
Submitted:
30 May 2024
Posted:
31 May 2024
You are already at the latest version
Abstract
The emergence of antimicrobial resistance have represented a serious threats for public health and infections due to multidrug-resistant (MDR) microorganisms represent one of the most important causes of death worldwide. The renew of old antimicrobials, such as colistin, have been proposed as valuable therapeutic alternative to the emergence of the MDR microorganisms. Although colistin is well known to presents several adverse toxic effect, its usage in clinical practice have been reconsidered due to the broad spectrum of activity to gram-negative (GN) bacteria and to the important role of “last resort” agent against MDR-GN. Despite the revolutionary prospective of treatment of this old antimicrobial molecule, many questions remain open regarding the emergence of novel phenotypic traits of resistance and the optimal usage of the colistin in clinical practice. In the last years, several forward steps have been done in the understanding of resistance determinants, clinical usage and pharmacological dosage of this molecule, however, different points regarding the role of colistin in clinical practice and the optimal pharmacokinetic/pharmacodynamic targets are not well defined yet. In this review, we summarize the mode of action, the emerging resistance determinants and its optimal administration in the treatment of difficult-to-treat infections due to MDR Gram-negative bacteria.
Keywords:
lipopeptide
; antimicrobials
; antimicrobial resistance
1. Introduction
Antibiotic resistance represents a serious public health and it is associated to to million of deaths annually [1]. Since the discovery of first antimicrobial molecules, the emergence of novel traits of resistance to antimicrobials have been observed concomitantly [2]. It’s well known that antimicrobial resistance have been associated with their misuse and overuse in different field of applications (humans, animals and plants). Indeed, the presence of antimicrobial rich environments create a favourable conditions that allow the selection of resistant subpopulations in opposition to sensitive microorganisms [3].
With the diffusion and rapid increase of antimicrobial-resistance, the development of microorganisms resistant to multiple antimicrobial classes of compounds have been observed subsequently [4]. The emergence of multi-drug resistant (MDR) microorganisms posed different limitations to the clinicians by reducing the available antimicrobial armamentarium. In the last years, the diffusion of MDR strains have been considered an urgent threat especially among gram-negative bacteria that requires a prompted response. To overcome these limitations, several strategies have been adopted including new schemes of treatment by combining antimicrobial molecules with no activity alone and the development of novel antimicrobial molecules [5,6]. At the same time, the revival of older antibiotics considered as last resort drug have posed new prospective in treatment of difficult-to-treat (DTR) infections due to MDR strains [5].
Colistin, also known as polymyxin E, is an old antimicrobial molecule that it was discovered in the middle of 19th century in Japan from a culture of Paenibacillus polymyxa subspecies [7]. Colistin is a cyclic oligopeptides antimicrobials belonging to the class of polycationic antibiotic and it’s active against most Gram-negative bacteria by binding to the lipopolysaccharide (LPS) of the outer cell membranes by electrostatic interaction. The linkage between colistin and outer membrane create a disorganization of the outer membrane structure thus resulting in an alteration of the outer membrane and consequently intracellular contents release and bacterial death [7].
In the last years, renew of older antibiotics such as colistin have created new prospective in treatment of DTR infections [6,8]. However, the emergence of new traits of resistance to this drug and the adverse toxic effects to mammalian cells have mitigated its use in clinical practice [6,7]
In this review we discuss the principle of the mode of action, the emerging traits related to the resistance and the use of colistin in clinical practice from a pharmacological and clinical point of views.
2. Mechanisms of Action, Antibacterial Activity and Adverse Effects
2.1. Structure and Mode of Action
Colistin is an amphiphilic lipopeptide antibiotic discovered in 1947 by Koyama [9,10], produced by Paenibacillus polymyxa subspecies colistinus. Colistin, also called polymixyn E, is a member of polymyxin family of antibiotics. Since 1952, the first formulation in clinical use is a solution for intravenous administration and shown its bactericidal function against many Gram-negative bacteria, but not against Gram-positive, anaerobic bacteria or mycoplasmas. Due to potent antibacterial activity against Gram-negative bacteria colistin was initially considered a “miraculous molecule”. However, since 1970’s the use in clinical practice was mitigated due to severe its severe adverse effects [11]. The original molecule has been modified to reduce principally the nephrotoxicity effect and actually, two forms of colistin are clinically available for human treatment: colistin sulfate and colistin methanesulfonate, also called colistimethate sodium. Differences between these compounds are related to their use and toxicity. In particular, colistin sulfate is an active compound administered topically and orally, while colistimethate, is used in formulations administered by parenteral and nebulization routes.
Colistin’s basic structure consists of a core region formed by a hydrophobic portion, a cyclic heptapeptide linked by a tripeptide bridge, to a hydrophilic part, a fatty acid. The colistin molecule is positively charged due to the presence of five diaminobutyric acid residues linked to the core [12]. The prodrug form differs from this structure for the presence of methan-sulfonates linked to the diaminobutyric acids (Figure 1) [13].
It is noteworthy that the five diaminobutyric acid residues, which confer the positive charge to the molecule, play a determining role in the drug’s antibacterial effect, generally described with the Shai-Matsuzaki-Huang (SMH) model [14,15,16].
Colistin acts by competition and displacement of Ca2+ and Mg2+ from the negatively charged sulfate portion of the lipid A in the lipopolysaccharide molecule (LPS) of Gram-negative bacteria. This ionic dislocation by colistin seems necessary for forming pore-like structures [17,18,19]. The loss of ions binding and their substitution with colistin molecules alters the tertiary structure of LPS, creating the possibility for colistin itself to insert its own portion of fatty acids into the membrane, definitively compromising the permeability of the outer membrane. In addition, colistin acyl fat inserted in the bilayer alters the inner membrane stability, leading to bacterial membrane disruption and a bactericidal effect [20]. Colistin also plays a key role in preventing endotoxin-induced shock through its binding to lipide A portion [21]. This drug acts both at the surface and intracellular levels, in particular, altering vesicle-vesicle contact of bacterial cells. In brief, colistin crosses the membrane and causes the fusion of the inner leaflet of the outer membrane and the outer leaflet of the cytoplasmic membrane, disrupting the cytoplasmic bilayer, altering the osmotic balance and leading to cell death [22,23]. The antibacterial action of colistin was also reported at the molecular level, where it can induce oxidative stress and, consequently, DNA, protein, and lipid damage in bacteria through ROS production and could inhibit essential enzymes involved in the respiratory chain, such as the NADH-quinone oxidoreductase [24], leading to cell death.
2.2. Adverse Effects
Colistin treatment was dismissed in clinical use principally due to its nephrotoxicity effect, which is lower for the prodrug form. Adverse effects were principally due to its re-absorption by proximal tubule cells through an endocytotic process, mediated by megalin, and through a facilitative transport by two transporters located in the apical cell membrane, the human peptide transporter 2 (PEPT2) and the carnitine/organic cation transporter 2 (OCTN2) [25,26]. Intracellular accumulation of colistin induces mitochondrial and endoplasmic reticulum stress with consequent toxic cellular effects [27]. This mechanism leads to cellular lysis and acute tubular necrosis [28,29]. The incidence of colistin-induced acute kidney injury varies between 12.7 and 70% in intensive cure units patients [30,31,32,33]. A recent study by Kilic and colleagues demonstrated that the nephrotoxicity effect depends proportionally on the duration of treatment and is related to older patients [28,34].
Due to the high lipid content of neuronal cells, colistin could also exert its action in these cells, and some patients (with an incidence of about 7% [34]) experienced neurological adverse effects, such as paresthesia, seizures, confusion, ataxia, and visual disturbances [35]. The mechanism by which colistin induces these effects is a non-competitive presynaptic myoneuronal blockade of acetylcholine releasee [36]. Adverse effects on neuronal cells could be reverted by discontinuing the therapy.
2.3. In Vitro Antimicrobial Activity
In vitro activity of colistin was tested with success on Acinetobacter baumannii, a large part of Enterobacteriaceae and Pseudomonas aeruginosa [37]. In particular, for 106 non-duplicate isolates of A. baumannii was reported a minimum inhibiting concentration of 0.5 ug/mL for MIC50 and of 1.0 ug/mL for MIC90, in monotherapy [37]
Walkty et al. [38] analyzed the colistin antibacterial activity on 3,480 isolates of Gram-negative bacilli from patients recruited during 2 years in 12 hospitals in Canada (CANWARD Study). In this study authors reported a MIC90 value ≤2 μg/ml against a several clinically relevant gram-negative bacilli, such as Escherichia coli (1,732 isolates), Klebsiella spp. (515 isolates), Enterobacter spp., A. baumannii, and P. aeruginosa (561 isolates), including all 76 MDR P. aeruginosa isolates tested in CANWARD Study.
A cross-sectional and descriptive study conducted on 52 MDR P. aeruginosa isolates, collected from urine, pus specimens and respiratory tract, reported a MIC50 value of 1.0μg/mL and a MIC90 of 3.0μg/mL [39].
in the last years, several studies reported the activity colistin in combination other antimicrobial molecules. In particular, colistin, in combination with meropenem or tigecycline shown synergistic activity against colistin-resistant KPC-producing K. pneumoniae [40]. Kheshti and coworkers [41] on a study based on an in vitro checkerboard assay, reported a good synergistic activity of colistin treatment in combination with ciprofloxacin, levofloxacin (5%, the lowest level), imipenem, meropenem, ampicillin–sulbactan molecules and higher synergism in combination with rifampin (55%) tested on 20 isolated of A. baumanii.
A recent study [42], conducted on 219 K. pneumoniae isolates demonstrated the high synergy of minocycline and colistin on colistin-resistant and minocycline-intermediate or - resistant K. pneumoniae. This drug combination acts by disrupting the outer membrane (by colistin) without affecting the cytoplasmic membrane, allowing the entrance and accumulation at intracellular level of minocycline.
