
Submitted:
30 May 2024
Posted:
31 May 2024
You are already at the latest version
Abstract
There are four types of influenza viruses: A, B, C, and D. Cattle are susceptible to influenza D infection and serve as a reservoir for this seven-segmented influenza virus. It is generally thought that bovines are not a host of other types of influenza viruses including type A. This long-standing viewpoint has been challenged by the recent highly pathogenic avian influenza (HPAI) H5N1 outbreaks in dairy cows in the United States. To date, HPAI H5N1 has spread into nine states affecting 49 dairy herds and causing clinical symptoms including reduced appetite, fever, and a sudden drop in milk production. This review article describes the current epidemiological landscape of HPAI H5N1 in U.S. dairy cows and the recent interspecies transmission events of HPAI H5N1 in other mammals reported in other countries. The review also discusses adaptation mutations of HPAI H5N1 to mammalian hosts and vaccination strategies. Finally, it summarizes some immediate questions that need to be addressed towards a better understanding of infection biology, transmission, and immune response of HPAI H5N1 in bovines.
Keywords:
high pathogenic avian influenza (HPAI)
; H5N1
; cattle
; transmission
1. Introduction
Influenza viruses belong to the Orthomyxoviridae family and there are four known types of influenza viruses, A, B, C, and D. Influenza A and B viruses can cause seasonal influenza with influenza A virus (IAV) having the potential to trigger an influenza pandemic with high morbidity and mortality. Influenza A type consists of 16 hemagglutinin (HA) proteins and 9 neuraminidase (NA), and aquatic waterfowls are considered the natural reservoirs for IAV [1]. Since the 1900s, four pandemics have occurred with two pandemics caused by H1N1 viruses (1918, 2009), one pandemic caused by H2N2 virus (1957), and one caused by H3N2 virus (1968) [2]. These pandemic viruses are believed to originate from birds or pigs, and have acquired human adaptation mutations leading to the sustainable human-to-human transmission. Although influenza C and D viruses do not cause pandemic, their risk to human health cannot be ignored. Influenza C virus (ICV) can cause mild illness in humans, but severe symptoms, such as pneumonia, bronchitis, and bronchiolitis, can develop in children less than two years old [3,4]. Influenza D virus (IDV) uses cattle as the reservoir and can infect other animals, but the impact on human health is not fully understood [5,6,7]. Before the emergence of the IDV in cattle, cattle were not recognized as a susceptible host for influenza viruses [8].
The avian influenza (AI) virus is type A. Based on the intravenous pathogenicity test performed on specific pathogen-free (SPF) chickens, AI viruses are divided into highly pathogenic avian influenza (HPAI) virus and low pathogenic avian influenza (LPAI) virus. HPAI virus can replicate in multiple organs while LPAI cannot, which is mainly because of the difference in the cleavage site of the HA protein precursor. To produce infectious virus particles, the HA precursor needs to be cleaved into HA1 and HA2 by a host protease. The HPAI HA can be cleaved by furin-like proteases ubiquitously expressed in most cell types from the body whereas the LPAI HA can only be cleaved by certain trypsin-like proteases from host cells in the respiratory and enteric tracts [9]. The robust replication property allows HPAI virus replication in multiple organs, which results in more damage to domestic poultry and deleterious outcomes to the poultry industry.
According to the HA sequence difference, HPAI H5N1 viruses are divided into ten distinct clades [10]. Clade 2.3.4.4b emerged in 2020 and has spread to Africa, Asia, Europe, North America, and South America by migratory birds, leading to substantial outbreaks in domestic poultry and wild birds. HPAI H5 viruses were first detected in commercial poultry in January 2022 in the U.S. since 2016, and have affected more than 90 million poultry in 48 states [11]. HPAI viruses have caused considerable amounts of death in commercial poultry, resulting in significant economic loss worldwide. HPAI H5 or H7 virus infects chicken normally with mortality up to 90% to 100% [12].
2. Recent Bovine Influenza A Virus Outbreak in Domestic Ruminants in the U.S.
In March 2024, juvenile goats with neurologic symptoms in an agriculture farm in Minnesota were tested positive for the HPAI H5N1 virus, which marked the first case of HPAI H5N1 infection in domestic ruminant in the U.S. [13]. The deceased goats were five to nine days of age and the HPAI H5N1 virus was detected in the brain, as well as in other tissues [14]. The poultry on the same farm was infected by the HPAI H5N1 virus in February and the virus might have spilled to goats as they shared the space and water on the farm. HPAI H5N1 virus was also tested in sick cattle in Texas in March 2024, the initial genomic analysis of the virus did not detect known antiviral resistance markers or substitutions related to mammalian host adaptation signature mutations [15]. HPAI H5N1 virus-infected cattle demonstrated gastrointestinal symptoms and a significant drop in milk production. Furthermore, the HPAI H5N1 virus was detected in their raw milk at high concentrations, and milk was also thickened or colostrum-like. The sick cattle also showed a loss of appetite, and it has been suggested that dairy cows might have contracted the virus from wild birds, given that the dead birds were found on the same farm property. The HPAI H5N1 virus causing an outbreak in cattle belongs to the 2.3.4.4b clade. As of May 16, 2024, the H5N1 virus was tested in dairy cow herds in nine states, 49 herds in total in the U.S. (Figure 1) [16].
Two unusual host infections that happened in March highlighted that the spring migration of wild birds may have played an important role in the virus spread. It is still unclear how these viruses were transmitted from birds to domestic ruminants. HPAI H5N1 viruses can replicate in epithelial cells of the gastrointestinal and respiratory tracts in birds, enabling viruses to transmit by both fecal-oral route and aerosols. Farm water and fomites may get contaminated by infected birds, which may be a plausible way for H5N1 virus transmission from birds to cattle. Alternatively, goats and cattle might get affected by the aerosolized virus. More studies are needed to address questions on virus transmission to cattle and goats from infected birds.
