
Submitted:
30 May 2024
Posted:
30 May 2024
You are already at the latest version
Abstract
The gastrointestinal tract has a pivotal role in nutrient absorption, immune function, and overall homeostasis. The ileum segment of the small intestine plays respective roles in nutrient breakdown and absorption. The purpose of this study was to investigate the impact of heat-induced oxidative stress and the potential mitigating effects of an astaxanthin antioxidant treatment on the ileum of broilers. By comparing the growth performance and gene expression profiles among three groups- thermal neutral, heat stress and heat stress with astaxanthin, under two temperature conditions of 21-22°C and 32-35°C, this research aims to elucidate the role of astaxanthin in supporting homeostasis and cellular protection in the ileum. Results showed both treatments under heat stress experienced reduced growth performance, while the group treated with astaxanthin showed a slightly lesser decline. Results further showed the astaxanthin treatment group significantly upregulated in the cytoprotective gene expression for HSF2, SOD2, GPX3, and TXN, as well as the upregulation of epithelial integrity genes, LOX, CLDN1 and MUC2. In conclusion, our experimental findings demonstrate upregulation of cytoprotective and epithelial integrity genes suggests astaxanthin may effectively enhance the cellular response to heat stress to mitigate oxidative damage, and contribute to cytoprotective capacity.
Keywords:
astaxanthin
; cytoprotective capacity
; ileum
; heat stress
; oxidative stress
; broilers
1. Introduction
The gastrointestinal tract (GIT) is essential for digesting food into basic components for optimal nutrient absorption, crucial for poultry production and welfare. Bacteria in the GIT protect against disease, stimulate the immune system, and enhance growth rates by producing extra nutrients through fermentation of non-digestible fibers [1]. The chicken GIT consists of five regions, each with a specific role: the crop, proventriculus, gizzard, small intestine (duodenum, jejunum, ileum), and large intestine (caeca, colon, rectum). In the crop, food is partially fermented before entering the proventriculus, and acid and enzymes aid in protein breakdown. The gizzard grinds food into smaller particles for the small intestine, where most nutrient absorption occurs, leaving behind non-digestible components. The caeca further ferment these components, forming organic acids, short-chain fatty acids, and vitamins for absorption [2,3]. The small intestine, the primary site for nutrient absorption, is divided into the duodenum, jejunum, and ileum. Digestive enzymes and bile aid nutrient breakdown, and villi increase absorption. The ileum absorbs remaining nutrients and plays a role in immune function via Peyer’s patches [1,4]. Impaired GIT function leads to reduced nutrient absorption, poor feed conversion efficiency, and higher feed costs. Dysbacteriosis, an imbalance of gut bacteria, negatively affects chickens by reducing nutrient absorption, often causing increased feed consumption to meet nutritional demands [1,5,6].
Heat-induced oxidative stress profoundly affects intestinal ileum tissue and contributes to gastrointestinal disorders such as inflammatory bowel disease, intestinal ischemia, and intestinal permeability dysfunction [7,8,9]. The balance between reactive oxygen species (ROS) and antioxidant defenses in the ileum is crucial for cellular homeostasis and function. Enhancing cytoprotective capacity with compounds or substances can protect cells from harmful stimuli [10,11,12]. Cytoprotective capacity refers to the ability to shield cells from damage or death caused by various stressors, including oxidative stress, toxins, radiation, and inflammation [13,14]. Antioxidants, such as superoxide dismutase (SOD), catalase, and glutathione peroxidase (GPX), neutralize ROS, but their effectiveness can diminish under elevated temperatures, increasing ROS and oxidative stress [15,16,17]. Thioredoxin (Trx) plays a vital role in cellular redox homeostasis and oxidative stress defense by maintaining a reducing environment through thiol-disulfide exchange reactions. Under oxidative stress, thioredoxin activity is upregulated to enhance antioxidant defenses, protecting cells from ROS damage [18]. Oxidative stress in the gastrointestinal system can compromise intestinal mucosa integrity, disrupt cellular function, and cause inflammation. Understanding heat-induced oxidative stress mechanisms in the small intestine is essential for developing strategies to protect intestinal health [19]. Heat shock proteins, produced under stress, assist in protein folding and function, protecting cells from damage. Heat shock response, activated by elevated temperatures, involves heat shock factor 2 (HSF2) binding to heat shock elements in gene promoters. HSF2 helps manage protein misfolding and aggregation, maintaining cellular homeostasis and preventing cell damage or death [20,21,22].
The ileum, the final section of the small intestine, is crucial for nutrient absorption before passing digested material to the ceca. Its wall is lined with villi, tiny projections that increase the surface area for absorption. Villi contain capillaries that transport amino acids and glucose to the hepatic portal vein and liver [19,23,24]. The LOX gene encodes lysyl oxidase, an enzyme essential for crosslinking collagen and elastin fibers in the extracellular matrix (ECM), vital for connective tissue integrity and stability. Although not primarily expressed in the epithelium, LOX activity is important in epithelial function, particularly under oxidative stress, aiding in wound healing and tissue repair by reorganizing and strengthening the ECM [25,26]. Claudins (CLDN) are integral membrane proteins fundamental to forming and regulating tight junctions (TJs) in epithelial and endothelial cell barriers. TJs control the passage of ions, water, and solutes across cells, maintaining barrier integrity. Changes in CLDN expression can affect tight junction permeability, with certain isoforms enhancing or compromising barrier function [27]. The ileal mucosa protects and maintains epithelial integrity, partly through the mucin 2 (MUC2) gene, which synthesizes mucin 2 glycoprotein to maintain the mucus layer, protecting cells from oxidative damage. Disruption of these factors can increase epithelial permeability. The mucosa regulates gut inflammation, producing anti-inflammatory factors and specialized mediators to restore and maintain epithelial integrity. It interacts with gut microbiota to balance beneficial and harmful microbes, supporting a healthy mucosa. Mucosal immune responses defend against pathogens and support beneficial bacteria, contributing to a healthy and functional barrier in the small intestine [28,29].
Astaxanthin (AST), a potent lipid-soluble antioxidant from the carotenoid family, exhibits remarkable free radical scavenging and anti-inflammatory properties [30,31]. Its unique chemical structure, with hydroxyl and keto moieties on each ionone ring, contributes to its superior antioxidant activity, protecting cellular membranes against oxidation [32]. Although naturally occurring in aquatic organisms and birds, astaxanthin cannot be synthesized by animals and must be obtained from the diet [33]. Common natural sources include green algae, red yeast, and crustacean byproducts, with the highest concentration found in Haematococcus pluvialis [34]. Astaxanthin supplementation in humans has shown positive health benefits, including anti-inflammatory, immunomodulatory, cardiovascular, neuroprotective, and anticancer effects [31].
Our research question is, “Does astaxanthin reduce oxidative stress in the broiler ileum, and what genes are responsible for regulating its cytoprotective capacity and endothelial integrity?” We hypothesize that heat stress induces oxidative stress in the broiler ileum correlating with poor growth performance, and tissue structure degradation; and H. pluvialis-derived astaxanthin dietary supplementation mitigates the effects through upregulation of cytoprotective and endothelial integrity gene expression.
2. Materials and Methods
2.1. Ethics Statement
Animal protocol (Protocol No. 17-2605) used in this study was approved by the University of Hawaii Institutional Animal Care and Use Committee (IACUC). Animals were raised under animal welfare guidelines and euthanized in accordance with humane protocols in preparation for necropsy.
2.2. Experimental Animal Design
Cob 500 unsexed broiler chicks were obtained from Asagi Hatchery (Honolulu, Hawaii, USA). Several mitigation strategies were simultaneously tested in parallel and the findings reported separately based on treatment. In this trial the feed additive treatment is AST. The animals were raised from Day 0 to 6 weeks in deep litter pens with pine shavings on concrete flooring, four birds in each pen. The animals were reared under two temperature conditions: TN (n=24) at 21-22°C and 50% RH, and HS (n=36) at 32-35°C and 42-50% RH. Animals were provided a normal starter feed from 0 to 21 days, and a normal finisher feeds on 22-42 days with free access to feed and water. Nutritional compositions of the supplemented diets are listed in Table 1. After 14 days, the HS group was further divided into two dietary regimens, i.e., basal diet HS (n=18, treatment 1), and basal diet with 1.33 mg/kg AX supplement HSAX (n=18, treatment 2). The light cycle was set at 1:23 dark:light cycles throughout the trial. The broilers were euthanized on day 42, and ileum tissue samples from randomly selected 6 birds of each group were collected at necropsy.
2.3. Astaxanthin-Rich Dietary Supplement
The diet was supplemented with P25HB provided by AstaReal®, Inc. (Burlington, New Jersey, USA). PH25B contains 2.5% (w/w) dried Haematococcus pluvialis-algae, and other components including modified starch, gum Arabic, mixed tocopherols, L-ascorbyl palmitate, silicon dioxide, xanthan gum, γ –cyclodextrin, polysorbate 80, rosemary extract and ferulic acid. A comparable nutritional composition of H. pluvialis-algae is listed in Appendix A (Table A1). The natural forms of astaxanthin comprise of mainly mono-esterified, followed by di-esterified and free forms, 3,3’ –dihydroxy-β, β-carotene-4, 4’ –dione (C40H5204 free form).
