
Submitted:
30 May 2024
Posted:
31 May 2024
You are already at the latest version
Abstract
The mutation in NF2 is the most common alteration associated with meningioma oncogenesis, and it is related to the loss of a suppressing protein called merlin. At the same time, alterations in energy production are visible in cancer cells where increased demand for energy is observed. Fatty acid oxidation could be one of the ways cancer cells obtain energy. This metabolic pathway uses the acylcarnitine shuttle system, which is responsible for the acylation of fatty acids and their transport through mitochondria. Therefore, this study aimed to profile acylcarnitines with short-, medium- and long-acyl chain length in meningiomas to assess their changes in tumors with different NF2 mutation statuses. For the analysis, solid-phase microextraction (SPME) coupled with liquid chromatography high resolution mass spectrometry (LC-HRMS) was used. The presented sampling method enables low invasive and easy collection of the analytes from the studied lesions, which can be crucial for future analysis of potential biomarkers in the surgery room. It was observed that higher levels of these analytes characterized meningiomas with NF2 mutation. Moreover, increased energy consumption and elevated levels of acylcarnitines show that these analytes can be considered as a marker of increased fatty acid oxidation in NF2 mutated cells.
Keywords:
acylcarnitine
; solid-phase microextraction
; meningioma
; merlin
; NF2
1. Introduction
Meningiomas are the most common benign brain tumors. The treatment is based on the surgery; however, the tumor cannot be entirely removed or is inaccessible in some cases. Also, it should be mentioned that some percent of meningiomas can evolve into II or III-grade tumors [1,2,3]. In this case, treatment is much more complex, and alternative therapies like chemotherapy or radiotherapy must be applied [1,2,3]. Therefore, basic research in the direction that enables an understanding of the relation between genetic mutations, their translation to molecular biology, and, subsequently, the impact on mechanisms behind the sudden increase of malignancy of meningiomas or resistance to particular therapy is of great importance [2]. Until now, no genetic biomarker has been used in meningioma diagnosis. Research revealed that mutation in NF2 is the most common alteration associated with meningioma oncogenesis [4]. The protein encoded by this gene is merlin, which regulates cell adhesion and proliferation signaling and impacts tumorigenesis suppression [5]. The lack of merlin correlates with a higher incidence of multiple meningiomas and schwannomas in the central nervous system and a higher risk of developing malignant lesions [3,5].
Acylcarnitines are esters of carnitine and fatty acids. These analytes are direct intermediates of fatty acid oxidation in mitochondria [6,7]. However, the carnitine shuttle system evolved due to the impermeability of the mitochondrial membranes to fatty acids with long acyl chains [6,8]. Specialized enzymes, such as the carnitine palmitoyltransferase 1 (CPT1) and 2 (CPT2), the carnitine-acylcarnitine translocase (CACT), and the carnitine acetyltransferase (CrAT), catalyze the reactions enabling transport of acyl chains of fatty acids through mitochondrial membranes to matrix resulting in energy production as well as acyl chains catabolism [6,9]. Alterations in energy production are visible in cancer cells where increased demand for energy is observed [6]. Moreover, lipid metabolism is upregulated if glucose availability decreases to sustain growth and survival in unfavorable conditions [6]. Thus, the carnitine system plays an essential role in cancer metabolic plasticity. Studies on the carnitine shuttle system are usually related to assessing crucial enzyme activity, but reports on the profiling of direct acylcarnitine intermediates were also proposed [8,10]. However, the bottleneck of this approach is the need to homogenize studied tissue and time-consuming analysis. Therefore, methods that can cope with these problems are needed. Solid phase microextraction (SPME) was one of the methods applied for acylcarnitines analysis in brain tumors [8].
SPME is based on the interaction between sorbent coated on the small size support (ca. 200 µm diameter in the fiber format) and analytes present in the analyzed sample [11]. The probe is introduced into the tissue for a particular time, after which it is stored or proceeded to the next step: desorption of analytes into organic solution. Subsequently, samples can be analyzed using chromatography coupled with a mass spectrometer or other analytical instrumentation.
The current study aimed to apply SPME sampling for acylcarnitine profiling of meningiomas and to test in greater detail if the acylcarnitine profile changes along with the different NF2 mutation statuses.
2. Materials and Methods
2.1. Chemicals and Materials
External calibrant Pierce LTQ Velos ESI Positive Ion Calibration Solution was purchased from Thermo Scientific. Isopropanol, methanol, water, acetonitrile, and ammonium acetate were LC-MS grade and were purchased from Merck (Warsaw, Poland). SPME C18 fibers were kindly provided by Supelco (Bellefonte, PA, USA).
2.2. Biological Material
Brain tumors were obtained during neurosurgical procedures in the 10th Military Research Hospital and Polyclinic in Bydgoszcz. SPME sampling was conducted directly after tumor removal. Meningothelial meningiomas without mutations in AKT1, PIK3CA, TRAF, and KLF4 were selected for this study: 22 tumors with a mutation in NF2 (NF2mt) and 18 samples without this genetic alteration (NF2wt).
