Submitted:
28 May 2024
Posted:
29 May 2024
You are already at the latest version
Abstract
Mutualism between insects and symbiotic bacteria drives insect evolution, and insecticide resistance is affected by symbiotic bacteria, increasing the complexity of pest resistance. This study evaluated the sublethal and transgenerational effects of the neonicotinoid insecticide nitenpyram on the population dynamics of Acyrthosiphon gossypii and measured its impact on the symbiotic bacterial community. The results showed that LC20 (2.49 mg·L−1) of nitenpyram significantly reduced the fecundity and lifespan of the parent G0 generation of A. gossypii, and had similar inhibitory effects on its G1 and G2 generations. Compared with those of control group, the population parameters λ (finite rate of increase), R0 (net reproductive rate), and r (intrinsic increase rate) of G1 and G2 were significantly reduced, but mean generation time (T) was increased. Nitenpyram treatment affected the biological traits of A. gossypii, and also changed the relative abundance of Buchnera and Sphingomonas in three consecutive generations G0-G2. Under nitenpyram treatment, the relative abundance of Buchnera decreased in G0, while that of Sphingomonas increased. The relative abundance of Buchnera increased in G1 and G2, but that of Sphingomonas decreased. Our findings may provide a new perspective for evaluating the potential effects of pesticides on aphids.
Keywords:
nitenpyram
; Acyrthosiphon gossypii
; symbiotic bacteria
; sublethal effects
; transgenerational effects
1. Introduction
The Acyrthosiphon gossypii Mordviiko (Hemiptera: Aphididae) is an important economic pest, mainly damaging cotton and legume crops [1], It also damages Malvaceae, Brassicaceae, Cruciferae, Zygophyllaceae, Asteraceae, and Rosaceae plants. A. gossypii is known to be only distributed in Xinjiang and Gansu provinces in China [2]. At present, cotton aphids are one of the main pests in Xinjiang cotton region, the largest and main cotton-producing area in China. Adults and nymphs of A. gossypii suck the juice of cotton leaves, stems, and tender shoots during the seedling stage, causing serious crop damages through direct feeding, honeydew excretion, and damaging bud bolls, thus affecting the development of cotton plants [3]. Due to their small size, rapid reproduction, and large number, the management of cotton aphids poses a great challenge [4]. Chemical insecticides are widely used to control these pests. However, the frequent application and over-reliance on chemical insecticides resulted in the rapid evolution of resistance. Recent reports show that aphids have resistance to most commonly used insecticides in the field [5,6].
Nitenpyram is a second-generation neonicotinoid insecticide developed and commercialized by Takeda Agro [7,8,9]. Nitenpyram acts on the nicotinic acetylcholine receptor (nAChR), and it has a nerve-blocking effect on synaptic receptors in the central nervous system of insects, thereby achieving insecticidal effects [10], Nitenpyram has been widely used in the prevention and control of piercing-sucking mouthpart pests such as Aphis gossypii [11], Nilaparvata lugens [12], Bemisia tabaci [7]. However, the resistance of pests to nitenpyram has been widely reported, which will lead to a resurgence of pests [13].
Driven by biotic and abiotic factors, pesticides continue to degrade, and surviving individuals exposed to sublethal concentrations of pesticides will experience sublethal effects [14,15]. Exposure to sublethal concentrations of pesticides directly affects the development, reproduction, survival rate, feeding behavior, of insects [16,17,18], and indirectly affects their descendants through transgenerational inheritance, thus causing changes in population dynamics [19,20]. The sublethal effects vary with different insecticide types. The sublethal effects of pesticides often have negative effects on the growth, development and reproduction of individual pests. For example, under treatment with flonicamid, the longevity and fecundity of Schizaphis graminum parents are significantly reduced, and the population growth parameters λ (finite rate of increase), R0 (net reproductive rate), r (intrinsic increase rate), longevity and fecundity of the F1 generation are also significantly reduced [21].
Low concentrations of chlorpyrifos and spirotetramat can significantly reduce fecundity, oviposition period, and female and male longevity of Aenasius bambawalei [22]. However, pesticides have been reported to promote population number growth of pests. For example, flupyradifurone stimulates the fecundity of Aphis craccivora and induces its transgenerational hormesis effects [20]. Hormesis effects have been also reported in cotton aphids under low-concentration treatments of Thiamethoxam [23]. This hormesis effect not only results in the ineffectiveness of chemical control, but also promotes the rapid development of insecticide resistance.
Insecticides affect both the insect life cycle characteristics and their symbiotic bacteria [24,25]. It has been reported that symbiotic bacteria are directly or indirectly involved in the resistance of insects to insecticide [26]. Citrobacter freundii, an intestinal symbiont of Bactrocera dorsalis, degrades trichlorphon into chloral hydrate and dimethyl phosphite, thus enhancing the host’s resistance to trichlorphon [27]. The density of symbiotic bacteria Serratia and Acinetobacter in insecticide-resistant Aedes albopictus is higher than that in sensitive individuals, indicating that Serratia and Acinetobacter promote the development of resistance in Aedes albopictus [28]. Similarly, Stenotrophomonas enhances silkworm tolerance to chlorpyrifos, thus contributing to the host resistance to its toxic effects [29]. Therefore, it is of great significance to explore the role of insect symbionts in the development of insecticide resistance during pest management.
This study determined the toxicity of nitenpyram to A. gossypii, and investigated the sublethal effect of nitenpyram sublethal concentrationon A. gossypii based on life table analysis. In addition, we further revealed the mechanism of sublethal effects using 16S rRNA sequencing and examined the effect of nitenpyram on the diversity and composition of the bacterial community of A. gossypii. This study provides an insight into the potential impact of nitenpyram on A. gossypii, and offers valuable references of the development of prevention and control strategies against A. gossypii.
2. Materials and Methods
2.1. Insect Source
The insect in this study was A. gossypii. Sensitive populations of A. gossypii were collected from the cotton experimental field of Agricultural Experiment Station, the Tarim University, Alar City, Xinjiang, China in 2020. Since collection, they were raised on cotton (Zhong 49) at 22±1°C, a relative humidity of 60% ± 5%, and a photoperiod of light:dark=16 h:8 h. During this period, A. gossypii was not exposed to any insecticide.
2.2. Toxicity Bioassays
The leaf-dip bioassay method was used to determine the toxicity of nitenpyram to A. gossypii [30,31]. Nitenpyram (purity > 95%) was purchased from Shandong Hailier Chemical Co, Ltd (Shangdong province, China), and Triton X-100 was provided by Beijing Solebao Technology Co, Ltd (Beijing, China). Distilled water containing 0.05% TritonX-100 was prepared. A small amount of acetone was added into the original drug nitenpyram to fully dissolve nitenpyram, and then the distilled water was added so as to serially dilute drug solution and obtain six pesticides concentration gradients for bioassay. Fresh cotton leaves with a diameter of 90 mm were soaked in distilled water (control group) or drug solution (treatment group) for 15 seconds, dried in the shade, and placed in a petri dish (diameter = 90 mm, height = 20 mm) containing 1.8% agar, with leaf front side touching agar. The 30 apterous adults of A. gossypii were carefully put into each dish, and each treatment was repeated three times. The mortality rate was recorded at hour 48 post treatment. The toxicity of nitenpyram at different concentrations (20%, 50%, and 90% lethal concentrations; LC20, LC50, and LC90) on A. gossypii was analyzed using IBM SPSS Statistics 25.0 software.
2.3. Sublethal Effect of Nitenpyram on A. gossypii
No less than 300 adult apterous aphids were put in a climate chamber with constant temperature and humidity, and the adults were transferred after 24 hours. The G0 generation of the experimental population referred to those which grew from the newborn nymphs to adults in the climate chamber, and the G1 generation referred to the offspring born by adults of the G0 generation. The toxicity was evaluated by the sublethal effect of nitenpyram at LC20 on A. gossypii. Thirty adult aphids were placed on cotton leaves treated with nitenpyram at LC20 in each petri dish and treated for 48 hours, and surviving aphids were carefully collected and raised on fresh cotton leaves in a climate chamber. There were no less than 30 newborn nymphs in each group with 3 biological replicates for each treatment. One single aphid was raised in one single petri dish, with the aphid in each petri dish numbered, and their life table parameters were recorded every 3 hours.
The experiment started from the G0 and ended up with the G3. Cotton leaves and 1.8% agar were renewed every 4 days, and newborn nymphs were counted and eliminated every day. The life table data were input into TWOSEX-MSChart software for analysis [32]. The mean value and standard error of the experimental data were estimated by 100,000 random samplings in the bootstrap program, and the significant differences between the control group and the treatment groups were compared and analyzed by the paired bootstrap test [33]. Sigmaplot 14.0 software was used to plot the curves of age-specific survival rate, reproductive value, age-stage-specific life expectancy, and others.
2.4. Changes in Microbial Diversity of A. gossypii under Nitenpyram
The G0 generation adult A. gossypii feeding on cotton leaves treated with LC20 nitenpyram or distilled water and the offspring (G1 and G2) feeding on cotton leaves without chemicals were collected into cryopreservation tubes and stored them in -80°C refrigerator for 16S rRNA sequencing. Each treatment was repeated three times.
The total genomic DNA of the samples was extracted using the MagPure Soil DNA LQ kit (Magen, D6356-02). The samples were loaded into the 1% agarose gel wells for electrophoresis at 120V for 15 min, and the concentration and quality of the DNA were detected using a NanoDrop2000 (Thermo Fisher) spectrophotometer. The 16S rRNA V3-V4 region fragments were PCR amplified using universal primers (343F: TACGGRAGGCAGCAG) (798R: AGGGTATCTAATCCT) [34] for PCR amplification with genomic DNA as template. A total of 30 μL PCR amplification system contained 15 μL 2×Gflex PCR Buffer, 1 μL 5 pmol/μL primer F, 1 μL 5 pmol/μL primer R, 1 μL template DNA, 0.6 μL Tks Gflex DNA Polymerase (1.25 U/μL), and 11.4 μL of H2O. The PCR amplification program was as follows: 94 °C for 5 min, followed by 26 cycles of 94 °C for 30 S, 56 °C for 30 S, and 72 °C for 20 S, ending up with an extension at 72 °C for 5 min. PCR amplification was performed with Bio-rad PCR instrument. The PCR amplification product was detected by 1% agarose gel electrophoresis, and then magnetic beads were purified and used as a template for secondary PCR amplification. The secondary PCR amplification product was detected by 1% agarose gel electrophoresis, and purified again using magnetic beads. The purified PCR product was quantified using Qubit dsDNA Assay Kit (Life Technologies).
According to the PCR product concentration, the samples were pooled in equimolar and sequenced by Shanghai Ouyi Biomedical Technology Co, Ltd (Shanghai, China). The obtained raw sequencing data were saved in the FASTQ format. The adapter sequences of raw reads were cut using cutadapt software. According to the QIIME 2 (2020.11) default parameters, the qualified paired-end raw reads obtained from the previous step were subjected to the quality control including noise reduction, splicing, and chimera sequence removal, and others, using the DADA2 software [35,36], to obtain the representative sequences and amplicon sequence variant (ASV) abundance table. Based on the Silva database (version138), the representative sequences were further selected from each ASV using the QIIME 2 software, aligned, and annotated using q2-feature-classifier software. Raw sequence data were deposited in the GenBank Short Read Archive (SRA) database with the project accession number of PRJNA1114060.