2.4. Growth Performance
Weekly feed intake was recorded, and the average daily feed intake (ADFI), average daily gain ratio (ADGR), and feed conversion ratio (FCR) were calculated. The body weight (BW) of each bird was recorded using a Mettler Toledo scale before heat stress treatment and at the end of the heat treatment.
2.5. Tissue Sample Collection
Immediately after euthanizing, ileum tissues were collected from randomly selected 6 birds of each group, snapped-frozen in liquid nitrogen, and stored at −80°C.
2.6. Total RNA extraction and cDNA preparation
Total RNA was isolated from frozen tissues (50–100mg) using TRIzol reagent (Invitrogen, Carlsbad, CA) according to the manufacturer’s instructions. The concentration of total RNA was determined using NanoPhotometer® P330 (IMPLEN, Los Angeles, CA). Complementary DNA (cDNA) was synthesized from 1 µg of total RNA (20 µl reaction of RT mixture) using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA), and further diluted with nuclease free–water (1:25) for the qPCR reaction below.
2.7. Bioinformatics: Genome Assembly and Gene Primer Design
The National Center for Biotechnology Information (NCBI) genome browser was used to search and compile genes for Gallus gallus domesticus related to heat stress, oxidative stress, cytoprotective, epithelial integrity, transcription factors and housekeeping genes. The NCBI-Basic Local Alignment Search Tool (BLAST) was used to design primers for polymerase chain reaction (PCR) from the accession numbers obtained from the list of genes (Table 2). The primer parameters were set for a PCR product size between a minimum of 100 and maximum of 250 for 5 primers to return. The primer melting temperatures were set for a minimum of 55°C, optimum of 57°C, and maximum of 60°C with a maximum Tm difference of 3°C. The exon junction span was set that the primers must span an exon-exon junction. The organism specified was Gallus gallus (taxid 9031). The forward and reverse primer sequences (5’->3’) were then submitted to Integrated DNA Technologies (Coralville, IA, USA) for synthesis.
2.8. Quantitative Real-Time RT-PCR (qPCR)
The qPCR was performed using PowerUp SYBR Green Master Mix (Applied Biosystems, Foster City, CA, USA) on a StepOne Plus real-time PCR system (Applied Biosystems, Foster City, CA, USA). The qPCR reaction mixture consisted of 3 µl of cDNA, 5 µl PowerUp SYBR Green Master Mix, and 1 µl of each forward and reverse primers (5 µmol concentration) to make a final reaction mixture of 10 µl. Specific primer pairs for the detection of each gene were designed using the NCBI Primer-Blast tool. The qPCR reaction was carried out following standard cycling mode. The amplification conditions were 50°C for 2 min (hold), 95°C for 2 min (hold), followed by 40 repeat cycles of 95°C for 15 s (denaturation), 60°C for 15 s (annealing), and 72°C for 1 min (extension). A melting curve was also generated to confirm SYBR Green-based objective amplicon, and further qPCR products were confirmed using 2% agarose gel electrophoresis. Three house-keeping genes Glyceraldehyde 3-phosphate dehydrogenase (GAPDH), Beta-actin (β-actin), and TATA-Box Binding Protein (TBP) were analyzed in triplicates in each bird to determine the most stable house-keeping gene. Based on the uniformity of expression level across samples, β-actin was chosen as the housekeeping gene. Gene expression level was determined using cycle threshold (Ct) values following the standard curve method after normalization with housekeeping genes. Fold change for each gene was calculated using the 2−ΔΔCt method. Data for fold change were presented as mean ± standard error [35].
2.9. Gene Ontology
Significantly differentially expressed genes identified from the qPCR procedure were searched in the Ensembl genome database for chicken (GRCg6a) species to obtain Gene ontologies (GO) information (https://uswest.ensembl.org/Gallus_gallus/Info/Index) [36]. The GO included the cellular component, molecular function and biological process of these genes identified with ENSGAL Transcript IDs.
2.10. Tissue Histology Using Hematoxylin and Eosin Staining
A section of the ileum was placed in a scintillating vial with 10% neutral buffered formalin (NBF) and stored at ambient temperature in preparation for Hematoxylin and Eosin (H&E) staining slides. Cross-sectional pieces of ileum were also prepared and fixed overnight in 4% Paraformaldehyde (PFA), then incubated in 15% sucrose solution for 3 hr., embedded in Optimal Cutting Temperature (OCT) compound, and stored at −80°C. The ileum tissues were sectioned at 6μm thickness, stained with H&E, and mounted to slides at the Histology Core Facility at the John A. Burns School of Medicine, University of Hawaii at Manoa. Slides were observed under both 5X and 20X objective lens and images were captured on a Leica Thunder microscope (Danaher, Illinois, USA). The criterion for villus selection was based on the presence of intact lamina propria, and fifteen villi were assessed per sample. Villus height (VH) was measured from the tip of the villus to the crypt, crypt depth (CD) from the villus base to the submucosa, villus surface area (VSA) calculated as the villus base width plus the villus apical width divided by two, and the ratio of villus height to crypt depth (VH:CD) [19,23,37]. The measurements were recorded using Image J Fiji software ver 1.54i [38].
2.11. Statistical Analysis
The statistical analysis was performed using the Kruskal-Wallis rank sum test with statistical significance set at p<0.05, followed by the Dunn post hoc test for comparison between three groups: TN, HS, and HSAX, and the p-value adjusted using the Bonferroni method. Growth performance measurements were calculated based on the data collected at the end of the 42-day trial period. Analysis was conducted using the R open source program, libraries ‘FSA’, ‘dunn.test’, and ‘gplots’, R Core Team (2023) (https://www.R-project.org/) [39].
3. Results
3.1. Growth Performance
At the end of the 42-day poultry trial, the TN group growth performance indicators were found to be significantly higher for BW, ADFI, ADGR, compared to the HS and HSAX groups by conducting a Kruskal-Wallis test to evaluate differences among the three groups (p<0.01). Although the results clearly showed negative impacts of heat on the HS and HSAX groups, when performing the post hoc pairwise comparisons using Dunn’s test and applying the Bonferroni correction to control increased risk of error due to multiple comparisons, the negative impacts were more statistically significant comparing HS vs TN (p<0.01), rather than HSAX vs TN (p<0.05). In addition, the FCR showed the TN group was significantly lower in feed conversion, requiring less feed to maintain body weight compared to the HS vs TN, and HSAX vs TN (p<0.05). (Table 3, Figure 1).
3.2. Quantitative Real-time RT-PCR (qPCR) Gene Expression
For gene expression studies, three genes were considered for housekeeping genes, GAPDH (glyceraldehyde-3-phosphate dehydrogenase), ACTB (β-actin), and TBP (TATA-box binding protein). β-actin was selected for its high and relatively stable expression under experimental conditions performed, making it a reliable reference for normalizing the gene expression data collected.
3.2.1. Statistically Significant Genes Identified in Differential Expression Analysis

Table 4.
Genes exhibiting significant differential expression for the ileum tissue of Cob-500 broilers with β-actin as the housekeeping gene for normalization. Three groups were compared: TN, HS, and HSAX. Statistical analysis based on the Kruskal-Wallis rank sum test, followed with the Dunn post hoc test for comparisons, and p-value adjusted with the Bonferroni method. * 0.05>p>0.01, ** 0.01>p>0.001.
Table 4.
Genes exhibiting significant differential expression for the ileum tissue of Cob-500 broilers with β-actin as the housekeeping gene for normalization. Three groups were compared: TN, HS, and HSAX. Statistical analysis based on the Kruskal-Wallis rank sum test, followed with the Dunn post hoc test for comparisons, and p-value adjusted with the Bonferroni method. * 0.05>p>0.01, ** 0.01>p>0.001.
| Kruskal-Wallis | Dunn/Bonferroni p-value | |||
| Gene | p-value | TN vs HS | TN vs HSAX | HS vs HSAX |
| HSF1 | 0.1495 | ns | ns | ns |
| HSF2 | 0.0222 | ns | ns | 0.0174 |
| HSF3 | 0.2199 | ns | ns | ns |
| HSP90 | 0.1299 | ns | ns | ns |
| HIFIA | 0.8948 | ns | ns | ns |
| SOD1 | 0.1990 | ns | ns | ns |
| SOD2 | 0.0030 | 0.0916 | ns | 0.0024 |
| ROS-1 | 0.3722 | ns | ns | ns |
| GPX1 | 0.0745 | ns | ns | ns |
| GPX3 | 0.0274 | ns | 0.0331 | ns |
| HMOX1 | 0.8490 | ns | ns | ns |
| CAT | 0.8342 | ns | ns | ns |
| CASPAS3 | 0.1299 | ns | ns | ns |
| GSTTI | 0.5225 | ns | ns | ns |
| TXN | 0.0018 | ns | 0.0520 | 0.0016 |
| TXN2 | 0.1034 | ns | ns | ns |
| PRDX1 | 0.0515 | ns | ns | ns |
| LOX | 0.0131 | ns | 0.0105 | ns |
| MYB | 0.0589 | ns | ns | ns |
| IL-4 | 0.1169 | ns | ns | ns |
| CLDN1 | 0.0214 | ns | ns | 0.0174 |
| OCLN | 0.3679 | ns | ns | ns |
| MUC2 | 0.0035 | 0.0798 | ns | 0.0029 |
| CDH1 | 0.5671 | ns | ns | ns |
Figure 2.