2.4. Antimcirobial Susceptibility Testing
The chemical structure of colistin and its cationic charge, make difficult the use of classical susceptibility test, like E-tests and disc diffusion. To overcome this limitation and to provide a pharmacological alternative to the numerous multi-resistant bacterial species, classical diagnostic protocols have been modified allowing the measurement of colistin susceptibility. Broth microdilution, the gold standard for colistin susceptibility test, is modified using a cation-adjusted Muller-Hinton broth without adding surfactant [43,44]. Another method approved by CLSI only for Enterobactarales and Pseudomonas spp, is a broth disc elution modified by Simner and colleagues [45]. The test, renamed as Colistin Broth Disc Elution (CBDE), is easier than the Broth microdilution and is based on analysis of the efficacy of a graded concentration of colistin (of 1,2,4 ug/mL) obtained from colistin disc elution in 10 mL of cation adjusted Muller-Hinton Broth, tested on a 0,5 Mc Farland of bacteria. EUCAST colistin breakpoints table, version 14.0 reports the following cut-off value for the detection of phenotypic resistance: MIC of 2 mg/L for Enterobacterales and for Acinetobacter spp., and MIC of 4 mg/L for P. aeruginosa.
3. Mechanisms of Colistin Resistance
A variety of mechanisms may be involved in the acquisition of colistin resistance in Gram-negative bacteria, and they can be summarized into four groups: modification of LPS structure by chromosomal mutations (i), modification of LPS structure by acquisition of plasmids (ii), loss of LPS structure (iii), overexpression of efflux pumps (iiii).
3.1. Modification of LPS Structure by Chromosomal Mutations
Reduction of negative charge of lipid A of LPS lead to loss of electrostatic interaction with colistin and consequently to resistance [46]. Many genes and operons are involved in LPS modifications: 1. pmrC and pmrE genes and the pmrHFIJKLM operon, which promote addition of phosphoethanolamine (PEtn) and/or 4-amino-4-deoxy-L-arabinose (L-Ara4N) to lipidA. 2. regulatory two-component systems such as PmrAB, PhoPQ, and crrAB, 3. mgrB negative regulator gene.
Addition of L-Ara4N and/or PEtn to lipidA changes the negative charge of the cell membrane by neutralizing the negatively charged phospholipids [47,48,49,50]. In detail, the addition of PEtn to the 1′- or 4′-phosphate group of lipid A is carried out by PmrC, a putative membrane protein with phosphoethanolamine transferase activity encoded by pmrABC operon [11,51,52,53,54]. The synthesis of L-Ara4N from uridine diphosphate glucuronic and its addition to lipid A is promoted by pmrHIJKLM operon (also called arnBCADTEF) and PmrE activity [51]. PmrB is a cytoplasmic membrane-bound which activates PmrA by phosphorylation, and PmrA in turns activates regulation of the pmrABC and pmrHFIJKLM operons and the pmrE gene. Subsequently, these operons and genes lead to LPS modification by adding PEtn and L-Ara4N to lipid A [55]. Although the L-Ara4N modification of LPS has been described as a common mechanism of colistin resistance among Gram-negative bacteria (Klebsiella pneumoniae, Escherichia coli, Salmonella enterica, and Pseudomonas aeruginosa), it does not occur in Acinetobacter baumannii because lacks all the genes required for L-Ara4N biosynthesis [51]. Alternatively, addition of galactosamine to the 1′-phosphate position of lipid A, following activation of the sensor kinase PmrB, has been associated with moderate levels of colistin resistance in A. baumannii [52].
Mutation of pmrA/pmrB results in upregulation of the pmrABC and pmrFHIJKLM operons and pmrE gene which lead to PEtN modification of lipid A, and in turn, results in colistin resistance. Several mutations have been reported in many Gram-negative bacteria, such as Salmonella enterica [56,57], K. pneumoniae [58,59,60], A. baumannii [61,62,63], P. aeruginosa [64,65], and E. coli [57,66,67].
Trascription of pmrFHIJKLM operon is also activated by PhoPQ regulatory two-component system. PhoQ is a sensor kinase that promote espression of the regulator protein PhoQ, which promote pmrFHIJKLM operon transcription via phosphorylation. Furthermore, PhoP indirectly activates pmrA through the PmrD connector protein, which subsequently activates the transcription of the pmrHFIJKLM operon. This then leads to synthesizes and transfer of PEtn to lipid A [11,49,50]. Mutation of the phoP/Q genes that led to acquired colistin resistance has been identified in K. pneumoniae and E. coli [68,69,70]. Higher polymyxin MICs have been observed in PhoQ-deficient P. aeruginosa mutants when additional alterations affected other regulatory two-component systems (CprRS and ColRS) [71].
More evidence has accumulated on the role played by mgrB, a gene encoding a small regulatory transmembrane protein, MgrB, that exerts negative feedback on the kinase activity of PhoQ [72]. Inactivation of mgrB leads to the ‘activation of a phosphorylation cascade involving at chain PhoQ, PhoP, PmrD and/or PmrAB and finally triggering the expression of the pmrHIJKLM operon resulting in LPS modification. Mutations of mgrB including points mutations, deletion, nonsense, and insertion sequences (IS5-like, IS1F, ISKpn14, ISKpn13, IS10R) represents the most common mechanism of colistin resistance in clinical K. pneumoniae isolates [69,73,74,75,76,77]. The wide range of resistance level showed by Gram-negative strains harbouring mutations in the genes pmrAB, phoPQ, or mgrB suggests a role for other genetic loci. Mutations in the CrrAB two-component system has been associated with increased level of colistin resistance in strains of K. pneumoniae [74,78]. Mutation/inactivation of the crrB gene led to activation of the pmrHFIJKLM operon and the pmrC and pmrE genes through overexpression of the pmrAB operon [77,78]. Furthermore, various of PEtn-coding genes, such as eptA (pmrC), eptB (pagC), and eptC (cptA), are able to add PEtn to LPS and can be involved in colistin resistance [79]. Overespression of eptA has been associated with colistin resistance in A. baumannii [61,80]. Gerson et al. showed that mutations in the eptA gene (R127L and ISAba1 insertion) was associated with overexpression of EptA and colistin resistance in A. baumannii [61].
3.2. Loss of LPS Structure
The complete loss of lipid A or LPS core leading to colistin resistance has been observed in A. baumannii. Analysis of laboratory-induced colistin-resistant A. baumannii showed that high level of resistance to colistin was caused by the inactivation of LPS biosynthesis genes lpxA, lpxC, lpxD and lpsB [81]. Various nucleotide substitutions, deletions, and insertions that cause frameshifts or result in truncated proteins have been reported from in vitro mutants and clinical isolates [81,82,83,84]. Moreover, disruption of lpxC and lpxD by insertion of IS elements, was described in colistin-resistant A. baumannii isolates [81,82,83,84,85,86]. Although LPS loss is an effective mechanism of colistin resistance, it has significant fitness costs and this explains why such mutants are rarely encountered in the clinical setting [87].
3.3. Plasmid-Mediated Colistin Resistance
Since the first report of the mcr gene encoding for phosphoethanolamine transferase (mcr-1) in E. coli in China in 2015 [88], several reports worldwide have demonstrated the presence of mcr-1 and additional 9 families (mcr-2 to mcr-10) with more than 100 overall variants in different Gram-negative species distributed worldwide [11,49,50,51,52,53,89,90,91,92,93,94]. MCR is a member of the PETN enzyme family, and its activity results in the modification of lipid A by PETN addition. The enzyme has a domain inserted in the inner membrane and a periplasmic C-terminal sulfatase catalytic domain.
In 2018, Partridge et al. proposed a nomenclature for mcr genes. Several variants have been identified, especially for MCR-3 and followed by MCR-1 [95]. MCR-1 and MCR-2 share 81% identity at the amino acid sequence level. Sequence identity suggests that these two variants originated from Moraxella spp [89], as mcr-3, mcr-4 and mcr-7 from Aeromonas spp. and Shewanella frigidimarina, respectively[90,91,92,93]
The mcr-1 variant can be connected to various types of plasmids, including IncHI2, IncI2, IncX4, IncP, IncX, and IncFIP. The mcr-2 gene was detected on an IncX4 plasmid. The presence of insertion sequence (ISApl1, IS1595) on the genetic environment of mcr genes explains the possibility to integrate on bacterial chromosomes [94].
Plasmid-mediated colistin resistance represents the mechanism of greatest concern because of the ease of intra- and inter-species spread. Despite most of MCR-harboring microorganisms belong to the Enterobacterales order, such as E.coli, Salmonella spp., and K. pneumoniae, several reports showed the presence of mcr-genes in non-fermenting Gram-negative species such as P. aeruginosa and A. baumannii complex [96,97,98,99,100,101,102,103,104,105,106,107,108,109,110]. The mcr-1 gene is the most commonly detected in P. aeruginosa both in clinical [96,97,98,99] and animal setting [100,101,102,103], and followed by mcr-5 [104,105]. In A. baumannii complex, the mcr-1 and mcr-4.3 are the major variants observed in clinical isolates from Asia and Europe [98,99,106,107,108,109]. Other mcr genes found in A. baumannii include mcr-2 and mcr-3 [110].
3.4. Overexpression of Efflux Pumps
The role of efflux pump in colistin resistance is suggested by few studies. Efflux pumps, such as the KpnEF and AcrAB have been reported in Enterobactericeae. The ΔKpnEF mutants showed increased susceptibility to various cationic antimicrobial peptides such as colistin [111]. On the other hand, AcrAB is a part of the AcrAB–TolC complex and its overespression has been observed in colistin-resistant E.coli, K. pneumoniae and Salmonella strains [112,113,114].
The contribution of EmrAB efflux system to colistin resistance in A. baumannii was shown by in vitro experiments with the ΔemrB mutant [115]. Moreover, the upregulation of genes encoding protein components of efflux pumps (adeI, adeC, emrB, mexB, and macAB) was also observed in colistin-resistant A. baumannii strains [83].
