
Submitted:
24 May 2024
Posted:
24 May 2024
You are already at the latest version
Abstract
Exosomes, natural nanovesicles that contain a cargo of biologically active molecules such as lipids, proteins, and nucleic acids, are released from cells to the extracellular environment. They then act as autocrine, paracrine, or endocrine mediators of communication between cells by delivering their cargo into recipient cells and causing downstream effects. Exosomes are greatly enriched in miRNAs, which are small non-coding RNAs that act as cytoplasmic posttranscriptional repression agents, modulating the translation of mRNAs into proteins, as well as nuclear transcriptional gene activators. Neuronal exosomal miRNAs have important physiologic functions in the Central Nervous System (CNS), including cell-to-cell communication, synaptic plasticity, and neurogenesis, as well as modulating stress and inflammatory responses. Stress-induced changes of exosomal functions include effects on neurogenesis and neuroinflammation, which can lead to the appearance of various neuropsychiatric disorders, such as schizophrenia, major depression, bipolar disorder, and Alzheimer's and Huntington’s's diseases. The current knowledge regarding the roles of exosomes in the pathophysiology of common mental disorders is discussed in this review.
Keywords:
exosomes
; microRNAs
; stress response
; synaptic plasticity
; neurogenesis
; major depression
; bipolar disorder
; schizophrenia
; Alzheimer disease
; Huntington’s disease
1. Introduction
1.1. Exosomes and Their Functions
As part of their physiological functions, virtually all cells release nanovesicles known as exosomes. These belong to the broader category of extracellular vesicles (EVs), a group of vesicles surrounded by a lipid bi-layer membrane and released from cells to their extracellular environment. Apart from exosomes (40-200nm), other types of EVs are microvesicles (950-1000 nm) and apoptotic bodies (500-2000nm) [1].
Exosomes contain a cargo of biologically active molecules, such as lipids, proteins, and nucleic acids. Their main role is to act as autocrine, paracrine, or endocrine mediators of communication between cells [2,3]. Another important role of exosomal is to discard useless lysosomal materials from the cell, such as misfolded proteins or cytoplasmic DNA [4]. Exosomes have crucial roles in a wide variety of biological processes, namely, the stress and immune responses, as well as cell differentiation, angiogenesis, and tissue regeneration. Last but not least, exosomes are part of the pathophysiologic mechanisms of various diseases, such as heart failure, cancer, and neurodegenerative disorders [5].
1.2. The Biogenesis of Exosomes
The process by which exosomes are manufactured starts inside the cell, within the endosomes. The constant folding of the endosomal envelope gives rise to a large number of intraluminal vesicles (ILVs). The resultant mature endosomes, called multivesicular bodies (MVBs), act by transferring molecules marked by mono-ubiquitination or by tetraspanins. The endosomal sorting complex required for transport (ESCRT), a multi-subunit, cytoplasmic protein machinery that takes part in membrane budding and tearing [6], coordinates the assembly of the mono-ubiquitinated proteins into the vesicles, while the ESCRT-independent pathway, composed of tetraspanins and the lipid ceramide, controls the incorporation of their cargo into the vesicles and the configuration of their membrane curvature [7,8]. Following their assembly, the MVBs utilize the cell cytoskeleton to carry themselves to the plasma membrane and fuse with it (Figure 1). The fusion of the MVBs with the plasma membrane is regulated by the cell-specific GTPase family of Rab proteins and the SNARE protein complex (SNAP receptor proteins) [9,10].
By the time the ILVs are released by the MVBs into the extracellular space, they are called exosomes. The exosomes then can be identified by protein biomarkers, such as the previously mentioned tetraspanins (CD9, CD63, CD81, CD82), the ALG2-interacting protein X (Alix), the heat-shock protein 70 (Hsp70), and the tumor susceptibility gene 101 (TSG101) protein [11,12]. The specific composition of exosomes is affected by a variety of factors, such as the cell origin and metabolic status, the cell release site, and the physiologic context of the cell [13]. Exosomes may be transported through biological fluids, such as plasma, to deliver their cargo into recipient cells, and to cause downstream effects, including the triggering of signaling cascades [10,14].
Exosomes may interact with and transfer their cargo to their target cells through cell surface adhesion molecules (phosphatidylserine receptors, lectins, integrins, etc.) in one of the following ways:
- Exosomal membrane proteins directly activate membrane receptors of target cells without being internalized,
- Exosomal membrane proteins are cleaved by proteases, and the resultant soluble fragments bind to membrane receptors of the target cells and
- Exosomes are internalized into the target cells, via endocytosis, phagocytosis, or direct fusion with the plasma membrane of the target cells.
1.3. The Cargo of Exosomes
As mentioned above, exosomes contain a complex cargo of proteins, lipids, and nucleic acids [2]:
- (a)
- Their protein cargo consists of the most commonly expressed endosomal proteins incorporated during the biogenesis of the MVBs (Alix, TSG101, heat shock proteins Hsc/Hsp 70 and 90), the vesicular transport and fusion proteins (Rab GTPases, SNAREs, annexins, and flotillin), and the proteins that interact with the target cell integrins and tetraspanins. Apart from the above, however, the protein composition of each exosome is diverse and depends on the cell type of origin and its physiologic or pathologic state [11].
- (b)
- Their lipid cargo includes cholesterol, sphingolipids (e.g., ceramide), and glycerophospholipids [15].
- (c)
- Their nucleic acid cargo includes DNA, mRNA, and non-coding RNAs (ncRNAs). The latter subdivided into long non-coding RNAs (lncRNAs) and short ncRNAs -including microRNAs (miRNAs)-react with other nucleic acids and proteins of the target cell and dynamically modify its gene expression and protein translation.
1.4. Exosomal miRNAs: Biogenesis and Function as Modulators of Gene Expression
Interestingly, in comparison to other biological fluids, exosomes are greatly enriched in miRNAs [16]. MicroRNAs are short ncRNAs with an average length of 19-24 nucleotides [17,18]. They comprise one of the most substantial subcategories of ncRNAs and are carried throughout the body mainly as exosomal cargos [19,20]. The miRNAs may vary significantly in terms of nucleotide sequence. However, they all function as posttranscriptional repression agents; thus, they down- or up-regulate gene transcription through the formation of complexes that interact with gene promoter regions and through synergistic interaction with other miRNAs. They can also bind complementarily to the 3′-untranslated region (3- UTR) of other mRNAs and regulate protein translation (Figure 2) [21].
The biogenesis of miRNAs begins in the nucleus, where RNA polymerases II and III transcribe the corresponding genes into several kilobases long, hairpin-loop pri-miRNAs [22]. Following this, pri-miRNAs are cut up by the enzymatic complex Drosha-DGCR8 (DiGeorge syndrome Critical Region gene 8) into stem-loop pre-miRNAs [18,23]. The pre-miRNAs are transferred to the cytoplasm by exportin 5 and Ran GTPase, where they are processed by the RNAse III enzyme Dicer and the transactivation-responsive RNA binding protein (TRBP). These enzymes produce a mature, 22nt-long, double-stranded miRNA with a 5′ phosphate end [24]. The double-stranded miRNA becomes unwinded, producing two strands. One strand is degraded, while the other strand represents the mature miRNA, which is integrated into the RNA-mediated silencing complex (RISC), acquiring the ability to repress translation (Figure 3) [25].
The incorporation of miRNAs into the exosomes can be exosome type-specific, in which case processes such as the neural sphingomyelinase 2-dependent pathway [26] and the miRNA-induced silencing complex-related pathway regulate the sorting of miRNAs [27]. It can also be independent of the miRNA sequence, mainly controlled by enzymes and other function-related proteins [28].
Taking into account all miRNA subcategories, a very broad class is represented by neuronal miRNAs, which comprise about 70% of all miRNAs in our body [29]. Neuronal miRNAs have very important functions in the CNS, among which are neuronal development, neurogenesis, and synaptic plasticity. A single neuronal miRNA molecule can interact with hundreds of other mRNA molecules, even without full complementarity [30,31]. As a result, neuronal miRNAs are integral regulatory molecules in both physiological conditions of the CNS and in disease states, making them an important research area to focus on.
2. The Role of Exosomes and Exosomal miRNAs in the CNS
In the brain, exosomes are involved in various, often overlapping processes. Such processes include cell-to-cell communication, neurogenesis, synaptic plasticity, and stress response.
2.1. Cell-to-Cell Communication
Neurons contain a high amount of MVBs in their soma and dendrites but not along their axons [32]. This observation led to the hypothesis that exosomes traveling along the neuraxes have a specific role in the neural synapses, taking thus part in cell-to-cell communication. Indeed, exosomes are critical for signal transmission in the CNS, as they act as vehicles of communication between cells of the same or different types, particularly between neurons and glia. The release of exosomes from CNS cells is a process controlled by synaptic glutamatergic activity and calcium influx [33,34].
2.2. Neurogenesis
Exosomes may both promote and hinder adult hippocampal neurogenesis (AHN), as shown by Luarte et al. [35]. Furthermore, the injection of cultured exosomes containing known pathogens into the dentate gyrus of mouse hippocampi caused significant impairment of AHN [36].
Of all miRNAs, the most prevalent in the brain is miR-124 since it is a key regulator of adult neurogenesis by targeting cAMP-response element binding protein (CREB) [37,38]. If dysregulated, miR-124 can cause significant neurodegeneration. Also involved in neurogenesis are miR-137, which regulates the differentiation of neural stem cells into mature neurons [39], and miR-125b, which mediates axon outgrowth [40,41].
2.3. Synaptic Plasticity
Synaptic plasticity is of great importance because it is involved in the processes of learning and memory, and if disrupted, it can cause a range of psychiatric disorders. As revealed by protein analysis of exosomes in the CNS, it is greatly influenced by exosomes [42,43]. Goldie et al. showed that neuron-to-glia signaling via exosomes helps active synapses stimulate the pruning of neighboring synapses that are inactive. More specifically, they identified microtubule-associated protein 1B (MAP1b)), a protein essential to synaptic plasticity, in exosomes from depolarized human neurons in culture [44]. A deficit in the synaptic signal transduction and over-abundance of cortical miRNAs in patients with schizophrenia suggested that miRNAs are an important component in the regulation of neuronal plasticity [45,46]. Furthermore, Bahrini et al. showed that when microglial cells were incubated with neuron-derived exosomes, there was an increase in the expression of complement component 3 (C3) in the microglial cells, which facilitated the removal of neurites [47].
