
Submitted:
29 May 2024
Posted:
30 May 2024
You are already at the latest version
Abstract
There are hundreds, if not thousands, of pathways in a unicellular prokaryotic organism, and all these pathways will have to be controlled, regulated, and coordinated to ensure their proper functioning. As organisms evolve from unicellular eukaryotic organisms to multicellular organisms such as the metazoans, more pathways have evolved to support more diverse biological functions. As a result, fine-tuned control, regulation, and coordination of these pathways becomes an immensely complex and demanding task. Yet, consistent with Darwin’s insight that “nature is prodigal in variety, though niggard in innovation,” nature has produced only a few master coordination hubs for these pathways. Moreover, multicellular organisms such as metazoans have evolved two broader means for coordinating various pathways: cross-talk and communication. Cross-talk operates via having shared components and links between intracellular pathways whereas communication operates via transmitting intercellular signals such as hormones and other ligands with membrane receptors and downstream intracellular signal transductions pathways. Our framework points to a more complex network-based approach for understanding biological pathways and the consequences of their disruption, escape, and malfunction such as cancer, aging, and other diseases.
Keywords:
Pathway
; Coordination
; Network
; Metazoans
; Crosstalk
; Communication
Introduction
About 800-600 million years ago, simple and then complex multicellular organisms emerged, leading to the Cambrian explosion of animals and the Devonian explosion of plants (Knoll 2011). Genuine multicellularity had evolved multiple times in different clades (Ruzi-Trillo & Nedelcu 2015; Niklas & Newman 2016; Ros-Rocher et al., 2021).1 As a result, control and regulatory machineries of different biological pathways in different clads are necessarily different despite sharing striking similarities. Here, we focus on (eu-)metazoans, with the hallmark of temporal-and-spatial differentiation of soma and germ.
Overwhelming evidence suggests that an elaborate network that coordinates the various pathways must exist. Despite a voluminous literature on specific pathways and coordination of several pathways, however, no integrated framework that brings together functional, molecular, and comparative mechanisms with an evolutionary perspective exists. Most existing studies focus on individual pathways or provide only limited integrative pictures, thus capturing trees or small bushes within the forest, but not the whole forest. With more and more components, pathways, and cross-talks and communications among pathways being discovered every day, (clusters of) trees grow bigger and ever more entangled with each other. As a result, the whole forest becomes so vast and dense that it is impossible to see without a map.
This contribution seeks to provide such a map, by synthesizing existing findings, propositions, and scattered evidence into an integrated framework. Taking modularity in the evolution of biological systems as the starting point (e.g., Simon 1962 [1966]; Kaufmann 1993; Hartwell et al. 1999; Bhalla 2003; McAdams et al. 2004; Saez-Rodriguez et al. 2005; Davidson & Erwin 2006; Del Vecchio et al. 2008; Andrews et al. 2024), we contend that the coordination of the various pathways constitutes a key task that organisms have evolved to cope with modularity so that organisms can survive and reproduce under stress. The resulting framework brings together key aspects of cell cycle, metabolism, differentiation, development, cancer, and aging into a more integrated framework. Most critically, the new framework calls for an integrative network approach toward understanding the control, regulation, and coordination of pathways (Green et al. 2018; Bechtel 2019; 2020; Forster et al. 2022). The framework also points to new directions for future inquiries.
Two caveats are in order. First, our focus is the evolution of control, regulation, and coordination networks in metazoans rather than the evolution of individual control and regulation machineries and pathways or the origins of metazoans. Second, although the control and regulation of the key pathways and the coordination network of them hold extensive implications for understanding aging, cancer, and lifespan of metazoans, we can only address these implications in a separate paper. Here, suffice to say that any disruption, escape, and malfunction in the network of control, regulation, and coordination brings undesired consequences that can vary from metabolism disorder, to cancer, aging (both the cellular and the organism level), and even death to an organism. Third and most obviously, there are hundred pathways even in a unicellular organism, and we cannot possibly address all of them. Rather, our discussion focuses on only the most central pathways that are indispensable for survival and reproduction and how they are coordinated.
In network and graph theory terms, every biological pathway is a network and hence can be captured in a graph, with entities being nodes while interactions and connections between entities being edges. A coordination hub therefore is a common node that integrates inputs (e.g., signals and other information) and coordinates outputs (e.g., gene expressions, functions) from at least two pathways. A major coordination hub is a machinery that integrates information and coordinates outputs from more than two major pathways in an organism. For metazoans, a master coordination hub is a hub that coordinates at least two or more major pathways that are critical for the proper function of an organism, from cell cycle control, to metabolism, homeostasis, growth, stress response, differentiation, development, programmed cell death (PCD), cancer, and reproduction. Typical examples include the well-known RB-E2F machinery and the p53-MDM2/4 machinery (see below for details).
The rest of the paper unfold in four sections. Section 2 identifies the foundational pathways from LUCA (the last universal common ancestor) to LECA (the last eukaryotic common ancestor) and then discusses how coordination of the various key pathways is achieved in LECA. Section 3 focuses on new coordination tasks and key pathways in metazoans and then addresses how coordination of the various key pathways is achieved. Section 4 highlights some key features of the coordination network in metazoans. Section 5 draws some initial implications of our framework for understanding cancer, aging, and lifespan.
2. The Foundations: From LUCA to LECA
Eukaryogenesis via endosymbiosis was a major transition that paved the way for the evolution of metazoans. By any measure, LECA was already a very complex cell (Domazet-Lošo & Tautz 2010; Niklas & Newman 2016; 2020; Ruiz-Trillo 2016; Sebé-Pedrós et al. 2017). In fact, many key control and regulatory pathways had evolved in LECA before the coming of pseudo-multicellularity in eukaryotes. These pathways all have a similar structure with three sets of components: sensor/monitor, trigger/regulator (including check points), and executor/effector, often with feedback loops (see Figure 1 for a schematic representation). Here, we focus on six key pathways that are essential for LECA to survive and reproduce.
Evidently, all these pathways need to be coordinated. Remarkably, consistent with Darwin’s insight that “nature is prodigal in variety, though niggard in innovation,” nature has produced only a few master coordination hubs for these pathways. We focus on the first three master coordinator hubs that must have evolved in LECA, the RB-E2F machinery, the PTEN machinery, and the TOR complex machinery.
Moreover, nature has evolved two broader mechanisms for coordinating these pathways, with each broader categories containing one or two sub-categories (Table 1). The first is cross-talk, which operates only intracellularly. The second category is communication, which operates both intracellularly and extracellularly.
In unicellular eukaryotic organisms, cross-talk operates via having shared components and (indirect) links via coordination hubs that integrate signals from different pathways. In contrast, regular communication operates via signals between different cellular apparatuses (e.g., chromatin) and organelles (e.g., the nucleus, the mitochondria) under normal conditions, whereas irregular communication operates via sensing extracellular stresses, damages, and genetic invasions. They are schematically summarized in Table 1.
2.1. Six Central Pathways to Be Coordinated
DNA replication & repair system. Even before LECA, key components and machineries involved in DNA replication and repair had evolved because they are indispensable for reproduction as replication (Trigos et al. 2017; 2018; 2019). Within this replication and repair system, some of the key components are the first set of the so-called “caretakers” in oncogenesis in metazoans (Domazet-Loso and Tautz 2010; Aktipis et al. 2015). These genes thus had not evolved to prevent cancer per se: cancer does not exist in unicellular organisms.
Notably, chromosome, as a coordinator of replicators, might have evolved quite early, most likely in LUCA and before the bifurcation of bacteria and Archaea (Smith and Szathmáry 1993).
In eukaryotes, circular chromosomes become chromatins with a linear structure. The coming of chromatin makes the maintaining of genome integrity and regulation of chromatin activities in different phases of cell cycle and responding to different stresses far more elaborate and fine-grained. This was largely achieved via cross-talk and communication with other major pathways (e.g., cell cycle control, autophagy) discussed below.
Heat shock response and the Ubiquitin(-like) System. Both the heat shock response (HSR) system and the ubiquitin (Ub) system are essential for cellular homeostasis and survival under stress. These two machineries must have come to exist very early on and all subsequent organisms contain these two machineries. In fact, some components in the HSR must have been some of the earliest molecules to be firmly integrated and retained by LUCA. Unsurprisingly, major heat-shock proteins (HSPs), such as Hsp100, Hsp90, Hsp70, Hsp60, and other small HSPs are highly conserved across the three domains (Akerfelt et al. 2010; Richter et al. 2010).
Like HSR, the Ub system is also indispensable for survival under stress via maintaining intracellular homeostasis (Gwong et al. 2021; Krause et al. 2022; Sheng et al. 2024). Indeed, the Ub system plays a key role in the regulation of almost all cellular processes because it is central to intracellular protein degradation (Varshavsky 2012).
Ubiquitin is universally conserved in eukaryotes while UBLs (Ub-like proteins) and URMs (Ub-related proteins) are highly conserved in Bacteria and Archaea (Hochstrasser 2009; Richter et al. 2010). Whereas bacteria and most Archaea do not possess the Ub system, at least one particular Archaea group, Caldiarchaeum subterraneum does, and it does not appear to be a result of horizontal gene transfer (Nunoura et al. 2011; Grau-Bové et al. 2014; Koonin and Yutin 2014; James et al. 2017). Thus, UBLs and UBMs might have been early inventions that were essential for the survival of LUCA, and an Ub or UBL system had already existed in some Archaea lineages and LECA, and the Ub system then had a major evolutionary burst, from Archaea to LECA (Grau-Bové et al. 2014).
SUMOs (small ubiquitin-like modifiers) are also present in all eukaryotes and regulate a host of cellular functions (Vertegaal 2022; Hendriks & Vertegaal 2024). Remarkably but unsurprisingly, the Ub system and the SUMO system not only have extensive cross-talks but share a similar configuration with the E1/E2/E3 cascade and DUBs (de-ubiquitinase) for writing and erasing the Ub/Ub-like codes (Komander & Rape 2012; Hunsnjak & Dikic 2012).
Both the Ub system and the UB-like SUMO system regulates cell cycle, gene expression (via modifying the chromatin structure, e.g., by Ub-modification of histones), autophagy, metabolism, mitochondria stress response, and other key functions via highly specific post-translational modifications of key proteins such as histones and the histone chaperon FACT (PTMs; Eifer & Vertegaal 2015; Mattiroli & Penengo 2021; Vaughan et al. 2021; Vertegaal 2022; Luo et al. 2023).
Moreover, HSR and the Ub system crosstalk with each other extensively. In fact, ubiquitinization is essential for recovery after heat shock (Gwong et al. 2021; Maxwell et al. 2021). Later on, these two systems will be co-opted and integrated with other control and regulatory pathways. For example, the Ub system and Ub-like systems are intimately connected with autophagy, which had evolved only in eukaryotes while both Ub and UBL systems play critical role in chromatin configuration, genome stability, and gene expression (Mattiroli & Penengo 2021; Vaughan et al. 2021; Vertegaal 2022; Luo et al. 2023).
Cell cycle control. Even unicellular eukaryotic organisms have a tightly controlled cell cycle. Hence, key machineries of eukaryotic cell cycle control must have already been present in LECA (Hernansaiz-Ballesteros et al. 2021). The overall picture of cell cycle control is now well understood. The cell cycle consists of four phases: G1, S1, G2, and M. Entry into each phase is governed by checkpoints that are designed to ensure that proper preparations are in place for a cell to entry the next phase. Unsurprisingly, cell cycle control is tightly coordinated with DNA replication and repair, gene expression, cell size growth, and extracellular signals.
Cell cycle control is essentially a stress-response system: a eukaryotic organism cannot survive or reproduce without a normal cell cycle that is tightly coupled with DNA replication and repair (Basu et al. 2022). Thus, while cell cycle control almost inevitably breaks down in cancer cells (Matthews et al. 2022), it did not evolve for preventing cancer.
Mitochondria as the energy and metabolism hub. Undoubtedly, a central feature of eukaryotes is the coming of mitochondria as the energy hub and hence the center for metabolism (Spinelli & Hagis 2018). Although both the exact origin of mitochondria and the rough timing of its acquisition by the Archean ancestor of LECA remain unsolved issues (Poole & Gribaldo 2014; Roger et al. 2016; Zachar & Szathmáry 2017), LECA must have possessed the mitochondria (Dacks et al. 2016). The introduction of mitochondria also brought some key ancestral components of the apoptosis pathway into LECA (Ameisen 2002; Klim et al. 2018).
With mitochondria, eukaryotes can now support far more structural complexities and functional activities than prokaryotic organisms (Lane and Martin 2010; Lane 2015; cf. Booth & Doolittle 2015). More critically, metabolism and energy production by the mitochondria must now be closely coordinated with the cell cycle (Franco & Serrano-Marin 2022).
Most critically, because there are now two sub-systems of inheritance (i.e., nucleus and mitochondria), LECA needs to control, regulate, and coordinate the two sub-systems.
Autophagy. Autopage is essentially a housekeeping machinery for homeostasis. Autophagy is central to the integrated stress response apparatus, including mitochondria stresses, heat shock, and other stresses such as starvation and oxidative stress (White et al. 2021).
A complete machinery of autophagy is fully present in unicellular eukaryotes (Kroemer & Levin 2008; Kroemer et al. 2010; Jiang et al. 2010; Rabinowitz & White 2010; Mizushima & Komatsu 2011; Shen & Codogno 2012). In fact, key components of ATG already existed in the Archean ancestors of LECA (Zhang et al. 2021). Moreover, the Ub-system plays a critical role in the autophagic process of stress responses (Maxwell et al. 2021; Gwong et al. 2021). Indeed, both ATG8 and ABT12 in autophagy are UBL proteins, thus connecting ATG with the UBL system (Zhang et al. 2021).
2.2. Coordination Hub-1 in LECA: The RB-E2F Pathway
The coordination of six major pathways noted above must have evolved along with the pathways. Most likely, coordination of the DNA replication-repair system, HSR, and the Ub system had already been achieved between LUCA and LECA. By LECA, a fairly tight coupling of DNA replication-repair, cell growth, and cell cycle control must have existed, although the exact mechanism of the coordination remains somewhat unclear (Ǿvrebǿ et al. 2023).
Both chromatin (as the apparatus hosting genetic material) and the mitochondria (as the metabolism and energy hub) must have evolved in LECA. The former is central for genetic replication and genomic stability whereas the latter is for survival. Moreover, the mitochondria first came to present as an alien object in LECA. For direct controlling of the mitochondria, LECA gradually moved many genes of the original alpha-proteobacteria to the nucleus. The host LECA cell, however, also evolved mechanisms for the mitochondria to control, regulate, and coordinate activities and functions in other parts of the cell (Monaghan & Whitmarsh 2015).
Besides direct control, a central coordination hub or machinery in eukaryotes emerged in LECA, and this is the RB-E2F system. RB is the first master coordinator in eukaryotes. Three sets of evidence suggest that the RB-E2F system is the first master coordination hub.
First, RB exists in almost all eukaryotic organisms (Takemura 2005; Cao et al. 2007; Hallman 2009), and its ancestor might have even existed in Archaea and poxvirus. Second, RB-E2F can regulate cell cycle, mitochondria, HSR, the Ub system, and autophagy, by directly and indirectly interacting and regulating with more than several hundred proteins, many of them are key components in more specific control and regulation pathways (Engeland 2022).
The exact mechanisms through which RB and RBL1/RBL2 control, regulate, and coordinate a vast array of critical activities via the central pathways noted above are now well understood (Giacinti & Giordano 2006; Sadasivam & DeCaprio 2013; Engeland 2022), although new discoveries have continued to emerge. Overall, RB-E2F exerts its effects through six distinct yet overlapping mechanisms.
First, RB controls cell cycle (including replication and division) via a very intuitive mechanism: cell size via growth. Cell growth dilutes the concentration of RB as the cell cycle inhibitor to trigger cell division in human cells (Zatulovskiy et al. 2020). Remarkably, although yeast Whi5 has no sequence homology with Rb, yeast Whi5 also controls cell division via diluting the level of Whi5 protein in cells (Huang et al. 2009; Desvoyes et al. 2014; Hasan et al. 2014; Schmoller et al. 2015). Whi5 also interacts with SBF/MBF, an E2F analogue in yeast. Whi5 and RB are thus functionally equivalent.3 This suggests that controlling cell division with a cell size sensor (either RB or Whi5) has been an ancient and foundational mechanism of regulating cell cycle in eukaryotic organisms.
