| DNA | Mutagenic oxidation | 2-Hydroxy adenine 8-Oxoadenine 5-Hydroxycytosine Cytosine glycol Thymine Glycol 8-OHGua 8-OHdG |
[45] |
AGEs = advanced glycation end products (N-ε-carboxymethyl-lysine pentosidine glucosepane); 4-HNE = 4-hydroxynonenal; MDA = malondialdehyde; F2-IsoP = F2-Isoprostanes; Ox-LDL = oxidized low-density lipoprotein; PC = protein carbonyl; 8-OHGua; 8-hydroxyguanine; 8OHdG—8-hydroxy-2′-deoxyguanosine; * linoleic, arachidonic acids; NT = nitro tyrosine.
Table 4.
Endogenous and exogenous molecules which can counteract free radicals and reactive species toxicity. ↑ Means increase; ↓ means decrease/reduction.
Table 4.
Endogenous and exogenous molecules which can counteract free radicals and reactive species toxicity. ↑ Means increase; ↓ means decrease/reduction.
| Process | Endogenous Molecules | Actions | Exogenous Molecules | Effect |
|---|---|---|---|---|
| NE | Vitamin E Vitamin C Carotenes Ferritin Ceruloplasmin Selenium GSH Manganese Ubiquinone Zinc Flavonoids Coenzyme Q Melatonin Bilirubin Taurine Cysteine Albumin Uric acid |
Interact with ROS and RNS and terminate the free radical chain reactions | Vitamin C | ↓O2• ↓•OH |
| Vitamin E | ↓Lipid peroxidation | |||
| Resveratrol Phenolic acids Flavonoids |
↓O2• ↓•OH ↓Lipid peroxidation |
|||
| Oil Lecithin |
↓O2• ↓•OH ↓Lipid peroxidation |
|||
| Selenium Zinc |
Antioxidant | |||
| Acetylcysteine | Antioxidant | |||
| E | SOD | Converts O2 to H2O2 ↓Hydroxyl radical production |
||
| CAT | Decomposes H2O2 to H2O+O2 ↓Hydroxyl radical production |
|||
| GSH-Px | Converts peroxides and hydroxyl radicals into nontoxic forms by the oxidation of GSH into GSSG | |||
| GR | Converts glutathione disulphide to GSH | |||
| GSTs | Catalyses the conjugation of GSH to xenobiotic substrate | |||
| G6PD | Catalyses the dehydrogenation of G6P to 6-phosphoglucono-Δ-lactone | |||
| Nrf2 | Regulates the expression of antioxidant proteins | |||
| ARE | Encodes for detoxification enzymes and cytoprotective proteins | |||
| NQO1 | Catalyses the reduction of quinones and quinonoids to hydroquinone molecules | |||
| MSR | Carries out the enzymatic reduction of the oxidized form of methionine to methionine | |||
NE = not enzymatic; E = enzymatic; CAT = catalase; GSH-Px = Glutathione peroxidase; GR = glutathione reductase; GSH = reduced glutathione; Nrf2 = erythroid nuclear transcription factor-2; ARE = antioxidant response element; NQO1 = NAD (P)H quinone oxidoreductase 1; GSTs = glutathione S-transferases; G6P = glucosio-6-fosfato; G6PD = glucosio-6-fosfato dehydrogenase; GSSG = glutathione disulfide; MSR = methionine sulfoxide reductase.GSH-Px, GR, and MSR are the main intermediaries in the processes for repairing oxidative damage.
Table 5.
Conventional and alternative antimicrobials including some antibiotics, nanoparticles, and natural compounds, which exert their effects by generating ROS.
Table 5.
Conventional and alternative antimicrobials including some antibiotics, nanoparticles, and natural compounds, which exert their effects by generating ROS.