Schratt et al. as well as Le et al. suggested that miR-132, -134, and -let-7 were important in the regulation of synaptic plasticity [41,40]. Castrén and Rantamäki showed that several miRNAs influenced the expression of the gene encoding brain-derived neurotrophic factor (BDNF), which in turn promoted the synthesis of miR-132, increasing neurogenesis [48]. They proposed that BDNF is involved in synaptic plasticity and that this is important for recovery from depression.
2.4. Stress Response
CNS signaling via exosomes is involved in important processes, including the stress response. Indeed, neurons release various neurotransmitters, which stimulate the secretion of exosomes from the oligodendrocytes. In turn, neurons internalize these oligodendrocyte-derived exosomes, which offer them cellular protection, as their cargo increases the tolerance of neurons to stress [49].
Neurons are also protected from stress via astrocyte-derived exosomes, through the prion protein (PrP), and other neuroprotective molecules. According to Guitart et al., the PrP contained in astrocyte-derived exosomes protects neurons from stress conditions like ischemia, hypoxia, hypoglycemia, and oxidative stress [50]. Apart from PrP, astrocyte-derived exosomes contain synapsin 1, heat shock protein 70, and matrix metalloproteinases [51,52]. As a result, upon heat, hypoxia, or any other stressful stimulus, astrocyte exosomes increase the viability of neural cells [53].
2.5. Mitochondrial Function in Brain
The mitochondria play a pivotal role in brain activity, therefore in recent years escalating research has focused on the profound impact of mitochondrial dysfunction on the pathogenesis of mental health conditions. Beyond their role as primary energy source of the cell, mitochondria oversee many processes vital for cellular survival, including redox signaling and calcium transportation. This intricate web of functions underscores the indispensability of mitochondria in various network processes, emphasizing their central role in CNS homeostasis and pathological states. The “mitochondrial hypothesis” posits that mitochondrial dysfunction is intricately linked to a broad spectrum of processes influencing the progression and severity of mental disorders [54].
Impaired mitochondrial functioning may result from several causes, including alterations in mitochondrial gene expression. In that respect, a subcategory of miRNAs is called mitochondrial miRNAs (mitomiRs). Named after their site of action, mitomiRs can originate from the nucleic DNA and then be transported into the mitochondria or can be transcribed directly from the mitochondrial DNA. The highly coiled, circular human mitochondrial DNA houses 37 genes and approximately 150 miRNA sequences [55]. However, due to the absence of the DICER and Drosha enzymes, the transcription and biogenesis of mitomiRs in mitochondria remain largely unknown [56]. There is evidence that mitomiRs are involved in several critical mitochondrial functions, such as oxidative phosphorylation (OXPHOS), electron transport chain (ETC) components, redox signaling, lipid metabolism, mitochondrial membrane potential, and transportation of calcium [57]. In addition, mitomiRs may influence nuclear gene expression and mRNA processing through mitochondrial retrograde signaling in the host cell or though their incorporation in exosomes in other cells [58].
3. Stress-Induced Changes in CNS Exosomal Functions
According to Chrousos: “All organisms must preserve a complex dynamic equilibrium, or homeostasis, which is repeatedly challenged by internal or external adverse forces termed stressors. Stress occurs when homeostasis is threatened or perceived to be so; homeostasis is re-established by various physiological and behavioral adaptive changes” [59].
Although stress forces virtually all biological systems away from a physiological steady state, it is a fundamental element of the natural world, which may lead to adaptational changes and progressive individual and species improvement. Thus, in acute, transient, motivating, and surmountable stress states, adaptive responses are beneficial. In contrast, in prolonged stress, when the adjustive capacity of the organism is exceeded, the adaptive changes may become detrimental, damaging cerebral functions and favoring the development of mental diseases [59].
3.1. Neurogenesis
As reported by Luarte et al., miRNAs derived from astrocytes possibly regulate neurogenesis under stress. In fact, astrocyte-derived miRNAs normally take part in neurogenic processes and are modulated by different stimuli. Under stress conditions, they may be up- or down-regulated and secreted via exosomes into the interstitial glymphatic space [60], contributing to cellular communication between astrocytes and neurons [35].
Focusing on microglia, Fan et al. showed that miR-146a-5p hinders neurogenesis in depression by targeting Krüppel-like factor 4 (KLF4) [61]. Furthermore, Wei et al. demonstrated both in vivo and in vitro that miR-139-5p is a negative regulator of neural stem cell proliferation and differentiation. As a result, exosomes from patients with major depressive disorder might have contributed to depressive-like behaviors in mice through miR-139-5p-suppressed neurogenesis [62].
3.2. Neuroinflammation
There is extensive evidence for the roles of exosomes in stress-induced neuroinflammation. First, Frühbeis et al. showed that reactive microglia released exosomes and microvesicles (MVs) carrying the pro-inflammatory cytokine interleukin-1β (IL-1β), the IL-1β processing enzyme caspase-1, and the purinergic receptor P2X 7 (P2RX7), all of which may induce and propagate inflammatory reactions throughout the brain [63]. Moreover, according to Dozio & Sanchez, the exosomal protein cargo from brain endothelial cells is modified after exposure to the proinflammatory cytokine tumor necrosis factor-alpha (TNF-α) [64]. They carry proteins involved in the signaling pathways of TNF-α and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), which may cause the low-level, chronic neuroinflammation associated with depression [65].
In addition, monocytes activated by interferon alpha (IFNα) release exosomes with altered miRNA profiles that could influence brain macrovascular endothelial cells (BMECs) and trigger an inflammatory reaction [66]. Taken into consideration, along with analyses of blood-brain barrier (BBB) permeability in mental disorders, the above evidence suggests that changes in BMEC could result in a leaky BBB, extended neuroinflammation, and eventually in the onset or progression of mental disorders [67].
Another miRNA, miR-9-5p, is capable of causing significant neuronal injury. When this miRNA was transferred as an exosomal cargo from neurons to microglia, it led to polarization of the latter into M1-type, damaging brain tissue [68].
3.3. Stress-Induced Epigenetic Modifications
Exosomes, by carrying genetic cargo like miRNA, serve as vehicles of transcriptional and translational regulators. As exosomes fuse with the recipient cells, they inject into them their miRNA cargo components, which act as targeted epigenetic modifiers leading to biochemical changes that may be associated with mental disorders [69,70]. Recent research has revealed the involvement of several miRNAs in the molecular mechanisms that contribute to the clinical manifestations of mental disorders.
4. The Role of Exosomes in the Pathogenesis of Mental Disorders
Mental disorders comprise major causes of misery and disability in humans. Their prevalence has been increasing significantly during the last few years, especially during and after the COVID-19 pandemic. Recent research in the field of exosomes and exosomal miRNAs allows some insight into CNS pathophysiology and may result in the development of new diagnostic and therapeutic methods. The possible roles of exosomes and miRNAs in the pathophysiology of common neuropsychiatric disorders, namely schizophrenia, major depression, bipolar disorder, and Alzheimer's and Huntington’s's diseases (summarized in Table 1), are discussed.
4.1. Schizophrenia
Schizophrenia (SCZ) is a chronic mental disorder that is characterized by motivational, cognitive, and affective dysfunction and psychosis. The pathogenesis of SCZ has a diverse genetic, epigenetic, and neurobiological basis, which involves alterations in brain structure and function, where dopamine dysregulation is shown to play a central role [71].
Exosomes may be involved in the pathophysiology of schizophrenia (SCZ) in multiple ways, mainly by causing neurotoxicity. Glial cells secrete, through exosomes, N-methyl D-aspartate receptor C (NMDAR-C) C-C motif chemokine receptor 2 (CCR2), NMDAR-C-X-C chemokine receptor type 4 (CXCR4), and NMDAR- interleukin 1 receptor type II (IL1R2) heteromers, which may form complexes with NMDARs, reducing NMDAR signaling [72]. In addition, analysis of blood exosomes of SCZ patients showed up-regulation of miR-206, which suppresses the expression of BDNF mRNA and protein [73].
Furthermore, mitochondrial dysfunction and oxidative stress contribute to SCZ pathogenesis [74,75,76,77]. In the blood of early psychosis patients, oxidative stress causes upregulation of the miR-137, resulting in decreased cytochrome c oxidase subunit 6A2 (COX6A2) and accumulation of impaired mitochondria [78]. Mitochondrial dysfunction was correlated with social behavioral impairment via the sequestration of gamma-aminobutyric acid (GABA) and lower GABAergic signaling [77].
Tan et al. recently showed that the expression of four circular RNAs (circRNAs) is modified in SCZ. These RNAs contain binding sites for miRNAs, such as miR-34a-5p and miR-499a, and take part in metabolic processes and histone ubiquitination, all of which have been associated with SCZ. Other circRNAs were also enriched in signaling pathways crucial for adult neurogenesis, such as the Notch and MAPK pathways [79,80,81].
4.2. Major Depressive Disorder
Major depressive disorder (MDD) is a mood disorder characterized by feelings of sadness, emptiness, or irritable mood, as well as cognitive and somatic changes. Although extensively studied, a large part of the pathophysiology of depression remains elusive. There is a need to identify new research targets, such as the exosomes and the exosomal miRNAs, whose levels are greatly dysregulated in depressed patients.
In depression, exosomes containing miRNAs target the serotoninergic system (SERT, 5HT1A, 5HT1D), while there is a consistent alteration in the levels of miRNAs that regulate the MAPK (Mitogen-activated protein kinase) and Wnt (Wingless-related integration site) pathways. Some miRNAs are shared with SCZ and target BDNF, Ataxin 1 (ATXN1), the glutamatergic system, and other signaling pathways [82]. miRNAs unique for depression pathophysiology are miR-451a, miR-1202, miR-135a, miR-425-3p, miR-34a-5p, miR-335-5p, miR-26a, miR-24-3p, miR-221-3p, miR-182, and let-7d [83,84,85,86,87,88,89,90,91].
