Submitted:
30 May 2024
Posted:
31 May 2024
You are already at the latest version
Abstract
Current sampling methods for detecting the presence of the invasive freshwater hydrozoan Craspedacusta sowerbii rely mainly on visual confirmation of the medusa stage. Confirming the presence of the polyp stage is equally important to observing medusae since typical late summer/early fall occurrences or observations of medusae are sporadic though becoming more frequent. The polyp stage is important as it is the organism’s primary stage and is present throughout the year depending on water temperatures. Therefore, sampling methods for the polyp stage are commonly the collection of substrates such as rocks, plants or pieces of wood in a given body of water and can be cumbersome to examine. Based on preliminary culturing of the polyp stage on glass and plastic microscope slides in the lab, we designed a novel sampling methodology based on submerging 4 substrate types (small glass petri dishes, Hester-Dendy plates, plastic and glass microscope slides) to confirm the presence of C. sowerbii polyps in the field. We tested this method in various lakes in the Illinois-Indiana region (USA). The sampling method we designed was effective in that C. sowerbii polyps (and sometimes colonies) were found on both plastic and glass slides. While this method can be sufficient for detection of the polyp stage, it also shows significant potential for improvement; we highlight water flow and competition and predation from other organisms as significant factors influencing the collection of C. sowerbii polyps for future methodologies.
Keywords:
Invasive hydrozoan
; sampling
; polyp
; frustule
1. Introduction
Invasive species have become more prevalent across the globe due to rising temperatures and human activity; these factors increase the available range of invasive species and allow them to reach more places, respectively [1,2,3]. While total elimination of an invasive species is often impossible, the most successful efforts to curb the spread of certain invasive species has been partly due to increased awareness and knowledge of the species in question [4,5,6,7]. Since the spread of invasive species poses a major threat to global biodiversity, it is critical to understand the impact of invasive species to develop control protocols as needed [8,9,10].
Aquatic ecosystems are especially vulnerable to invasive species given increases in aquatic temperatures and competition with native species [11,12,13]. More specifically, invasive cnidaria have already done recorded damage to marine ecosystems [14,15,16,17]. However, much less work has been done on freshwater cnidaria despite the fact that they may pose just as much of a threat as their marine counterparts [18,19]. While many species of invasive cnidaria exist in freshwater and marine environments, we focus specifically on the hydrozoan Craspedacusta sowerbii Lankester, 1880. Craspedacusta sowerbii, commonly known as the peach blossom jellyfish, originates from the Yangtze River in China [20,21] and has since spread to every continent except Antarctica [22,23,24,25,26]. In the United States, C. sowerbii has been reported in 44 out of 50 states across a range of freshwater ecosystems [27].
The life cycle of Craspedacusta sowerbii consists of an asexual portion and a sexual portion [28,29] (Figure 1); both portions start with sessile colonies of polyps, the most predominant form of C. sowerbii [30,31]. In the asexual portion, polyp colonies bud rod-like structures known as frustules, which then transform into a new polyp colony [32,33]. The sexual portion, on the other hand, involves polyp colonies budding motile medusae, which mature and produce gonads [19]; much like other cnidaria, these gonads combine to form a planula larva, which develops into a new polyp colony. These medusae are the larger stage of the life cycle of C. sowerbii and are most often the form identified when confirming the presence of C. sowerbii in a given body of water.
Sampling of Craspedacusta sowerbii has mainly been done on the medusa stage. This is for a variety of reasons; the medusae are larger and easier to see, so they can be easily acquired in several different ways, including scooping them directly out of the water with a jar or spoon [22,34,35,36,37], or towing a plankton net [18,24,37,38,39,40,41,42]. However, medusa sightings alone cannot confirm the establishment of C. sowerbii in a given environment [43]. Medusa appearances are sporadic [31] and can be influenced by a variety of environmental factors. We are aware of reports citing their appearance yearly at certain sites, while other sites can go many years without their appearance. Additionally, Duggan and Eastwood [43] report finding C. sowerbii polyp colonies in bodies of water where the medusae had not been previously sighted, as well as finding no polyp colonies in bodies where the medusae had been sighted. As such, Duggan and Eastwood [43] concluded that medusa sightings alone are not sufficient to establish the presence of C. sowerbii in a body of water and highlight the importance of determining the presence of polyp colonies. With rising temperatures, we might expect more appearances of the medusa stage [1], but medusa sightings cannot be guaranteed even if the polyp stage is confirmed to exist in a given environment. As such, reliable detection of the presence of C. sowerbii depends on confirmation of the polyp stage.
Polyp sampling poses several difficulties, however: namely, the polyp colonies are sessile and must be sampled by pulling substrates out of the water. As such, current sampling methods mainly involve removing natural substrates (including rocks, plants, and wood) from the water and examining them under a dissecting microscope [22,43,44,45]. Polyp colonies are also transparent and can be hard to detect, especially with untrained eyes [44]. Few studies rigorously sample to obtain Craspedacusta sowerbii polyp colonies, and far less work has been done to collect and conduct experiments with the colony and frustule stage of C. sowerbii compared to the medusa stage [33,43,46].
Due to the difficulty and importance of sampling the polyp stage of Craspedacusta sowerbii, more insight into the efficacy of sampling is warranted. However, to our knowledge, there exists no systematic method to sample for C. sowerbii polyps in the field. As such, we sought to create a methodical and effective sampling method and provide quantitative data on sampling efficacy for use in future sampling and research.
2. Materials and Methods
Our work aims to build on previous difficulties involved with polyp stage sampling; we constructed a methodical protocol to sample for the polyp stage of Craspedacusta sowerbii that allows for ease of deployment, retrieval, and subsequent analysis.
2.1. Laboratory Pilot Substrate Study
While various literature had described Craspedacusta sowerbii polyp colonies grown in glass [28] and plastic [47], we found no comparison between the two on its effects on the growth and spread of the colonies. As such, we performed an experiment to assess the ability of polyp colonies to grow on glass versus plastic slides in aquaria. We cultured frustules from Coal City, Illinois, USA on glass slides and plastic slides microscope slides (all 7.6 cm × 2.5 cm [3 in × 1 in]): to do this, we seeded the slides with ten frustules each in dishes of Hydra Medium (HM) [29]. Once two or more frustules differentiated into polyps, we put the slides into slide boxes (14.1 cm × 9.2 cm × 3.6 cm) with the front and back walls cut out in a technique adapted from Folino-Rorem and Renken [48]. These boxes were then placed into a 2.5L tank of HM with filtration by an AquaClear 20 filter and aeration by air stone and bubbler. To feed the polyps, these boxes were transferred to interim containers of HM and flooded with brine shrimp rinsed in DI water before being returned to the original tank. Colonies were grown on slides in the tanks for 2 months before being counted; colony numbers, frustule numbers, and the number of polyps per colony were recorded and analyzed with Mann-Whitney U-tests and a t-test.
2.2. Construction of Sampling Apparatus
After observing high growth levels on both glass and plastic slides in aquaria, we sought to further experiment with substrates as methods for field sampling. We chose four different substrates for sampling: glass slides, plastic slides, masonite (Hester-Dendy discs), and borosilicate glass petri dishes (6 cm diameter × 1.5 cm depth) in a metal cage (Dr. Tim Wood, personal communication). These were selected based on our own experience with culturing Craspedacusta sowerbii colonies in a laboratory setting, and sampling techniques used for other organisms, as well as the ability to easily place each substrate type into a small dish of water and examine it underneath a dissecting microscope. For our glass and plastic substrates, we used the aformentioned slide box apparatus (Figure 2). Each slide box also had six glass microscope slides at the bottom of the box to help with weighing the boxes down; any organisms on these slides were not quantified.