The obtained high-quality sequences were further analyzed. Bacterial diversity was analyzed according to the ASV characteristics table. The alpha diversity analysis was conducted to reveal the degree of species diversity within a biological environment [37]. The beta diversity analysis was also performed to reflect the differences among different sample groups. Principal coordinates analysis (PCoA) was implemented based on the binary_jaccard distance matrix analysis results to visualize the data of the treatment group and the control group [38]. The differences in microbiota species among different groups were determined through LEfSe analysis (LDA value>2) [39]. The differences in bacterial composition among groups were analyzed using SPSS 26.0 software. P < 0.05 was considered as statistically significant.
3. Results
3.1. Toxicity of Nitenpyram to A. gossypii
The acute toxicity of A. gossypii adults was determined after 48 hour exposure to nitenpyram (Table 1). The LC20 and LC50 of nitenpyram against A. gossypii were 2.49 mg·L−1 with a confidence interval (CI) of 0.78-4.31 mg·L−1 and 10.12 mg·L−1 with a CI of 6.34-17.08 mg·L−1, respectively. Nitenpyram LC20 sub-lethal concentration (2.49 mg·L−1) was selected for subsequent experiments.
3.2. Sublethal Effect of Nitenpyram on A. gossypii Parents
Nitenpyram LC20 affected the fecundity and lifespan of the A. gossypii G0 generation after 48 hour treatment. As shown in Figure 1, the fecundity and lifespan of the nitenpyram-treated group were significantly lower than that of the control group without exposure to nitenpyram.
3.3. Transgenerational Sublethal Effect of Nitenpyram on G1-G3 Generations of A. gossypii
Table 2 and 3 presented the sublethal effects of nitenpyram on the life table parameters of G1-G3 generations of A. gossypii. The 48 hour exposure of A. gossypii G0 generation individuals to LC20 nitenpyram had a negative effect on their offsprings. Compared with the control group, the G1 and G2 in treatment group exhibited an extended nymph developmental period. Accordingly, LC20 nitenpyram also significantly prolonged the total preovipositional period (TPOP) and pre-adult stage of offspring aphids, and fecundity and lifespan were significantly shorter in the treatment group than in control group.
Additionally, compared with those of the control group, the net reproductive rate (R0), intrinsic rate of increase (rm), and weekly finite rate of increase (λ) of A. gossypii populations in the treatment group were significantly reduced. However, LC20 nitenpyram significantly prolonged the mean generation time (T) of G1 generation aphids. No significant differences in parameters such as development period, population growth, and fecundity of G3 A. gossypii were observed between control and treatment group (Table 3).
In the G1, the age-specific survival rate (lx) of A. gossypii was decreased with the increase in age, and on day 18 after treatment, the survival rate of treatment group was significantly lower than that of the control group (Figure 2(a)). The age-specific fecundity (mx) and age specific maternity (lxmx) curves of the total population showed the control adults reached the peak (4.47) on day 11, whereas the treatment group reached the peak (3.63) on day 12, which was one day later than the control group (Figure 2(b) and (d)). The age-specific reproductive value (Vx) curve showed a first upward and then downward trend. Compared with the control group, the treatment group displayed a decrease in Vx from day 0 to day 14, but an increase in Vx from day 15 to day 24 (Figure 2(c)). Nitenpyram treatment shortened the age stage specific life expectancy (exj) of nymphs in each instar and adult (Figure 3(b) and (d)). The age stage specific survival rate (sxj) curve of the control group displayed a later decrease in the late adult stage than that of the treatment group (Figure 3(a) and (c)).
In the G2, the lx of the control group was higher than that of the treatment group (Figure 4 (a)). The peak values of mx and lxmx curves were lower in the treatment group than in the control group (Figure 4(b) and (d)). The mx and lxmx values were lower in treatment group than in control group on day 6-15, but they were higher in the treatment group than the control group on day 16, 17, 19, and 21. The maximum Vx of the control group (14.39 on day 8) was higher than that of the treatment group (13.17 on day 9), occurring one day earlier than the treatment group (Figure 4 (c)). The exj of adults was shortened after treatment with nitenpyram(Figure 5(b) and (d)). In the adult stage, the sxj curve of the treatment group exhibited an earlier decrease than that of the control group (Figure 5 (a) and (c)).
3.4.16. S rRNA Sequencing Data and Sample Characteristics
After A. gossypii was directly or indirectly exposed to nitenpyram, the 16S rRNA gene amplicons in the bacterial communities of the G0, G1, and G2 were sequenced and analyzed. The bacterial diversity of the 16S V3-V4 hypervariable regions was detected in A. gossypii. A total of 79,587 raw reads were obtained from CK-G0, N-G0 (nitenpyram-treated G0), CK-G1, N-G1, CK-G2, and N-G2 samples, and a total of 462,365 clean reads were obtained after sequence filtering. The dilution curves of the six sample groups all tended to be stable with a Good’s coverage of 1 for each sample, indicating that the amount and sequencing depth of sequencing data were sufficient and sequencing data were qualified for subsequent analysis (Table 4, Figure 6 (a-c)).
3.5. Changes in Microbial Community of A. gossypii under Nitenpyram Treatment
Microbiome analysis showed that the bacterial community of A. gossypii was mainly Proteobacteria at phylum level, and it was Buchnera and Acinetobacter at the genus level (Table S1). The principal coordinates analysis (PCoA) was performed to investigate the differences in bacterial community at the genus level in the G0-G2 A. gossypii between control group and the treatment group (Figure 6 (d)). The results showed that in all the generations, the nitenpyram treatment group and the control group were significantly separated, indicating the differences in the bacterial community structure at the genus level. The LC20 nitenpyram stress changed significantly the bacterial community of G0-G1 A. gossypii, but the G2 treatment group and the G0 control group exhibited the similar bacterial communities. Further, α diversity analysis of bacterial community of A. gossypii was conducted. Chao1 and ACE data showed that after 48-hour nitenpyram treatment, the bacterial richness was increased in the G0 treatment group, but it was decreased in the offspring G1-G2 treatment group. Correspondingly, the species diversity represented by Shannon and Simpson indexes showed the same trend (Table 4).
The differential bacterial species in A. gossypii treated with nitenpyram at sublethal doses were analyzed based on the relative abundance clustering heatmap, and the 15 most abundant species were analyzed at the family level. The results showed that 48-hour nitenpyram treatment at sublethal concentrations affected the relative abundance of the top 15 symbiotic bacteria in A. gossypii. As shown in Figure 7(a) and Table S2, Morganellaceae is the dominant bacterial family, with a relative abundance of up to 95.98%. The relative abundance of Morganellaceae and Xanthomonadaceae in the G0 generation was lower in the treatment group than in the control group, while the relative abundance of Moraxellaceae and Sphingomonadaceae was higher in the treatment group. The relative abundance of Morganellaceae in the offspring (G1-G2) of A. gossypii was increased, compared with that in the control group, while the relative abundance of Moraxellaceae and Sphingomonadaceae was decreased, compared with that in the control group (Figure 7(a), Table S2).
At the genus level, the relative abundance of the observed genera in the treatment and control groups was evaluated. The results showed that the bacterial community of A. gossypii was mainly composed of Buchnera, Acinetobacter, Arsenophonus and Sphingomonas, with relative abundances accounting for 46.97%-88.14%, 0.04%-32.72%, 5.47%-10.19%, and 0.17%-6.18%, respectively. Compared with the control group, the relative abundance of Buchnera in the G0 A. gossypii in the LC20 nitenpyram treatment group was significantly reduced (Figure 8(a)), while that of Sphingomonas was significantly increased (Figure 8(b)). After G0 was treated with nitenpyram, the relative abundance of Buchnera was increased in both their G1 and G2 A. gossypii, while the relative abundance of Sphingomonas was decreased in two consecutive generations of their offsprings. The relative abundance of Arsenophonus exhibited no significant difference between the control group and treatment group. The relative abundance of Acinetobacter was increased in the G0 in the treatment group, and the relative abundance of their G1and G2 was decreased, compared with that in the control group, but the difference was not statistically significant (Figure 7(b), Table S3).
The biomarkers of all the A. gossypii samples were analyzed through LEfSe (LDA score > 2.0) to investigate the composition of biological communities of differential species and nitenpyram-associated bacteria. After G0 was treated with nitenpyram, Acinetobacter, Pseudomonas, Paracoccus, Bacteroides, Sphingomonas, Dongia, and Massilia were identified in the treatment group, while Buchnera was identified in the control group. The abundance of Comamonas, Acinetobacter, Sphingomonas, Escherichia_Shigella, Muribaculaceae, Lactobacillus, Blautia, Paracoccus, and Arsenophonus was significantly increased in the G1 control group, whereas the abundance of Buchnera was increased in nitenpyram-treated G1. In addition, Buchnera and Arsenophonus were identified as the biomarkers of the G2 treatment group, while Acinetobacter, Blautia, Pseudomonas, and Stenotrophomonas as the biomarkers of the G2 control group (Figure 9 (a-c)).
4. Discussion
In the natural environment, pesticides continue to degrade after their initial use, and individual insects are constantly exposed to these pesticides, causing sublethal effects on various physiology and behaviors such as fecundity, lifespan, immunity, and biochemistry[15,40,41]. Which further causes pest resurgence, and this is known as the adaptive mechanisms of organisms for their survival [42]. In this study, the population growth of A. gossypii was affected by LC20 nitenpyram treatment, and its symbiotic bacterial community also changed significantly.
In this study, the fecundity and lifespan of G0 A. gossypii adults were significantly reduced after 48-hour direct exposure to LC20 nitenpyram. Similar adverse effects have been reported when Schizaphis graminum is exposed to sublethal concentrations of flonicamid [21]. Consistently, imidacloprid at LC15, LC25, and LC35 significantly reduced the fecundity, lifespan, and reproductive period of the G0 adults of Metopolophium dirhodum [43]. The results showed that nitenpyram inhibited the growth of the G0 population of A. gossypii.
The 48-hour nitenpyram treatment on the parent G0 adults resulted in transgenerational sublethal effects on the development period, the adult pre-oviposition period (APOP), and the total pre-ovipositional period (TPOP), and other parameters of the offspring G1-G2. After the parent G0 adults were exposed to pesticides, the development period of each instar of the G1-G2 was prolonged. This developmental delay might be a detoxification mechanism at the expense of development and reproduction[44,45]. For example, LC25 flupyradifurone significantly prolonged the 4th instar nymphal period and adult period of the F1 generation of cotton aphid [46],and LC10 and LC25 broflanilide significantly prolonged the 3rd instar nymphal period of Myzus persicae [47]. In this study, compared with those in the control group, the fecundity and life span, and APOP of the G1-G2 in nitenpyram treatment group were significantly reduced.
In addition, the population growth parameter analysis results showed that the values of rm, λ, and R0 of the G1-G2 in nitenpyram treatment group were significantly lower than those in the control group. On the contrary, under the stress of sublethal concentration of nitenpyram, the mean generation time (T) of A. gossypii was significantly higher than that in the control group. This might be attributed to the energy shortage induced by insects’ adaptation to the pesticide stress under the exposure to pesticides [48]. Furthermore, the reduced fecundity and lifespan indicate hormesis deficiency, which is an important manifestation of the sublethal effects of pesticides [46]. Similarly, sublethal concentration of afidopyropen has been reported to significantly reduce the key parameters rm, λ, R0, lifespan, and fecundity of the offspring population of Aphis gossypii [49]. The sublethal effects of afidopyropen on Myzus persicae at sublethal concentrations is in line with our results [50]. Therefore, it could be concluded that sublethal concentrations of nitenpyram can change the population structure of A. gossypii offspring G1-G2 and slow down their population growth. However, there are also different findings in the previous studies. For example, after treatment with sublethal concentrations of imidacloprid, insecticide-induced hormesis is detected in the progeny of Metopolophium dirhodum[43] and A. gossypii [51], indicating that imidacloprid treatment significantly promoted the population growth of progeny aphids. This reported difference in the sublethal effects of pesticides on insects may be attributed to the differences in the structure and efficacy of different types of pesticides, different application times and concentrations of the same type of pesticides, different insect species, actual application conditions, and others [52,53,54].
