
Submitted:
27 May 2024
Posted:
28 May 2024
You are already at the latest version
Abstract
Predictive studies are essential to establish fundamental data for monitoring and controlling Xylella fastidiosa infections in our country. Limited knowledge exists about how Xylella fastidiosa interacts with the plant host's immune system, leaving numerous unanswered questions regarding Xylella fastidiosa biology and its interactions with national plant hosts. Key inquiries include understanding of how the plant immune system interfaces with Xylella fastidiosa in the xylem and identifying the specific or shared mechanisms underlying the susceptibility of different hosts. Factors such as the presence of insect vectors, abundance of host plants, and climate suitability play pivotal roles in Xylella fastidiosa dispersal. We recommend that countries identified with relatively high risk, like Portugal, undertake thorough individual risk analyses. The implementation of preventive measures and, if necessary, the enhancement of surveillance systems for early detection of Xylella fastidiosa in plants and insect vectors are crucial steps. Additionally, raising awareness is of equal importance to mitigate socioeconomic and ecological impacts on their ecosystems.
Keywords:
Philaenus spumarius
; olive quick decline syndrome
; citrus variegated chlorosis
; bacterial leaf scorch
; pierce disease
1. Introduction
The introduction of invasive species can have severe effects on the economy and ecosystems, leading to significant financial burdens. This is especially noteworthy in the agricultural sector, which is highly susceptible to the infiltration of pathogenic organisms. These invasions can lead to a decline in food supply, increased prices, compromised food quality, and potential concerns regarding food safety and security [1]. It is crucial to evaluate the consequences of invasive plant pathogens in terms of their impact on agriculture and also by considering the broader range of services provided by the agroecological system in its landscape context [2]. Invasive plant pests and diseases, like Xylella fastidiosa (Xf), can have a profound impact on the ecosystems by significantly disrupting its flow.
Xf is an aerobic bacterium[3], transmitted through xylem-sucking insects [4], that multiplies in the xylem [5], causing occlusion of xylem vessels and water and nutrient stress. It has been considered a quarantine pest, for affecting economically important agricultural crops [6]. It is imperative to understand the real incidence of this bacterium and the corresponding risk factors, which are, to date, poorly understood.
This plant pathogen, originally from the Americas, has been detected in various European and Near Eastern countries [2]. The initial outbreak of this pathogen in the European Union was identified in olive trees in Italy’s Apulia region, in 2013 [7,8]. Subsequent occurrences of the bacterium have been reported in France, starting with an outbreak in Corsica in 2015, followed by Provence in 2019 and Occitania in 2020 [9]. Those countries have reported infected coffee plants that came from Latin American countries such as Mexico, Ecuador, Costa Rica, and Honduras [10], indicating that the global spread of this pathogen was expanding due to the exchange of commodities and plant materials.
The initial outbreak of the plant pathogen in Spain occurred in the Balearic Islands in 2016 [11,12]. This outbreak involved three subspecies of the bacterium, affecting more than 20 plant species such as grapevine, almond, olive, and fig [13]. By December 2020, over 600 olive trees in the Balearic archipelago were confirmed to be infected. As the situation progressed into 2021, the affected area expanded, covering 2292 hectares, and resulting in the destruction of over 100,000 almond trees [6].
The detection of Xylella fastidiosa in Portugal occurred through prospection, in 2019 in Vila Nova de Gaia, Porto, in plants of Lavandula dentata L. [14]. Currently, in Portugal there are 18 Xf demarcated zones (DZ), as we can see in Figure 1, and 1 deleted DZ in Tavira in the Region of Algarve [15].
In Porto, once the presence of the bacterium was confirmed in 2019, measures were immediately taken to prevent its spread and ensure eradication [16]. Following the detection of Xf, a “Demarcated Zone” was promptly established. This zone encompasses the “Infected Zone”, which includes all susceptible plants within a 50m radius around the contaminated plants. Additionally, there is a surrounding “Buffer Zone” extending to a radius of 2.5 km [14]. Subsequently a series of control or eradication measures were implemented. These measures included uprooting infected trees, prohibiting the planting of susceptible plant species, restricting the transportation of plants from the infected areas, as well as conducting vector control through vegetation removal and the application of chemical treatments [17].
Following the guidelines stipulated in Implementing Regulation (EU) 2020/1201, all European countries must prospect for quarantine pests [16]. The objective of prospecting for Xf is primarily geared towards identifying the presence of this pest. This is achieved through the inspection and collection of samples, followed by the implementation of control or eradication measures [2].
The General Directorate of Food and Veterinary (DGAV), as the National Phytosanitary Authority, is tasked with defining and coordinating the nationwide survey for Xf. As part of the program, several key components are incorporated. These include identifying priority sites that require monitoring, establishing specific timelines for observations and sampling, compiling a comprehensive list of host plants that need to be monitored, describing the symptomatology associated with Xf infections, and establishing procedures for collecting samples for laboratory analysis [14,18]. This responsibility extends to various entities involved in the prospecting program. These entities include the Regional Directorates for Agriculture and Fisheries (DRAP), the Directorates of Agriculture of the Autonomous Regions of Madeira and Azores (DRA), and the Institute of Conservation of Nature and Forests (ICNF) [19]. Collaboratively, they work together to ensure the effective implementation of the survey across Portugal. The critical task of analyzing these samples in the laboratory is entrusted to the National Institute for Agrarian and Veterinary Research (INIAV). They play a vital role in providing accurate and reliable results for further assessment and action [19].
The main objective of this study is to gain a comprehensive understanding of this problem within the defined DZs in the North Region. The focus is on analyzing the available prospection data from DRAP North, since 2019 to June 2023, within these zones. By doing so, we aim to improve our knowledge about the host biology, dispersion of symptoms, control methods, and associated risks of those Xf outbreaks in our country. This information will serve as a foundation for formulating hypotheses for further investigation. The study findings will be valuable in assisting public authorities and private entities, in making informed decisions and taking decisive actions related to this issue. Moreover, this preliminary assessment will contribute to the current decision-making process regarding the planning and execution of control operations, providing valuable insights for future data-driven evaluations.
2. Materials and Methods
In this study we will analyze DRAP North prospection data, since 2019 to June of 2023, from DZ of Oporto Metropolitan Area; DZ Baião and DZ Bougado; and from several DZ of the interior North part of the country, including Sabrosa; Alijó; Mirandela; Mirandela II. This work comprehends two phases: the prospection phase on Figure 2 (inspecting plants, sampling and submission of samples to the laboratory) and the research phase (collecting and organizing data and statistical treatment).
2.1. Prospection Phase
Prospecting for Xf includes, inspecting and carrying out sample collection and in the event of their presence, implement control or eradication measures. The first step includes identifying the places to be prospected, the times when observations and sample collections are carried out, the list of host plants subject to observation, the description of symptoms and the establishment of sample collection procedures for analysis. This procedure is subject to detailed instructions prepared by DGAV. Following the EFSA guidelines for territorial prospecting based on statistical data (Guidelines for statistically sound and risk-based surveys of Xf), the number of plants to be sampled is determined for each area with homogeneous characteristics, using the statistical tool RIBESS+. Thus, resorting to the land use classification of the Corine Land Cover database, the territory is subdivided into four classes with epidemiological significance regarding Xf- agricultural, urban, forestry and semi-natural - to obtain areas with homogeneous characteristics. For each of these areas, the number of host plants per hectare is estimated to calculate the size of the target population subject to prospecting, from which the number of plants to be sampled through RIBESS+ is determined.
After DGAV determined the places to be prospected and the number of sample collections, DRAP North can implement the reinforcement. DRAP North prospect with observations of symptoms and collect samples for laboratory analysis whenever suspicious symptoms were observed and in plants without symptoms. Then all prospected plants and insects must be georeferenced and registered by each prospector on the GESFITO platform for coding.
2.1.1. Sample Collection
In asymptomatic and symptomatic plants, we collected all biological material in the four quadrants of the host plant; 2 branches or 5 branches with 8-10 leaves, respectively. The parts of plants that make up the sample were previously shaken, wrapped in newsprint paper and placed in a plastic bag, carefully closed to protect the dispersion of possible insect vectors during transport.
Then, after sample coding, the samples were sent, within a maximum period of 24 hours after collection, directly to INIAV Plant Health Laboratory.
2.1.2. Results Reception
Once the presence of the bacteria is confirmed by INIAV this result is communicated to DGAV and measures are immediately taken to prevent its spread and ensure eradication. To ensure the enforcement and adherence to these measures, the national phytosanitary authority (DGAV), pursuant to Decree-Law No. 67/2020 of 15 September, designates the Demarcated Zone (DZ), outlining the eradication measures for the bacteria and the restrictions on the movement of plants intended for planting within the Infected Zone and Buffer Zone [18,19]. Within this DZ, destruction measures are subsequently established and executed by DRAPN (Figure 3):
- In situ destruction of infected plants and other plants of the same species.
- In situ destruction of all plants listed in Annexes I and II of Implementing Regulation (EU) 2020/1201.
2.2. Collecting, Organizing and Statistical Treatment
Following the laboratory results, data from DZ in Área Metropolitana do Porto, DZ Sabrosa, DZ Alijó, DZ Baião, DZ Mirandela, DZ Mirandela II, and DZ Bougado were compiled. Subsequently, we selected variables for statistical analysis, including the number of locations observed per district, types of locations observed, presence of symptoms, and observed plant hosts. All selected variables underwent statistical analysis using the JMP statistical program.
3. Results and Discussion
3.1. Number of Locations Observed
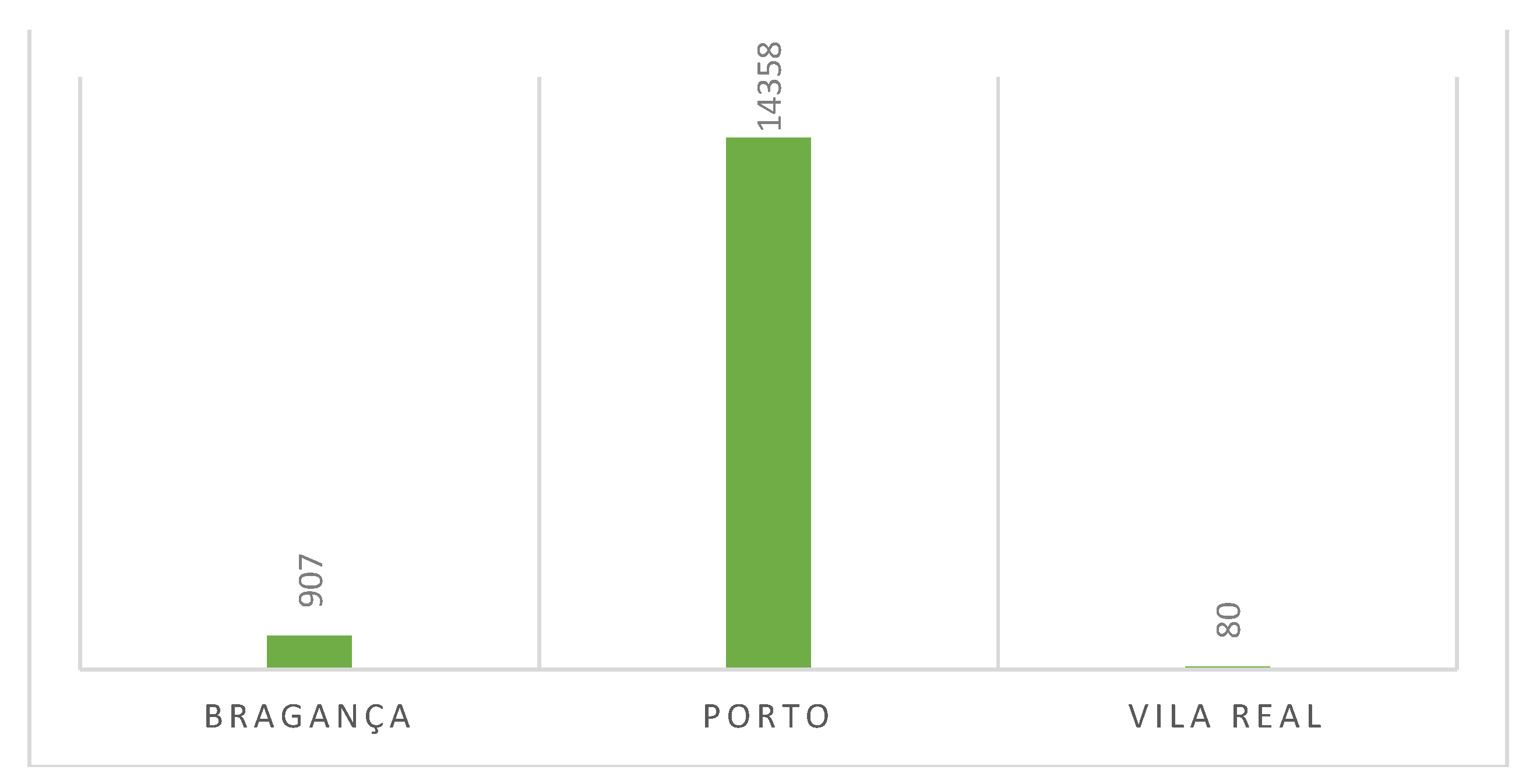
Bar Scheme 1 presents the count of locations surveyed from 2019 to June 2023. In the Porto district, there are the DZ of Área Metropolitana do Porto, DZ Baião, and DZ Bougado. In the Bragança district, you’ll find DZ Mirandela and DZ Mirandela II. Lastly, in the Vila Real district, there are DZ Sabrosa and DZ Alijó. Notably, 14,358 out of 15,345 locations were examined in Porto (93.6%). This discrepancy can be attributed to the establishment of the first DZ of Porto, in 2019. The regions of Bragança (5.9%) and Vila Real (0.5%) began sample observations in 2022, reflecting a lower number of observations.