| Type | Name | Mechanism of Action | Clinical Application | Target | Ref. | ||||
|---|---|---|---|---|---|---|---|---|---|
| Antibiotics | Nitrofurantoin | Autooxidation of nitroaromatic anion radicals * in presence of O2 providing O2− and then ROS thus causing OS and toxicity to bacteria | UTI | E. coli | [18] | ||||
| Polymyxin B | Accumulation of OH• | Untreatable infections |
Acinetobacter baumannii Pseudomonas aeruginosa Enterobacteriaceae ** |
[58] | |||||
| Nalidixic acid | Mutagenesis by oxygen free radical generation |
Gastroenteritis Enteric fever Bacteremia |
Salmonella typhimurium | [59] | |||||
| Norfloxacin | |||||||||
| Norfloxacin Ampicillin Kanamycin Fluoroquinolones β-Lactams Aminoglycosides |
↑ Superoxide levels ↑ H2O2 ↑ Lethality by accumulation of OH• |
UTI | E. coli | [60] | |||||
| Nalidixic acid Trimethoprim Ampicillin Aminoglyoside |
Post-stress ROS-mediated toxicity | [61,62] | |||||||
|
Alternative antimicrobials |
Organo Metals |
OSECs | Ebselen *** | By inhibiting TrxR in bacteria laching glutathione thus triggering OS. |
Skin infections Bacteremia Endocarditis Food poisoning Pneumonia TSS |
S. aureus | [63] | ||
| Tuberculosis | M. tuberculosis | [64] | |||||||
| Nanomaterials | NPs | MSNP-maleamic | ↑ ROS (40% E. coli, 50% S. aureus) | UTI | E. coli | [65] | |||
| MSNPs-maleamic-Cu | ↑ ROS (40% E. coli, 30% S. aureus) | Skin infections Bacteremia Endocarditis Food poisoning Pneumonia TSS |
S. aureus | ||||||
| Metal oxide NPs | ↑ ROS ↑ RNS |
Skin infections Bacteremia Endocarditis Food poisoning Pneumonia TSS UTI |
E. coli S. aureus S. epiderdimis Photobacterium hosphoreum |
[64] | |||||
| NZs | AgPd0.38 | By ROS produced on Ag and Pd bimetallic alloy |
Severe infections |
S. aureus Bacillus subtilis E. coli P. aeruginosa |
[66] | ||||
| Metal based | AgRuSCs | AGXX® | By ROS catalytical production | UTI | Enterococcus faecalis | [54,67,68,69] | |||
| Skin infections Bacteremia Endocarditis Food poisoning Pneumonia TSS |
MRSA | ||||||||
| Natural Compounds | Allicin | OS by ↑ ROS via ↓ of low MW thiols | Skin infections Bacteremia Endocarditis Food poisoning Pneumonia TSS |
S. aureus | [70] | ||||
UTI = urinary tract infections; * by a NADH-dependent reduction; ** carbapenemase-producing; ↑ Means increase; ↓ means decrease/reduction; TSS = toxic shock syndrome; NPs = nanoparticles; *** used also in combination with ROS-producing antimicrobials such as silver nanoparticles (AgNPs}; NZs = nanozimes; OSECs = organo-selenium compounds; AgRuSCs = silver and ruthenium-based surface coatings; MRSA = methicillin-resistant S. aureus; MW = molecular weight.3.1.1. ROS Formation Induced by Antibiotics.
Table 6.
Most common photosensitizers used in clinical trials.
| Class | Compound | Description | Target | Abs max (nm) |
Impact in the field | Refs. |
|---|---|---|---|---|---|---|
| Phenotiazinium derivatives |
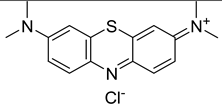 Methylene Blue |
3-ring p-system Auxochromic side groups Single positive charge SOQY < 0.5 Type I reactions |
Dental plaque | 632 | 1st clinically approved PS (dentistry) Standard PS in vitro |
[94] |
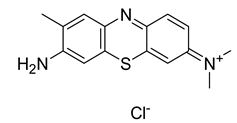 Toluidine Blue |
Streptococcus mutans | 410 | [95] | |||
| E. coli | [96] | |||||
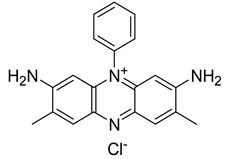 Safranine O |