Second, with a network of interactions and post-translation modifications (PTMs), RB regulates cell size, chromatin modification, and transcription via cyclin complexes, cyclin-dependent kinases (CDKs), and CDK inhibitors (CKIs). The key regulatory mechanism is via phosphorylation of different sites of the RB protein: CDKs phosphorylate RB and release E2Fs from the RB-F2F complex, and E2F is a master transcription factor that controls the expression of a host of genes critical for cell cycle progression. For instance, the expression of CDC6 (cell division cycle protein 6) gene, a key cell cycle checkpoint that is highly conserve from yeast to human, is also regulated by E2F (Yan et al. 1998). Meanwhile, p16INK4a (CDK inhibitor 2A) and p15INK4b (CDK inhibitor 2B) as key CKIs, can block CDKs from phosphorylating RB thus lock E2F within the RB-E2F complex (Kim & Sharpless 2006; Kent & Leone 2019).
Third, RB regulates mitochondria functions and protects mitochondria under stresses. Loss of RB impacts mitochondria functions extensively (Nicolay et al. 2015), suggesting that RB was an early protector of mitochondria functions.
Fourth, RB controls and coordinates survival (including metabolism) and replication, including stress responses (Dick & Rubin 2013; Desvoyes et al. 2014; Harashima & Sugimoto 2016). DNA damages can arrest cell cycle via the RB pathway. In C. elegans, Lin-53, the RB homologue, responds to starvation-induced stress (Cui et al. 2013; Lavie et al. 2018; Thomas et al. 2018; see also Zhu et al. 2020).
Fifth, E2F, as a family of transcription factors, can regulate the expression of many genes that are critical for a host of cellular pathways and functions. Thus, by regulating the activity of E2F, the RB-E2F complex can regulate, control, and coordinate numerous pathways (Kent & Leone 2019; Emanuele et al. 2020). Moreover, E2F and p53 (see below) often work together in regulating key pathways (Polager & Ginsberg 2009).
Finally, RB might have been the original germline protector before the divergence of fungi, plant, and animals. When under stress, the RB homologue (lin-35) in C. elegans regulates germ cell apoptosis by blocking ced-9, the homologue of bcl-2 in C. elegans (Schertel & Conradt 2007). Furthermore, RBBP-6, an RB-binding protein that is an E3-Ub ligase is highly conserved from unicellular organisms to plants and animals, regulates fertility by controlling cell proliferation in the germline in C. elegans (Huang et al. 2013).
Unlike metazoans, plants do not have a differentiated germline in early embryogenesis. Instead, the germline of plants is established de novo from somatic cells in reproductive organs. Yet, RBR1, the RB homologue in plants, plays similar regulatory roles in plants (Park et al. 2005; Biederman et al. 2017). Most strikingly, RBR1 also regulates germline entry in Arabidopsis (Zhao et al. 2017), and RBR1 also functions as a coordination hub of diverse pathways in plants (Desvoyes et al. 2014; Harashima and Sugimoto 2016).
Because plants have an RB pathway but not the p53/p63/p73 pathway (which is unique to animals), these findings suggest that RB had germ line protection function before the divergence of fungi, plant, and animals, and this function had only been gradually weakened after the coming of p63/p73/p53 genes in metazoans (see the discussion below). Indeed, CEP-1, the only p53 gene in C. elegans, does not regulate germline (Rutkowskiu et al. 2010).
2.3. Two Additional Coordination Hubs in LECA: PTEN and TOR
PTEN (phosphatase and tensin-homologue) and TOR (target of rapamycin) are two other master coordinators evolved in LECA. Originally discovered as a key tumor suppressor in human (Yin and Shen 2008; Hopkins et al. 2014; Chen et al. 2018; Parsons 2020; Fan et al. 2020), PTEN is a non-redundant lipid phosphatase with additional non-enzymatic functions. PTEN has been shown to play key roles in the control, regulation, and coordination of cell growth, cell size, cell (hyper-) proliferation, aging, apoptosis, development, metabolism, chromosome stability, and tumor suppression from yeast (known as TEP1 in yeast) to human. The origin of PTEN is therefore at least comparable to RB.
In planarians, PTEN is key to cellular (hyper-)proliferation in regeneration after lesion, which is a form of stress response (Oviedo et al. 2008). Thus, PTEN is also a key component of another stress-response apparatus for survival (Alvarado 2012). Furthermore, PTEN has been hypothesized to be a thrifty gene in needy time (Venniyoor 2020).
PTEN can reside in both the cytoplasm and the nuclei, and it interacts with a host of proteins to regulate diverse functions. Moreover, not only the transcription of PTEN gene is tightly regulated during transcription, PTEN protein is also finely regulated by various PTMs, including acetylation, phosphorylation, and ubiquitination (Lee et al. 2018). Nuclear PTEN protein can interact with a host of key regulators of genome stability and transcription, including the centromere-specific binding protein-C (CENP-C) and the anaphase-promoting complex (APC/C) E3 ligase. Meanwhile, cytoplasmic PTEN protein regulates AKT, a serine-threonine protein kinase, which in turn regulates the PI3K (phosphatidylinositol 3-kinase), the anchor of the PI3K pathway (Chalhoub & Baker 2009; Parsons 2020). Thus, PTEN interacts with the TOR pathway, another crucial coordination hub in metazoans.
The TOR complexes (TORC-1 and TORC-2) were discovered as the target of rapamycin, with the TOR protein being its anchor (for a historical recount, see Sabatini 2017). Whereas PTEN is a lipids phosphatase, TOR protein is a serine/threonine protein kinase. Like PTEN, the TOR gene is highly conserved in all eukaryotes. Moreover, key components of the TORC-1 and TORC-2 complexes such as TSC-1 and TSC-2 are also highly conserved (Serfontein et al. 2010; van Dam et al. 2011; Beauchamp & Platanias 2013; Tatebe & Shiozaki 2017).
TORC-1 and TORC-2 control, regulate, and coordinate a host of functions and pathways, from nutrition, metabolism, homeostasis, growth, proliferation, aging, to cancer. TORC-1 and TORC-2 possess different but some overlapping functions. The TORC-1 pathway integrates signals from growth factors and mitogens. TORC-1 also responds to amino acid and other nutrients, via the Rag-GTPase within GATOR1 and GATOR2 complexes. TORC-1 thus is key to nutrient sensing (Goul et al. 2022). Together, the TORC1 pathway integrates signals from growth factors, mitogens, and nutrients with biosynthesis and autophagy.
Meanwhile, the TORC-2 pathway is critically regulated by the AKT-PI3K pathway. The AKT-PI3K-TORC-2 pathway thus integrates not only extracellular signals ranging from stress, oxygen, energy, nutrients, and growth factors but also intracellular pathways such as autophagy, metabolism, stress-response, homeostasis, and biosynthesis. Together, the TORC-1 and TORC-2 pathways transduce a host of signals to downstream pathways via numerous upstream regulators and cross-talks with other pathways (for reviews, see Gonzalez & Rallis 2017; Liu & Sabatini 2020; Thorner 2022).
While PTEN and TORC-1/2 coordinate pathways that are different from RB and p53, there are striking similarities between how PTEN and TORC-1/2 operate and how RB and P53 operate as coordinators. Moreover, PTEN cross-talks with the TORC-1/2 pathway via the AKT-PI3K pathway, which also regulates mitochondrial functions (Oviedo et al. 2008; Sengupta et al. 2010; Millela et al. 2017). Meanwhile, autophagy and mitochondria are connected via the TORC-1 (Thomas et al. 2018), and TORC-1 also regulates Mitochondrial Integrated Stress (MIS; Khan et al. 2017). Finally, there are extensive cross-talks among RB, PTEN, TORC-1/2, and p53 (see section 3 below). Altogether, these coordination hubs have managed to coordinate all the pathways in eukaryotes since LECA (Figure 1).
Later on, these coordination networks were co-opted and integrated into a more complex control, regulation, and coordination network in more complex multicellular organisms, including metazoans.
3. New Coordination Challenges and Solutions in Metazoans
The evolution of simple and then complex multicellularity, including metazoans, has been a major transition (Sathmáry & Smith 1995). A hallmark of this transition has been the transition from temporal differentiation (i.e., a life cycle) to spatiotemporal differentiation and integration of cell types, tissues, and organs (Mikhailov et al. 2009; Sogabe et al. 2019; see also Michod et al. 2006). More complex development and differentiation in metazoans then led to the next landmark breakthrough: the spatial separation of soma and germline (Woodland 2016, 7-8). This soma-germline differentiation requires a new coordination machinery.
3.1. Sex and Soma-Germline Differentiation as Stress Response
In simple animals (e.g., planaria, sponges, Hydra), soma and germline are not so clearly differentiated: both cell types can be generated from similar or identical stem-cell like populations (Woodland 2016, 9-11). In contrast, soma and germline are clearly differentiated both temporally and spatially in complex animals. By relieving soma cells of the duty of reproduction, soma cells can differentiate into all kinds of tissues and organs that fulfill many diverse functions, and thus allow organisms to adapt to various new niches.
Protecting reproduction (i.e., the germline) under stress, however, remains a central task. In fact, sex itself might have originated as a stress response, often induced by damages (Michod 1998; 2005; 2011; Michod & Nedelcu 2003; Nedelcu et al. 2004; Nedelcu 2005; see also Lane 2015).4 This holds true even in in bacteria: fundamentally, sex in bacteria is a repairing system.
The spatial separation of soma and germline almost always involves putting the germline within the organism and preventing it from facing the environment directly: Reproduction is now more protected. The protected germline, foremost embodied in macrogametes (i.e., eggs) has been an adaption for retaining reproduction potential under stress, including limitation or depletion of food resources, genetic damages, external invasions, and other inimical conditions (Kerszberg & Wolpert 1998; Wolpert & Szathmary 2002; Woodland 2016).
Unfortunately, the spatial separation of soma and germ also entails a partial de-coupling of survival and reproduction, which makes the coordination of survival and reproduction much more demanding. More concretely, the spatial separation of the soma specialized in survival and the germline specialized in reproduction requires the reorganization of the two fitness components, namely, survival and reproduction (Michod 2003; 2005; 2011; Michod & Nedelcu 2003). In complex metazoans, therefore, a new challenge results: the control, regulation, and coordination of cellular growth/division, development, and reproduction rather than of growth/division alone as in unicellular organisms.
In other words, life has evolved from a state of little or no conflict between survival and reproduction in unicellular organisms to a state of serious conflict between survival and reproduction in simple multicellular organisms but more so in complex metazoans. This results in a critical tradeoff: even though survivability came before replicability, organisms now can, and often need to, sacrifice some soma for the germ when under stress. In short, some soma becomes disposable to reproduction (Kirkwood 1977; 2005; Michod et al. 2006; Michod and Nedelcu 2003). The spatial differentiation of germ and soma in metazoans thus requires the coordination of survival as an organism and reproduction of the organism via controlling, regulating, and coordinating the major pathways under normal and stressful conditions. This new coordination task is a fundamentally new challenge.
Besides de novo innovations (e.g., PCD, p53), metazoans cope with this new challenge by coopting existing components and pathways and adding new twinkles. For effective and efficient coordination, this is the most efficient way (Simon 1962 [1996]). Thus, some key components for cell cycle control, PCD, and germline-soma differentiation had evolved in LECAs and unicellular animals, but were later recruited into other key pathways such as PCD and soma-germline regulation in metazoans (Fairclough et al. 2013; de Mendoza et al. 2015; Ruiz-Trillo 2016; Sebé-Pedrós et al. 2017). This is most evident in the (co-)evolution of the PCD, which is indispensable to proper germ-soma differentiation.
3.2. Programmed Cell Death as Stress Response
The second major new pathway in metazoans is regulated/programmed cell death (R/PCD), which includes apoptosis, necropotosis, and pyropotosis, collectively known as PANoptosis (Tang et al. 2019; Bertheloot et al. 2021; Tang & Kang 2022).5 For metazoans, PCD is essentially a more fine-tuned and versatile homeostasis-maintaining system than autophagy. And because PCD is a homeostasis-maintaining system, PCD came to play a key role in tumor suppression later on in metazoans with long lifespan, as a byproduct.
Most likely, soma-germ differentiation and PCD had co-evolved together as stress response for multicellular organisms (Raff 1992; Kirkwood 1997; 2005; Conlin & Ratcliff 2016), because PCD is indispensable to development and differentiation of metazoans, including the normal differentiation and development of germline and protecting the germline under stress (Baum et al. 2005; Aitken et al. 2011). Fundamentally, PCD is also a stress response machinery (Hofmann 2020), much like HSR that had evolved earlier in LUCA.
Again, while PCD is a new pathway to metazoans, PCD builds upon components and pathways existing in unicellular organisms. Here, it is critical to point out that although some PCD components may be present in prokaryotes, unicellular eukaryotes (e.g., fungus), and unicellular animals (e.g., choanoflagellates) with important functions (Ameisen 2002; Koonin & Aravind 2002; Nedelcu 2009; Nedelcu et al. 2011; Klim et al. 2018; Aouacheria et al. 2013; Aktipis et al. 2015; Banjara et al. 2020), PCD becomes only indispensable in multicellular organisms (Conlin & Ratcliff 2016; Hofmann 2020). This is so because for unicellular organisms without at least a multicellular stage, PCD cannot be evolutionary meaningful (Raff 1992; Ameisen 1996; Koonin & Aravind 2002; Bayles 2014; Durrand & Ramsey 2019).
Thus, although some components of PCD pathways may be present in yeast, it is unclear whether PCD occurs in natural environment for yeast. In lab conditions, (induced) apoptosis as the only form of PCD in yeast is quite different from PCD in metazoans. Moreover, components of PCD in yeast do not have the same repertoires of control and regulation as it has in complex metazoans (Hofmann 2020).6 In contrast, apoptosis in Hydra is almost indistinguishable from apoptosis in higher animals, with both caspases and members of the Bcl-2 family participating in the process (Böttger & Alexandrova 2007, 139-140).
Apparently, the intrinsic pathway of PCD, centered upon mitochondria and cytochrome-c, came earlier than the extrinsic pathway centered upon tumor necrosis factor (TNF) receptors and death receptors: these receptors came to exist only in metazoans whereas mitochondria came to exist in LECA. After the coming of metazoans, PCD becomes more and more complex and elaborate as metazoan organisms becomes more and more complex.
3.3. The New Coordination Hub in Metazoans: p53
The evolution of new coordinating hubs must have accompanied the evolution of control and regulation pathways themselves. Metazoans did evolve a new coordination hub, the p53/p63/p73 machinery (Levine 2020). Moreover, the new hub and old hubs crosstalk with each other extensively. Altogether, these coordination hubs connect all the key pathways to ensure a close coordination for organism’s survival and reproduction under stress, partly by managing the inherent tradeoffs between survival and reproduction at the cellular and the organism level in multicellular organisms (Michod 2003; 2005; 2007; Michod et al. 2006).
The p63/p73/p53 gene exists only in animals, including unicellular choanoflagellates, with Capsapora owczarzaki containing one gene (p63) while Monosigia brevicollis contains a hybrid of p63 and p73. This suggests that p63 had already been duplicated to produce p73 in M. brevicollis (Nedelcu & Tan 2007; Levine 2020).
Quite likely, the hybrid p63/p73 gene had first evolved as a germline protector and not played any role in soma regulation in early metazoans. Early metazoans, including Placozoans and Cnidarias, all contain the p63/p73 hybrid gene that clearly functions as a key protector of the germline (Pankow& Bamberger 2007). CEP-1, the only p53 family gene in C. elegans, also functions in protecting germline by regulating apoptosis (Belyi et al. 2010; Dötsch et al. 2010; Rutkwoski et al. 2010). In mammalians, p63 still protects the female germ line (Suh et al. 2006). All these findings suggest that p63 and then p63/73 are the more ancient members of the p53 family and their function as a germline protector has been extremely conserved.
Moreover, p63/p73 does not seem to play a role in regulating soma functions. MDM2, an E3 ubiquitin ligase highly specific for p53, is a critical partner of p53 in regulating somatic functions (Siau et al. 2016). In contrast, the p63/p73 gene does not require MDM2 to function.