In contrast, r, λ, R0, and fecundity of G3 adults were increased after treatment of the parent A. gossypii G0 with sublethal concentration of nitenpyram, but there were no significant differences between treatment group and the control group. The population growth rate of A. gossypii G3 was increased, compared with that of G1 and G2, which might be a compensation mechanism to restore the initial population density after the mother generation was treated with nitenpyram [55]. This result indicated that the G3 population of A. gossypii returned to a stable state.
Insect symbiont interactions are critical for insects’ survival and growth [56,57,58]. It has been widely reported that insect-related symbiotic bacteria are involved in the detoxification of chemical pesticides [59,60,61]. In this study, the bacterial community of three successive generations (G0-G2) of A. gossypii was mainly composed of Proteobacteria, Bacteroidota, and Firmicutes, and Proteobacteria, Morganellaceae, and Buchnera are the dominant bacteria at the phylum, family, and genus levels respectively. Our result is consistent with the bacterial composition of cotton aphids reported in previous studies [51,62]. Our results also showed that nitenpyram treatment significantly reduced the relative abundance of Buchnera in the G0 A. gossypii after 48 h, but increased the relative abundance of this genus in G1 and G2, compared with the control group (Table S3). Numerous studies have shown that Buchnera, as the primary symbiont of aphids, affects the nutritional metabolism and normal development of the host [63,64], and it is also involved in the development of insecticide resistance [65,66]. Our data showed the changes in the relative abundance of Buchnera in three consecutive generations (G0-G2), which might be attributed to the stimulation of pesticides, or the reduction in population adaptability of A. gossypii.
Sphingomonas is a kind of degradation bacteria [67], and its relative abundance in the intestine of imidacloprid-resistant cotton aphids is higher than that of imidacloprid-sensitive cotton aphids. Sphingomonas isolated from the intestine of imidacloprid resistant cotton aphids can effectively metabolize imidacloprid [68]. In this study, nitenpyram treatment significantly increased the relative abundance of Sphingomonas in the G0, but decreased it in the G1 and G2 (Table S3). The direct exposure of A. gossypii to nitenpyram increased the relative abundance of Sphingomonas in the G0, which might be due to the stress response of A. gossypii to nitenpyram, and Sphingomona might be involved in the development of resistance to nitenpyram in A. gossypii. The reduced relative abundance of Sphingomonas in offspring A. gossypii might be affected by the balance of the host bacterial community.
5. Conclusions
Our study confirmed that low concentration nitenpyram (LC20) had a transgenerational sublethal effect on A. gossypii. Treatment with nitenpyram had a negative impact on the fitness of its population, in which the fecundity of G1 and G2 A. gossypii was decreased, and the average generation time (T) was increased. Likewise, nitenpyram also affected the bacterial community of A. gossypii. The relative abundance of the endosymbiotic bacteria Buchnera and Sphingomonas changed significantly in three consecutive generations (G0-G2). Furthermore, different biomarkers were identified in different generations. In summary, nitenpyram can affect the biological characteristics of cotton aphid and is closely related to bacterial symbionts. Our findings contribute to understanding the adaptability of A. gossypii to pesticides and the relationship between cotton aphids and symbiotic bacteria under insecticide stress, and provide theoretical basis for future comprehensive management of cotton aphids.
Supplementary Materials
Effect of nitenpyram symbiotic bacteria of A. gossypii G0-G2 generation (PDF).
Author Contributions
Conceptualization, Y.Y.; methodology, Y.W. and Y.S.; software, Y.W, W. L. and T. C.; validation, Y.W. and Y.S.; formal analysis, Y.W. and X.H.; investigation, Y.W. and Y.S.; resources, Y.W.; data curation, Y.W. and W.L.; writing—original draft preparation, Y.W.; writing—review and editing, Y.Y., Y.W. and X.H.; visualization, Y.W.; supervision, Y.S.; project administration, Y.Y.; funding acquisition, Y.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was sponsored by the National Natural Science Foundation of China (No. 32360671) and the National Key R&D Program of China (2022YFD1400300) .
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data Availability Statement: No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Müller, F.P. Incidence of the aphid Acyrthosiphon gossypii Mordvilko on legumes and on cotton (Homoptera: Aphididae). Beitrage Entomol 1975, 25, 257–260. [Google Scholar]
- Gao, G.Z.; Perkins, L.E.; Zalucki, M.P.; Lu, Z.Z.; Ma, J.H. Effect of temperature on the biology of Acyrthosiphon gossypii Mordvilko (Homoptera: Aphididae) on cotton. J. Pest Sci. 2013, 86, 167–172. [Google Scholar] [CrossRef]
- Lu, Z.Z.; Tian, C.Y.; Song, Y.D. Relationship between Aphis gossypii and Acyrthosiphon gossypii on cotton in Xinjiang. China Cotton 2002, 29, 11–12. [Google Scholar]
- Liu, D.; Jiang, H.X.; Wang, Z.F.; Cao, Y.; Zhang, S.E.; Zhai, G.Y. The prevention and control of alfalfa aphid. Shandong J. Anim. Husb. Vet 2012, 33, 94–96. [Google Scholar]
- Bass, C.; Puinean, A.M.; Zimmer, C.T.; Denholm, I.; Field, L.M.; Foster, S.P.; Gutbrod, O.; Nauen, R.; Slater, R.; Williamson, M.S. The evolution of insecticide resistance in the peach potato aphid, Myzus persicae. Insect Biochem. Mol. Biol. 2014, 51, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Mirza, F.K.; Yarahmadi, F.; Jalal Abadi, A.L.; Meraaten, A.A. Enzymes mediating resistance to chlorpyriphos in Aphis fabae (Homoptera: Aphididae). Ecotoxicol. Environ. Saf. 2020, 206, 111335. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.; Tian, Y.A.; Biondi, A.; Desneux, N.; Gao, X.W. Short-term and transgenerational effects of the neonicotinoid nitenpyram on susceptibility to insecticides in two whitefly species. Ecotoxicology 2012, 21, 1889–1898. [Google Scholar] [CrossRef] [PubMed]
- Elbert, A.; Haas, M.; Springer, B.; Thielert, W.; Nauen, R. Applied aspects of neonicotinoid uses in crop protection. Pest Manage. Sci. 2008, 64, 1099–1105. [Google Scholar] [CrossRef]
- Minamida, I.; Iwanaga, K.; Tabuchi, T.; Uneme, H.; Dantsuji, H.; Oksuchi, T. Synthesis and insecticidal activity of acyclic nitroethene compounds containing a 3-pyridylmethylamino group. J Pestic Sci. 1993, 18, 31–40. [Google Scholar] [CrossRef]
- Bass, C.; Denholm, I.; Williamson, M.S.; Nauen, R. The global status of insect resistance to neonicotinoid insecticides. Pestic. Biochem. Physiol. 2015, 121, 78–87. [Google Scholar] [CrossRef]
- Wang, S.Y.; Qi, Y.F.; Desneux, N.; Shi, X.Y.; Biondi, A.; Gao, X.W. Sublethal and transgenerational effects of short-term and chronic exposures to the neonicotinoid nitenpyram on the cotton aphid Aphis gossypii. J Pest Sci. 2017, 90, 389–396. [Google Scholar] [CrossRef]
- Zhu, X.H.; Wei, Q.; Wan, P.J.; Wang, W.X.; Lai, F.X.; He, J.C.; Fu, Q. Effect of Paclobutrazol Application on Enhancing the Efficacy of Nitenpyram against the Brown Planthopper, Nilaparvata lugens. Int. J. Mol. Sci. 2023, 24, 10490. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Mao, K.; Jin, R.; Cai, T.; Qin, Y.; Zhang, Y.; He, S.; Ma, K.; Wan, H.; Ren, X.; et al. miRNA novel_268 targeting NlABCG3 is involved in nitenpyram and clothianidin resistance in Nilaparvata lugens. Int. J. Biol. Macromol. 2022, 217, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Stapel, J.O.; Cortesero, A.M.; Lewis, W.J. Disruptive sublethal effects of insecticides on biological control: altered foraging ability and life span of a parasitoid after feeding on extrafloral nectar of cotton treated with systemic insecticides. Biol. Control 2000, 17, 243–249. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Desneux, N.; Rafalimanana, H.; Kaiser, L. Dose–response relationship in lethal and behavioural effects of different insecticides on the parasitic wasp Aphidius ervi. Chemosphere 2004, 54, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Ceuppens, B.; Eeraerts, M.; Vleugels, T.; Cnops, G.; Roldan-Ruiz, I.; Smagghe, G. Effects of dietary lambda-cyhalothrin exposure on bumblebee survival, reproduction, and foraging behavior in laboratory and greenhouse. J Pest Sci. 2015, 88, 777–783. [Google Scholar] [CrossRef]
- Wu, H.M.; Feng, H.L.; Wang, G.D.; Zhang, L.L.; Zulu, L.; Liu, Y.H.; Zheng, Y.L.; Rao, Q. Sublethal effects of three insecticides on development and reproduction of Spodoptera frugiperda (Lepidoptera: Noctuidae). Agronomy 2022, 12, 1334. [Google Scholar] [CrossRef]
- Cheng, Z.; Qin, Q.; Wang, D.; Han, S.; Zhang, S.; He, Y. Sublethal and transgenerational effects of exposures to the thiamethoxam on the seven-spotted lady beetle, Coccinella septempunctata L. (Coleoptera: Coccinellidae). Ecotoxicol. Environ. Saf. 2022, 243, 114002. [Google Scholar] [CrossRef]
- Fouad, E.A.; El-Sherif, S.A.N.; Mokbel, E.S.M.S. Flupyradifurone induces transgenerational hormesis effects in the cowpea aphid, Aphis craccivora. Ecotoxicology 2022, 31, 909–918. [Google Scholar] [CrossRef]
- Gul, H.; Ul Haq, I.; Ullah, F.; Khan, S.; Yaseen, A.; Shah, S.H.; Tariq, K.; Güncan, A.; Desneux, N.; Liu, X. Impact of sublethal concentrations of flonicamid on key demographic parameters and feeding behavior of Schizaphis graminum. Ecotoxicology 2023, 32, 756–767. [Google Scholar] [CrossRef] [PubMed]
- Hadian, S.; Zandi-Sohani, N.; Yarahmadi, F.; Sohrabi, F. Determination of sub-lethal effects of spirotetramat and chlorpyrifos on Aenasius bambawalei Hayat (Hymenoptera: Encyrtidae) by using life table. Iran Agric. Res. 2022, 40, 33–40. [Google Scholar]
- Ullah, F.; Gul, H.; Tariq, K.; Desneux, N.; Gao, X.; Song, D. Thiamethoxam induces transgenerational hormesis effects and alteration of genes expression in Aphis gossypii. Pestic. Biochem. Physiol. 2020, 165, 104557. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Li, X.Q.; Wang, W.R.; Zhang, S.C.; Cui, J.J.; Peng, Y.; Zhao, Y. Impact of Sulfoxaflor Exposure on Bacterial Community and Developmental Performance of the Predatory Ladybeetle Propylea japonica. Microb Ecol. 2023, 86, 1226–1239. [Google Scholar] [CrossRef] [PubMed]