Interestingly, mitomiRs are also involved in the pathophysiology of MDD. It is known that in states of cellular stress such as inflammation, mitochondrial genes are fragmented and released to the peripheral blood, resulting in circulating cell-free mitochondrial DNA (ccf-mtDNA) [92]. Ccf-mtDNA acts as a damage-associated molecular pattern (DAMP) in the plasma and triggers innate immunity [93]. Indeed, in untreated MDD patients, there was a positive correlation between the expression levels of five plasma miRNAs (hsa-miR-6068, hsa-miR-939-5p, hsa-miR-187-5p, hsa-miR7110-5p, and hsa-miR-4707-3p) with plasma ccf-mtDNA copy number, indicating a common mechanism of action [94]. These mitomiRs are hypothesized to elevate reactive oxygen species (ROS) and produce ccf-mtDNA by impairing cytochrome oxidase complexes I and III. However, additional research is required to confirm it.
4.3. Bipolar Disorder
Bipolar Disorder (BD) is a chronic mood disorder in which affected patients experience alternating depressive and manic episodes with intermittent periods of mood stability (euthymia). Recent evidence indicates that dysregulated miRNAs in BD target pathways such as neural signaling and synaptic plasticity, and there is promising evidence that exosomes, which primarily function as miRNA carriers, play a key role in the pathogenesis of BD. This is confirmed both by the observation that miRNA levels are altered in patients and by genetic research that has found MIR genes within loci of susceptibility for BD [95].
In their review, Fries et al. suggest that the most promising miRNAs in BD are miR-34a and miR-137 [95,96,97,98]. Ceylan et al., who performed miRNA sequencing of plasma exosomes from BD patients and healthy individuals, identified 13 aberrant miRNAs. Among them, the level of miR-185-5p was significantly increased, while the levels of miR-484, miR-652-3p, and miR142-3p were decreased [99]. MiR-484 takes part in regulatory pathways of neurogenesis, mitochondrial network, and redox modulations, processes that are closely related to the pathophysiology of BD [100,101]. Furthermore, miR-142–3p has an important signaling function during embryonic development, homeostasis, and disease. This miRNA, which has been shown to be dysregulated in exosomal samples of patients with BD, had previously been shown to be downregulated in unipolar depression patients [102,103,104]. Another down-regulated miRNA in the patients with BD was miR652–3p, which is involved in immune processes and oxidative stress mechanisms [82,105]. In addition, miR185–5p, the only upregulated miRNA, has a possible involvement in the regulation of the tyrosine kinase receptor type 2 [106], which has been related to anxiety, depression, and suicide [107].
4.4. Alzheimer Disease
Alzheimer's disease (AD), the most common type of dementia, is a neurodegenerative disorder characterized by progressive cognitive and functional decline related to age and accompanied by a unique neuropathology. Deposition of hyperphosphorylated Tau protein and APP fragment beta-amyloid (Aβ) results in synaptic loss and neuronal atrophy, mainly in the hippocampus and cerebral cortex [108]. Except for memory loss and impairment in executive functioning, AD involves a range of neuropsychiatric symptoms like apathy, disinhibition, agitation, and psychosis, which significantly impair behavior and mental health.
The role of exosomes in AD is dual, as they can have both neurotoxic and neuroprotective effects. On the one hand, exosomes induce apoptosis of neurons, as they take part in the secretion, transfer, and propagation of toxic Aβ and tau proteins, which are taken up by microglia and neurons. Also, Gao et al. showed that there is increased expression of glutaminase C (GAC) in the microglia of AD mice. GAC activates microglia by enhancing the secretion of exosomes and by changing the exosomal content to contain more pro-inflammatory miRNAs. [109]
On the other hand, the neural stem cells of the hippocampus produce exosomes that protect synapses against Aβ oligomer toxicity, restoring functions like memory and long-term potentiation. More specifically, exosomes absorb Aβ and transport neuroprotective substances between cells [110,111,112].
Impaired synaptic activity is one of the hallmarks of AD and mitochondria, and generating ATP and buffering calcium ions have a central role in neurotransmission [113]. Today, approximately 20 mitomiRs have been associated with AD. They are both upregulated and downregulated and participate in various mitochondrial functions. For example, the upregulation of miR-15a targets the β-site APP-cleaving enzyme 1 (BACE1) and unbalances mitochondrial potential, whereas the downregulation of miR-132 inhibits complex 1 of mitochondria, reducing aerobic respiration [114].
4.5. Huntington’s Disease
Huntington’s’s disease (HD) is an autosomal dominant neurodegenerative disorder that affects patients in the fourth to sixth decade of their life and is characterized by involuntary chorea-like movements, along with cognitive and behavioral impairment. In HD, there is a loss of medium spiny neurons of the CNS caused by the misfolding and successive intracellular accumulation of a mutant form of huntingtin (HTT). The huntingtin gene, located in the short arm of chromosome 4p16.3, contains a polymorphic CAG repeat, which becomes pathological if it comprises more than 35 CAG trinucleotide repeats [115]. The mutant range CAG repeats result in a polyglutamine tract in huntingtin, which is responsible for the toxicity of the protein [115]. The more the CAG repeats, the earlier the disease onset (116).
Exosomes mediate the progression of neurodegenerative diseases by integrating pathogenic proteins and RNAs into ILVs and releasing them in the extracellular environment [117]. In HD, the pathogenic protein is mutant huntingtin (mHTT), which is spread through cells via exosomes. This was shown by animal experiments that demonstrated the transmission of mHTT aggregates between cells [115,118] and by experiments of cytoplasmic aggregate formation [119].
Some miRNAs are commonly associated with HD. Among them, miR-9, which participates in transcriptional dysregulation, and miR-146a, which rescues cell cycle and apoptosis abnormalities and regulates HTT and TATA-binding protein (TBP) expression in cell culture, are down-regulated [120,121]. MiR-10b-5p, which physiologically regulates the expression of BDNF, miR-196a, which regulates CREB protein and peroxisome proliferator-activated receptor-γ coactivator-1α (PGC-1α) expression, and miR-214, which suppresses aggregates of mHTT and regulate mitofusin 2 (MFN2) expression in cell culture, are up-regulated [122,123].
Some mitomiRs may also be involved in HD by modulating mitochondrial dynamics, biogenesis, and mitophagy. Approximately 10 mitomiRs have been identified to be associated with HD, and most of them are downregulated. For example, the downregulation of miR128, which targets Fas-associated death domain (FADD) protein, prevents apoptosis in damaged mitochondria [114].
4.6. Critical View of Neuropsychiatric Diseases That Share Similarities in Alterations of miRNA Levels
In MDD, miRNAs, such as miR-146b-5p, involved in different MAPK and Wnt signaling pathways, have been observed to be consistently altered. Interestingly, the same miRNAs are altered in BD [82]. It is known that the MAPK pathway is involved in neurological processes like initiation of cortical neurogenesis and plasticity, while the Wnt pathway plays a crucial role in diverse stages of neurogenesis during early development and adulthood. The alteration of the levels of the same miRNAs for these pathways in MDD and BD confirms our knowledge that neurogenesis and plasticity are directly related to both of these mood disorders [82].
Both in MDD and SCZ there are alterations in the levels of the same miRNAs that target BDNF signaling pathways. The role of BDNF in the pathogenesis of MDD has been well-established, as it is associated with maladaptive neuroplasticity [124]. Meanwhile, human and animal studies have shown that BDNF is related to SCZ-like phenotypes, and that dysregulated BDNF signaling causes alterations in neurodevelopment. However, although a sub-threshold association exists, the BDNF locus has not been officially correlated with SCZ from genome-wide association studies. A better comprehension of BDNF signaling during development will possibly explain the role of this gene in SCZ.
Furthermore, miR-146a-5pa was up-regulated in both AD and HD. It has been proposed that in AD, miR-146a-5pa activates MAPK signaling, triggers oxidative stress, and, via this mechanism, increases Aβ deposition [125]. MiR-146a also targets human and mouse HTT genes, while decreased miR-146a expression increases the expression of Checkpoint Kinase 1 (CHEK1) and Cyclin A2 (CCNA2), which rescue the cell cycle. In addition, dysregulated miR-146a-5pa may contribute to HD pathogenesis by targeting TBP [123]. MiR-146a-5pa may also be involved in several other pathways, possibly common for AD and HD, a potentiality that might explain the amnesia-like and neuropsychiatric manifestations that appear in both of these CNS diseases.
5. Conclusion
Whereas the exosomes were first discovered in the 1980s, it was only over the past decade that they attracted the research interest of scientists and physicians. The field started to increase especially after the discovery of functional ncRNAs and miRNAs, and the fact that they exist in abundance in body fluids and cell culture media.
Today, we can conclude that:
(a) Exosomes constitute an evolutionarily conserved, universal mechanism of communication between cells,
(b) Exosomal miRNAs regulate a wide range of biological processes by binding to complementary sequences and modulating gene expression and mRNA translation,
(c) Exosomal miRNAs control the response to stress in neurons and glial cells by modifying their morphology and function in response to different stimuli [6,18]
(d) Exosomal miRNAs have been associated with neurodegeneration and psychopathology. They have crucial regulatory roles in the interface between genetic information, external (environmental) stimuli, and brain structure and function,
(e) The current knowledge of the exosomal ability to transfer genetic information between cells is cultivating a fertile ground for the design of therapeutic interventions to alleviate disease and,
(f) New discoveries regarding the role of miRNAs in the CNS promise to be unpredictable and astonishing, while they could help decode the very essence of life.
Author Contributions
M.C. conceived the study, researched the literature, designed the figures and authored the original draft of the manuscript. G.P.C., E.K. and D.V. thoroughly reviewed and revised the manuscript. C.Y. conceived the study, designed and supervised the study, as well as authored the final draft of the manuscript. All authors have reviewed and consented to the published version of the manuscript.
Funding
This research did not receive any funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Conflicts of Interest
It is declared by the authors that they have no conflicts of interest.