Notably, poriferas (sponges) lack both the p53/p63/p73 gene and the MDM2 gene.7 In sponges, choanocytes retain totipotency (i.e., they can regenerate the whole organism). As such, sponges do not have irreversible soma-germ differentiation. When this is the case, p63/p73 has no indispensable role in maintaining the soma-germ differentiation.
Meanwhile, Cnidaria (hydra and sea anemone) has p63/p73, which does not seem to interact with MDM2. C. elegans does not have MDM2 either (Åberg et al. 2017). Moreover, the single p53 family gene in Hydra, C. elegans, and Drosophila melanogaster is closer to the p63/p73 gene, and neither Hydra nor C. elegans develops cancer while Hydra does not seem to experience aging (Danko et al. 2015). D. melanogaster also lacks the MDM2 gene.
The p53 gene came to exist via another gene duplication: this marks the birth of a dedicated master protector and coordinator for the (disposable) soma in metazoans. MDM2 came to exist only in metazoans since choanoflagellates do not possess MDM2 (Åberg et al. 2017). Thus, unicellular animal organisms lack the p53/MDM2 pathway, at least a complete one. In contrast, Trichoplax possesses a single p63/p73 hybrid gene and a single MDM2 gene, and the p53/MDM2 pathway is already functional (Siau et al. 2016). Together, these sets of evidence suggest that MDM2 is essential for p53’s function of regulating soma.
When in place, the p53-MDM2 machinery came to occupy a central place in the regulation of soma, and p53-MDM2 is thus the first master controlling, regulating, and coordinating hub dedicated to the protection of the “disposable” soma (Alvarado 2012; Åberg et al. 2017; Panatta et al. 2021). The many roles of the p53-MDM2 coordinating machinery in diverse pathways of cell cycle, genome stability, apoptosis, aging, metabolism and cancer etc. have been a byproduct of its functions in the control, regulation, and coordination of survival and reproduction under stress (for reviews, see Lazano & Levine 2016; Levine 2020).
First, p53-MDM2 controls and regulates stress response in soma, including HSR and PCD. Different stress stimuli, including chronic low-dose DNA damage and nutrient starvation, can activate p53 (Mello & Attardi 2018). p53-MDM2 also promotes homeostasis by regulating autophagy in stress response (Jiang et al. 2010; Maiuri et al. 2010; White 2016; Shi et al. 2021) and metabolism (Rabinowitz & White 2010). As a result, p53-MDM2 connects key homeostasis pathways (i.e., HSR, autophagy, PCD) with metabolism and the cell cycle. Very critically, p53 proteins in two different locations performs different but interconnected roles. Whereas nuclear p53 regulates transcription of autophagy genes (ATGs), cytoplasmic p53 on the membrane of mitochondria induces apoptosis. Intuitively, all these pathways and functions can be understood as stress responses (Broz et al. 2013; Mello & Attardi 2018).
Second, p53 controls stem cell proliferation with PCD (Belyi et al. 2010). In planarian, the single p53 gene also regulates the proliferation and self-renewal of the adult stem-cell lineage (Pearson & Alejandro 2010; Siau et al. 2016). The stem-cell system is essentially a system for repair and renewal of the soma after stresses and lesions.
Altogether, the p53/MDM2 machinery is a central hub for controlling, regulating, and coordinating stress responses for the soma in complex metazoans, responding to almost the entire spectrum of stresses. Because the p53-MDM2 pathway is so central in the control, regulation, and coordination of almost all the central pathways (e.g., PCD, metabolism, autophagy) in soma, its disruption and malfunction unsurprisingly often lead to aging, cancer, and other undesired outcomes (Mello & Attardi 2018; Levine 2020).
The p53-MDM2 machinery, however, did not originate as tumor suppressors. Instead, it had first evolved as a stress-response machinery for protecting the soma in order to protect reproduction capacity even under stress. Indeed, p53’s tumor suppressing function does not have to coincide with its stress response function. After all, the single p53 gene in both C. elegans and D. melanogaster does not have any tumor suppressor function yet still retains stress response function (Alvarado 2012). Interestingly, while both C. elegans and D. melanogaster lack MDM2 (Lane & Verma 2012), the latter develops cancer while the former does not. This suggests that MDM2 is not essential for p53’s tumor-related functions.
4. Coordination in Metazoans: Cross-Talk and Communication
Adding the two new major pathways (i.e., soma-germ differentiation and PCD) in metazoans to the six major pathways already present in LECA (i.e., DNA replication and repair, cell cycle control, HSR, Ub, mitochondria, and autophagy), the coordinating task for these pathways becomes even more demanding. Unsurprisingly, metazoans have evolved a far more complex and elaborate network for coordinating these key pathways. Moreover, metazoans have evolved more complex means of cross-talk and communication, by coopting and extending machineries from its unicellular ancestors, in addition to de novo innovations. Metazoans thus retain all the means of cross-talk and communications but add many new weaponries to its arsenal, especially for communication. Most prominently, there is now regular extracellular communications: different cell types, tissues, and organs have to communicate with each other via hormones and other ligands with membrane receptors and downstream intracellular signal transductions to specific pathways. Obviously, the level of “intra-body but extra-cellular communication” is the one that distinguishes genuine multicellular organisms such as metazoans from unicellular organisms. They are schematically summarized in Table 2.
4.1. Crosstalk: Direct
Direct crosstalk is a primary means for coordination. To begin with, all the stress response pathways are coordinated by an elaborate network with built-in extensive crosstalk between the pathways. Because after a period of stress, some cells may not recover, these cells will have to be cleansed from an organism’s body, Thus, DNA repair, HSR, Ub/UBL, and autophagy, as stress response pathways already evolved in LECA, must be coordinated with PCD to form the core network for survival under stress (Eisenberg-Lerner et al. 2009; Shen et al. 2012; Ouyang et al. 2012; Yonekawa & Thorburn 2013; Fuchs & Steller 2015). Together, they also protect the germline (Bialik et al. 2010; Kroemer et al. 2010). Unsurprisingly, the coordination of these pathways is achieved by a network with shared components (e.g., caspases, HSPs, and Bcl-2) and overlapping (sub-)pathways, with new pathways building upon existing ones and drawing from existing components.
PCD, HSR, and autophagy are intimately connected in several ways to form the more complex and more sophisticated homeostasis system in metazoans (Kennedy et al. 2014; Fuchs & Steller 2015). For instance, HSPs interact and regulate all three PCD pathways, by interacting with Bcl-2 and Bax (Tang and Kang 2022). Indeed, Hsp70, as one of the most conserved proteins across the three domains, is a key regulator of apoptosis (Beere et al. 2000). Meanwhile, the chaperon-mediated autophagy pathway centers upon Hsc70, a Hsp70-homologue with 80% homology to Hsp70. Beclin-1, a Bcl-2/BclXl interacting protein, is a homologue of a yeast autophagy protein ATG6. Moreover, as a scaffolding molecule of the class III phosphoinositol-3-kinase (PtdIns3K) complex, Beclin-1 also plays critical roles in apoptosis, autophagy, cancer, and metabolism (Sinha & Levine 2009).9 Finally, DRAM1 (DNA damage regulated autophagy modulator-1), a lysosomal membrane protein that is found in C. elegans and other higher metazoans but not in simpler metazoans, is central to both PCD and autophagy (Crighton et al. 2006; 2007a; 2007b, Napoli & Flores 2013). More specifically, DRAM1 regulates PCD via regulating the level and location of BAX (Guan et a. 2015). Together, these direct cross-talks via shared components and direct interactions provide metazoans with an effective and efficient means for coordination.
Another way of direct cross-talk is via direct regulation of gene transcription or location. The most prominent example is E2F, as a master transcription factor, controls the expression of a host of genes critical for cell cycle progression. Also, while many nuclear and cytoplasmic proteins can regulate mitochondria functions, many proteins in mitochondria can also go to nuclear and regulate nuclear gene expressions (Monaghan & Whitemarsh 2015).
4.2. Crosstalk: Indirect
Indirect cross-talks primarily operate via the four coordination hubs, RB, PTEN, TOR, and p53, which in turn coordinate many diverse pathways (Figure 1). To save space, we focus on the extensive indirect cross-talks between the RB-E2F hub and the p53-MDM2/4 hub
The RB-E2F and the p53-MDM2 machinery form a larger network with extensive built-in crosstalk: this is the most efficient way for coordinating the various pathways already controlled, regulated, and coordinated by the two machineries (Simon 1962 [1996]). Through the two machineries, a metazoan can then control, regulate, and coordinate almost all the key pathways for survival and reproduction, from metabolism, homeostasis, cell cycle, growth, development, and reproduction. Most prominently, Key components in the p53-MDM2 machinery are downstream targets of the Rb-E2F machinery: RB-E2F regulates the p53-MDM2 pathway via regulating MDM2, which in turn can target p53 for Ub-mediated degradation (Watson & Irvin 2006; Brooks & Gu 2011). Also, P19ARF in mice (p14ARF in human), a tumor suppressor gene only found in mammalians, not only regulates the p53-MDM2/MDM4 pathway but is itself a downstream target of the RB-E2F pathway (Kim & Sharpless 2006).
The significant overlapping between the RB-E2F pathway and the DREAM pathway that centers on RB-like (RBL) proteins (RBL-1/p107 & RBL-2/p130) suggests that the latter plays overlapping but non-redundant functions. The RBL-centric DREAM pathway regulates genes that are different from genes regulated by the RB-E2F pathway (Sadasivam & DeCaprio 2013; Engeland 2022; Hoareau et al. 2023).
Both RB and p53 regulate PCD and autophagy (Attardi and Sage 2013; Kroemer et al. 2010; Jiang et al. 2010; Maiuri et al. 2010; Broz et al. 2013; Napoli & Flores 2013). RB regulates autophagy via regulating and interacting with CDKI (CDK inhibitors) and E2F1 (Jiang et al. 2010). Meanwhile, p53 regulates both the Ub pathway and PCD via transcription (Watson & Irwin 2006; Brooks& Gu 2011; Lomonosova & Chinnadurai 2009).
P53 can be modified by both Ub and UBL proteins. Also, both DUPs (de-ubiquitination enzymes) and USP7/HAUSP (Herpesvirus-associated ubiquitin-specific protease) regulate the p53-MDM2 pathway via their de-ubiquitinase activity (Bhattacharya & Ghosh 2014). Meanwhile, PUMA, a potent proapoptotic protein regulated by p53, play a critical role in both p53-dependent and p53-independent apoptosis (Yu & Zhang 2009). Indeed, PUMA promotes apoptosis by releasing cytoplasmic p53 from the anti-apoptotic BCL-xL (Chipuk et al. 2005). Of course, the mitochondria are a key hub for apoptosis, with the BNIP3 proteins being key sensors for mitochondria stress (Chinnadurai et al. 2009). These communications connect pathways of UB, HSR, PCD, autophagy, metabolism, and proliferation into a highly integrated machinery, with mitochondria as a central node (Lavie et al. 2018; Thomas et al. 2018).
Because of p53’s centrality in all these pathways for controlling and regulating soma (Levine 2020), p53 came to play critical roles in regulating cell division, aging, and cancer in metazoans with long life span, as unintended consequences. Other components that interact with the p53/MDM2 pathway can also have tumor regulating functions. Similarly, the PCD pathways and its interactions with RB-E2F and p53 via Bcl-2/Bax also play important roles in aging and tumor suppression.
p53 has a dual role in regulating metabolism, autophagy, and PCD (White 2016), thus also in cancer and aging as unintended consequences (Sherr & McCromick 2002; Maiuri et al. 2010; Ouyang et al. 2012; Napoli & Flores 2013; Guo & White 2016; White et al. 2021). Also, IAPs (inhibitors of apoptosis) all contain a ubiquitin E3-ligase domain, which is essential to the activity of IAPs (Berthelet & Dubrez 2013). Finally, caspase inhibition leads to cell death by autophagy, which involves reactive oxygen species (ROS) accumulation, membrane lipid oxidation, and loss of plasma membrane integrity. Altogether, via PCDs, autophagy is connected with aging, cancer-suppression, inflammation, immunity, and many other processes as byproducts (Hayat 2014; Zhong et al. 2016).
RB can regulate PCD and autophagy via p53 but also independently from p53 (Attardi & Sage 2013; Kroemer et al. 2010; Maiuri et al. 2010). Because autophagy and PCD are connected, p53 and autophagy are also connected with chromatin and mitochondria, often via RB. Thus, PCD and autophagy are also intimately linked via p53, RB, and mitochondria. RB also indirectly interacts with HSR via Ub/UBL/UBM. Finally, E2F1 and p53 both regulate the expression of genes of proapoptotic proteins (Lomonosova & Chinnadurai 2009). Together, the crosstalk and communication of autophagy and PCD protects the survival of organisms via protecting soma cells, which only become “disposable” after reproduction.
4.3. Communications
In addition to intracellular communications already present in unicellular eukaryotes, complex metazoans also deploy many extracellular communications between different cell types, tissues, organs, and systems. Perhaps the most studied communication has been the one between the neural system and the immune system (Dantza 2018; Schiller et al. 2021; special collections of Nature Immunology and Nature Neuroscience, 2017).
Similar communications must also lie between other systems (e.g., between the digestive system and the immune system; see Fung et al. 2017; Veiga-Fernades & Pachnis 2017). Internal organs of vertebrates must also communicate with each other quite extensively during development and under normal and stressful conditions.
Unfortunately, we know little about these possible communications. The molecular foundation for such communication, however, is partly known. Five key signaling pathways, FGF, Hedgehog, Notch, TGF-beta, and Wnt, underpin much of the long-distance communications during development in eumetazoans (Adamska 2015). Also, some receptors are nuclear ones that can function directly as transcription factors (e.g., retinoic acid receptors; Berenguer & Duester 2022). In contrast, other receptors can only regulate gene expression via a longer signal transduction pathway that goes through tyrosine kinases (Hubbard & Miller 2007). Brain nuclear receptors seem to regulate many cardiovascular functions (Wang et al. 2023). Meanwhile, the effect of glucocorticoids on inflammation might operate via metabolism rewiring (Auger et al. 2024). Still, much remains unknown.
4.4. Why Are the Mitochondria So Central to the Coordination Network in Metazoans?
The mitochondria have emerged as a central node in the network that coordinates the various pathways in metazoans (Chandel 2015; Mottis et al. 2019). So why are mitochondria so central? We believe that the centrality of mitochondria is partially underpinned by its central role in regulating metabolism and homeostasis pathways (i.e., HSR, autophagy, and PCD) as the key hub of survival machinery under stress.
To begin with, mitochondria play a central role in coordinating replication and survival under stress since LECA. Mitochondria are central to cellular metabolism, including metabolism under stress (Harputlugil et al. 2014; Matilainen et al. 2017). For instance, stress such as oxidative stress and starvation can include mitochondria unfolded proteins response (UPR), and mitochondria is central to UPR as a form of stress response (Haynes & Ron 2010). More strikingly, mitochondrial stress induces chromatin remodeling via UPR (Y. Tian et al. 2016; 2020). In addition, the Ub pathway also regulates metabolism by regulating stress response-induced mitochondria protein degradation (Lavie et al. 2018; Thomas et al. 2018).
In C. elegans, low levels of mitochondrial stress caused by exposure to RNAi or xenobiotics can restore HSF-1 function, thereby maintaining cytosolic proteostasis, enhancing stress resistance, and prolonging lifespan, all without detrimental effects on development or reproduction (Labbadia & Morimoto 2015; Labbadia et al. 2017).
Meanwhile, heat shock induces HSR, and mitochondrial SSBP1 (single-strained DNA binding protein-1, which is a homologue of E. coli SSBP) protects cells from proteotoxic stresses by translocating SSBP1 to the nuclear and potentiating stress induced HSF1 (heat-shock factor-1) transcriptional activity (K. Tan et al. 2015). Moreover, HSF1 and hence SSBP1 and ISW-1 (a chromatin modeling factor) regulate ssDNA binding and transcription (Matilainen et al. 2017), thus coordinating extensive communications between chromatin configuration, transcription, and HSR. Similarly, RAD9, a DNA damage repair protein that functions as a cell cycle check point, contains a BH3 motif that allows it to interact with BCL-2 and BCL-xL to promote apoptosis after DNA damage (Komatsu et al., 2000). RAD9 thus plays important roles in tumorigenesis (Lieberman et al. 2011). Moreover, RAD9 and p53 work together to regulate DNA damage response (DDR) by selectively regulating expressions of downstream genes within the DDR pathway (Lieberman et al. 2017). In short, there is extensive crosstalk between mitochondria and the nucleus, especially under stress (Mottis et al. 2019; Vizioli et al. 2020; Monaghan & Whitmarsh 2015).