2.3. Genetic Testing
The genetic testing of mutations in the following genes: NF2, AKT1, PIK3CA (rs104886003, rs121913273, and rs121913279), TRAF (N520C, R653Q, R641C, and K615E), and KLF4 was carried out. Analysis was performed using polymerase chain reaction (PCR), multiplex ligation-dependent probe amplification (MLPA), and real-time quantitative PCR (RQ-PCR). Tests were proceeded according to the manufacturer’s protocol.
2.4. Chemical Biopsy (Solid-Phase Microextraction) Protocol and LC-HRMS Analysis
Solid-phase microextraction probes coated with 7mm C18 sorbent were used to sample brain tumors removed during the neurosurgical procedure. The fibers were preconditioned overnight in methanol: water (1:1 v/v) solution, and then directly before sampling, they were rinsed in water. Subsequently, the probe was inserted into the tissue for 30 minutes (extraction), and after this time, it was rinsed briefly in water. Probes were stored in a freezer at −30 °C until instrumental analysis. Then, the fibers were desorbed into 150μl of isopropanol: methanol (1:1 v/v) solution using silanized inserts. Desorption was conducted for 1 hour under agitation at 850 rpm [12]. Pooled quality control (QC) and extraction blanks were also prepared [12].
The liquid chromatography-high resolution mass spectrometry (LC–HRMS) platform consisted of a Dionex UltiMate 3000 RS autosampler, a Dionex Ultimate 3000 RS pump (Thermo Fisher Scientific, Dionex, Bremen, Germany), and a Q Exactive Focus high-resolution mass spectrometer (Thermo Fisher Scientific, Bremen, Germany) was used for instrumental analysis.
LC analysis was conducted using 5 mM ammonium acetate in water as phase A and acetonitrile as phase B. Column: SeQuantZIC-cHILIC (3 μm 100 × 2.1 mm) was used, and the injection volume was set at 10 μL. The hhydrophilic interaction chromatography (HILIC) was used in the analysis. The detailed parameters were given elsewhere [8]. The study was conducted in positive ion mode in a 100–1000 m/z scan range. Acylcarnitines were identified by matching their fragmentation patterns with spectra libraries at a mass accuracy of <3 ppm (the presence of characteristic m/z: 85.0290 in MS/MS spectra). Full MS/dd-MS2 discovery mode was used for this purpose, and the detailed parameters of the fragmentation protocol were given elsewhere [8].
2.5. Data Processing and Statistical Analysis
Acylcarnitine identification was performed using XCalibur software (Thermo Fisher Scientific, San Jose, CA, USA) based on m/z and characteristic fragmentation pattern. Statistical analysis was conducted using MetaboAnalyst 6.0 [13]. Chemometric analysis and box-plot visualization were prepared. The average peak area, coefficient of variation, and the ratio of compared study groups for all analytes were calculated; the Mann–Whitney U Test was applied to compare the variables. The p-value lower than 0.05 was set as a statistical significance threshold. Apart from analyzing acylcarnitine levels without normalization, normalization on the summary peak area was also analyzed.
Finally, ChlorTox was calculated along with the recommendation given by Nowak et al. [14].
3. Results and Discussion
A set of several acylcarnitines was extracted from brain tumors using SPME fibers, and a relative standard deviation (RSD) was below 30%. Among detected analytes were short-chain acylcarnitines (SCAC): AC C2:0, AC C3:0, AC C4:0 and AC C5:0, medium-chain acylcarnitines (MCAC): AC C6:0, AC C8:0, AC C10:0, AC C10:1, AC C12:0, and long-chain acylcarnitines (LCAC): AC C14:0, AC C14:1, AC C16:0, AC C16:1, AC C18:0, AC C18:1 (Table 1). A similar set of analytes was extracted in a study on acylcarnitine profiling in gliomas, where SPME probes were also used for sampling [8].
Firstly, a chemometric analysis using a principal component analysis (PCA) was conducted. Visualization of meningioma samples showed that NF mutant tumors were more dispersed in the plot than NFwt samples, which created a more concentrated group (Figure 1). The lack of merlin, a suppressor and microtubule stabilizer protein in NFmt meningiomas, could be related to more heterogenous metabolism and energy demands, impacting the acylcarnitine profile. The presence of NF-2 mutation can relate to a higher possibility of developing new foci of cancer origin or lead to tumor transformation or the occurrence of multiple tumors [15].
A trend of higher acylcarnitine levels in the samples lacking merlin (NF2mt) than in the wildtype was observed. This observation corresponds to ratios of peak areas in NF2 mutant to NF2 wildtype meningiomas in Table 1. Nevertheless, significantly different levels of acylcarnitines were observed for ten of them (p<0.05) (Figure 2). The lack of merlin, can be related to higher malignancy and increased energy consumption, which can be observed in cancerous cells [16]. This demand could be fulfilled by changing glucose metabolism into aerobic glycolysis– Warburg effect, elevated glutamine metabolism, or changes in fatty acid oxidation [5,6,17]. Indeed, it was reported in the literature that NF2 mutant cells are characterized by higher dependence on lipid metabolism [5]. Stepanova et al. observed that cells with NF mutation have significantly higher fatty acid synthesis, as described by the higher activity of FASN, ACC 1 and 2, and other enzymes in this metabolic pathway [5]. The authors also suspected some fatty acid oxidation alteration could be observed in samples lacking merlin. The explanation of this observation could be related to the phenomena described by Melone et al. as a “futile cycle” where two metabolic cycles going in opposite directions could be used by cancer cells. Fatty acid biosynthesis supplies appropriate fatty acid levels, while their oxidation in mitochondria provides energy for proliferating cells [6]. Thus, elevated levels of acylcarnitines could explain increased energy consumption and elevated levels of acylcarnitines as markers of increased fatty acid oxidation in NF2 mutated cells. Thus, increased fatty acid oxidation is related to an enhanced carnitine shuttle system, which produces higher acylcarnitine levels, as observed in this study. It should be mentioned herein that higher levels of acylcarnitines were observed in various types of cancerous lesion glioma and breast cancer hepatocellular carcinoma in comparison to respective non-cancerous samples [8,10,18,19,20].