- Shang, J.; Yao, Y.S.; Chen, L.L.; Zhu, X.Z.; Niu, L.; Gao, X.K.; Luo, J.Y.; Ji, J.C.; Cui, J.J. Sublethal exposure to deltamethrin stimulates reproduction and alters symbiotic bacteria in Aphis gossypii. J Agric Food Chem. 2021, 69, 15097–15107. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Lin, X.Y.; Guo, X.R. The role of insect symbiotic bacteria in metabolizing phytochemicals and agrochemicals. Insects 2022, 13, 583. [Google Scholar] [CrossRef]
- Cheng, D.F.; Guo, Z.J.; Riegler, M.; Xi, Z.Y.; Liang, G.W.; Xu, Y.J. Gut symbiont enhances insecticide resistance in a significant pest, the oriental fruit fly Bactrocera dorsalis (Hendel). Microbiome 2017, 5, 1–12. [Google Scholar] [CrossRef]
- Wang, H.Y.; Zhang, C.X.; Cheng, P.; Wang, Y.; Liu, H.M.; Wang, H.F.; Wang, H.W.; Gong, M.Q. Differences in the intestinal microbiota between insecticide-resistant and-sensitive Aedes albopictus based on full-length 16S rRNA sequencing. Microbiology Open 2021, 10, e1177. [Google Scholar] [CrossRef]
- Chen, B.; Zhang, N.; Xie, S.; Zhang, X.; He, J.; Muhammad, A.; Sun, C.; Lu, X.; Shao, Y. Gut bacteria of the silkworm Bombyx mori facilitate host resistance against the toxic effects of organophosphate insecticides. Environ. Int. 2020, 143, 105886. [Google Scholar] [CrossRef]
- Moores, G.D.; Gao, X.W.; Denholm, I.; Devonshire, A.L. Characterisation of insensitive acetylcholinesterase in insecticide-resistant cotton aphids, Aphis gossypii glover (homoptera: Aphididae). Pestic. Biochem. Physiol. 1996, 56, 102–110. [Google Scholar] [CrossRef]
- Chen, X.W.; Li, F.; Chen, A.Q.; Ma, K.S.; Liang, P.Z.; Liu, Y.; Song, D.I.; Gao, X.W. Both point mutations and low expression levels of the nicotinic acetylcholine receptor β1 subunit are associated with imidacloprid resistance in an Aphis gossypii (Glover) population from a Bt cotton field in China. Pestic. Biochem. Physiol. 2017, 141, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Johnson, R.W. An introduction to the bootstrap. Teach Stat. 2001, 23, 49–54. [Google Scholar] [CrossRef]
- Nossa, C.W.; Oberdorf, W.E.; Yang, L.Y.; Aas, J.A.; Paster, B.J.; DeSantis, T.Z.; Brodie, E.L.; Malamud, D.; Poles, M.A.; Pei, Z.H. Design of 16S rRNA gene primers for 454 pyrosequencing of the human foregut microbiome. World J. Gastroenterol. 2010, 16, 4135. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods. 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Du, P.Q.; Wu, X.H.; Xu, J.; Dong, F.S.; Liu, X.G.; Zheng, Y.Q. Effects of trifluralin on the soil microbial community and functional groups involved in nitrogen cycling. J. Hazard. Mater. 2018, 353, 204–213. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, 1–18. [Google Scholar] [CrossRef]
- Simon-Delso, N.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Chagnon, M.; Downs, C.; Furlan, L.; Gibbons, D.W.; Giorio, C.; Girolami, V. Systemic insecticides (neonicotinoids and fipronil): trends, uses, mode of action and metabolites. Environ. Sci. Pollut. Res. 2015, 22, 5–34. [Google Scholar] [CrossRef]
- Dai, C.; Ricupero, M.; Puglisi, R.; Lu, Y.; Desneux, N.; Biondi, A.; Zappalà, L. Can contamination by major systemic insecticides affect the voracity of the harlequin ladybird? Chemosphere 2020, 256, 126986. [Google Scholar] [CrossRef] [PubMed]
- Sial, M.U.; Zhao, Z.Z.; Zhang, L.; Zhang, Y.N.; Mao, L.G.; Jiang, H.Y. Evaluation of insecticides induced hormesis on the demographic parameters of Myzus persicae and expression changes of metabolic resistance detoxification genes. Sci. Rep. 2018, 8, 16601. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, Y.; Zhu, X.; Li, X.; Cheng, D.; Zhang, Y. Effects of imidacloprid-induced hormesis on the development and reproduction of the rose-grain aphid Metopolophium dirhodum(Hemiptera: Aphididae). Front Physiol. 2023, 14, 1113464. [Google Scholar] [CrossRef] [PubMed]
- Skouras, P.J.; Stathas, G.J.; Voudouris, C.C.; Darras, A.I.; Tsitsipis, J.A.; Margaritopoulos, J.T. Effect of synthetic insecticides on the larvae of Coccinella septempunctata from Greek populations. Phytoparasitica 2017, 45, 165–173. [Google Scholar] [CrossRef]
- Hannig, G.T.; Ziegler, M.; Marçon, P.G. Feeding cessation effects of chlorantraniliprole, a new anthranilic diamide insecticide, in comparison with several insecticides in distinct chemical classes and mode-of-action groups. Pest Manage. Sci. 2009, 65, 969–974. [Google Scholar] [CrossRef] [PubMed]
- Liang, P.Z.; Ma, K.S.; Chen, X.W.; Tang, C.Y.; Xia, J.; Chi, H.; Gao, X.W. Toxicity and Sublethal Effects of Flupyradifurone, a Novel Butenolide Insecticide, on the Development and Fecundity of Aphis gossypii (Hemiptera: Aphididae). J. Econ. Entomol. 2019, 112, 852–858. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.W.; Zeng, Q.H.; Yang, H.; Zhang, C.; Ding, B.; Yang, H.Z.; Yang, M.F. Sublethal and transgenerational effects of broflanilide on Myzus persicae (Sulzer)(Hemiptera: Aphididae). Crop Prot. 2023, 174, 106421. [Google Scholar] [CrossRef]
- Iftikhar, A.; Hafeez, F.; Aziz, M.A.; Hashim, M.; Naeem, A.; Yousaf, H.K.; Saleem, M.J.; Hussain, S.; Hafeez, M.; Ali, Q.; et al. Assessment of sublethal and transgenerational effects of spirotetramat, on population growth of cabbage aphid,Brevicoryne brassicae L. (Hemiptera: Aphididae). Front Physiol. 2022, 13, 1014190. [Google Scholar] [CrossRef]
- Tang, Q.L.; Liang, P.Z.; Li, J.H.; Gao, X.W. A sublethal concentration of afidopyropen suppresses the population growth of the cotton aphid, Aphis gossypii Glover (Hemiptera: Aphididae). J. Integr. Agric. 2022, 21, 2055–2064. [Google Scholar]
- Liu, X.; Fu, Z.; Zhu, Y.; Gao, X.; Liu, T.X.; Liang, P. Sublethal and transgenerational effects of afidopyropen on biological traits of the green peach aphid Myzus persicae (Sluzer). Pestic. Biochem. Physiol. 2022, 180, 104981. [Google Scholar] [CrossRef]
- Wei, Y.D.; Su, Y.; Han, X.; Guo, W.F.; Zhu, Y.; Yao, Y.S. Evaluation of Transgenerational Effects of Sublethal Imidacloprid and Diversity of Symbiotic Bacteria on Acyrthosiphon gossypii. Insects 2023, 14, 427. [Google Scholar] [CrossRef] [PubMed]
- Han, W.S.; Zhang, S.F.; Shen, F.Y.; Liu, M.; Ren, C.C.; Gao, X.W. Residual toxicity and sublethal effects of chlorantraniliprole on Plutella xylostella (lepidoptera: plutellidae). Pest Manage. Sci. 2012, 68, 1184–1190. [Google Scholar] [CrossRef] [PubMed]
- Afza, R.; Afzal, A.; Riaz, M.A.; Majeed, M.Z.; Idrees, A.; Qadir, Z.A.; Afzal, M.; Hassan, B.; Li, J. Sublethal and transgenerational effects of synthetic insecticides on the biological parameters and functional response of Coccinella septempunctata (Coleoptera: Coccinellidae) under laboratory conditions. Front Physiol. 2023, 14, 1088712. [Google Scholar] [CrossRef] [PubMed]
- Xiao, D.; Yang, T.; Desneux, N.; Han, P.; Gao, X.W. Assessment of Sublethal and Transgenerational Effects of Pirimicarb on the Wheat Aphids Rhopalosiphum padi and Sitobion avenae. PloS one 2015, 10, e0128936. [Google Scholar] [CrossRef]
- Szabó, B.; Seres, A.; Bakonyi, G. Distinct changes in the life-history strategies of Folsomia candida Willem (Collembola: Isotomidae) due to multi-and transgenerational treatments with an insecticide. Appl Soil Ecol. 2020, 152, 103563. [Google Scholar] [CrossRef]
- Wang, H.; Xian, X.Q.; Gu, Y.Y.; Castañé, C.; Arnó, J.; Wu, S.R.; Wan, F.H.; Liu, W.X.; Zhang, G.F.; Zhang, Y.B. Similar bacterial communities among different populations of a newly emerging invasive species, Tuta absoluta (Meyrick). Insects 2022, 13, 252. [Google Scholar] [CrossRef] [PubMed]
- Naveed, W.A.; Liu, Q.; Lu, C.C.; Huang, X.L. Symbiotic Bacterial Communities of Insects Feeding on the Same Plant Lineage: Distinct Composition but Congruent Function. Insects 2024, 15, 187. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.M.; Li, D.L.; Desneux, N.; Gatti, J.L.; Hu, Z.Q.; Luo, C. Facultative symbiont provides fitness benefits to the grain aphid, but not to parasitoid offspring. Entomol Gen. 2024, 44, 163–170. [Google Scholar] [CrossRef]
- Ye, Q.T.; Gong, X.; Liu, H.H.; Wu, B.X.; Peng, C.W.; Hong, X.Y.; Bing, X.L. The symbiont Wolbachia alleviates pesticide susceptibility in the two-spotted spider mite Tetranychus urticae through enhanced host detoxification pathways. Insect Sci. 2024. [Google Scholar] [CrossRef]
- Xia, X.; Sun, B.; Gurr, G.M.; Vasseur, L.; Xue, M.; You, M. Gut Microbiota Mediate Insecticide Resistance in the Diamondback Moth, Plutella xylostella(L.). Front Microbiol. 2018, 9, 25. [Google Scholar] [CrossRef]
- Zeng, B.; Zhang, F.; Liu, Y.T.; Wu, S.F.; Bass, C.; Gao, C.F. Symbiotic bacteria confer insecticide resistance by metabolizing buprofezin in the brown planthopper, Nilaparvata lugens (Stål). PLoS Pathog. 2023, 19, e1011828. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Luo, J.; Wang, L.; Zhang, L.; Zhu, X.; Jiang, W.; Cui, J. Bacterial communities in natural versus pesticide-treated Aphis gossypii populations in North China. Microbiology Open 2019, 8, e00652. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.F.; Ren, Y.J.; Chen, J.C.; Cao, L.J.; Qiao, G.H.; Zong, S.X.; Hoffmann, A.A.; Wei, S.J.; Yang, Q. Effects of fungicides on fitness and Buchnera endosymbiont density in Aphis gossypii. Pest Manage. Sci. 2023, 79, 4282–4289. [Google Scholar] [CrossRef] [PubMed]
- Shigenobu, S.; Wilson, A.C.C. Genomic revelations of a mutualism: the pea aphid and its obligate bacterial symbiont. Cell. Mol. Life Sci. 2011, 68, 1297–1309. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.K.; Gong, Y.J.; Chen, J.C.; Shi, P.; Cao, L.J.; Yang, Q.; Hoffmann, A.A.; Wei, S.J. Increased density of endosymbiotic Buchnera related to pesticide resistance in yellow morph of melon aphid. J. Pest Sci. 2020, 93, 1281–1294. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, C.Y.; Li, R.; Liang, P.Z.; Gu, S.H.; Zhang, L.; Gao, X.W. Endosymbiosis change under the stress of omethoate and four plant allelochemicals in cotton aphid, Aphis gossypii Glover (Hemiptera: Aphididae). 2023.