F. nucleatum P. gingivalis |
520 | [97] | |||
| Porphyrin derivatives |
 Porphyrin |
Four pyrrole cycles Up to eight positive charges SOQY = 0.5-0.8 Occurring in nature Type II reactions |
S. aureus P. aeruginosa E. faecalis |
446 | Widely used as standard PS in vitro | [98,99,100] |
 TMPyP * |
MRSA ESBL K. pneumoniae |
421 | N.R. | [101] | ||
 XF-73 * |
Staphylococci Enterococci Streptococci S. aureus biofilm |
380-480 | [102] | |||
| Chlorin derivatives |
 Chlorin e6 |
Like heterocyclic-macrocyclic compounds Neutral or up to eight positive charges SOQY = 0.5-0.8 3 pyrrole and 1 pyrroline subunits Type II reactions |
S. aureus E. coli |
660 (neutral) |
[103] | |
| E. coli | 532 (cationic) |
[104] | ||||
 Photodithazine® |
MRSA MSSA |
660 | [105] | |||
| Phthalocyanin derivatives |
 Phthalocyanine |
4 pyrrole cycles Hydrophobic and uncharged Type II reactions |
A. hidrophila | 670 | [106] | |
| Xanthene derivatives |
 Eosin Y |
Anionic xanthene dyes Fluorescein derivatives SOQY = 0.5-0.6 Type II reactions |
MRSA MRSA biofilm S. aureus |
N.R. | Sparse studies in recent years | [107] |
 Erythrosine |
S. mutans Lactobacillus casei Candida albicans |
470 | [108] | |||
 Rose Bengal |
E. faecalis P. aeruginosa |
532 | [109] | |||
| Nanomaterials |
 Fullerene C60 *** |
Soccer-ball shaped cage molecules Made exclusively from carbon atoms Neutral Extended p-conjugated system Type I and II reactions |
S. aureus E. coli |
532 | Unique class of PS Sparse studies on effect on biofilms | [110] [111] |
| Phenalenones |
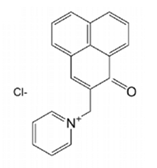 SAPYR |
Biosynthesized by plants to defend against pathogens using sun to generate singlet oxygen SOQY > 0.9 Positively charged pyridinium-methyl moiety Type II reactions |
E. faecalis Actinomyces naeslundii |
360-420 | First water-soluble exclusive type-II PS | [112] |
| Riboflavin derivatives |
 Vitamin B2 |
In PDT cationic derivatives with up to eight positive charges SOQY = 0.7-0.8 Type II reactions |
MRSA EHEC |
450 (cationic) |
N.R.° | [113] |
| Curcumins |
 Curcumin (neutral) |
Naturally occurring yellow dye from the rootstocks of Curcuma longa Approved as food additive (E100) For use in PDT positive charges have been included in derivatives structures Type I reactions |
S. mutans L. acidophilus |
547 | Novel positively charged derivatives with enhanced water solubility |
[114] [115] |
| Natural compounds |
 Hypericin (neutral) |
Naturally occurring For use in PDT positive charges have been included in derivatives structures Type II reactions |
S. aureus E. coli |
593 | N.R. | [116] [117] |
| 5-ALA |
 5-amminolevulinic acid |
δ-amino acid in which the hydrogens at the γ position are replaced by an oxo group Metabolized to protoporphyrin IX |
MRSA | 410 | Optical imaging agent | [118] |
SOQY = Singlet oxygen quantum yield * derivatives of porphyrin; ** derivative of chlorine e6; S = Abs max, Soret band; Q = Q band; *** functionalized in multiple ways by adding positively charged moieties; ESBL = extended spectra β-lactamases; MRSA = methicillin resistant S. aureus; MSSA = methicillin sensitive S. aureus; MRSE = methicillin resistant S. epidermidis; EHEC = Enterohemorrhagic E. coli; 5-ALA = 5-aminolevulinic acid.
Table 9.
Summary of a clinical register of the use of SHRO in various complex infections.