Abbreviations
| AD | Alzheimer’s disease |
| AHN | adult hippocampal neurogenesis |
| Alix protein | ALG2-interacting protein X |
| ATXN1 | Ataxin 1 |
| Aβ | beta-amyloid |
| BACE1 | β-site APP-cleaving enzyme 1 |
| BBB | Blood-Brain Barrier |
| BD | Bipolar disorder |
| BDNF | Brain-Derived Neurotrophic Factor |
| BMECs | Brain Macrovascular Endothelial Cells |
| C3 | complement component 3 |
| ccf-mtDNA | circulating cell-free mitochondrial DNA |
| CCNA2 | Cyclin A2 |
| CCR2 | C-C motif chemokine receptor 2 |
| CHEK1 | Checkpoint Kinase 1 |
| circRNAs | Circular RNAs |
| CNS | Central Nervous System |
| COX6A2 | Cytochrome c Oxidase Subunit 6A2 |
| CREB | cAMP-response element binding protein |
| CXCR4 | C-X-C chemokine receptor type 4 |
| DAMP | damage-associated molecular pattern |
| Drosha-DGCR8 | DiGeorge syndrome Critical Region gene 8 |
| ECT | Electron Transport Chain |
| ESCRT | Endosomal Sorting Complex Required for Transport |
| EV | Extracellular vesicle |
| FADD protein | Fas-associated death domain protein |
| GABA | Gamma-aminobutyric acid |
| GAC | glutaminase C |
| HD | Huntington’s’s disease |
| Hsp70 | heat-shock protein 70 |
| HTT | Huntingtin |
| IFNα | Interferon alpha |
| IL1R2 | Interleukin 1 receptor type II |
| IL-1β | Interleukin-1β |
| ILV | Intraluminal vesicle |
| KLF4 | Krüppel-like factor 4 |
| lncRNA | long non-coding RNA |
| MAP1b | microtubule-associated protein 1B |
| MAPK pathway | Mitogen-activated protein kinase |
| MDD | Major Depressive Disorder |
| MFN2 | mitofusin 2 |
| mHTT | mutant Huntingtin |
| miRNA | microRNA |
| MV | microvesicle |
| MVB | Multivesicular body |
| ncRNA | non-coding RNA |
| NF-Κb | Nuclear Factor kappa-light-chain-enhancer of activated B cells |
| NMDAR-C | N-methyl D-aspartate receptor C |
| OXPHOS | Oxidative Phosphorylation |
| P2RX7 | Purinergic Receptor P2X 7 |
| PGC-1α | peroxisome proliferator-activated receptor-γ coactivator-1α |
| PrP | Prion Protein |
| RISC | RNA-mediated silencing complex |
| ROS | reactive oxygen species |
| SCZ | Schizophrenia |
| SNARE protein | SNAP Receptor protein |
| TBP | TATA-binding protein |
| TNF-α | Tumor Necrosis Factor-alpha |
| TRBP | Transactivation-Responsive RNA Binding Protein |
| TSG101 | tumor susceptibility gene 101 protein |
| Wnt pathway | Wingless-related integration site |
References
- Lee Y, El Andaloussi S, Wood MJA. Exosomes and microvesicles: extracellular vesicles for genetic information transfer and gene therapy. Hum Mol Genet. 2012 Oct 15;21(R1):R125-134. [CrossRef]
- Zduriencikova M, Gronesova P, Cholujova D, Sedlak J. Potential biomarkers of exosomal cargo in endocrine signaling. Endocr Regul. 2015 Jul;49(3):141–50. [CrossRef]
- Théry C, Ostrowski M, Segura E. Membrane vesicles as conveyors of immune responses. Nat Rev Immunol. 2009 Aug;9(8):581–93. [CrossRef]
- Takahashi A, Okada R, Nagao K, Kawamata Y, Hanyu A, Yoshimoto S, et al. Exosomes maintain cellular homeostasis by excreting harmful DNA from cells. Nat Commun. 2017 May 16;8(1):15287.
- Isola AL, Chen S. Exosomes: The Messengers of Health and Disease. Curr Neuropharmacol. 2017;15(1):157–65.
- Makrygianni EA, Chrousos GP. Extracellular Vesicles and the Stress System. Neuroendocrinology. 2023;113(2):120–67.
- Zhang Y, Liu Y, Liu H, Tang WH. Exosomes: biogenesis, biologic function and clinical potential. Cell & Bioscience. 2019 Feb 15;9(1):19.
- Maligianni I, Yapijakis C, Bacopoulou F, Chrousos G. The Potential Role of Exosomes in Child and Adolescent Obesity. Children (Basel). 2021 Mar 6;8(3):196.
- Catalano M, O’Driscoll L. Inhibiting extracellular vesicles formation and release: a review of EV inhibitors. J Extracell Vesicles. 9(1):1703244.
- Vlachakis D, Mitsis Τ, Nicolaides N, Efthimiadou A, Giannakakis A, Bacopoulou F, et al. Functions, pathophysiology and current insights of exosomal endocrinology (Review). Molecular Medicine Reports. 2021 Jan 1;23(1):1–1.
- Simons M, Raposo G. Exosomes – vesicular carriers for intercellular communication. Current Opinion in Cell Biology. 2009 Aug 1;21(4):575–81.
- Budnik V, Ruiz-Cañada C, Wendler F. Extracellular vesicles round off communication in the nervous system. Nat Rev Neurosci. 2016 Mar;17(3):160–72.
- Kita S, Maeda N, Shimomura I. Interorgan communication by exosomes, adipose tissue, and adiponectin in metabolic syndrome. J Clin Invest. 2019 Oct 1;129(10):4041–9. [CrossRef]
- Urbanelli L, Magini A, Buratta S, Brozzi A, Sagini K, Polchi A, et al. Signaling Pathways in Exosomes Biogenesis, Secretion and Fate. Genes (Basel). 2013 Mar 28;4(2):152–70. [CrossRef]
- Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol. 2013 Feb 18;200(4):373–83. [CrossRef]
- Gallo A, Tandon M, Alevizos I, Illei GG. The Majority of MicroRNAs Detectable in Serum and Saliva Is Concentrated in Exosomes. PLOS ONE. 2012 Mar 9;7(3):e30679.
- Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004 Jan 23;116(2):281–97.
- Yapijakis C. Regulatory Role of MicroRNAs in Brain Development and Function. Adv Exp Med Biol. 2020;1195:237–47.
- Sohel MH. Extracellular/Circulating MicroRNAs: Release Mechanisms, Functions and Challenges. Achievements in the Life Sciences. 2016 Dec 1;10(2):175–86.
- Cheng L, Sharples RA, Scicluna BJ, Hill AF. Exosomes provide a protective and enriched source of miRNA for biomarker profiling compared to intracellular and cell-free blood. J Extracell Vesicles. 2014;3.
- Catalanotto C, Cogoni C, Zardo G. MicroRNA in Control of Gene Expression: An Overview of Nuclear Functions. IJMS. 2016 Oct 13;17(10):1712.
- Kim VN, Han J, Siomi MC. Biogenesis of small RNAs in animals. Nat Rev Mol Cell Biol. 2009 Feb;10(2):126–39.
- Mouillet-Richard S, Baudry A, Launay JM, Kellermann O. MicroRNAs and depression. Neurobiology of Disease. 2012 May 1;46(2):272–8.
- Gregory RI, Shiekhattar R. MicroRNA Biogenesis and Cancer. Cancer Research. 2005 May 2;65(9):3509–12. [CrossRef]
- Valencia-Sanchez MA, Liu J, Hannon GJ, Parker R. Control of translation and mRNA degradation by miRNAs and siRNAs. Genes Dev. 2006 Mar 1;20(5):515–24. [CrossRef]
- Kosaka N, Iguchi H, Hagiwara K, Yoshioka Y, Takeshita F, Ochiya T. Neutral Sphingomyelinase 2 (nSMase2)-dependent Exosomal Transfer of Angiogenic MicroRNAs Regulate Cancer Cell Metastasis*. Journal of Biological Chemistry. 2013 Apr 12;288(15):10849–59. [CrossRef]
- Frank F, Sonenberg N, Nagar B. Structural basis for 5′-nucleotide base-specific recognition of guide RNA by human AGO2. Nature. 2010 Jun;465(7299):818–22.
- Brites D, Fernandes A. Neuroinflammation and Depression: Microglia Activation, Extracellular Microvesicles and microRNA Dysregulation. Front Cell Neurosci. 2015;9:476.
- Nowak JS, Michlewski G. miRNAs in development and pathogenesis of the nervous system. Biochem Soc Trans. 2013 Aug;41(4):815–20.
- Guo H, Ingolia NT, Weissman JS, Bartel DP. Mammalian microRNAs predominantly act to decrease target mRNA levels. Nature. 2010 Aug;466(7308):835–40.
- Gao W, Lu X, Liu L, Xu J, Feng D, Shu Y. MiRNA-21: a biomarker predictive for platinum-based adjuvant chemotherapy response in patients with non-small cell lung cancer. Cancer Biol Ther. 2012 Mar;13(5):330–40.
- Altick AL, Baryshnikova LM, Vu TQ, von Bartheld CS. Quantitative Analysis of Multivesicular Bodies (MVBs) in the Hypoglossal Nerve: Evidence that Neurotrophic Factors do not use MVBs for Retrograde Axonal Transport. J Comp Neurol. 2009 Jun 20;514(6):641–57.
- Glebov K, Löchner M, Jabs R, Lau T, Merkel O, Schloss P, et al. Serotonin stimulates secretion of exosomes from microglia cells. Glia. 2015 Apr;63(4):626–34.
- Lachenal G, Pernet-Gallay K, Chivet M, Hemming FJ, Belly A, Bodon G, et al. Release of exosomes from differentiated neurons and its regulation by synaptic glutamatergic activity. Mol Cell Neurosci. 2011 Feb;46(2):409–18.
- Luarte A, Cisternas P, Caviedes A, Batiz LF, Lafourcade C, Wyneken U, et al. Astrocytes at the Hub of the Stress Response: Potential Modulation of Neurogenesis by miRNAs in Astrocyte-Derived Exosomes. Stem Cells Int. 2017;2017:1719050.