The overexpression of the efflux pumps MexXY (RND family) under exposure to ribosome-targeting antibiotics was found to correlate with increased level of colistin resistance in P. aeruginosa [116]. However, the heterogeneity of MexXY expression observed in clinical isolates of P. aeruginosa showing variable levels of colistin resistance suggested that contribution of the efflux pumps to colistin resistance might also be related to other specific genetic backgrounds [117].
Further evidence for the role of efflux pumps in colistin resistance is the suppression of resistance by efflux pump inhibitor (EPI), cyanide-3-chlorophenylhydrazone (CCCP) in A. baumannii, P. aeruginosa, K. pneumoniae, and S. maltophilia [118]. However, a possible explanation is that CCCP-mediated depolarization of the electrochemical gradient may restore the negative charge of the outer membrane and lead to increased susceptibility to colistin [48,118]. Furthermore, various studies suggested a complex regulatory relationship between the efflux pumps and their transcriptional regulators and LPS synthesis, transport, and modification [48].
4. Pharmacokinetic/Pharmacodynamic Features
According to several preclinical evidence, the free area under the concentration-to-time curve to minimum inhibitory concentration ratio (fAUC/MIC) was defined as the best pharmacokinetic/pharmacodynamic (PK/PD) target for colistin efficacy in infections caused by P. aeruginosa and A. baumannii [119]. In a neutropenic murine thigh and lung infection model against three P. aeruginosa strains, Dudhani et al. [120] found that the fAUC/MIC ratio was the best PK/PD index correlating with colistin efficacy both in thigh (R2=0.87) and lung infection model (R2=0.89). The colistin fAUC/MIC targets required to achieve 1-log and 2-log kill against the three strains were 15.6 to 22.8 and 27.6 to 36.1, respectively, in the thigh infection model, whereas a fAUC/MIC ratio ranging from 12.2 to 16.7 and from 36.9 to 45.9 was found in the lung infection model for achieving 1-log and 2-log kill [120]. In a neutropenic murine thigh and lung infection model against three A. baumannii strains (of which two were colistin heteroresistant), Dudhani et al. [121] reported that the fAUC/MIC ratio was the best PK/PD index correlating with colistin efficacy both in thigh (R2=0.90) and lung infection model (R2=0.80). The colistin fAUC/MIC targets required to achieve stasis and 1-log kill against the three strains were 1.89–7.41 and 6.98–13.6 in the thigh infection model, respectively, and 1.57–6.52 and 8.18–42.1, respectively, in the lung infection model [121]. Notably, these colistin PK/PD targets against P. aeruginosa and A. baumannii were consistent with those retrieved in a recent murine thigh and lung infection model [122]. Indeed, the fAUC/MIC ratio was confirmed as the best PK/PD target for predicting colistin efficacy, being desired fAUC/MIC ratios for achieving 2-log kill against Pseudomonas aeruginosa and A. baumannii strains of 7.4–13.7 and 7.4–17.6, respectively [122]. It should be noticed that these PK/PD targets could be attained only in two P. aeruginosa strains and in one A. baumannii strain in the lung infection model even at the highest colistin dose tolerated [122].
In Enterobacterales, an in vitro model investigated the best PK/PD target of colistin efficacy against three K. pneumoniae strains exhibiting MIC values of 0.5, 1, and 4 mg/L, respectively [123]. The fAUC/MIC ratio emerged as the best PK/PD target for colistin efficacy, being an fAUC/MIC ≥25 more predictive for a bactericidal effect [123]. Notably, this PK/PD target may be attained at standard colistin dose of 9 MU in 100%, 5-70%, and 0% of K. pneumoniae isolates showing an MIC value of 0.5, 1, and 2 mg/L, respectively [123]. These findings may suggest on the one hand the need for revising current colistin clinical breakpoint against Enterobacterales, and on the other hand the potential relevance of implementing a therapeutic drug monitoring (TDM)-guided approach for personalizing colistin dosage.
It should be noticed that evidence investigating the relationship between optimal PK/PD target attainment for colistin retrieved in preclinical studies and clinical outcome are currently limited. A prospective observational study investigated the relationship between PK/PD target attainment of colistin and microbiological/clinical outcome in nine patients affected by multidrug-resistant (MDR) Gram-negative infections (eight caused by A. baumannii and one by K. pneumoniae) [124]. After the fifth colistin dose of 2 MU, the AUC0-8/MIC ranged from 35.5 to 126- Although no significant relationship between AUC/MIC ratio and microbiological/clinical cure was found, a positive trend was observed at logistic regression (p=0.28) [124]. A prospective observational study including 33 patients affected by urinary tract infections and/or pyelonephritis caused by extremely drug-resistant P. aeruginosa reported no significant difference in fAUC/MIC ratio between cases exhibiting favourable clinical outcome and those with clinical failure (21.5 vs. 47.4; p=0.85) or in proportion of attainment of an AUC/MIC ratio ≥60 mg/L (32.3% vs. 50.0%; p=0.99) [125]. At multivariate analysis, average steady-state colistin concentration showed a trend towards statistical significance for acute kidney injury occurrence at the multivariate analysis (OR 4.36; 95%CI 0.86-20.0; p=0.07 (Sorlí et al., 2019).
Studies assessing colistin penetration in different sites of infection are reported in Table 1. Currently, data are available only for lung, central nervous system (CNS), and eye (Table 1). Specifically, a prospective observational study investigating epithelial lining fluid (ELF) penetration of intravenous colistin administered at a dosage of 2 MU every 8 hours in 13 critically ill patients affected by ventilator-associated pneumonia reported undetectable colistin concentrations in ELF [126].
A prospective observational study including five critically ill patients assessed colistin penetration in cerebrospinal fluid (CSF) administered intravenously at a dosage of 2-3 MU every 8 hours [127]. Colistin CSF-to-plasma ratio was 0.05, with absolute concentrations retrieved in CSF allowing to attain optimal PK/PD target only against P. aeruginosa and A. baumannii strains showing an MIC value up to 0.06 mg/L [127]. In regard to ocular penetration, only a preclinical animal model currently assessed this issue in twenty rabbits receiving intravenous colistin at a dosage of 5 mg/kg [128]. Overall, absolute colistin concentrations were extremely low in aqueous humor and undetectable in vitreous humor in most of included cases [128].
Overall, these findings may strongly support the implementation of alternative agents in case of deep-seated infections, according to the limited penetration colistin penetration rate in lung and CSF and the failure in attaining optimal PK/PD targets. Notably, these findings may be expected according to the physicochemical and PK features of colistin, namely hydrophilic properties, large molecular weight, and limited volume of distribution [129].
5. Conclusions
In the last years, the renewed of older antimicrobial molecules have revolutionized the treatment of infections due to MDR-GN microorganisms. At the same time, novel approaches including the therapeutic drug monitoring (TDM) for personalizing antimicrobial dosage of the different antimicrobial molecules and new therapeutic schemes of treatment by combining antibiotics with limited antimicrobial activity have revolutionized the treatment of infections due to MDR pathogens. In this context, the clinical usage of colistin alone and in combination with other antimicrobials with scarce and/or limited antimicrobial activity have recently reinvented its role in clinical practice. Also, considering the limited antimicrobial options against these pathogens, colistin was defined as the “last-hope resource” for the treatment of DTR infections especially among critical-ill patients.
On the other side, the adverse toxic effects and the limited tissue penetrations in different anatomical districts prompted to mitigate its role in clinical setting by limiting its use. In addition, the widespread of colistin resistant strains poses a serious limitation in the sue of this molecule especially to the light of the new antimicrobial molecules recently developed with high bactericidal activity against MDR microorganisms (i.e., cefiderocol, ceftazidime/avibactam, meropenem/vaborbactam, etc.).
Author Contributions
Authorship must be limited to those who have contributed substantially to the work reported.