The symptoms of dead goats and sick cattle also need more attention. Influenza virus can cause gastrointestinal and respiratory illness in infected animals, but the goats and cattle demonstrated illness beyond that. The fact that goats showed neurologic symptoms and the virus was detected in the brain, suggests that the brain is a viral target brain. High levels of the H5N1 virus were detected in the raw milk of sick dairy cows which suggests that the virus can replicate efficiently in dairy cow mammary glands. Recently, Kristensen et al. reported that both human-type receptors and avian-type receptors are widely expressed in cattle mammary glands [17], which provides new insights into the HPAI H5N1 tropism in cow mammary glands.
There are some earlier studies showing that dairy cows can be infected by H1N1 and H3N2 viruses in the late 1990s [18,19]. The affected dairy cows showed a significant drop in milk production. In addition to the reduction in milk yield, IAV infected cattle also developed other clinical symptoms, like fever, lack of appetite, and discomfort. Some infected dairy cows also manifested nasal discharge or respiratory noise. Gunning et al. performed HI assays to evaluate the prevalence of IAV infections in cattle. Of 40 cattle investigated, 60% of cattle showed elevated antibody titers to the H1N1 virus and 65% of cattle showed elevated antibody titers to the H3N2 virus [18]. Crawshaw et al. further demonstrated the association of elevated titers to H1N1 and H3N2 viruses with reduced milk yield in cattle [20]. In this regard, the H5N1 infection in cattle with a significant drop in milk yield we observed today is not entirely new.
HPAI H5N1 emerging in the U.S. dairy cattle has posed a significant health threat to dairy farm workers. High levels of H5N1 viruses were detected in raw milk, which may be transmitted to the workers who have access to the raw milk. Besides, the infected cattle may shed the virus in feces and in the surrounding environment, further enhancing the health risk to dairy farm workers and other animals in the property. More importantly, the virus-contaminated milking machine and the environment may spread viruses to healthy cattle. Cattle transportation may allow the virus to further spread to other herds. Currently, the tested samples are limited in size and there is a concern that the H5N1 virus may spread in cattle more widely than initially thought as the virus was detected in lung sample of an asymptomatic cow. To date, there is no information available about the infection in beef and other cattle species, which needs immediate attention.
3. HPAI H5N1 Virus Infection in Mammals
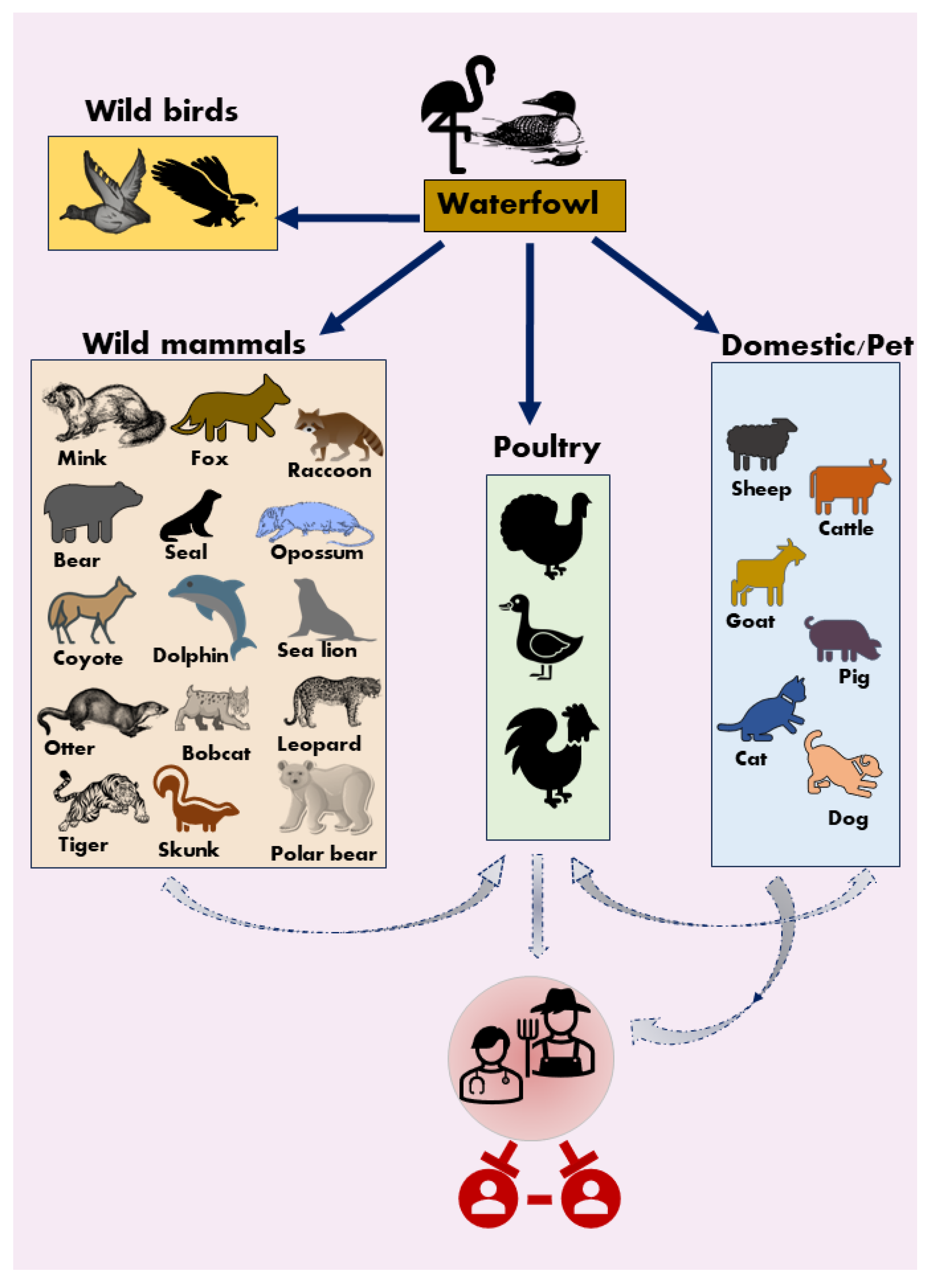
HPAI H5N1 virus can infect birds and poultry, it also can infect wild and domestic mammal species, such as sea lion, dolphin, fox, bobcat, Virginia opossum, raccoon, coyotes, striped skunk, seal, black bear, polar bear and tiger (Figure 2) [21,22,23,24,25,26,27]. Wild mammals may get infected by feeding on virus-infected birds and HPAI H5N1 virus infections can be fatal to these mammals. In 2004, two tigers and two leopards in a zoo in Thailand were reported to be infected by H5N1 HPAI virus after eating raw chicken possibly infected with the virus. These zoo animals showed high body temperature, increased nasal discharge, and neurologic manifestations before death [28]. In 2022, minks on a farm in Spain were detected positive for HPAI H5N1 virus, and further examination of the infected minks found lesions in the lungs [29]. In Finland, an otter was found dead because of HAPI H5N1 virus infection and the microscopic examination showed meningoencephalitis in the brain. Furthermore, foxes can be infected after eating HPAI H5N1 virus-infected birds and exhibit respiratory and neurologic symptoms [30,31]. In 2022, dolphin and sea lion were found positive for HPAI H5N1 in Peru and these animals were either dead or showed respiratory symptoms and/or neurological signs [32]. As discussed above, the goat infected by H5N1 HPAI virus also demonstrated neurologic symptoms. This study together with published work indicates that HPAI H5N1 virus can be neurotropic to mammals and cause a disorder in central nervous system (CNS). Similarly, a study performed in ferrets showed that HPAI H5N1 can enter the CNS through the olfactory nerve and replicate in this organ [33]. Viruses may acquire substitutions to adapt to the CNS and substantial virus production in the brain can result in severe meningoencephalitis in ferrets.