More critically, both the RB-E2F machinery and the p53-MDM2 machinery operate via their interaction with the mitochondria (Attardi & Sage 2013). RB was an early protector of mitochondria function because mitochondria is key to the survival of LECA: loss of RB protein impacts mitochondria function extensively (Nicolay et al. 2015). RB can also activate Bax directly on the membrane of mitochondria (Attardi & Sage 2013; Hilgendorf et al. 2013). p53 can also interact with Bcl-2, Bcl-XL, and Bax on the membrane of mitochondria. In fact, p53 can open the mitochondrial permeability transition pore to trigger necrosis (Vaseva et al. 2012). In addition, both E2F1 and p53 regulates the expression of key genes in the PCD pathways (Lomonosova & Chinnadurai 2009). Finally, mitochondria are a central coordinator hub of the various PCD pathways (Bock & Tait 2020). In sum, because RB and p53 are key coordinating hubs for all the major pathways and their coordination relies on mitochondria, mitochondria is central to the coordination network in metazoans.
5. Discussion
We have outlined a new framework for understanding the coordination network of the numerous pathways in metazoans, a topic that has received inadequate attention. We highlight five key points.
First, an organism’s structure and function are underpinned by numerous biological pathways. For survival and reproduction, these pathways must be finely controlled and regulated. Control and regulation, however, are insufficient: these pathways must also be coordinated.
Second and quite remarkably, evolution has produced only a few master coordination hubs Moreover, evolution has produced essentially two broader means for coordination: (direct and indirect) cross-talk and (intra-cellular and extra-cellular) communication.
Third, RB, p53, PTEN, TOR are master coordinating hubs, and they also have extensive cross-talks and communications among themselves. This fact alone explains why they have been so critical in regulating so many key biological functions, from cell cycle control to homeostasis, metabolism, development, PCD, reproduction, cancer, and aging.
Fourth, because these coordination hubs have evolved in different time, their roles have been stratified or stacked upon even though they also have extension cross-talk and communication among themselves: this is the most efficient and effective way for building a coordination network (Simon 1962 [1996]).
Finally, consistent with the gist of evolutionary theory, much of the innovation was driven by evolutionary pressure to survive and reproduce under stress (Nedelcu & Michod 2004; Michod 2007; Wagner et al. 2019). As a result, all the pathways and their coordination hubs bear the imprints of coping with stresses, ranging from genetic to nutritional.
Self-evidently, our new framework holds extensive implications for understanding a host of issues, including cancer, aging, and lifespan in metazoans. Unfortunately, addressing these issues in any detail is beyond the scope of this paper. Here, we merely highlight a few key points for organizing evidence and drawing further implications. We start with principal implications and then proceed to a brief discussion of aging, cancer, and lifespan.
5.1. Principal Implications
Control and regulation of the key pathways are essential for the survival and reproduction of a metazoan. Both the inability to perform required roles due to defects in and escaping from the pathways produce harmful outcomes. Control and regulation of the key pathways, however, is not enough: coordination of them via crosstalk and communication is also absolutely necessary for the survival and reproduction of a metazoan. Unsurprisingly, disruption or loss of the proper coordination within this network of coordination is also harmful for a metazoan.
Remarkably, coordination in metazoans has centered upon only a few master coordination hubs: RB-E2F, p53-MDM2, PTEN, and TOR. As more complex animals evolve, more components and hence additional pathways for control, regulation, and coordination that support more complex and intricate processes of development, differentiation, and morphogenesis have also evolved. Lacking a better term, I call these additional pathways and hubs as “secondary”. Inevitably, all these secondary pathways or hubs are also coordinated by the four master coordination hubs, directly or indirectly. Hence, there is a hierarchical structure that governs these different coordination hubs and pathways (Simon 1962 [1996]).
For example, both forkhead (FOX) proteins and polycomb proteins can be understood as critical but secondary regulators downstream of RB and p53. In Hydra, p53-MDM2 already interacts with FOX proteins to control and regulate many diverse pathways (Belyi et al. 2010; Lam et al. 2013; Rutkowski et al. 2010). Polycomb proteins, as epigenetic regulators that are responsible for the evolution of complexity in both plants (including green algae) and metazoans, repress p14/p19ARF and hence impacts the RB-E2F machinery (Sowpati et al. 2015). Pathways centered upon other key signal and regulatory proteins, including JAK3, hedgehog (HH), Hippo, homoetic box (Hox), insulin, insulin-like growth factors (IGFs), transforming growth factor beta (TGF-beta), c-myc, Notch, and Wnt, are additional examples of such secondary pathways (Adamska 2015; Loh et al. 2016; Mortzfeld et al. 2019; Biglou et al. 2021). Together, these developmental signaling pathways regulate key pathways in development.
Similarly, c-myc, a transcription factor that plays critical roles in cell cycle control and apoptosis, is regulated by the SUMO system (Sun et al. 2018). Since the SUMO system also cross-talks with the Ub-system and many other key pathways, some of the key functions of c-myc might have been exerted via regulation by the SUMO system. Thus, when the regulation of c-myc by the SUMO system breaks down, cancer and other diseases become far more likely (Eifer & Vertegaal 2015; Vertegaal 2022; Claessens & Vertegaal 2024). Likewise, CYLD (Cylindromatosis Tumor Suppressor Protein), a mammalian de-uniquitinase, regulates p53-dependent genotoxic stress response (Fernández-Majada et al. 2016). CYLD is also an essential negative regulator of the NF-kB pathway and mitogen-activated protein kinase pathways.
Our framework thus calls for a more integrated network approach for understanding the control, regulation, and coordination of the various pathways in all organisms, and in metazoans in particular. In fact, with more and more discoveries that point to extensive cross-talk and communication among the different pathways, only a more complex network-based approach can lead us to a more integrated picture of these pathways. While scientists have made much progress in this direction (e.g., Forster et al. 2022; see also Bechtel 2019; Green et al. 2018 for interesting philosophical discussions), much more remains to be done.
5.2. Implications for Understanding Aging, Cancer, and Lifespan
Within so many pathways, there will never be perfect control and regulation. Moreover, with such a complex network of pathways, coordination will never be perfect either. With wear-and-tear during an organism’s life course and imperfect repairment, disruption and malfunction in and escape from these pathways become inevitable, and so does breakdown of coordination of these pathways. Unsurprisingly, these disruptions, malfunctions, escapes, and breakdowns will be associated with defects and escapes from the pathways, which in turn result in undesired outcomes, although the specific manifestations of defects and escapes may differ significantly in different tissues in different organisms. Among these undesired outcomes, aging and cancer are the two most prominent. Our network framework implies some key lessons for understanding aging and cancer.
Unicellular organism only needs to take care of the cell within the environment: the cell and the organism are equivalent. Moreover, without the possibility of cancer, unicellular organisms only need to take care of cellular senescence as aging at the organism level.
In contrast, cells and the organisms are no longer equivalent in metazoans as multicellular organisms. Thus, while cellular senescence still underpins aging at the organism level in metazoans, they are no longer equivalent. Indeed, cellular senescence and PCD are critical to mammalian development, partly because cellular senescence can trigger PCD (Wanner et al. 2021). When senescence does not trigger PCD due to malfunctions or disruptions, senescent cells accumulate, leading to either bodily aging or cancer, or both (Van Deursen 2014). Meanwhile, uncontrolled multiplication by some cells (as cancer) poses serious threat to the survival of an organism. As a result, the loss of control, regulation, and coordination at the cellular level threat eventually threatens proper function at the organism level and potentially endangers survival and reproduction.
In metazoans, therefore, there is a conflict between survival (soma) and reproduction (germ). As such, mediation of this conflict is also necessary, as Michod and his collaborators have long argued (e.g., Michod 1997; 2007; Michod & Nedelcu 2003; Nedelcu & Michod 2003; Michod & Herron 2006). When this is the case, some (disposable) soma cells can be sacrificed for the sake of survival and then reproduction under stress, via stress responses such as HSR, autophagy, and PCD. Indeed, during development and differentiation, PCD is necessary for limiting soma’s selfishness for the whole organisms to survive and the germ to reproduce (Aktipis et al. 2015).
As a result, a multi-level selection (MLS) perspective is essential for understanding the evolution of multicellularity (e.g., Maynard Smith & Szathmáry 1995; Michod 1997; 2007; Okasha 2006; Szathmáry 2015). From the MLS perspectives, the disposable soma theory essentially captures the underlying logic of the tradeoff between survival and reproduction (Kirkwood 1977; 2005; Kirkwood & Austadi 2000; Gaddy et al. 2021). The notion that aging and cancer are the results of antagonistic pleitrophy essentially captures the same insight.
Multicellularity is the foundation of aging and cancer at the organism level (Trigos et al. 2018; Nedelcu 2020). Hence, aging and cancer are byproducts of multicellularity. Most critically, the control and regulation of aging and cancer came very late in the evolution of metazoans. Cancer is the outcome of breakdown of and escape from control and regulation, which in turn leads to the breakdown of cooperation by individual cells within a multicellular organism. Cancer, however, faces little selection pressure until reproduction requires maturity and grooming (e.g., in mammalians). Indeed, tumors in plants are rarely life threatening. As a result, there is little selection pressure against tumor in plants (Doonan & Sablowski 2010).
For metazoans with a relatively short lifespan, selection pressure for controlling aging will be minimal because these organisms have completed their reproduction before their maximum lifespan or even before the middle point of their maximum lifespan (Campisi 2003). For example, more than 80% of cancer in humans are found in people over the age of 55, by which most reproduction tasks have been completed. An organism’s long lifespan does not contribute to its fitness unless successful reproduction requires maturity (e.g., in mammals) and grooming (including social grooming). Cancer follows the same logic. Fundamentally, cancer is the inevitable outcome of wear-and-tear: with damages constantly accumulating, some organisms simply cannot check and repair all the damages.
Thus, although defects in and escapes from the key pathways can lead to aging and cancer, aging, cancer, and the connection between them, are evolutionary byproducts (Finkel et al. 2007; cf. Campisi 2003). In other words, although disruption of functions within the key pathways are often associated with aging and cancer, most pathways have not evolved for regulating or suppressing aging or cancer. This is also why aging at cell, organ, and organism levels are not tightly connected. Likewise, there is no neat connection between PCD and cancer (Labi & Erlacher 2015). In short, aging, cancer, and other malfunctions are byproducts of the loss of control, regulation, and coordination: they cannot have genuine evolutionary explanations. And because aging and cancer are fundamentally underpinned by the loss of control, regulation, and coordination of the key machineries and pathways outlined above, prevention and curing of caner has to focus on the terminals of these pathways.
Finally, in light of the network of control, regulation, and coordination, the differentiation of caretakers (e.g., RecQ in DNA repair) and gatekeepers or type I and type II tumor suppressor (e.g., CDC6) is a useful heuristic, but only to a degree (cf. Kinzler& Vogelstein 1997; Campisi 2003). While many caretakers that are essential for survival and reproduction had evolved before eukaryotes whereas gatekeepers such as the RB-E2F machinery evolved only in eukaryotes for regulating chromatin-based gene expression and hence coupling with cell cycle in LECA and then the p53-MDM2 machinery in metazoans (Domazet-Lošo & Tautz 2010), their functions are intertwined extensively. A multilevel selection (MLS) perspective is also essential for understanding aging and cancer (Lean & Plutynski 2016; see also Okasha 2006; Goldfrey-Smith 2009; Michod 2005; 2007; Weissman 2015).
References
- Åberg, E.; Saccoccia, F.; Grabherr, M.; Ore, W.Y.J.; Jemth, P.; Hultqvist, G. Evolution of the p53-MDM2 pathway. BMC Evolutionary Biology 2017, 17, 177. [Google Scholar] [CrossRef] [PubMed]
- Adamska, M. (2015). Developmental signaling and the emergence of animal multicellularity. In I. Ruiz-Trillo, & A. M. Nedelcu. (Eds.), Evolutionary transition to multicellular life: Principles and mechanisms (pp. 425–450). Springer.