| Clinical site | Dosing regimen | Clinical details | Outcome | Adverse reports |
|---|---|---|---|---|
| Respiratory tract | Daily nebulized SHRO in respiratory nebulizer | Bronchiectasis, Several patients One patient—recurrent exacerbations with secondary infection with Mycobacterium avium |
Reduction in bacterial load and temporary eradication of M. avium (1 patient) | None |
| Scalp | Daily topical application for 6 weeks | Fungal kerion, T. tonsurans Patient intolerant to oral antifungals |
Complete resolution | None |
| Intraperitoneal | 50–100 g daily via abdominal drain | Severe four-quadrant peritonitis following intraabdominal infection and corrective surgery Patients also on systemic antibiotics Often polymicrobial infections with MDR strains and Candida spp. |
Variable, but general peritonitis Control |
N.R. to SHRO use |
| Abdominal wall Deep soft tissue |
SHRO into open cavity with each dressing | Used both prophylactically and therapeutically in around 20 patients | Prevention of infection and effective therapy in infected cavities | None |
| Prosthetic joints | Single dose around prosthetic joint at surgery Numerous patients |
Mixed microbiology including S. ludenensis Given in conjunction with systemic antibiotics | Good adjunct to existing management | None |
| Prepatellar bursitis |
Single application at debridement |
On immunosuppression for psoriasis M. malmoense isolated from prepatellar pus Put on clarithromycin, rifampicin and ethambutol + SHRO topically |
Complete healing and no further isolation of M. malmoense | None |
| Bladder | Twice-weekly instillation via suprapubic catheter | Several patients with long-term urethral or suprapubic catheters | Reduction in urosepsis | None |
| External auditory canal |
Daily with wick or cotton wool | Pseudomonas otitis externa | Resolved | None |
| Oral infections | Daily oral application of 10 g SHRO |
Recurrent aphthous ulcer, gingivitis, geographic tongue. No microbiology |
Reported reduction in duration of symptoms | None |
|
Helicobacter gastritis |
Once daily 10 g SHRO for 10 days | Confirmed Helicobacter pylori gastritis MDR strain and no response to antibiotic eradication regimens |
Continuation of symptoms Therapeutic failure |
None |
N.R. = Not related.
Table 10.
Commercially available honey-based wound healing products.
| Product | Description | Indications | Mechanism of Action | Ref. | Clinical Evidence |
|---|---|---|---|---|---|
| Activon® Manuka Honey Tube AM |
100% MGMH | Sloughy, necrotic wounds # Malodorous wounds # |
Debrides necrotic tissue Can be used in dressings or directly into cavities |
[190] | Blistering and cellulitis on a type 2 diabetic patient Pediatric burn Foot ulceration Grade 5 sacral wound [190] |
| Activon® Tulle AM | Knitted viscose mesh dressing with 100% MH | Granulating or shallow wounds Debriding or de-sloughing small areas of necrotic or sloughy tissue |
Creates a moist healing environment Eliminates wound odor Antibacterial action |
[190] | Over-granulated grade 3 and 4 pressure ulcers Extensive leg cellulitis Venous ulcer, chronic wound Infections, necrotic foot [190] |
| Algivon® Plus AM | Reinforced alginate dressing with 100% MH |
Cavities, sinuses, pressure, leg, diabetic ulcers Surgical, infected wounds Burns, graft sites Ideal for wetter wounds |
Absorbs exudate Debrides, removes slough Reduces bacterial load |
[190] | Chronic wounds [191] Burn wound management [192] |
| Algivon® Plus Ribbon AM | [190] | Autoamputation of fingertip necrosis [193] |
|||
| Aurum® ostomy bags WM | MGMH added to hydrocolloids |
Stoma care | Kills bacteria Suppresses inflammation Promote healthy skin around the stoma |
[194] | Pyoderma gangrenosum around ileostomy [195] |
| L-Mesitran® Border AME | Hydrogel + honey (30%) pad on a fixation layer | Chronic wounds $ | Exudate absorption Re-hydration of dry tissue Antibacterial properties |
[196] | Pediatric minor burns and scalds [197] |
| L-Mesitran® Hydro AME | Sterile, semi-permeable hydrogel dressing ## |
Chronic wounds * Superficial and acute wounds ** Superficial and partial-thickness burns *** Fungating wounds, donor sites Surgical wounds, cuts, abrasions |
Donates moisture to rehydrate dry tissue Antibacterial properties |
[196] | Pediatric minor burns and scalds [197] Fungating wounds [198] |
| L-Mesitran® Ointment AME |
Ointment $$ | Aids debridement and reduce bacterial colonization | [196] | Skin tears; irritation and inflammation [198] | |