Funding
This research received no external funding
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
None
Conflicts of Interest
The authors declare no conflict of interest
References
- Salam, M.A.; Al-Amin, M.Y.; Salam, M.T.; Pawar, J.S.; Akhter, N.; Rabaan, A.A.; Alqumber, M.A.A. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Healthcare (Basel) 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, M.I.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, Present and Future. Curr Opin Microbiol 2019, 51, 72–80. [Google Scholar] [CrossRef]
- Urban-Chmiel, R.; Marek, A.; Stępień-Pyśniak, D.; Wieczorek, K.; Dec, M.; Nowaczek, A.; Osek, J. Antibiotic Resistance in Bacteria-A Review. Antibiotics (Basel) 2022, 11. [Google Scholar] [CrossRef]
- Muteeb, G.; Rehman, M.T.; Shahwan, M.; Aatif, M. Origin of Antibiotics and Antibiotic Resistance, and Their Impacts on Drug Development: A Narrative Review. Pharmaceuticals (Basel) 2023, 16. [Google Scholar] [CrossRef]
- Walsh, C.T.; Wencewicz, T.A. Prospects for New Antibiotics: A Molecule-Centered Perspective. J Antibiot (Tokyo) 2014, 67, 7–22. [Google Scholar] [CrossRef] [PubMed]
- Gaibani, P.; Giani, T.; Bovo, F.; Lombardo, D.; Amadesi, S.; Lazzarotto, T.; Coppi, M.; Rossolini, G.M.; Ambretti, S. Resistance to Ceftazidime/Avibactam, Meropenem/Vaborbactam and Imipenem/Relebactam in Gram-Negative MDR Bacilli: Molecular Mechanisms and Susceptibility Testing. Antibiotics (Basel) 2022, 11. [Google Scholar] [CrossRef]
- El-Sayed Ahmed, M.A.E.-G.; Zhong, L.-L.; Shen, C.; Yang, Y.; Doi, Y.; Tian, G.-B. Colistin and Its Role in the Era of Antibiotic Resistance: An Extended Review (2000–2019). Emerg Microbes Infect 2020, 9, 868–885. [Google Scholar] [CrossRef]
- Mondal, A.H.; Khare, K.; Saxena, P.; Debnath, P.; Mukhopadhyay, K.; Yadav, D. A Review on Colistin Resistance: An Antibiotic of Last Resort. Microorganisms 2024, 12, 772. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y. A New Antibiotic “Colistin” Produced by Spore-Forming Soil Bacteria. J Antibiot. 1950, 3, 457–458. [Google Scholar]
- Stansly, P.G.; Schlosser, M.E. Studies on Polymyxin: Isolation and Identification of Bacillus Polymyxa and Differentiation of Polymyxin from Certain Known Antibiotics. J Bacteriol 1947, 54, 549–556. [Google Scholar] [CrossRef]
- Hamel, M.; Rolain, J.-M.; Baron, S.A. The History of Colistin Resistance Mechanisms in Bacteria: Progress and Challenges. Microorganisms 2021, 9, 442. [Google Scholar] [CrossRef] [PubMed]
- Bergen, P.J.; Li, J.; Rayner, C.R.; Nation, R.L. Colistin Methanesulfonate Is an Inactive Prodrug of Colistin against Pseudomonas Aeruginosa. Antimicrob Agents Chemother 2006, 50, 1953–1958. [Google Scholar] [CrossRef] [PubMed]
- Ehrentraut, S.F.; Muenster, S.; Kreyer, S.; Theuerkauf, N.U.; Bode, C.; Steinhagen, F.; Ehrentraut, H.; Schewe, J.-C.; Weber, M.; Putensen, C.; et al. Extensive Therapeutic Drug Monitoring of Colistin in Critically Ill Patients Reveals Undetected Risks. Microorganisms 2020, 8, 415. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, K. Why and How Are Peptide–Lipid Interactions Utilized for Self-Defense? Magainins and Tachyplesins as Archetypes. Biochimica et Biophysica Acta (BBA) - Biomembranes 1999, 1462, 1–10. [Google Scholar] [CrossRef]
- Shai, Y. Mechanism of the Binding, Insertion and Destabilization of Phospholipid Bilayer Membranes by α-Helical Antimicrobial and Cell Non-Selective Membrane-Lytic Peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes 1999, 1462, 55–70. [Google Scholar] [CrossRef]
- Yang, L.; Weiss, T.M.; Lehrer, R.I.; Huang, H.W. Crystallization of Antimicrobial Pores in Membranes: Magainin and Protegrin. Biophys J 2000, 79, 2002–2009. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Lee, W.; Kwa, A.L. Polymyxin B versus Colistin: An Update. Expert Rev Anti Infect Ther 2015, 13, 1481–1497. [Google Scholar] [CrossRef] [PubMed]
- Trimble, M.J.; Mlynárčik, P.; Kolář, M.; Hancock, R.E.W. Polymyxin: Alternative Mechanisms of Action and Resistance. Cold Spring Harb Perspect Med 2016, 6, a025288. [Google Scholar] [CrossRef] [PubMed]
- Andrade, F.F.; Silva, D.; Rodrigues, A.; Pina-Vaz, C. Colistin Update on Its Mechanism of Action and Resistance, Present and Future Challenges. Microorganisms 2020, 8. [Google Scholar] [CrossRef]
- Velkov, T.; Thompson, P.E.; Nation, R.L.; Li, J. Structure−Activity Relationships of Polymyxin Antibiotics. J Med Chem 2010, 53, 1898–1916. [Google Scholar] [CrossRef]
- Falagas, M.E.; Kasiakou, S.K.; Saravolatz, L.D. Colistin: The Revival of Polymyxins for the Management of Multidrug-Resistant Gram-Negative Bacterial Infections. Clinical Infectious Diseases 2005, 40, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Kaye, K.S.; Pogue, J.M.; Tran, T.B.; Nation, R.L.; Li, J. Agents of Last Resort. Infect Dis Clin North Am 2016, 30, 391–414. [Google Scholar] [CrossRef] [PubMed]
- Gurjar, M. Colistin for Lung Infection: An Update. J Intensive Care 2015, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Deris, Z.Z.; Akter, J.; Sivanesan, S.; Roberts, K.D.; Thompson, P.E.; Nation, R.L.; Li, J.; Velkov, T. A Secondary Mode of Action of Polymyxins against Gram-Negative Bacteria Involves the Inhibition of NADH-Quinone Oxidoreductase Activity. J Antibiot (Tokyo) 2014, 67, 147–151. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Chan, T.; Xu, C.; Zhu, L.; Zhou, Q.T.; Roberts, K.D.; Chan, H.-K.; Li, J.; Zhou, F. Human Oligopeptide Transporter 2 (PEPT2) Mediates Cellular Uptake of Polymyxins. J Antimicrob Chemother 2016, 71, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Visentin, M.; Gai, Z.; Torozi, A.; Hiller, C.; Kullak-Ublick, G.A. Colistin Is Substrate of the Carnitine/Organic Cation Transporter 2 (OCTN2, SLC22A5). Drug Metabolism and Disposition 2017, 45, 1240–1244. [Google Scholar] [CrossRef] [PubMed]
- Gai, Z.; Samodelov, S.; Kullak-Ublick, G.; Visentin, M. Molecular Mechanisms of Colistin-Induced Nephrotoxicity. Molecules 2019, 24, 653. [Google Scholar] [CrossRef]
- Kilic, I.; Ayar, Y.; Ceylan, İ.; Kaya, P.K.; Caliskan, G. Nephrotoxicity Caused by Colistin Use in ICU: A Single Centre Experience. BMC Nephrol 2023, 24, 302. [Google Scholar] [CrossRef]
- Nation, R.L.; Li, J. Colistin in the 21st Century. Curr Opin Infect Dis 2009, 22, 535–543. [Google Scholar] [CrossRef]
- Akajagbor, D.S.; Wilson, S.L.; Shere-Wolfe, K.D.; Dakum, P.; Charurat, M.E.; Gilliam, B.L. Higher Incidence of Acute Kidney Injury With Intravenous Colistimethate Sodium Compared With Polymyxin B in Critically Ill Patients at a Tertiary Care Medical Center. Clinical Infectious Diseases 2013, 57, 1300–1303. [Google Scholar] [CrossRef]
- Dalfino, L.; Puntillo, F.; Ondok, M.J.M.; Mosca, A.; Monno, R.; Coppolecchia, S.; Spada, M.L.; Bruno, F.; Brienza, N. Colistin-Associated Acute Kidney Injury in Severely Ill Patients: A Step Toward a Better Renal Care? A Prospective Cohort Study. Clinical Infectious Diseases 2015, 61, 1771–1777. [Google Scholar] [CrossRef]
- ÖZKARAKAŞ, H.; KÖSE, I.; ZİNCİRCİOĞLU, Ç.; ERSAN, S.; ERSAN, G.; ŞENOĞLU, N.; KÖSE, Ş.; ERBAY, R.H. Risk Factors for Colistin-Associated Nephrotoxicity and Mortality in Critically Ill Patients. Turk J Med Sci 2017, 47, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Petrosillo, N.; Giannella, M.; Antonelli, M.; Antonini, M.; Barsic, B.; Belancic, L.; Inkaya A., C.; De Pascale, G.; Grilli, E.; Tumbarello, M.; et al. Clinical Experience of Colistin-Glycopeptide Combination in Critically Ill Patients Infected with Gram-Negative Bacteria. Antimicrob Agents Chemother 2014, 58, 851–858. [Google Scholar] [CrossRef]