In addition to wild mammals, cats and dogs, can be infected by HPAI H5N1 virus and these infections can be fatal in some cases [34,35,36]. In June 2023, cats in Poland tested positive for HPAI H5N1 virus and the possible infection route is cat food [37]. Cats infected by HPAI H5N1 virus were also reported in Germany, the U.S., and South Korea [38,39,40]. In April 2024, cats on an H5N1 positive dairy cow farm in the U.S. were also infected with HPAI H5N1 virus [41]. Infection of cats might be due to consuming unpasteurized milk from virus-infected dairy cows. The virus infection is fatal to cats and the deceased cats from a Texas dairy farm showed meningoencephalitis, pneumonia, myocarditis and chorioretinitis. An experimental study on cats demonstrated that cats can be infected with the HPAI H5N1 virus by intratracheally inoculated virus or consuming virus-infected chickens [42]. The study also found that virus-infected cats can transmit the virus to other cats. Furthermore, the HPAI H5N1 virus can also infect dogs. One dog in Canada was confirmed to be infected by this virus in 2023 after chewing on a wild goose [43]. The infection in this dog is fatal and the necropsy demonstrated severe respiratory pathology. Taken together, these clinical cases and studies emphasize that consuming raw meat from dead birds can be an important source of cat and dog infections. The infections in these companion animals pose a potential health risk to their owners and veterinarians.
4. HPAI H5N1 Virus Infection in Humans
According to WHO, 873 persons have been infected by H5N1 virus worldwide and 458 of them lost their lives with the virus since 2003 [44]. The patients can develop respiratory symptoms, ranging from asymptotic or mild to severe respiratory symptoms including pneumonia. Besides respiratory symptoms, gastrointestinal symptoms such as diarrhea, was also frequently reported in patients. In a fatal case of H5N1 virus infection in a child in southern Vietnam, the patient showed severe diarrhea but no respiratory symptoms [45]. Further investigation demonstrated the virus infection resulted in acute encephalitis in the patient.
In April 2024, a dairy farm worker in Texas was tested positive for HPAI H5N1 virus after exposure to presumed H5N1 virus-infected dairy cows [46]. The only symptom is mild conjunctivitis. The first H5N1 human infection case was reported in 2022 in Colorado. The patient had direct exposure to H5N1 virus-infected poultry and fatigue was the only symptom noticed [47]. Conjunctivitis is a typical symptom in humans infected with HPAI viruses. Fouchier et al reported that 78 of 86 H7N7 virus-infected patients showed conjunctivitis [48]. Conjunctival route can serve as an entry portal for HPAI viruses as the ocular surface harbors alpha 2,3 linked sialic acid (avian-type receptor) [49]. Therefore, avian influenza virus can bind to these alpha 2,3 linked sialic acid and initiate the infection in eyes.
It is worth noting that the human specimen collected in 2024 from a H5N1 positive patient in Texas harbors a substitution (E627K) in PB2 which is not present in Texas dairy cattle and birds. The PB2 E627K substitution may emerge during replication in that patient. The substitution can enhance viral polymerase activity in human and other mammalian cells and is considered an important mammalian-adapting substitution [50]. Substitution PB2-E627K was also detected in the H5N1-infected otter and red fox. PB2-E627K was reported as the main determinant of virulence in the H5N1 virus [51]. A study on a mouse model suggested that PB2-627K enables the virus to replicate efficiently in the mammals’ upper and lower respiratory tracts [52].
Substitution PB2-D701N was detected in the virus from a dead lynx in Finland and it was also associated with enhanced viral replication in mammals [53]. This substitution was also detected in the H5N1-infected sea lion in Peru [32]. On March 29, 2023, a 53-year-old man was reported infected with HPAI H5N1 virus and the genomic analysis detected a D701N substitution in the virus PB2 segment [54]. PB2-D701N was reported to enhance viral polymerase activity in human cells as determined in a luciferase-minigene replication assay [55]. In addition to E627K and D701N in PB2, other substitutions such as T271A [29,56], may also increase viral polymerase activity in mammalian cells and are important for virus transmissibility in mammals including humans. The increased polymerase activity may facilitate HPAI virus replication in a new host. Of note, these adaptation substitutions can facilitate virus replication in mammals, but there is no clear evidence that these substitutions contribute to the transmission among mammals.