A summary peak area normalization was applied to assess the relative changes between acylcarnitines inside the profile. This normalization method (also known as total peak area or constant sum normalization) normalizes the peak areas to total peak area, the sum of the areas of all detected peaks in a studied sample [21]. This type of normalization is often used in metabolomics, but it should be noted that it converts analytes into a fraction of the whole profile [21,22,23]. Thus, an increase or decrease of one analyte will be related to the opposite change of different analyte/analytes from the normalized profile. In the data presented herein, decreased levels of AC C16:0, AC C18:1, and AC C18:2 were observed in the NF mutant samples compared to wildtype (p<0.05, FDR > 0.05) (Table 1). This observation indicates that the mutation in NF2 is related to the impairment in fatty acid oxidation. The possible alteration could be related to CPT-2 activity. This enzyme catalyzes the transfer of the acyl chain from long and very-long-chain acylcarnitines to coenzyme A[6]. The activity of CPT-2 can be estimated using the formula based on the concentration of the following acylcarnitines: (AC C16:0 + AC C18:1)/ AC C2:0 [24]. It indicates that the activity of CPT-2 is directly proportional to the levels of AC C16:0 and AC C 18:1 and is inversely proportional to the concentration of AC C2:0. Thus, decreased levels of long acylcarnitines, such as AC C16:0, AC C18:1, and AC C18:2 with a simultaneous increase short acylcarnitines level, such as AC C2:0 suggest downregulation of CPT-2 activity. However, it should be noted that AC C2:0 upregulation was statistically insignificant (p>0.05) (Table 1). The observed alterations may be related to changes in the activity of enzymes engaged in the carnitine shuttle system or an increase in the levels of short acyl chain acylcarnitines, whose transport is independent of the carnitine shuttle system enzymes. Downregulation of CPT-2 activity were observed in different types of cancers, such as ovarian cancer, renal cancer, hepatocellular carcinoma, and colorectal cancer[25,26,27,28]. There are also reports that patients with obesity are more prone to cancer development due to the downregulation of CPT-2 [29].
A similar SPME analysis coupled with LC-HRMS and total peak area normalization was applied to analyze the acylcarnitine profile in gliomas with different grades, IDH1/2 mutations, and 1p/19q codeletion status [30]. Tumors with worse prognosis had significantly higher levels of a few acylcarnitines. The AC C16:0, AC C18:1, and AC C18:2 were downregulated in high-grade gliomas. It was also observed that IDH1/2 wild-type tumors were generally characterized by higher acylcarnitine levels, which corresponds to a worse prognosis for the patient[8]. However, AC C16:0, AC C18:1, and AC C18:2 were slightly downregulated in wild-type samples, similar to the research presented herein – lower levels of mentioned analytes in samples with a higher proliferation risk. In the case of 1p/19q, the samples without co-deletion had slightly lower or almost equal levels of the mentioned normalized acylcarnitines as in tumors with co-deletion, which can be connected with worse response to radiotherapy patients without 1p/19q codeletion [8].
Results presented herein show changes in acylcarnitine levels in meningioma with different mutation NF2 status. However, enriching this research with the assessment of carnitine shuttle enzyme activity and the expression of genes responsible for their production would be beneficial. It would help to select the most important acylcarnitines in cancer diagnosis, especially considering LCAC alteration explained in the literature by the downregulation of CPT-2. Moreover, a method enabling fast and quantitative analysis should be introduced. The chromatographic analysis takes about half an hour to analyze one sample. Moreover, this time is even longer if sample preparation is counted. Thus, to increase the chances of clinical use of acylcarnitine analysis in meningioma diagnosis, it would be useful to optimize the method, enabling fast, quantitative, and reliable analysis of potential biomarkers.
An additional advantage would be low invasiveness, as represented by the methods based on SPME. Therefore, technology, such as coated blade spray mass spectrometry (CBS) or microfluidic open interface (MOI), could be applied [31]. The CBS sampling is conducted with the probe in the shape of a sword coated with the sorbent at the tip. Then, the blade is mounted in the interface installed in the ion source. Subsequently, the drop of desorption solvent is added to the surface of the probe, and high voltage is applied. Results could be acquired in a few seconds. CBS was tested for carnitine analysis in glioma homogenate [32]. Another solution could be microfluidic open ion source MOI mass spectrometry based on the coated fiber sampling [33]. However, instead of desorption followed by instrumental analysis, the probe is put to the interface installed on the mass spectrometer. The interface consists of a chamber filled with desorption solvent. The probe is introduced to this chamber for a few seconds, during which desorption is conducted. Then, the solution with desorbed analytes is directly injected into the mass spectrometer. The combination of desorption and instrumental analysis allows a reduction of analysis time.