For our use of Hester-Dendys, we used three discs of 7.6 cm (3in) diameter evenly spaced apart with discs of 2.5 cm (1in) diameter (Figure 2). Additionally, we adapted a technique used to sample for freshwater bryozoans (Dr. Tim Wood, personal communication, 2023) and placed four glass petri dishes (bottoms or tops) in a plastic coated metal cage (a bird suet cage; 13.5 cm X 12 cm X 5 cm) with the open ends facing the outside (Figure 2).
To combine and deploy these, we first secured a zip tie through a cinder block. We then attached one slide box of glass slides, one slide box of plastic slides, one Hester-Dendy with three discs, and one metal cage to this cable with carabiners (Figure 2). We also attached a rope to the zip tie to hold a buoy for collection of these setups later on.
2.3. Field Site Descriptions
We sampled three sites in the Illinois-Indiana (USA) region (Figure 3, Table 1), two of which had been confirmed to have both polyp colonies and medusae (Inverness and Coal City, IL). These three sites have different characteristics in Secchi depth, conductivity and temperature.
Site 1 is a drainage lake, Lake Harrowgate, in a neighborhood in Inverness, Illinois. The site’s abiotic conditions include a cold input of water from the bottom of the lake, bringing higher dissolved oxygen with it. Medusae have been confirmed at the Inverness site (Phyllis Trostrud, personal communication, 2022), as well as the polyp stage.
Site 2 is a lake in a private club in Coal City, Illinois. The grounds consist of several lakes where Craspedacusta sowerbii medusae regularly have been sighted, including the Coal City Area Club Catfish Pond, where we sampled. We have also confirmed the presence of polyp colonies in prior samples of wood and stems from this lake. Coal City colonies (referred to as simply CC) have been cultured in a laboratory setting with very little trouble; The waters in the club are relatively shallow with low turbidity (Secchi depth = 2m) and reflect seasonal temperatures. In addition, the Coal City Area Club Catfish Pond does not exhibit high levels of movement, but the waters are regularly used for human recreation, especially for fishing during the summer months.
Site 3 is a public lake, Wolf Lake, in Hammond, Indiana. The lake is regularly used for recreation (especially fishing) and has high turbidity; similar to Coal City, the waters do not exhibit high levels of movement. To our knowledge, neither the polyp or medusa stage of Craspedacusta sowerbii has been confirmed at Wolf Lake.
2.4. Setup Deployments and Counting
We deployed four setups at each of the three locations. At each location, two different sites in the water were identified, and two setups were placed at each of these sites. In the cases of Coal City and Inverness, previous sampling for Craspedacusta sowerbii polyp colonies had been conducted, so we placed the setups at those sites known to have colonies. Hammond (Wolf Lake), on the other hand, has not had extensive sampling work for polyps, so we selected spots where we hypothesized colonies to be based on available substrate. Setups were left at each location for two months before we retrieved them; while our setups at Coal City and Inverness were retrieved without difficulty, we lost two of the setups at Hammond due to vandalism.
Once removed from the water, we removed the four substrates from the cinder block and placed them into separate containers filled with water from the site. These samples were then transported back to Wheaton College (Wheaton, Illinois, USA) for examination; we used a dissecting microscope to search all slides, Hester-Dendy discs, and petri dishes for polyp colonies and frustules. For slides and Hester-Dendy discs, we examined both sides of the slide or disc. We also looked at both sides of the petri dishes although one side of the dish, the dish portion versus the bottom, was more directly exposed to the water. Additionally, any sediment accumulation on the slides was removed by gently puffed away using a pipette.
Once the number of polyp colonies and frustules on each substrate piece was counted, we standardized their numbers so as to get a per square inch (6.4516 cm2) count of colonies for each substrate type. Following this, we performed Kruskal-Wallis and Dunn post-hoc tests using R and the dunn.test package [49].
2.5. Follow-Up Laboratory Experiments
Following our recording of data, we performed a small follow-up experiment relating to the sampling procedure. Firstly, while the ability of frustules to attach and develop into colonies was tested on plastic and glass, we had not confirmed it on Hester-Dendy discs; as such, we seeded Inverness frustules on Hester-Dendy discs suspended in HM and monitored them for both attachment and development into colonies. We had 3 replicate bowls each with one disc that was seeded with 15 frustules from the Inverness population. This was done with older discs and with recently purchased discs to see if that was a factor in establishment. Secondly, we kept the sample slides in HM and fed them Artemia brine shrimp rinsed in DI water [29] for continued monitoring and examination. We found that some of the microscope slides had podocysts; we kept these slides in HM at room temperature and monitored them for polyp development. Additionally, we identified general types of other invertebrates observed on the various substrate types.
3. Results
3.1. Pilot Laboratory Substrates
In general, we observed more colonies and frustule presence on glass slides with more polyps per colony (Table 2). We used a Mann-Whitney U-Test to test the difference between colony number, frustule number, and polyps per colony between samples on glass slides versus on plastic slides, and we report no significant difference in the number of colonies per slide () and no significant difference in the number of frustules per slide (). However, since polyps per colony is closer to a normal distribution, we used the t-test and again observed no significant difference in the number of polyps per colony ().
3.2. Field Results
Concerning the usage of substrates in the field, our results showed that our method is capable of detection of the frustule and polyp stage; we found frustules and polyp colonies on both glass and plastic slides, but we did not find them on Hester-Dendy discs or on petri dishes. For a detection context, this may be sufficient to confirm the presence of colonies in a given body of water. However, we generally had low colony and frustule numbers overall; despite this, a Kruskal-Wallis test and Dunn post-hoc test indicated a significant difference between the number of colonies per square inch between all methods (); additionally, plastic and glass slides had more colonies settle on them versus on Hester-Dendy discs and petri dishes (Figure 4). A similar result was observed when quantifying the differences in the number of frustules per square inch between substrates () (Figure 4). Additionally, while we found specimens at the Inverness and Wolf sites, we did not find any at the Coal City site; this produced a statistically significant difference in sampled animals between the sites ().
3.3. Follow-Up Results
Follow-up experiments provide additional nuance to some of our findings; when Craspedacusta sowerbii frustules were seeded on Hester-Dendy discs, we found that the frustules failed to attach to the discs; instead, the Hester-Dendy discs had tinted the water brown and the frustules disintegrated. This occurred with both the new and older discs.
Furthermore, we were able to identify various types of invertebrates that had also settled on the slides, including oligochaetes, Hydra, bryozoan colonies, dreissenids, and chironomids, indicating that our gathering of Craspedacusta sowerbii specimens presents a competition for space and may thus have led to reduced numbers. This was especially pronounced in the Hester-Dendy discs; our data indicate a significant difference in the number of other invertebrates per square inch (), with Hester-Dendy discs containing the highest number of invertebrates compared to all other groups (Figure 5).
During the analyses of all substrate types, we found podocysts on glass slides and petri dishes from all three sites, as well as on plastic slides from Wolf Lake. While we did not collect any quantitative data on podocyst numbers, we found that some of the podocysts had regenerated into colonies after being cultured at room temperature.