- Zheng T, Pu J, Chen Y, Guo Z, Pan H, Zhang L, et al. Exosomes Secreted from HEK293-APP Swe/Ind Cells Impair the Hippocampal Neurogenesis. Neurotox Res. 2017 Jul;32(1):82–93.
- Cheng LC, Pastrana E, Tavazoie M, Doetsch F. miR-124 regulates adult neurogenesis in the subventricular zone stem cell niche. Nat Neurosci. 2009 Apr;12(4):399–408.
- Rajasethupathy P, Fiumara F, Sheridan R, Betel D, Puthanveettil SV, Russo JJ, et al. Characterization of Small RNAs in Aplysia Reveals a Role for miR-124 in Constraining Synaptic Plasticity through CREB. Neuron. 2009 Sep 24;63(6):803–17. [CrossRef]
- Szulwach KE, Li X, Smrt RD, Li Y, Luo Y, Lin L, et al. Cross talk between microRNA and epigenetic regulation in adult neurogenesis. J Cell Biol. 2010 Apr 5;189(1):127–41. [CrossRef]
- Le MTN, Xie H, Zhou B, Chia PH, Rizk P, Um M, et al. MicroRNA-125b Promotes Neuronal Differentiation in Human Cells by Repressing Multiple Targets. Molecular and Cellular Biology. 2009 Oct 1;29(19):5290–305. [CrossRef]
- Schratt GM, Tuebing F, Nigh EA, Kane CG, Sabatini ME, Kiebler M, et al. A brain-specific microRNA regulates dendritic spine development. Nature. 2006 Jan;439(7074):283–9.
- Bátiz LF, Castro MA, Burgos PV, Velásquez ZD, Muñoz RI, Lafourcade CA, et al. Exosomes as Novel Regulators of Adult Neurogenic Niches. Front Cell Neurosci. 2015;9:501.
- Lafourcade C, Ramírez JP, Luarte A, Fernández A, Wyneken U. MiRNAs in Astrocyte-Derived Exosomes as Possible Mediators of Neuronal Plasticity. J Exp Neurosci. 2016;10(Suppl 1):1–9.
- Goldie BJ, Dun MD, Lin M, Smith ND, Verrills NM, Dayas CV, et al. Activity-associated miRNA are packaged in Map1b-enriched exosomes released from depolarized neurons. Nucleic Acids Res. 2014 Aug;42(14):9195–208.
- Beveridge NJ, Gardiner E, Carroll AP, Tooney PA, Cairns MJ. Schizophrenia is associated with an increase in cortical microRNA biogenesis. Mol Psychiatry. 2010 Dec;15(12):1176–89.
- Santarelli DM, Beveridge NJ, Tooney PA, Cairns MJ. Upregulation of dicer and microRNA expression in the dorsolateral prefrontal cortex Brodmann area 46 in schizophrenia. Biol Psychiatry. 2011 Jan 15;69(2):180–7. [CrossRef]
- Bahrini I, Song J hoon, Diez D, Hanayama R. Neuronal exosomes facilitate synaptic pruning by up-regulating complement factors in microglia. Sci Rep. 2015 Jan 23;5(1):7989. [CrossRef]
- Castrén E, Rantamäki T. The role of BDNF and its receptors in depression and antidepressant drug action: Reactivation of developmental plasticity. Developmental Neurobiology. 2010;70(5):289–97. [CrossRef]
- Frühbeis C, Fröhlich D, Kuo WP, Amphornrat J, Thilemann S, Saab AS, et al. Neurotransmitter-Triggered Transfer of Exosomes Mediates Oligodendrocyte–Neuron Communication. PLOS Biology. 2013 Jul 9;11(7):e1001604.
- Guitart K, Loers G, Buck F, Bork U, Schachner M, Kleene R. Improvement of neuronal cell survival by astrocyte-derived exosomes under hypoxic and ischemic conditions depends on prion protein. Glia. 2016 Jun;64(6):896–910.
- Wang S, Cesca F, Loers G, Schweizer M, Buck F, Benfenati F, et al. Synapsin I Is an Oligomannose-Carrying Glycoprotein, Acts As an Oligomannose-Binding Lectin, and Promotes Neurite Outgrowth and Neuronal Survival When Released via Glia-Derived Exosomes. J Neurosci. 2011 May 18;31(20):7275–90.
- Sbai O, Ould-Yahoui A, Ferhat L, Gueye Y, Bernard A, Charrat E, et al. Differential vesicular distribution and trafficking of MMP-2, MMP-9, and their inhibitors in astrocytes. Glia. 2010;58(3):344–66.
- Taylor AR, Robinson MB, Gifondorwa DJ, Tytell M, Milligan CE. Regulation of heat shock protein 70 release in astrocytes: Role of signaling kinases. Developmental Neurobiology. 2007;67(13):1815–29.
- Kato T. Neurobiological basis of bipolar disorder: Mitochondrial dysfunction hypothesis and beyond. Schizophrenia Research. 2017 Sep 1;187:62–6. [CrossRef]
- Rosolen D, Nunes-Souza E, Marchi R, Tofolo MV, Antunes VC, Berti FCB, et al. MiRNAs Action and Impact on Mitochondria Function, Metabolic Reprogramming and Chemoresistance of Cancer Cells: A Systematic Review. Biomedicines. 2023 Feb 24;11(3):693. [CrossRef]
- Macgregor-Das AM, Das S. A microRNA’s journey to the center of the mitochondria. American Journal of Physiology-Heart and Circulatory Physiology. 2018 Aug;315(2):H206–15.
- Duarte FV, Palmeira CM, Rolo AP. The Emerging Role of MitomiRs in the Pathophysiology of Human Disease. Adv Exp Med Biol. 2015;888:123–54.
- Pearce WJ. Mitochondrial influences on smooth muscle phenotype. American Journal of Physiology-Cell Physiology. 2024 Feb;326(2):C442–8.
- Chrousos GP. Stress and disorders of the stress system. Nat Rev Endocrinol. 2009 Jul;5(7):374–81.
- Gayen M, Bhomia M, Balakathiresan N, Knollmann-Ritschel B. Exosomal MicroRNAs Released by Activated Astrocytes as Potential Neuroinflammatory Biomarkers. Int J Mol Sci. 2020 Mar 27;21(7):2312.
- Fan C, Li Y, Lan T, Wang W, Long Y, Yu SY. Microglia secrete miR-146a-5p-containing exosomes to regulate neurogenesis in depression. Molecular Therapy. 2022 Mar 2;30(3):1300–14.
- Wei ZX, Xie GJ, Mao X, Zou XP, Liao YJ, Liu QS, et al. Exosomes from patients with major depression cause depressive-like behaviors in mice with involvement of miR-139-5p-regulated neurogenesis. Neuropsychopharmacol. 2020 May;45(6):1050–8.
- Frühbeis C, Fröhlich D, Kuo WP, Krämer-Albers EM. Extracellular vesicles as mediators of neuron-glia communication. Front Cell Neurosci. 2013 Oct 30;7:182.
- Dozio V, Sanchez JC. Characterisation of extracellular vesicle-subsets derived from brain endothelial cells and analysis of their protein cargo modulation after TNF exposure. J Extracell Vesicles. 2017;6(1):1302705.
- Maes M. The cytokine hypothesis of depression: inflammation, oxidative & nitrosative stress (IO&NS) and leaky gut as new targets for adjunctive treatments in depression. Neuro Endocrinol Lett. 2008 Jun;29(3):287–91.
- Dalvi P, Sun B, Tang N, Pulliam L. Immune activated monocyte exosomes alter microRNAs in brain endothelial cells and initiate an inflammatory response through the TLR4/MyD88 pathway. Sci Rep. 2017 Aug 30;7(1):9954.
- Saeedi S, Israel S, Nagy C, Turecki G. The emerging role of exosomes in mental disorders. Transl Psychiatry. 2019 Mar 28;9(1):1–11.
- Xian X, Cai LL, Li Y, Wang RC, Xu YH, Chen YJ, et al. Neuron secrete exosomes containing miR-9-5p to promote polarization of M1 microglia in depression. Journal of Nanobiotechnology. 2022 Mar 9;20(1):122.
- Baudry A, Mouillet-Richard S, Schneider B, Launay JM, Kellermann O. miR-16 targets the serotonin transporter: a new facet for adaptive responses to antidepressants. Science. 2010 Sep 17;329(5998):1537–41. [CrossRef]
- Beveridge NJ, Tooney PA, Carroll AP, Gardiner E, Bowden N, Scott RJ, et al. Dysregulation of miRNA 181b in the temporal cortex in schizophrenia. Hum Mol Genet. 2008 Apr 15;17(8):1156–68. [CrossRef]
- Kahn RS, Sommer IE, Murray RM, Meyer-Lindenberg A, Weinberger DR, Cannon TD, et al. Schizophrenia. Nat Rev Dis Primers. 2015 Nov 12;1:15067.
- Borroto-Escuela DO, Tarakanov AO, Bechter K, Fuxe K. IL1R2, CCR2, and CXCR4 May Form Heteroreceptor Complexes with NMDAR and D2R: Relevance for Schizophrenia. Front Psychiatry. 2017;8:24.
- Du Y, Yu Y, Hu Y, Li XW, Wei ZX, Pan RY, et al. Genome-Wide, Integrative Analysis Implicates Exosome-Derived MicroRNA Dysregulation in Schizophrenia. Schizophrenia Bulletin. 2019 Oct 24;45(6):1257–66.
- Fraguas D, Díaz-Caneja CM, Ayora M, Hernández-Álvarez F, Rodríguez-Quiroga A, Recio S, et al. Oxidative Stress and Inflammation in First-Episode Psychosis: A Systematic Review and Meta-analysis. Schizophr Bull. 2019 Jun 18;45(4):742–51.