5. HA, NA, and NS also Determine Virus Tropism and Host Range
Viral hemagglutinin (HA) can bind sialic acid (Sia) receptors on the cell surface to initiate the infection process and substitutions in HA can alter the receptor binding affinity, therefore affecting viral replication, transmission, and host range. Influenza viruses circulating in avian species usually bind to alpha-2,3 linkage Sia receptors whereas viruses in humans usually bind to alpha-2,6 linkage. Substitutions in HA may change the virus preference of Sia receptor linkage, which is exemplified by the virus strain caused the human pandemic in 1968. The avian virus can infect humans after acquiring six amino acid substitutions in the HA [57]. The loss of glycosylation at HA position 158 can increase alpha-2,6 sialic acid receptor binding affinity. Gao et al. reported that T160A substitution in HA causing the loss of the glycosylation site at HA-158-160 can affect the viral pathogenicity in mice [58]. Furthermore, Jang et al. showed that another substitution in H5 HA, N193D, can enhance the binding to alpha-2,6 linage [59].
Neuraminidase (NA) is another glycoprotein on the virus surface, and it acts as a sialidase enzyme to cleave sialic acid from the host cell surface and facilitate the release of newly assembled progeny virions from infected cells. NA possesses a second sialic acid-binding site (2SBS) which is important for HA-NA balance and virus replication [60]. Of note, 2SBS is highly conserved in avian IAVs whereas it is not present in human viruses. 2SBS binds to alpha-2,3 linked Sia receptors, but substitution in 2SBS can reduce the binding to alpha-2,3 linked Sia receptors and may enhance the binding to alpha-2,6 linked Sia receptors. The HPAI H5N1 virus responsible for numerous deaths of farmed mink has disrupted 2SBS [61]. The loss of 2SBS results in the enhanced binding to human-type receptors, therefore, 2SBS serves as an important determinant of virus host range.
The non-structural protein 1 (NS1) has been demonstrated to be important for virulence, pathogenicity, and host range [62,63]. NS1 can enhance the expression level of cytokines and limit the induction of interferon (IFN)-beta by multiple pathways, thereby inhibiting the host antiviral responses. The deletion of the NS1 gene in the influenza A virus (A/PR/8/34) results in reduced replication in embryonated chicken eggs because of the IFN-mediated antiviral effects [64]. A recombinant H7N1 HPAI virus with NS segment replaced by the H5N1 virus NS replicates more efficiently in human cell lines and induces stronger pathogenicity in chicken embryos compared to the wild-type virus [60]. In another study, a recombinant H1N1 virus with the H5N1 NS gene conferred more prolonged clinical manifestations in pigs compared to the wild-type virus. The effect was conferred by a D92E substitution in NS1 [65,66]. The amino acid at position 149 of NS1 was also reported to impact the pathogenicity of the H5N1 virus in chickens. A recombinant H5N1 virus A/goose/Guangdong/1/96 with Ala at 149 position of NS1 can limit the interferon protein expression in chicken embryo fibroblasts, whereas virus with Val at 149 position cannot [66]. More studies are needed to investigate the NS1 determinants of adaptation to mammals.
6. Vaccines against HPAI H5N1
The reverse genetics approach provides a novel and important way to investigate virus gene functions, virus virulence, transmission, and antivirals [6,67,68,69,70,71,72,73]. Moreover, the reverse genetics approach is also critical to rapid generation of vaccine strains [74,75,76]. Reverse genetics-based vaccines were developed and demonstrated high protective efficacy in influenza viruses, including HPAI H5N1 viruses [74,76,77,78,79,80]. Webby et al. used a reverse genetics approach to generate a candidate reference virus strain in response to a pandemic alert issued by WHO in 2003 [76].
Commercial vaccines against H5N1 for poultry are currently not used in US poultry. The HPAI H5N1 virus affected chickens were culled to eradicate viruses and its poultry reservoir. Vaccines were used to protect endangered California condor from HPAI H5N1 infections, which marked the first case that U.S. vaccinated any endangered bird species to curb HPAI virus infections [81]. Vaccinations are used against H5N1 HPAI in China, Egypt, Indonesia, and Vietnam [82,83,84,85]. Because of virus evolution, vaccine strains need to be updated timely to maintain vaccine protective efficacy [86]. Vaccines against HPAI H5N1 viruses were shown to prevent illness in chickens, geese, and ducks, and these vaccines can reduce virus replication in gastrointestinal and respiratory tracts [87]. The implementation of vaccines against HPAI H5N1 can reduce outbreaks in poultry farm, decreasing the chance of human and wild bird exposure to HPAI virus [82]. Poultry vaccination can help control of HPAI in wild birds as vaccine-mediated protective immune responses can reduce viral shedding in the environment. USDA has worked on the evaluation of the efficacy of four vaccine candidates against HPAI in poultry [88].
It is feared that the adaptation of the HPAI H5N1 virus to dairy cattle may result in human-to-human transmission. Influenza vaccines against seasonal influenza viruses cannot provide any protection against H5N1 virus infections, highlighting the need for a vaccine against H5N1 infections in humans. Although no substitutions linked to human-to-human transmission were detected in the virus sequences, the pandemic potential of the H5N1 virus cannot be overlooked given the high mutation rate that can readily occur and accumulate during virus replication over time. As an important activity of pandemic preparedness, the Centers for Disease Control and Prevention (CDC) has developed an H5 candidate vaccine virus (CVV) which can be used for the production of human vaccines if needed [89]. It should be kept in mind that vaccines against HPAI H5N1 viruses may need to be updated due to antigenic drift or shift associated with mobile influenza viruses. Therefore, virus surveillance and antigenic analysis are critically needed to select a candidate vaccine strain and maintain vaccine optimal efficacy.
7. Conclusion and Future Directions
Suddenly emerging of HPAI H5N1 clade 2.3.4.4b viruses in dairy cows, and in milk samples in multiple U.S. states raise several immediate questions that need to be addressed in future research.