Gene expression fold change heatmap for ileum tissue of Cob-500 broilers. Three groups were compared: TN, HS, and HSAX. The HSAX group expression values show shades of red with significant higher expression, whereas, blue indicates lower levels of expression in the TN and HS groups.
Figure 2.
Gene expression fold change heatmap for ileum tissue of Cob-500 broilers. Three groups were compared: TN, HS, and HSAX. The HSAX group expression values show shades of red with significant higher expression, whereas, blue indicates lower levels of expression in the TN and HS groups.
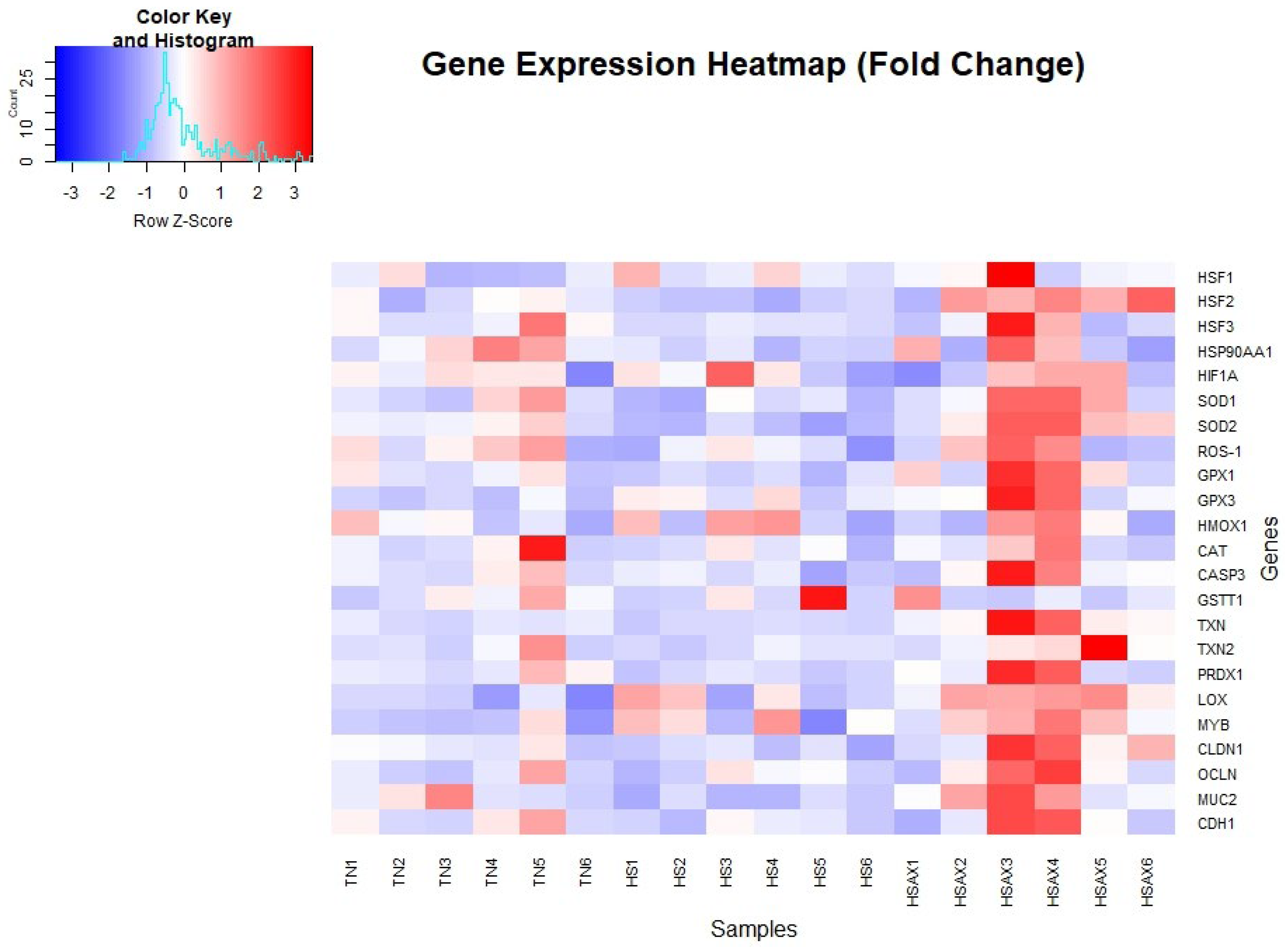
3.2.2. Cytoprotective Capacity Genes
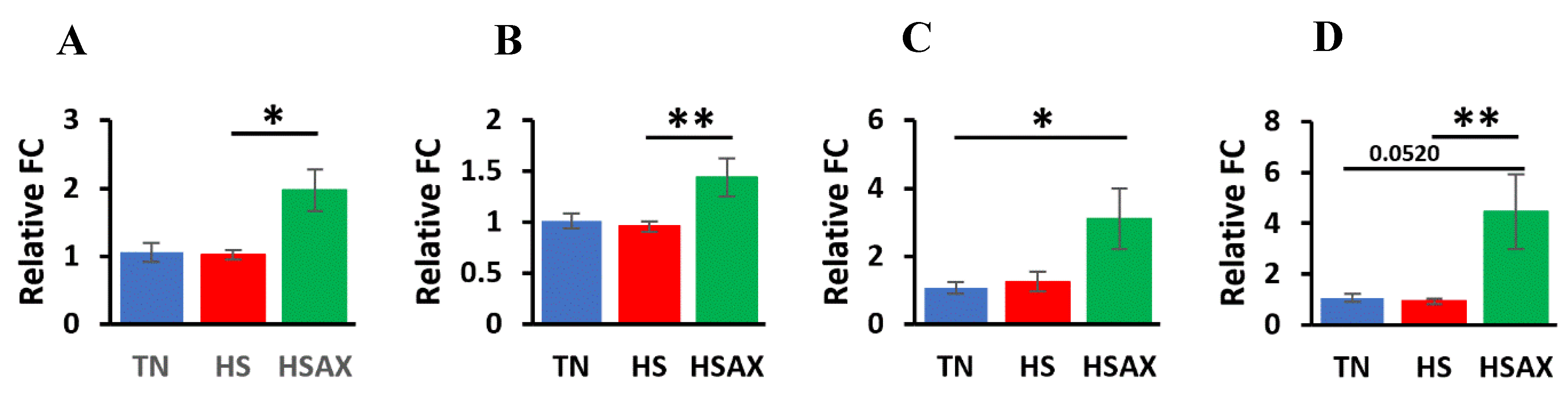
The studies showed the impact of HS and AOX treatment expressed through the cytoprotective capacity genes which resulted in the HSAX treatment group upregulated as compared to both the TN and HS groups. Gene expression results of the Heat Shock Factor 2 (HSF2), HS vs HSAX (p=0.0174), Superoxide Dismutase 2 (SOD2), HS vs HSAX (p=0.0024) and Thioredoxin (TXN), HS vs HSAX (p=0.0016) genes showed significant upregulation observed in the HSAX group as compared to the HS group. While there was minimal significance of TXN upregulation comparing TN vs HSAX (p=0.0520), Glutathione Peroxidase (GPX3), TN vs HSAX (p=0.0331) showed significant upregulation observed in the HSAX as compared to the TN group.
Figure 3.
Effects of heat stress and astaxanthin treatment in the cytoprotective capacity gene expression in the ileum tissue of Cob-500 broilers. Three groups were compared: TN, HS, and HSAX. Kruskal-Wallis test and Dunn post hoc test for statistical significance represented: (A) HSF2, KW p=0.0222, HS vs HSAX (p=0.0174); (B) SOD2, KW p=0.0030, HS vs HSAX (p=0.0024); (C) GPX3, KW p=0.0274, TN vs HSAX (p=0.0331); and (D) TXN, KW p=0.0018, TN vs HSAX (p=0.0520) and HS vs HSAX (p=0.0016).
Figure 3.
Effects of heat stress and astaxanthin treatment in the cytoprotective capacity gene expression in the ileum tissue of Cob-500 broilers. Three groups were compared: TN, HS, and HSAX. Kruskal-Wallis test and Dunn post hoc test for statistical significance represented: (A) HSF2, KW p=0.0222, HS vs HSAX (p=0.0174); (B) SOD2, KW p=0.0030, HS vs HSAX (p=0.0024); (C) GPX3, KW p=0.0274, TN vs HSAX (p=0.0331); and (D) TXN, KW p=0.0018, TN vs HSAX (p=0.0520) and HS vs HSAX (p=0.0016).