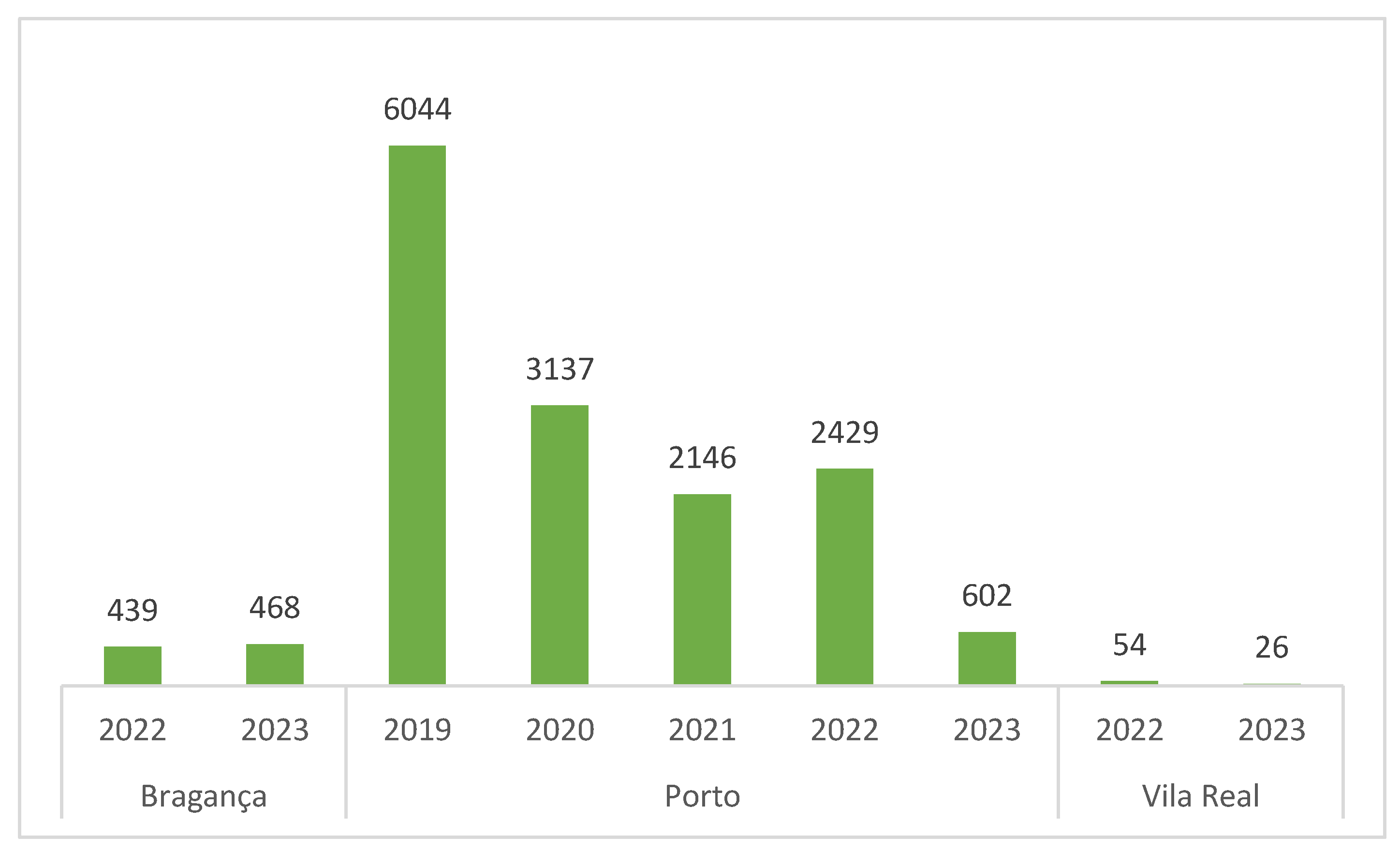
The bars of Scheme 2 illustrate a general decrease in the number of prospected points in the DZ over the years. This reduction can be attributed to several factors. In 2020, a weekly limit on the collection of plant material samples was imposed, leading to a consequently decrease in the number of observed locations and collected samples. Additionally, during the first quarter of 2021, in locations marked for prospecting, only georeferencing/survey of addresses was feasible due to restrictions on entering residents’ gardens and backyards amid Covid-19 confinement. In 2023, the authorization for prospecting was granted late (in August), contributing to the diminished number of collected data.
3.2. Type of Locations Observed
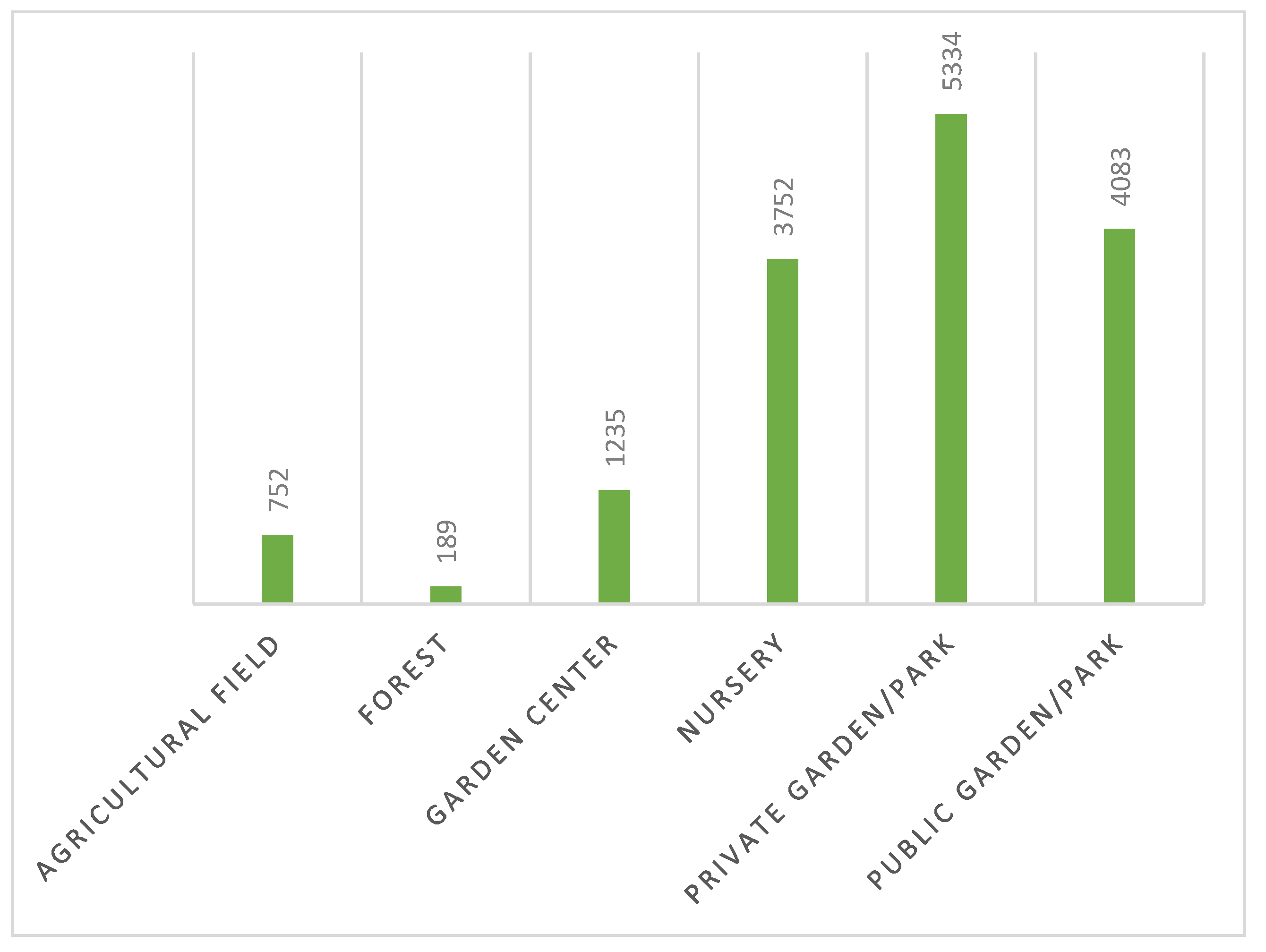
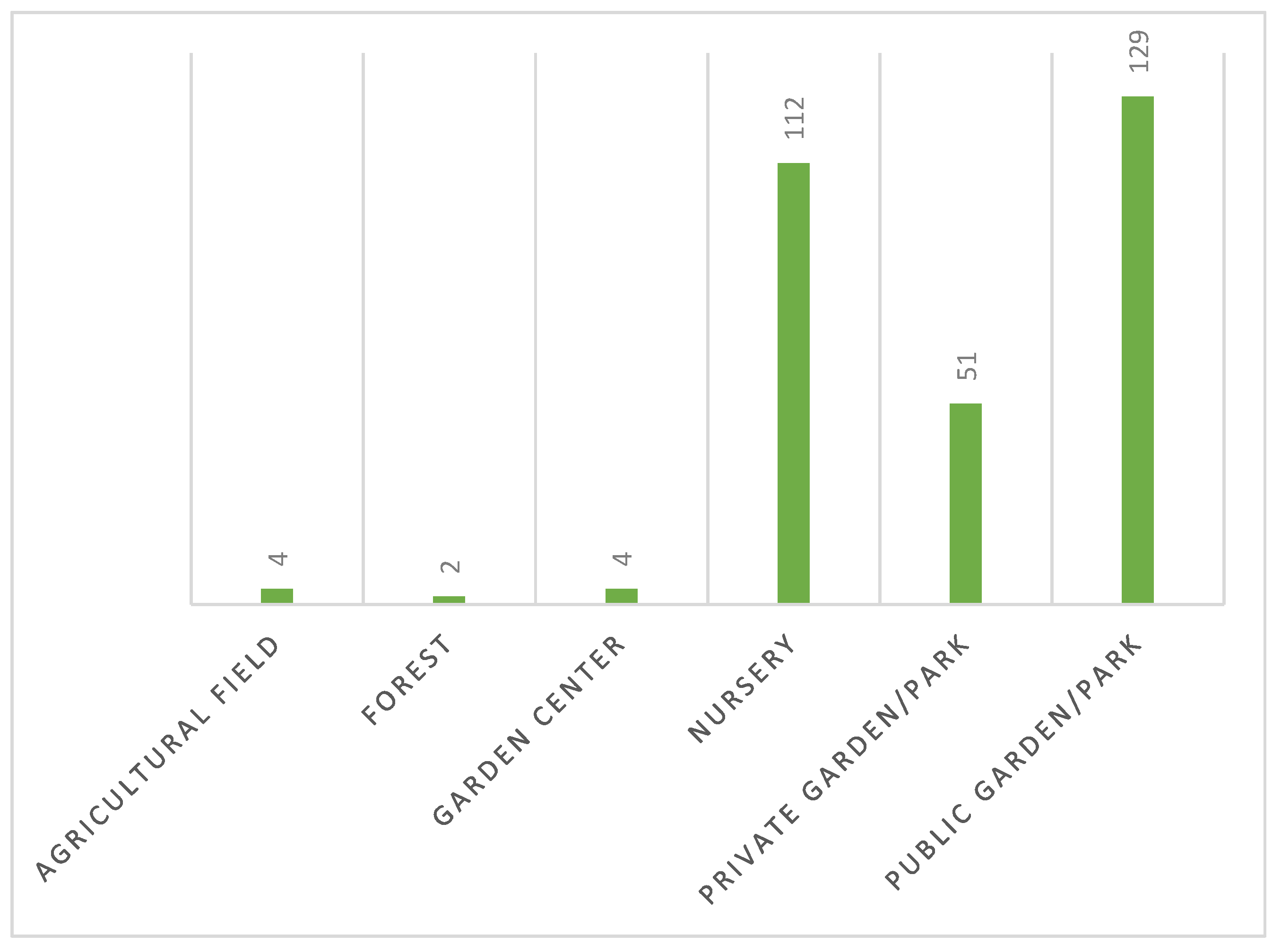
Scheme 3 illustrates the various types of locations prospected in the North of Portugal since 2019. The majority of prospections were carried out in private gardens/parks (34.8%), followed by public gardens/parks (26.6%) and nurseries (24.5%). Garden centers, agricultural fields, and forests constituted a limited sample of observations. This distribution could be explained by the fact that since a DZ was stablished, it is necessary to identify the places to be prospected. This procedure is subject to detailed instructions prepared by DGAV. Following the EFSA guidelines for territorial prospecting based on statistical data (Guidelines for statistically sound and risk-based surveys of Xf), the number of plants to be sampled is determined for each area with homogeneous characteristics, using the statistical tool RIBESS+. Thus, resorting to the land use classification of the Corine Land Cover database, the territory is subdivided into four classes with epidemiological significance with regard to Xf- agricultural, urban, forestry and semi-natural - in order to obtain areas with homogeneous characteristics. For each of these areas, the number of host plants per hectare is estimated to calculate the size of the target population subject to prospecting, from which the number of plants to be sampled through RIBESS+ is determined [15]. Subsequently, DGAV identifies the locations designated for prospection, assigning priorities to areas such as nurseries, private gardens, and public gardens. Adhering to these guidelines, DRAP North implements reinforcement measures, resulting in the majority of prospected results in these locations.