4. Discussion
Overall, our findings are indicative of a sampling method for Craspedacusta sowerbii frustules, polyp colonies and even podocysts that can be useful for detection and collection of these stages given the challenges and time of finding them compared to the medusa stage. Furthermore, this method will increase the ability to definitely locate more life cycle stages of C. sowerbii in various locations globally. It can also be effective for obtaining specific populations of C. sowerbii. Additionally, our results indicate that past research cultivating colonies in plastic versus glass vessels may have an impact on growth rates, though this impact is not likely to be large and would require further investigation to confirm any substantial difference.
One key further point of improvement for this work is to place additional controls on timing of apparatus deployment; i.e., how long the apparatuses are left in the water, and when they are first put in. We were unable to control for timing as well as we would have liked; natural next directions in this research would be to leave the apparatuses in the water for shorter and varying periods of time. Deploying several setups at a given location in the spring and retrieving a portion of those throughout the summer and fall months would provide a more complete documentation of frustule and polyp colony presence. In conjunction, recording zooplankton and benthos food availability along with abiotic parameters at a given location would provide valuable information for food availability for the various life cycle stages. Additionally, another natural next research direction would be the investigation of other substrates. While we had rationale for our four substrates of choice, we cannot rule out possibilities of other substrates being more efficient in enhancing frustule settlement and colony growth. Four other substrates recorded in the literature are wood, stones, plant material, and dreissenid shells. One of the first recorded sightings of Craspedacusta sowerbii in Japan was by Amemiya [50], who reported polyp colonies in an aquarium tank; these colonies were on plants brought to Japan from San Francisco, California (USA). Gasith et al. [51] reported a similar observation in Israel, where polyp colonies were growing on Elodea in an axolotl tank, and Bushnell and Porter [52] reported similar occurrences at the University of Colorado (USA). Craspedacusta sowerbii is thought to be introduced to new environments primarily by the transport of plants. As such, it is possible to sample for C. sowerbii polyp colonies and frustules by collecting plants from any aquatic environment. While most papers cite the polyp colonies as growing on Elodea [32,50,51], a variety of plants could be gathered for sampling of the polyp stage; Bushnell and Porter [52] reference many plants in the same environment with C. sowerbii; we suspect that many of these could be conducive to the establishment of the polyp stage. However, sampling and looking over plant stems and leaves is very labor intensive and our sampling method would save a great deal of time. Therefore, further research would involve laboratory experiments with plants and the other substrates mentioned to determine which types promote frustule settlement and polyp colony establishment.
In addition to plants, some literature reports sampling for Craspedacusta sowerbii polyp colonies and frustules on rocks [44,45]. Previous research describes the collection of stones from habitats where C. sowerbii medusae were sighted, with Duggan and Eastwood [43] also collecting stones from lakes with no known sightings of medusae. In this study, these stones were kept in the laboratory in lake water while being scanned for polyp colonies and frustules. However, stones presented a challenge in finding the colonies: Duggan and Eastwood [43] reported examining stones for two weeks, four times a week, to find polyp colonies on the stones, and we suspect that some samples may take even longer to look through to confirm the presence of C. sowerbii.
Wood is another natural substrate on which Craspedacusta sowerbii polyp colonies have been found [22,43,44,45]. Folino-Rorem et al. [29] collected a piece of wood from Gatun Lake (Panama), from which they sourced the animals used in their studies on laboratory rearing of C. sowerbii. Additionally, Lewis et al. [22] sampled on both wood and concrete in freshwater systems throughout Japan. While not extensively commented on in the literature, we find from personal experience that wood is a common substrate for polyp colonies and have found colonies on wood pieces from various freshwater systems throughout Illinois (USA). Additionally, the size of the wood does not seem to play a role in attachment; we have found colonies on twigs around 1-2 cm in diameter and on logs more than 10 cm in diameter. However, similar to other natural substrates, wood as a collection method poses similar challenges to the aforementioned stones, namely in that it takes a long time to confirm the presence or absence of polyp colonies on a piece of wood. Wood can also bring the challenges of transport and storage (especially for large pieces of wood).
For all aforementioned substrates, we find that our method mitigates many of the problems posed by the use of natural substrates. The use of glass slides, petri dishes, and Hester-Dendy discs allows for examiners to quickly and easily search for frustules and polyp colonies since these substrates easily fit in dishes for viewing using a dissecting scope compared to larger natural rocks and pieces of wood. While we were unable to acquire stones, wood, and plants in neat slides or discs to easily deploy and retrieve them, a systematic method of using these substrates would be a natural direction for future experimentation.
We also suggest that the impact of flow on the spread and establishment of Craspedacusta sowerbii is important. As indicated by our results, we observed more organisms on the slides in the slide boxes compared to other substrate types, and we observed more organisms on the substrates at the Inverness site. This difference in collected organisms between glass slides and glass petri dishes may be attributed to flow; the glass petri dishes in the metal cages, while commonly used to collect freshwater bryozoa (Tim Wood, personal communication, 2023), seemed to diminish large amounts of water flow through the apparatus, while the slide box setup did not. It could also be that the cages with petri dishes were positioned in the setup too far from the source of frustules since they were above the slide boxes. From personal experience, we have observed frustules flowing in water and settling on new substrates as a method of spreading; Matthews [32] documents frustules spreading via thin mucus threads, and we have observed this phenomenon in aquaria. Additionally, aquaria with flow have shown to be an effective method to culture all stages of the life cycle [28] (personal observations). As such, it may be that C. sowerbii frustules were able to attach to the microscope slides due to water flow. This may also be an explanation as to why no colonies or frustules were found on the setups deployed at Coal City, as the site features smaller lakes with relatively still waters. In addition to aspects of flow, the fact that the slide boxes were closer to the cinder block and closer to the benthic substrate where frustules were produced in all of the setups may explain the minimal occurrences of frustules and polyp colonies on the petri dishes and similarly for the Hester-Dendy setups. Future studies where all 4 substrates are alternated in position relative to the cinder block would address this issue.
Furthermore, our results indicate that further investigations into the specific nature of interactions between Craspedacusta sowerbii and other organisms are warranted for future study. The organisms we collected on our substrates include many that prey on C. sowerbii frustules and polyps and may also compete with polyp colonies for space. Previous studies comment on the richness of zooplankton in the environments where C. sowerbii are found, as well as the diet habits of the medusa stage [23,24,34,53], and some discuss these predatory relationships as well [36,42], though the feeding habits of the polyp stage are not commented as frequently in the literature, with Lytle [54] and Acker and Muscat [30] being two that demonstrate the impact of diet on polyp behavior. In addition to feeding habits, various organisms also prey on C. sowerbii polyps; from our own experience with culturing C. sowerbii colonies and frustules in the lab, we have observed various mites and chironomids eating colonies and even Hydra eating frustules. Additionally, we observe that mites are often brought into cultures through wood and other new natural substrates on which the colonies are present. As such, it is possible that various other organisms got to the substrates first and outcompeted C. sowerbii colonies for space or ate any frustules and polyps on the substrates. This may especially have been the case for Hester-Dendy disk samples, which featured higher numbers of other organisms such as oligochaetes, chironomids and potentially other invertebrates compared to all other substrates, though the Hester-Dendy disks we used could also have been less suitable for the settlement of frustules due to some contamination (as stated in Results 3.3).