First, among influenza A subtypes, is H5N1 clade 2.3.4.4b only a group of the virus capable of jumping to dairy cows from birds and causing sustainable cow-to-cow transmission? Several studies published in the late 1990s have provided good evidence that other subtypes of influenza A virus such as H3 could infect dairy cows [19] and cause a drop in milk production and other symptoms like what has been observed in the U.S. dairy cows infected with HPAI H5N1 virus. In this regard, infection of dairy cows by an influenza A virus is not entirely new. The uniqueness of this influenza endemic in cattle lies in BSL3 agent, HPAI H5N1 with the potential to cause a human pandemic if human adaptation mutations are acquired. A comprehensive investigation of other subtypes of IAV in addition to H5N1 is needed to determine their ability in infecting and causing a drop in milk production in dairy cows. Identifying the potential differences between different strains or subtypes may lay the foundation for future research toward identifying cattle adaptation mutations.
Second, despite the mammary gland being one of the established in vivo targets for H5N1 clade 2.3.4.4b, the virus was detected in nasal shedding as well as in the lungs of infected cattle. Asymptomatic cows appear to be responsible for the widespread of this disease in U.S. dairy cow farms. It has been also shown that the virus was detected in the lungs of clinically normal cows. It can be envisioned that the H5N1 clade 2.3.4.4b has a very broad tropism ranging from the respiratory and gastrointestinal tracts to the mammary gland tissues, which warrants further investigation. Also, it remains unclear whether beef cattle breeds and heifers are susceptible to HPAI H5N1 infection. More work is needed to define tissue tropism and the host range of this emerging virus in cattle.
Third, some gathered evidence suggests that milk or contaminated equipment in dairy cow farms plays an important role in cow-to-cow transmission. Whether the major mode of transmission occurs through the fecal-oral route or aerosol droplet during the milk collection process is not well known. Considering the virus detected from nasal shedding and lung tissues, addressing the transmission through the classical respiratory tract is also needed. A significant amount of viral load is found in milk indicating that the mammary gland is a primary target organ where HPAI H5N1 replicates. Whether the virus reaches the mammary gland through the blood-milk barrier (systemic infection and viremia) or a local route warrants future investigation.
Finally, bovine immunity especially its antibody responses to influenza virus infection has not been well studied in the past due to a long-time concept that cattle are not a susceptible host to influenza A virus. Several studies on bovine influenza D viruses showed that persistent infections frequently occur in bovine herds and preexisting immunity is somehow short-lived [90]. Investigation of the durability and potency of protective antibody responses against HPAI H5N1 in dairy cows should address this important question, which will provide insights into the successful implementation of vaccine strategies, which is critical to the control and prevention of future influenza epidemics in the cattle industry.
Author Contributions
Writing-original draft preparation, R.G.; Writing-review and editing, C.C.S., F.L. and D.W.; Illustrations: C.C.S. and C.F.; Supervision, F.L. and D.W. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by the William Robert Mills Endowed Chair Fund.
Acknowledgement
The members of the Wang-Li lab are particularly appreciated for their comments of this manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Yoon, S.W.; Webby, R.J.; Webster, R.G. Evolution and ecology of influenza A viruses. Curr Top Microbiol Immunol 2014, 385, 359–375. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y. Pathogenicity and virulence of influenza. Virulence 2023, 14, 2223057. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, Y.; Katsushima, N.; Nagai, Y.; Shoji, M.; Itagaki, T.; Sakamoto, M.; Kitaoka, S.; Mizuta, K.; Nishimura, H. Clinical features of influenza C virus infection in children. J Infect Dis 2006, 193, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Sheng, Z.; Lin, T.; Sreenivasan, C.; Gao, R.; Thomas, M.; Druce, J.; Hause, B.M.; Kaushik, R.S.; Li, F.; et al. Genetic and antigenic characteristics of a human influenza C virus clinical isolate. J Med Virol 2020, 92, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Sheng, Z.; Huang, C.; Wang, D.; Li, F. Influenza D virus. Curr Opin Virol 2020, 44, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Liu, R.; Zhou, B.; Chou, T.W.; Ghedin, E.; Sheng, Z.; Gao, R.; Zhai, S.L.; Wang, D.; Li, F. Development and Characterization of a Reverse-Genetics System for Influenza D Virus. J Virol 2019, 93. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasan, C.C.; Liu, R.; Gao, R.; Guo, Y.; Hause, B.M.; Thomas, M.; Naveed, A.; Clement, T.; Rausch, D.; Christopher-Hennings, J.; et al. Influenza C and D Viruses Demonstrated a Differential Respiratory Tissue Tropism in a Comparative Pathogenesis Study in Guinea Pigs. J Virol 2023, 97, e0035623. [Google Scholar] [CrossRef] [PubMed]
- Sreenivasan, C.C.; Thomas, M.; Kaushik, R.S.; Wang, D.; Li, F. Influenza A in Bovine Species: A Narrative Literature Review. Viruses 2019, 11. [Google Scholar] [CrossRef] [PubMed]
- Suarez, D.L.; Schultz-Cherry, S. Immunology of avian influenza virus: a review. Dev Comp Immunol 2000, 24, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Neumann, G.; Chen, H.; Gao, G.F.; Shu, Y.; Kawaoka, Y. H5N1 influenza viruses: outbreaks and biological properties. Cell Res 2010, 20, 51–61. [Google Scholar] [CrossRef] [PubMed]
- H5N1 Bird Flu Detections across the United States (Backyard and Commercial). Availabe online: https://www.cdc.gov/flu/avianflu/data-map-commercial.html (accessed on April, 23,2024).
- Avian Influenza in Birds. Availabe online: https://www.cdc.gov/flu/avianflu/avian-in-birds.htm (accessed on April,23, 2024).
- Goat in Minnesota tests positive for HPAI. Availabe online: https://www.avma.org/news/goat-minnesota-tests-positive-hpai (accessed on April,24,2024).
- United States of America - Influenza A viruses of high pathogenicity (Inf. with) (non-poultry including wild birds) (2017-) - Follow up report 43. Availabe online: https://wahis.woah.org/#/in-review/4451?reportId=166488&fromPage=event-dashboard-url (accessed on April, 24,2024).
- United States of America - Influenza A viruses of high pathogenicity (Inf. with) (non-poultry including wild birds) (2017-) - Follow up report 44. Availabe online: https://wahis.woah.org/#/in-review/4451?reportId=166639&fromPage=event-dashboard-url (accessed on April 24,2024).