Upon analyzing Scheme 4, it becomes apparent that public gardens (42.7%) accounted for the majority of positive plant results, followed by nurseries (37.1%) and private gardens (16.9%).
These observations can be attributed to two main factors: the higher number of observations (or samples) potentially leading to an increased number of positive results, and the fact that, when delineating DZ and prospecting plants within a 5km radius of positive findings, public garden/parks are commonly included in the DZ. This inclusion tends to result in a higher number of positive outcomes. Also, these locations often feature herbaceous vegetation, creating shade and humidity that serve as ideal conditions for the development of Philaenus spumarius, one of the main vectors of Xf. In nurseries, a notable occurrence of plants positive for Xf has been observed (37.1%). Nurseries engage in the buying and selling of annual plants, perennial plants and trees, and related products, encompassing activities like cultivating plants for sale and conducting transactions with customers. Due to their nature, nurseries can pose as potential sources for the dissemination of plant diseases, acting as reservoirs for pathogens such as Xf. The sale or distribution of infected plants within this context can significantly contribute to the spread of diseases. The current phytosanitary regulations and inspection systems in some parts of the region may have vulnerabilities, potentially allowing the transport of infected planting materials. This raises the possibility of Xf invading new districts, with the transport infrastructure inadvertently facilitating the movement of Xf insect vectors. Additionally, the skepticism surrounding the efficacy of eradication measures by Economic Operators, the financial burden of analysis costs on economic operators, and the delayed submission of analytical results to the Economic Operators (leading to economic impact) contribute to plant transactions occurring without official controls and the consequent positive results.
Consequently, a genuine risk exists that Xf could continue to breach geographical barriers and potentially triggering significant disease outbreaks [20].
3.3. Presence of Symptoms
On Bar Scheme 5, the presence of symptoms in the prospected plants is depicted. It can be inferred that the majority of samples did not exhibit any symptoms (91.1%) of bacterial presence.
After conducting the chi-square test to analyze the relationship between the species variable and the presence or absence of symptoms (Table 1), we determined a statistically significant association (p < 0.0001). This suggests that the occurrence and expression of symptoms are influenced by distinctive factors characteristic of the Portuguese Northern region hosts.
Although infection in certain host species leads to noticeable visual changes, there are several species in which these bacteria establish colonization without producing any symptoms [21]. In the context of North Portugal, it was observed that the majority of positive reports for Xf did not exhibit any symptoms (255 cases) on Citrus, Lavandula, Olea europaea and Prunus. In contrast, 45 positive reports did show symptoms on Lavandula dentata. It seems that Xf infection in perennial plants has the potential to evolve into a chronic state and endure over extended periods. The longevity of the bacterium within perennial plants is contingent upon factors such as the specific host species, cultivars [22,23,24], bacteria strain [25], time of infection [23,26] and winter temperatures.
Furthermore, the timing of infection can impact the presence or absence of symptoms. Plants infected early in the growing season were more likely to develop persistent infections and exhibit symptoms. This is because the pathogen had sufficient time to reproduce and spread within the plant’s perennial tissues, allowing it to endure through the winter [23,26]. Xf was previously reported to translocate and colonize the roots of perennial plants such as olive [27,28] almond [29], peach [30,31] blueberry [32], sycamore [33] and citrus [34,35]. As trees transition into dormancy, there is a decrease in the presence and concentration of Xf within their leaves. This decline can be ascribed to leaf senescence, bacterial mortality, and the migration of viable cells into perennial tissues, where they endure the winter season [36]. Consequently, these factors likely contribute to the presence or absence of symptoms. These results are in accordance with the report of Amanifar et al. (2016) in almond leaves [29]. Upon the emergence of leaves in the spring, overwintering bacterial cells initiate their migration from perennial tissues towards the developing leaves. Substantial colonization of the leaves by Xf typically occurs approximately two months after leaf emergence, and the pathogen’s concentration continues to rise over time, reaching its peak in mid to late summer, coinciding with the period when foliar symptoms become most pronounced.
This pattern may clarify the absence of symptoms in our research findings. Among the 255 positive reports of Xf that didn’t exhibit any symptoms, 170 were collected in the first quarter of the year. According to the authors, Xf migrated into perennial tissues, causing the consequent absence of symptoms.
Additionally, Purcell (2013) suggested that infection or colonization by Xf does not invariably lead to disease development. Xf can effectively colonize its hosts to ensure its survival and facilitate transmission through vectors. This pathogen can also mitigate harm to the host by modulating its gene expression in response to external signals.[37]. Bragard et al. (2019) reported that the duration of the asymptomatic period following infection can differ significantly, spanning from 1 month in ornamental plants to as long as 3-4 years in other hosts. [38]. Queiroz-Voltan et al. (2004) observed that symptom severity varied even within the same variety grown under identical soil and climate conditions, cultural practices, and management [39]. Hopkins et al. (2007) and Kadel et al. (2017) discovered that less infectious subspecies of the bacteria provide vine protection against more aggressive subspecies, all the while exhibiting reduced symptom manifestation [40,41]. On a recent investigation on Quercus pyrenaica, the xylem vessels composed with intact pit membranes and fully coated with lignin, made them more restrictive to bacterial migration. Considering the oak’s limited manifestation of symptoms and the identification of a solitary bacterium, the authors conclude that the formation of tyloses may hinder the pathogen’s dissemination from the initial point(s) of introduction. Also, the presence of starch grains in stems may acted as a refilling mechanism, preventing the loss of plant hydraulic conductivity and potentially serving as a means to entrap the bacteria. All of these mechanisms can be considered examples of constitutive defense systems of the plant against Xf.
It’s apparent that external symptoms aren’t solely determined by the presence or absence of Xf. The way symptoms respond and manifest is shaped by a multitude of physiological and environmental factors. These include variances in plant resistance or tolerance, fluctuating levels of bacterial concentration, diverse transpiration rates, and the occlusion capacity across different plant subspecies. Nevertheless, the presence of perennial plants in the Northern region of Portugal may account for our findings, as Xf infection in perennial plants have the capacity to persist in a chronic state over prolonged periods without exhibiting symptoms.
3.4. Hosts Observed
The initial step in organizing the observed host species and preparing the data for statistical analysis was to group all species with fewer than 10 observations into a single category, referred to others as “outros” (Table 2).
Our findings (listed on Table 2) indicated that, in a universe of 15345 samples, the most frequently prospected host genders or species included Olea europeae (5.5%), Citrus (2.7%), Laurus nobilis (2.5%), Rosa spp. (2.3%), Nerium oleander L. (2.2%), Pelargonium sp. (2.1%), Hedera helix L. (2.1%), and Lavandula dentata L. (2.1%). This observation aligns with the directives from DGAV, which emphasize the prospection of species listed in Annex I and II of Implementing Regulation (EU) 2020/1201, including the aforementioned species [16].
Analyzing the positive plant species in Table 3, it is evident that Lavandula dentata (18.9%), Hebe (2.0%), Citrus (1.7%), Lavandula angustifolia (1.7%), and Olea europaea L. (1.7%) were the most commonly observed species with positive results.
Our results are not surprising because all those species are vulnerable to infection and can develop diseases caused by these bacteria. The plant genera or species referred are identified as susceptible to subspecies of the bacteria in Annex I or II on Commission Implementing Regulation (EU) 2020/1201 [16].
On the other hand, Citrus sinensis and Vitis vinifera exhibited limited positive results, despite being also susceptible species. This outcome can be attributed to the genetic characteristics of the identified subspecies. In Portugal is commonly found the prevailing Sequence Type 7 of Xf subsp. multiplex (in Santolina chamaecyparissus, Quercus suber, Lavandula dentata, Hebe sp., Coprosma repens, Cytisus scoparius) [42]. The Sequence Type with a closer affinity to Citrus belongs to Xf subsp. pauca [43] never found in our country, while for Vitis vinifera, belongs to Xf subsp. fastidiosa [44].
According to data from EFSA’s 2023 report, there are currently 690 plant species that have been identified as susceptible to the bacteria, corresponding to 306 genus and 88 families [42]. The most common host species are Olea europaea, Vitis vinífera, Prunus dulcis, Citrus sinensis, Nerium oleander and Polygala myrtifolia. In comparison with the previous database, published in December 2022, 12 new species (and two genera) have been identified as Xf hosts, 9 of them were identified naturally infected in Portugal (Castanea sativa, Genista triacanthos, Grevillea rosmarinifolia, Lonicera periclymenum, Mentha suaveolens, Platanus hispanica, Pyracantha coccínea, Quercus pyrenaica, Ulex micranthus) [6,42].
The list of all known plant species that have been identified in Portugal are explained on Table 4 [45].
It seems that introgression events and its inherent ability to assimilate new genetic material have enabled Xf to expand its host range in our country [42,46]. However, also trophic interactions between insect vectors and their host plants can shape the epidemiology of Xf in North of Portugal. To understand the increase of Xf one must consider the behavior of the insect vectors.
Xf spreads naturally among plants through insect vectors from the order Hemiptera, primarily involving cycads, aphrophids, and cercopids [15]. Among the key European insect vectors are members of the Aphrophoridae family, such as Philaenus spumarius (Ps), Philaenus italosignus, and Neophilaenus campestris [5,47]. These insects are widely distributed in tropical and temperate climates, inhabiting forests, meadows, and grasslands [48,49,50]. They acquire the bacteria while feeding on infected plants, becoming hosts themselves. Subsequently, they transmit the pathogen into the vascular system of host plants by piercing the leaf petiole with their stylets [46], facilitating the spread of bacteria into the xylem of branches and stems [5].
These insects undergo a biological cycle wherein they overwinter as eggs. Following a dormancy period of around 100 days, the eggs hatch in early spring. The nymphs then progress through five developmental stages over 5-8 weeks, all the while enveloped in a protective mucilaginous foam [51]. Adults usually emerge in April or May and engage in mating during early summer, after which they tend to linger in the nearby vegetation [48]. During spring, nymphs are typically found in weeds, while adults predominantly inhabit the canopy from May throughout the summer months. Come autumn, the adults retreat to the weeds within the plot and surrounding areas, or migrate to other nearby plants [4]. According to Drosopoulos and Asche (1991) in Greece [52], Godefroid and Durán (2022) [53] in southern Spain, and Karban and Strauss (2004) in California [54] humidity and temperature exerts a significant influence on the developmental duration of immature Philaenus spumarius stages [55]. Cooler and humid conditions seem to be more advantageous for egg hatching and larval development with the temperature ranging between 4 and 10°C [55,56]. Adequate humidity is conducive to both hatching and the survival of nymphs, as they rely on an ample sap supply for nourishment. Consequently, severe or prolonged summer droughts can significantly diminish Philaenus spumarius populations [54,57]. This phenomenon may account for the transition in host plants, as the insect shifts from arid vegetation to shrubs and trees that are less water-stressed or migrates to cooler locales, such as areas near rivers. This underscores the significant influence of humidity and temperature in the ecological dynamics of this insect [57].
In southern Italy, for instance, the summer migration of adult insects from ground-level vegetation to the canopy has been noted in conjunction with the desiccation of herbaceous plants [49,58]. In particular, adults of Philaenus spumarius were observed from May to August in the vicinity of olive orchards, and on groves of Quercus ilex, Quercus crenata, Pistacia lentiscus, P. terebinthus, Hedera helix and Myrtus communis [49,59]. In the Apulia region, Philaenus spumarius adults have also been found on Pinus halepensis from May to September and have been reported once on Ulmus minor in October [59].
The nymphal stages of Philaenus spumarius exhibits a preference for specific plant species, primarily those belonging to families such as Asteraceae (Sonchus, Crepis, Picris), Fabaceae (Medicago, Vicia, Lathyrus), Rubiaceae (Galium), and Apiaceae (Daucus, Foeniculum), with G. album and F. vulgare being the favored species. On the other hand, certain species were categorized as “non-preferred” or actively avoided, including those from the Poaceae family (Avena, Lolium, Hordeum), Brassicaceae (Raphanus), Papaveraceae (Papaver), Fumariaceae (Fumaria), and Oxalidaceae (Oxalis) [60]. On the other hand, in Belgium, Hasbroucq et al. (2020), reported adults of Philaenus spumarius on Ranunculaceae, Prunus sp., Rubus sp., Crataegus sp., Alnus sp., Picea sp., Quercus sp., Salix sp., Carex sp. and Urtica sp. [61]. Recent studies performed in Corsica, France, suggest that Philaenus spumarius, at least locally, has affinity with Cistus monspeliensis [62,63]. Chatois et al. (2003), highlighted a positive relationship between Ps abundance and the density of Cistus monspeliensis. They also found larger populations of Ps in cooler and moister plots [58]. Lester et al. observed that among the various botanical families, those with the highest occurrence of host plants harboring Philaenus spumarius nymphs included Lamiaceae (10%), Rosaceae (10%), Onagraceae (9%), and Poaceae (9%). Lavender (Lavandula sp.) emerged as the most frequently encountered host plant in gardens, while in unmanaged areas like waste grounds, parks, and natural areas, grasses (Poaceae) were the predominant hosts. Additionally, adult Philaenus spumarius were also collected from trees and hedges, such as birch (Betula sp.), hawthorn (Crataegus sp.), and willow (Salix sp.) during the month of August [64].