A key subset of interactions work would be further research into the specific relationship between Craspedacusta sowerbii polyps and dreissenids. Stanković and Ternjej [55] describe C. sowerbii polyps on Dreissenia polymorpha shells, and we have often observed frustules and polyps on dreissenids in samples from Lake Michigan (N. Folino-Rorem, personal observation, 2022). In this study, we found small mussels and polyps on the substrates deployed at Wolf Lake but no polyps on the mussels perhaps due to the small size of the mussels. Dreissenids and other freshwater bivalves may provide additional substrate for polyp colonies to become established as seen with empty bivalve shells providing a means of locating polyps and podocysts with marine jellyfish (scyphzoans) [56]. However, with live mussels, additional factors could be at play, including flow created by dreissenid filtration and predation by chironomids and mites in clumps of dreissenids.
Finally, our sampling method could be beneficial to studies conducting eDNA assessment of invasive species. Recent studies utilizing eDNA samples in conjunction with hydrobiological surveys have detected Craspedacusta sowerbii via eDNA surveys but have not located the morphological stages of frustules, polyps or medusae [57,58] (Drs. Sarah Brown and John Darling, personal communication, 2024). It is unclear if the eDNA for C. sowerbii is from frustule, polyp or medusa cells. Our sampling method could enhance the eDNA assessment efforts for the invasive hydrozoan Craspedacusta sowerbii by locating frustules and polyps in sites where they have never been collected but eDNA indicates the presence of this invasive hydroid. Furthermore, our sampling protocol could aid in understanding how often introductions have occurred and which species of Craspedacusta may be present in specific habitats. This information would prove valuable in the overall understanding of the extent of invasion in the hydrozoan where observing the medusa stage is unpredictable and the frustule and polyp stages are so small and difficult to detect. Furthermore, our method of detecting the pre-medusa stage will enhance our understanding of which aspects of the polyp biology influence the medusa occurrences and biology [59,60]. This invasive species may ecologically impact aquatic systems as we consider the various factors important in potential jellyfish blooms with increasing water temperatures [1,61].
5. Conclusions
While we presented and tested an effective methodology for detecting and collecting the frustule and polyp stage of the invasive freshwater hydrozoan Craspedacusta sowerbii, that being immersion of a cut-open microscope slide box with glass or plastic microscope slides in a body of water, our method nonetheless has room for improvement. We particularly mention deployment timing, substrate positioning, and substrate types. Our results also open up key areas for further research regarding C. sowerbii, particularly in the realm of flow and interactions with other organisms.
Nonetheless,we hope to further utilize this method to detect and collect the polyp stage at more locations and believe that this tool can be instrumental in determining the full range of Craspedacusta sowerbii life cycle stages since medusa sightings alone cannot adequately capture this range. Enhanced information regarding the presence and ability to survive and bud medusae will potentially clarify the sporadic observance of medusae at given locations.
Author Contributions
Conceptualization, J.Z.; methodology, J.Z.; software, J.Z.; validation, N.F.-R.; formal analysis, J.Z.; investigation, J.Z., N.F.-R.; resources, J.Z.; data curation, J.Z., N.F.-R.; writing—original draft preparation, J.Z, N.F.-R.; writing—review and editing, J.Z, N.F.-R.; visualization, J.Z., N.F.-R.; supervision, N.F.-R.; project administration, N.F.-R.; funding acquisition, N.F.-R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Illinois-Indiana Sea Grant, project number R/22-1-04 and the Wheaton College Department of Biological and Health Sciences. Wheaton College Science Division, Wheaton, IL. The APC was funded by the same grant.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
All data gathered in this project, as well as the code used for subsequent analysis, can be found at the following GitHub repository: https://github.com/jonazhu/craspedacustasampling.
Acknowledgments
We thank Jensyn Langguth, Chloe Davis, and Micah Perry for their aid in setup and deployment of the sampling substrates, Doug Rorem and Dr. Kristen Page for their additional aid in deployment, retrieval, and data collection, and Ryan Cummings and Phyllis Trostrud for granting us access to the Coal City and Inverness sites, respectively.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Marchessaux, G.; Lüskow, F.; Bejean, M.; Pakhomov, E.A. Increasing temperature facilitates polyp spreading and medusa appearance of the invasive hydrozoan Craspedacusta sowerbii. Biology 2022, 11, 1100. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, J.A.; Westerman, E.L.; Harris, L.G. Elevated seasonal temperatures eliminate thermal barriers of reproduction of a dominant invasive species: A community state change for northern communities? Diversity and Distributions 2017, 23, 1182–1192. [Google Scholar] [CrossRef]
- Walsh, J.R.; Hansen, G.J.; Read, J.S.; Vander Zanden, M.J. Comparing models using air and water temperature to forecast an aquatic invasive species response to climate change. Ecosphere 2020, 11, e03137. [Google Scholar] [CrossRef]
- Beric, B.; MacIsaac, H.J. Determinants of rapid response success for alien invasive species in aquatic ecosystems. Biological Invasions 2015, 17, 3327–3335. [Google Scholar] [CrossRef]
- Coughlan, N.E.; Lyne, L.; Cuthbert, R.N.; Cunningham, E.M.; Lucy, F.E.; Davis, E.; Caffrey, J.M.; Dick, J.T. In the black: Information harmonisation and educational potential amongst international databases for invasive alien species designated as of union concern. Global Ecology and Conservation 2020, 24, e01332. [Google Scholar] [CrossRef]
- Simberloff, D. Maintenance management and eradication of established aquatic invaders. Hydrobiologia 2021, 848, 2399–2420. [Google Scholar] [CrossRef]
- Soto, I.; Cuthbert, R.N.; Ahmed, D.A.; Kouba, A.; Domisch, S.; Marquez, J.R.; Beidas, A.; Amatulli, G.; Kiesel, J.; Shen, L.Q.; et al. Tracking a killer shrimp: Dikerogammarus villosus invasion dynamics across Europe. Diversity and Distributions 2023, 29, 157–172. [Google Scholar] [CrossRef]
- Linders, T.E.W.; Schaffner, U.; Eschen, R.; Abebe, A.; Choge, S.K.; Nigatu, L.; Mbaabu, P.R.; Shiferaw, H.; Allan, E. Direct and indirect effects of invasive species: Biodiversity loss is a major mechanism by which an invasive tree affects ecosystem functioning. Journal of Ecology 2019, 107, 2660–2672. [Google Scholar] [CrossRef]
- Britton, J.R.; Lynch, A.J.; Bardal, H.; Bradbeer, S.J.; Coetzee, J.A.; Coughlan, N.E.; Dalu, T.; Tricarico, E.; Gallardo, B.; Lintermans, M.; et al. Preventing and controlling nonnative species invasions to bend the curve of global freshwater biodiversity loss. Environmental Reviews 2023, 31, 310–326. [Google Scholar] [CrossRef]
- Scalici, M.; Chiesa, S.; Mancinelli, G.; Rontani, P.M.; Voccia, A.; Nonnis Marzano, F. Euryhaline aliens invading Italian inland waters: The case of the Atlantic blue crab Callinectes sapidus Rathbun, 1896. Applied Sciences 2022, 12, 4666. [Google Scholar] [CrossRef]
- Lord, J.P. Impact of seawater temperature on growth and recruitment of invasive fouling species at the global scale. Marine ecology 2017, 38, e12404. [Google Scholar] [CrossRef]
- Emery-Butcher, H.E.; Beatty, S.J.; Robson, B.J. The impacts of invasive ecosystem engineers in freshwaters: A review. Freshwater Biology 2020, 65, 999–1015. [Google Scholar] [CrossRef]
- Ricciardi, A.; Iacarella, J.C.; Aldridge, D.C.; Blackburn, T.M.; Carlton, J.T.; Catford, J.A.; Dick, J.T.; Hulme, P.E.; Jeschke, J.M.; Liebhold, A.M.; et al. Four priority areas to advance invasion science in the face of rapid environmental change. Environmental Reviews 2021, 29, 119–141. [Google Scholar] [CrossRef]
- Angel, D.L.; Edelist, D.; Freeman, S. Local perspectives on regional challenges: Jellyfish proliferation and fish stock management along the Israeli Mediterranean coast. Regional Environmental Change 2016, 16, 315–323. [Google Scholar] [CrossRef]
- González-Duarte, M.M.; Megina, C.; López-González, P.J.; Galil, B. Cnidarian alien species in expansion. The Cnidaria, Past, Present and Future: The world of Medusa and her sisters 2016, pp. 139–160.