- Falagas, M.E.; Kasiakou, S.K.; Tsiodras, S.; Michalopoulos, A. The Use of Intravenous and Aerosolized Polymyxins for the Treatment of Infections in Critically Ill Patients: A Review of the Recent Literature. Clin Med Res 2006, 4, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Spapen, H.; Jacobs, R.; Van Gorp, V.; Troubleyn, J.; Honoré, P.M. Renal and Neurological Side Effects of Colistin in Critically Ill Patients. Ann Intensive Care 2011, 1, 14. [Google Scholar] [CrossRef]
- KUBIKOWSKI, P.; SZRENIAWSKI, Z. THE MECHANISM OF THE NEUROMUSCULAR BLOCKADE BY ANTIBIOTICS. Arch Int Pharmacodyn Ther 1963, 146, 549–560. [Google Scholar] [PubMed]
- Wang, Y.; Li, H.; Xie, X.; Wu, X.; Li, X.; Zhao, Z.; Luo, S.; Wan, Z.; Liu, J.; Fu, L.; et al. In Vitro and in Vivo Assessment of the Antibacterial Activity of Colistin Alone and in Combination with Other Antibiotics against Acinetobacter Baumannii and Escherichia Coli. J Glob Antimicrob Resist 2020, 20, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Walkty, A.; DeCorby, M.; Nichol, K.; Karlowsky, J.A.; Hoban, D.J.; Zhanel, G.G. In Vitro Activity of Colistin (Polymyxin E) against 3,480 Isolates of Gram-Negative Bacilli Obtained from Patients in Canadian Hospitals in the CANWARD Study, 2007-2008. Antimicrob Agents Chemother 2009, 53, 4924–4926. [Google Scholar] [CrossRef] [PubMed]
- Gill, M.M.; Rao, J.U.; Kaleem, F.; Hassan, A.; Khalid, A.; Anjum, R. In Vitro Efficacy of Colistin against Multi-Drug Resistant Pseudomonas Aeruginosa by Minimum Inhibitory Concentration. Pak J Pharm Sci 2013, 26, 7–10. [Google Scholar]
- Gaibani, P.; Lombardo, D.; Lewis, R.E.; Mercuri, M.; Bonora, S.; Landini, M.P.; Ambretti, S. In Vitro Activity and Post-Antibiotic Effects of Colistin in Combination with Other Antimicrobials against Colistin-Resistant KPC-Producing Klebsiella Pneumoniae Bloodstream Isolates. Journal of Antimicrobial Chemotherapy 2014, 69, 1856–1865. [Google Scholar] [CrossRef]
- Kheshti, R.; Pourabbas, B.; Mosayebi, M.; Vazin, A. In Vitro Activity of Colistin in Combination with Various Antimicrobials against <em>Acinetobacter Baumannii</Em> Species, a Report from South Iran. Infect Drug Resist 2018, Volume 12, 129–135. [Google Scholar] [CrossRef]
- Brennan-Krohn, T.; Grote, A.; Rodriguez, S.; Kirby, J.E.; Earl, A.M. Transcriptomics Reveals How Minocycline-Colistin Synergy Overcomes Antibiotic Resistance in Multidrug-Resistant Klebsiella Pneumoniae. Antimicrob Agents Chemother 2022, 66. [Google Scholar] [CrossRef] [PubMed]
- Rout, B.; Dash, S.K.; Sahu, K. kumar; Behera, B.; Praharaj, I.; Otta, S. Evaluation of Different Methods for in Vitro Susceptibility Testing of Colistin in Carbapenem Resistant Gram-Negative Bacilli. Access Microbiol 2023, 5. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI) Performance and Standards for Antimicrobial Susceptibility Testing. CLSI Supplement M100; CLSI: 950 West Wiley Road, Suite 2500. [Google Scholar]
- Simner, P.J.; Bergman, Y.; Trejo, M.; Roberts, A.A.; Marayan, R.; Tekle, T.; Campeau, S.; Kazmi, A.Q.; Bell, D.T.; Lewis, S.; et al. Two-Site Evaluation of the Colistin Broth Disk Elution Test To Determine Colistin In Vitro Activity against Gram-Negative Bacilli. J Clin Microbiol 2019, 57. [Google Scholar] [CrossRef] [PubMed]
- Needham, B.D.; Trent, M.S. Fortifying the Barrier: The Impact of Lipid A Remodelling on Bacterial Pathogenesis. Nat Rev Microbiol 2013, 11, 467–481. [Google Scholar] [CrossRef]
- Novović, K.; Jovčić, B. Colistin Resistance in Acinetobacter Baumannii: Molecular Mechanisms and Epidemiology. Antibiotics 2023, 12, 516. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Hao, J.; Xiao, W.; Ye, C.; Xiao, X.; Jian, C.; Tang, M.; Li, G.; Liu, J.; Zeng, Z. Role of Efflux Pumps, Their Inhibitors, and Regulators in Colistin Resistance. Front Microbiol 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Aghapour, Z.; Gholizadeh, P.; Ganbarov, K.; bialvaei, A.Z.; Mahmood, S.S.; Tanomand, A.; Yousefi, M.; Asgharzadeh, M.; Yousefi, B.; Samadi Kafil, H. <p>Molecular Mechanisms Related to Colistin Resistance in Enterobacteriaceae</P>. Infect Drug Resist 2019, Volume 12, 965–975. [Google Scholar] [CrossRef]
- Zhang, H.; Srinivas, S.; Xu, Y.; Wei, W.; Feng, Y. Genetic and Biochemical Mechanisms for Bacterial Lipid A Modifiers Associated with Polymyxin Resistance. Trends Biochem Sci 2019, 44, 973–988. [Google Scholar] [CrossRef]
- Olaitan, A.O.; Morand, S.; Rolain, J.-M. Mechanisms of Polymyxin Resistance: Acquired and Intrinsic Resistance in Bacteria. Front Microbiol 2014, 5. [Google Scholar] [CrossRef]
- Pelletier, M.R.; Casella, L.G.; Jones, J.W.; Adams, M.D.; Zurawski, D. V.; Hazlett, K.R.O.; Doi, Y.; Ernst, R.K. Unique Structural Modifications Are Present in the Lipopolysaccharide from Colistin-Resistant Strains of Acinetobacter Baumannii. Antimicrob Agents Chemother 2013, 57, 4831–4840. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Rafailidis, P.I.; Matthaiou, D.K. Resistance to Polymyxins: Mechanisms, Frequency and Treatment Options. Drug Resistance Updates 2010, 13, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Ly, N.S.; Yang, J.; Bulitta, J.B.; Tsuji, B.T. Impact of Two-Component Regulatory Systems PhoP-PhoQ and PmrA-PmrB on Colistin Pharmacodynamics in Pseudomonas Aeruginosa. Antimicrob Agents Chemother 2012, 56, 3453–3456. [Google Scholar] [CrossRef] [PubMed]
- Gunn, J.S. The Salmonella PmrAB Regulon: Lipopolysaccharide Modifications, Antimicrobial Peptide Resistance and More. Trends Microbiol 2008, 16, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, A.O.; Dia, N.M.; Gautret, P.; Benkouiten, S.; Belhouchat, K.; Drali, T.; Parola, P.; Brouqui, P.; Memish, Z.; Raoult, D.; et al. Acquisition of Extended-Spectrum Cephalosporin- and Colistin-Resistant Salmonella Enterica Subsp. Enterica Serotype Newport by Pilgrims during Hajj. Int J Antimicrob Agents 2015, 45, 600–604. [Google Scholar] [CrossRef]
- Quesada, A.; Porrero, M.C.; Téllez, S.; Palomo, G.; García, M.; Domínguez, L. Polymorphism of Genes Encoding PmrAB in Colistin-Resistant Strains of Escherichia Coli and Salmonella Enterica Isolated from Poultry and Swine. Journal of Antimicrobial Chemotherapy 2015, 70, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Cannatelli, A.; Di Pilato, V.; Giani, T.; Arena, F.; Ambretti, S.; Gaibani, P.; D’Andrea, M.M.; Rossolini, G.M. In Vivo Evolution to Colistin Resistance by PmrB Sensor Kinase Mutation in KPC-Producing Klebsiella Pneumoniae Is Associated with Low-Dosage Colistin Treatment. Antimicrob Agents Chemother 2014, 58, 4399–4403. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-H.; Lin, T.-L.; Pan, Y.-J.; Wang, Y.-P.; Lin, Y.-T.; Wang, J.-T. Colistin Resistance Mechanisms in Klebsiella Pneumoniae Strains from Taiwan. Antimicrob Agents Chemother 2015, 59, 2909–2913. [Google Scholar] [CrossRef] [PubMed]
- Jayol, A.; Poirel, L.; Brink, A.; Villegas, M.-V.; Yilmaz, M.; Nordmann, P. Resistance to Colistin Associated with a Single Amino Acid Change in Protein PmrB among Klebsiella Pneumoniae Isolates of Worldwide Origin. Antimicrob Agents Chemother 2014, 58, 4762–4766. [Google Scholar] [CrossRef]