Several studies have explored host associations of the insect vector in olive groves in Italy [65,66], Portugal [67], Spain[68] and Greece [69].
Philaenus spumarius may have limited flight ranges, confining the spread of the disease to a localized area as well as specific host preferences for feeding. Variations in these preferences can impact the range of plant species that may serve as hosts for Xf, influencing the types of ecosystems and agricultural crops that are at greater risk of infection. If a vector has a limited flight range and prefers specific host plants, the disease may be prevalent in certain localized areas but constrained from spreading to distant regions. The interplay of these vector behaviors directly impacts the geographic range of Xf.
In addition to the insect vector, it is recognized that each subspecies of the bacteria also exhibits a preference for a specific group of hosts. However, their genetic plasticity allows them to infect other plant species to some extent [14]. The taxonomic classification of Xf strains is a crucial step for identifying susceptible hosts. Additionally, in Europe, it holds practical significance as the management strategies mandated by the European Commission are tailored to the specific subspecies present in each outbreak.
The techniques employed for detecting the bacterium and determining its sequence type (ST) in plants align with the guidelines outlined in the EPPO protocol PM7/244 [10]. Briefly, the detection is performed using the real-time PCR and the identification of the ST by applying an Multilocus Sequence Typing (MLST) scheme of seven housekeeping genes and using the PubMLST database to identify allele numbers and ST [10,70].
Europe has reported four subspecies of the pathogen (Xyllela fastidiosa subsp. fastidiosa, multiplex, sandyi, and pauca) [42]. The initial outbreak in the European Union was detected in 2013, affecting olive trees near Gallipoli, located in the Lecce province of Italy’s Puglia region [7,8]. Genetic analysis of the subspecies prevalent in this area suggests a strong correlation with the subspecies found in Costa Rica. Nunney et al. (2012) conducted an analysis using MLST on isolates from grapevines across the United States and identified ST2, which differed from ST1 by a single base pair. This distinct pattern strongly suggests a recent common ancestry and challenges the previous assumption that the bacterial pathogen responsible for Pierce’s disease of grapevine (PD) is native to the United States. Instead, it indicates that the pathogen descended from a single genotype introduced from Central America [71]. Their invasion hypothesis is supported by five pieces of compelling evidence: a genome-wide lack of genetic diversity in Xf subsp. fastidiosa in the U.S., which is consistent with a recent common ancestor; evidence suggesting historical separation (allopatry) of the North American subspecies Xf subsp. multiplex and Xf subsp. fastidiosa; indications that Xf subsp. fastidiosa evolved in a more tropical climate compared to Xf subsp. multiplex; a considerably higher level of genetic variability in the proposed source population in Central America, within which the U.S. genotypes are phylogenetically nested; circumstantial evidence of the importation of known hosts (coffee plants) from Central America directly into southern California, just prior to the first documented outbreak of the disease [71,72]. Also, Yuan et al. (2010), using MLST scheme for the bacterial plant pathogen had consistent results with the hypothesis that Xf subsp. fastidiosa were introduced into the United States [70].
In 2015, the subspecies multiplex ST6 and ST7 genotypes were detected in France and annual official surveys with plant and vector samplings confirmed the presence and dissemination of these STs in Corsica and PACA regions [73,74]. In 2020, a new region, Occitanic (Aude), was found to be impacted by the presence of this genotype. Landa et al. (2020) constructed a phylogenetic tree using the available genome sequences of Xf subsp. multiplex and incorporated Xf subsp. fastidiosa strains. They also conducted ancestral state reconstruction using the geographic location of the strains as a character. The findings suggest that the ancestral nodes within Xf subsp. multiplex originated in the southeastern United States, implying that isolates from California, Europe, and Brazil likely resulted from introductions [75].
Up to now, 89 sequence types (ST) of Xf have been reported all around the world [6]. The sequence types most found in the world are described on Table 5 [42].
In Europe, the most common sequence types are ST53 in Italy [3,7], ST06 found in Spain and France [73], sequence type 07 in Portugal and France [76]. All these records of different STs in distant areas are clear indications of various independent introductions of Xf strains. Most of these STs had previously been described in the Americas hence that represents their most probable source of introduction in Europe [75]. These hypotheses were tested and confirmed by a first analysis of a limited number of strains from most EU outbreaks [77].
In Portugal, until now, we can only find the subspecies multiplex ST7 [76], and subspecies fastidiosa ST2 [78]. We found majoritarian de ST7 (in Santolina chamaecyparissus, Quercus suber, Lavandula dentata, Hebe sp., Coprosma repens, Cytisus scoparius) [42]. This indicated the potential maintenance of this variant over time in our country. The ST7 is closely related to STs already found in Europe, such as ST6 (France, and Spain mainland), ST81 (Balearic Islands) and ST87 (Italy) [27,79]. We can find this ST also in France, Balearic Islands and US [42]. Moreover, five samples of ST2 were found to be infected. That means additionally, this variant was introduced. We interpret these results as a consequence of commercial trade of plant material infected with Xf. The first reason is due to Xf, as a species, infects hundreds of plants, initially symptomless, and simultaneously the insect vectors are unlikely to disseminate at continental scales. So, it is reasonable to assume that anthropogenic activities are linked to the introduction of these pathogen strains into Europe. According to Yoon et al. (2023), current phytosanitary regulations and airport inspection systems in some parts of the region may have vulnerabilities, potentially allowing passengers to unknowingly transport infected planting materials. This raises the possibility of Xf invading new countries, with the transport infrastructure inadvertently facilitating the movement of Xf insect vectors. In the vicinity of the Demarcated Area of Porto, two major entry points can be identified: Francisco Sá Carneiro Airport and the Harbor of Leixões, close to Porto city. The plant nurseries within this Demarcated Area could represent a potential source of outbreaks. Consequently, a genuine risk exists that Xf could continue to breach geographical barriers and potentially triggering significant disease outbreaks [20,50].
In their study, Godefroid et al. (2018) examined the potential spread of three Xf subspecies by utilizing datasets delineating both native and recently colonized territories alongside four distinct species distribution models. The findings suggest that the subspecies multiplex and fastidiosa pose a significant threat to vast swathes of Europe, whereas climatically suitable regions for the subspecies pauca are primarily confined to the Mediterranean basin. The potential distribution of French strains ST6 and ST7, belonging to subsp. multiplex, is further influenced by a gradual shift of suitable areas. These areas are transitioning from Southern France, Italy, and Portugal toward Northern France, Belgium, the Netherlands, and Southern England. The potential expansion of these strains, particularly in Corsica and Southern France, is substantial. However, the dynamics of their expansion rely more on plant exchanges and disease management than solely on climate suitability. Furthermore, Godefroid et al. (2018) have predicted a moderate to highly severe risk of Pierce’s Disease (subspecies fastidiosa) in numerous lowlands and coastal regions of the Mediterranean areas, encompassing Spain, Italy, the Balearic Islands, and North Africa. This risk extends along the Atlantic coasts of France, Northern Spain, and Portugal [80]. Yoon et al. (2023) sought to determine the shared climatic traits of Xf. Furthermore, they calculated the spatiotemporal distances between occurrence areas and categorized them into either natural or anthropogenic spread. Among countries without ongoing outbreaks, those in the Mediterranean basin, notably Turkey at the highest level of risk, followed by Greece, Morocco, and Tunisia at a high-risk level, are most vulnerable to Xf’s arrival and establishment. The ranking model also underscores the susceptibility of southern European countries (Italy, Portugal, and Spain), where the pathogen has previously been detected [50].
According to models’ predictions, the currently reported distribution of Xf in Europe is small compared to the large extent of climatically suitable areas [80]. Species distribution models serve as valuable tools for forecasting the presence of species within a particular location and facilitating investigations related to fields such as biogeography, species preservation, invasive species ecology, and the management of natural resources [81]. These models empower researchers to untangle the influence of ecological and environmental factors on species distribution.
In their research, Hoddle et al. (2004) employed the CLIMEX algorithm to predict the global severity potential of PD. Their model indicated that while most Mediterranean regions are conducive to PD, the cold winter temperatures in certain areas might impede the expansion of Xf into several economically crucial wine-producing regions of France, as well as central and northern parts of Spain and Italy. Based on the model predictions, Xf had a high probability of colonizing areas characterized by low altitude (0–150 m a.s.l.), precipitations in the driest month < 10 mm, in the wettest month ranging between 80–110 mm and during the warmest quarter < 60 mm; mean temperature of coldest quarter ≥ 8°C; forest (essentially oak woodland) and Mediterranean shrubland [82].
The main reason for the exclusion of Xf from coldest and high altitudes is that cold stress on high altitudes can affect both, the bacterium and its vector [83]. The pathogen is probably less harmful where winter temperatures are colder, such as at higher altitudes, because freezing events can strongly decrease Xf infections [82,84]. European, and especially Euro-Mediterranean areas, are characterized by a broad spectrum of climatic conditions and plant species, providing the bacterium high chances to encounter hosts and environmental conditions favoring invasion and establishment. Conversely, the prospective behavior of Xf in regions characterized by exceptionally high summer temperatures, such as southern and central Spain, remains largely ambiguous due to the limited understanding of the effects of extreme heat on Xf. While elevated temperatures during spring and summer promote Xf proliferation in plants, research suggests that Xf populations decline in grapevines subjected to temperatures exceeding 37 °C [85]. According to the National Institute of Statistics, in 2022, the Northern region of Portugal encountered average temperatures of 9.8ºC in the coldest quarter and around 22°C in the hottest quarter, indicating favorable conditions for Xf [86].
Bosso et al. (2016), concluded that the analysis of single variable contribution showed that precipitation of the driest (40.3%) and wettest (30.4%) months were the main factors influencing model performance. Altitude, precipitation of warmest quarter, mean temperature of coldest quarter, and land cover provided a total contribution of 19.5% [87]. Yoon et al. (2023), reported that 90 % of precipitation density ranged from 19 to 112 mm, was suitable for Xf occurrence. Moreover, the average precipitation and high standard deviation suggested that the infections occurred easily in environments with regular precipitation with relatively large seasonal variations [50]. According to the National Institute of Statistics, in 2022, the average precipitation during the first and second quarters of the year was 37.2mm and 35.4mm respectively, creating favorable conditions for the establishment of Xf on North of Portugal.
Forest coverage or Mediterranean shrubland was associated to a higher probability of detecting the insect vector of Xf. This habitat likely provides a shelter for Ps, especially adults, which may move into woodland areas when the conditions in olive orchards or vineyards are less favorable (i.e., during hot days or crop management practices). Moreover, forests can provide nutritious hosts such as actinorhizal plants, which are characterized by a reliable supply of xylem-borne organic nitrogen compounds and are exploitable by spittlebugs when plants within crops are under stress [88]. According to Hoddle et al. (2004), the oak woodland creates a high probability of Xf colonization. Quercus pyrenaica Willd dominates nearly 95% of its natural distribution area in Spain, covering approximately 600,000 hectares, and 62,000 hectares in Portugal. In Portugal, the primary concentration of this species is observed in the Bragança region, encompassing around 40% of the total rangeland area [89]. It stands out as one of the most prevalent and distinctive species in Northern Portugal, making it favorable for Xf infection.
Godefroid et al. (2018) meticulously documented the natural and invaded territories of Xf subspecies fastidiosa, pauca, and multiplex. They employed bioclimatic species distribution models (SDMs) to gauge the potential climatic adaptability of these pathogens across Europe. Their findings highlight regions at heightened risk, including the coastal Mediterranean zones of Spain, Greece, Italy, and France, along with the Atlantic coastal regions of France, Portugal, and Spain, as well as the southwestern areas of Spain and low-lying regions in southern Italy. The subspecies multiplex, and to some extent fastidiosa, pose significant threats to much of Europe, whereas areas climatically suitable for subspecies pauca are primarily confined to the Mediterranean basin [80]. Bosso et al. (2016), developed a Maxent model to detect the current and future potential distribution of Xf in the Mediterranean under climate change. The potential distribution obtained comprises Portugal, Spain, Italy, Corsica, Albania, Montenegro, Greece and Turkey as well as all countries of Northern Africa and the Middle East [90]. That means that the bacterium could colonize those areas whose climatic regime is especially favorable for it. In contrast, cold stress would not allow the bacterium to spread [82,91]. Xf was not predicted to change its distribution in the Basin in response to climate change. Also, Cardone et al. (2022), confirmed the vulnerability of Euro-Mediterranean countries in terms of Xf dispersal. In the MENA and Balkans regions, countries with fragmented and small farms are likely to face the worst social impacts, whereas the Euro-Mediterranean region runs the highest economic losses on the target crops [92].
It seems that this disease occurs in a wide range of climatic zones, including tropical, subtropical, and continental areas, but is concentrated in temperate climate zones [38]. Similar to the studies mentioned earlier, our findings provide essential insights, suggesting that the Northern region of Portugal is highly conducive to Xf. The presence of a bacterial species with highly infectious genetic capabilities, along with hosts that are ideal for the bacteria, is evident. External conditions affecting the interaction between bacteria and hosts, such as environmental factors (temperature, humidity), are also present. This, combined with anthropogenic activities like the transport of infected planting materials from the first DZ of the Area Metropolitana of Porto, could be linked to the introduction and spread of Xf, potentially triggering new disease outbreaks in our country. Also, different subspecies of Xf and insect vector distributions must be considered. Another significant area of uncertainty lies in the potential adaptation of Xf to novel environmental pressures within its invaded territories, possibly through mechanisms like genetic recombination. Additionally, microclimate variations, yearly climate fluctuations, host plant susceptibility, interactions between hosts and pathogens, landscape configurations, and the spatiotemporal distribution of potential vectors all play crucial roles in shaping the epidemiology of the bacteria.