- Purcell, J.E. Successes and challenges in jellyfish ecology: Examples from Aequorea spp. Marine Ecology Progress Series 2018, 591, 7–27. [Google Scholar] [CrossRef]
- Rodrigues, T.; Domínguez-Pérez, D.; Almeida, D.; Matos, A.; Antunes, A. Medusozoans reported in Portugal and its ecological and economical relevance. Regional studies in marine science 2020, 35, 101230. [Google Scholar] [CrossRef]
- Smith, A.S.; Alexander Jr, J.E. Potential effects of the freshwater jellyfish Craspedacusta sowerbii on zooplankton community abundance. Journal of plankton research 2008, 30, 1323–1327. [Google Scholar] [CrossRef]
- Minchin, D.; Caffrey, J.M.; Haberlin, D.; Germaine, D.; Walsh, C.; Boelens, R.; Doyle, T.K. First observations of the freshwater jellyfish Craspedacusta sowerbii Lankester, 1880 in Ireland coincides with unusually high water temperatures 2016.
- Parent, G. Une page d’histoire des sciences contemporaines: Un siècle d’observations sur la méduse d’eau douce, Craspedacusta sowerbii Lank. Publications de la Société Linnéenne de Lyon 1982, 51, 47–63. [Google Scholar] [CrossRef]
- Didžiulis, V. Fact Sheet–Craspedacusta sowerbyi. Online database of the North European and Baltic Network on Invasive Alien Species–NOBANIS. Invasive Alien Species 2006, pp. 1–7.
- Lewis, C.; Migita, M.; Hashimoto, H.; Collins, A.G. On the occurrence of freshwater jellyfish in Japan 1928–2011: eighty-three years of records of mamizu kurage (Limnomedusae, Olindiidae). Proceedings of the Biological Society of Washington 2012, 125, 165–179. [Google Scholar] [CrossRef]
- Caputo, L.; Fuentes, R.; Woelfl, S.; Castañeda, L.E.; Cárdenas, L. Phenotypic plasticity of clonal populations of the freshwater jellyfish Craspedacusta sowerbii (Lankester, 1880) in Southern Hemisphere lakes (Chile) and the potential role of the zooplankton diet. Austral Ecology 2021, 46, 1192–1197. [Google Scholar] [CrossRef]
- Pérez-Bote, J.L.; Muñoz, A.; Morán, R.; Roso, R.; Romero, A.J. First record of Craspedacusta sowerbyi Lankester, 1880 (Cnidaria: Limnomedusae: Olindiidae) in the Proserpina Reservoir (Extremadura, SW Spain) with notes on their feeding habits. Belgian Journal of Zoology 2006, 136, 163. [Google Scholar]
- Fish, G.R. Craspedacusta sowerbyi Lankester (Coelenterata: Limnomedusae) in New Zealand lakes. New Zealand Journal of Marine and Freshwater Research 1971, 5, 66–69. [Google Scholar] [CrossRef]
- Rayner, N.A. First record of Craspedacusta sowerbyi Lankester (Cnidaria: Limnomedusae) from Africa. Hydrobiologia 1988, 162, 73–77. [Google Scholar] [CrossRef]
- McKercher, E.; O; Connell, D.; Fuller, P.; Liebig, J.; Larson, J.; Makled, T.; Fusaro, A.; .; Daniel, W. Craspedacusta sowerbii Lankester, 1880. https://nas.er.usgs.gov/queries/factsheet.aspx?SpeciesID=1068, 2004. Accessed: 2024-05-09.
- Marchessaux, G.; Bejean, M. From frustules to medusae: A new culture system for the study of the invasive hydrozoan Craspedacusta sowerbii in the laboratory. Invertebrate biology 2020, 139, e12308. [Google Scholar] [CrossRef]
- Folino-Rorem, N.C.; Reid, M.; Peard, T. Culturing the freshwater hydromedusa, Craspedacusta sowerbii under controlled laboratory conditions. Invertebrate Reproduction & Development 2016, 60, 17–27. [Google Scholar] [CrossRef]
- Acker, T.S.; Muscat, A.M. The ecology of Craspedacusta sowerbii Lankester, a freshwater hydrozoan. American Midland Naturalist 1976, pp. 323–336.
- Payne, F. Further studies on the life history of Craspedacusta ryderi, a fresh-water hydromedusan. The Biological Bulletin 1926, 50, 433–443. [Google Scholar] [CrossRef]
- Matthews, D.C. A comparative study of Craspedacusta sowerbyi and Calpasoma dactyloptera life cycles 1966.
- Payne, F. A study of the fresh-water medusa, Craspedacusta ryderi. Journal of Morphology 1924, 38, 387–429. [Google Scholar] [CrossRef]
- Davis, C.C. Notes on the food of Craspedacusta sowerbii in Crystal Lake, Ravenna, Ohio. Ecology 1955, 36, 364–366. [Google Scholar] [CrossRef]
- Lucas, K.; Colin, S.P.; Costello, J.H.; Katija, K.; Klos, E. Fluid interactions that enable stealth predation by the upstream-foraging hydromedusa Craspedacusta sowerbyi. The Biological Bulletin 2013, 225, 60–70. [Google Scholar] [CrossRef]
- Dodson, S.I.; Cooper, S.D. Trophic relationships of the freshwater jellyfish Craspedacusta sowerbyi Lankester 1880. Limnology and Oceanography 1983, 28, 345–351. [Google Scholar] [CrossRef]
- Stefani, F.; Leoni, B.; Marieni, A.; Garibaldi, L. A new record of Craspedacusta sowerbii, Lankester 1880 (Cnidaria, Limnomedusae) in northern Italy. Journal of Limnology 2010, 69, 189. [Google Scholar] [CrossRef]
- DeVries, D.R. The freshwater jellyfish Craspedacusta sowerbyi: A summary of its life history, ecology, and distribution. Journal of Freshwater Ecology 1992, 7, 7–16. [Google Scholar] [CrossRef]
- Gophen, M.; Shealtiel, L.; et al. Record of the alien species Craspedacusta sowerbyi Lankester, 1880 (Cnidaria: Limnomedusae) in Lake Kinneret catchment area. BioInvasions Records 2012, 1, 29–31. [Google Scholar] [CrossRef]
- Akçaalan, R.; Isinibilir, M.; Gürevin, C.; Sümer, A. A new contribution of biodiversity of Sapanca lake: Craspedacusta sowerbyi Lankester, 1880 (Cnidaria: Hydrozoa). Journal of FisheriesSciences. com 2011, 5, 43. [Google Scholar] [CrossRef]
- Jakovčev-Todorović, D.; Đikanović, V.; Skorić, S.; Cakić, P. Slatkovodna meduza Craspedacusta sowerbyi Lankester, 1880 (Hydrozoa, Olindiidae)-50 godina istraživanja u Srbiji. Archives of Biological Sciences 2010, 62, 123–127. [Google Scholar] [CrossRef]
- Jankowski, T.; Strauss, T.; Ratte, H.T. Trophic interactions of the freshwater jellyfish Craspedacusta sowerbii. Journal of plankton research 2005, 27, 811–823. [Google Scholar] [CrossRef]
- Duggan, I.C.; Eastwood, K.R. Detection and distribution of Craspedacusta sowerbii: observations of medusae are not enough 2012.