- Gerson, S.; Betts, J.W.; Lucaßen, K.; Nodari, C.S.; Wille, J.; Josten, M.; Göttig, S.; Nowak, J.; Stefanik, D.; Roca, I.; et al. Investigation of Novel PmrB and EptA Mutations in Isogenic Acinetobacter Baumannii Isolates Associated with Colistin Resistance and Increased Virulence In Vivo. Antimicrob Agents Chemother 2019, 63. [Google Scholar] [CrossRef]
- Park, Y.K.; Choi, J.Y.; Shin, D.; Ko, K.S. Correlation between Overexpression and Amino Acid Substitution of the PmrAB Locus and Colistin Resistance in Acinetobacter Baumannii. Int J Antimicrob Agents 2011, 37, 525–530. [Google Scholar] [CrossRef]
- Dahdouh, E.; Gómez-Gil, R.; Sanz, S.; González-Zorn, B.; Daoud, Z.; Mingorance, J.; Suárez, M. A Novel Mutation in PmrB Mediates Colistin Resistance during Therapy of Acinetobacter Baumannii. Int J Antimicrob Agents 2017, 49, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Moskowitz, S.M.; Brannon, M.K.; Dasgupta, N.; Pier, M.; Sgambati, N.; Miller, A.K.; Selgrade, S.E.; Miller, S.I.; Denton, M.; Conway, S.P.; et al. PmrB Mutations Promote Polymyxin Resistance of Pseudomonas Aeruginosa Isolated from Colistin-Treated Cystic Fibrosis Patients. Antimicrob Agents Chemother 2012, 56, 1019–1030. [Google Scholar] [CrossRef] [PubMed]
- Owusu-Anim, D.; Kwon, D.H. Differential Role of Two-Component Regulatory Systems (≪I≫PhoPQ≪/I≫ and ≪I≫PmrAB≪/I≫) in Polymyxin B Susceptibility of ≪I≫Pseudomonas Aeruginosa≪/I≫ Adv Microbiol 2012, 02, 31–36. Adv Microbiol. [CrossRef]
- Olaitan, A.O.; Morand, S.; Rolain, J.-M. Emergence of Colistin-Resistant Bacteria in Humans without Colistin Usage: A New Worry and Cause for Vigilance. Int J Antimicrob Agents 2016, 47, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-H.; Siu, L.K.; Chang, F.-Y.; Tsai, Y.-K.; Lin, Y.-T.; Chiu, S.-K.; Huang, L.-Y.; Lin, J.-C. A Novel Deletion Mutation in PmrB Contributes to Concurrent Colistin Resistance in Carbapenem-Resistant Escherichia Coli Sequence Type 405 of Clinical Origin. Antimicrob Agents Chemother 2020, 64. [Google Scholar] [CrossRef] [PubMed]
- Jayol, A.; Nordmann, P.; Brink, A.; Poirel, L. Heteroresistance to Colistin in Klebsiella Pneumoniae Associated with Alterations in the PhoPQ Regulatory System. Antimicrob Agents Chemother 2015, 59, 2780–2784. [Google Scholar] [CrossRef] [PubMed]
- Olaitan, A.O.; Diene, S.M.; Kempf, M.; Berrazeg, M.; Bakour, S.; Gupta, S.K.; Thongmalayvong, B.; Akkhavong, K.; Somphavong, S.; Paboriboune, P.; et al. Worldwide Emergence of Colistin Resistance in Klebsiella Pneumoniae from Healthy Humans and Patients in Lao PDR, Thailand, Israel, Nigeria and France Owing to Inactivation of the PhoP/PhoQ Regulator MgrB: An Epidemiological and Molecular Study. Int J Antimicrob Agents 2014, 44, 500–507. [Google Scholar] [CrossRef]
- Nordmann, P.; Jayol, A.; Poirel, L. Rapid Detection of Polymyxin Resistance in Enterobacteriaceae. Emerg Infect Dis 2016, 22, 1038–1043. [Google Scholar] [CrossRef] [PubMed]
- Gutu, A.D.; Sgambati, N.; Strasbourger, P.; Brannon, M.K.; Jacobs, M.A.; Haugen, E.; Kaul, R.K.; Johansen, H.K.; Høiby, N.; Moskowitz, S.M. Polymyxin Resistance of Pseudomonas Aeruginosa PhoQ Mutants Is Dependent on Additional Two-Component Regulatory Systems. Antimicrob Agents Chemother 2013, 57, 2204–2215. [Google Scholar] [CrossRef]
- Lippa, A.M.; Goulian, M. Feedback Inhibition in the PhoQ/PhoP Signaling System by a Membrane Peptide. PLoS Genet 2009, 5, e1000788. [Google Scholar] [CrossRef]
- Cannatelli, A.; Giani, T.; D’Andrea, M.M.; Di Pilato, V.; Arena, F.; Conte, V.; Tryfinopoulou, K.; Vatopoulos, A.; Rossolini, G.M. MgrB Inactivation Is a Common Mechanism of Colistin Resistance in KPC-Producing Klebsiella Pneumoniae of Clinical Origin. Antimicrob Agents Chemother 2014, 58, 5696–5703. [Google Scholar] [CrossRef] [PubMed]
- Wright, M.S.; Suzuki, Y.; Jones, M.B.; Marshall, S.H.; Rudin, S.D.; van Duin, D.; Kaye, K.; Jacobs, M.R.; Bonomo, R.A.; Adams, M.D. Genomic and Transcriptomic Analyses of Colistin-Resistant Clinical Isolates of Klebsiella Pneumoniae Reveal Multiple Pathways of Resistance. Antimicrob Agents Chemother 2015, 59, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Cannatelli, A.; D’Andrea, M.M.; Giani, T.; Di Pilato, V.; Arena, F.; Ambretti, S.; Gaibani, P.; Rossolini, G.M. In Vivo Emergence of Colistin Resistance in Klebsiella Pneumoniae Producing KPC-Type Carbapenemases Mediated by Insertional Inactivation of the PhoQ/PhoP MgrB Regulator. Antimicrob Agents Chemother 2013, 57, 5521–5526. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Camacho, E.; Gomez-Gil, R.; Tobes, R.; Manrique, M.; Lorenzo, M.; Galvan, B.; Salvarelli, E.; Moatassim, Y.; Salanueva, I.J.; Pareja, E.; et al. Genomic Analysis of the Emergence and Evolution of Multidrug Resistance during a Klebsiella Pneumoniae Outbreak Including Carbapenem and Colistin Resistance. Journal of Antimicrobial Chemotherapy 2014, 69, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; Jayol, A.; Bontron, S.; Villegas, M.-V.; Ozdamar, M.; Turkoglu, S.; Nordmann, P. The MgrB Gene as a Key Target for Acquired Resistance to Colistin in Klebsiella Pneumoniae. Journal of Antimicrobial Chemotherapy 2015, 70, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.-H.; Lin, T.-L.; Lin, Y.-T.; Wang, J.-T. Amino Acid Substitutions of CrrB Responsible for Resistance to Colistin through CrrC in Klebsiella Pneumoniae. Antimicrob Agents Chemother 2016, 60, 3709–3716. [Google Scholar] [CrossRef] [PubMed]
- Baron, S.; Hadjadj, L.; Rolain, J.-M.; Olaitan, A.O. Molecular Mechanisms of Polymyxin Resistance: Knowns and Unknowns. Int J Antimicrob Agents 2016, 48, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Trebosc, V.; Gartenmann, S.; Tötzl, M.; Lucchini, V.; Schellhorn, B.; Pieren, M.; Lociuro, S.; Gitzinger, M.; Tigges, M.; Bumann, D.; et al. Dissecting Colistin Resistance Mechanisms in Extensively Drug-Resistant Acinetobacter Baumannii Clinical Isolates. mBio 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Moffatt, J.H.; Harper, M.; Harrison, P.; Hale, J.D.F.; Vinogradov, E.; Seemann, T.; Henry, R.; Crane, B.; St. Michael, F.; Cox, A.D.; et al. Colistin Resistance in Acinetobacter Baumannii Is Mediated by Complete Loss of Lipopolysaccharide Production. Antimicrob Agents Chemother 2010, 54, 4971–4977. [Google Scholar] [CrossRef]
- Carretero-Ledesma, M.; García-Quintanilla, M.; Martín-Peña, R.; Pulido, M.R.; Pachón, J.; McConnell, M.J. Phenotypic Changes Associated with Colistin Resistance Due to Lipopolysaccharide Loss in Acinetobacter Baumannii. Virulence 2018, 9, 930–942. [Google Scholar] [CrossRef]
- Boinett, C.J.; Cain, A.K.; Hawkey, J.; Do Hoang, N.T.; Khanh, N.N.T.; Thanh, D.P.; Dordel, J.; Campbell, J.I.; Lan, N.P.H.; Mayho, M.; et al. Clinical and Laboratory-Induced Colistin-Resistance Mechanisms in Acinetobacter Baumannii. Microb Genom 2019, 5. [Google Scholar] [CrossRef]
- Lean, S.-S.; Yeo, C.C.; Suhaili, Z.; Thong, K.-L. Comparative Genomics of Two ST 195 Carbapenem-Resistant Acinetobacter Baumannii with Different Susceptibility to Polymyxin Revealed Underlying Resistance Mechanism. Front Microbiol 2016, 6. [Google Scholar] [CrossRef]
- Moffatt, J.H.; Harper, M.; Adler, B.; Nation, R.L.; Li, J.; Boyce, J.D. Insertion Sequence IS Aba11 Is Involved in Colistin Resistance and Loss of Lipopolysaccharide in Acinetobacter Baumannii. Antimicrob Agents Chemother 2011, 55, 3022–3024. [Google Scholar] [CrossRef] [PubMed]