4. Conclusions
Our findings provide essential insights, suggesting that the Northern region of Portugal is highly conducive to Xf.
Introgression events have facilitated the ability of Xf to enter and infect national hosts. Its highly infectious genetic characteristics and natural competence to acquire new genes position makes it as a phytopathogen equipped with the tools necessary for adaptation to a wide range of ecological niches. This adaptability to changing climates leads us to anticipate that, without the implementation of a robust prevention plan grounded in local data, the Portuguese economy may suffer significant setbacks, including yield losses and other forms of damage. Mitigating these potential impacts will likely be necessary the implementation of expensive control measures.
Additionally, our country has preferred hosts, with priority given to specific plant species due to their economic significance or the expansion of production areas. Notably, plants such as Laurus nobilis, Rosa spp., Nerium oleander L., Pelargonium sp., Hedera helix L. and Lavandula dentata L., are selected for sampling. Olive trees, almond trees, vine and Citrus crops are also selected for prospecting, given their economic significance and uncertainties regarding their susceptibility to the subspecies of Xf present in the EU. Considering these factors, prospecting efforts are concentrated on monitoring and sampling these priority plant species. Nevertheless, it is crucial to consider variations in the behavior of insect vectors, as these variations can limit the prevalence of the disease. Ps may have limited flight ranges, confining the spread of the disease to a localized area. Also has specific host preferences for feeding. Variations in these preferences can impact the range of plant species that may serve as hosts for Xf, influencing the types of ecosystems and agricultural crops that are at greater risk of infection. The interplay of these vector behaviors directly impacts the geographic range of Xf impact.
Our adequate environmental factors such as temperature and humidity play a role in influencing the interaction between bacteria and hosts. The climatic conditions suitable for plant cultivation align with the climatic preferences of Xf outbreaks and insect vectors, posing a high risk of spread. Historical patterns of disease and insect vector spread indicate the potential for dispersion to various regions with favorable climatic conditions, facilitated by both anthropogenic and natural factors. Activities like the transport of infected planting materials from the first DZ of the Area Metropolitana do Porto could be associated with the introduction and spread of Xf, potentially triggering new disease outbreaks in our country. Trade networks are implicated in the increasing rates of spread of plant diseases through the transportation of live plants and other plant-related products. However, the risk of significant outbreaks is contingent on the presence of a suitable climate and land use to support the establishment and spread. Finally, it is worth noting that microclimate conditions, inter-annual climate variability, host-plant sensitivity, host-pathogen interactions, landscape structure and the spatiotemporal structure of the community of potential vectors can also affect the bacteria epidemiology.
Also, we can conclude that the presence of perennial plants in the Northern region of Portugal may account for absence of symptoms, as Xf infection in perennial plants has the capacity to persist in a chronic state over prolonged periods without exhibiting symptoms. The expression and appearance of symptoms are shaped by a myriad of physiological and environmental factors, encompassing variations in plant resilience or tolerance, diverse rates of transpiration, fluctuating levels of bacterial concentration, and differences in occlusion abilities among plant subspecies.
Our findings affirm the alarming spread of Xf across Portugal. Moreover, existing evidence suggests that the conditions in central and southern Europe are conducive to the introduction and further dissemination of Xf. Emphasizing the importance of proactive risk prediction for early detection to minimize damage, predictive studies are essential to establish fundamental data for monitoring and controlling Xf infections in our country. This entails analyzing the climatic characteristics and distribution patterns of Xf and its associated insect vectors.
As there are currently no effective control measures targeting the bacterium itself, this remains an open area of research. Limited knowledge exists about how Xf interacts with the plant host’s immune system, leaving numerous unanswered questions regarding Xf biology and its interactions with national plant hosts. Key inquiries include understanding how the plant immune system interfaces with Xf in the xylem and identifying the specific or shared mechanisms underlying the susceptibility of different hosts. Additionally, given that many Portuguese Xf plant hosts are non-symptomatic, there is potential to glean insights from these interactions to enhance the tolerance of other susceptible hosts.
This study introduces a straightforward statistical method to analyze the dispersion characteristics of Xf in the Northern region of Portugal, coupled with spatial data selection and extraction. This foundational data is essential for informing strategies to prevent the spread of Xf. Recognizing the interconnected nature of our world, where international plant trade and human travel act as potential pathways for Xf introduction into new areas, it becomes imperative to adopt comprehensive approaches considering multiple risk drivers. Factors such as the presence of insect vectors, abundance of host plants, and climate suitability play pivotal roles in Xf dispersal. We recommend that countries, especially those identified with relatively high risk, like Portugal, undertake thorough individual risk analyses. The implementation of preventive measures and, if necessary, the enhancement of surveillance systems for early detection of Xf in plants and insect vectors are crucial steps. Additionally, raising awareness is of equal importance to mitigate socioeconomic and ecological impacts on their ecosystems.
Author Contributions
Conceptualization, T.L., I.C.; methodology: T.L., I.C.; validation: T.L., I.C.; formal analysis: A.M., I.C., P.P.; investigation: T.L. and L.S.; data curation: T.L. and L.S.; writing—original draft preparation: T.L. and L.S.; writing—review and editing: T.L., I.C., P.P., A.M. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the projects UIDB/00772/2020(Doi:10.54499/UIBD/00772/2020) funded by the Portuguese Foundation for Science and Technology (FCT). The authors are grateful for all the conditions made available by the Foundation for Science and Technology (FCT, Portugal) and FEDER under Programme PT2020 for financial support to CITAB (UIDB/04033/2020)(https://doi.org/10.54499/UIDB/04033/2020).
Acknowledgments
This work had the support of the Northern Regional Directorate of Agriculture and Fisheries of the Ministry of Agriculture and Food. The author would like to thanks to Eng. Maria Manuel Mesquita, Eng. José Alves and to all colleagues involved in this work. Without their effort, this could not have been achieved.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- K. Schneider, M. Mourits, W. van der Werf, and A. O. Lansink, “On consumer impact from Xylella fastidiosa subspecies pauca,” Ecological Economics, vol. 185, p. 107024, Jul. 2021. [CrossRef]
- B. M. Ali, W. van der Werf, and A. Oude Lansink, “Assessment of the environmental impacts of Xylella fastidiosa subsp. pauca in Puglia,” Crop Protection, vol. 142, p. 105519, Apr. 2021. [CrossRef]
- V. Montilon et al., “Xylella fastidiosa subsp. pauca ST53 exploits pit membranes of susceptible olive cultivars to spread systemically in the xylem,” Plant Pathol, vol. 72, no. 1, pp. 144–153, Jan. 2023. [CrossRef]
- E. Food et al., “Pest survey card on Xylella fastidiosa,” EFSA Supporting Publications, vol. 16, no. 6, p. 1667E, Jun. 2019. [CrossRef]
- G. Petit et al., “Susceptibility to Xylella fastidiosa and functional xylem anatomy in Olea europaea: revisiting a tale of plant-pathogen interaction,” AoB Plants, vol. 13, no. 4, 2021. [CrossRef]
- Delbianco, D. Gibin, L. Pasinato, D. Boscia, and M. Morelli, “Update of the Xylella spp. host plant database – systematic literature search up to 30 June 2022,” EFSA Journal, vol. 21, no. 1, Jan. 2023. [CrossRef]
- Baù, A. Delbianco, G. Stancanelli, and S. Tramontini, “Susceptibility of Olea europaea L. varieties to Xylella fastidiosa subsp. pauca ST53: systematic literature search up to 24 March 2017,” EFSA Journal, vol. 15, no. 4, Apr. 2017. [CrossRef]
- G. P. Martelli, D. Boscia, F. Porcelli, and M. Saponari, “The olive quick decline syndrome in south-east Italy: a threatening phytosanitary emergency,” Eur J Plant Pathol, vol. 144, no. 2, pp. 235–243, Feb. 2016. [CrossRef]
- D. Martinetti and S. Soubeyrand, “Identifying lookouts for epidemio-surveillance: Application to the emergence of xylella fastidiosa in France,” Phytopathology, vol. 109, no. 2, pp. 265–276, Feb. 2019. [CrossRef]
- EPPO Bulletin, “PM 7/24 (4) Xylella fastidiosa,” EPPO Bulletin, vol. 49, no. 2, pp. 175–227, Aug. 2019. [CrossRef]
- G. Gilioli et al., “An eco-epidemiological model supporting rational disease management of Xylella fastidiosa. An application to the outbreak in Apulia (Italy),” Ecol Modell, vol. 476, Feb. 2023. [CrossRef]
- O. G. Hernández and L. V. García, “Incidencia de Xylella fastidiosa en las Islas Baleares y distribución potencial en la península ibérica,” Investigaciones Geográficas, no. 69, pp. 55–72, Jun. 2018. [CrossRef]
- D. Olmo et al., “Landscape Epidemiology of Xylella fastidiosa in the Balearic Islands,” Agronomy 2021, Vol. 11, Page 473, vol. 11, no. 3, p. 473, Mar. 2021. [CrossRef]
- DGAV, Plano de Contingência Xylella fastidiosa e seus vetores. 2022.
- “Xylella fastidiosa – DGAV.” Accessed: Mar. 25, 2023. [Online]. Available: https://www.dgav.pt/plantas/conteudo/sanidade-vegetal/inspecao-fitossanitaria/informacao-fitossanitaria/xylella-fastidiosa/.
- RE (UE) 2020/1201, “REGULAMENTO DE EXECUÇÃO (UE) 2020/1201 DA COMISSÃO de 14 de agosto de 2020 relativo às medidas para impedir a introdução e a propagação na União de Xylella fastidiosa (Wells et al.),” Accessed: Nov. 06, 2022. [Online]. Available: https://eur-lex.europa.eu/legal-content/PT/TXT/PDF/?uri=CELEX:32020R1201&from=EN.
- Paula, A. Cruz De Carvalho, A. Cruz De Carvalho Dn: C=pt, Title=subdiretora Geral, G. de Alimentação E Veterinária, P. de Almeida, and C. de Carvalho, “PLANO DE AÇÃO PARA ERRADICAÇÃO DE Xylella fastidiosa e controlo dos seus vetores ZONA DEMARCADA DA ÁREA METROPOLITANA DO PORTO Atualizado em fevereiro de 2022 Aprovado,” 2022.
- “Portaria n.o 243/2020, de 14 de outubro | DRE.” Accessed: Apr. 23, 2023. [Online]. Available: https://dre.pt/dre/detalhe/portaria/243-2020-145359683.
- “Decreto-Lei n.o 67/2020, de 15 de setembro | DRE.” Accessed: Apr. 23, 2023. [Online]. Available: https://dre.pt/dre/detalhe/decreto-lei/67-2020-142870334.
- M. Frem et al., “Xylella fastidiosa invasion of new countries in Europe, the Middle East and North Africa: Ranking the potential exposure scenarios,” NeoBiota 59: 77-97, vol. 59, pp. 77–97, Aug. 2020. [CrossRef]
- H. Purcell and S. R. Saunders, “Fate of Pierce’s Disease Strains of Xylella fastidiosa in Common Riparian Plants in California,” https://doi.org/10.1094/PDIS.1999.83.9.825, vol. 83, no. 9, pp. 825–830, Feb. 2007. [CrossRef]
- “Phyto71n04_429.pdf | Enhanced Reader.