- Goetzl EJ, Srihari VH, Guloksuz S, Ferrara M, Tek C, Heninger GR. Neural cell-derived plasma exosome protein abnormalities implicate mitochondrial impairment in first episodes of psychosis. FASEB J. 2021 Feb;35(2):e21339. [CrossRef]
- Rajasekaran A, Venkatasubramanian G, Berk M, Debnath M. Mitochondrial dysfunction in schizophrenia: pathways, mechanisms and implications. Neurosci Biobehav Rev. 2015 Jan;48:10–21. [CrossRef]
- Kanellopoulos AK, Mariano V, Spinazzi M, Woo YJ, McLean C, Pech U, et al. Aralar Sequesters GABA into Hyperactive Mitochondria, Causing Social Behavior Deficits. Cell. 2020 Mar 19;180(6):1178-1197.e20. [CrossRef]
- Khadimallah I, Jenni R, Cabungcal JH, Cleusix M, Fournier M, Beard E, et al. Mitochondrial, exosomal miR137-COX6A2 and gamma synchrony as biomarkers of parvalbumin interneurons, psychopathology, and neurocognition in schizophrenia. Mol Psychiatry. 2022 Feb;27(2):1192–204.
- Tan G, Wang L, Liu Y, Zhang H, Feng W, Liu Z. The alterations of circular RNA expression in plasma exosomes from patients with schizophrenia. J Cell Physiol. 2021 Jan;236(1):458–67.
- He K, Guo C, Guo M, Tong S, Zhang Q, Sun H, et al. Identification of serum microRNAs as diagnostic biomarkers for schizophrenia. Hereditas. 2019 Jun 27;156(1):23.
- Xu C, Zhang Y, Zheng H, Loh HH, Law PY. Morphine modulates mouse hippocampal progenitor cell lineages by upregulating miR-181a level. Stem Cells. 2014 Nov;32(11):2961–72.
- Gruzdev SK, Yakovlev AA, Druzhkova TA, Guekht AB, Gulyaeva NV. The Missing Link: How Exosomes and miRNAs can Help in Bridging Psychiatry and Molecular Biology in the Context of Depression, Bipolar Disorder and Schizophrenia. Cell Mol Neurobiol. 2019 Aug;39(6):729–50.
- Issler O, Chen A. Determining the role of microRNAs in psychiatric disorders. Nat Rev Neurosci. 2015 Apr;16(4):201–12.
- Fiori LM, Lopez JP, Richard-Devantoy S, Berlim M, Chachamovich E, Jollant F, et al. Investigation of miR-1202, miR-135a, and miR-16 in Major Depressive Disorder and Antidepressant Response. Int J Neuropsychopharmacol. 2017 Jun 6;20(8):619–23.
- Camkurt MA, Acar Ş, Coşkun S, Güneş M, Güneş S, Yılmaz MF, et al. Comparison of plasma MicroRNA levels in drug naive, first episode depressed patients and healthy controls. J Psychiatr Res. 2015 Oct;69:67–71.
- Wan Y, Liu Y, Wang X, Wu J, Liu K, Zhou J, et al. Identification of Differential MicroRNAs in Cerebrospinal Fluid and Serum of Patients with Major Depressive Disorder. PLOS ONE. 2015 Mar 12;10(3):e0121975. [CrossRef]
- Kuang WH, Dong ZQ, Tian LT, Li J. MicroRNA-451a, microRNA-34a-5p, and microRNA-221-3p as predictors of response to antidepressant treatment. Braz J Med Biol Res. 2018;51(7):e7212. [CrossRef]
- Lopez JP, Fiori LM, Cruceanu C, Lin R, Labonte B, Cates HM, et al. MicroRNAs 146a/b-5 and 425-3p and 24-3p are markers of antidepressant response and regulate MAPK/Wnt-system genes. Nat Commun. 2017 May 22;8:15497. [CrossRef]
- Enatescu VR, Papava I, Enatescu I, Antonescu M, Anghel A, Seclaman E, et al. Circulating Plasma Micro RNAs in Patients with Major Depressive Disorder Treated with Antidepressants: A Pilot Study. Psychiatry Investig. 2016 Sep;13(5):549–57.
- Capitano F, Camon J, Licursi V, Ferretti V, Maggi L, Scianni M, et al. MicroRNA-335-5p modulates spatial memory and hippocampal synaptic plasticity. Neurobiol Learn Mem. 2017 Mar;139:63–8.
- Li Y, Li S, Yan J, Wang D, Yin R, Zhao L, et al. miR-182 (microRNA-182) suppression in the hippocampus evokes antidepressant-like effects in rats. Prog Neuropsychopharmacol Biol Psychiatry. 2016 Feb 4;65:96–103.
- Chiu RWK, Chan LYS, Lam NYL, Tsui NBY, Ng EKO, Rainer TH, et al. Quantitative Analysis of Circulating Mitochondrial DNA in Plasma. Clinical Chemistry. 2003 May 1;49(5):719–26.
- Zhang M, Zhang B, Guo Y, Zhang L, Yang S, Yin L, et al. Alteration of circulating mitochondrial DNA concentration after irradiation. Adv Exp Med Biol. 2013;765:371–7.
- Ogata H, Higasa K, Kageyama Y, Tahara H, Shimamoto A, Takekita Y, et al. Relationship between circulating mitochondrial DNA and microRNA in patients with major depression. Journal of Affective Disorders. 2023 Oct 15;339:538–46.
- Fries GR, Carvalho AF, Quevedo J. The miRNome of bipolar disorder. J Affect Disord. 2018 Jun;233:110–6.
- Bavamian S, Mellios N, Lalonde J, Fass DM, Wang J, Sheridan SD, et al. Dysregulation of miR-34a links neuronal development to genetic risk factors for bipolar disorder. Mol Psychiatry. 2015 May;20(5):573–84.
- Azevedo JA, Carter BS, Meng F, Turner DL, Dai M, Schatzberg AF, et al. The microRNA network is altered in anterior cingulate cortex of patients with unipolar and bipolar depression. J Psychiatr Res. 2016 Nov;82:58–67.
- Mokhtari MA, Sargazi S, Saravani R, Heidari Nia M, Mirinejad S, Hadzsiev K, et al. Genetic Polymorphisms in miR-137 and Its Target Genes, TCF4 and CACNA1C, Contribute to the Risk of Bipolar Disorder: A Preliminary Case-Control Study and Bioinformatics Analysis. Dis Markers. 2022 Sep 22;2022:1886658.
- Ceylan D, Tufekci KU, Keskinoglu P, Genc S, Özerdem A. Circulating exosomal microRNAs in bipolar disorder. J Affect Disord. 2020 Feb 1;262:99–107.
- Wang K, Long B, Jiao JQ, Wang JX, Liu JP, Li Q, et al. miR-484 regulates mitochondrial network through targeting Fis1. Nat Commun. 2012 Apr 17;3(1):781.
- Andreazza AC, Young LT. The neurobiology of bipolar disorder: identifying targets for specific agents and synergies for combination treatment. Int J Neuropsychopharmacol. 2014 Jul;17(7):1039–52.
- Shrestha A, Mukhametshina RT, Taghizadeh S, Vásquez-Pacheco E, Cabrera-Fuentes H, Rizvanov A, et al. MicroRNA-142 is a multifaceted regulator in organogenesis, homeostasis, and disease. Dev Dyn. 2017 Apr;246(4):285–90.
- Fries GR, Lima CNC, Valvassori SS, Zunta-Soares G, Soares JC, Quevedo J. Preliminary investigation of peripheral extracellular vesicles’ microRNAs in bipolar disorder. J Affect Disord. 2019 Aug 1;255:10–4.
- Smalheiser NR, Lugli G, Rizavi HS, Torvik VI, Turecki G, Dwivedi Y. MicroRNA expression is down-regulated and reorganized in prefrontal cortex of depressed suicide subjects. PLoS One. 2012;7(3):e33201.
- Gilicze AB, Wiener Z, Tóth S, Buzás E, Pállinger É, Falcone FH, et al. Myeloid-derived microRNAs, miR-223, miR27a, and miR-652, are dominant players in myeloid regulation. Biomed Res Int. 2014;2014:870267.
- Maussion G, Yang J, Yerko V, Barker P, Mechawar N, Ernst C, et al. Regulation of a truncated form of tropomyosin-related kinase B (TrkB) by Hsa-miR-185* in frontal cortex of suicide completers. PLoS One. 2012;7(6):e39301.
- Kohli MA, Salyakina D, Pfennig A, Lucae S, Horstmann S, Menke A, et al. Association of Genetic Variants in the Neurotrophic Receptor–Encoding Gene NTRK2 and a Lifetime History of Suicide Attempts in Depressed Patients. Arch Gen Psychiatry. 2010 Apr;67(4):348–59.
- Soria Lopez JA, González HM, Léger GC. Alzheimer’s disease. Handb Clin Neurol. 2019;167:231–55.
- Gao G, Zhao S, Xia X, Li C, Li C, Ji C, et al. Glutaminase C Regulates Microglial Activation and Pro-inflammatory Exosome Release: Relevance to the Pathogenesis of Alzheimer’s Disease. Front Cell Neurosci. 2019;13:264.
- Losurdo M, Pedrazzoli M, D’Agostino C, Elia CA, Massenzio F, Lonati E, et al. Intranasal delivery of mesenchymal stem cell-derived extracellular vesicles exerts immunomodulatory and neuroprotective effects in a 3xTg model of Alzheimer’s disease. Stem Cells Translational Medicine. 2020 Sep 1;9(9):1068–84.
- Jahangard Y, Monfared H, Moradi A, Zare M, Mirnajafi-Zadeh J, Mowla SJ. Therapeutic Effects of Transplanted Exosomes Containing miR-29b to a Rat Model of Alzheimer’s Disease. Front Neurosci. 2020;14:564.
- Qi Y, Guo L, Jiang Y, Shi Y, Sui H, Zhao L. Brain delivery of quercetin-loaded exosomes improved cognitive function in AD mice by inhibiting phosphorylated tau-mediated neurofibrillary tangles. Drug Delivery. 2020 Jan 1;27(1):745–55.
- Cai Q, Tammineni P. Mitochondrial Aspects of Synaptic Dysfunction in Alzheimer’s Disease. Journal of Alzheimer’s Disease. 2017;57(4):1087–103.
- John A, Kubosumi A, Reddy PH. Mitochondrial MicroRNAs in Aging and Neurodegenerative Diseases. Cells. 2020 May 28;9(6):1345.