| ManukaDress IG MPI | Wound dressing made with 100% Leptospermum scoparium @ | Leg and pressure ulcers First- and second-degree burns Diabetic foot ulcers, surgical and trauma wounds |
Osmotic activity that promotes autolytic debridement and helps maintain a moist wound environment | [152] | Burn management [152] Difficult-to-debride wounds [199] Necrotic pressure ulcer Recurrent venous leg ulceration [200] |
| Medihoney® Antibacterial Honey DSC |
100% sterilized MGMH | Deep, sinus, necrotic, infected surgical and malodorous wounds |
Creates an antibacterial environment Debridement on sloughy and necrotic tissue, removes malodor Provides a moist environment |
[201] | Wound healing [202] Prevention of catheter-associated infections in hemodialyzed patients [203] |
| Medihoney® Apinate Dressing DSC |
Calcium alginate dressing with 100% MGMH |
Diabetic foot, leg, pressure ulcers First- and second-degree partial-thickness burns Donor sites, traumatic, surgical wounds. |
Provides a moist environment Osmotic potential Draws fluid through the wound to the surface Low pH of 3.5–4.5. |
[204] | Venous leg ulcers [205] |
| Medihoney® Barrier Cream DSC |
Barrier cream with 30% MGMH | Protects skin damaged by irradiation treatment or in wet areas Prevents damage caused by shear and friction |
Maintains skin moisture and pH. | [206] | Treatment for intertrigo in large skin folds [207] |
| Medihoney® Antibacterial Wound Gel™ DSC |
Antibacterial wound gel & | Burns, cuts, grazes, and eczema wounds | Creates a moist, low-pH environment Cleans the wound by osmotic effect Reduces the risk of infection |
[208] | Reduction in incidence of wound infection after microvascular free tissue reconstruction [209] |
| SurgihoneyRO™ MH | Antimicrobial wound gel && | Infected, chronic wounds | Antimicrobial activity by controlled release of H2O2 Promotes debridement and new tissue growth |
[210] | Prevention of caesarean wound infection Prevention/eradication of bacterial colonies in dressing oncology long vascular lines; ulcers, surgical wounds and trauma wounds [183,185,211] In vitro activity against biofilm-producing clinical bacterial isolates [178] |
MGMH = Medical grade manuka honey; AM Advances Medical manufacturer; WM Welland Medicals Ltd manufacturer; AME Aspen Medical Europe Ltd manufacturer; MPI Medicareplus International manufacturer; DSC Derma Science-Comvita manufacturer; MH Matoke Holdings Ltd manufacturer; # pressure ulcers, leg ulcers, diabetic ulcers, surgical wounds, burns, graft sites, infected wounds, cavity wounds and sinuses; $ pressure ulcers; superficial and partial-thickness burns; venous, arterial, and diabetic ulcers; ## contains 30% honey with vitamin C and E, as well as an acrylic polymer gel and water, with a polyurethane film backing; * pressure ulcers, venous and diabetic ulcers; ** cuts, abrasions and donor sites; ***first- and second-degree; $$ made of 48% medical-grade honey, medical-grade hypoallergenic lanolin, oils, and vitamins; @sterile honey from New Zealand. Non-adherent impregnated gauze; & 80% medical-grade manuka honey with natural waxes and oils; && utilizes bioengineered honey to deliver Reactive Oxygen® (RO™).3.3.2. Antibacterial Mechanisms of Honey.
Table 11.
Components that confer honey its antibacterial effects and would healing properties.
| Antibacterial factors | Sources | Mechanism of formation | Effects on Bacteria | Ref. |
|---|---|---|---|---|
| H2O2 | Glu | Oxidation of Glu deriving by sucrose captured by bee from flowers | OS DNA damage |
[152] |
| Bee Def-1 | Bee’s hypopharyngeal gland | Innate immune response | Create pores in membrane Interferes with bacterial adhesion Alters the production of EPSs |
[215] |
| Acidic pH (3.4-6.1) |
GluLac GluA |
By enzymatic oxidation of Glu | Prevents bacterial growth | [152] |
| MGO * | Dihydroxyacetone | Heating | Alters bacteria fimbria and flagella |
[220] |
| Osmotic pressure |
Super concentration of sugars | N.A. | ↓ Availability of free water molecules ↓ Bacterial growth |
[221] |
| Polyphenols | Flowers as secondary metabolites | Flowers metabolism | Pro-oxidative properties Accelerate HO• formation Oxidative DNA breakage Non-enzymatic ↑ H2O2 |
[222] |
Glu = glucose; GluLac = gluconolattone; GluA = gluconic acid; EPSs = extracellular polymeric substances; Bee Def 1 = Bee defensin 1; MGO = methyl glyoxal; * exclusively in Leptospermum honeys (e.g. manuka); ↑ means increase, more; ↓ means decrease/reduction, less, minor.
Table 12.