- Klotz, R.U. Hidden neozoans in macrozoobenthos: The polyp stage of the freshwater jellyfish Craspedacusta sowerbii. PhD thesis, Dissertation, München, Ludwig-Maximilians-Universität, 2022, 2022.
- McClary, A. The effect of temperature on growth and reproduction in Craspedacusta sowerbii. Ecology 1959, 40, 158–162. [Google Scholar] [CrossRef]
- Siquier, M.F.; Alanis, W.S.; Debat, C.M. First record of Craspedacusta sowerbii Lankester, 1880 (Hydrozoa, Limnomedusae) in a natural freshwater lagoon of Uruguay, with notes on polyp stage in captivity. Brazilian Journal of Biology 2017, 77, 665–672. [Google Scholar] [CrossRef]
- Wang, Y. Genetic population structure and environmental impact on Craspedacusta at the medusa and polyp stages. PhD thesis, Ludwig Maximilian University of Munich, 2022.
- Folino-Rorem, N.C.; Renken, C.J. Effects of salinity on the growth and morphology of the invasive, euryhaline hydroid Cordylophora (Phylum Cnidaria, Class Hydrozoa). Invertebrate biology 2018, 137, 78–90. [Google Scholar] [CrossRef]
- Dinno, A.; Dinno, M.A. Package ‘dunn. test’. CRAN Repos 2017, 10, 1–7. [Google Scholar]
- Amemiya, I. Fresh-water Medusa found in the Tank of my Laboratory. Japanese Journal of Zoology 1929, 3, 3. [Google Scholar]
- Gasith, A.; Gafny, S.; Hershkovitz, Y.; Goldstein, H.; Galil, B.S.; et al. The invasive freshwater medusa Craspedacusta sowerbii Lankester, 1880 (Hydrozoa: Olindiidae) in Israel. Aquatic Invasions 2011, 6, S147–S152. [Google Scholar] [CrossRef]
- Bushnell Jr, J.H.; Porter, T.W. The occurrence, habitat, and prey of Craspedacusta sowerbyi (particularly polyp stage) in Michigan. Transactions of the American Microscopical Society 1967, pp. 22–27.
- Galarce, L.C.; Riquelme, K.V.; Osman, D.Y.; Fuentes, R.A.; et al. A new record of the non indigenous freshwater jellyfish Craspedacusta sowerbii Lankester, 1880 (Cnidaria) in Northern Patagonia (40 S, Chile). BioInvasions Records 2013, 2, 263–270. [Google Scholar] [CrossRef]
- Lytle, C. Patterns of budding in the freshwater hydroid Craspedacusta. The Biology of Hydra and Some Other Coelenterates; University of Miami Press: Miami, FL, USA, 1961; pp. 317–336. [Google Scholar]
- Stanković, I.; Ternjej, I. New ecological insight on two invasive species: Craspedacusta sowerbii (Coelenterata: Limnomedusae) and Dreissenia polymorpha (Bivalvia: Dreissenidae). Journal of Natural History 2010, 44, 2707–2713. [Google Scholar] [CrossRef]
- van Walraven, L.; van Bleijswijk, J.; van der Veer, H.W. Here are the polyps: In situ observations of jellyfish polyps and podocysts on bivalve shells. PeerJ 2020, 8, e9260. [Google Scholar] [CrossRef]
- Jeunen, G.J.; Lipinskaya, T.; Gajduchenko, H.; Golovenchik, V.; Moroz, M.; Rizevsky, V.; Semenchenko, V.; Gemmell, N.J. Environmental DNA (eDNA) metabarcoding surveys show evidence of non-indigenous freshwater species invasion to new parts of Eastern Europe. Metabarcoding and Metagenomics 2022, 6, e68575. [Google Scholar] [CrossRef]
- Blackman, R.C.; Carraro, L.; Keck, F.; Altermatt, F. Measuring the state of aquatic environments using eDNA—upscaling spatial resolution of biotic indices. Philosophical Transactions of the Royal Society B 2024, 379, 20230121. [Google Scholar] [CrossRef]
- Dumont, H.J. The distribution and ecology of the fresh-and brackish-water medusae of the world. Studies on the ecology of tropical zooplankton 1994, pp. 1–12.
- Gießler, S.; Strauss, T.; Schachtl, K.; Jankowski, T.; Klotz, R.; Stibor, H. Trophic Positions of Polyp and Medusa Stages of the Freshwater Jellyfish Craspedacusta sowerbii Based on Stable Isotope Analysis. Biology 2023, 12, 814. [Google Scholar] [CrossRef]
- Oualid, J.A.; Iazza, B.; Tamsouri, N.M.; El Aamri, F.; Moukrim, A.; López-González, P.J. Hidden diversity under morphology–based identifications of widespread invasive species: The case of the ‘well–known’hydromedusa Craspedacusta sowerbii Lankester 1880. Animal Biodiversity and Conservation 2019, 42, 301–316. [Google Scholar] [CrossRef]
Short Biography of Authors
 |
Jonathan A. Zhu (B.S., Biology, Mathematics with Statistics, Wheaton College, 2023) is a Master’s student studying computational biology at Carnegie Mellon University. He has worked as a research assistant, laboratory assistant, and teaching assistant at Wheaton College. Jonathan has a wide range of research interests but is particularly interested in mathematical modeling and computational inference regarding invasive species spread and marine/freshwater invertebrate zoology. |
 |
Nadine C. Folino-Rorem (PhD, University of New Hampshire, 1989) is a professor in the Biological and Health Sciences Department at Wheaton College, IL, where she has served on faculty since 1993. She received both her doctorate and Master’s degree from the University of New Hampshire, Durham, N.H. Nadine’s teaching responsibilities range from introductory biology, research methods, invertebrate biology, to environmental ethics. Her research students and colleagues focus on the taxonomic and ecological aspects of invasive, freshwater cnidarians. |
Figure 1.
A modified life cycle from Folino-Rorem et al. [29] of the asexual, sexual, and dormancy stages of the invasive freshwater hydrozoan Craspedacusta sowerbii. Asexual reproduction occurs via polyps budding new polyps (approx. 1 mm in height) or by producing frustules (avg. length, 0.43 mm) that become polyps. Dioecious medusae are also budded from a polyp. Immature medusae are <1 mm but once sexually mature (approx. 15-20 mm) spawn either egg or sperm leading to a planula stage. A dormant stage, a podocyst (avg. diameter, 0.18 mm) occurs in unfavorable conditions. The sketch is not drawn to scale.
Figure 1.