- Aitken, R.J.; Findlay, J.K.; Hutt, K.J.; Kerr, J.B. Apoptosis in the germ line. Reproduction 2011, 141, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Åkerfelt, M.; Morimoto, R.I.; Lea Sistonen. Heat shock factors: integrators of cell stress, development, and lifespan. Nature Reviews, Molecular Cell Biology 2010, 11, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Aktipis, C.A.; Boddy, A.M.; Jansen, G.; Hibner, U.; Hochberg, M.E.; Maley, C.C.; et al. Cancer across the tree of life: Cooperation and cheating in multicellularity. Philosophical Transactions of the Royal Society B: Biological Sciences 2015, 370, 20140219. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, A.S. Cellular hyperproliferation and cancer as evolutionary variables. Current Biology 2012, 22, R772–R778. [Google Scholar]
- Ameisen, J.C. The origin of programmed cell death. Science 1996, 272, 1278–1279. [Google Scholar] [CrossRef] [PubMed]
- Ameisen, J.C. On the origin, evolution, and nature of programmed cell death: A timeline of four billion years. Cell Death Differ 2002, 9, 367–393. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S.S.; Wiley, H.S.; Sauro, H.M. Design patterns of biological cells. BioEssays 2024, 46, 2300188. [Google Scholar] [CrossRef] [PubMed]
- Aouacheria, A.; de Laval, W.R.; Combet, C.; Hardwick, J.M. Evolution of Bcl-2 homology motifs: homology versus homoplasy. Trends in Cell Biology 2013, 23, 103–111. [Google Scholar] [CrossRef]
- Attardi, L.D.; Sage, J. RB goes mitochondrial. Genes & Development 2013, 27, 975–979. [Google Scholar]
- 12. Auger, J-P.; et al. 2024. Metabolic rewiring promotes anti-inflammatory effects of glucocorticoids. Nature. [CrossRef]
- Banjara, S.; Suraweera, C.D.; Hinds, M.G.; Kvansakul, M. The Bcl-2 Family: Ancient Origins, Conserved Structures, and Divergent Mechanisms. Biomolecules 2020, 10, 128. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Greenwood, J.; Jones, A.W.; et al. Core control principles of the eukaryotic cell cycle. Nature 2022, 607, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Baum, J.S.; George, J.P. S.; Mccall, K. Programmed cell death in the germline. Seminars in Cell & Developmental Biology 2005, 16, 245–259. [Google Scholar]
- Bayles, K.W. Bacterial programmed cell death: Making sense of a paradox. Nature Reviews Microbiology 2014, 12, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, E.M.; Platanias, L.C. The evolution of the TOR pathway and its role in cancer. Oncogene 2013, 32, 3923–3932. [Google Scholar] [CrossRef] [PubMed]
- Bechtel, W. Analysing network models to make discoveries about biological mechanisms. The British Journal for the Philosophy of Science 2019, 70, 459–484. [Google Scholar] [CrossRef]
- Bechtel, W. Hierarchy and levels: analysing networks to study mechanisms in molecular biology. Philosophical Transactions of the Royal Society, B 2020, 375, 20190320. [Google Scholar] [CrossRef]
- Beere, H.M.; Wolf, B.B.; Cain, K.; Mossert, D.D.; Mahboubi, A.; Kuwana, T.; Tailor, P.; Morimoto, R.I.; Cohen, G.M.; Green, D.R. Heat-shock protein 70 inhibits apoptosis by preventing recruitment of procaspase-9 to the Apaf-1 apoptosome. Nature Cell Biology 2010, 2, 469–475. [Google Scholar] [CrossRef]
- Belyi, V.A.; Ak, P.; Markert, E.; Wang, H.; Hu, W.; Puzio-Kuter, A.; et al. The origins and evolution of the p53 family of genes. Cold Spring Harbor Perspectives in Biology 2010, 2, 968–976. [Google Scholar] [CrossRef]
- 22. Berenguer, Marie, & Duester, G. Retinoic Acid, RARs and early development. Journal of Molecular Endocrinology, 2022; 69, T59–T67. [CrossRef]
- Berthelet, J.; Dubrez, L. Regulation of apoptosis by inhibitors of apoptosis (IAPs). Cells 2013, 2, 163–187. [Google Scholar] [CrossRef]
- Bertheloot, D.; Latz, E.; Franklin, B.S. Necroptosis, pyroptosis and apoptosis: An intricate game of cell death. Cellular & Molecular Immunology 2021, 18, 1106–1121. [Google Scholar]
- Bhalla, U.S. Understanding complex signaling networks through models and metaphors. Progress in Biophysics and Molecular Biology 2003, 81, 45–65. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Ghosh, M.K. HAUSP, a novel deubiquitinase for Rb-MDM2 the critical regulator. FEBS Journal 2014, 281, 3061–3078. [Google Scholar] [CrossRef]
- Bialik, S.; Zalckvar, E.; Ber, Y.; Rubinstein, A.D.; Kimchi, A. Systems biology analysis of programmed cell death. Trends in Biochemical Sciences 2010, 35, 556–564. [Google Scholar] [CrossRef]
- Biederman, S.; Harashima, H.; Chen, P.; Heese, M.; Bouyer, D.; Sofroni, K.; Schnittger, A. The retinoblastoma homolog RBR1 mediates localization of the repair protein RAD51 to DNA lesions in Arabidopsis. EMBO Journal 2017, 36, 1279–1297. [Google Scholar] [CrossRef]
- Biglou, S.G.; Bendena, W.G.; Chin-Sang, I. An overview of the insulin signaling pathway in model organisms drosophila melanogaster and caenorhabditis elegans. Peptides 2021, 145, 170640. [Google Scholar] [CrossRef]
- Bock, F.J.; Tait, S.W. G. Mitochondria as multifaceted regulators of cell death. Nature Review Molecular Cell Biology 2020, 21, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Booth, A.; Doolittle, W.F. Eukaryogenesis, how special really? Proceedings of the National Academy of Sciences, USA, 2015; 112, 10278–10285. [Google Scholar]
- Böttger, A.; Alexandrova, O. Programmed cell death in Hydra. Seminars in Cancer Biology 2007, 17, 134–146. [Google Scholar] [CrossRef] [PubMed]
- Brooks, C.; Gu, W. P53 regulation by ubiquitin. Febs Letters 2011, 585, 2803–2809. [Google Scholar] [CrossRef]
- Broz, D.K.; Mello, S.S.; Bieging, K.T.; Jiang, D.; Dusek, R.L.; Brady, C.A.; et al. Global genomic profiling reveals an extensive p53-regulated autophagy program contributing to key p53 responses. Genes & Development 2013, 27, 1016–1031. [Google Scholar]
- Brunet, T.; Larson, B.T.; Linden, T.A.; Vermeij, M.J. A.; Mcdonald, K.; King, N. Light-regulated collective contractility in a multicellular choanoflagellate. Science 2019, 366, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Campisi, J. Cancer and ageing: rival demons? Nature Reviews, Cancer 2003, 3, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Peng, B.; Yao, L.; Zhang, X.; Sun, K.; Yang, X.; Yu, L. The ancient function of RB-E2F Pathway: Insights from its evolutionary history. Biology Direct 2010, 5, 55. [Google Scholar] [CrossRef] [PubMed]
- Chalhoub, N.; Baker, S.J. PTEN and the PI3-Kinase Pathway in Cancer. Annual. Review of Pathology: Mechanisms of Disease 2009, 4, 127–50. [Google Scholar] [CrossRef] [PubMed]
- Chandel, N.S. Evolution of mitochondria as signaling organelles. Cell Metabolism 2015, 22, 204–206. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, J.; He, L. & Stiles, B.L. PTEN: Tumor suppressor and metabolic regulator. Frontiers in Endocrinology 2018, 9, 338. [Google Scholar] [PubMed]
- Chinnadurai, G.; Vijayalingam, S.; Gibson, S.B. BNIP3 subfamily BH3-only proteins: mitochondrial stress sensors in normal and pathological functions. Oncogene 2008, 27, S114–S127. [Google Scholar] [CrossRef] [PubMed]
- Chipuk JE, Bouchier-Hayes L, Kuwana T, Newmeyer DD, Green DR. PUMA couples the nuclear and cytoplasmic proapoptotic function of p53. Science 2005, 309, 1732–1735. [Google Scholar] [CrossRef]
- Claessens, L.A.; Vertegaal, A.C. O. ; (2024). SUMO proteases: from cellular functions to disease. Trends in Cell Biology, 20. [CrossRef]
- Conlin, P.L.; Ratcliff, W.C. (2016). Trade-offs drive the evolution of increased complexity in nascent multicellular digital organisms. In K. J. Niklas, & S. A. Newman (Eds.), Multicellularity: Origins and evolution (pp. 131–149). MIT Press.
- Crighton, D.; O' Prey, J.; Bell, H.S.; Ryan, K.M. P73 regulates DRAM-independent autophagy that does not contribute to programmed cell death. Cell Death & Differentiation 2007, 14, 1071–1079. [Google Scholar]
- Crighton, D.; Wilkinson, S.; Ryan, K.M. DRAM links autophagy to p53 and programmed cell death. Autophagy 2007, 3, 72–74. [Google Scholar] [CrossRef]
- Crighton, D.; Wilkinson, S.; O' Prey, J.; Syed, N.; Smith, P.; Harrison, P.R.; et al. DRAM, a p53-induced modulator of autophagy, is critical for apoptosis. Cell 2006, 126, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Cohen, M.L.; Teng, C.; Han, M. The tumor suppressor Rb critically regulates starvation-induced stress response in C. elegans. Current Biology 2013, 23, 975–980. [Google Scholar] [CrossRef] [PubMed]
- Dacks, J.B.; Mark C., F.; Buick, R.; Eme, L.; Gribaldo, S.; Roger, A.J.; Brochier-Armanet, C.; Devos, D.M. The changing view of eukaryogenesis – fossils, cells, lineages and how they all come together. Journal of Cell Science 2016, 2016, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dam, T.J. P. V.; Zwartkruis, F.J. T.; Bos, J.L.; Snel, B. Evolution of the TOR pathway. Journal of Molecular Evolution 2011, 73, 209–220. [Google Scholar] [PubMed]
- Danko, M.; Kozlowski, J.; Schaible, R. Unraveling the non-senescence phenomenon in Hydra. Journal of Theoretical Biology 2015, 382, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Dantzer, R. Neuroimmune Interaction: from the Brain to the Immune System and vice versa. Physiology Review 2018, 98, 477–504. [Google Scholar] [CrossRef] [PubMed]
- Davidson, E.H.; Erwin, D.H. Gene Regulatory Networks and the Evolution of Animal Body Plans. Science 2006, 311, 796–800. [Google Scholar] [CrossRef] [PubMed]
- de Mendoza, A.; Suga, H.; Permanyer, J.; Irimia, M.; Ruiz-Trillo, I. Complex transcriptional regulation and independent evolution of fungal-like traits in a relative of animals. eLife, 2015, 4, e08904. [Google Scholar] [CrossRef] [PubMed]
- Del Vecchio, D.; Ninfa, A.J.; Sontag, E.D. Modular cell biology: Retroactivity and insulation. Molecular Systems Biology 2008, 4, 161. [Google Scholar] [CrossRef]
- Desvoyes, B.; Fernández-Marcos, M.; Sequeira-Mendes, J.; Otero, S.; Vergara, Z.; Gutierrez, C. Looking at plant cell cycle from the chromatin window. Frontiers in Plant Science 2014, 5, 369. [Google Scholar] [CrossRef]
- Dick, F.A.; Rubin, S.M. Molecular mechanisms underlying RB protein function. Nature Reviews Molecular Cell Biology 2013, 14, 297–306. [Google Scholar] [CrossRef]
- Domazet-Lošo, T.; Tautz, D. Phylostratigraphic tracking of cancer genes suggests a link to the emergence of multicellularity in metazoan. BMC Biology 2010, 8, 66. [Google Scholar] [CrossRef]
- Doonan, J.H.; Sablowski, R. 2010. Walls around tumors — why plants do not develop cancer. Nature Reviews Cancer, 10, 794–802.
- Dötsch, V.; Bernassola, F.; Coutandin, D.; Candi, E.; Melino, G. P63 and p73, the ancestors of p53. Cold Spring Harbor Perspectives in Biology 2010, 2, a004887. [Google Scholar] [CrossRef] [PubMed]
- Durrand, P.M.; Ramsey, G. The nature of programmed cell death. Biological Theory 2019, 14, 30–41. [Google Scholar] [CrossRef]
- Eifer, K. & Vertegaal, A.C.O. SUMOylation-Mediated Regulation of Cell Cycle Progression and Cancer. Trends in Biochemical Sciences 2015, 40, 779–793. [Google Scholar]
- Eisenberg-Lerner, A.; Bialik, S.; Simon, H-U. ; Kimchi, A. Life and death partners: Apoptosis, autophagy and the cross-talk between them. Cell Death & Differentiation 2009, 16, 966–975. [Google Scholar]
- Emanuele, M.J.; Enrico, T.P.; Mouery, R.D.; Wasserman, D.; Nachum, S.; Tzur, A. (2020) Complex Cartography: Regulation of E2F transcription factors by Cyclin F and Ubiquitin. Trends in Cell Biology, 30 (8), 640–652. [CrossRef]
- Engeland, K. Cell cycle regulation: P53-p21-RB signaling. Cell Death & Differentiation 2022, 29, 946–960. [Google Scholar]
- Fairclough, S.R.; Chen, Z.; Kramer, E.; Zeng, Q.; Young, S.; Robertson, H.M.; et al. Premetazoan genome evolution and the regulation of cell differentiation in the choanoflagellate Salpingoeca rosetta. Genome Biology 2013, 14, R15. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Kraynak, J.; Knisely, J.P. S.; Formenti, S.C.; Shen, W.H. PTEN as a guardian of the genome: Pathways and targets. Cold Spring Harbor Perspectives in Medicine 2020, 10, a036194. [Google Scholar] [CrossRef]
- Fernández-Majada, V.; Welz, P.S.; Ermolaeva, M.A.; Schell, M.; Adam, A.; Dietlein, F.; et al. The tumor suppressor CYLD regulates the p53 DNA damage response. Nature Communications 2016, 7, 12508. [Google Scholar] [CrossRef]
- Finkel, T.; Serrano, M.; Blasco, M.A. The common biology of cancer and ageing. Nature 2007, 448, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Forster, D.T.; Li, S.C.; Yashiroda, Y.; Yoshimura, M.; Li, Z.; Isuhuaylas, L.A. V.; et al. BIONIC: Biological network integration using convolutions. Nature Methods 2022, 19, 1250–1261. [Google Scholar] [CrossRef] [PubMed]
- Franco, R.; Serrano-Marin, J. The unbroken Krebs cycle. Hormonal-like regulation and mitochondrial signaling to control mitophagy and prevent cell death. BioEssays 2022, 45, 2200194. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, Y.; Steller, H. Live to die another way: Modes of programmed cell death and the signals emanating from dying cells. Nature Review Molecular Cell Biology 2015, 16, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.C.; Olson, C.A.; Hsiao, E.Y. Interactions between the microbiota, immune and nervous systems in health and disease, Nature Neuroscience 2017, 20, 145–155. 20. [CrossRef]
- Gaddy, M.A.; Kuang, S.; Alfhili, M.A.; Lee, M.H. The soma-germline communication: Implications for somatic and reproductive aging. BMB Reports, 2021, 54, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Giacinti, C.; Giordano, A. RB and cell cycle progression. Oncogene 2006, 25, 5220–5227. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, L.; Kroemer, G. Common and divergent functions of Beclin 1 and Beclin 2. Cell Research 2013, 23, 1341–1342. [Google Scholar] [CrossRef] [PubMed]
- Godfrey-Smith, P. (2009). Darwinian Populations and Natural Selection. Cambridge University Press.
- Gonzalez, S.; Rallis, C. The TOR signaling pathway in spatial and temporal control of cell size and growth. Frontiers in Cell and Developmental Biology 2017, 5, 61. [Google Scholar] [CrossRef] [PubMed]
- Goul, C.; Peruzzo, R.; Zoncu, R. The molecular basis of nutrient sensing and signaling by mTORC1 in metabolism regulation and disease. Nature Reviews Molecular Cell Biology 2023, 24, 857–875. [Google Scholar] [CrossRef]
- Grau-Bové, X.; Sebé-Pedrós, A.; Ruiz-Trillo, I. The Eukaryotic ancestor had a complex ubiquitin signaling system of archaeal origin. Molecular Biology and Evolution 2014, 32, 726–739. [Google Scholar] [CrossRef]
- Green, S.; Şerban, M.; Scholl, R.; Jones, N.; Brigandt, I.; Bechtel, W. Network analyses in systems biology: New strategies for dealing with biological complexity. Synthese 2018, 195, 1751–1777. [Google Scholar] [CrossRef]
- Guan, J.J.; Zhang, X.D.; Sun, W.; Qi, L. ; Wu, J-C.; and Qin, Z-H. (2015). DRAM1 regulates apoptosis through increasing protein levels and lysosomal localization of BAX. Cell Death & Disease, 2015; 6, e1624. [Google Scholar] [CrossRef]
- Guo, J.Y.; White, E. Autophagy, metabolism, and cancer. Cold Spring Harbor Symposia on Quantitative Biology 2016, 81, 73–78. [Google Scholar] [CrossRef]
- Gwon, Y.; Maxwell, B.A.; Kolaitis, R.; Zhang, P.; Kim, H.J. & Taylor, J.P. (2021) Ubiquitination of G3BP1 mediates stress granule disassembly in a context-specific manner. Science, 2021; 372, eabf6548. [Google Scholar] [CrossRef]
- Hallman, A. Retinoblastoma-related proteins in lower eukaryotes. Communicative & Integrative Biology 2009, 2, 538–544. [Google Scholar]
- Harashima, H.; Sugimoto, K. Integration of developmental and environmental signals into cell proliferation and differentiation through retinoblastoma-related 1. Current Opinion in Plant Biology 2016, 29, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Harputlugil, E.; Hine, C.; Vargas, D.; Robertson, L.; Manning, B.D.; Mitchell, J.R. The TSC complex is required for the benefits of dietary protein restriction on stress resistance in vivo. Cell Reports 2014, 8, 1160–1170. [Google Scholar] [CrossRef] [PubMed]
- Hartwell, L.H.; Hopfield, J.J.; Leibler, S.; Murray, A.W. From molecular to modular cell biology. Nature 1999, 402 (suppl 6761), 47–52. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.M.; Brocca, S.; Sacco, E.; Spinelli, M.; Papaleo, E.; Lambrughi, M.; et al. A comparative study of Whi5 and retinoblastoma proteins: From sequence and structure analysis to intracellular networks. Frontiers in Physiology 2014, 4, 315. [Google Scholar] [CrossRef] [PubMed]
- Hayat, M.A. Chapter 1- Introduction to autophagy: Cancer, other pathologies, inflammation, immunity, infection and aging, vol. 1–4. Autophagy: Cancer, Other Pathologies, Inflammation, Immunity, Infection, and Aging 2014, 35, 1–36. [Google Scholar]
- Hayes, C.S.; Low, D.A. Signals of growth regulation in bacteria. Current Opinion in Microbiology 2009, 12, 667–673. [Google Scholar] [CrossRef]
- Haynes, C.M.; Ron, D. The mitochondrial UPR-protecting organelle protein homeostasis. Journal of Cell Science 2010, 123, 3849–3855. [Google Scholar] [CrossRef]
- He, C.; et al. Beclin 2 functions in autophagy, degradation of G protein-coupled receptors, and metabolism. Cell 2013, 154, 1085–1099. [Google Scholar] [CrossRef]
- Heaton, L.L. M.; Jones, N.S.; Fricker, M.D. A mechanistic explanation of the transition to simple multicellularity in fungi. Nature Communications 2020, 11, 2594. [Google Scholar] [CrossRef] [PubMed]
- Hendriks, I.A. & Vertegaal, A. C. O. A comprehensive compilation of SUMO proteomics Nature Reviews, Cell and Molecular Biology 2022, 23, 715–731. [Google Scholar]
- Hernansaiz-Ballesteros, R.D.; Földi, C.; Cardelli, L.; et al. (2021) Evolution of opposing regulatory interactions underlies the emergence of eukaryotic cell cycle checkpoints. Scientic Report, 11, 11122. [CrossRef]
- Herron, M.D. What are the major transitions. Biology & Philosophy 2021, 36, 2. [Google Scholar]
- Herron, M.D.; Borin, J.M.; Boswell, J.C.; Walker, J.; Chen, I-C. K.; Knox, C.A.; et al. (2019). De novo origins of multicellularity in response to predation. Scientific Reports 9, 2328. [PubMed]
- Hilgendorf, K.I.; Leshchiner, E.S.; Nedelcu, S.; Maynard, M.A.; Calo, E.; Ianari, A.; Walensky, L.D. & Lees, J.A. The retinoblastoma protein induces apoptosis directly at the mitochondria. Genes & Development 2013, 27, 1003–1015. [Google Scholar]
- Hoareau, M.; Rincheval-Arnold, A. Gaumer, & S. Guénal, I. DREAM a little dREAM of DRM: Model organisms and conservation of DREAM-like complexes. Bioessays 2023, 46, 2300125. [Google Scholar]
- Hochstrasser, M. Origin and function of ubiquitin-like proteins. Nature 2009, 458, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Hoffmeyer, T.T.; Burkhardt, P. Choanoflagellate models- Monosiga brevicollis and Salpingoeca rosetta. Current Opinion in Genetics & Development 2016, 39, 42–47. [Google Scholar]
- Hofmann, K. The evolutionary origins of programmed cell death Signaling. Cold Spring Harbor Perspectives in Biology 2020, 12, a036442. [Google Scholar] [CrossRef]
- Hopkins, B.D.; Hodakoski, C.; Barrows, D.; Mense, S.M.; Parsons, R.E. PTEN function, the long and the short of it. Trends in Biochemical Sciences 2014, 39, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Kaluarachchi, S.; van Dyk, D.; Friesen, H.; Sopko, R.; Ye, W.; et al. Dual regulation by pairs of cyclin-dependent protein kinases and histone deacetylases controls G1 transcription in budding yeast. PLoS Biology 2009, 7, e1000188. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Ma, X.; Zhao, Y.; Miao, L. The C. elegans homolog of RBBP6 (RBPL-1) regulates fertility through controlling cell proliferation in the germline and nutrient synthesis in the Intestine. PLoS ONE 2013, 8, e58736. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, S.R.; Miller, W.T. Receptor tyrosine kinases: mechanisms of activation and signaling. Current Opinion in Cell Biology 2007, 19, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Husnjak, K.; Dikic, I. Ubiquitin-Binding Proteins: Decoders of Ubiquitin-Mediated Cellular Functions. Annual Review of Biochemistry 2012, 81, 291–322. [Google Scholar] [CrossRef] [PubMed]
- James, R.H.; Caceres, E.F.; Escasinas, A.; Alhasan, H.; Howard, J.A.; Deery, M.J.; Ettema, T.I. J.; Robinson, N.P. (2017) Functional reconstruction of a eukaryotic-like E1/E2/(RING) E3 ubiquitylation cascade from an uncultured archaeon. Nature Communications, 8, 1120. [CrossRef]
- Jékely, G. Evolution: How not to become an animal. Current Biology 2019, 29, R1240–R1242. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Martin, V.; Gomez-Manzano, C.; Johnson, D.G.; Alonso, M.; White, E.; et al. The RB-E2F1 pathway regulates autophagy. Cancer Research 2010, 70, 7882–7893. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, D. (2015). In I. Ruiz-Trillo, & A. M. Nedelcu. (Eds.), Evolutionary transition to multicellular life: Principles and mechanisms (pp. 451-468). Springer.