Diseases treatable with HBOT.
| Disease | Cause */Circumstances ° | Ref |
|---|---|---|
| CO poisoning | [233] | |
| Acute anemia | [234] | |
| Rheumatoid arthritis Cell hypoxia |
Polarization of Th17 cells to T reg * | [235] |
| Inflammatory disorders (by ↓ inflammatory mediators) |
Ischemic circumstances ° Compartment syndrome ° |
[227] |
| Microcirculatory disorder | ||
| ↓ Leucocyte chemotaxis and adhesion ↑ Proliferation of neutrophiles |
[236] | |
| Autoimmune syndrome Immune reactions in antigens Autoimmune symptoms. |
Proteinuria ° Facial erythema ° Lymphadenopathy ° |
[237,238] |
| ↓ Lymphocytes and leukocytes | [234] | |
| ↑ Mitochondrial Function | [239] | |
| Chronic skin damage healing (by angiogenesis) |
[240] | |
| Recalcitrant infections | Necrotizing fasciitis ° Osteomyelitis ° Chronic soft tissue infections ° Infective endocarditis ° Acute or chronic wounds ° Diabetic foot ulcers ° |
[227,241,242,243,244,245] [246,247] |
| Ocular disorders | Cystoid macular edema ° Scleral thinning ° Necrosis faced after pterygium surgery ° Nonhealing corneal edema ° Anterior segment ischemia ° Some blinding diseases ° |
[248,249] |
| Brain/cerebral injuries | Ischemic-reperfusion damage ° | [250] |
| Cancer | [251] | |
| Complications by radiotherapy | Radiation-induced skin necrosis ° | [252] |
↑ Means increase, improvement; ↓ means decrease/reduction.
Table 13.
Overview of some clinical studies investigating the application of HBOT for different infections [227].
Table 13.
Overview of some clinical studies investigating the application of HBOT for different infections [227].
| Infections | Study papulation | Treatment sessions * | Pressure ** | Exposure Time (min) | Main findings | Ref |
|---|---|---|---|---|---|---|
| Burns | 53 | Based on outcomes | 2.5 | 90 | All patients survived | [264] |
| Burn | 40 | 10 | 2.5 | 80 | Faster healing, shorter hospitalization | [265] |
| Brain abscess | 41 | 4–52 | 2.5–2.8 | 25– | ↓ Treatment failures, ↑ outcomes | [266] |
| SSIs | 42 | 30 | 2.4 | 90 | ↓ Post-surgical deep infections in CSD | [267] |
| SSIs | 32 | Based on outcomes | 2-3 | 90 | Valuable AT for PO organ/space S-SSI | [268] |
| SSIs | 6 | 28–106 | 2.5–2.8 | 75 | AT for early PO deep infections | [269] |
| NSTI | 48 | Based on outcomes | 3 | 90 | Not ↓ mortality rate, number of debridement, hospital stay, antibiotic use |
[270] |
| NSTI | 44 | 2.8 | 60 | ↑ Survival and limb salvage | [271] | |
| NSTI | 32 | 2.8 | 45 | AT | [272] | |
| NSTI | 37 | 2.5 | 45 | Doubtful advantage of using HBOT as AT for NF in ↓ mortality and morbidity | [273] | |
| DFIs | 100 | 20 to 30 | 2–3 | 90 | Useful AT for nonhealing DFIs | [274] |
| DFIs | 42 | Group1:<10 Group 2:>10 |
2.5 | 120 | ↓ Amputation rate | [247] |
| DFIs | 94 | 40 | 2.5 | 85 | Facilitates healing of chronic DFIs | [275] |
| DFIs | 35 | 38±8 | 2.2–2.5 | 90 | ↓ Amputations | [276] |
| DFIs | 28 | 20 | 2.5 | 90 | ↑ Healing rate of nonischemic chronic DFIs | [277] |
| DFIs | 36 | 2.5 | 90 | Healing response in chronic DFIs | [278] | |
| DFIs | 38 | 40–60 | 2.5 | 90 | ↑ Healing rate ↓ Amputation rate |
[279] |
| DFIs | 18 | 30 | 2.4 | 90 | AT when reconstructive surgery is not possible | [280] |
| Osteomyelitis | 6 | 2.0–2.4 | 30 | Effective following failure of primary therapy of osteomyelitis | [281] | |
| Osteomyelitis | 14 | 2.5 | 120 | Effective and safe for chronic refractory osteomyelitis |
[282] | |
| Osteomyelitis | 1 | 2 | – | Early use of HBOT for a compromised host who develops recurrent osteomyelitis | [283] | |
| Osteomyelitis | 12 | Based on outcomes | 2.5 | 90 | AT for patients who develop S-SSI and osteomyelitis after CTS |
[284] |
* Days; ** ATA = atmospheres absolute; DFIs = diabetic foot infections; HBOT = hyperbaric oxygen therapy; NSTI = necrotizing soft tissue infections; SSIs = surgical site infections; S-SSIs sternal surgical site infections; NF = neurofibromatosis; ↑ means increase, improvement, improved, high, higher; ↓ means decrease/reduction, low, lower, less; CTS = cardiothoracic surgery; CSD = complex spine deformity; PO = postoperative; AT = adjuntive treatment.