- Pinyakong, O.; Habe, H.; Kouzuma, A.; Nojiri, H.; Yamane, H.; Omori, T. Isolation and characterization of genes encoding polycyclic aromatic hydrocarbon dioxygenase from acenaphthene and acenaphthylene degrading Sphingomonas sp. strain A4. FEMS Microbiol. Lett. 2004, 238, 297–305. [Google Scholar]
- Lv, N.; Li, R.; Cheng, S.; Zhang, L.; Liang, P.; Gao, X. The gut symbiont Sphingomonas mediates imidacloprid resistance in the important agricultural insect pest Aphis gossypii Glover. BMC Biol. 2023, 21, 86. [Google Scholar] [CrossRef]
Figure 1.
The fecundity (a) and longevity (b) of A. gossypii (G0 generation) directly exposed for 48 h to LC20 of nitenpyram. Asterisk above the error bars show significant differences at P< 0.05 (t-test).
Figure 1.
The fecundity (a) and longevity (b) of A. gossypii (G0 generation) directly exposed for 48 h to LC20 of nitenpyram. Asterisk above the error bars show significant differences at P< 0.05 (t-test).
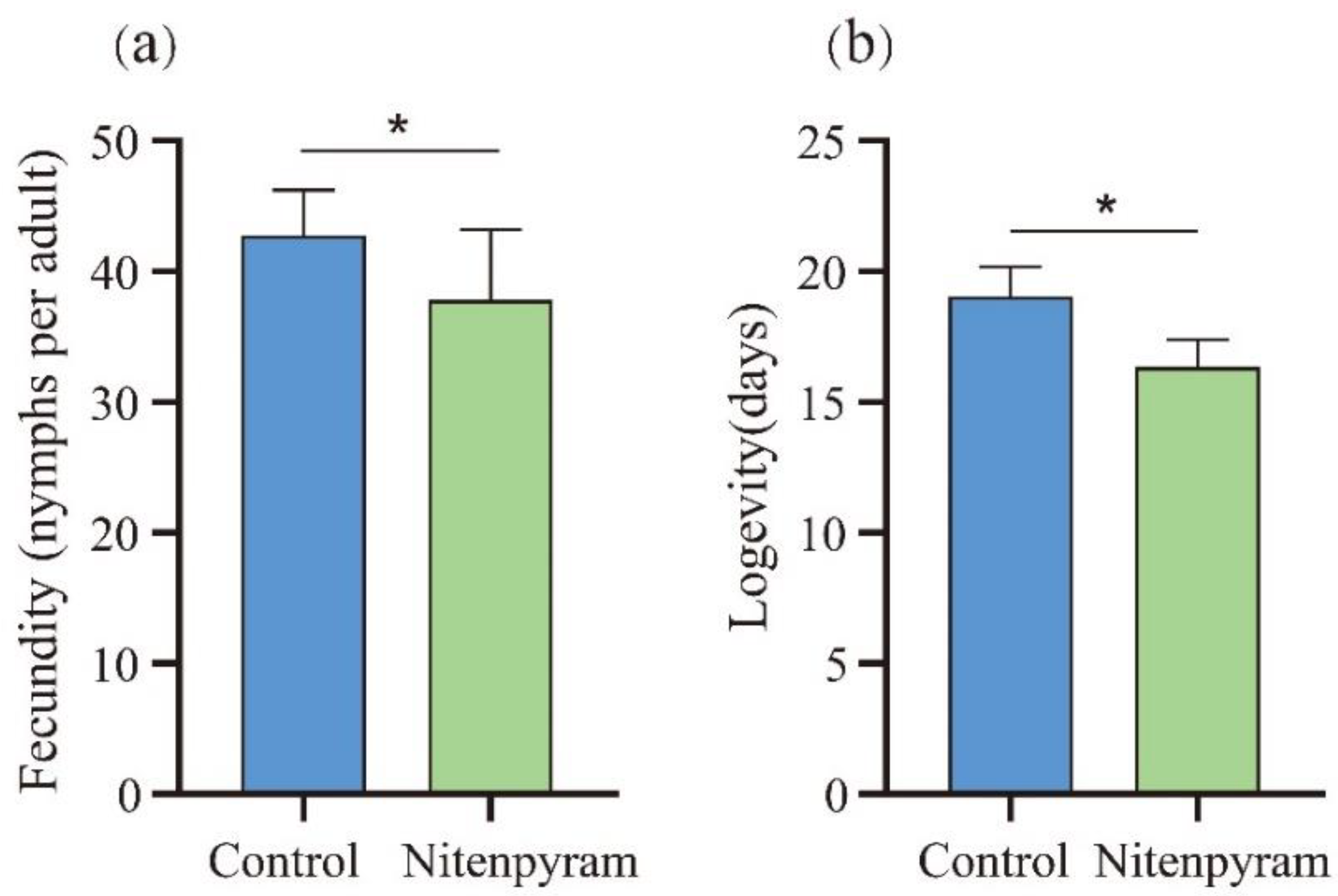
Figure 2.
Age-specific survival rate (lx), Age-specific fecundity (mx), Age-specific maternity (lxmx) , and Age-specific reproductive value (Vx) of A. gossypii in G1 generation after G0 treated with LC20 of nitenpyram.
Figure 2.
Age-specific survival rate (lx), Age-specific fecundity (mx), Age-specific maternity (lxmx) , and Age-specific reproductive value (Vx) of A. gossypii in G1 generation after G0 treated with LC20 of nitenpyram.
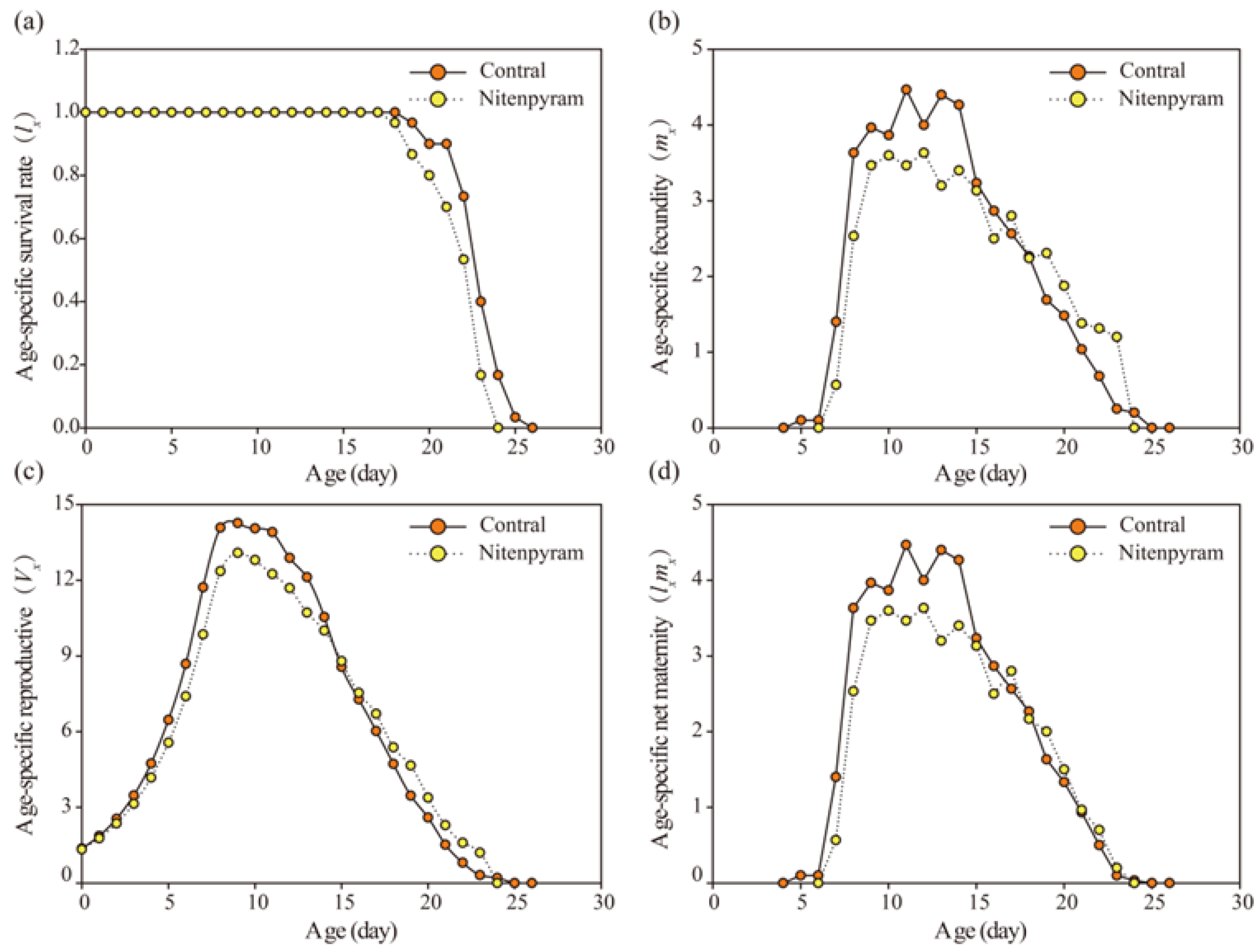
Figure 3.
Age-stage-specific survival rates (sxj) and Age-stage-specific life expectancy (exj) of A. gossypii in G1 generation after G0 treated with LC20 of nitenpyram. Figures (a) and (b) are the control group, and Figures (c) and (d) are the nitenpyram group.
Figure 3.
Age-stage-specific survival rates (sxj) and Age-stage-specific life expectancy (exj) of A. gossypii in G1 generation after G0 treated with LC20 of nitenpyram. Figures (a) and (b) are the control group, and Figures (c) and (d) are the nitenpyram group.