As important as low invasiveness and the possibility of rapid analysis, introducing methods harmless to the environment can be crucial. Therefore, the objective factor, such as the ChlorTox, was calculated for the studied analytical platform. This parameter enables estimation of substance toxicity in comparison to the standard substance – chloroform [34]. It was shown that ChlorTox for solvents used for instrumental analysis is comparable with other methods presented in the literature[14]. Due to the wide application of homogenization followed by liquid-liquid extraction (LLE) in tissue analysis, the liquid-liquid extraction coupled with high-performance chromatography (LLE/HPLC) method given by Nowak et al. [14,31] was used as a reference. For instance, the ChlorTox for HPLC analysis was 3.36g, while in the results presented herein, it was 3.87g (Table 2) [14]. On the other hand, ChlorTox for sample preparation protocol was significantly different. The ChlorTox was 2.78 for the LLE-HPC method compared to 0.13 for the SPME method [14]. This observation shows that SPME as a sample preparation method is more environment-friendly than LLE. It should be noted that ChlorTox per sample was calculated based on the number of studied samples, blanks, and QC samples. Data could also be biased due to limited information on the analytical methods for LLE/HPLC.
4. Conclusions
Application of SPME enabled simple profiling of a wide range of acylcarnitines in meningiomas and showed that the presence of NF2 could alter acylcarnitine profile. The loss of merlin coded by NF2 was related to a higher heterogeneity of acylcarnitine profile and increased levels of detected carnitine esters. These results suggest that alteration in the acylcarnitine system could be crucial in assessing energy usage in cancerous cells and could be potential biomarkers of neoplastic changes. Moreover, the application of SPME as a sampling and sample preparation method opens new possibilities for future application but also enables the reduction of toxicity in the environment in comparison with usually used methods, such as LLE.
Author Contributions
Conceptualization: J.B., M.H. and B.B.; Methodology: J.B.; Investigation: J.B., K.S., A.M., A.R., Resources: J.F., M.B., M.H. and B.B.; Data curation: J.B, Writing, original draft preparation: J.B.; Writing, review, and editing: B.B.; Visualization: J.B.; Supervision: B.B.; Project administration: J.B. and B.B.; Funding acquisition: J.B. and B.B. All authors have read and agreed to the published version of the manuscript.
Funding
Genetic tests were funded by the National Science Centre Poland within research grant No. 2019/33/N/ST4/00286. The National Science Centre Poland supported acylcarnitine profiling within research grant No. 2015/18/M/ST4/00059.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Bioethical Committee in Bydgoszcz (KB 628/2015).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Acknowledgments