Table 14.
Studies finalized to investigating the application of HBOT in treatment of different surgical site infections (SSI) [229].
Table 14.
Studies finalized to investigating the application of HBOT in treatment of different surgical site infections (SSI) [229].
| Study | Surgery | SSI | Population ***/# |
ATA Time (min) |
Outcomes and Conclusion | Refs. |
|---|---|---|---|---|---|---|
| Case report | CTS | S-SSI | 1 | 2.40 90 |
Rapid healing and epithelialization * | [285] |
| Retrospective | Sternotomy | S-SSI | 55 | 2.50 90 |
S-SSI cured in all patients within an average of 8 weeks No in-hospital death ↑ Clinical outcome in patients with sterno-mediastinis and post-sternotomy wound infection after CTS |
[286] |
| Prospective trial | CTS | S-SSI | 32; 14/18 | 2–3 90 |
S. aureus was the most common pathogen #, *** Infection duration was similar #,***,** Infection relapse rate was significantly ↓ in *** Intravenous antibiotic use duration ↓ in *** Total hospital stay ↓ in *** HBOT could be a valuable AT for treating PO organ/space S-SSI |
[268] |
| Case report | CTS | S-SSI | 1 | 2.50 90 |
Sternal wounds totally healed and epithelized in 9 weeks HBOT with TNP dressing is a good alternative method for patients who cannot tolerate or refuse any surgical reconstruction |
[287] |
| Retrospective | NMSS | DWI | 6 | 2.50 3 × 25 |
All infections resolved Wound healing in an average of 3 months Minor side effects of HBOT HBOT is a safe AT for early deep PO infections in case of spinal implants in HR pediatric patients |
[288] |
| Retrospective | CTS | S-SSI | 12; 6/6 | 2.50 90 |
No treatment-related complication in *** Length of stay in ICU ↓ in *** Invasive and noninvasive positive pressure ventilation ↓ in *** Hospital mortality ↓ in *** HBOT may be used as a safe AT to ↑ clinical outcomes in patients with S-SSI and osteomyelitis after sternotomy and CTS |
[284] |
| Retrospective | CABS | Mediastinitis | 18 | 2.50 90 |
1 HBOT-unrelated death caused by sepsis HBOT was well-tolerated Favorable clinical outcomes using HBOT as AT for treating mediastinis patients after CABS |
[289] |
| Retrospective | NMSS | DWI | 42; 18/24 | 2.40 90 |
11.9% (5/42) Incidence of infection in both ***,# 5.5% (1/18) Infection rate in *** 6.6% (4/24) Infection rate in # HBOT significantly ↓ PO infections in NMSS patients HBOT is a safe AT to prevent PO deep infections in complex spine deformity in HR NM patients |
[267] |
| Retrospective | CTS | S-SSI | 10 | 2.50 92 |
70% complete wound healing with fibrous scar formation in 4 weeks HBOT *** HBOT had 80% success as AT in DSWI *** No complications were observed |
[290] |
| Retrospective | NM | HRI | 14 | 2.0–2.8 75 |
86% of HRI successfully treated without hardware removal *** Hardware was removed following HBOT failure in 2 infections Observed intrathecal pump malfunction caused by HBOT (2.8 bars) HBOT has potential as AT in the treatment of HRI in NM Diminished need for hardware removal and treatment interruption |
[291] |
| Retrospective | CTS | S-SSI | 53 | Readmitted with infected sternotomies discharged in 2–39 days Time for wounds healing using NPWT alone was 21–42 days in # Time for wounds healing using NPWT+HBOT was 28–42 days in *** HBOT was 5–35 days HBO treatments were 22.6 (+11.06) Restore time 21–98 days in *** (NPWT+HBOT) Multimodality therapy of incision and drainage using NPWT+HBOT+antibiotics is successful for treat complex deep sternal wound infections in the pediatric population after congenital heart surgery |
[292] | |