A modified life cycle from Folino-Rorem et al. [29] of the asexual, sexual, and dormancy stages of the invasive freshwater hydrozoan Craspedacusta sowerbii. Asexual reproduction occurs via polyps budding new polyps (approx. 1 mm in height) or by producing frustules (avg. length, 0.43 mm) that become polyps. Dioecious medusae are also budded from a polyp. Immature medusae are <1 mm but once sexually mature (approx. 15-20 mm) spawn either egg or sperm leading to a planula stage. A dormant stage, a podocyst (avg. diameter, 0.18 mm) occurs in unfavorable conditions. The sketch is not drawn to scale.
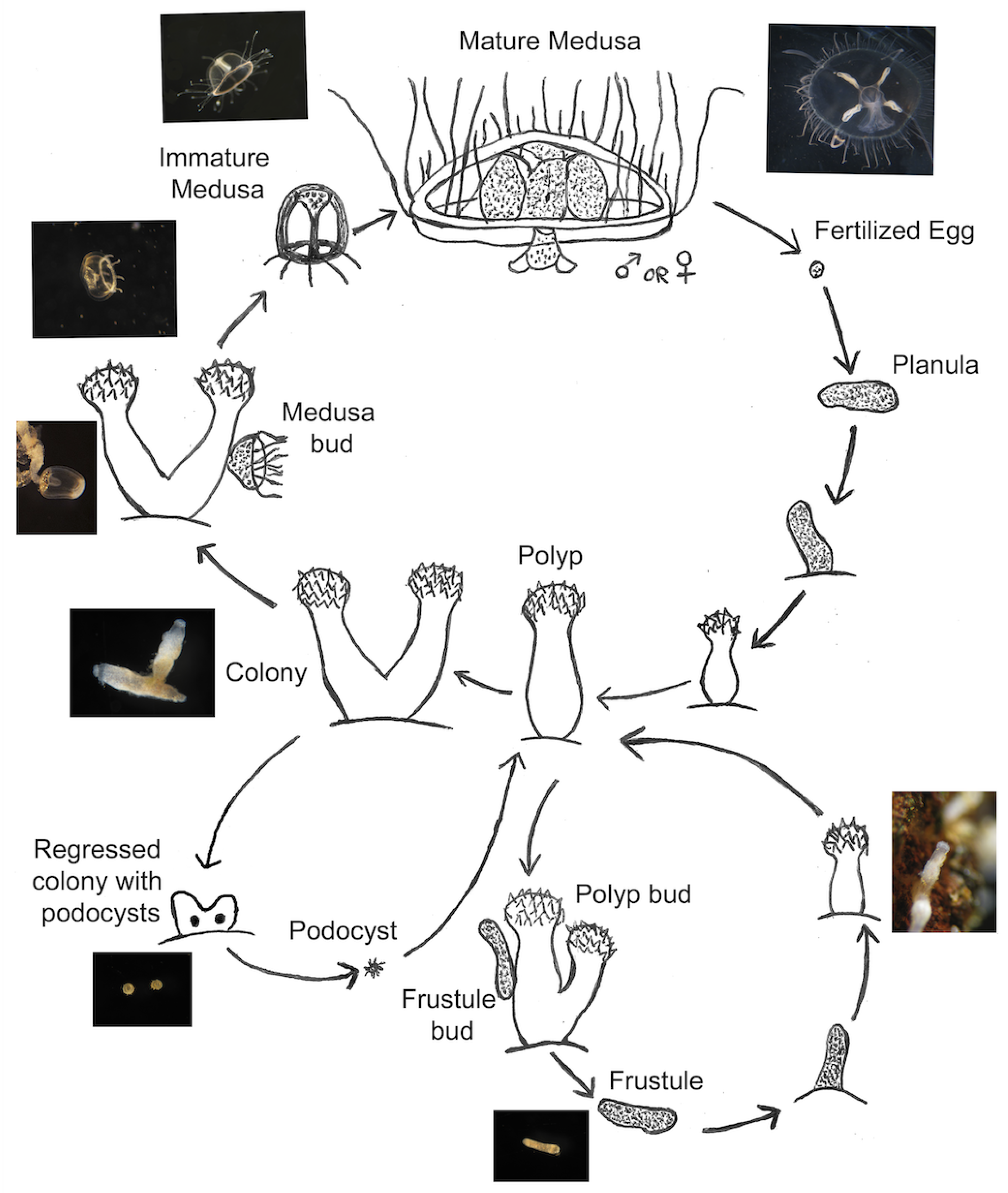
Figure 2.
Construction of the sampling apparatus and substrate deployments used to collect polyp colonies of Craspedacusta sowerbii. (a, left) The substrates utilized in the apparatus, including a Hester-Dendy apparatus with three evenly spaced masonite discs, separated by smaller masonite discs, microscope slide boxes with the sides cut out used to hold glass and plastic slides, and a metal cage holding eight petri dishes, with four visible. (b, right) The entire setup with all substrates attached to a zip tie looped through a cinder block.
Figure 2.
Construction of the sampling apparatus and substrate deployments used to collect polyp colonies of Craspedacusta sowerbii. (a, left) The substrates utilized in the apparatus, including a Hester-Dendy apparatus with three evenly spaced masonite discs, separated by smaller masonite discs, microscope slide boxes with the sides cut out used to hold glass and plastic slides, and a metal cage holding eight petri dishes, with four visible. (b, right) The entire setup with all substrates attached to a zip tie looped through a cinder block.

Figure 3.
Approximate locations of each sampling site, with major cities in the Northern Illinois-Indiana region labeled. (A) Inverness (IL), (B) Coal City (IL), (C) Hammond (IN).
Figure 3.
Approximate locations of each sampling site, with major cities in the Northern Illinois-Indiana region labeled. (A) Inverness (IL), (B) Coal City (IL), (C) Hammond (IN).
Figure 4.
Graphical representation of the mean number of Craspedacusta sowerbii polyp colonies and frustules per square inch that settled on various substrates. (a, left) Comparison of mean polyp colony number. (b, right) Comparison of mean frustule number. All error bars represent SEM.
Figure 4.
Graphical representation of the mean number of Craspedacusta sowerbii polyp colonies and frustules per square inch that settled on various substrates. (a, left) Comparison of mean polyp colony number. (b, right) Comparison of mean frustule number. All error bars represent SEM.