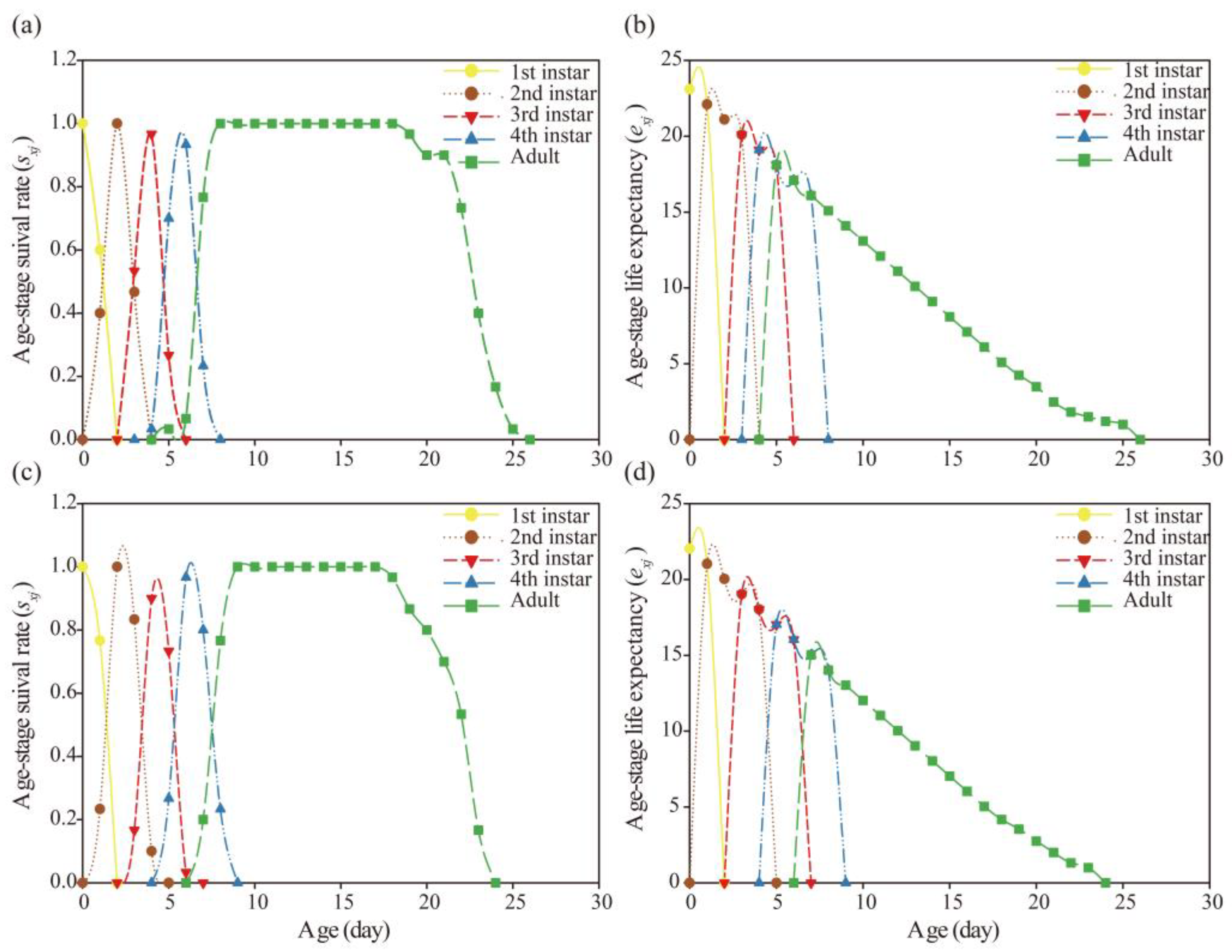
Figure 4.
Age-specific survival rate (lx), Age-specific fecundity (mx), Age-specific maternity (lxmx) , and Age-specific reproductive value (Vx) of A. goss ypii in G2 generation after G0 treated with LC20 of nitenpyram.
Figure 4.
Age-specific survival rate (lx), Age-specific fecundity (mx), Age-specific maternity (lxmx) , and Age-specific reproductive value (Vx) of A. goss ypii in G2 generation after G0 treated with LC20 of nitenpyram.
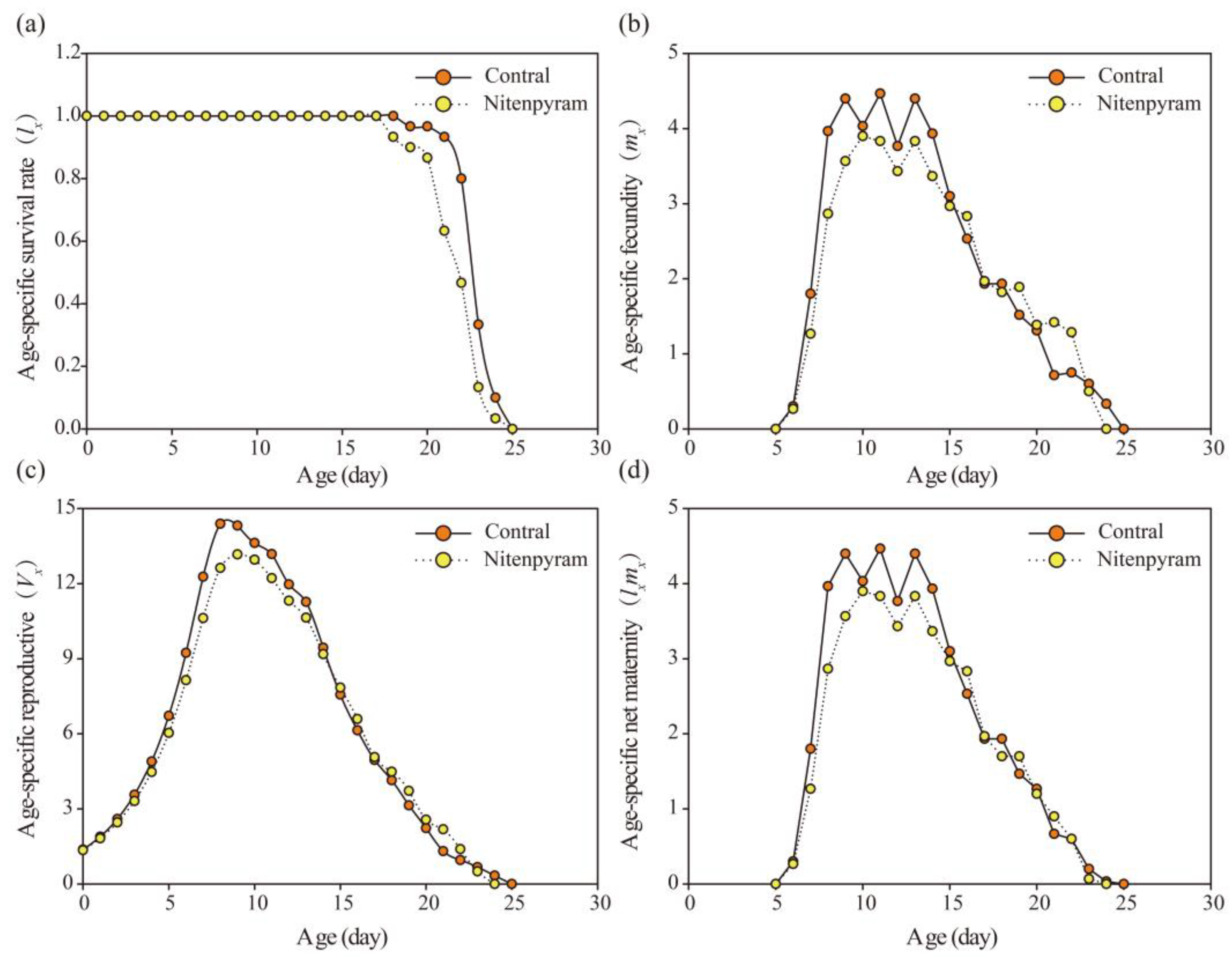
Figure 5.
Age-stage-specific survival rates (sxj) and Age-stage-specific life expectancy (exj) of A. gossypii in G2 generation after G0 treated with LC20 of nitenpyram. Figures (a) and (b) are the control group, and Figures (c) and (d) are the nitenpyram group.
Figure 5.
Age-stage-specific survival rates (sxj) and Age-stage-specific life expectancy (exj) of A. gossypii in G2 generation after G0 treated with LC20 of nitenpyram. Figures (a) and (b) are the control group, and Figures (c) and (d) are the nitenpyram group.
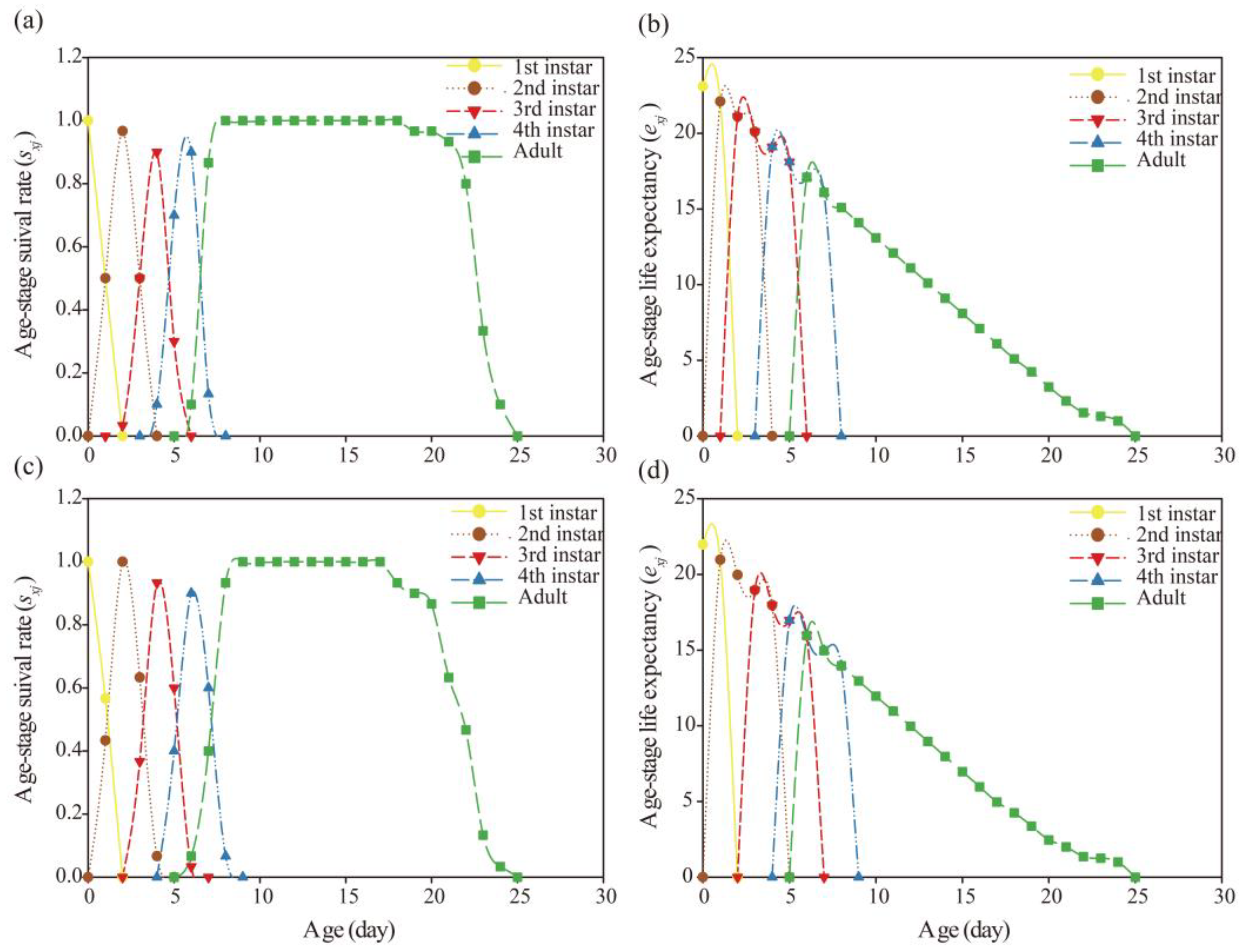
Figure 6.