| Retrospective | MtF-GAS | SSI | 33; 15/18 | 2.2–3.0/90 | 100% Complete wound healing *** 94.4% Complete wound healing # ↓ Duration of antibiotic therapy *** ↓ Perineal drain time *** ↓ Bladder catheter time *** ↓ Hospital stay *** HBOT as effective adjuvant treatment for SSIs in patients undergoing MtF GAS |
[293] |
* First reported case of HBOT use for the treatment of deep sternal (surgical site infections) SSI in a heart transplant recipient; S-SSI = sternal surcical site infections; ** 31.8 ± 7.6 vs. 29.3 ± 5.7 days, respectively, p = 0.357); ↑ means increase, improvement, improved, high, higher; ↓ means decrease/reduction, low, lower; MtF-GAS = male-to-female gender affirmation surgery; *** in HBOT group; # in not HBOT group, AT = adjuvant therapy; PO = postoperative; TNP = topical negative pressure; CTS = cardiothoracic surgery; CABS = coronary artery bypass surgery; DSWI = deep sternal wound infections; HRI = hardware-related infections; NMSS = neuromuscular sclerosis surgery; NM = neuromodulation; DWI = deep wound infections; ICU = intensive cure units; HR = high risk; NPWT = negative pressure wound care therapy.
Table 15.
Factors influencing PFRs formation in BC.
| Parameter | Influencing Factors | Specifications | Observations | Ref. |
|---|---|---|---|---|
| PFRs concentration |
Biomass type | Cow manure, rice husk, others (< 500°C) | ≠ Concentrations | [305,306] |
| Non-lignocellulosic biomass with ⇓ H/C and O/C | ⇓ Concentration | [307] | ||
| Lignocellulosic biomass | ⇑ Concentration | |||
| Temperature | 300°C, 700°C | ≠ Concentrations | [305] | |
| Maximum concentration at 600°C | [308] | |||
| Maximum of concentration at 500-600°C | [309] | |||
| Transition metals | Adsorb onto biomass and transfers electrons from polymer to metal center during pyrolysis | ⇑ Concentration | [25] | |
| Type of PFRs | Temperature | 200-300°C | OCR | [309] |
| 400°C | OCR + CCR | |||
| 500-700°C | CCR |
OCR = Oxygen centered radicals; CCR = carbon centered radicals [24].
Table 16.
BC-derived PFRs and their applications as ROS- forming antibacterial agents reported in the years 2019-2023.
Table 16.
BC-derived PFRs and their applications as ROS- forming antibacterial agents reported in the years 2019-2023.
| Biomass | Pyrolysis °C/Time | BC-name | Active radicals | Radical Mechanisms | Degraded compound | Refs. |
|---|---|---|---|---|---|---|
|
Anaerobic
digestion sludge |
400°C 600°C 800°C 1000°C |
ADSBC 400 ADSBC 600 ADSBC 800 ADSBC 1000 |
SO4•- PFRs OH• | BC-mediated activation of PDS | Dyes, Estrogens Sulfonamides, E. coli Others |
[30] |
|
Caragana korshinskii |
650 °C/3h | ACB-K-gC3N | PFRs h+•OH• O2- | Electron photogeneration and PFRs mediated H2O and O2 activation |
S. aureus, E. coli RhB, TC, NOR, CAP |
[31] |
| Pinewood | 600°C | Ag0-PBC | PFRs •OH • O2- | UV-light promoted excitation of the electron-hole pairs and subsequently, the production of ROS Enhanced ROS generation by PFRs |
MB, E. coli | [32] |
BCs = biochar; TC = tetracycline; RhB = rhodamine B; PDS = peroxydisulfate; CAP = chloramphenicol; MB = methylene blue; NOR = norfloxacin.The employed BC was derived from pyrolysis of sludge, Caragana korshinskii plant and pinewood. In these processes, the electron transfer promoted by PFRs of diverse nature, generated ROS such as SO4•-, •OH, •O2-, •O2H, which carried out the oxidative degradation of different organic pollutants, including drugs, dyes, antibiotics, hormones, and showed antibacterial effects against E. coli and S. aureus.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