3.2.3. Epithelial Integrity Genes
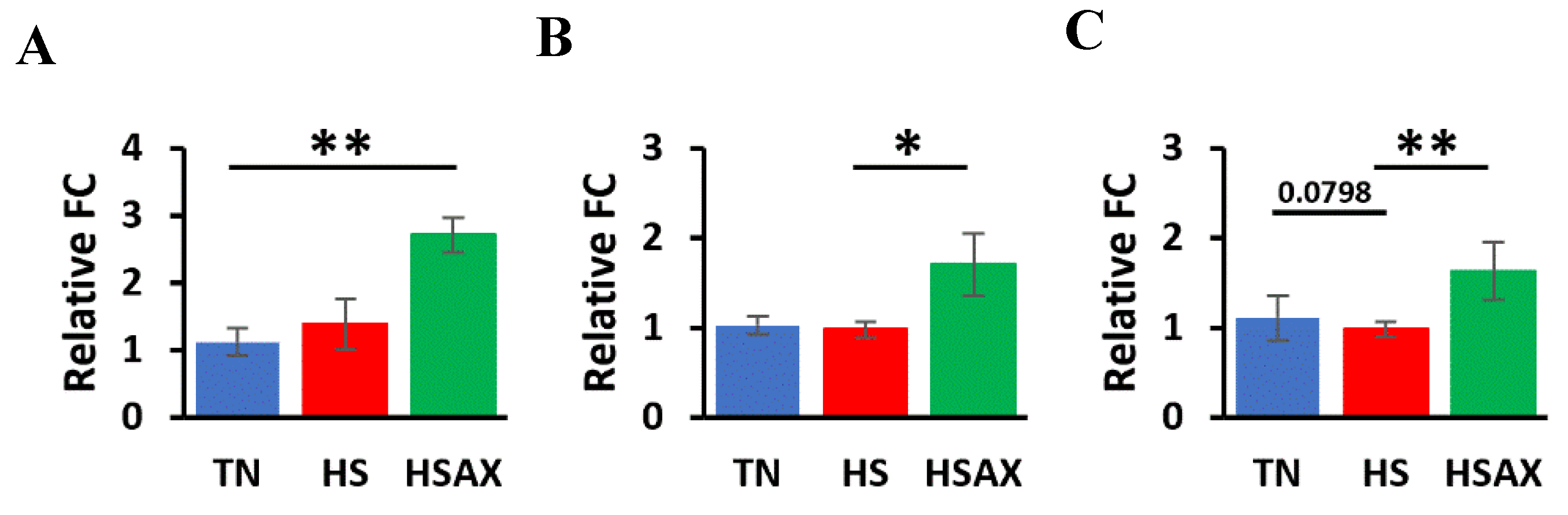
The studies showed the impact of HS and AOX treatment on the expression of the epithelial integrity genes which resulted in the HSAX treatment group upregulated as compared to both the TN and HS groups. Gene expression results of Lysyl Oxidase (LOX), TN vs HSAX (p=0.0105), showed significant upregulation observed in the HSAX as compared to the TN group, while there was no significance between the TN and HS groups. Gene expression results of Claudin 1 (CLDN1), HS vs HSAX (p=0.0174), and Mucin 2 (MUC2), HS vs HSAX (p=0.0029), showed significant upregulation observed in the HSAX as compared to the HS group, and while there was no significance between the TN and HS groups for CLDN1, the TN group was marginally upregulated over the HS group in the MUC2 TN vs HS (p=0.0798) gene expression.
Figure 4.
Effects of heat stress and astaxanthin treatment in the epithelial integrity gene expression in ileum tissue of Cob-500 broilers. Three groups were compared: TN, HS, and HSAX. Kruskal-Wallis test and Dunn post hoc test for statistical significance represented: (A) LOX, KW p=0.0131, TN vs HSAX (p=0.0105); (B) CLDN1, KW p=0.0214, TN vs HSAX (p=0.0174); (C) MUC2, KW p=0.0035, HS vs HSAX (p=0.0029).
Figure 4.
Effects of heat stress and astaxanthin treatment in the epithelial integrity gene expression in ileum tissue of Cob-500 broilers. Three groups were compared: TN, HS, and HSAX. Kruskal-Wallis test and Dunn post hoc test for statistical significance represented: (A) LOX, KW p=0.0131, TN vs HSAX (p=0.0105); (B) CLDN1, KW p=0.0214, TN vs HSAX (p=0.0174); (C) MUC2, KW p=0.0035, HS vs HSAX (p=0.0029).
3.2.4. Gene Ontology (GO) Gene Enrichment
Table 5.
Gene ontology of differentially expressed genes in the ileum of 6-week Cob-500 broilers exposed to 21 days of heat stress and astaxanthin treatment (Ensembl)[36].
Table 5.
Gene ontology of differentially expressed genes in the ileum of 6-week Cob-500 broilers exposed to 21 days of heat stress and astaxanthin treatment (Ensembl)[36].
| Gene | Cellular component | Molecular function |
Biological process |
Transcript IDs |
| HSF2 (heat shock transcription factor 2) |
nucleus, cytoplasm | DNA-binding transcription factor activity, DNA-binding transcription activator activity, RNA polymerase II-specific, RNA polymerase II cis-regulatory region sequence-specific DNA binding | cellular response to heat, positive regulation of transcription from RNA polymerase II promoter in response to heat stress, regulation of transcription, DNA-templated | ENST00000368455 ENST00000452194 |
| SOD2 (superoxide dismutase 2, mitochondrial) | mitochondrion | superoxide dismutase activity, oxidoreductase activity, manganese ion binding, metal ion binding, identical protein binding | response to oxidative stress, oxidation-reduction process, negative regulation of oxidative stress-induced intrinsic apoptotic signaling pathway, response to hydrogen peroxide, removal of superoxide radicals | ENSGALT00000019062 |
| GPX3 (glutathione peroxidase 3) | extracellular space | glutathione peroxidase activity, selenium binding, identical protein binding | response to oxidative stress, hydrogen peroxide catabolic process | ENSGALG00010016480 |
| TXN (thioredoxin) | nucleus, cytosol, extracellular region | protein disulfide oxidoreductase activity | oxidation-reduction process, cell redox homeostasis, negative regulation of hydrogen peroxide-induced cell death, positive regulation of peptidyl-serine phosphorylation, positive regulation of peptidyl-cysteine S-nitrosylation, negative regulation of transcription by RNA polymerase II, positive regulation of DNA binding | ENSGALT00000025326 |
| LOX (lysyl oxidase) |
extracellular space | protein-lysine 6-oxidase activity, copper ion binding, collagen binding | peptidyl-lysine oxidation, collagen fibril organization | https://www.ncbi.nlm.nih. gov/gene/396474 |
| CLDN1 (claudin 1) | cell junction, bicellular tight junction, integral component of membrane, cytoplasm | structural molecule activity | bicellular tight junction assembly, cell adhesion, regulation of ion transport | ENSGALT00000077095 ENSGALT00000030190 |
| MUC2 (mucin 2) |
extracellular matrix, intracellular membrane-bounded organelle, supramolecular fiber | protein binding, virion binding, antigen binding | cholesterol homeostasis, intestinal cholesterol absorption, maintenance of gastrointestinal epithelium, negative regulation of cell growth, macrophage activation involved in immune response | https://www.ncbi.nlm.nih. gov/gene/414878 |
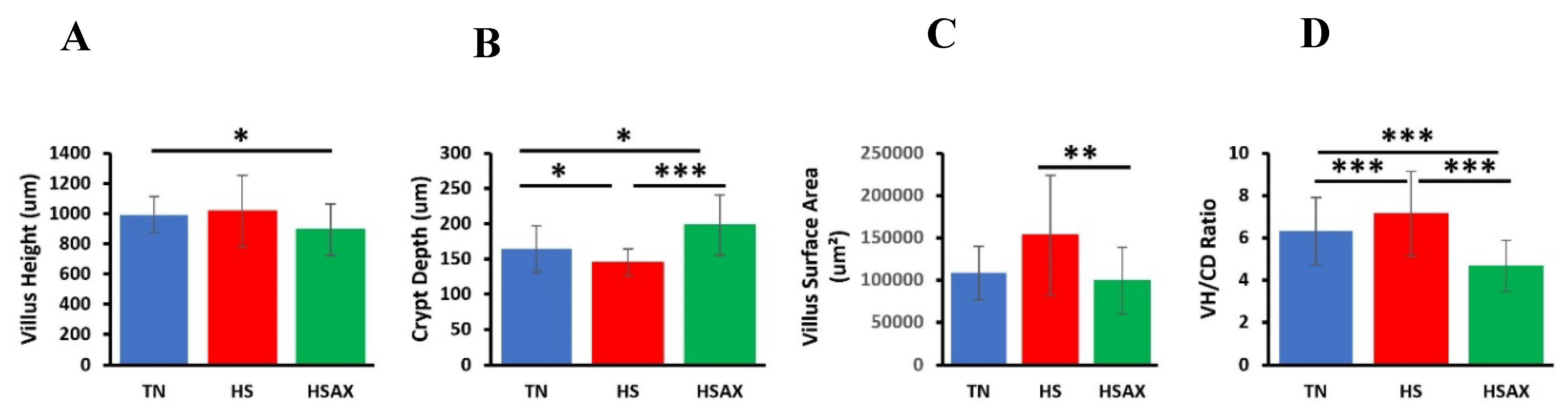
3.3. Ileum Histomorphology
The studies showed the impact of HS and AOX treatment on the ileum histomorphology. The villus height (VH) measurements showed a marginal significant difference comparing the TN vs HSAX (p=0.0552), where the TN measurements were longer. The crypt depth (CD) measurements were significant in all measurements, TN vs HS (p=3.77e-02), TN vs HSAX (p=2.63e-02), and HS vs HSAX (p=9.30e-07), where HSAX showed the highest measurements of all. The villus surface area (VSA) calculation, HS vs HSAX (p=0.0042) was the only significant group comparison, with HS significantly higher than HS. The overall VH:CD ratio resulted in significant difference between all groups, TN vs HSAX (p=3.59e-01), TN vs HSAX (p=4.17e-04), and HS vs HSAX (p=2.41e-07).