- Kauffman, S.A. (1993). The origins of order: Self-organization and selection in evolution. Oxford University Press.
- Kawabe, Y.; Schilde, C. ; Chen, Z-H.; Du, Q.; Schaap. P.; et al. (2015). The Evolution of Developmental Signaling in Dictyostelia from an Amoebozoan Stress Response. In I. Ruiz-Trillo, & A. M. Nedelcu. (Eds.), Evolutionary transition to multicellular life: Principles and mechanisms (pp. 451-468). Springer.
- Kennedy, D.; Jäger, R.; Mosser, D.D.; Afshin Samali. Regulation of apoptosis by heat shock proteins. IUMBM Life 2014, 66, 327–38. [Google Scholar] [CrossRef] [PubMed]
- Kent, L.N.; Leone, G. The broken cycle: E2F dysfunction in cancer. Nature Review Cancer 2019, 19, 326–336. [Google Scholar] [CrossRef]
- Kerszberg, M.; Wolpert, L. The Origin of Metazoa and the Egg: a Role for Cell Death. Journal of Theoretical Biology 1998, 193, 535–537. [Google Scholar] [CrossRef]
- Khan, N.A.; Nikkanen, J.; Yatsuga, S.; Jackson, C.; Wang, L.; Pradhan, S.; et al. MTORC1 regulates mitochondrial integrated stress response and mitochondrial myopathy progression. Cell Metabolism 2017, 26, 419–428. [Google Scholar] [CrossRef]
- Kim, W.Y.; Sharpless, N.E. The regulation of INK4/ARF in cancer and aging. Cell 2006, 127, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Kinzler, K.W.; Vogelstein, B. Cancer-susceptibility genes. Gatekeepers and caretakers. Nature 1997, 386, 761–763. [Google Scholar] [CrossRef] [PubMed]
- Kirkwood, T.B. L. The evolution of aging. Nature 1977, 270, 301–304. [Google Scholar] [CrossRef] [PubMed]
- Kirkwood, T.B. L. The odd science of aging. Cell 2005, 120, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Kirkwood, T.B. L.; Austad, S.N. Why do we age? Nature 2000, 408, 233–238. [Google Scholar] [CrossRef] [PubMed]
- Klim, J.; Gladki, A.; Kucharczyk, R.; Zielenkiewicz, U.; Kaczanowski, S. Ancestral state reconstruction of the apoptosis machinery in the common ancestor of eukaryotes. G3 (Bethesda) 2018, 8, 2121–2134. [Google Scholar] [CrossRef] [PubMed]
- Knoll, A.H. The multiple origins of complex multicellularity. Annual Review of Earth & Planetary Sciences 2011, 39, 217–239. [Google Scholar]
- Komander, D.; Rape, M. The Ubiquitin Code. Annual Review of Biochemistry 2012, 81, 203–229. [Google Scholar] [CrossRef]
- Komatsu, K.; Miyashita, T.; Hang, H.; Hopkins, K.M.; Zheng, W.; Cuddeback, S.; et al. Human homologue of S. pombe Rad9 interacts with BCL-2/BCL-xL and promotes apoptosis. Nature Cell Biology 2000, 2, 1–6. [Google Scholar] [CrossRef]
- König, S.G.; Nedelcu, A.M. (2016). The mechanistic basis for the evolution of soma during the transition to multicellularity in the volvocine algae. In K. J. Niklas, & S. A. Newman (Eds.), Multicellularity: Origins and evolution (pp. 43–71). The MIT Press.
- König, S.G.; Nedelcu, A.M. The genetic basis for the evolution of soma: Mechanistic evidence for the co-option of a stress-induced gene into a developmental master regulator. Proceedings of the Royal Society. Biological sciences 2020, 287, 20201414. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Aravind, L. Origin and evolution of eukaryotic apoptosis: The bacterial connection. Cell Death and Differentiation 2002, 9, 394–404. [Google Scholar] [CrossRef] [PubMed]
- Koonin, E.V.; Yutin, N. The dispersed archaeal eukaryome and the complex archaeal ancestor of eukaryotes. Cold Spring Harbor Perspectives in Biology 2014, 6, a016188. [Google Scholar] [CrossRef] [PubMed]
- Krause, L.J.; Herrera, M.G.; Winkhofer, K.F. The Role of Ubiquitin in Regulating Stress Granule Dynamics. Frontiers in Physiology 2022, 13, 910759. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G.; Levine, B. Autophagic cell death: The story of a misnomer. Nature Reviews Molecular Cell Biology 2008, 9, 1004–1010. [Google Scholar] [CrossRef] [PubMed]
- Kroemer, G.; Marino, G.; Levine, B. Autophagy and the integrated stress response. Molecular Cell 2010, 40, 280–293. [Google Scholar] [CrossRef] [PubMed]
- Labbadia, J.; Morimoto, R.I. Repression of the Heat Shock Response Is a Programmed Event at the Onset of Reproduction. Molecular Cell 2015, 59, 639–50. [Google Scholar] [CrossRef] [PubMed]
- Labbadia, J.; Brielmann, R.M.; Neto, M.F.; Lin, Y.F.; Haynes, C.M.; Morimoto, R.I. Mitochondrial stress restores the heat shock response and prevents proteostasis collapse during aging. Cell Reports 2017, 21, 1481–1494. [Google Scholar] [CrossRef] [PubMed]
- Labi, V.; Erlacher, M. How cell death shapes cancer. Cell Death & Disease 2015, 6, e1675. [Google Scholar]
- Lam, E.W.; Brosens, J.J.; Gomes, A.R.; Koo, C. Forkhead box proteins: Tuning forks for transcriptional harmony. Nature Reviews Cancer 2013, 13, 482–495. [Google Scholar] [CrossRef]
- Lane, D.P.; Verma, C. Mdm2 in evolution. Genes& Cancer 2012, 3, 320–324. [Google Scholar] [CrossRef]
- Lane, N. (2015) Lane, N. (2015). The vital question: Energy, evolution, and the origins of complex life. W. W. Norton & Co.
- Lane, N.; Martin, W. The energetics of genome complexity. Nature 2010, 467, 929–934. [Google Scholar] [CrossRef] [PubMed]
- Lavie, J.; De Belvalet, H.; Sonon, S.; Ion, A.M.; Dumon, E.; Melser, S.; et al. Ubiquitin-dependent degradation of mitochondrial proteins regulates energy metabolism. Cell Reports 2018, 23, 2852–2863. [Google Scholar] [CrossRef] [PubMed]
- Lean, C.; Plutynski. A. The evolution of failure: explaining cancer as an evolutionary process. Biology and Philosophy 2016, 31, 39–57. [Google Scholar] [CrossRef]
- Lee, Y.R.; Chen, M, & Pandolfi, P. P. The functions and regulation of the PTEN tumor suppressor: New modes and prospects. Nature Reviews Molecular Cell Biology 2018, 19, 547–562. [Google Scholar] [CrossRef] [PubMed]
- Levin, T. C, & King, N. Evidence for sex and recombination in the choanoflagellate salpingoeca rosetta. Current Biology 2013, 23, 2176–2180. [Google Scholar] [PubMed]
- Levin, T.C.; Greaney, A.J.; Wetzel, L.; King, N. The rosetteless gene controls development in the choanoflagellate S. rosetta. eLife 2014, 3, e04070. [Google Scholar] [CrossRef] [PubMed]
- Levine, A.J. ; p53: 800 million years of evolution and 40 years of discovery. Nature Reviews Cancer 2020, 20, 471–480. [Google Scholar] [CrossRef]
- Lieberman, H.B.; et al. The role of RAD9 in tumorigenesis. Journal of Molecular Cell Biology 2011, 3, 39–43. [Google Scholar] [CrossRef]
- Lieberman, H.B.; et al. p53 and RAD9, the DNA Damage Response, and Regulation of Transcription Networks. Radiation Research 2017, 187, 424–432. [Google Scholar] [CrossRef]
- Liu, G.Y.; Sabatini, D.M. MTOR at the nexus of nutrition, growth, ageing and disease. ” Nature Reviews Molecular Cell Biology 2020, 21, 183–203. [Google Scholar] [CrossRef] [PubMed]
- Loh, K.M.; van Amerongen, R.; Nusse, R. Generating cellular diversity and spatial form: Wnt signaling and the evolution of multicellular animals. Developmental Cell 2016, 38, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Lomonosova, E.; and Chinnadurai, G. BH3-only proteins in apoptosis and beyond: an overview. Oncogene 2009, 27, S2–S19. [Google Scholar] [CrossRef] [PubMed]
- Lozano, G.; Levin, A.J. (2016) The p53 protein: from cell regulation to cancer. Cold Spring Harbor Perspectives in Medicine.
- Luo, A.; et al. H2B ubiquitination recruits FACT to maintain a stable altered nucleosome state for transcriptional activation. Nature Communications 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Maiuri, M.C.; Galluzzi, L.; Morselli, E.; Kepp, O.; Malik, S.A.; Kroemer, G. Autophagy regulation by p53. Current Opinion in Cell Biology 2010, 22, 181–185. [Google Scholar] [CrossRef]
- Matilainen, O.; Quirós, P.M.; Auwerx, J. Mitochondria and epigenetics– Crosstalk in homeostasis and stress. Trends in Cell Biology 2017, 27, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Matilainen, O.; Sleiman, M.S. B.; Quiros, P.M.; Garcia, S.M. D. A.; Auwerx, J. The chromatin remodeling factor ISW-1 integrates organismal responses against nuclear and mitochondrial stress. Nature Communications 2017, 8, 1818. [Google Scholar] [CrossRef]
- Matthews, H.K.; Bertoli, C. & de Bruin, R.A.M. Cell cycle control in cancer. Nat Rev Mol Cell Biol 2022, 23, 74–88. [Google Scholar] [CrossRef]
- Mattiroli, F.; Penengo, L. Histone Ubiquitination: An Integrative Signaling Platform in Genome Stability. Trends in Genetic 2021, 37, 566–5812. [Google Scholar] [CrossRef]
- Maxwell, B.A.; Gwon, G.; Mishra, A.; Peng, J.; Nakamura, H.; Zhang,K. ; Kim, H.J.; Taylor, J.P. Ubiquitination is essential for recovery of cellular activities following heat shock. Science 2021, 372, eabc3593. [Google Scholar] [CrossRef]
- McAdams HH, Srinivasan B, Arkin AP. The evolution of genetic regulatory systems in bacteria. Nature Review Genetics 2004, 5, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Mello, S.S.; Attardi, L.D. Deciphering p53 signaling in tumor suppression. Current Opinion in Cell Biology 2018, 51, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Michod, R.E. Cooperation and conflict in the evolution of individuality. I. Multilevel selection of the organism. The American Naturalist 1997, 149, 607–645. [Google Scholar] [CrossRef]
- Michod, R.E. (2003). Cooperation and conflict mediation during the origin of multicellularity. In P. Hammerstein (Ed.), Genetic and cultural evolution of cooperation (pp. 291–307). MIT Press.
- Michod, R.E. On the transfer of fitness from the cell to the multicellular organism. Biology & Philosophy 2005, 20, 967–987. [Google Scholar]
- Michod, R.E. Evolution of individuality during the transition from unicellular to multicellular life. Proceedings of the National Academy of Sciences, USA 2007, 104, 8613–8618. [Google Scholar] [CrossRef] [PubMed]
- Michod, R.E. (2011). Evolutionary transition in individuality: Multicellularity and sex. In B. Calcott, & K. Sterelny (Eds.), The major transitions in evolution revisited (pp. 169–198). MIT Press.
- Michod, R.E.; Herron, M.D. Cooperation and conflict during evolutionary transitions in individuality. Journal of Evolutionary Biology 2006, 19, 1406–1409. [Google Scholar] [CrossRef] [PubMed]
- Michod, R.E.; Nedelcu, A.M. On the reorganization of fitness during evolutionary transitions in individuality. Integrative & Comparative Biology 2003, 43, 64–73. [Google Scholar]
- Michod, R.E. ; (1998). Origin of sex for error repair: III. selfish sex. Theoretical Population Biology, 53, 60–74.