The authors acknowledge Supelco/MilliporeSigma for kindly supplying the SPME probes. The authors would like to thank Magdalena Gaca-Tabaszewska and Paulina Szeliska for their technical support in the laboratory. The authors would like to acknowledge Paulina Zofia Goryńska, Krzysztof Goryński and Karol Jaroch for their help with sampling the first cases.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Goldbrunner, R.; Minniti, G.; Preusser, M.; Jenkinson, M.D.; Sallabanda, K.; Houdart, E.; von Deimling, A.; Stavrinou, P.; Lefranc, F.; Lund-Johansen, M.; et al. EANO guidelines for the diagnosis and treatment of meningiomas. Lancet Oncol. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Nowosielski, M.; Galldiks, N.; Iglseder, S.; Kickingereder, P.; Von Deimling, A.; Bendszus, M.; Wick, W.; Sahm, F. Diagnostic challenges in meningioma. Neuro. Oncol. 2017, 19. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Bi, W.L.; Dunn, I.F. Medical management of meningioma in the era of precision medicine. Neurosurg. Focus 2018, 44. [Google Scholar] [CrossRef]
- Ghalavand, M.A.; Asghari, A.; Farhadi, M.; Taghizadeh-Hesary, F.; Garshasbi, M.; Falah, M. The genetic landscape and possible therapeutics of neurofibromatosis type 2. Cancer Cell Int. 2023, 23. [Google Scholar] [CrossRef] [PubMed]
- Stepanova, D.S.; Semenova, G.; Kuo, Y.M.; Andrews, A.J.; Ammoun, S.; Hanemann, C.O.; Chernoff, J. An essential role for the tumor-suppressor merlin in regulating fatty acid synthesis. Cancer Res. 2017, 77. [Google Scholar] [CrossRef]
- Melone, M.A.B.; Valentino, A.; Margarucci, S.; Galderisi, U.; Giordano, A.; Peluso, G. The carnitine system and cancer metabolic plasticity review-article. Cell Death Dis. 2018, 9, 228. [Google Scholar] [CrossRef]
- McCoin, C.S.; Knotts, T.A.; Adams, S.H. Acylcarnitines-old actors auditioning for new roles in metabolic physiology. Nat. Rev. Endocrinol. 2015, 11. [Google Scholar] [CrossRef]
- Bogusiewicz, J.; Burlikowska, K.; Jaroch, K.; Gorynska, P.Z.; Gorynski, K.; Birski, M.; Furtak, J.; Paczkowski, D.; Harat, M.; Bojko, B. Profiling of carnitine shuttle system intermediates in gliomas using solid-phase microextraction (SPME). Molecules 2021, 26, 6112. [Google Scholar] [CrossRef]
- Kant, S.; Kesarwani, P.; Prabhu, A.; Graham, S.F.; Buelow, K.L.; Nakano, I.; Chinnaiyan, P. Enhanced fatty acid oxidation provides glioblastoma cells metabolic plasticity to accommodate to its dynamic nutrient microenvironment. Cell Death Dis. 2020, 11. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, X.; Zhang, Y.; Zhang, K.; Zhan, C.; Shi, X.; Li, Y.; Zhao, J.; Bai, Y.; Wang, Y.; et al. Metabolic profiling analysis upon acylcarnitines in tissues of hepatocellular carcinoma revealed the inhibited carnitine shuttle system caused by the downregulated carnitine palmitoyltransferase 2. Mol. Carcinog. 2019, 58, 749–759. [Google Scholar] [CrossRef]
- Reyes-Garcés, N.; Gionfriddo, E.; Gómez-Ríos, G.A.; Alam, M.N.; Boyacı, E.; Bojko, B.; Singh, V.; Grandy, J.; Pawliszyn, J. Advances in Solid Phase Microextraction and Perspective on Future Directions. Anal. Chem. 2018, 90, 302–360. [Google Scholar] [CrossRef] [PubMed]
- Bogusiewicz, J.; Kupcewicz, B.; Goryńska, P.Z.; Jaroch, K.; Goryński, K.; Birski, M.; Furtak, J.; Paczkowski, D.; Harat, M.; Bojko, B. Investigating the Potential Use of Chemical Biopsy Devices to Characterize Brain Tumor Lipidomes. Int. J. Mol. Sci. 2022, 23. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Zhou, G.; Ewald, J.; Chang, L.; Hacariz, O.; Basu, N.; Xia, J. Using MetaboAnalyst 5.0 for LC–HRMS spectra processing, multi-omics integration and covariate adjustment of global metabolomics data. Nat. Protoc. 2022, 17. [Google Scholar] [CrossRef] [PubMed]