- T. Cao, J. H. Connell, M. Wilhelm, and B. C. Kirkpatrick, “Influence of Inoculation Date on the Colonization of Xylella fastidiosa and the Persistence of Almond Leaf Scorch Disease Among Almond Cultivars,” https://doi.org/10.1094/PDIS-05-10-0327, vol. 95, no. 2, pp. 158–165, Jan. 2011. [CrossRef]
- A. Ledbetter, J. Chen, S. Livingston, and R. L. Groves, “Winter curing of prunus dulcis cv ‘Butte,’ P. webbii and their interspecific hybrid in response to xylella fastidiosa infections,” Euphytica, vol. 169, no. 1, pp. 113–122, May 2009. [CrossRef]
- R. P. P. Almeida and A. H. Purcell, “Biological Traits of Xylella fastidiosa Strains from Grapes and Almonds,” Appl Environ Microbiol, vol. 69, no. 12, pp. 7447–7452, Dec. 2003. [CrossRef]
- H. Feil, W. S. Feil, and A. H. Purcell, “Effects of Date of Inoculation on the Within-Plant Movement of Xylella fastidiosa and Persistence of Pierce’s Disease Within Field Grapevines,” Phytopathology, vol. 93, no. 2, pp. 244–251, Feb. 2003. [CrossRef]
- M. Saponari, A. Giampetruzzi, G. Loconsole, D. Boscia, and P. Saldarelli, “Xylella fastidiosa in olive in apulia: Where we stand,” Phytopathology, vol. 109, no. 2, pp. 175–186, Feb. 2019. [CrossRef]
- de N. G. dos Santos, M. Anguita-Maeso, and H. D. Coletta-Filho, “Transmission and distribution of Xylella fastidiosa subsp. pauca in olive trees as a parameter for managing olive quick decline syndrome,” Plant Pathol, vol. 71, no. 9, pp. 1849–1858, Dec. 2022. [CrossRef]
- N. Amanifar, M. Taghavi, and M. Salehi, “<em>Xylella fastidiosa</em> from almond in Iran: overwinter recovery and effects of antibiotics,” Phytopathol Mediterr, vol. 55, no. 3, pp. 337–345, Jul. 2016. [CrossRef]
- M. J. Davis, W. J. French, and N. W. Schaad, “Axenic culture of the bacteria associated with phony disease of peach and plum leaf scald,” Curr Microbiol, vol. 6, no. 5, pp. 309–314, 1981. [CrossRef]
- J. H. Aldrich, “ Distribution of Xylella fastidiosa Within Roots of Peach,” Plant Dis, vol. 76, no. 9, p. 885, 1992. [CrossRef]
- R. M. Holland, R. S. C. Christiano, E. Gamliel-Atinsky, and H. Scherm, “Distribution of Xylella fastidiosa in Blueberry Stem and Root Sections in Relation to Disease Severity in the Field,” https://doi.org/10.1094/PDIS-06-13-0680-RE, vol. 98, no. 4, pp. 443–447, Mar. 2014. [CrossRef]
- T. S. M. Henneberger, K. L. Stevenson, K. O. Britton, and C. J. Chang, “Distribution of Xylella fastidiosa in Sycamore Associated with Low Temperature and Host Resistance,” https://doi.org/10.1094/PDIS.2004.88.9.951, vol. 88, no. 9, pp. 951–958, Feb. 2007. [CrossRef]
- L. Hopkins, “ Seasonal Fluctuation in the Occurrence of Xylella fastidiosa in Root and Stem Extracts from Citrus with Blight,” Plant Dis, vol. 75, no. 2, p. 145, 1991. [CrossRef]
- X. He, W. B. Li, A. J. Ayres, J. S. Hartung, V. S. Miranda, and D. C. Teixeira, “Distribution of Xylella fastidiosa in Citrus Rootstocks and Transmission of Citrus Variegated Chlorosis Between Sweet Orange Plants Through Natural Root Grafts,” https://doi.org/10.1094/PDIS.2000.84.6.622, vol. 84, no. 6, pp. 622–626, Feb. 2007. [CrossRef]
- J. N. Rapicavoli et al., “Lipopolysaccharide O-antigen delays plant innate immune recognition of Xylella fastidiosa,” Nature Communications 2018 9:1, vol. 9, no. 1, pp. 1–12, Jan. 2018. [CrossRef]
- Purcell, “Paradigms: Examples from the Bacterium Xylella fastidiosa,” https://doi.org/10.1146/annurev-phyto-082712-102325, vol. 51, pp. 339–356, Aug. 2013. [CrossRef]
- Bragard et al., “Update of the Scientific Opinion on the risks to plant health posed by Xylella fastidiosa in the EU territory,” EFSA Journal, vol. 17, no. 5, May 2019. [CrossRef]
- R. B. Queiroz-Voltan, L. Perosin Cabral, and O. Paradela Filho, “Severidade do sintoma da bactéria Xylella fastidiosa em cultivares de cafeeiro,” Bragantia, vol. 63, no. 3, pp. 395–404, 2004. [CrossRef]
- L. Hopkins, “Biological Control of Pierce’s Disease in the Vineyard with Strains of Xylella fastidiosa Benign to Grapevine,” https://doi.org/10.1094/PD-89-1348, vol. 89, no. 12, pp. 1348–1352, Feb. 200. [CrossRef]
- P. P. Kandel, R. P. P. Almeida, P. A. Cobine, and L. De La Fuente, “Natural competence rates are variable among xylella fastidiosa strains and homologous recombination occurs in vitro between subspecies fastidiosa and multiplex,” Molecular Plant-Microbe Interactions, vol. 30, no. 7, pp. 589–600, Jul. 2017. [CrossRef]
- Gibin, L. Pasinato, and A. Delbianco, “Update of the Xylella spp. host plant database – systematic literature search up to 31 December 2022,” EFSA Journal, vol. 21, no. 6, p. e08061, Jun. 2023. [CrossRef]
- Niza, H. D. Coletta-Filho, M. V. Merfa, M. A. Takita, and A. A. de Souza, “Differential colonization patterns of Xylella fastidiosa infecting citrus genotypes,” Plant Pathol, vol. 64, no. 6, pp. 1259–1269, Dec. 2015. [CrossRef]
- Baccari, E. Antonova, and S. Lindow, “Biological control of Pierce’s disease of grape by an endophytic bacterium,” Phytopathology, vol. 109, no. 2, pp. 248–256, Feb. 2019. [CrossRef]
- Direção Geral de Alimentação e Veterinária, “XYLELLA FASTIDIOSA GÉNEROS E ESPÉCIES VEGETAIS DETETADOS INFETADOS NA ZONA DEMARCADA DA ÁREA METROPOLITANA DO PORTO,” Mar. 2023.
- P. S. Pereira, “Xylella fastidiosa - a new menace for Portuguese agriculture and forestry.,” Revista de Ciências Agrárias (Portugal), vol. 38, no. 2, pp. 149–154, 2015.
- V. Cavalieri et al., “Transmission of Xylella fastidiosa Subspecies Pauca Sequence Type 53 by Different Insect Species,” Insects, vol. 10, no. 10, Oct. 2019. [CrossRef]
- H. Dietrich, “KEYS TO THE FAMILIES OF CICADOMORPHA AND SUBFAMILIES AND TRIBES OF CICADELLIDAE (HEMIPTERA: AUCHENORRHYNCHA)”, Accessed: Apr. 23, 2023. [Online]. Available: https://bioone.org/journals/florida-entomologist/volume-88/issue-4/0015-4040_2005_88_502_KTTFOC_2.0.CO_2/KEYS-TO-THE-FAMILIES-OF-CICADOMORPHA-AND-SUBFAMILIES-AND-TRIBES/10.1653/0015-4040(2005)88[502:KTTFOC]2.0.CO;2.full.
- N. Bodino et al., “Spittlebugs of Mediterranean Olive Groves: Host-Plant Exploitation throughout the Year,” Insects 2020, Vol. 11, Page 130, vol. 11, no. 2, p. 130, Feb. 2020. [CrossRef]
- S. Yoon and W. H. Lee, “Spatial analysis of climatic and dispersion characteristics of Xylella fastidiosa outbreak by insect vectors,” J Asia Pac Entomol, vol. 26, no. 1, p. 102011, Mar. 2023. [CrossRef]
- Lago, D. Cornara, S. A. Minutillo, A. Moreno, and A. Fereres, “Feeding behaviour and mortality of Philaenus spumarius exposed to insecticides and their impact on Xylella fastidiosa transmission,” Pest Manag Sci, vol. 78, no. 11, p. 4841, Nov. 2022. [CrossRef]
- S. DROSOPOULOS and M. ASCHE, “Biosystematic studies on the spittlebug genus Philaenus with the description of a new species,” Zool J Linn Soc, vol. 101, no. 2, pp. 169–177, Feb. 1991. [CrossRef]
- M. Godefroid and J. M. Durán, “Composition of landscape impacts the distribution of the main vectors of Xylella fastidiosa in southern Spain,” Journal of Applied Entomology, vol. 146, no. 6, pp. 666–675, Jul. 2022. [CrossRef]
- R. Karban and S. Y. Strauss, “Physiological tolerance, climate change, and a northward range shift in the spittlebug, Philaenus spumarius,” Ecol Entomol, vol. 29, no. 2, pp. 251–254, Apr. 2004. [CrossRef]
- S. M. Chmiel and M. Curtis Wilson, “Estimation of the Lower and Upper Developmental Threshold Temperatures and Duration of the Nymphal Stages of the Meadow Spittlebug, Philaenus spumarius,” Environ Entomol, vol. 8, no. 4, pp. 682–685, Aug. 1979. [CrossRef]
- D. Ahmed and R. H. Davidson, “Life History of the Meadow Spittlebug in Ohio,” J Econ Entomol, vol. 43, no. 6, pp. 905–908, Dec. 1950. [CrossRef]
- S. Drosopoulos, “New data on the nature and origin of colour polymorphism in the spittlebug genus Philaenus (Hemiptera: Aphorophoridae),” Annales de la Société Entomologique de France, vol. 39, no. 1, pp. 31–42, Jan. 2003. [CrossRef]
- M. Chartois et al., “Environmental factors driving the abundance of Philaenus spumarius in mesomediterranean habitats of Corsica (France),” Scientific Reports 2023 13:1, vol. 13, no. 1, pp. 1–11, Feb. 2023. [CrossRef]
- Cornara et al., “Natural areas as reservoir of candidate vectors of Xylella fastidiosa,” Bull Insectology, vol. 74, no. 2, pp. 173–180, 2021.
- Dongiovanni et al., “Plant Selection and Population Trend of Spittlebug Immatures (Hemiptera: Aphrophoridae) in Olive Groves of the Apulia Region of Italy,” J Econ Entomol, vol. 112, no. 1, pp. 67–74, Feb. 2019. [CrossRef]
- H. Séverine and N. Casarin, “Belgian Journal of Entomology Distribution, adult phenology and life history traits of potential insect vectors of Xylella fastidiosa in Belgium”, Accessed: Oct. 25, 2023. [Online]. Available: https://www.researchgate.net/publication/352903222.
- X. Mesmin, M. Chartois, S. Borgomano, J. Y. Rasplus, J. P. Rossi, and A. Cruaud, “Interaction networks between spittlebugs and vegetation types in and around olive and clementine groves of Corsica; implications for the spread of Xylella fastidiosa,” Agric Ecosyst Environ, vol. 334, p. 107979, Aug. 2022. [CrossRef]
- J. Albre, J. M. G. Carrasco, and M. Gibernau, “Ecology of the meadow spittlebug Philaenus spumarius in the Ajaccio region (Corsica) – I: spring,” Bull Entomol Res, vol. 111, no. 2, pp. 246–256, Apr. 2021. [CrossRef]
- “(PDF) Xylella fastidiosa: AN OVERVIEW OF RESEARCH AT SASA.” Accessed: Oct. 24, 2023. [Online]. Available: https://www.researchgate.net/publication/344376341_Xylella_fastidiosa_AN_OVERVIEW_OF_RESEARCH_AT_SASA.
- N. Bodino et al., “Phenology, seasonal abundance and stage-structure of spittlebug (Hemiptera: Aphrophoridae) populations in olive groves in Italy,” Scientific Reports 2019 9:1, vol. 9, no. 1, pp. 1–17, Nov. 2019. [CrossRef]
- Dongiovanni et al., “Plant Selection and Population Trend of Spittlebug Immatures (Hemiptera: Aphrophoridae) in Olive Groves of the Apulia Region of Italy,” J Econ Entomol, vol. 112, no. 1, pp. 67–74, Feb. 2019. [CrossRef]
- M. Villa, I. Rodrigues, P. Baptista, A. Fereres, and J. A. Pereira, “Populations and Host/Non-Host Plants of Spittlebugs Nymphs in Olive Orchards from Northeastern Portugal,” Insects 2020, Vol. 11, Page 720, vol. 11, no. 10, p. 720, Oct. 2020. [CrossRef]
- M. Morente et al., “Distribution and Relative Abundance of Insect Vectors of Xylella fastidiosa in Olive Groves of the Iberian Peninsula,” Insects 2018, Vol. 9, Page 175, vol. 9, no. 4, p. 175, Dec. 2018. [CrossRef]
- S. Antonatos, D. P. Papachristos, K. Varikou, P. Vahamidis, A. Kapranas, and P. Milonas, “Seasonal Appearance, Abundance, and Host Preference of Philaenus spumarius and Neophilaenus campestris (Hemiptera: Aphrophoridae) in Olive Groves in Greece,” Environ Entomol, vol. 50, no. 6, pp. 1474–1482, Dec. 2021. [CrossRef]
- X. Yuan, L. Morano, R. Bromley, S. Spring-pearson, R. Stouthamer, and L. Nunney, “Multilocus sequence typing of Xylella fastidiosa causing Pierce’s disease and oleander leaf scorch in the United States,” Phytopathology, vol. 100, no. 6, pp. 601–611, Jun. 2010. [CrossRef]
- L. Nunney, S. Elfekih, and R. Stouthamer, “The importance of multilocus sequence typing: cautionary tales from the bacterium Xylella fastidiosa,” Phytopathology, vol. 102, no. 5, pp. 456–462, May 2012. [CrossRef]
- L. Nunney et al., “Population Genomic Analysis of a Bacterial Plant Pathogen: Novel Insight into the Origin of Pierce’s Disease of Grapevine in the U.S.,” PLoS One, vol. 5, no. 11, p. e15488, 2010. [CrossRef]
- N. Denancé et al., “Several subspecies and sequence types are associated with the emergence of Xylella fastidiosa in natural settings in France,” Plant Pathol, vol. 66, no. 7, pp. 1054–1064, Sep. 2017. [CrossRef]
- Cunty et al., “Update of the Xylella fastidiosa outbreak in France: two new variants detected and a new region affected,” Eur J Plant Pathol, vol. 163, no. 2, pp. 505–510, Jun. 2022. [CrossRef]
- B. Landa et al., “Emergence of a plant pathogen in europe associated with multiple intercontinental introductions,” Appl Environ Microbiol, vol. 86, no. 3, Feb. 2020. [CrossRef]
- Direção Geral de Alimentação e Veterinária, “Despacho no6/G/2022 - ATUALIZAÇÃO DA ZONA DEMARCADA PARA Xylella fastidiosa DA ÁREA METROPOLITANA DO PORTO.