- Michod, R.E.; Viossat, Y.; Solari, C.A.; Hurand, M.; Nedelcu, A.M. Life-history evolution and the origin of multicellularity. Journal of Theoretical Biology 2006, 239, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Mikhailov, K.V.; Konstantinova, A.V.; Nikitin, M.A.; Troshin, P.V.; Rusin, L.Y.; Lyubetsky, V.A. The origin of Metazoa: A transition from temporal to spatial cell differentiation. Bioessays 2009, 31, 758–768. [Google Scholar] [CrossRef]
- Milella, M.; Falcone, I.; Conciatori, F.; Matteoni, S.; Sacconi, A.; Luca, T.D.; et al. PTEN status is a crucial determinant of the functional outcome of combined MEK and mTOR inhibition in cancer. Scientific Reports 2017, 7, 43013. [Google Scholar] [CrossRef]
- Milo, R.; Itzkovitz, S.; Kashtan, N.; Levitt, R.; Shen-Orr, S.; Ayzenshtat, I.; et al. Superfamilies of evolved and designed networks. Science 2004, 303, 1538–1542. [Google Scholar] [CrossRef] [PubMed]
- Milo, R.; Shen-Orr, S.; Itzkovitz, S.; Kashtan, N.; Chklovskii, D.; Alon, U. Network motifs: Simple building blocks of complex networks. Science 2002, 298, 824–827. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of cells and tissues. Cell 2011, 147, 728–741. [Google Scholar] [CrossRef]
- Monaghan, R.M.; Whitmarsh, A.J. Mitochondrial Proteins Moonlighting in the Nucleus. Trends in Biochemical Sciences 2022, 40, 728–735. [Google Scholar] [CrossRef] [PubMed]
- Mortzfeld, B.M.; Taubenheim, J.; Klimovich, A.V.; Fraune1, S.; Rosenstiel, P.; Bosch, T.C. G. Temperature and insulin signaling regulate body size in Hydra by the Wnt and TGF-beta pathways. Nature Communications 2019, 10, 3257. [Google Scholar] [CrossRef]
- Mottis, A. Sébastien Herzig, & Auwerx, J. Mitocellular communication: Shaping health and disease. Science 2019, 366, 827–832. [Google Scholar]
- Nanjundiah, V. (2016). Cellular slime mold development as a paradigm for the transition from unicellular to multicellular life. In K. J. Niklas, & S. A. Newman (Eds.), Multicellularity: Origins and evolution (pp 105–130). MIT Press.
- Napoli, M.; Flores, E.R. The family that eats together stays together: New p53 family transcriptional targets in autophagy. Genes & Development 2013, 27, 971–974. [Google Scholar]
- Nedelcu, A.M. Sex as a response to oxidative stress: Stress genes co-opted for sex. Proceedings of the Royal Society of London. Series B: Biological Sciences 2005, 272, 1935–1940. [Google Scholar] [CrossRef]
- Nedelcu, A.M. Comparative genomics of phylogenetically diverse unicellular eukaryotes provide new insights into the genetic basis for the evolution of the programmed cell death machinery. Journal of Molecular Evolution 2009, 68, 256–268. [Google Scholar] [CrossRef]
- Nedelcu, A.M. The evolution of multicellularity and cancer: Views and paradigms. Biochemical Society Transactions 2020, 48, 1505–1518. [Google Scholar] [CrossRef]
- Nedelcu, A.M.; Michod, R.E. Sex as a response to oxidative stress: The effect of antioxidants on sexual induction in a facultatively sexual lineage. Proceedings of the Royal Society of London. Series B: Biological Sciences 2003, 270, S136–S139. [Google Scholar] [CrossRef] [PubMed]
- Nedelcu, A.M.; Michod, R.E. The evolutionary origin of an altruistic gene. Molecular Biology and Evolution 2006, 23, 1460–1464. [Google Scholar] [CrossRef] [PubMed]
- Nedelcu, A.M.; Michod, R.E. (2011). Molecular mechanisms of life history trade-offs and the evolution of multicellular complexity in Volvo calean green algae. In T. Flatt, & A. Heyland (Eds.), Mechanisms of life history evolution (pp. 271–283). Oxford University Press.
- Nedelcu, A.M.; Tan, C. Early diversification and complex evolutionary history of the p53 tumor suppressor gene family. Development Genes and Evolution 2007, 217, 801–806. [Google Scholar] [CrossRef] [PubMed]
- Nedelcu, A.M.; Driscoll, W.W.; Durand, P.M.; Herron, M.D.; Rashidi, A. On the paradigm of altruistic suicide in the unicellular world. Evolution 2011, 65, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Nedelcu, A.M.; Marcu, O.; Michod, R.E. Sex as a response to oxidative stress: A twofold increase in cellular reactive oxygen species activates sex genes. Proceedings of the Royal Society of London. Series B: Biological Sciences 2004, 271, 1591–1596. [Google Scholar] [CrossRef] [PubMed]
- Nicolay, B.N.; Danielian, P.S.; Kottakis, F.; Lapek, J.D.; Sanidas, I.; Miles, W.O.; et al. Proteomic analysis of pRb loss highlights a signature of decreased mitochondrial oxidative phosphorylation. Genes & Development 2015, 29, 1875–1889. [Google Scholar]
- Niklas, K.J.; Newman, S.A. The origins of multicellular organisms. Evolution & Development 2013, 15, 41–52. [Google Scholar]
- Niklas, K.J.; Newman, S.A. (2016). Multicellularity: Origins and evolution, The MIT Press.
- Niklas, K.J.; Newman, S.A. The many roads to and from multicellularity. Journal of Experimental Botany 2020, 71, 3247–3253. [Google Scholar] [CrossRef] [PubMed]
- Nunoura, T.; Takaki, Y.; Kakuta, J.; Nishi, S.; Takami, H. Insights into the evolution of archaea and eukaryotic protein modifier systems revealed by the genome of a novel archaeal group. Nucleic Acids Research 2011, 39, 3204–3223. [Google Scholar] [CrossRef]
- Okasha, S. (2006). Evolution and the levels of selection.
- Ouyang, L.; Shi, Z.; Zhao, S.; Wang, F-T. ; Zhou, T-T.; Liu, B.; et al. Programmed cell death pathways in cancer: A review of apoptosis, autophagy and programmed necrosis. Cell Proliferation 2012, 45, 487–498. [Google Scholar] [CrossRef]
- Oviedo, N.J.; Pearson, B.J.; Levin, M.; Alvarado, A.S. Planarian PTEN homologs regulate stem cells and regeneration through TOR signaling. Disease Models and Mechanisms 2008, 1, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Ǿvrebǿ, J.I.; Ma, Y.; Edgar, B.A. Cell growth and the cell cycle: new insights about persistent questions. BioEssays, 2022, 44, 2200150. [Google Scholar] [CrossRef] [PubMed]
- Panatta, E.; Zampieri, C.; Melino, G.; Amelio, I. Undertanding p53 tumour surressor network. Biology Direct 2021, 16, 14. [Google Scholar] [CrossRef] [PubMed]
- Pankow, S.; Bamberger, C. The p53 tumor suppressor-like protein nvp63 mediates selective germ cell death in the sea anemone Nematostella vectensis. PLoS ONE 2007, 2, e782. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Ahn, J-W. ; Kim, Y-K.; Kim, S.J.; Kim, J.K.; Kim, W.T.; Pai, H-S. Retinoblastoma protein regulates cell proliferation, differentiation, and endoreduplication in plants. The Plant Journal 2005, 42, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Parsons, R. Discovery of the PTEN tumor suppressor and its connection to the PI3K and AKT oncogenes. Cold Spring Harbor Perspectives in Medicine 2020, 10, a036129. [Google Scholar] [CrossRef] [PubMed]
- Pearson, B.J.; Alejandro, A.S. A planarian p53 homolog regulates proliferation and self-renewal in adult stem cell lineages. Development 2010, 137, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Polager, S.; Ginsberg, D. P53 and E2F: partners in life and death. Nature Reviews Cancer 2009, 9, 738–748. [Google Scholar] [CrossRef]
- Poole, A.M.; Gribaldo, S. Eukaryotic origins: how and when was the mitochondrion acquired? Cold Spring Harbor Perspectives in Biology 2014, 6, a015990. [Google Scholar] [CrossRef]
- Quiles, J.M.; et al. Deciphering functional roles and interplay betweenBeclin1and Beclin2in autophagosome formation and mitophagy. Science Signaling 2023, 16. [Google Scholar] [CrossRef]
- Rabinowitz, J.D.; White, E. Autophagy and metabolism. Science 2010, 330, 1344–1348. [Google Scholar] [CrossRef] [PubMed]
- Raff, M.C. Social controls on cell survival and cell death. Nature 1992, 356, 397–400. [Google Scholar] [CrossRef] [PubMed]
- Richter, K.; Haslbeck M, Buchner J. The heat shock response: life on the verge of death. Molecular Cell 2010, 40, 253–266. [Google Scholar] [CrossRef]
- Roger, A.J.; Munoz-Gomez, S.A.; Kamikawa, R. The origin and diversification of mitochondria. Current Biology 2016, 27, R1177–R1192. [Google Scholar] [CrossRef]
- Ros-Rocher, N.; Pérez-Posada, A.; Leger, M.M.; Ruiz-Trillo, I. The origin of animals: An ancestral reconstruction of the unicellular-to-multicellular transition. Open Biology 2021, 11, 200359. [Google Scholar] [CrossRef]
- Ruiz-Trillo, I. (2016). What are the genomes of premetazoan lineages telling us about the origin of Metazoa? In K. J. Niklas, & S. A. Newman (Eds.), Multicellularity: Origins and evolution (pp. 171–184). The MIT Press. Springer.
- Ruiz-Trillo, I.; Nedelcu, A.M. (2015). Evolutionary transitions to multicellular life: Principles and mechanisms.
- Rutkowski, R.; Hoffman, K.; Gartner, A. Phylogeny and Function of the Invertebrate p53 Superfamily. Cold Spring Harbor Perspectives in Biology 2010, 2, a001131. [Google Scholar] [CrossRef] [PubMed]
- Sabatini, D.S. Twenty-five years of mTOR: Uncovering the link from nutrients to growth. Proceedings of the National Academy of Sciences, USA 2017, 114, 11818–11825. [Google Scholar] [CrossRef]
- Sadasivam, S.; DeCaprio, J.A. The DREAM complex: Master coordinator of cell cycle-dependent gene expression. Nature Reviews Cancer 2013, 13, 585–595. [Google Scholar] [CrossRef]
- Saez-Rodriguez, J.; Kremling, A.; Gilles, E.D. Dissecting the puzzle of life: modularization of signal transduction networks. Computers & Chemical Engineering 2005, 29, 619–629. [Google Scholar] [CrossRef]
- Saggere, R.M. S.; Lee, C.W. J.; Chan, I.C. W.; Durnford, D.G.; Nedelcu, A.M. A life-history trade-off gene with antagonistic pleiotropic effects on reproduction and survival in limiting environments. Proceedings of the Royal Society B, Biological Sciences 2022, 289, 20212669. [Google Scholar] [CrossRef]
- Schaap, P. Evolution of developmental signaling in Dictyostelid social amoebas. Current Opinion in Genetics & Development 2016, 39, 29–34. [Google Scholar]
- Schertel, C.; Conradt, B.C. elegans orthologs of components of the RB tumor suppressor complex have distinct pro-apoptotic functions. Development 2007, 134, 3691–3701. [Google Scholar] [CrossRef]
- Schiller, M.; Ben-Shannan T., L.; Rolls, A. Neuronal regulation of immunity: why, how and where? Nature Reviews Immunology 2021, 21, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Schmoller, K.M.; Turner J., J.; Kõivomägi, M.; Skotheim, J.M. Dilution of the cell cycle inhibitor Whi5 controls budding-yeast cell size. Nature 2015, 526, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Schultz, D.T.; Haddock, S.H. D.; Bredeson, J.V.; Green, R.E.; Simakov, O.; Rokhsar, D.S. Ancient gene linkages support ctenophores as sister to other animals. Nature 2023, 618, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Sebé-Pedrós, A.; Degnan, B.M.; Ruiz-Trillo, I. The origin of Metazoa: A unicellular perspective. Nature Reviews Genetics volume 2017, 18, 498–512. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, S.; Peterson, T.R.; Sabatini, D.M. Regulation of the mTOR complex 1 pathway by nutrients, growth factors, and stress. Molecular Cell 2010, 40, 310–322. [Google Scholar] [CrossRef]
- Serfontein, J.; Nisbet, R.E. R.; Howe, C.J.; de Vries, P.J. Evolution of the TSC1/TSC2-TOR Signaling Pathway. Science Signaling 2010, 3, ra49. [Google Scholar] [CrossRef]
- Shen, H.M.; Codogno, P. Autophagy is a survival force via suppression of necrotic cell death. Experimental Cell Research 2012, 318, 1304–1308. [Google Scholar] [CrossRef]
- Sheng, X.; Xia, Z.; Yang, H.; Hu, R. The ubiquitin codes in cellular stress response. Protein & Cell 2024, 15, 157–190. [Google Scholar] [CrossRef]
- Sherr, C.J.; McCormick, F. The RB and p53 pathways in cancer. Cancer Cell 2002, 2, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Norberg, E.; Vakifahmetoglu-Norberg, H. Mutant p53 as a regulator and target of autophagy. Frontiers in Oncology 2021, 10, 607149. [Google Scholar] [CrossRef] [PubMed]
- Siau, J.W.; Coffill, C.R.; Zhang, W.V.; Tan, Y.S.; Hundt, J.; Lane, D.; et al. Functional characterization of p53 pathway components in the ancient metazoan. Scientific Reports 2016, 6, 33972. [Google Scholar] [CrossRef] [PubMed]
- Simon, Herbert A. 1962[1996]. The Architecture of Complexity. In H. A. Simon (Ed.), The sciences of the artificial (pp. 183–216). MIT Press.
- Sinha, S.; Levine, N. The autophagy effector Beclin-1: a novel BH3-only protein. Oncogene 2009, 27, S137–S148. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.M.; Szathmáry, E. The origin of chromosomes I. Selection for linkage. Journal of Theoretical Biology 1993, 164, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Sogabe, S.; Hatleberg, W.L.; Kocot, K.M.; Say, T.E.; Stoupin, D.; Roper, K.E.; et al. Pluripotency and the origin of animal multicellularity. Nature 2019, 570, 519–522. [Google Scholar] [CrossRef]
- Sowpati, D.T.; Ramamoorthy, S.; Mishra, R.K. Expansion of the polycomb system and evolution of complexity. Mechanisms of Development 2015, 138, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, J.B.; Haigs, M.C. The multifaceted contributions of mitochondria to cellular metabolism. Nature Cell Biology 2018, 20, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Stielow, B.; Zhou, Y.; Cao, Y.; Simon, C.; Pogoda, H.M.; Jiang, J.; et al. The SAM domain-containing protein 1 (SAMD1) acts as a repressive chromatin regulator at unmethylated CpG islands. Science advances 2021, 7, eabf2229. [Google Scholar] [CrossRef]
- Suga, H.; Ruiz-Trillo, I. Development of ichthyosporeans sheds light on the origin of metazoan multicellularity. Developmental Biology 2013, 377, 284–292. [Google Scholar] [CrossRef]
- Suga, H.; Ruiz-Trillo, I. (2015). Filastereans and Ichthyosporeans: Models to Understand the Origin of Metazoan Multicellularity. In I. Ruiz-Trillo, & A. M. Nedelcu. (Eds.), Evolutionary transition to multicellular life: Principles and mechanisms (pp. 117-128). Springer.
- Suh, E.K.; Yang, A.; Kettenbach, A.; Bamberger, C.; Michaelis, A.H.; Zhu, Z.; et al. p63 protects the female germ line during meiotic arrest. Nature 2006, 444, 624–628. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.X.; et al. SUMO protease SENP1 deSUMOylates and stabilizes c-Myc. Proceedings of the National Academy of Sciences, USA, 2018; 115, 10983–10988. [Google Scholar]
- Szathmáry, E. Toward major evolutionary transitions theory 2.0. Proceedings of the National Academy of Sciences, USA 2015, 112, 10104–10111. [Google Scholar] [CrossRef] [PubMed]
- Szathmáry, E.; Smith, J.M. The major evolutionary transitions. Nature 2003, 374, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Szathmáry, E.; Wolpert, L. (2003). The transition from single cells to multicellularity. In P. Hammerstein (Ed.), Genetic and Cultural Evolution of Cooperation (pp. 271–290). MIT Press.