- Kamoshida, G.; Yamada, N.; Nakamura, T.; Yamaguchi, D.; Kai, D.; Yamashita, M.; Hayashi, C.; Kanda, N.; Sakaguchi, M.; Morimoto, H.; et al. Preferential Selection of Low-Frequency, Lipopolysaccharide-Modified, Colistin-Resistant Mutants with a Combination of Antimicrobials in Acinetobacter Baumannii. Microbiol Spectr 2022, 10. [Google Scholar] [CrossRef]
- Beceiro, A.; Moreno, A.; Fernández, N.; Vallejo, J.A.; Aranda, J.; Adler, B.; Harper, M.; Boyce, J.D.; Bou, G. Biological Cost of Different Mechanisms of Colistin Resistance and Their Impact on Virulence in Acinetobacter Baumannii. Antimicrob Agents Chemother 2014, 58, 518–526. [Google Scholar] [CrossRef]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet Infect Dis 2016, 16, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Kieffer, N.; Nordmann, P.; Poirel, L. Moraxella Species as Potential Sources of MCR-Like Polymyxin Resistance Determinants. Antimicrob Agents Chemother 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Xavier, B.B.; Lammens, C.; Ruhal, R.; Kumar-Singh, S.; Butaye, P.; Goossens, H.; Malhotra-Kumar, S. Identification of a Novel Plasmid-Mediated Colistin-Resistance Gene, Mcr-2, in Escherichia Coli, Belgium, June 2016. Euro Surveill 2016, 21. [Google Scholar] [CrossRef]
- Yin, W.; Li, H.; Shen, Y.; Liu, Z.; Wang, S.; Shen, Z.; Zhang, R.; Walsh, T.R.; Shen, J.; Wang, Y. Novel Plasmid-Mediated Colistin Resistance Gene Mcr-3 in Escherichia Coli. mBio 2017, 8. [Google Scholar] [CrossRef]
- Carattoli, A.; Villa, L.; Feudi, C.; Curcio, L.; Orsini, S.; Luppi, A.; Pezzotti, G.; Magistrali, C.F. Novel Plasmid-Mediated Colistin Resistance Mcr-4 Gene in Salmonella and Escherichia Coli, Italy 2013, Spain and Belgium, 2015 to 2016. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef]
- Borowiak, M.; Fischer, J.; Hammerl, J.A.; Hendriksen, R.S.; Szabo, I.; Malorny, B. Identification of a Novel Transposon-Associated Phosphoethanolamine Transferase Gene, Mcr-5, Conferring Colistin Resistance in d-Tartrate Fermenting Salmonella Enterica Subsp. Enterica Serovar Paratyphi B. Journal of Antimicrobial Chemotherapy 2017, 72, 3317–3324. [Google Scholar] [CrossRef] [PubMed]
- Zurfluh, K.; Tasara, T.; Poirel, L.; Nordmann, P.; Stephan, R. Draft Genome Sequence of Escherichia Coli S51, a Chicken Isolate Harboring a Chromosomally Encoded Mcr-1 Gene. Genome Announc 2016, 4. [Google Scholar] [CrossRef] [PubMed]
- Partridge, S.R.; Di Pilato, V.; Doi, Y.; Feldgarden, M.; Haft, D.H.; Klimke, W.; Kumar-Singh, S.; Liu, J.-H.; Malhotra-Kumar, S.; Prasad, A.; et al. Proposal for Assignment of Allele Numbers for Mobile Colistin Resistance (Mcr) Genes. Journal of Antimicrobial Chemotherapy 2018, 73, 2625–2630. [Google Scholar] [CrossRef] [PubMed]
- Nitz, F.; de Melo, B.O.; da Silva, L.C.N.; de Souza Monteiro, A.; Marques, S.G.; Monteiro-Neto, V.; de Jesus Gomes Turri, R.; Junior, A.D.S.; Conceição, P.C.R.; Magalhães, H.J.C.; et al. Molecular Detection of Drug-Resistance Genes of BlaOXA-23-BlaOXA-51 and Mcr-1 in Clinical Isolates of Pseudomonas Aeruginosa. Microorganisms 2021, 9, 786. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Baky, R.M.; Masoud, S.M.; Mohamed, D.S.; Waly, N.G.; Shafik, E.A.; Mohareb, D.A.; Elkady, A.; Elbadr, M.M.; Hetta, H.F. <p>Prevalence and Some Possible Mechanisms of Colistin Resistance Among Multidrug-Resistant and Extensively Drug-Resistant <em>Pseudomonas Aeruginosa</Em></P>. Infect Drug Resist 2020, Volume 13, 323–332. [Google Scholar] [CrossRef]
- Shabban, M.; Fahim, N.A.E.; Montasser, K.; El Magd, N.M.A. Resistance to Colistin Mediated by Mcr-1 among Multidrug Resistant Gram Negative Pathogens at a Tertiary Care Hospital, Egypt. J Pure Appl Microbiol 2020, 14, 1125–1132. [Google Scholar] [CrossRef]
- Hameed, F.; Khan, M.A.; Muhammad, H.; Sarwar, T.; Bilal, H.; Rehman, T.U. Plasmid-Mediated Mcr-1 Gene in Acinetobacter Baumannii and Pseudomonas Aeruginosa: First Report from Pakistan. Rev Soc Bras Med Trop 2019, 52. [Google Scholar] [CrossRef] [PubMed]
- Martins, E.; Maboni, G.; Battisti, R.; da Costa, L.; Selva, H.L.; Levitzki, E.D.; Gressler, L.T. High Rates of Multidrug Resistance in Bacteria Associated with Small Animal Otitis: A Study of Cumulative Microbiological Culture and Antimicrobial Susceptibility. Microb Pathog 2022, 165, 105399. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, Z.S.; Elshafiee, E.A.; Khalefa, H.S.; Kadry, M.; Hamza, D.A. Evidence of Colistin Resistance Genes (Mcr-1 and Mcr-2) in Wild Birds and Its Public Health Implication in Egypt. Antimicrob Resist Infect Control 2019, 8, 197. [Google Scholar] [CrossRef]
- Tartor, Y.H.; Gharieb, R.M.A.; Abd El-Aziz, N.K.; El Damaty, H.M.; Enany, S.; Khalifa, E.; Attia, A.S.A.; Abdellatif, S.S.; Ramadan, H. Virulence Determinants and Plasmid-Mediated Colistin Resistance Mcr Genes in Gram-Negative Bacteria Isolated From Bovine Milk. Front Cell Infect Microbiol 2021, 11. [Google Scholar] [CrossRef]
- Javed, H.; Saleem, S.; Zafar, A.; Ghafoor, A.; Shahzad, A. Bin; Ejaz, H.; Junaid, K.; Jahan, S. Emergence of Plasmid-Mediated Mcr Genes from Gram-Negative Bacteria at the Human-Animal Interface. Gut Pathog 2020, 12, 54. [Google Scholar] [CrossRef] [PubMed]
- Fujihara, H.; Yamazoe, A.; Hosoyama, A.; Suenaga, H.; Kimura, N.; Hirose, J.; Watanabe, T.; Futagami, T.; Goto, M.; Furukawa, K. Draft Genome Sequence of Pseudomonas Aeruginosa KF702 (NBRC 110665), a Polychlorinated Biphenyl-Degrading Bacterium Isolated from Biphenyl-Contaminated Soil. Genome Announc 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Snesrud, E.; Maybank, R.; Kwak, Y.I.; Jones, A.R.; Hinkle, M.K.; McGann, P. Chromosomally Encoded Mcr-5 in Colistin-Nonsusceptible Pseudomonas Aeruginosa. Antimicrob Agents Chemother 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Martins-Sorenson, N.; Snesrud, E.; Xavier, D.E.; Cacci, L.C.; Iavarone, A.T.; McGann, P.; Riley, L.W.; Moreira, B.M. A Novel Plasmid-Encoded Mcr-4.3 Gene in a Colistin-Resistant Acinetobacter Baumannii Clinical Strain. Journal of Antimicrobial Chemotherapy 2020, 75, 60–64. [Google Scholar] [CrossRef] [PubMed]
- Fan, R.; Li, C.; Duan, R.; Qin, S.; Liang, J.; Xiao, M.; Lv, D.; Jing, H.; Wang, X. Retrospective Screening and Analysis of Mcr-1 and BlaNDM in Gram-Negative Bacteria in China, 2010–2019. Front Microbiol 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Kalová, A.; Gelbíčová, T.; Overballe-Petersen, S.; Litrup, E.; Karpíšková, R. Characterisation of Colistin -Resistant Enterobacterales and Acinetobacter Strains Carrying Mcr Genes from Asian Aquaculture Products. Antibiotics 2021, 10, 838. [Google Scholar] [CrossRef] [PubMed]
- Bitar, I.; Medvecky, M.; Gelbicova, T.; Jakubu, V.; Hrabak, J.; Zemlickova, H.; Karpiskova, R.; Dolejska, M. Complete Nucleotide Sequences of Mcr-4.3 -Carrying Plasmids in Acinetobacter Baumannii Sequence Type 345 of Human and Food Origin from the Czech Republic, the First Case in Europe. Antimicrob Agents Chemother 2019, 63. [Google Scholar] [CrossRef] [PubMed]
- Al-Kadmy, I.M.S.; Ibrahim, S.A.; Al-Saryi, N.; Aziz, S.N.; Besinis, A.; Hetta, H.F. Prevalence of Genes Involved in Colistin Resistance in Acinetobacter Baumannii: First Report from Iraq. Microbial Drug Resistance 2020, 26, 616–622. [Google Scholar] [CrossRef]
- Srinivasan, V.B.; Rajamohan, G. KpnEF, a New Member of the Klebsiella Pneumoniae Cell Envelope Stress Response Regulon, Is an SMR-Type Efflux Pump Involved in Broad-Spectrum Antimicrobial Resistance. Antimicrob Agents Chemother 2013, 57, 4449–4462. [Google Scholar] [CrossRef]