- Highly Pathogenic Avian Influenza (HPAI) Detections in Livestock. Availabe online: https://www.aphis.usda.gov/livestock-poultry-disease/avian/avian-influenza/hpai-detections/livestock (accessed on May,2nd, 2024).
- Kristensen, C.; Larsen, L.E.; Trebbien, R.; Jensen, H.E. The avian influenza A virus receptor SA-alpha2,3-Gal is expressed in the porcine nasal mucosa sustaining the pig as a mixing vessel for new influenza viruses. Virus Res 2024, 340, 199304. [Google Scholar] [CrossRef] [PubMed]
- Gunning, R.F.; Brown, I.H.; Crawshaw, T.R. Evidence of influenza A virus infection in dairy cows with sporadic milk drop syndrome. Vet Rec 1999, 145, 556–557. [Google Scholar] [CrossRef] [PubMed]
- Brown, I.H.; Crawshaw, T.R.; Harris, P.A.; Alexander, D.J. Detection of antibodies to influenza A virus in cattle in association with respiratory disease and reduced milk yield. Vet Rec 1998, 143, 637–638. [Google Scholar] [PubMed]
- Crawshaw, T.R.; Brown, I.H.; Essen, S.C.; Young, S.C. Significant rising antibody titres to influenza A are associated with an acute reduction in milk yield in cattle. Vet J 2008, 178, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Ulloa, M.; Fernandez, A.; Ariyama, N.; Colom-Rivero, A.; Rivera, C.; Nunez, P.; Sanhueza, P.; Johow, M.; Araya, H.; Torres, J.C.; et al. Mass mortality event in South American sea lions (Otaria flavescens) correlated to highly pathogenic avian influenza (HPAI) H5N1 outbreak in Chile. Vet Q 2023, 43, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Murawski, A.; Fabrizio, T.; Ossiboff, R.; Kackos, C.; Jeevan, T.; Jones, J.C.; Kandeil, A.; Walker, D.; Turner, J.C.M.; Patton, C.; et al. Highly pathogenic avian influenza A(H5N1) virus in a common bottlenose dolphin (Tursiops truncatus) in Florida. Commun Biol 2024, 7, 476. [Google Scholar] [CrossRef] [PubMed]
- Keawcharoen, J.; Oraveerakul, K.; Kuiken, T.; Fouchier, R.A.; Amonsin, A.; Payungporn, S.; Noppornpanth, S.; Wattanodorn, S.; Theambooniers, A.; Tantilertcharoen, R.; et al. Avian influenza H5N1 in tigers and leopards. Emerg Infect Dis 2004, 10, 2189–2191. [Google Scholar] [CrossRef] [PubMed]
- Jakobek, B.T.; Berhane, Y.; Nadeau, M.S.; Embury-Hyatt, C.; Lung, O.; Xu, W.; Lair, S. Influenza A(H5N1) Virus Infections in 2 Free-Ranging Black Bears (Ursus americanus), Quebec, Canada. Emerg Infect Dis 2023, 29, 2145–2149. [Google Scholar] [CrossRef]
- Puryear, W.; Sawatzki, K.; Hill, N.; Foss, A.; Stone, J.J.; Doughty, L.; Walk, D.; Gilbert, K.; Murray, M.; Cox, E.; et al. Highly Pathogenic Avian Influenza A(H5N1) Virus Outbreak in New England Seals, United States. Emerg Infect Dis 2023, 29, 786–791. [Google Scholar] [CrossRef] [PubMed]
- Elsmo, E.J.; Wunschmann, A.; Beckmen, K.B.; Broughton-Neiswanger, L.E.; Buckles, E.L.; Ellis, J.; Fitzgerald, S.D.; Gerlach, R.; Hawkins, S.; Ip, H.S.; et al. Highly Pathogenic Avian Influenza A(H5N1) Virus Clade 2.3.4.4b Infections in Wild Terrestrial Mammals, United States, 2022. Emerg Infect Dis 2023, 29, 2451–2460. [Google Scholar] [CrossRef] [PubMed]
- Plaza, P.I.; Gamarra-Toledo, V.; Rodriguez Eugui, J.; Rosciano, N.; Lambertucci, S.A. Pacific and Atlantic sea lion mortality caused by highly pathogenic Avian Influenza A(H5N1) in South America. Travel Med Infect Dis 2024, 59, 102712. [Google Scholar] [CrossRef] [PubMed]
- Thanawongnuwech, R.; Amonsin, A.; Tantilertcharoen, R.; Damrongwatanapokin, S.; Theamboonlers, A.; Payungporn, S.; Nanthapornphiphat, K.; Ratanamungklanon, S.; Tunak, E.; Songserm, T.; et al. Probable tiger-to-tiger transmission of avian influenza H5N1. Emerg Infect Dis 2005, 11, 699–701. [Google Scholar] [CrossRef] [PubMed]
- Aguero, M.; Monne, I.; Sanchez, A.; Zecchin, B.; Fusaro, A.; Ruano, M.J.; Del Valle Arrojo, M.; Fernandez-Antonio, R.; Souto, A.M.; Tordable, P.; et al. Highly pathogenic avian influenza A(H5N1) virus infection in farmed minks, Spain, October 2022. Euro Surveill 2023, 28. [Google Scholar] [CrossRef]
- Bordes, L.; Vreman, S.; Heutink, R.; Roose, M.; Venema, S.; Pritz-Verschuren, S.B.E.; Rijks, J.M.; Gonzales, J.L.; Germeraad, E.A.; Engelsma, M.; et al. Highly Pathogenic Avian Influenza H5N1 Virus Infections in Wild Red Foxes (Vulpes vulpes) Show Neurotropism and Adaptive Virus Mutations. Microbiol Spectr 2023, 11, e0286722. [Google Scholar] [CrossRef] [PubMed]
- Reperant, L.A.; van Amerongen, G.; van de Bildt, M.W.; Rimmelzwaan, G.F.; Dobson, A.P.; Osterhaus, A.D.; Kuiken, T. Highly pathogenic avian influenza virus (H5N1) infection in red foxes fed infected bird carcasses. Emerg Infect Dis 2008, 14, 1835–1841. [Google Scholar] [CrossRef] [PubMed]