Figure 5.
Effect of chronic heat stress of Cob-500 broiler ileum histomorphology from hematoxylin and Eosin (H&E) stained images (scale bar = 200μm). Three groups were measured and compared: (A) TN, (B) HS, and (C) HSAX. The VH was measured from the tip of the villus to the junction of villus and crypt, where the TN was slightly longer than the HSAX. CD was defined from the villus base to the submucosa, where statistical differences occurred in all pairwise comparisons and particularly significant the HSAX vs HS. VSA determined as the villus base width plus the villus apical width divided by two in which case the comparison between HS vs HSAX was most significant.
Figure 5.
Effect of chronic heat stress of Cob-500 broiler ileum histomorphology from hematoxylin and Eosin (H&E) stained images (scale bar = 200μm). Three groups were measured and compared: (A) TN, (B) HS, and (C) HSAX. The VH was measured from the tip of the villus to the junction of villus and crypt, where the TN was slightly longer than the HSAX. CD was defined from the villus base to the submucosa, where statistical differences occurred in all pairwise comparisons and particularly significant the HSAX vs HS. VSA determined as the villus base width plus the villus apical width divided by two in which case the comparison between HS vs HSAX was most significant.
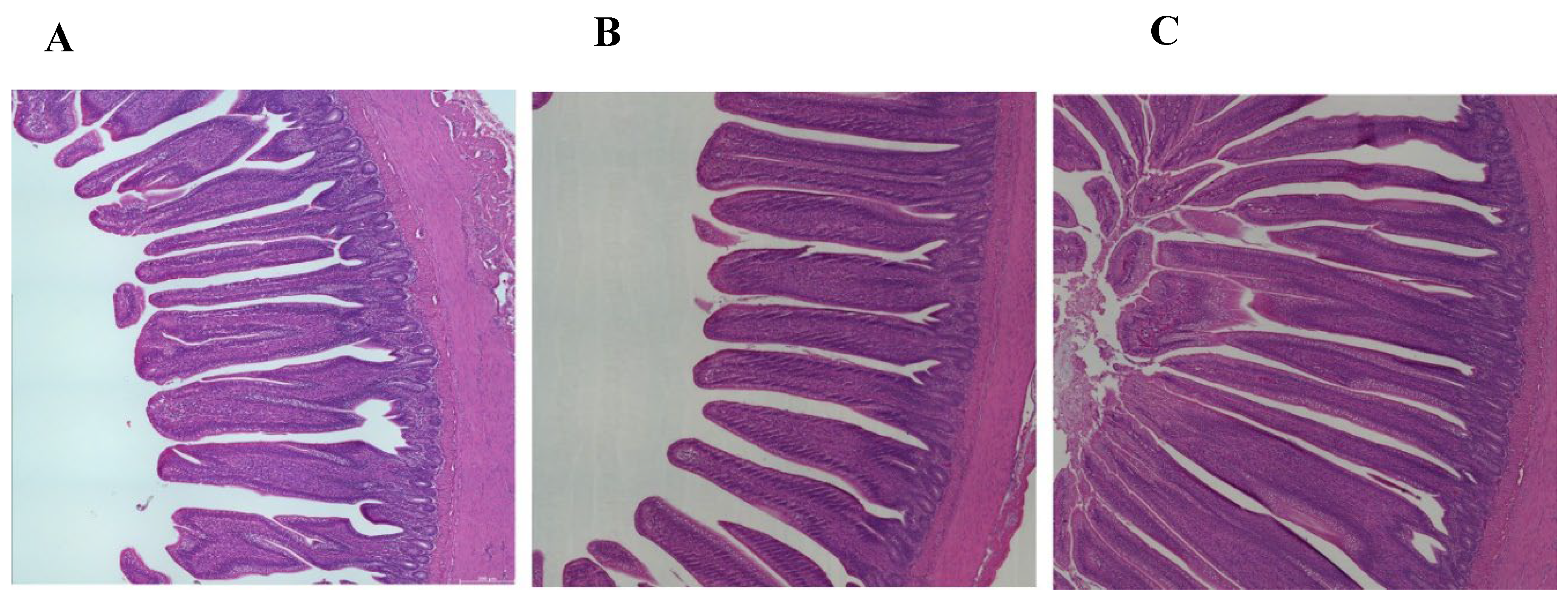
Table 6.
Ileum histomorphology and statistical significance of Cob-500 broilers villus and crypt measurements. Measurements for Villis Height (VH), Crypt Depth (CD), Villus Surface Area (VSA), and VH:CD Ratio were obtained in μm units. Three groups were compared: TN, HS, and HSAX. Statistical analysis based on the Kruskal-Wallis rank sum test, followed with the Dunn post hoc test for comparisons, and p-value adjusted with the Bonferroni method. * 0.05>p>0.01, ** 0.01>p>0.001.
Table 6.
Ileum histomorphology and statistical significance of Cob-500 broilers villus and crypt measurements. Measurements for Villis Height (VH), Crypt Depth (CD), Villus Surface Area (VSA), and VH:CD Ratio were obtained in μm units. Three groups were compared: TN, HS, and HSAX. Statistical analysis based on the Kruskal-Wallis rank sum test, followed with the Dunn post hoc test for comparisons, and p-value adjusted with the Bonferroni method. * 0.05>p>0.01, ** 0.01>p>0.001.
| Kruskal-Wallis |
Dunn/Bonferroni p-value |
||||||
| Measurements | TN | HS | HSAX | p-value | TN vs HS | TN vs HSAX | HS vs HSAX |
| VH (μm) | 992.79 ± 118.97 |
1061.89 ± 235.73 | 893.08 ± 170.00 | 0.0471 | 1 | 0.0552 | 0.2046 |
| CD (μm) |
164.28 ± 32.67 |
145.08 ± 19.09 |
197.87 ± 42.86 |
2.05e-06 | 3.77e-02 | 2.63e-02 | 9.30e-07 |
| VSA (μm) |
108602.30 ± 31508.84 |
152946.60 ± 70507.57 |
99497.84 ± 39296.00 |
0.0050 | 0.0995 | 0.8640 | 0.0042 |
| VH:CD (μm2) |
6.31 ± 1.59 |
7.13 ± 2.00 |
4.67 ± 1.21 |
2.39e-07 | 3.59e-01 | 4.17e-04 | 2.41e-07 |
Figure 6.
Effects of heat stress and astaxanthin treatment on Cob-500 broiler ileum tissue morphology. Three groups were compared: TN, HS, and HSAX. Kruskal-Wallis test and Dunn post hoc test for statistical significance represented: (A) VH, KW p=0.0471, TN vs HSAX (p=0.0552); (B) CD, KW p=2.05e-06, TN vs HS (p=3.77e-02), HS vs HSAX (p=9.30e-07); (C) VSA, KW p=0.0050, HS vs HSAX (p=0.0042); and (D) VH:CD, KW p=2.39e-07, TN vs HS (p=3.59e-01), TN vs HSAX (p=4.17e-04), HS vs HSAX (p=2.41e-07).
Figure 6.
Effects of heat stress and astaxanthin treatment on Cob-500 broiler ileum tissue morphology. Three groups were compared: TN, HS, and HSAX. Kruskal-Wallis test and Dunn post hoc test for statistical significance represented: (A) VH, KW p=0.0471, TN vs HSAX (p=0.0552); (B) CD, KW p=2.05e-06, TN vs HS (p=3.77e-02), HS vs HSAX (p=9.30e-07); (C) VSA, KW p=0.0050, HS vs HSAX (p=0.0042); and (D) VH:CD, KW p=2.39e-07, TN vs HS (p=3.59e-01), TN vs HSAX (p=4.17e-04), HS vs HSAX (p=2.41e-07).
4. Discussion
Results showed both treatments under HS experienced reduced growth performance in comparison to the TN group, while the group treated with AST showed a slightly lesser decline. Results further showed the AST treatment group was significantly upregulated in the cytoprotective gene expression for HSF2, SOD2, GPX3, and TXN, as well as the upregulation of epithelial integrity genes, LOX, CLDN1 and MUC2. Elucidating the molecular mechanisms underlying cellular responses to environmental stressors such as heat stress contributes to our understanding of how animals adapt and survive adverse conditions and paves the way for targeted therapeutic strategies aimed at mitigating harmful effects on poultry exposed to environmental challenges.
4.1. Growth Performance
Growth performance is anticipated to suffer under any stress condition, and in particular HS [40]. Although the growth performance indexes did not show any significant benefit from an AOX supplement, the health of the poultry was further elucidated though the gene expression and tissue histomorphology studies providing insights into mechanisms affecting the health and wellbeing of the broilers.