- Herrera F, Tenreiro S, Miller-Fleming L, Outeiro TF. Visualization of cell-to-cell transmission of mutant huntingtin oligomers. PLoS Curr. 2011 Feb 11;3:RRN1210.
- Yapijakis C. Ancestral Concepts of Human Genetics and Molecular Medicine in Epicurean Philosophy. In: Petermann HI, Harper PS, Doetz S, editors. History of Human Genetics: Aspects of Its Development and Global Perspectives [Internet]. Cham: Springer International Publishing; 2017 [cited 2023 Sep 14]. p. 41–57. [CrossRef]
- Vella LJ, Sharples RA, Nisbet RM, Cappai R, Hill AF. The role of exosomes in the processing of proteins associated with neurodegenerative diseases. Eur Biophys J. 2008 Mar 1;37(3):323–32. [CrossRef]
- Pearce MMP, Spartz EJ, Hong W, Luo L, Kopito RR. Prion-like transmission of neuronal huntingtin aggregates to phagocytic glia in the Drosophila brain. Nat Commun. 2015 Apr 13;6(1):6768. [CrossRef]
- Ren PH, Lauckner JE, Kachirskaia I, Heuser JE, Melki R, Kopito RR. Cytoplasmic penetration and persistent infection of mammalian cells by polyglutamine aggregates. Nat Cell Biol. 2009 Feb;11(2):219–25. [CrossRef]
- Packer AN, Xing Y, Harper SQ, Jones L, Davidson BL. The Bifunctional microRNA miR-9/miR-9* Regulates REST and CoREST and Is Downregulated in Huntington’s’s Disease. J Neurosci. 2008 Dec 31;28(53):14341–6.
- Sinha M, Ghose J, Das E, Bhattarcharyya NP. Altered microRNAs in STHdhQ111/HdhQ111 cells: miR-146a targets TBP. Biochemical and Biophysical Research Communications. 2010 Jun 4;396(3):742–7.
- Hoss AG, Labadorf A, Latourelle JC, Kartha VK, Hadzi TC, Gusella JF, et al. miR-10b-5p expression in Huntington’s’s disease brain relates to age of onset and the extent of striatal involvement. BMC Medical Genomics. 2015 Mar 1;8(1):10.
- Dong X, Cong S. MicroRNAs in Huntington’s’s Disease: Diagnostic Biomarkers or Therapeutic Agents? Front Cell Neurosci. 2021 Aug 6;15:705348.
- Yang T, Nie Z, Shu H, Kuang Y, Chen X, Cheng J, et al. The Role of BDNF on Neural Plasticity in Depression. Front Cell Neurosci. 2020 Apr 15;14:82.
- Zhan-Qiang H, Hai-Hua Q, Chi Z, Miao W, Cui Z, Zi-Yin L, et al. miR-146a aggravates cognitive impairment and Alzheimer disease-like pathology by triggering oxidative stress through MAPK signaling. Neurologia (Engl Ed). 2021 Mar 11;S0213-4853(21)00022-0.
Figure 1.
Biogenesis of exosomes: (1) the cell membrane invaginates and forms the early endosome, (2) the early endosome is loaded with its cargo, which can be DNA, RNA, or proteins, (3) the multivesicular body is formed, (4) the multivesicular body is either degraded by lysosomes or fuses with the cell membrane and releases its intracavitary vesicles in the extracellular space. Once released, the intracavitary vesicles are named exosomes.
Figure 1.
Biogenesis of exosomes: (1) the cell membrane invaginates and forms the early endosome, (2) the early endosome is loaded with its cargo, which can be DNA, RNA, or proteins, (3) the multivesicular body is formed, (4) the multivesicular body is either degraded by lysosomes or fuses with the cell membrane and releases its intracavitary vesicles in the extracellular space. Once released, the intracavitary vesicles are named exosomes.
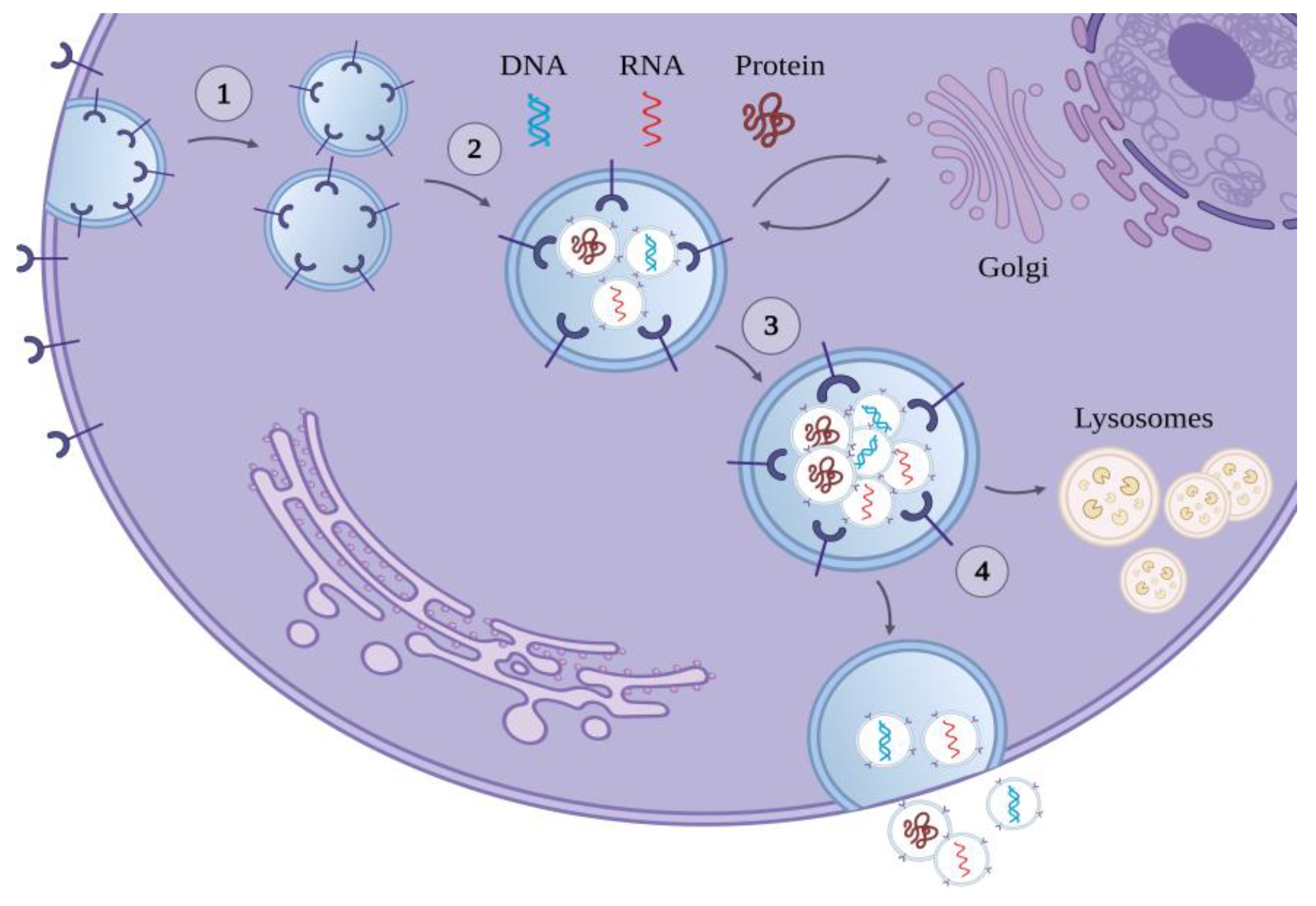
Figure 2.
Nucleic and cytoplasmic actions of miRNAs: In the nucleus, the miRNA-RISC complex binds with complementary sequences of promoters or enhancers and causes transcriptional repression (a) or activation (b, c). In the cytoplasm, the binding of the miRNA-RISC complex to the target mRNA can result in (a) cleavage of the 5’ cap of the target mRNA and reduced stability, (b) deadenylation and subsequent degradation of the target mRNA, (c) detachment of the ribosome and inhibition of the translation process.
Figure 2.
Nucleic and cytoplasmic actions of miRNAs: In the nucleus, the miRNA-RISC complex binds with complementary sequences of promoters or enhancers and causes transcriptional repression (a) or activation (b, c). In the cytoplasm, the binding of the miRNA-RISC complex to the target mRNA can result in (a) cleavage of the 5’ cap of the target mRNA and reduced stability, (b) deadenylation and subsequent degradation of the target mRNA, (c) detachment of the ribosome and inhibition of the translation process.