- Leguia, M.; Garcia-Glaessner, A.; Munoz-Saavedra, B.; Juarez, D.; Barrera, P.; Calvo-Mac, C.; Jara, J.; Silva, W.; Ploog, K.; Amaro, L.; et al. Highly pathogenic avian influenza A (H5N1) in marine mammals and seabirds in Peru. Nat Commun 2023, 14, 5489. [Google Scholar] [CrossRef] [PubMed]
- Siegers, J.Y.; Ferreri, L.; Eggink, D.; Veldhuis Kroeze, E.J.B.; Te Velthuis, A.J.W.; van de Bildt, M.; Leijten, L.; van Run, P.; de Meulder, D.; Bestebroer, T.; et al. Evolution of highly pathogenic H5N1 influenza A virus in the central nervous system of ferrets. PLoS Pathog 2023, 19, e1011214. [Google Scholar] [CrossRef] [PubMed]
- Ly, H. Highly pathogenic avian influenza H5N1 virus infection of companion animals. Virulence 2024, 15, 2289780. [Google Scholar] [CrossRef] [PubMed]
- Thiry, E.; Zicola, A.; Addie, D.; Egberink, H.; Hartmann, K.; Lutz, H.; Poulet, H.; Horzinek, M.C. Highly pathogenic avian influenza H5N1 virus in cats and other carnivores. Vet Microbiol 2007, 122, 25–31. [Google Scholar] [CrossRef]
- Szalus-Jordanow, O.; Golke, A.; Dzieciatkowski, T.; Czopowicz, M.; Kardas, M.; Mickiewicz, M.; Moroz-Fik, A.; Lobaczewski, A.; Markowska-Daniel, I.; Frymus, T. Upper Respiratory Tract Disease in a Dog Infected by a Highly Pathogenic Avian A/H5N1 Virus. Microorganisms 2024, 12. [Google Scholar] [CrossRef] [PubMed]
- Rabalski, L.; Milewska, A.; Pohlmann, A.; Gackowska, K.; Lepionka, T.; Szczepaniak, K.; Swiatalska, A.; Sieminska, I.; Arent, Z.; Beer, M.; et al. Emergence and potential transmission route of avian influenza A (H5N1) virus in domestic cats in Poland, June 2023. Euro Surveill 2023, 28. [Google Scholar] [CrossRef] [PubMed]
- Klopfleisch, R.; Wolf, P.U.; Uhl, W.; Gerst, S.; Harder, T.; Starick, E.; Vahlenkamp, T.W.; Mettenleiter, T.C.; Teifke, J.P. Distribution of lesions and antigen of highly pathogenic avian influenza virus A/Swan/Germany/R65/06 (H5N1) in domestic cats after presumptive infection by wild birds. Vet Pathol 2007, 44, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Yeom, M.; Vu, T.T.H.; Do, H.Q.; Na, W.; Lee, M.; Jeong, D.G.; Cheon, D.S.; Song, D. Characterization of highly pathogenic avian influenza A (H5N1) viruses isolated from cats in South Korea, 2023. Emerg Microbes Infect 2024, 13, 2290835. [Google Scholar] [CrossRef]
- Sillman, S.J.; Drozd, M.; Loy, D.; Harris, S.P. Naturally occurring highly pathogenic avian influenza virus H5N1 clade 2.3.4.4b infection in three domestic cats in North America during 2023. J Comp Pathol 2023, 205, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Burrough, E.; Magstadt, D.; Petersen, B.; Timmermans, S.; Gauger, P.; Zhang, J.; Siepker, C.; Mainenti, M.; Li, G.; Thompson, A.; et al. Highly Pathogenic Avian Influenza A(H5N1) Clade 2.3.4.4b Virus Infection in Domestic Dairy Cattle and Cats, United States, 2024. Emerging Infectious Disease journal 2024, 30. [Google Scholar] [CrossRef]
- Kuiken, T.; Rimmelzwaan, G.; van Riel, D.; van Amerongen, G.; Baars, M.; Fouchier, R.; Osterhaus, A. Avian H5N1 influenza in cats. Science 2004, 306, 241. [Google Scholar] [CrossRef] [PubMed]
- Domestic dog tests positive for avian influenza in Canada Availabe online: https://www.canada.ca/en/food-inspection-agency/news/2023/04/domestic-dog-tests-positive-for-avian-influenza-in-canada.html (accessed on April 25,2024).
- Human Infection caused by Avian Influenza A (H5N1) - Chile. Availabe online: https://www.who.int/emergencies/disease-outbreak-news/item/2023-DON461 (accessed on April 25, 2024).
- de Jong, M.D.; Bach, V.C.; Phan, T.Q.; Vo, M.H.; Tran, T.T.; Nguyen, B.H.; Beld, M.; Le, T.P.; Truong, H.K.; Nguyen, V.V.; et al. Fatal avian influenza A (H5N1) in a child presenting with diarrhea followed by coma. N Engl J Med 2005, 352, 686–691. [Google Scholar] [CrossRef] [PubMed]
- Uyeki, T.M.; Milton, S.; Abdul Hamid, C.; Reinoso Webb, C.; Presley, S.M.; Shetty, V.; Rollo, S.N.; Martinez, D.L.; Rai, S.; Gonzales, E.R.; et al. Highly Pathogenic Avian Influenza A(H5N1) Virus Infection in a Dairy Farm Worker. N Engl J Med, 2024. [Google Scholar] [CrossRef]
- State health officials investigate a detection of H5 influenza virus in a human in Colorado. Availabe online: https://cdphe.colorado.gov/press-release/state-health-officials-investigate-a-detection-of-h5-influenza-virus-in-a-human (accessed on April 25, 2024).