4.2. Gene Ontology Enrichment and Expression Analysis
4.2.1. Cytoprotective Capacity Genes
HSF2 is noted to be found in the nucleus and cytoplasm, where the molecular function of RNA polymerase II cis-regulatory sequence-specific in DNA-binding transcription factor activity occurs. HSF2 affects the cellular response through positive regulation of transcription form RNA polymerase II promoter thereby regulating the transcription of the DNA-template (Table 5) [20,21,22,36]. The results suggest AST treatment has a positive effect on the significant upregulation of HSF2 in the HSAX over the HS group.
SOD2 is located in the mitochondrion and functions as an oxidoreductase enzyme involving manganese ion binding in response to hydrogen peroxide negative regulation of OS and the removal of superoxide radicals (Table 5) [15,16,18,36]. The results support AST having a positive effect on AOX production to mitigate free radical damage from OS by the significant upregulation of HSAX over the HS group.
GPX3 is located in the extracellular space involved in selenium and identical protein binding in response to hydrogen peroxide catabolic process negative regulation of OS (Table 5) [15,16,18,36]. The study results indicate the endogenous capability of producing GPX3 in response to OS, however the application of AST provides a significant advantage in the upregulation of HSAX over the TN.
TXN is found in the nucleus, cytosol and extracellular region supporting protein disulfide oxidoreductase activity for cell redox homeostasis through the balancing of biomolecules positively and negatively affecting the regulation of transcription by RNA polymerase II and DNA binding (Table 5)[18,36]. The results are consistent with AST having a positive AOX effect in supporting cell redox homeostasis to reduce free radical damage from OS in the significant upregulation of HSAX over the HS group.
4.2.2. Epithelial Integrity Genes
LOX is found in the extracellular space responsible for collagen fibril organization and binding from peptidyl-lysine oxidase enzyme activity involving copper ion binding (Table 5) [25,26,36]. The results show AST has a positive correlation with promoting tissue integrity in the significant upregulation of the HSAX over the TN. group.
CLDN1 is an integral component of the cell membrane, particularly within the bicellular tight junction and provides structural biomolecular activity for the assembly, adhesion and regulation of ion transport (Table 5) [27,36]. The results of the study imply that AST has a positive effect in support of membrane integrity through the upregulation of the HSAX over the HS group.
MUC2 is located both intracellular and extracellular of the membrane as a supramolecular fiber for protein, virion and antigen binding for maintenance of the gastrointestinal epithelium through intestinal cholesterol homeostasis and macrophage activation involving the immune response (Table 5)[28,29,36]. The results support that AST may help maintain the integrity of the mucus barrier in the gastrointestinal tract for protection of the epithelial surfaces as indicated by the upregulation of the HSAX over the HS group.
4.3. Ileum Histomorphology
The ileum is the final section of the small intestine where enzyme molecules may adsorb and attach to the surface area, and the final absorption of nutrients occurs before exiting to the ceca. The wall of the ileum is made up of folds of villi, tiny projections that increases the surface area for adsorption and absorption that transport amino acids and glucose produced through digestion into the hepatic portal vein and the liver.
The villus height (VH) of the HSAX measured significantly less than the TN, while HS was higher, though not by a significant amount. This may be the result of physiological changes altering the epithelial cell and morphology, and may indicate a heat-induced stress and adaptive mechanism to preserve intestinal function and optimize nutrient absorption [19,23,37].
There were significant differences in the crypt depth (CD) which correlates to the intestinal stem cells and progenitor cells responsible for replenishing the epithelial lining. Crypts also play a crucial role in maintaining the mucosal barrier function helping to prevent entry of luminal pathogens, toxins and antigens. This research study showed a highly significant difference in the HSAX group in comparison to the HS group, but not as significant to the TN group. The statistical difference in crypt depth between the TN and HS indicates potential damage for epithelial regeneration, mucosal barrier function and immune regulation in the ileum.
The villus surface area (VSA) showed a significant difference where the HSAX calculated lower than the HS, indicating some adaptive mechanism enhanced by AOX supplementation to maintain efficient nutrient absorption. The higher measurements in the HS group may be the result of increased permeability of the intestinal epithelium, known as a “leaky gut” and increased susceptibility to negative health impacts for inflammation, immune dysregulation, digestive disorders, nutrient malabsorption and increased susceptibility to infections.
The overall VH:CD ratio can be used to assess structural integrity functional status of the ileum, providing valuable insights into the balance between epithelial regeneration and turnover. This study results show the HSAX group is significantly lower than both the TN and HS groups as a result of AOX treatment, indicating a substantive correlation in intestinal health.
5. Conclusions
In conclusion, our experimental findings indicate AST, a potent antioxidant, shows promise in mitigating heat-induced oxidative stress in the ileum, suggesting therapeutic potential. This research study integrates current knowledge, providing insights into the molecular mechanisms and potential interventions for protecting epithelial integrity by increasing cytoprotective capacity during thermal stress. Nonetheless, knowledge gaps remain in the understanding of the endogenous cellular mechanisms involved in cellular protection from heat-induced oxidative stress. Further research is needed to elucidate the mechanisms underlying the gene expression and observed histomorphology to assess the effects of heat-induced oxidative stress on intestinal physiology and function.
Supplementary Materials
Supplementary data is available in an open-source repository Github as referenced in the Data Availability Statement below.
Author Contributions
DLK conceived the study, collected samples, performed laboratory assays, analyzed the data, performed data preparation and figures and wrote the manuscript. YF, MN, HY and VSK conceived the study and performed analysis. YD conceived the study and contributed resources. All authors edited the manuscript and approved the final draft.
Funding
This project was supported by grants from the National Institute of Health (NIH) grants T32DK137523, U54GM138062, U54MD007601, P30GM114737, P20GM103466, P20GM139753, U54HG013243, National Science Foundation (NSF) 1745130, American Indian Science and Engineering Society, Advancing Agricultural Science Opportunities for Native Americans (AASONA), and University of Hawaii, Hilinehu Educational Leadership Advancement (S362A210110). The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH.
Institutional Review Board Statement
The study was conducted according to the guidelines and approved by the University of Hawaii Institutional Animal Care and Use Committee (IACUC) under protocol number 17-2605.
Informed Consent Statement
“Not applicable.”
Data Availability Statement
The original data presented in this study are openly available in Github at (https://github.com/sweetiek/Broiler_ileum_astaxanthin).
Acknowledgments
Special acknowledgments and sincere gratitude to Socorro Tauyan, Facility Manager of the Small Animal Facility, Magoon Research Station, Honolulu, Hawaii, for her support and oversight throughout the animal nutrition experiment, and coordination with the Animal Physiology Lab at the University of Hawaii, Human Nutrition, Food and Animal Sciences.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
Table A1. Common components of Haematococcus algae (Lorenz; Cyanotech 1999).
References
- Ricke, S. C. Improving gut health in poultry. 1st Edition 2019. Burleigh Dodds Science Publishing. University of Arkansas, USA. [CrossRef]
- Bailey, Richard A. “Gut Health in Poultry: The World within - Update | The ...” Gut Health in Poultry:the World within-Update, 1 Oct. 2019, 8:30am, thepoultrysite.com/articles/gut-health-in-poultry-the-world-within-1.
- Glendinning, L., Stewart, R. D., Pallen, M. J., Watson, K.A. “Assembly of Hundreds of Novel Bacterial Genomes from the Chicken Caecum.” Genome Biology, 2019. [CrossRef]
- Kers, Jannigje G.; et al. Host and Environmental Factors Affecting the Intestinal Microbiota in Chickens. Frontiers in Microbiology, vol. 9, 2018. [CrossRef]
- Carrasco, Juan M. Diaz; et al. Microbiota, Gut Health and Chicken Productivity: What Is the Connection? Microorganisms, vol. 7, no. 10, 2019, p. 374.. [CrossRef]
- Surai, P.F., and V.I. Fisinin. Vitagenes in Poultry Production: Part 3. Vitagene Concept Development. World’s Poultry Science Journal, vol. 72, no. 4, 1 Dec. 2016, pp. 793–804. [CrossRef]
- Farag, M. R., Alagawany, M. Physiological Alterations of Poultry to the High Environmental Temperature. Journal of Thermal Biology, vol. 76, 2018, pp. 101–106.. [CrossRef]
- Lara, L., Rostagno, M. Impact of Heat Stress on Poultry Production. Animals, vol. 3, no. 2, 24 Apr. 2013, pp. 356–369, www.ncbi.nlm.nih.gov/pmc/articles/PMC4494392/. [CrossRef]
- Collier, R. J., & Gebremedhin, K. G. Thermal biology of domestic animals. Annual Review of Animal Biosciences, 2015, 3, 513–532. [CrossRef]
- Sies, H. Oxidative stress: Oxidants and antioxidants. Experimental Physiology, 1997, 82, 291–295.
- Belhadj Slimen, I., Najar, T., Ghram, A., & Abdrrabba, M. Heat stress effects on livestock: Molecular, cellular and metabolic aspects, a review. Journal of Animal Physiology and Animal Nutrition, 2016, 100(3), 401–412. [CrossRef]
- Cano-Europa, E. et al. Regulation of the redox environment. Intech, 2015. https://www.intechopen.com/books/basic-principles-and-clinical-significance-of-oxidative-stress/regulation-of-the-redox-environment.