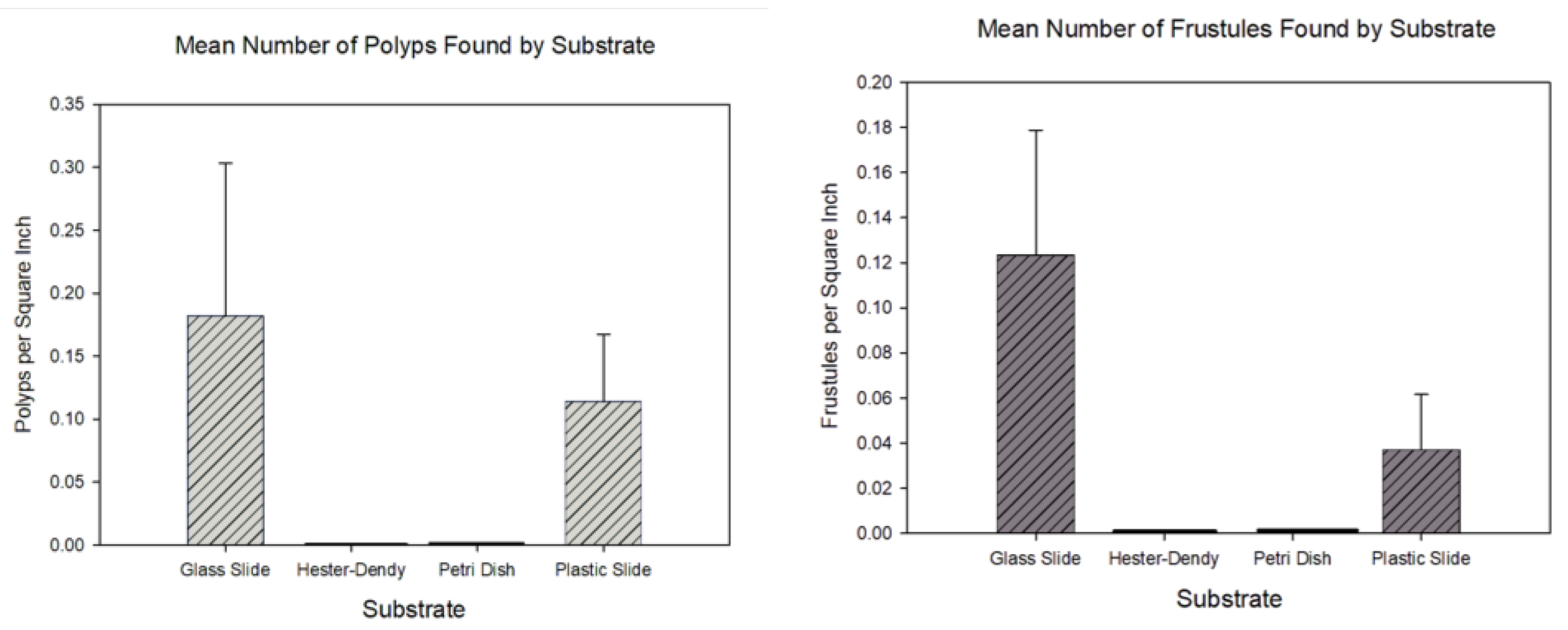
Figure 5.
Graphical representation of the mean number of total invertebrates (excluding Craspedacusta sowerbii) per square inch that settled on various substrates.
Figure 5.
Graphical representation of the mean number of total invertebrates (excluding Craspedacusta sowerbii) per square inch that settled on various substrates.
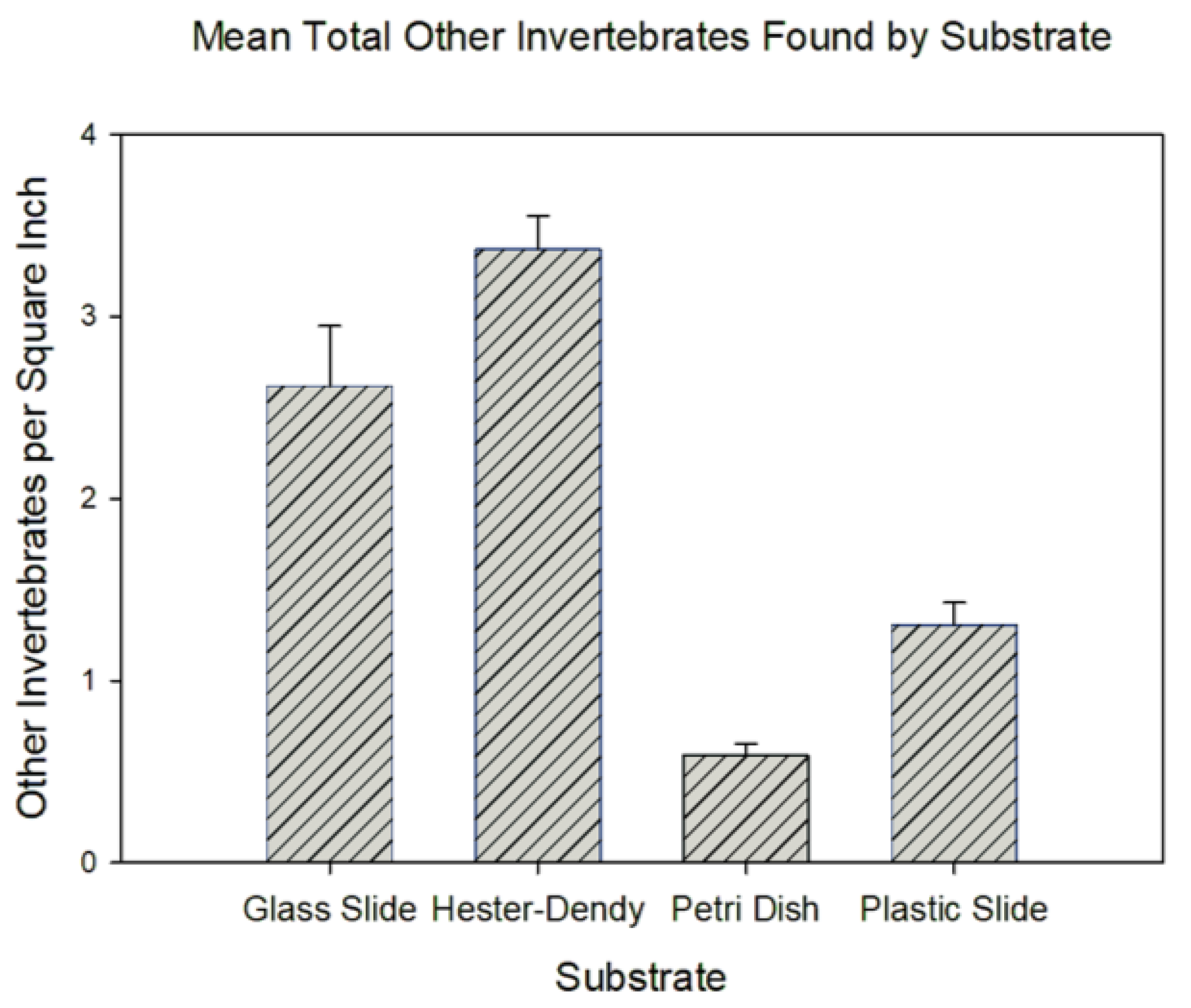

Table 1.
Water parameters, latitude, and longitude for each sampling setup.
| Location | Secchi Depth (m) | Conductivity (S/cm) | Temperature | pH | Setup Site | Latitude | Longitude |
|---|---|---|---|---|---|---|---|
| Inverness (IL) | 0.75 | 1055 | 25.8ºC | 8.26 | 1 | 42.116540°N | 88.122080°W |
| 2 | 42.116540°N | 88.122060°W | |||||
| Coal City (IL) | 2 | 228.6 | 28.3ºC | 7.93 | 1 | 41.316213°N | 88.271704°W |
| 2 | 41.316053°N | 88.271673°W | |||||
| Hammond (IN) | 1 | 927.0 | 25.2ºC | 8.60 | 1 | 41.671930°N | 87.511810°W |
| 2 | 41.672540°N | 87.512360°W |
Table 2.
Summary of polyps/colonies for cultures of Craspedacusta sowerbii grown on glass slides versus plastic slides.
Table 2.
Summary of polyps/colonies for cultures of Craspedacusta sowerbii grown on glass slides versus plastic slides.
| Substrate | Polyps and/or Colonies per slide | Frustules per slide | Polyps per colony |
|---|---|---|---|
| Glass | 28.29 | 24.71 | 3.237 |
| Plastic | 17.93 | 19.00 | 2.933 |
| p-value | 0.3734 | 0.7414 | 0.1759 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