- Warner, D.M.; Levy, S.B. Different Effects of Transcriptional Regulators MarA, SoxS and Rob on Susceptibility of Escherichia Coli to Cationic Antimicrobial Peptides (CAMPs): Rob-Dependent CAMP Induction of the MarRAB Operon. Microbiology (N Y) 2010, 156, 570–578. [Google Scholar] [CrossRef]
- Koutsolioutsou, A.; Peña-Llopis, S.; Demple, B. Constitutive SoxR Mutations Contribute to Multiple-Antibiotic Resistance in Clinical Escherichia Coli Isolates. Antimicrob Agents Chemother 2005, 49, 2746–2752. [Google Scholar] [CrossRef] [PubMed]
- Parra-Lopez, C.; Baer, M.T.; Groisman, E.A. Molecular Genetic Analysis of a Locus Required for Resistance to Antimicrobial Peptides in Salmonella Typhimurium. EMBO J 1993, 12, 4053–4062. [Google Scholar] [CrossRef]
- Lin, M.-F.; Lin, Y.-Y.; Lan, C.-Y. Contribution of EmrAB Efflux Pumps to Colistin Resistance in Acinetobacter Baumannii. Journal of Microbiology 2017, 55, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Poole, K.; Lau, C.H.-F.; Gilmour, C.; Hao, Y.; Lam, J.S. Polymyxin Susceptibility in Pseudomonas Aeruginosa Linked to the MexXY-OprM Multidrug Efflux System. Antimicrob Agents Chemother 2015, 59, 7276–7289. [Google Scholar] [CrossRef]
- Goli, H.R.; Nahaei, M.R.; Ahangarzadeh Rezaee, M.; Hasani, A.; Samadi Kafil, H.; Aghazadeh, M. Emergence of Colistin Resistant Pseudomonas Aeruginosa at Tabriz Hospitals, Iran. Iran J Microbiol 2016, 8, 62–69. [Google Scholar] [PubMed]
- Ni, W.; Li, Y.; Guan, J.; Zhao, J.; Cui, J.; Wang, R.; Liu, Y. Effects of Efflux Pump Inhibitors on Colistin Resistance in Multidrug-Resistant Gram-Negative Bacteria. Antimicrob Agents Chemother 2016, 60, 3215–3218. [Google Scholar] [CrossRef] [PubMed]
- Bergen, P.J.; Li, J.; Nation, R.L. Dosing of Colistin—Back to Basic PK/PD. Curr Opin Pharmacol 2011, 11, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Dudhani, R. V.; Turnidge, J.D.; Coulthard, K.; Milne, R.W.; Rayner, C.R.; Li, J.; Nation, R.L. Elucidation of the Pharmacokinetic/Pharmacodynamic Determinant of Colistin Activity against Pseudomonas Aeruginosa in Murine Thigh and Lung Infection Models. Antimicrob Agents Chemother 2010, 54, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- Dudhani, R. V.; Turnidge, J.D.; Nation, R.L.; Li, J. FAUC/MIC Is the Most Predictive Pharmacokinetic/Pharmacodynamic Index of Colistin against Acinetobacter Baumannii in Murine Thigh and Lung Infection Models. Journal of Antimicrobial Chemotherapy 2010, 65, 1984–1990. [Google Scholar] [CrossRef]
- Cheah, S.-E.; Wang, J.; Nguyen, V.T.T.; Turnidge, J.D.; Li, J.; Nation, R.L. New Pharmacokinetic/Pharmacodynamic Studies of Systemically Administered Colistin against Pseudomonas Aeruginosa and Acinetobacter Baumannii in Mouse Thigh and Lung Infection Models: Smaller Response in Lung Infection. Journal of Antimicrobial Chemotherapy 2015, dkv267. [Google Scholar] [CrossRef]
- Tsala, M.; Vourli, S.; Georgiou, P.-C.; Pournaras, S.; Tsakris, A.; Daikos, G.L.; Mouton, J.W.; Meletiadis, J. Exploring Colistin Pharmacodynamics against Klebsiella Pneumoniae: A Need to Revise Current Susceptibility Breakpoints. Journal of Antimicrobial Chemotherapy 2018, 73, 953–961. [Google Scholar] [CrossRef]
- Gautam, V.; Shafiq, N.; Mouton, J.; Malhotra, S.; Kaur, S.; Ray, P. Pharmacokinetics of Colistin in Patients with Multidrug-Resistant Gram-Negative Infections: A Pilot Study. Indian Journal of Medical Research 2018, 147, 407. [Google Scholar] [CrossRef] [PubMed]
- Sorlí, L.; Luque, S.; Li, J.; Campillo, N.; Danés, M.; Montero, M.; Segura, C.; Grau, S.; Horcajada, J.P. Colistin for the Treatment of Urinary Tract Infections Caused by Extremely Drug-Resistant Pseudomonas Aeruginosa: Dose Is Critical. Journal of Infection 2019, 79, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Imberti, R.; Cusato, M.; Villani, P.; Carnevale, L.; Iotti, G.A.; Langer, M.; Regazzi, M. Steady-State Pharmacokinetics and BAL Concentration of Colistin in Critically Ill Patients After IV Colistin Methanesulfonate Administration. Chest 2010, 138, 1333–1339. [Google Scholar] [CrossRef] [PubMed]
- Markantonis, S.L.; Markou, N.; Fousteri, M.; Sakellaridis, N.; Karatzas, S.; Alamanos, I.; Dimopoulou, E.; Baltopoulos, G. Penetration of Colistin into Cerebrospinal Fluid. Antimicrob Agents Chemother 2009, 53, 4907–4910. [Google Scholar] [CrossRef] [PubMed]
- Ozcimen, M.; Ozcimen, S.; Sakarya, Y.; Sakarya, R.; Goktas, S.; Alpfidan, I.; Erdogan, E. Ocular Penetration of Intravenously Administered Colistin in Rabbit Uveitis Model. Journal of Ocular Pharmacology and Therapeutics 2014, 30, 681–685. [Google Scholar] [CrossRef]
- Li, J.; Nation, R.L.; Turnidge, J.D.; Milne, R.W.; Coulthard, K.; Rayner, C.R.; Paterson, D.L. Colistin: The Re-Emerging Antibiotic for Multidrug-Resistant Gram-Negative Bacterial Infections. Lancet Infect Dis 2006, 6, 589–601. [Google Scholar] [CrossRef]
Figure 1.
Colistindrug structure. 2D representation of colistin methanesulfonate molecule highlighted the five methanesulfonate groups (inside the purple dotted circles) responsible for the difference between active compound and its prodrug form. This 2D representation was performed with MolView v2.4 online tool (https://molview.org/).
Figure 1.
Colistindrug structure. 2D representation of colistin methanesulfonate molecule highlighted the five methanesulfonate groups (inside the purple dotted circles) responsible for the difference between active compound and its prodrug form. This 2D representation was performed with MolView v2.4 online tool (https://molview.org/).
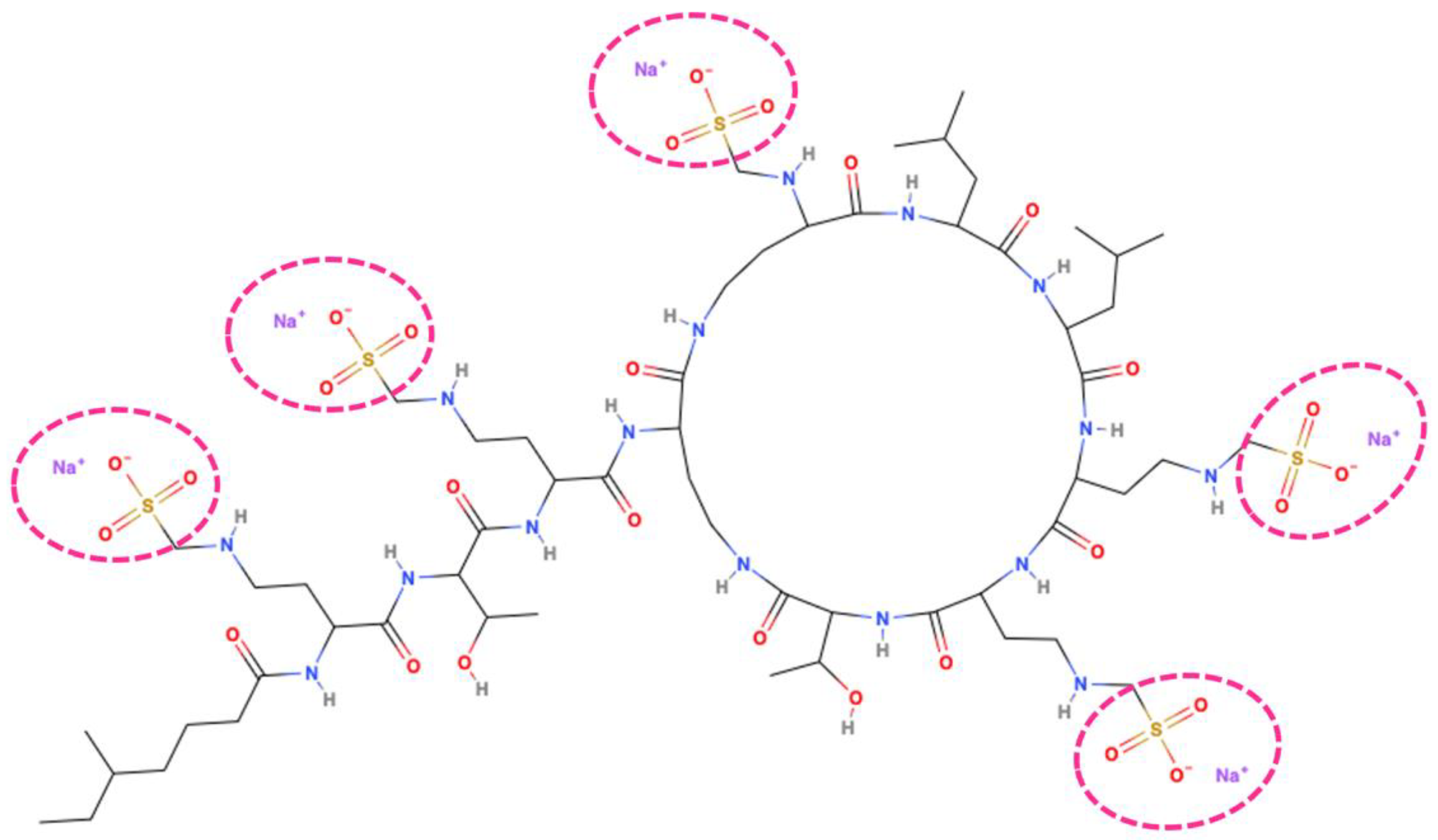

Table 1.
Colistin penetration and assessment of PK/PD target attainment in different sites of infection.
Table 1.
Colistin penetration and assessment of PK/PD target attainment in different sites of infection.
| Site of infection |
Study design | Number of patients | Setting | Dose | Absolute tissue concentrations | Absolute plasmatic concentrations | Penetration rate (AUCtissue/AUCplasma) |
PK/PD target attainment | Ref. |
|---|---|---|---|---|---|---|---|---|---|
| Lung | Prospective observational | 13 | ICU VAP |
2 MU q8h IV | Undetectable | Cmin 1.03±0.69 mg/L AUC/MIC ratio 17.3±9.3 (for MIC=2 mg/L) |
0.00 | Suboptimal in ELF | [126] |
| CSF | Prospective observational | 5 | ICU | 2-3 MU q8h IV | Cmin 0.47 mg/L AUC 0.53 mg*h/L |
Cmin 9.26 mg/L AUC 10.4 mg*h/L |
0.05 | Optimal PK/PD target attainment only for P. aeruginosa and A. baumannii strains exhibiting MIC values up to 0.06 mg/L | [127] |
| Ocular | Preclinical rabbit uveitis model | 20 | Uveitis induced after endotoxin injection | 5 mg/kg IV |
Aqueous humor 0.62±0.07 (at 0.5h) 0.45±0.05 (at 3h) 0.38±0.08 (at 6h) Vitreous humor 0.02±0.01 (at 3h) |
9.84±2.0 (at 0.5h) 0.93±0.07 (at 3h) 0.24±0.08 (at 6h) |
0.07 (aqueous humor at 0.5h) 0.48 (aqueous humor at 3h) 1.58 (aqueous humor at 6h) 0.02 (vitreous humor at 3h) |
Not assessable | [128] |
AUC: area under concentration-to-time curve; Cmin: trough concentrations; CSF: cerebrospinal fluid; ELF: epithelial lining fluid; ICU: intensive care unit; IQR: interquartile range; IV: intravenous; MIC: minimum inhibitory concentration; PK/PD: pharmacokinetic/pharmacodynamic; VAP: ventilator-associated pneumonia.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