- Nowak, P.M.; Wietecha-Posłuszny, R.; Płotka-Wasylka, J.; Tobiszewski, M. How to evaluate methods used in chemical laboratories in terms of the total chemical risk? – a ChlorTox Scale. Green Anal. Chem. 2023, 5. [Google Scholar] [CrossRef]
- Petrilli, A.M.; Fernández-Valle, C. Role of merlin/NF2 inactivation in tumor biology. Oncogene 2016, 35. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Karas, P.J.; Hadley, C.C.; Bayley, V.J.C.; Basit Khan, A.; Jalali, A.; Sweeney, A.D.; Klisch, T.J.; Patel, A.J. The role of merlin/NF2 loss in meningioma biology. Cancers (Basel) 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Zhu, X.; Wu, Y. Effects of Glucose Metabolism, Lipid Metabolism, and Glutamine Metabolism on Tumor Microenvironment and Clinical Implications. Biomolecules 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Zoni, E.; Minoli, M.; Bovet, C.; Wehrhan, A.; Piscuoglio, S.; Ng, C.K.Y.; Gray, P.C.; Spahn, M.; Thalmann, G.N.; Kruithof-De Julio, M. Preoperative plasma fatty acid metabolites inform risk of prostate cancer progression and may be used for personalized patient stratification. BMC Cancer 2019, 19, 1–18. [Google Scholar] [CrossRef]
- Yaligar, J.; Teoh, W.W.; Othman, R.; Verma, S.K.; Phang, B.H.; Lee, S.S.; Wang, W.W.; Toh, H.C.; Gopalan, V.; Sabapathy, K.; et al. Longitudinal metabolic imaging of hepatocellular carcinoma in transgenic mouse models identifies acylcarnitine as a potential biomarker for early detection. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Yu, D.; Xuan, Q.; Zhang, C.; Hu, C.; Li, Y.; Zhao, X.; Liu, S.; Ren, F.; Zhang, Y.; Zhou, L.; et al. Metabolic Alterations Related to Glioma Grading Based on Metabolomics and Lipidomics Analyses. Metabolites 2020, 10, 478. [Google Scholar] [CrossRef]
- Nam, S.L.; Paulina de la Mata, A.; Dias, R.P.; Harynuk, J.J. Towards standardization of data normalization strategies to improve urinary metabolomics studies by gc×gc-tofms. Metabolites 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Xia, Y. Pretreating and normalizing metabolomics data for statistical analysis. Genes Dis. 2024, 11. [Google Scholar] [CrossRef]
- Nam, S.L.; Giebelhaus, R.T.; Tarazona Carrillo, K.S.; de la Mata, A.P.; Harynuk, J.J. Evaluation of normalization strategies for GC-based metabolomics. Metabolomics 2024, 20. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Zheng, X.; Yang, M.; Zhao, A.; Li, M.; Chen, T.; Panee, J.; Jia, W.; Ji, G. Serum lipid alterations identified in chronic hepatitis B, hepatitis B virus-associated cirrhosis and carcinoma patients. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, Z.; Liu, S.; Li, J.; Wu, L.; Lv, X.; Xu, J.; Chen, B.; Zhao, S.; Yang, H. CPT2 down-regulation promotes tumor growth and metastasis through inducing ROS/NFκB pathway in ovarian cancer. Transl. Oncol. 2021, 14. [Google Scholar] [CrossRef]