- Montzouris, K.C., Paraskeuas, V.V., Fegeros, K. Priming of intestinal cytoprotective genes and antioxidant capacity by dietary phytogenic inclusion in broilers. Anim Nutr. 2020 Sep;6(3):305-312. [CrossRef]
- Paraskeuas, V., Griela, E., Bouziotis, D., Fegeros, K., Antonissen, G. et al., Effects of Deoxynivalenol and Fumonisins on Broiler Gut Cytoprotective Capacity. Toxins (Basel). 2021 Oct 16;13(10):729. .pimd:34679022; PMCID: PMC8538483. [CrossRef]
- Tang, D., Wu, J., Jiao, H., Wang, X., Zhao, J. et al., The development of antioxidant system in the intestinal tract of broiler chickens. Poult Sci. 2019 Feb 1:98(2):664-678. . PMID: 30289502. [CrossRef]
- Surai F, Peter. “Antioxidant Systems in Poultry Biology: Superoxide Dismutase.” Journal of Animal Research and Nutrition, vol. 01, no. 01, 2016. [CrossRef]
- Surai, P.F., and V.I. Fisinin. Vitagenes in Poultry Production: Part 1. Technological and Environmental Stresses. World’s Poultry Science Journal, vol. 72, no. 4, 1 Dec. 2016, pp. 721–734. [CrossRef]
- Surai, P. F., & Fisinin, V. I. Vitagenes in poultry production: Part 3. Vitagene concept development. World’s Poultry Science Journal, 2016, 72(4), 793–804. [CrossRef]
- 19. Liu, Guanhui; et al. Effect of Chronic Cyclic Heat Stress on the Intestinal Morphology, Oxidative Status and Cecal Bacterial Communities in Broilers. Journal of Thermal Biology, vol. 91, July 2020, p. 102619. [CrossRef]
- Morimoto, R.I., Tissieres A., and Georgopoulos, C. The biology of heat shock proteins and molecular chaperones. Cold Spring Harbor, 1994: Cold Spring Harbor Laboratory Press.
- Pirkkala, Lila; et al. Roles of the Heat Shock Transcription Factors in Regulation of the Heat Shock Response and Beyond. The FASEB Journal, vol. 15, no. 7, May 2001, pp. 1118–1131. [CrossRef]
- Vabulas, R. M. et al. Protein folding in the cytoplasm and the heat shock response. Cold Spring Harbor Perspectives in Biology, 2010, 1-19.
- Javid, M. A., Abbas, G., Waqas, M. Y., Basit, M. A., Asif, M., Akhtar, M. S., Masood, S., Saleem, M. U., Qamar, S. H., & Kiani, F. A. Evaluation of comparative effect of feed additive of allium sativum and zingeber officinale on bird growth and histomorphometric characteristics of small intestine in broilers. Revista Brasileira de Ciencia Avicola, 2019, 21(2). [CrossRef]
- Awad, W. A., Ghareeb, K., Abdel-Raheem, S., & Böhm, J. Effects of dietary inclusion of probiotic and synbiotic on growth performance, organ weights, and intestinal histomorphology of broiler chickens. Poultry Science, 2009, 88(1), 49–55. [CrossRef]
- Hsiu Chung Ou; et al. Galectin-3 Aggravates Ox-LDL-Induced Endothelial Dysfunction through LOX-1 Mediated Signaling Pathway. Environmental Toxicology, vol. 34, no. 7, 9 Apr. 2019, pp. 825–835. [CrossRef]
- Laczko, Rozalia, and Katalin Csiszar. Lysyl Oxidase (LOX): Functional Contributions to Signaling Pathways. Biomolecules, vol. 10, no. 8, 22 July 2020, p. 1093. Accessed 22 Aug. 2020. [CrossRef]
- Santos, R. R., Awati, A., Roubos-van den Hil, P. J., van Kempen, T. A. T. G., Tersteeg-Zijderveld, M. H. G. et al. Effects of feed additive blend on broilers challenged with heat stress. Avian Pathology, 2019, 48:6, 582-601. [CrossRef]
- Apalowo, O.O., Minor, R.C., Adetunji, A.O., Ekunseitan, D.A., Fasina, Y.O. Effect of Ginger Root Extract on Intestinal Oxidative Status and Mucosal Morphometrics in Broiler Chickens. Animals 2024, 14, 1084. [CrossRef]
- Akagi, R. Role of Heme Oxygenase in Gastrointestinal. Epithelial Cells. Antioxidants, 2022. [CrossRef]
- Britton, G., Liaaen-Jensen, S., & Pfander, H. Carotenoids. Isolation and analysis. Basel: Birkhäuser Verlag, 1995.
- Shahidi, F., & Barrow, C. J. Marine nutraceuticals and functional foods. Boca Raton: CRC Press, 2008.
- Astaxanthin. (n.d.). Retrieved from https://pubchem.ncbi.nlm.nih.gov/compound/Astaxanthin. Accessed 02 Apr 2024.
- Heaney, R. P. Factors Influencing the Measurement of Bioavailability, Taking Calcium as a Model. The Journal of Nutrition, 2001,131(4). [CrossRef]
- Mularczyk, M., Michalak, I., Marycz, A. Astaxanthin and other Nutrients from Haematococcus pluvialis-Multifunctional Applications. Mar Drugs. 2020 Sep 7;18(9):459. PMID: 32906619; PMCID: PMC7551667. [CrossRef]
- Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt method. Methods, 2001, 25: 402-408.
- Ensembl.org. https://uswest.ensembl.org/Gallus_gallus/Info/Index Accessed 04 Apr 2024.
- Iji, P.A., Saki, A., Tivey, D.R. Body and intestinal growth of broiler chicks on a commercial starter diet. 1. Intestinal weight and mucosal development. Br Poult Sci. 2001 Sep;42(4):505-13. PMID: 11572627. [CrossRef]
- Schindelin, J., Arganda-Carreras, I., Frise, E.; et al. Fiji an open-source platform for biological-image analysis. Nature Methods, 2019, 9(7): 676-682. PMID 22743772. [CrossRef]
- R Core Team (2023) https://www.R-project.org/ Accessed 04 Apr 2024.
- Akbarian, A.; et al. Association between Heat Stress and Oxidative Stress in Poultry; Mitochondrial Dysfunction and Dietary Interventions with Phytochemicals. Journal of Animal Science and Biotechnology, vol. 7, no. 1, 28 June 2016. [CrossRef]
Figure 1.
Growth performance indicators of Cob-500 broilers. Three groups were compared: TN, HS, and HSAX. Kruskal-Wallis test and Dunn post hoc test for statistical significance represented: (A) BW(g), KW p=0.0047, TN vs HS (p=0.0095) and TN vs HSAX (p=0.0278); (B) ADFI(g), KW p=0.0038, TN vs HS (p=0.0047) and TN vs HSAX (p=0.0500); (C) ADGR(g), KW p=0.0047, TN vs HS (p=0.0095) and TN vs HSAX (p=0.0278); and (D) FCR, KW p=0.0049, TN vs HS (p=0.0226) and TN vs HSAX (p=0.0119).
Figure 1.
Growth performance indicators of Cob-500 broilers. Three groups were compared: TN, HS, and HSAX. Kruskal-Wallis test and Dunn post hoc test for statistical significance represented: (A) BW(g), KW p=0.0047, TN vs HS (p=0.0095) and TN vs HSAX (p=0.0278); (B) ADFI(g), KW p=0.0038, TN vs HS (p=0.0047) and TN vs HSAX (p=0.0500); (C) ADGR(g), KW p=0.0047, TN vs HS (p=0.0095) and TN vs HSAX (p=0.0278); and (D) FCR, KW p=0.0049, TN vs HS (p=0.0226) and TN vs HSAX (p=0.0119).
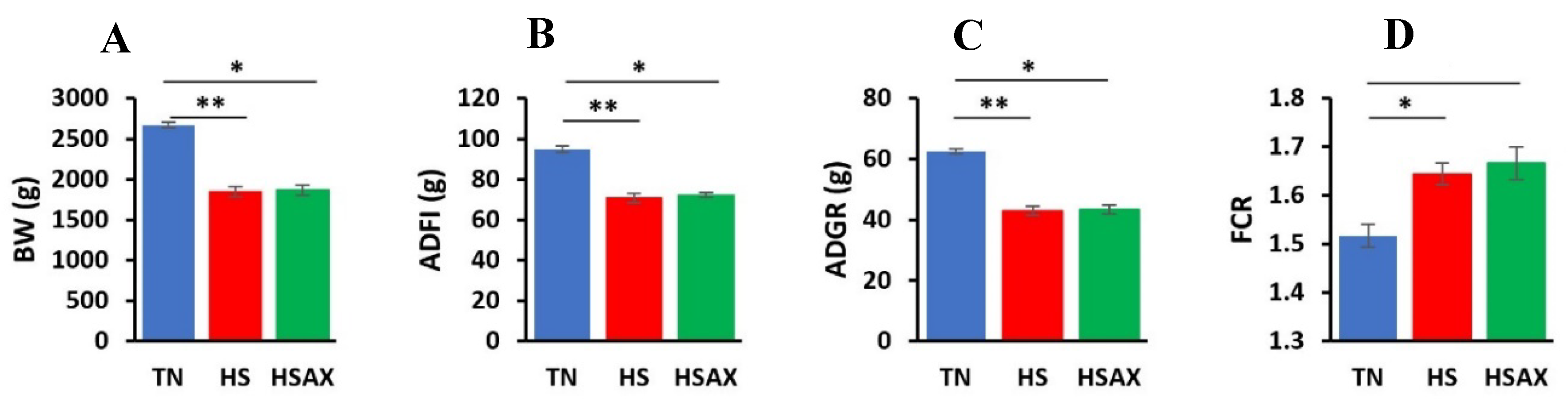
Table 1.