Rarefaction curves, Chao1diversity index and sample clustering analysis plot of bacterial communities in A. gossypii G0-G2. (a) Observed species. (b) Goods coverage diversity index. (c) Chao1 diversity index. (d) Principal coordinate analysis (PCoA) G0-G2 of bacterial communities of A. gossypii exposed to nitenpyram. Green, yellow and red circles represent control group (G0-G2) respectively. Purple, blue and brown circles represent treatment group (G0-G2) respectively.
Figure 6.
Rarefaction curves, Chao1diversity index and sample clustering analysis plot of bacterial communities in A. gossypii G0-G2. (a) Observed species. (b) Goods coverage diversity index. (c) Chao1 diversity index. (d) Principal coordinate analysis (PCoA) G0-G2 of bacterial communities of A. gossypii exposed to nitenpyram. Green, yellow and red circles represent control group (G0-G2) respectively. Purple, blue and brown circles represent treatment group (G0-G2) respectively.
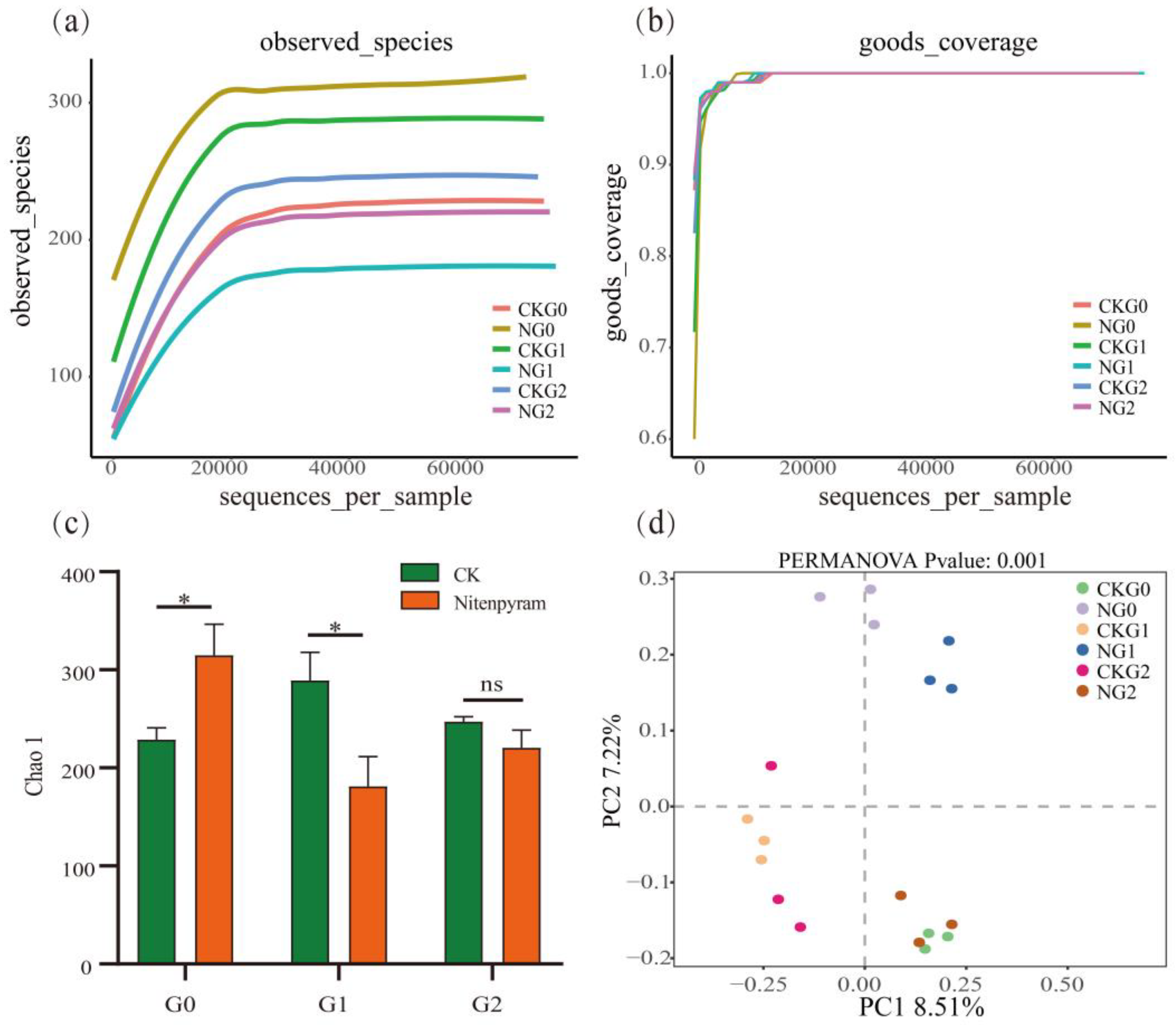
Figure 7.
Heat map of bacterial communities of A. gossypii to at different taxonomic levels after nitenpyram expose (a, b). (a) Heat map of the top 15 abundant symbiotic bacteria of A. gossypii in three successive generations at the family level. (b) Heat map of the top 15 abundant symbiotic bacteria of A. gossypii in three successive generations at the genus level. Orange indicates a high relative abundance of species, while blue indicates a low relative abundance of species. Generate heat map based on the data in Table S3.
Figure 7.
Heat map of bacterial communities of A. gossypii to at different taxonomic levels after nitenpyram expose (a, b). (a) Heat map of the top 15 abundant symbiotic bacteria of A. gossypii in three successive generations at the family level. (b) Heat map of the top 15 abundant symbiotic bacteria of A. gossypii in three successive generations at the genus level. Orange indicates a high relative abundance of species, while blue indicates a low relative abundance of species. Generate heat map based on the data in Table S3.
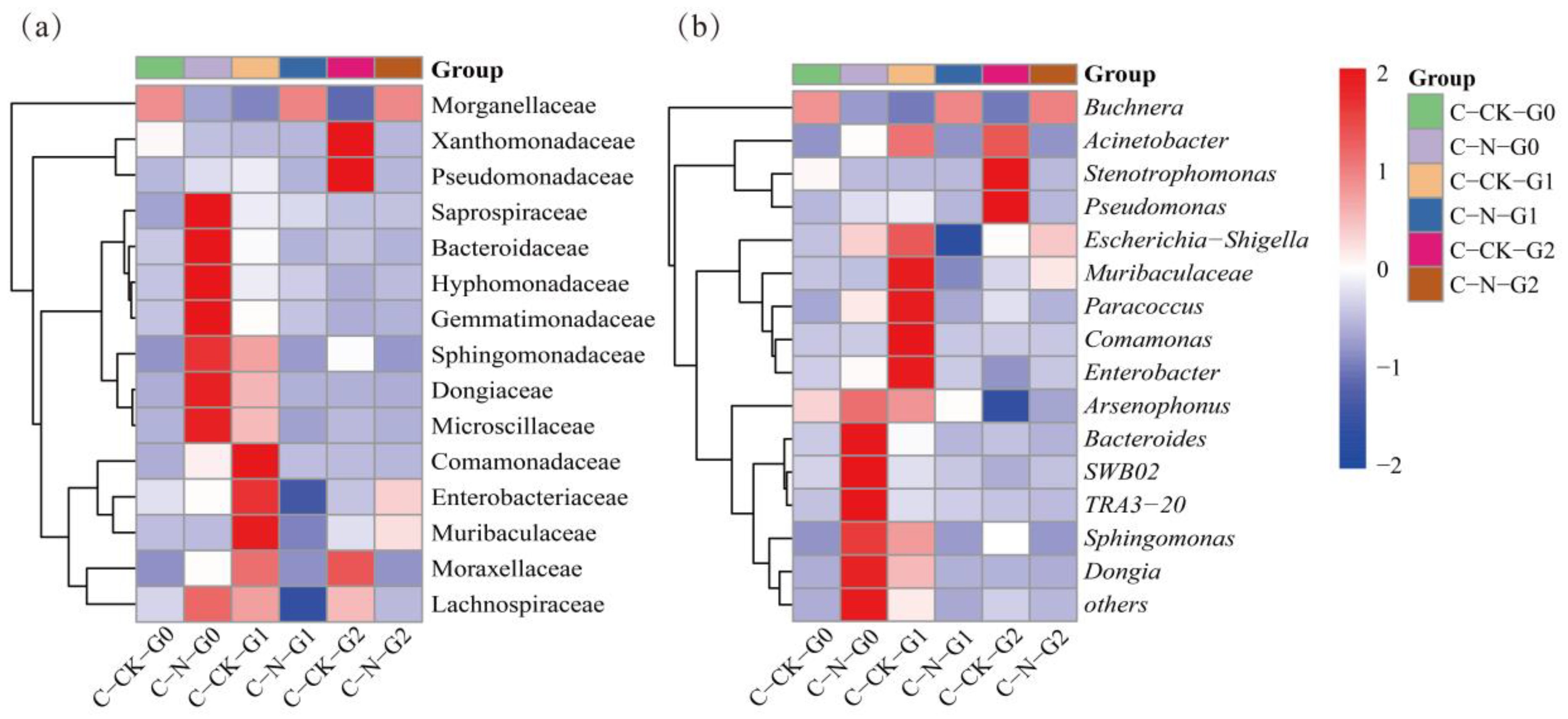
Figure 8.
Comparison of relative abundance of two typical symbiotic bacteria in A. gossypii control group and nitenpyram treatment group. (a) The relative abundance of Buchnera was significant different between the treatment group and the control group in G0 and G1 generation. (b) The relative abundance of Sphingomonas was significant different between the treatment group and the control group in G0 generation. An asterisk indicates a significant difference between the control group and the treatment group at the P < 0.05 level (t-test).
Figure 8.
Comparison of relative abundance of two typical symbiotic bacteria in A. gossypii control group and nitenpyram treatment group. (a) The relative abundance of Buchnera was significant different between the treatment group and the control group in G0 and G1 generation. (b) The relative abundance of Sphingomonas was significant different between the treatment group and the control group in G0 generation. An asterisk indicates a significant difference between the control group and the treatment group at the P < 0.05 level (t-test).