- Takemura, M. Evolutionary history of the retinoblastoma gene from archaea to eukarya. Biosystems 2005, 82, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.; Fujimoto, M.; Takii, R.; Takaki, E.; Hayashida, N.; Nakai, A. Mitochondrial SSBP1 protects cells from proteotoxic stresses by potentiating stress-induced HSF1 transcriptional activity. Nature Communications 2015, 6, 6580. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Kang, R. (2022). Cell death: Machinery and regulation. In (Eds.), Mechanisms of Cell Death and Opportunities for Therapeutic Development (pp. 47–64). Elsevier.
- Tang, D.; Kang, R.; Berghe, T.V.; Vandenabeele, P. & Kroemer, G. The molecular machinery of regulated cell death. Cell Research 2019, 29, 347–364. [Google Scholar] [PubMed]
- Tatebe, H.; Shiozaki, K. Evolutionary conservation of the components in the tor signaling pathways. Biomolecules 2017, 7, 77. [Google Scholar] [CrossRef] [PubMed]
- Thomas, H.E.; Zhang, Y.; Stefely, J.A.; Veiga, S.R.; Thomas, G.; Kozma, S.C.; et al. Mitochondrial complex I activity is required for maximal autophagy. Cell Reports 2018, 24, 2404–2417. [Google Scholar] [CrossRef] [PubMed]
- Thomer, J. TOR complex 2 is a master regulator of plasma membrane homeostasis. Biochemical Journal 2022, 479, 1917–1940. [Google Scholar] [CrossRef]
- Tian, Y.; Garcia, G.; Bian, Q.; Steffen, K.; Dillin, A. Mitochondrial stress induces chromatin reorganization to promote longevity and UPRmt. Cell 2016, 165, 1197–1208. [Google Scholar] [CrossRef]
- Trigos, A.S.; Pearson, R.B.; Papenfuss, A.T.; Goode, D.L. Somatic mutations in early metazoan genes disrupt regulatory links between unicellular and multicellular genes in cancer. eLife 2019, 8, e40947. [Google Scholar] [CrossRef] [PubMed]
- Trigos, A.S.; Pearson, R.B.; Papenfuss, A.T.; Goode, D.L. Altered interactions between unicellular and multicellular genes drive hallmarks of transformation in a diverse range of solid tumors. Proceedings of the National Academy of Sciences of the United States of America 2017, 114, 6406–6411. [Google Scholar] [CrossRef] [PubMed]
- Trigos, A.S.; Pearson, R.B.; Papenfuss, A.T.; Goode, D.L. How the evolution of multicellularity set the stage for cancer. British Journal of Cancer 2018, 118, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Van Deursen, J.M. The role of senescent cells in ageing. Nature 2014, 509, 439–46. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, R.M.; Kupai, A.; Rothbart, S.B. Chromatin regulation through ubiquitin and ubiquitin-like histone modifications. Trends in Biochemical Sciences 2021, 46, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Vaseva, A.V.; Marchenko, N.D.; Ji, K.; Tsirka, S.E.; Holzmann, S.; Moll, U.M. p53 opens the mitochondrial permeability transition pore to trigger necrosis. Cell 2012, 149, 1536–1548. [Google Scholar] [CrossRef] [PubMed]
- Veiga-Fernades, H.; Pachnis, V. Neuroimmune regulation during intestinal development and homeostasis. Nature Immunology 2017, 18, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Venniyoor, A. PTEN: A thrifty gene that causes disease in times of plenty? Frontiers in Nutrition 2020, 7, 81. [Google Scholar] [CrossRef] [PubMed]
- Ventura, R. Multicellular individuality: The case of bacteria. Biological Theory 2019, 14, 131–140. [Google Scholar] [CrossRef]
- Vertegaal, A.C. O. Signalling mechanisms and cellular functions of SUMO. Nature Reviews, Cell and Molecular Biology 2022, 23, 715–731. [Google Scholar] [CrossRef]
- Vizioli, M.G.; Liu, T.; Miller, K.N.; Robertson, N.A.; Gilroy, K.; Lagnado, A.B.; et al. Mitochondria-to-nucleus retrograde signaling drives formation of cytoplasmic chromatin and inflammation in senescence. Genes & Development 2020, 34, 428–445. [Google Scholar]
- Wagner, G.P.; Erkenbrack, E.M.; Love, A.C. Stress-induced evolutionary innovation: A mechanism for the origin of cell types. BioEssays 2019, 41, 1800188. [Google Scholar] [CrossRef] [PubMed]
- Walston, H.; Iness, A.N.; Litovchick, L. DREAM On: Cell Cycle Control in Development and Disease. Annual Review of Genetics 2021, 55, 309–329. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yang, Y.; Xu, Y. Brain nuclear receptors and cardiovascular Function. Cell & Bioscience 2023, 13, 14. [Google Scholar] [CrossRef]
- Wanner, E.; Thoppli, H.; Riabowol, K. Senescence and Apoptosis: Architects of Mammalian Development. Frontiers in Cell and Developmental Biology 2021, 8, 620089. [Google Scholar] [CrossRef] [PubMed]
- Watson, I.R.; Irwin, M.S. Ubiquitin and ubiquitin-like modifications of the p53 family. Neoplasia 2006, 8, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Weissman, Irving L. Stem cells are units of natural selection for tissue formation, for germline development, and in cancer development. Proceedings of the National Academy of Sciences USA 2015, 112, 8922–8928. [Google Scholar] [CrossRef] [PubMed]
- Whelan, N.V.; Kocot, K.M.; Moroz, T.P.; Mukherjee, K.; Williams, P.; Paulay, G.; et al. Ctenophore relationships and their placement as the sister group to all other animals. Nature Ecology & Evolution 2017, 1, 1737–1746. [Google Scholar]
- White, E. Autophagy and p53. Cold Spring Harbor Perspectives in Medicine 2016, 6, a026120. [Google Scholar] [CrossRef]
- White, E.; Lattime, E.C.; Guo, J.Y. (2021). Autophagy Regulates Stress Responses, Metabolism, and Anticancer Immunity. Trends in Cancer 2021, 7, 778–789. [Google Scholar] [CrossRef]
- Wolpert, L.; Szathmáry, E. Evolution and the egg. Nature 2002, 420, 745. [Google Scholar] [CrossRef] [PubMed]
- Woodland, H.R. The Birth of Animal Development: Multicellularity and the Germline. Current Topics in Developmental Biology 2016, 117, 609–630. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; DeGregori, J.; Shohet, R.; Leone, G.; Stillman, B.; Nevins, J.R.; Williams, R.S. Cdc6 is regulated by E2F and is essential for DNA replication in mammalian cells. Proceedings of the National Academy of Sciences USA 1998, 95, 3603–3608. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Shen, W.H. PTEN: A new guardian of the genome. Oncogene 2008, 27, 5443–5453. [Google Scholar] [CrossRef] [PubMed]
- Yonekawa, T.; Thorburn. A. Autophagy and cell death. Essays in Biochemistry 2013, 55, 105–117. [Google Scholar] [PubMed]
- Yu, J.; Zhang, L. PUMA, a potent killer with or without p53. Oncogene 2009, 27, S71–S83. [Google Scholar] [CrossRef] [PubMed]
- Zachar, I.; Szathmáry, E. Breath-giving cooperation: Critical review of origin of mitochondria hypotheses. Biology Direct 2017, 12, 19. [Google Scholar] [CrossRef] [PubMed]
- Zatulovskiy, E.; Zhang, S.; Berenson, D.F.; Topacio, B.R.; Skotheim, J.M. Cell growth dilutes the cell cycle inhibitor Rb to trigger cell division. Science 2020, 369, 466–471. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, H.; Li, M.; Rayburn, E.R.; Agrawal, S.; Zhang, R. Stabilization of E2F1 protein by MDM2 through the E2F1 ubiquitination pathway. Oncogene 2005, 24, 7238–7247. [Google Scholar] [CrossRef]
- Zhang, S.; Hama, Y.; Mizushima, N. The evolution of autophagy proteins-diversification in eukaryotes and potential ancestors in prokaryotes. Journal of Cell Science 2021, 134, jcs233742. [Google Scholar] [CrossRef]
- Zhao, X.; Bramsiepe, J.; Van Durme, M.; Komaki, S.; Prusicki, M.A.; Maruyama, D.; et al. Retinoblastoma related1 mediates germline entry in arabidopsis. Science 2017, 356, eaaf6532. [Google Scholar] [CrossRef]
- Zhong, Z.; Sanchez-Lopez, E.; Karin, M. Autophagy, inflammation, and immunity: A Troika governing cancer and its treatment. Cell 2016, 166, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Wu, X.; Zhou, J.; Li, X.; Tian, Y. NuRD mediates mitochondrial stress-induced longevity via chromatin remodeling in response to acetyl-CoA level. Science Advances 2020, 6, eabb2529. [Google Scholar] [CrossRef] [PubMed]
| 1 | Although pseudo- or quasi- multicellularity exists in bacteria and Archaea (e.g., myxobacteria, cyanobacteria), genuine multicellularity, simple and then complex, exists only in eukaryotes (cf. Ventura 2019). |
| 2 | For a complete picture, we also include “(extra-body) exchange”, denoting the relationship between an organism and its environment. |
| 3 | RB-E2F is perhaps more useful for aerobic than anaerobic metabolism, and (most) metazoans are aerobic. This also suggests that RB-E2F is indispensable for genuinely multicellularity in metazoans (Attardi & Sage 2013). |
| 4 | In this sense, sex itself should not be considered to be a major transition (Herron 2021; see also Michod 2011, 186-9). More appropriately, sex laid a key foundation for the emergence of more complex individuality in multicellular organisms, as an unintended consequence. |
| 5 | Regulated cell death is perhaps a more fitting term than the more widely known programmed cell death (PCD). It is now commonly accepted that ferroptosis is not a distinct form of PANoptosis. For instance, ferroptosis is also connected with mitochondria and the p53 pathway. |
| 6 | Thus, components of PCD pathways in unicellular eukaryotes may function primarily as part of the mitochondria regulating or protecting machinery. |
| 7 | This fact is consistent with the more recent finding that ctenophores (comb jelly fish), rather than poriferas are the more likely sister group to all other animals (Schultz et al. 2023). See also Whelan et al. 2017. |
| 8 | The level of “intra-body but extra-cellular communication” is highlighted to denote the one that distinguishes genuine multicellular organisms such as metazoans from unicellular organisms. |
| 9 | In mammalians, Beclin-2 (BECN2), a homologue of Beclin-1 (and ATG-6), plays similar but non-identical functions in key pathways (He et al. 2013; Galluzzi & Kroemer 2013; Quiles et al. 2023). |
Figure 1.
The schematic structure of coordination hubs and pathways in LECA.
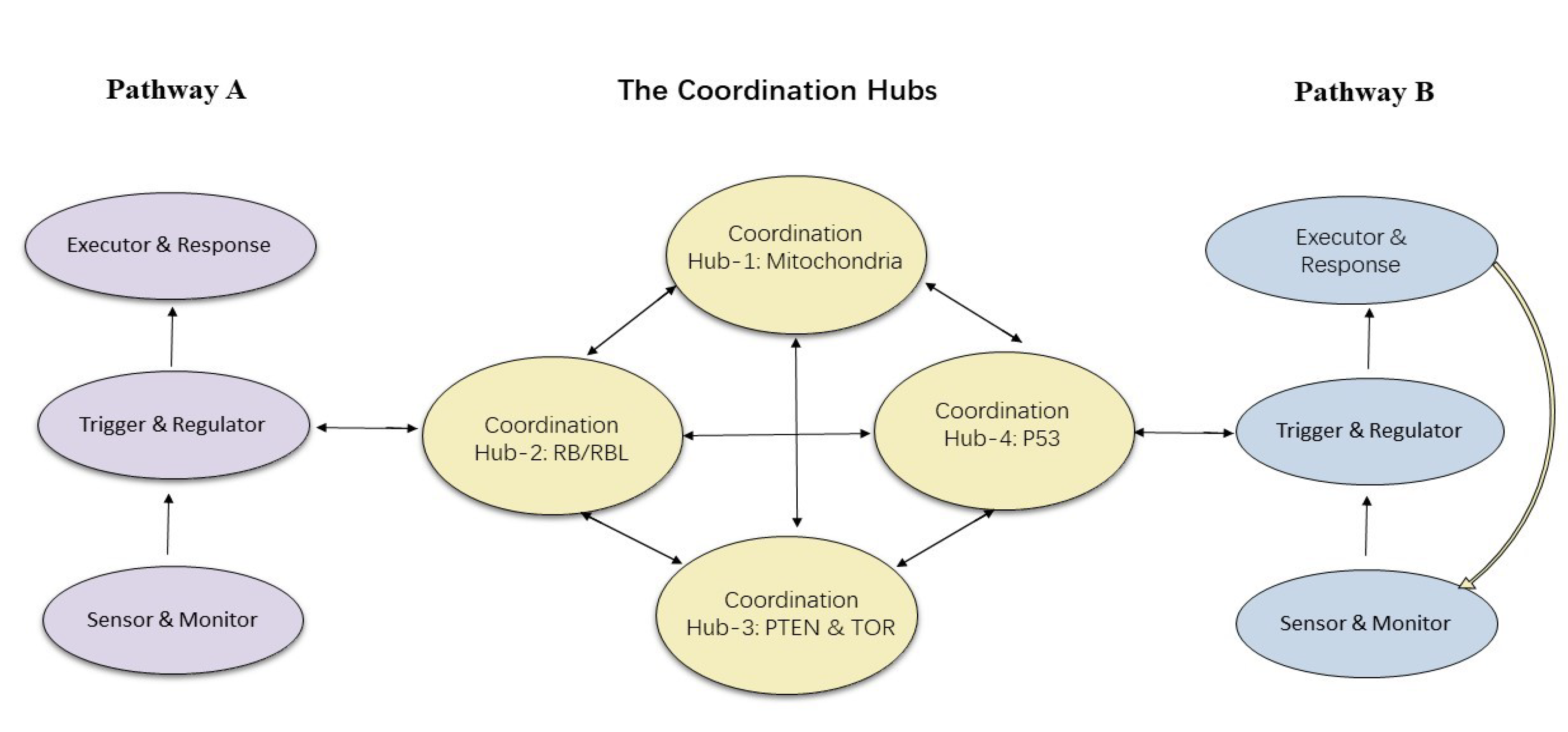

Table 1.
Means of coordination in unicellular eukaryotes since LECA.
| Type | Sub-category | Example |
|---|---|---|
| Cross-talk (intracellular) | Direct: via: 1) shared components and links, and 2) regulation of the expression of downstream genes. |
|
| Indirect: via coordination hubs. | RB-E2F, PTEN, TOR | |
| Intra-cellular (and intra-body) communication | Regular and irregular: between different cellular apparatuses and organelles | Between the nucleus, cytoplasm, and the mitochondria, vis targeting and regulation of gene expression. Intracellular stress signals. |
| Extra-cellular (and extra-body) exchange2 | Regular and irregular: between the organism and the external environment | Food and waste, nutritional changes, extra-body stresses, damages, and invasions. |
Table 2.
Cross-talks and communications in metazoans: a more complex picture.
| Type | Sub-category | Example |
|---|---|---|
| Cross-talk (intra-cellular) | Direct: via 1) gene expression, 2) via shared components and links | RB-E2F and p53-MDM2; apoptosis and autophagy |
| Indirect: via coordination hub. | RB-E2F, PTEN, TOR, p53-MDM2 | |
| Intra-body and intra-cellular communication | Regular and irregular: between different cellular apparatuses and organelles | Between the nucleus, cytoplasm, and the mitochondria, vis targeting and regulation of gene expression. Intracellular stress signals. |
| Intra-body but extra-cellular communication8 | Regular and irregular: between different cell types, tissues, and organs. | Hormone (e.g., insulin) and other extracellular ligands (e.g., lipids). Other intra-body stresses as signals |
| Extra-body (and extra-cellular) exchange | Regular and irregular: between the organism and the external environment | Food and waste, seasonal change, nocturnal cycles, and other extra-body inputs and outputs. Extra-body stresses, toxins, damages, and invasions. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