Basal diet ingredients and calculated analysis of broiler diets used in the mixture design for thermal neutral and heat stress including astaxanthin supplement.
Table 1.
Basal diet ingredients and calculated analysis of broiler diets used in the mixture design for thermal neutral and heat stress including astaxanthin supplement.
| Ingredients (%) | Starter | Finisher |
| Corn | 54.86 | 63.14 |
| Soybean Meal | 39.99 | 30.09 |
| Soybean oil | 2.00 | 4.50 |
| Limestone | 1.27 | 0.85 |
| Monocalcium phosphate | 0.75 | 0.50 |
| Lysine (%) | 0.23 | 0.18 |
| Methionine (%) | 0.14 | 0.12 |
| Threonine (%) | 0.20 | 0.16 |
| Sodium chloride (%) | 0.43 | 0.35 |
| Sodium bicarbonate | 0.12 | 0.10 |
| Vitamin-mineral premix | 0.50 | 0.50 |
| Astaxanthin supplement¹ | 0.01 | 0.01 |
| Total | ||
| Calculated analysis | ||
| ME (kcal/kg) | 2909 | 3203 |
| CP (%) | 22.09 | 18.07 |
| Calcium (%) | 0.75 | 0.52 |
| Total Phosphorus (%) | 0.57 | 0.47 |
| digPhosphorous (%) | 0.30 | 0.23 |
| Lysine (%) | 1.39 | 1.10 |
| dig Lysine (%) | 1.25 | 0.99 |
| Methionine (%) | 0.48 | 0.41 |
| dig Methionine (%) | 0.45 | 0.39 |
| Cysteine (%) | 0.43 | 0.38 |
| Threonine (%) | 1.03 | 0.85 |
| dig Threonine (%) | 0.85 | 0.69 |
| Tryptophan (%) | 0.33 | 0.26 |
| Methionine + cysteine (%) | 0.91 | 0.80 |
| Arginine (%) | 1.61 | 1.31 |
| Valine (%) | 1.22 | 1.03 |
| Isoleucine (%) | 0.93 | 0.76 |
| Leucine (%) | 1.89 | 1.63 |
| Neutral detergent fiber | 9.13 | 8.78 |
| Crude fiber | 3.97 | 3.46 |
| Sodium | 0.22 | 0.18 |
| Chloride | 0.30 | 0.25 |
| Choline (mg/kg) | 1419 | 1200 |
| Astaxanthin (mg/kg) | --- | 1.33 |
| ¹Astaxanthin was mixed with the soybean oil and supplemented in the diet during feed mixing. | ||
Table 2.
Gallus gallus oligonucleotide primers used for real-time RT-PCR analysis.
| Gene | NCBI Accession No. | Primer set (5’-3’) |
|---|---|---|
| GAPDH | X00182 | F:5' -CGCAAGGGCTAGGACGG |
| R:3' -GCGCTCTTGCGGGTACC | ||
| β-ACTIN | NM_205518.1 | F:5' -GAGAAATTGTGCGTGACATCA |
| R:3' -CCTGAACCTCTCATTGCCA | ||
| TBP | NM_205103.1 | F:5' -TAGCCCGATGATGCCGTAT |
| R:3' -GTTCCCTGTGTCGCTTGC | ||
| HSF1 | NM_001305256.1 | F:5’-AAGGAGGTGCTCCCAAAGTA |
| R:3’-TTCTTTATGCTGGACACGCTG | ||
| HSF2 | NM_001167764.2 | F:5’ -TCTTTTTACAAGCTCCGTGC |
| R:3’ -TCCCTTTGTCTCCATTTTGGT | ||
| HSF3 | NM_001305041.1 | F:5’ -TTCAGCGATGTGTTTAACCCT |
| R:3’ -GGAGGTCTTTTGGATCCTCT | ||
| HSP90AA1 | NM_001109785.1 | F:5’ -GATAACGGTGAACCTTTGGG |
| R:3’ -GGGTAGCCAATGAACTGAGA | ||
| HIF1A | XM_015287264.4 | F:5’ - GTCACCGACAAGAAGAGGAT |
| R:3’ -GTCTCTAGCTCACCAGCATC | ||
| SOD1 | NM_205064.1 | F:5' -CAACACAAATGGGTGTACCA |
| R:3' -CTCCCTTTGCAGTCACATTG | ||
| SOD2 | NM_204211.1 | F:5' -CCTTCGCAAACTTCAAGGAG |
| R:3' -AGCAATGGAATGAGACCTGT | ||
| ROS-1 | NM_205257.2 | F:5' -AAACTACAGCTGGTGTTCCC |
| R:3' -CTAAGTTCTCGGCCTTCCAT | ||
| GPX1 | NM_001277853.2 | F:5' -AATTCGGGCACCAGGAGAA |
| R:3' -CTCGAACATGGTGAAGTTGG | ||
| GPX3 | NM_001163232.3 | F:5' -AATTCGGGCACCAGGAGAA |
| R:3' -CTCGAACATGGTGAAGTTGG | ||
| HMOX1 | NM_205344.2 | F:5' -AATCGCATGAAAACAGTCCA |
| R:3' -CACATGGCAAATAAGCCCAC | ||
| CAT | NM_001031215.2 | F:5' -TGGCCAATTATCAGAGGGAC |
| R:3' -CTCGCACCTGAGACACATTA | ||
| CASP3 | NM_204725.1 | F:5' -GGTGGAGGTGGAGGAGC |
| R:3' -TGAGCGTGGTCCATCTTTTA | ||
| GSTT1 | NM_205365.1 | F:5' -AACATCCCGTTCGAGTTCAA |
| R:3' -CACTATTTGATGGCCCTGTG | ||
| TXN | NM_205453.1 | F:5' -GGCAATCTGGCTGATTTTGA |
| R:3' -ACCATGTGGCAGAGAAATCA | ||
| TXN2 | NM_001031410.1 | F:5' -CGATTGAGTACGAGGTGTCA |
| R:3' -CAGAAGAAAACCCCACAAACTT | ||
| PRDX1 | NM_001271932.1 | F:5' -GGTATTGCATACAGGGGTCT |
| R:3' -AGGGTCTCATCAACAGAACG | ||
| LOX | NM_205481.2 | F:5' -TACTTCCAGTACGGTCTGCC |
| R:3' -CTCTAACATCCGCCCGATAA | ||
| MYB | NM_205306.1 | F:5' -AGCATATACAGCAGCGATGA |
| R:3' -TTTCTCATCCTCTTCACGGG | ||
| CLDN1 | NM_001013611.2 | F:5' -CATCACTTCTCCTTCGTCAGC |
| R:3' -GCACAAAGATCTCCCAGGTC | ||
| OCLN | NM_205128.1 | F:5' -CTACAAGCAGGAGTTCGACA |
| R:3' -CTCTGCCACATCCTGGTATT | ||
| MUC2 | NM_001318434.1 | F:5' -TACAGGGAGTTCTCTGTCCA |
| R:3' -TAGGGTGTCTTGACAATCCG | ||
| CDH1 | NM_001039258.2 | F:5' -GAACTTCATCGACGAGAACC |
| R:3' -CGTTGAGGTAGTCGTAGTCC |
Table 3.
Growth performance indicators of Cob-500 broilers for body weight, adjusted daily feed intake, adjusted daily growth rate, and feed conversion. Three groups were compared: TN, HS, and HSAX. Statistical analysis was based on the Kruskal-Wallis rank sum test, followed with the Dunn post hoc test for comparisons, and p-value adjusted with the Bonferroni method. * 0.05>p>0.01, ** 0.01>p>0.001.
Table 3.
Growth performance indicators of Cob-500 broilers for body weight, adjusted daily feed intake, adjusted daily growth rate, and feed conversion. Three groups were compared: TN, HS, and HSAX. Statistical analysis was based on the Kruskal-Wallis rank sum test, followed with the Dunn post hoc test for comparisons, and p-value adjusted with the Bonferroni method. * 0.05>p>0.01, ** 0.01>p>0.001.
| Kruskal-Wallis |
Dunn/Bonferroni p-value |
||||||
| Measurements | TN | HS | HSAX | p-value | TN vs HS | TN vs HSAX | HS vs HSAX |
| BW (g) | 2673.68 ± 35.71 | 1848.85 ± 61.56 | 1867.83 ± 60.82 | 0.0047 | 0.0095 | 0.0278 | ns |
| ADFI (g) | 94.98 ± 1.45 |
70.68 ± 72.24 |
72.24 ± 1.14 |
0.0038 | 0.0047 | 0.0500 | ns |
| ADGR (g) | 62.65 ± 0.86 |
42.99 ± 1.46 |
43.45 ± 1.44 |
0.0047 | 0.0095 | 0.0278 | ns |
| FCR | 1.52 ± 0.02 |
1.64 ± 0.02 |
1.67 ± 0.03 |
0.0049 | 0.0226 | 0.0119 | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