- Zeng, K.; Li, Q.; Song, G.; Chen, B.; Luo, M.; Miao, J.; Liu, B. CPT2-mediated fatty acid oxidation inhibits tumorigenesis and enhances sorafenib sensitivity via the ROS/PPARγ/NF-κB pathway in clear cell renal cell carcinoma. Cell. Signal. 2023, 110. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Li, X.; Yan, H.; Wu, J.; Yang, Y.; He, J.; Chen, J.; Jiang, Z.; Wu, F.; Jiang, Z. Downregulation of CPT2 promotes proliferation and inhibits apoptosis through p53 pathway in colorectal cancer. Cell. Signal. 2022, 92. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Lv, D.; Zheng, Y.; Wu, M.; Xu, C.; Zhang, Q.; Wu, L. Downregulation of CPT2 promotes tumorigenesis and chemoresistance to cisplatin in hepatocellular carcinoma. Onco. Targets. Ther. 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, N.; Nakagawa, H.; Enooku, K.; Kudo, Y.; Hayata, Y.; Nakatsuka, T.; Tanaka, Y.; Tateishi, R.; Hikiba, Y.; Misumi, K.; et al. CPT2 downregulation adapts HCC to lipid-rich environment and promotes carcinogenesis via acylcarnitine accumulation in obesity. Gut 2018, 67. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: a summary. Neuro. Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef]
- Bogusiewicz, J.; Bojko, B. Insight into new opportunities in intra-surgical diagnostics of brain tumors. TrAC Trends Anal. Chem. 2023, 162, 117043. [Google Scholar] [CrossRef]
- Bogusiewicz, J.; Gaca-Tabaszewska, M.; Olszówka, D.; Jaroch, K.; Furtak, J.; Harat, M.; Pawliszyn, J.; Bojko, B. Coated Blade Spray-Mass Spectrometry as a New Approach for the Rapid Characterization of Brain Tumors. Molecules 2022, 27, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Tascon, M.; Alam, M.N.; Gómez-Ríos, G.A.; Pawliszyn, J. Development of a Microfluidic Open Interface with Flow Isolated Desorption Volume for the Direct Coupling of SPME Devices to Mass Spectrometry. Anal. Chem. 2018, 90. [Google Scholar] [CrossRef] [PubMed]
- Nowak, P.M.; Bis, A.; Zima, A. ChlorTox Base – a useful source of information on popular reagents in terms of chemical hazards and greenness assessment. Green Anal. Chem. 2023, 6. [Google Scholar] [CrossRef]
Figure 1.
The PCA presents a visualization of NFmt and NFwt meningiomas based on an acylcarnitine profile. A. The PCA with pooled QC, B. The PCA without pooled QC. QC- pooled quality control (red dots), NF2mt – NF2 mutated (grey dots), NF2wt – NF2 wildtype (cyan dots).
Figure 1.
The PCA presents a visualization of NFmt and NFwt meningiomas based on an acylcarnitine profile. A. The PCA with pooled QC, B. The PCA without pooled QC. QC- pooled quality control (red dots), NF2mt – NF2 mutated (grey dots), NF2wt – NF2 wildtype (cyan dots).
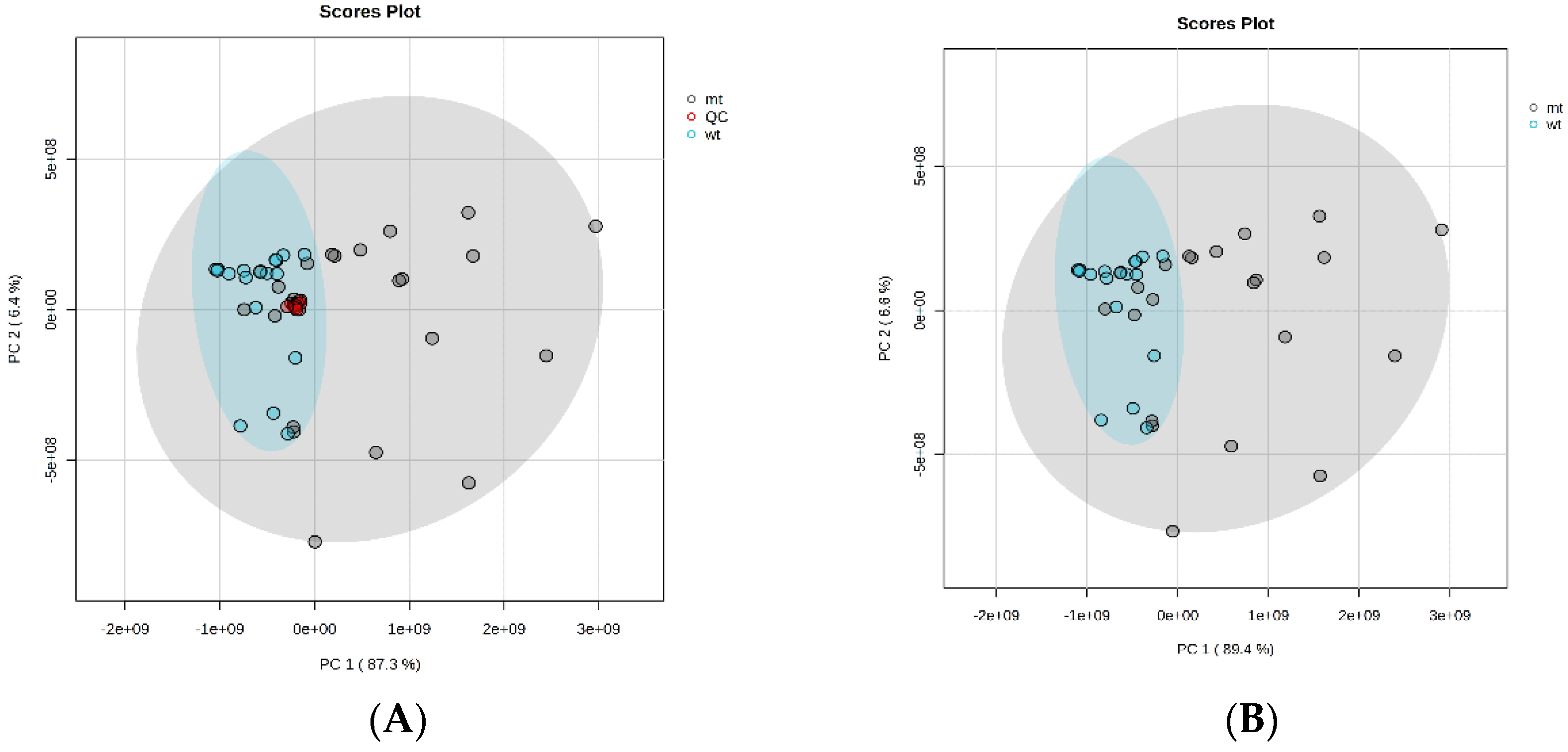
Figure 2.
Box-plots representing levels of significantly altered acylcarnitines in NF2 mutated and NF2 wildtype meningiomas analyzed using SPME coupled with LC-HRMS. Plots for raw data are given. NF2mt – NF2 mutated (grey boxes), NF2wt – NF2 wildtype (cyan boxes).
Figure 2.
Box-plots representing levels of significantly altered acylcarnitines in NF2 mutated and NF2 wildtype meningiomas analyzed using SPME coupled with LC-HRMS. Plots for raw data are given. NF2mt – NF2 mutated (grey boxes), NF2wt – NF2 wildtype (cyan boxes).
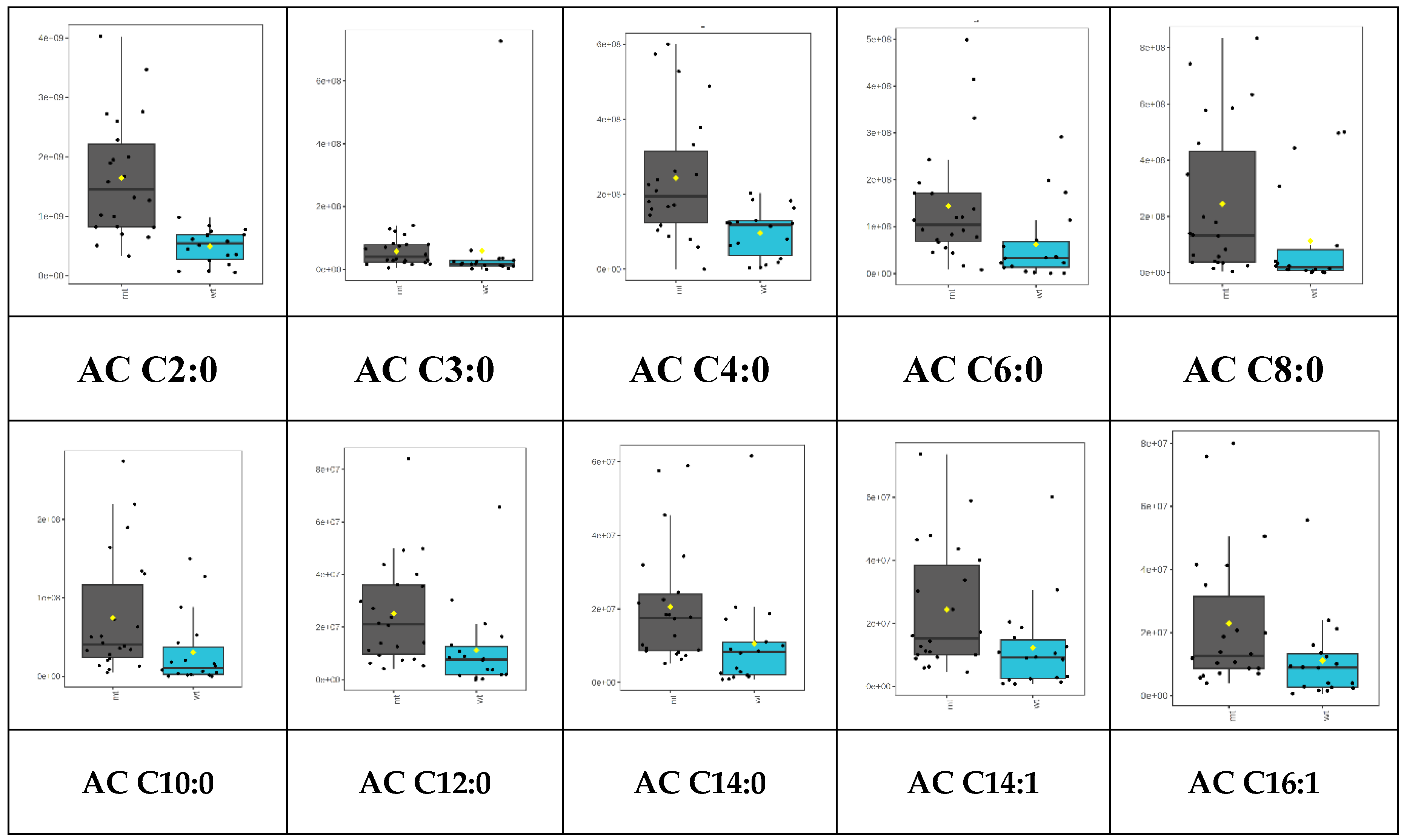

Table 1.