- Dupas et al., “Suspicions of two bridgehead invasions of Xylella fastidiosa subsp. multiplex in France,” Communications Biology 2023 6:1, vol. 6, no. 1, pp. 1–13, Jan. 2023. [CrossRef]
- Direção Geral de Alimentação e Veterinária, “Atualização da Zona Demarcada para Xylella fastidiosa da Área Metropolitana de Lisboa.
- Giampetruzzi et al., “Genome-Wide Analysis Provides Evidence on the Genetic Relatedness of the Emergent Xylella fastidiosa Genotype in Italy to Isolates from Central America,” Phytopathology, vol. 107, no. 7, pp. 816–827, Jul. 201. [CrossRef]
- M. Godefroid, A. Cruaud, J.-C. Streito, J.-Y. Rasplus, and J.-P. Rossi, “Climate change and the potential distribution of Xylella fastidiosa in Europe,” bioRxiv, p. 289876, Mar. 2018. [CrossRef]
- Piyapong, C. Tattoni, M. Ciolli, S. Dembski, and E. Paradis, “Modelling the geographical distributions of one native and two introduced species of crayfish in the French Alps,” Ecol Inform, vol. 60, p. 101172, Nov. 2020. [CrossRef]
- M. S. Hoddle, “The potential adventive geographic range of glassy-winged sharpshooter, Homalodisca coagulata and the grape pathogen Xylella fastidiosa: implications for California and other grape growing regions of the world,” Crop Protection, vol. 23, no. 8, pp. 691–699, Aug. 2004. [CrossRef]
- V. Chikwendu and H. Tochi, “Aspects of the ecology of spittlebugs (Homoptera: Cercopidae) in Nsukka, south east, Nigeria,” Animal Research International, vol. 7, no. 3, pp. 1242–1252, 2010. [CrossRef]
- H. Purcell and D. L. Hopkins, “FASTIDIOUS XYLEM-LIMITED BACTERIAL PLANT PATHOGENS,” https://doi.org/10.1146/annurev.phyto.34.1.131, vol. 34, pp. 131–151, Nov. 2003. [CrossRef]
- H. Feil and A. H. Purcell, “Temperature-Dependent Growth and Survival of Xylella fastidiosa in Vitro and in Potted Grapevines,” Plant Dis, vol. 85, no. 12, pp. 1230–1234, 2001. [CrossRef]
- “Temperaturas médias do ar em Portugal 2019-2022.” Accessed: Dec. 17, 2023. [Online]. Available: https://www.ine.pt/xportal/xmain?xpid=INE&xpgid=ine_indicadores&indOcorrCod=0009895&contexto=bd&selTab=tab2&xlang=pt.
- L. Bosso, D. Russo, M. Di Febbraro, G. Cristinzio, and A. Zoina, “Potential distribution of Xylella fastidiosa in Italy: A maximum entropy model,” Phytopathol Mediterr, vol. 55, no. 1, pp. 62–72, 2016. [CrossRef]
- S. Avosani, C. Tattoni, V. Mazzoni, and M. Ciolli, “Occupancy and detection of agricultural threats: The case of Philaenus spumarius, European vector of Xylella fastidiosa,” Agric Ecosyst Environ, vol. 324, p. 107707, Feb. 2022. [CrossRef]
- M. Castro, J. Castro, and A. Gómez Sal, “The role of black oak woodlands (Quercus pyrenaica Willd.) in small ruminant production in Northeast Portugal,” Sustainability of Agrosilvopastoral Systems, pp. 221–229, 2004, Accessed: Dec. 07, 2023. [Online]. Available: https://bibliotecadigital.ipb.pt/handle/10198/4447.
- L. Bosso, M. Di Febbraro, G. Cristinzio, A. Zoina, and D. Russo, “Shedding light on the effects of climate change on the potential distribution of Xylella fastidiosa in the Mediterranean basin,” Biol Invasions, vol. 18, no. 6, pp. 1759–1768, Jun. 2016. [CrossRef]
- H. Feil and A. H. Purcell, “Temperature-Dependent Growth and Survival of Xylella fastidiosa in Vitro and in Potted Grapevines,” https://doi.org/10.1094/PDIS.2001.85.12.1230, vol. 85, no. 12, pp. 1230–1234, Feb. 2007. [CrossRef]
- Cardone et al., “Socio-Economic Risks Posed by a New Plant Disease in the Mediterranean Basin,” Diversity (Basel), vol. 14, no. 11, p. 975, Nov. 2022. [CrossRef]
Figure 1.
Demarcated zones (DZ) of Xylella fastidiosa in Portugal.

Figure 2.
Resume for prospection phase.
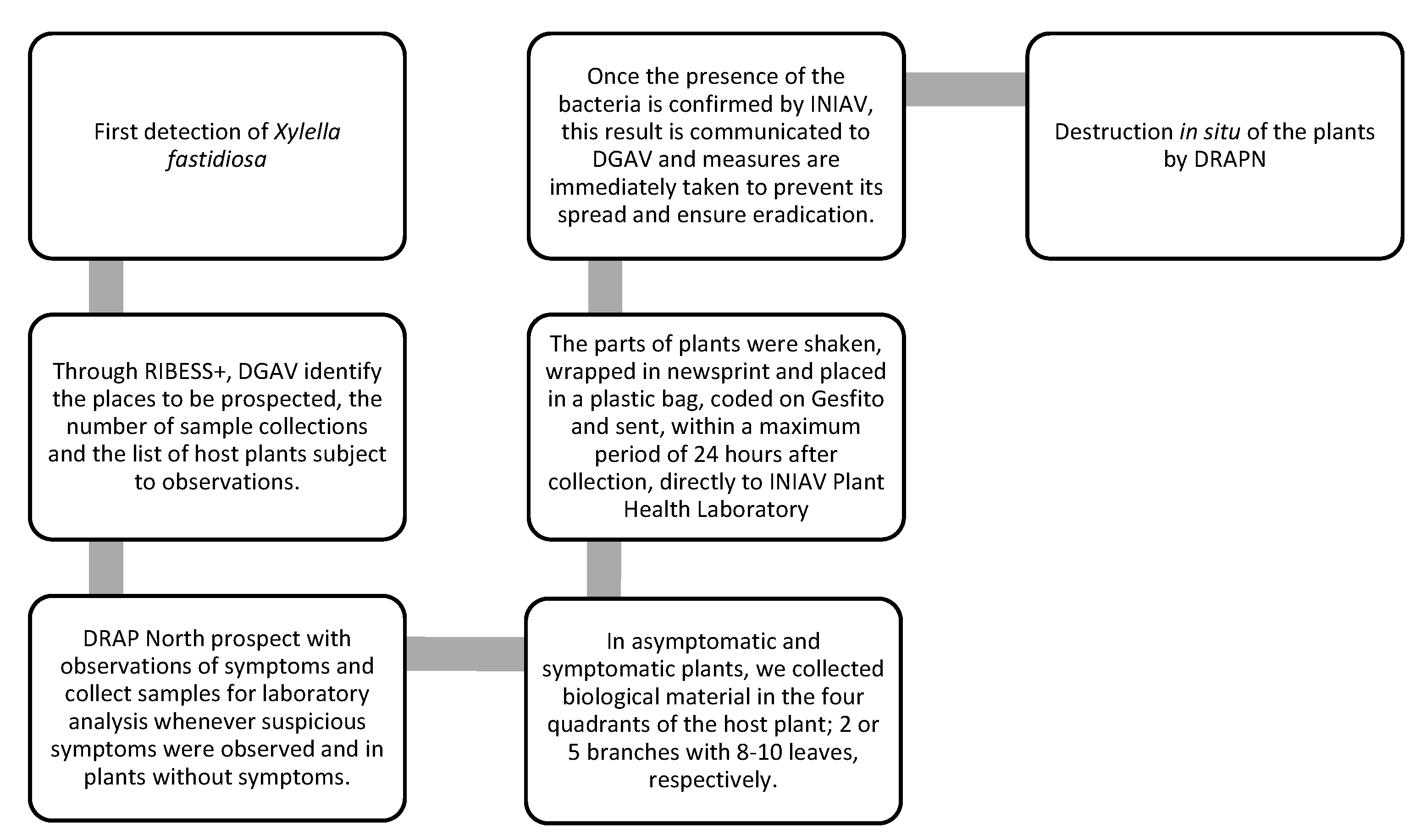
Figure 3.
Destruction of positive plants (author’s image).

Scheme 1.
Locations observed on North Districts of Portugal: Porto- 14358 observations (93.6%); Bragança– 907 observations (5.9%); Vila Real – 80 observations (0.5%).
Scheme 1.
Locations observed on North Districts of Portugal: Porto- 14358 observations (93.6%); Bragança– 907 observations (5.9%); Vila Real – 80 observations (0.5%).
Scheme 2.
Locations observed on DZs of Bragança, Porto and Vila Real since 2019 to 2023.
Scheme 3.
Type of local observed on North zone of Portugal. (1) - Private Garden /park, 5334 observations, 34.8%; (2) – Public Garden/park, 4083 observations, 26.6%; (3) – Nursery, 3752 observations, 24.5%; (4) – Garden center, 1235 observations, 8.1%; (5) – Agricultural field, 752 observations, 4.9%; (6) – Forest 189 observations, 1.2%.
Scheme 3.
Type of local observed on North zone of Portugal. (1) - Private Garden /park, 5334 observations, 34.8%; (2) – Public Garden/park, 4083 observations, 26.6%; (3) – Nursery, 3752 observations, 24.5%; (4) – Garden center, 1235 observations, 8.1%; (5) – Agricultural field, 752 observations, 4.9%; (6) – Forest 189 observations, 1.2%.
Scheme 4.
Positive plants distributed by type of local observed on North zone of Portugal. (1) - Private garden/park, 51 observations,16.9%; (2) – Public Garden/park, 129 observations, 42.7%; (3) – Nursery, 112 observations, 37.1%; (4) – Garden center, 4 observations, 1.3%; (5) – Agricultural field, 4 observations=1.3%;(6) – Forest, 2 observations, 0.7%.
Scheme 4.
Positive plants distributed by type of local observed on North zone of Portugal. (1) - Private garden/park, 51 observations,16.9%; (2) – Public Garden/park, 129 observations, 42.7%; (3) – Nursery, 112 observations, 37.1%; (4) – Garden center, 4 observations, 1.3%; (5) – Agricultural field, 4 observations=1.3%;(6) – Forest, 2 observations, 0.7%.
Scheme 5.
Presence of symptoms on the positive prospected plants on North of Portugal. (Absence) – 13984 observations, 91.1%; (Presence) – 1361 observations, 8.9%.
Scheme 5.
Presence of symptoms on the positive prospected plants on North of Portugal. (Absence) – 13984 observations, 91.1%; (Presence) – 1361 observations, 8.9%.
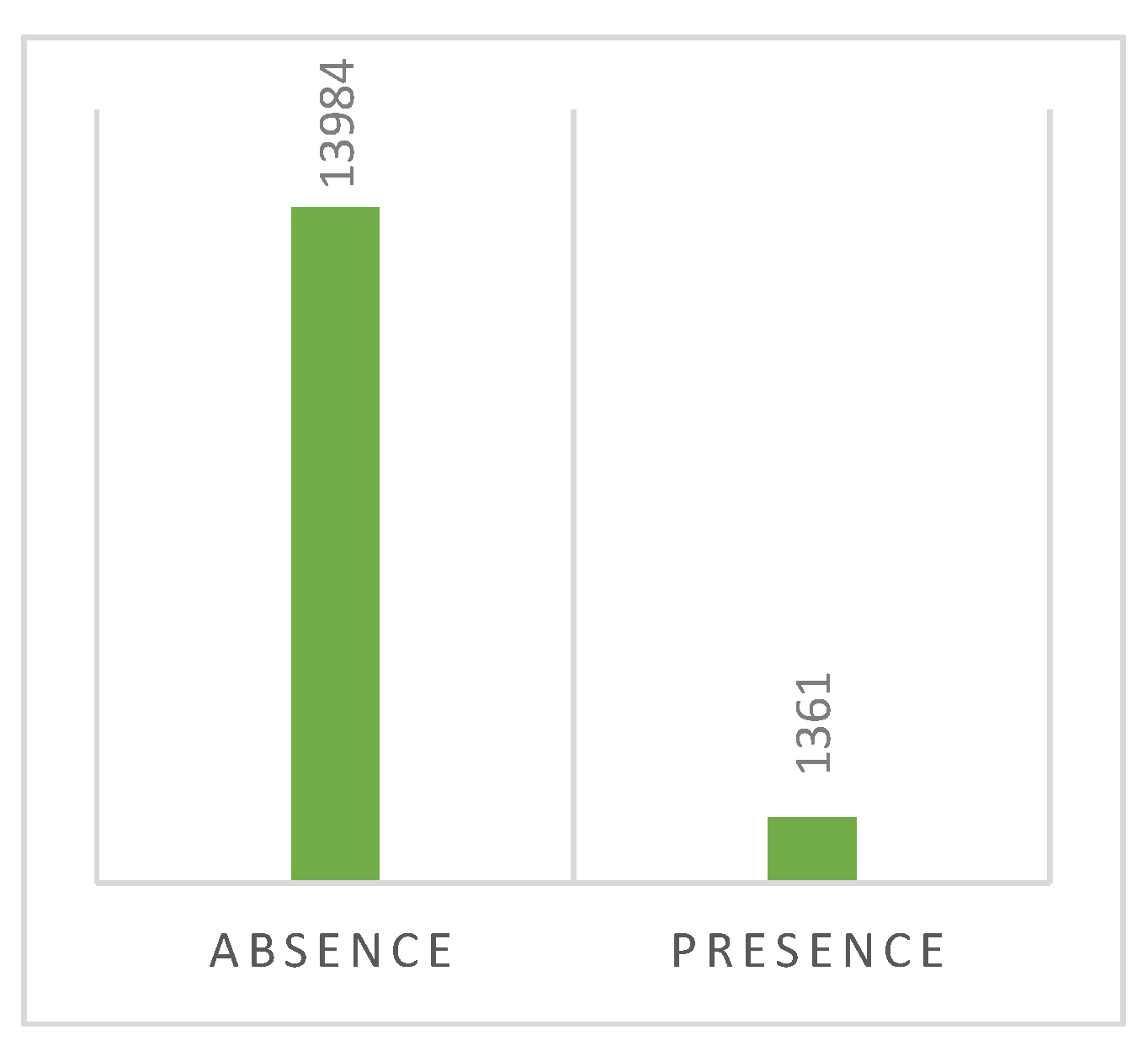

Table 1.