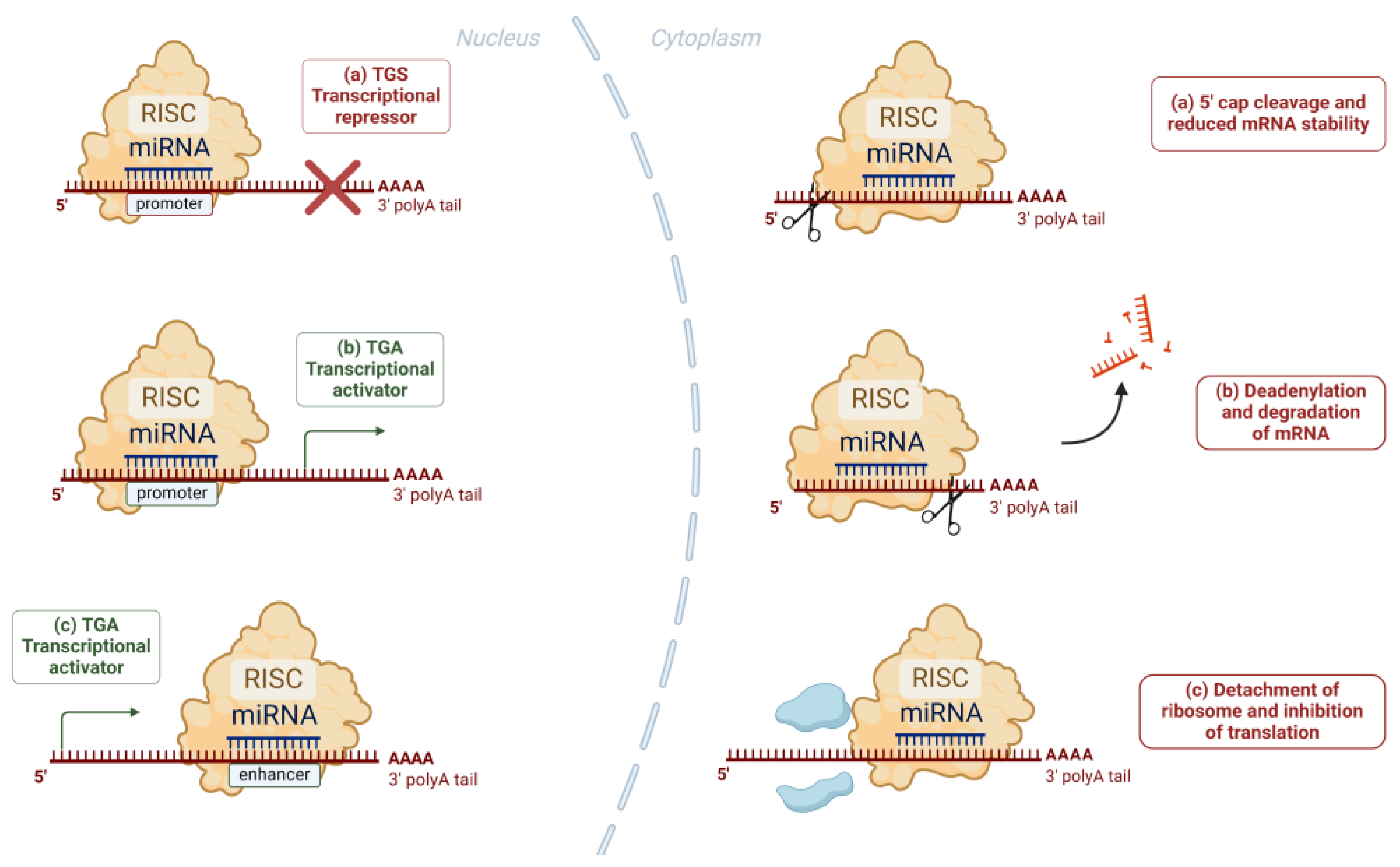
Figure 3.
Biogenesis of miRNA: (1) RNA polymerase II transcribes the miRNA gene into pri-miRNA, (2) RNase III endonuclease, Drosha, cleaves the pri-miRNA, creating the pre-miRNA, (3) pre-miRNA is exported to the cytoplasm, (4) the cytoplasmic RNase III enzyme, Dicer and the TAR RNA-binding protein (TRBP) converts the pre-miRNA into double-stranded miRNA, (5) the mature miRNA binds to RNA-induced silencing complex (RISC) and then gets unchained by the Argonaute 2 protein (AGO2), (6) the mature miRNA remains bound to RISC and binds complementary with one strand to the target mRNA, while the other strand is hydrolyzed, (7) the interaction of the miRNA with the mRNA results in mRNA degradation or translational repression.
Figure 3.
Biogenesis of miRNA: (1) RNA polymerase II transcribes the miRNA gene into pri-miRNA, (2) RNase III endonuclease, Drosha, cleaves the pri-miRNA, creating the pre-miRNA, (3) pre-miRNA is exported to the cytoplasm, (4) the cytoplasmic RNase III enzyme, Dicer and the TAR RNA-binding protein (TRBP) converts the pre-miRNA into double-stranded miRNA, (5) the mature miRNA binds to RNA-induced silencing complex (RISC) and then gets unchained by the Argonaute 2 protein (AGO2), (6) the mature miRNA remains bound to RISC and binds complementary with one strand to the target mRNA, while the other strand is hydrolyzed, (7) the interaction of the miRNA with the mRNA results in mRNA degradation or translational repression.
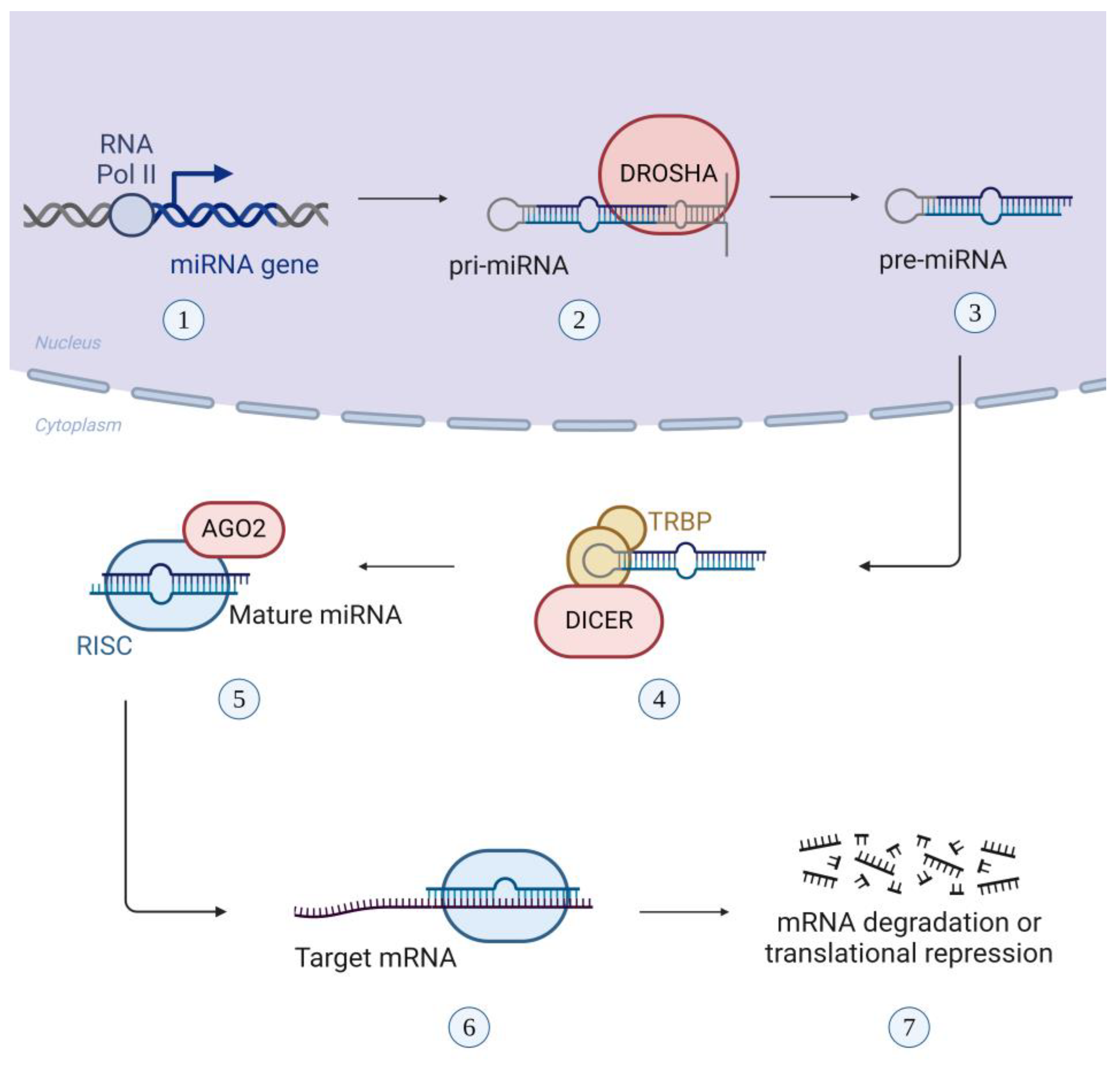

Table 1.
MicroRNA (miRNA) that have been documented to exhibit increased (↑) or decreased (↓) expression levels in Alzheimer’s disease (AD), Huntington’s disease (HD), Bipolar disorder (BD), Major depressive disorder (MDD), and Schizophrenia (SCZ).
Table 1.
MicroRNA (miRNA) that have been documented to exhibit increased (↑) or decreased (↓) expression levels in Alzheimer’s disease (AD), Huntington’s disease (HD), Bipolar disorder (BD), Major depressive disorder (MDD), and Schizophrenia (SCZ).
| Disease | miRNA | Alteration |
|---|---|---|
| Alzheimer’s disease | miR-15a | ↑ [114] |
| miR-132 | ↓ [114] | |
| miR-146a-5pa | ↑ [125] | |
| Huntington’s disease | miR-9 | ↓ [120] |
| miR-10b-5p | ↑ [122] | |
| miR-128 | ↓ [114] | |
| miR-146a | ↓ [121] | |
| miR-196a | ↑ [123] | |
| miR-214 | ↑ [123] | |
| Bipolar Disorder | miR-34a | ↑ [96]/ ↓ [97] |
| miR-137 | ↑ [98] | |
| miR-142-3p | ↓ [99] | |
| miR-185-5p | ↑ [99] | |
| miR-484 | ↓ [99] | |
| miR-652-3p | ↓ [99] | |
| Major Depressive Disorder | let-7d | ↑ [86] |
| miR-24-3p | ↑ [88] | |
| miR-26a | ↑ [89] | |
| miR-34a-5p | ↑ [86] | |
| miR-135a | ↓ [83] | |
| miR-182 | ↑ [91] | |
| miR-187-5p | Positive correlation with ccf-mtDNA copy number [94] | |
| miR-221-3p | ↑ [86] | |
| miR-335-5p | ↑ [90] | |
| miR-425-3p | ↑ [88] | |
| miR-451a | ↑ [85] | |
| miR-939-5p | Positive correlation with ccf-mtDNA copy number [94] | |
| miR-1202 | ↓ [84] | |
| miR-4707-3p | Positive correlation with ccf-mtDNA copy number [94] | |
| miR-6068 | Positive correlation with ccf-mtDNA copy number [94] | |
| miR-7110-5p | Positive correlation with ccf-mtDNA copy number [94] | |
| Schizophrenia | miR-34a-5p + miR-499a | ↑ [80] |
| miR-137 | ↑ [78] | |
| miR-206 | ↑ [73] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