The ratio of meningiomas with NF2 mutation to the tumor without this mutation was assessed using the SPME-LC-HRMS platform. AC – acylcarnitine, FDR – False discovery rate, LCAC – long-chain acylcarnitine, MCAC – medium-chain acylcarnitine, NF2mt – NF2 mutated, NF2wt – NF2 wildtype, SCAC – short-chain acylcarnitines, RT – retention time.
Table 1.
The ratio of meningiomas with NF2 mutation to the tumor without this mutation was assessed using the SPME-LC-HRMS platform. AC – acylcarnitine, FDR – False discovery rate, LCAC – long-chain acylcarnitine, MCAC – medium-chain acylcarnitine, NF2mt – NF2 mutated, NF2wt – NF2 wildtype, SCAC – short-chain acylcarnitines, RT – retention time.
| Acylcarnitine | m/z | RT | Raw data | Normalized data | |||||
|---|---|---|---|---|---|---|---|---|---|
| NF2mt/ NF2wt ratio | p-value | FDR | NF2mt/ NF2wt ratio | p-value | FDR | ||||
| SCAC | AC C2:0 | 204.1230 | 13.49 | 3.33 | <0.05 | <0.05 | 1.19 | 0.095 | 0.300 |
| AC C3:0 | 218.1387 | 11.94 | 2.50 | <0.05 | <0.05 | 0.51 | 0.737 | 0.789 | |
| AC C4:0 | 232.1543 | 10.70 | 2.50 | <0.05 | <0.05 | 0.93 | 0.527 | 0.719 | |
| AC C5:0 | 246.1700 | 9.92 | 2.28 | 0.206 | 0.219 | 0.77 | 0.100 | 0.300 | |
| MCAC | AC C6:0 | 260.1856 | 9.28 | 2.31 | <0.05 | <0.05 | 1.15 | 0.251 | 0.470 |
| AC C8:0 | 288.2169 | 8.60 | 2.16 | <0.05 | <0.05 | 1.22 | 0.155 | 0.388 | |
| AC C10:0 | 316.2484 | 8.24 | 2.41 | <0.05 | <0.05 | 1.29 | 0.219 | 0.468 | |
| AC C10:1 | 314.2326 | 8.29 | 1.77 | 0.066 | 0.066 | 0.69 | 0.946 | 0.989 | |
| AC C12:0 | 344.2796 | 7.95 | 2.21 | <0.05 | <0.05 | 1.21 | 0.657 | 0.758 | |
| LCAC | AC C14:0 | 372.3108 | 7.75 | 1.94 | <0.05 | <0.05 | 1.02 | 0.657 | 0.758 |
| AC C14:1 | 370.2952 | 7.73 | 2.00 | <0.05 | <0.05 | 1.07 | 0.459 | 0.688 | |
| AC C16:0 | 400.3423 | 7.63 | 1.63 | 0.055 | 0.074 | 0.60 | <0.05 | 0.196 | |
| AC C16:1 | 398.3266 | 7.65 | 2.06 | <0.05 | <0.05 | 1.08 | 0.367 | 0.611 | |
| AC C18:0 | 428.3734 | 7.63 | 1.11 | 0.219 | 0.219 | 0.42 | <0.05 | 0.176 | |
| AC C18:1 | 426.3579 | 7.49 | 1.68 | 0.119 | 0.137 | 0.69 | <0.05 | 0.178 | |
Table 2.
The calculation of the hazards of the SPME-LC-MS method in acylcarnitine analysis using HILIC chromatography and high-resolution mass spectrometry per sample.
Table 2.
The calculation of the hazards of the SPME-LC-MS method in acylcarnitine analysis using HILIC chromatography and high-resolution mass spectrometry per sample.
| Analysis step | Reagents | CAS | CHsub | ChlorTox [g] | Total ChlorTox [g] |
|---|---|---|---|---|---|
| SPME | Methanol | 67-56-1 | 4.81 | 0.08 | 0.13 |
| Isopropanol | 67-63-0 | 3.13 | 0.05 | ||
| Instrumental Analysis | Ammonium acetate | 631-61-8 | 0.00 | 0.00 | 3.87 |
| Acetonitrile | 75-05-8 | 2.25 | 3.87 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