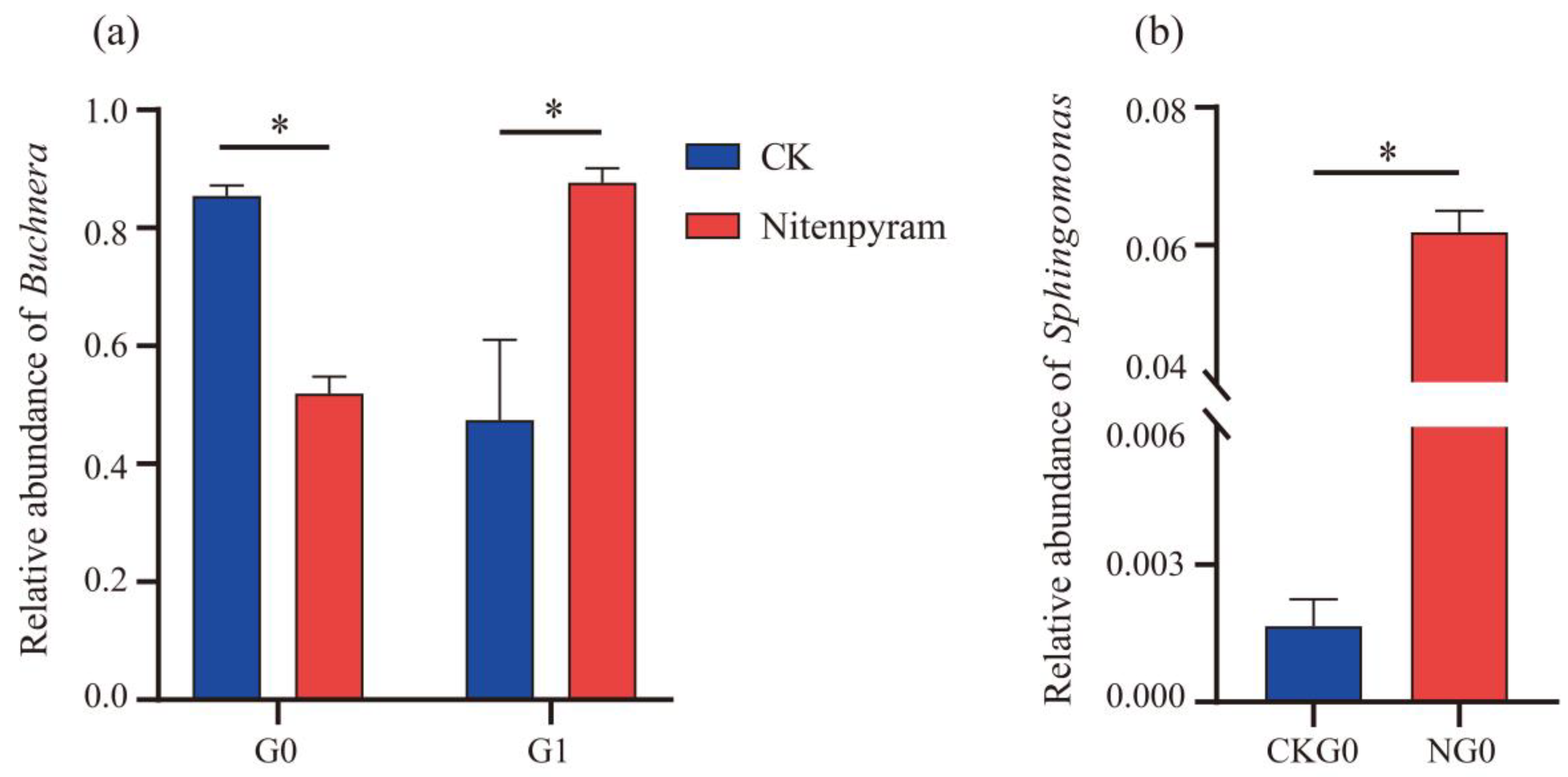
Figure 9.
Score map of different species of bacterial community of the A. gossypii (G0-G2). The green bars represent species that were relatively abundant in the control group, and the purple bars represent species that were relatively abundant in the treatment group.
Figure 9.
Score map of different species of bacterial community of the A. gossypii (G0-G2). The green bars represent species that were relatively abundant in the control group, and the purple bars represent species that were relatively abundant in the treatment group.
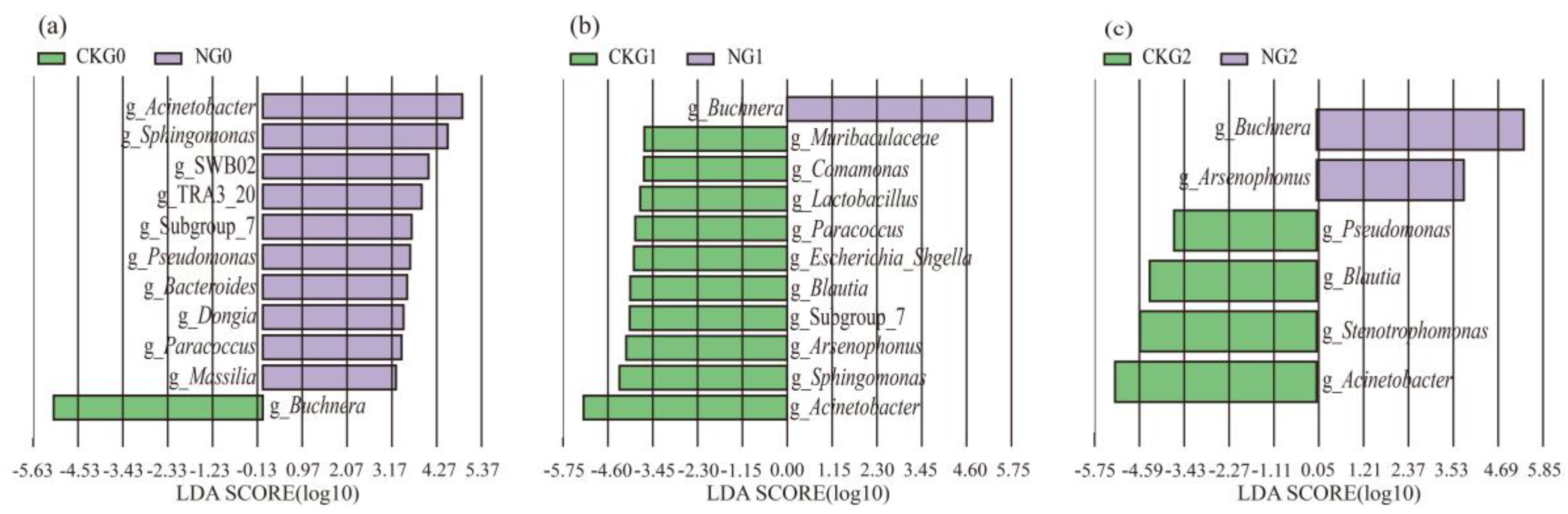

Table 1.
Acute toxicity of nitenpyram to A. gossypii adults after 48 h exposure.
| Treatment | Slope ± SEa | LC20 (mg·L−1) (95% CL)b |
LC50 (mg·L−1) (95% CL)b |
R2 |
|---|---|---|---|---|
| Nitenpyram | 1.41±0.31 | 2.49 (0.78-4.31) |
10.12 (6.34-17.08) |
0.98 |
a Standard error. b 95% confidence intervals.
Table 2.
Effect of nitenpyram LC20 on life history parameters of A. gossypii in the G1-G3 generation.
Table 2.
Effect of nitenpyram LC20 on life history parameters of A. gossypii in the G1-G3 generation.
| Treatments | First instar (d) | P | Second instar (d) | P | Third instar (d) | P | Fourth instar (d) | P | Pre-adult (d) | P | APOP (d) | P | TPOP (d) | P | Longevity (d) | P | Fecundity | P | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G1 | Control | 1.60±0.09 | 0.20 | 1.87±0.09 | 0.02 | 1.77±0.09 | 0.56 | 1.90±0.09 | 0.01 | 7.13±0.11 | <0.001 | 0.27±0.08 | <0.001 | 7.40±0.14 | <0.001 | 23.10±0.28 | 0.01 | 45.67±1.10 | <0.001 |
| Nitenpyram | 1.77±0.08 | 2.17±0.08 | 1.83±0.07 | 2.27±0.10 | 8.03±0.12 | 0.00±0.00 | 8.03±0.12 | 22.03±0.31 | 39.83±1.20 | ||||||||||
| G2 | Control | 1.50±0.09 | 0.70 | 1.97±0.11 | 0.19 | 1.73±0.08 | 0.08 | 1.83±0.08 | 0.29 | 7.03±0.09 | <0.001 | 0.13±0.06 | 0.69 | 7.17±0.11 | 0.004 | 23.10±0.22 | 0.002 | 44.80±0.99 | 0.001 |
| Nitenpyram | 1.57±0.09 | 2.13±0.06 | 1.93±0.07 | 1.97±0.07 | 7.60±0.13 | 0.10±0.05 | 7.70±0.14 | 21.97±0.30 | 40.27±1.03 | ||||||||||
| G3 | Control | 1.60±0.09 | 0.79 | 1.97±0.06 | 0.53 | 1.87±0.06 | 0.60 | 1.97±0.06 | 0.48 | 7.40±0.12 | 0.11 | 0.07±0.05 | 0.81 | 7.47±0.12 | 0.21 | 22.13±0.25 | 0.04 | 43.17±1.23 | 0.34 |
| Nitenpyram | 1.57±0.09 | 1.87±0.15 | 1.80±0.07 | 1.87±0.10 | 7.10±0.14 | 0.10±0.05 | 7.20±0.17 | 22.87±0.25 | 44.63±0.90 | ||||||||||
Note: Values in the table represent mean±SE. In the same generation, In the same generation, P value within the same list indicate significant differences among control and nitenpyram treated groups (at the P < 0.05 level, paired bootstrap test using TWOSEX MS chart).
Table 3.
Transgenerational effects of nitenpyram on the demographic index of G1-G3 generation A. gossypii descending from G0 adults exposed to LC20 concentrations.
Table 3.
Transgenerational effects of nitenpyram on the demographic index of G1-G3 generation A. gossypii descending from G0 adults exposed to LC20 concentrations.
| Treatments | R0 | P | rm | P | λ | P | T | P | |
|---|---|---|---|---|---|---|---|---|---|
| G1 | Control | 45.67±1.10 | <0.001 | 0.31±0.004 | <0.001 | 1.36±0.006 | <0.001 | 12.29±0.19 | 0.02 |
| Nitenpyram | 39.83±1.20 | 0.29±0.004 | 1.33±0.005 | 12.88±0.15 | |||||
| G2 | Control | 44.80±0.99 | 0.001 | 0.32±0.004 | 0.006 | 1.37±0.006 | 0.006 | 11.97±0.16 | 0.13 |
| Nitenpyram | 40.27±1.03 | 0.30±0.005 | 1.35±0.007 | 12.34±0.19 | |||||
| G3 | Control | 43.17±1.23 | 0.34 | 0.31±0.004 | 0.070 | 1.36±0.006 | 0.07 | 12.26±0.17 | 0.16 |
| Nitenpyram | 44.63±0.90 | 0.32±0.005 | 1.38±0.007 | 11.90±0.19 | |||||
Note: rm, intrinsic rate of increase (d-1); λ, finite rate of increase (d-1); R0, net reproductive rate (offspring per individual); T, mean generation time (days). Values in the table represent mean±SE. In the same generation, P value within the same list indicate significant differences among control and nitenpyram treated groups (at the P < 0.05 level, paired bootstrap test using TWOSEX MS chart).
Table 4.
Sequencing analysis of 16S rRNA of A gossypii with diversity indices.
| Samples | Raw_reads | filtered | ASV_counts | chao1 | goods_coverage | shannon | observed_species | simpson | ACE |
|---|---|---|---|---|---|---|---|---|---|
| CK-G0 | 79947 | 76925 | 229 | 228.96 | 1.00 | 1.22 | 228.60 | 0.27 | 228.06 |
| N-G0 | 79587 | 76499 | 315 | 314.75 | 1.00 | 3.86 | 314.63 | 0.72 | 316.08 |
| CK-G1 | 80480 | 77255 | 289 | 289.02 | 1.00 | 3.06 | 288.90 | 0.71 | 289.35 |
| N-G1 | 81553 | 78381 | 181 | 180.94 | 1.00 | 1.06 | 180.77 | 0.23 | 180.97 |
| CK-G2 | 79600 | 76584 | 247 | 247.34 | 1.00 | 2.37 | 247.30 | 0.64 | 247.43 |
| N-G2 | 79847 | 76721 | 220 | 220.54 | 1.00 | 1.10 | 220.23 | 0.22 | 220.24 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