Chi-square test of independence between species and symptoms.
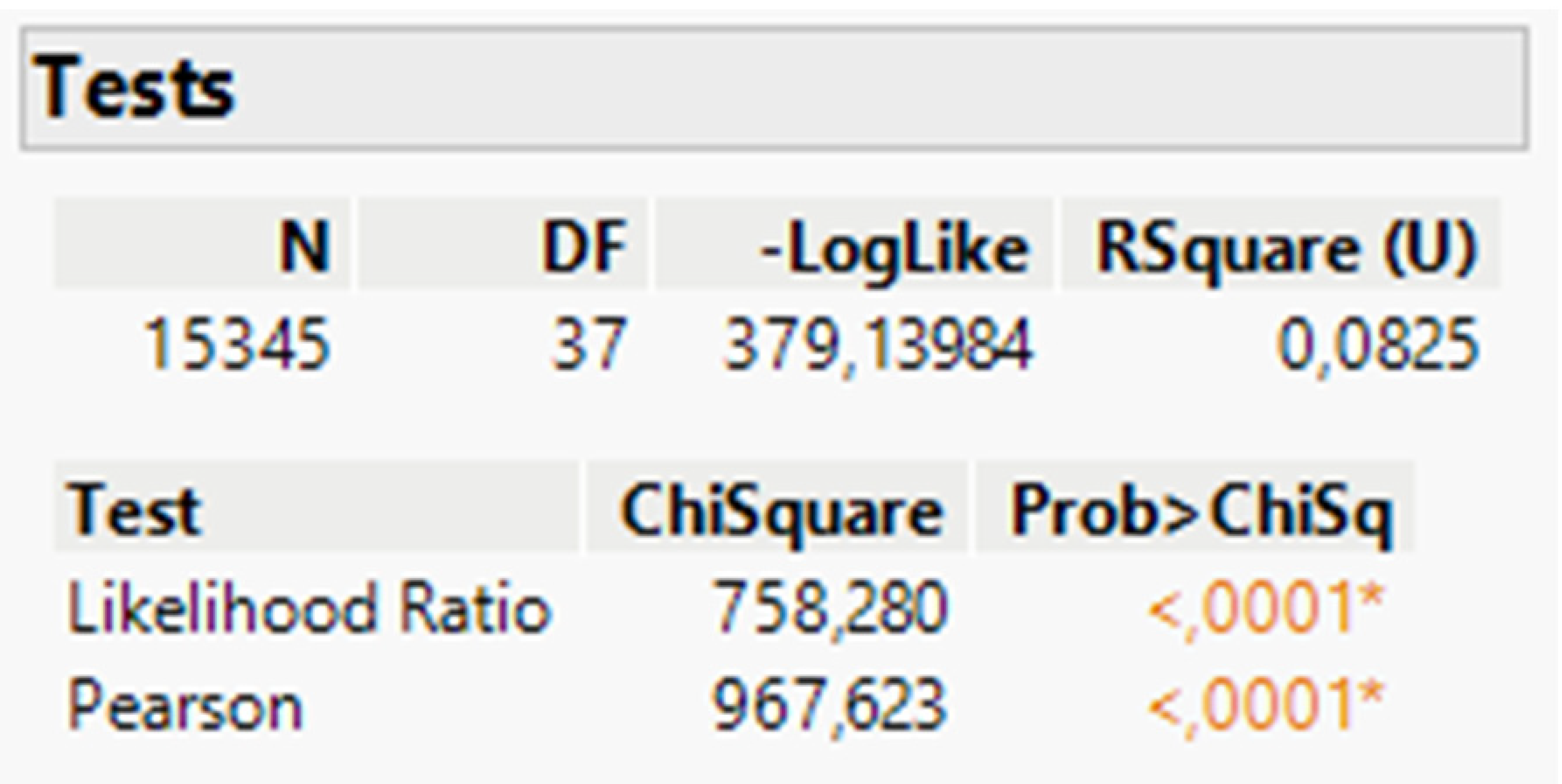 |
Table 2.
Most frequently prospected host species.
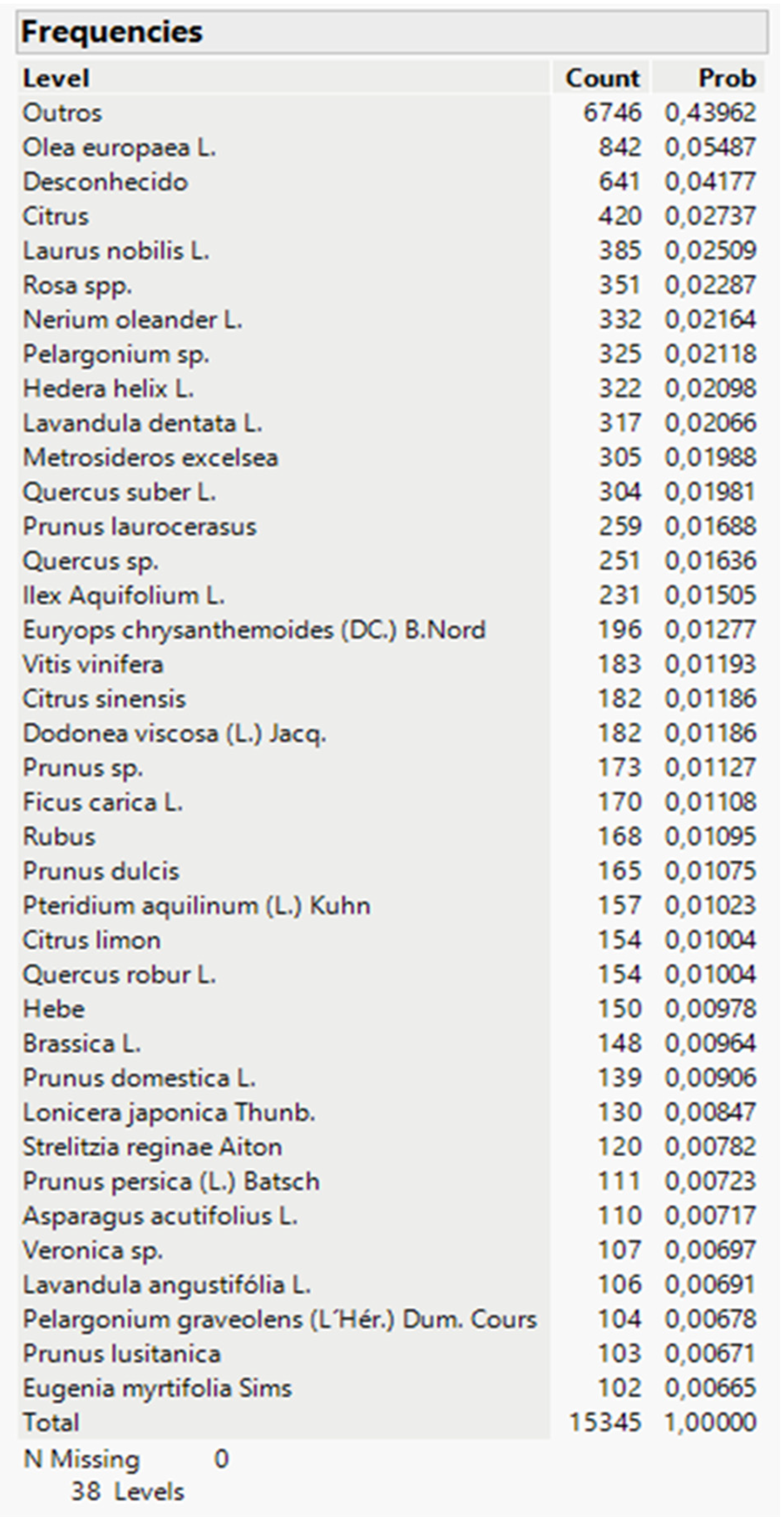 |
Table 3.
Positive Plant Species.
 |
Table 4.
Plant species identified in Portugal 2019-2023 [45.].
| 1. Acacia longifolia (Andrews) Wild. | 41. Lagerstroemia indica L. | 38. Hypericum androsaemum L. |
|---|---|---|
| 2. Acacia melanoxylon R. Br. | 42. Laurus nobilis | 39. Hypericum perforatum L. |
| 3. Acacia dealbata Link. | 43. Lavandula angustifólia L. | 40. Ilex aquifolium L. |
| 4. Adenocarpus lainzii (Castrov.) | 44. Lavandula dentata L. | 78. Vinca |
| 5. Argyranthemum frutescens L. | 45. Lavandula stoechas L. | 79. Vitis spp. |
| 6. Artemisia arborescens L. | 46. Lavatera cretica L. | |
| 7. Asparagus acutifolius L. | 47. Liquidambar styraciflua L. | |
| 8. Athyrium filix-femina (L.) Roth. | 48. Lonicera periclymenum L. | |
| 9. Berberis thunbergii DC. | 49. Magnolia grandiflora L. | |
| 10. Calluna vulgaris (L.) Hull. | 50. Magnolia x soulangeana Soul.-Bod. | |
| 11. Castanea sativa Mill. | 51. Mentha suaveolens Ehrh. | |
| 12. Cistus psilosepalus Sweet. | 52. Medicago sativa L. | |
| 13. Cistus salviifolius L. | 53. Metrosideros excelsea Sol. Ex Gaertn. | |
| 14. Citrus limon (L.) N. Burman | 54. Myrtus communis L. | |
| 15. Citrus paradisii Macfadyen | 55. Nerium oleander L. | |
| 16. Citrus reticulata Blanco | 56. Olea europaea L. | |
| 17. Citrus sinensis (L.) Osbeck | 57. Pelargonium graveolens (L´Hér.) Dum. | |
| 18. Coprosma repens A. Rich. | 58. Pyracantha coccinea M. Römer | |
| 19. Cortaderia selloana | 59. Plantago lanceolata L. | |
| 20. Cytisus scoparius (L.) Link. | 60. Platanus x hispanica | |
| 21. Dimorphoteca ecklonis (DC.) Norl. | 61. Prunus laurocerasus L. | |
| 22. Dodonea viscosa (L.) Jacq. | 62. Prunus persica (L.) Batsch | |
| 23. Echium plantagineum L. | 63. Prunus cerasifera Ehrh. | |
| 24. Elaeagnus x submacrophylla | 64. Pteridium aquilinum (L.) Kuhn | |
| 25. Erica cinerea L. | 65. Quercus coccinea Münchh. | |
| 26. Erigeron canadensis (L.) | 66. Quercus robur L. | |
| 27. Erodium moschatum (L.) L. Her. | 67. Quercus rubra L. | |
| 28. Euryops chrysanthemoides (DC.) B. Nord. | 68. Quercus suber L. | |
| 29. Frangula alnus Mill. | 69. Rosa | |
| 30. Gazania rigens (L.) Gaertn. | 70. Rubus idaeus L. | |
| 31. Genista triacanthos Brot. | 71. Rubus ulmifolius Schott. | |
| 32. Genista tridentata (L.) | 72. Ruta graveolans L. | |
| 33. Gleditsia triacanthos L. | 73. Salvia rosmarinus Spenn. | |
| 34. Grevillea rosmarinifolia | 74. Sambucus nigra L. | |
| 35. Hebe | 75. Santolina chamaecyparissus L. | |
| 36. Helichrysum italicum (Roth) G.Don | 76. Strelitzia reginae Ait. | |
| 37. Hibiscus syriacus L. | 77. Ulex spp. |
Table 5.
Sequence types of Xf most found in the world.
| Sequence Types | Subspecies | Country most frequently found | Number of records |
|---|---|---|---|
| ST53 | pauca | Italy, France, Costa Rica | 475 |
| ST11 | pauca | Brazil | 52 |
| ST1 | fastidiosa | US, Spain, Mexico | 201 |
| ST06 | multiplex | France, Spain, US | 160 |
| ST07 | multiplex | Portugal, US, France | 142 |
| ST81 | multiplex | Spain | 100 |
| ST87 | multiplex | Italy | 91 |
| ST29 | morus | US | 10 |
| ST05 | sandyi | US | 25 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