- Fouchier, R.A.; Schneeberger, P.M.; Rozendaal, F.W.; Broekman, J.M.; Kemink, S.A.; Munster, V.; Kuiken, T.; Rimmelzwaan, G.F.; Schutten, M.; Van Doornum, G.J.; et al. Avian influenza A virus (H7N7) associated with human conjunctivitis and a fatal case of acute respiratory distress syndrome. Proc Natl Acad Sci U S A 2004, 101, 1356–1361. [Google Scholar] [CrossRef]
- Belser, J.A.; Lash, R.R.; Garg, S.; Tumpey, T.M.; Maines, T.R. The eyes have it: influenza virus infection beyond the respiratory tract. Lancet Infect Dis 2018, 18, e220–e227. [Google Scholar] [CrossRef] [PubMed]
- Taft, A.S.; Ozawa, M.; Fitch, A.; Depasse, J.V.; Halfmann, P.J.; Hill-Batorski, L.; Hatta, M.; Friedrich, T.C.; Lopes, T.J.; Maher, E.A.; et al. Identification of mammalian-adapting mutations in the polymerase complex of an avian H5N1 influenza virus. Nat Commun 2015, 6, 7491. [Google Scholar] [CrossRef] [PubMed]
- Hatta, M.; Gao, P.; Halfmann, P.; Kawaoka, Y. Molecular basis for high virulence of Hong Kong H5N1 influenza A viruses. Science 2001, 293, 1840–1842. [Google Scholar] [CrossRef] [PubMed]
- Hatta, M.; Hatta, Y.; Kim, J.H.; Watanabe, S.; Shinya, K.; Nguyen, T.; Lien, P.S.; Le, Q.M.; Kawaoka, Y. Growth of H5N1 influenza A viruses in the upper respiratory tracts of mice. PLoS Pathog 2007, 3, 1374–1379. [Google Scholar] [CrossRef] [PubMed]
- Tammiranta, N.; Isomursu, M.; Fusaro, A.; Nylund, M.; Nokireki, T.; Giussani, E.; Zecchin, B.; Terregino, C.; Gadd, T. Highly pathogenic avian influenza A (H5N1) virus infections in wild carnivores connected to mass mortalities of pheasants in Finland. Infect Genet Evol 2023, 111, 105423. [Google Scholar] [CrossRef] [PubMed]
- Human Infection with highly pathogenic avian influenza A(H5N1) virus in Chile. Availabe online: https://www.cdc.gov/flu/avianflu/spotlights/2022-2023/chile-first-case-h5n1-addendum.htm (accessed on April 25, 2024).
- Gabriel, G.; Dauber, B.; Wolff, T.; Planz, O.; Klenk, H.D.; Stech, J. The viral polymerase mediates adaptation of an avian influenza virus to a mammalian host. Proc Natl Acad Sci U S A 2005, 102, 18590–18595. [Google Scholar] [CrossRef]
- Bussey, K.A.; Bousse, T.L.; Desmet, E.A.; Kim, B.; Takimoto, T. PB2 residue 271 plays a key role in enhanced polymerase activity of influenza A viruses in mammalian host cells. J Virol 2010, 84, 4395–4406. [Google Scholar] [CrossRef]
- Bean, W.J.; Schell, M.; Katz, J.; Kawaoka, Y.; Naeve, C.; Gorman, O.; Webster, R.G. Evolution of the H3 influenza virus hemagglutinin from human and nonhuman hosts. J Virol 1992, 66, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Gu, M.; Liu, K.; Li, Q.; Li, J.; Shi, L.; Li, X.; Wang, X.; Hu, J.; Liu, X.; et al. T160A mutation-induced deglycosylation at site 158 in hemagglutinin is a critical determinant of the dual receptor binding properties of clade 2.3.4.4 H5NX subtype avian influenza viruses. Vet Microbiol 2018, 217, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.G.; Kim, Y.I.; Casel, M.A.B.; Choi, J.H.; Gil, J.R.; Rollon, R.; Kim, E.H.; Kim, S.M.; Ji, H.Y.; Park, D.B.; et al. HA N193D substitution in the HPAI H5N1 virus alters receptor binding affinity and enhances virulence in mammalian hosts. Emerg Microbes Infect 2024, 13, 2302854. [Google Scholar] [CrossRef] [PubMed]
- Du, W.; de Vries, E.; van Kuppeveld, F.J.M.; Matrosovich, M.; de Haan, C.A.M. Second sialic acid-binding site of influenza A virus neuraminidase: binding receptors for efficient release. FEBS J 2021, 288, 5598–5612. [Google Scholar] [CrossRef] [PubMed]
- de Vries, E.; de Haan, C.A. Letter to the editor: Highly pathogenic influenza A(H5N1) viruses in farmed mink outbreak contain a disrupted second sialic acid binding site in neuraminidase, similar to human influenza A viruses. Euro Surveill 2023, 28. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Brenner, D.; Wang, Z.; Dauber, B.; Ehrhardt, C.; Hogner, K.; Herold, S.; Ludwig, S.; Wolff, T.; Yu, K.; et al. The NS segment of an H5N1 highly pathogenic avian influenza virus (HPAIV) is sufficient to alter replication efficiency, cell tropism, and host range of an H7N1 HPAIV. J Virol 2010, 84, 2122–2133. [Google Scholar] [CrossRef] [PubMed]
- Hale, B.G.; Randall, R.E.; Ortin, J.; Jackson, D. The multifunctional NS1 protein of influenza A viruses. J Gen Virol 2008, 89, 2359–2376. [Google Scholar] [CrossRef]
- Garcia-Sastre, A.; Egorov, A.; Matassov, D.; Brandt, S.; Levy, D.E.; Durbin, J.E.; Palese, P.; Muster, T. Influenza A virus lacking the NS1 gene replicates in interferon-deficient systems. Virology 1998, 252, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.H.; Hoffmann, E.; Webster, R.G. The NS1 gene of H5N1 influenza viruses circumvents the host anti-viral cytokine responses. Virus Res 2004, 103, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jiang, Y.; Jiao, P.; Wang, A.; Zhao, F.; Tian, G.; Wang, X.; Yu, K.; Bu, Z.; Chen, H. The NS1 gene contributes to the virulence of H5N1 avian influenza viruses. J Virol 2006, 80, 11115–11123. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Wang, Z.; Uprety, T.; Sreenivasan, C.C.; Sheng, Z.; Hause, B.M.; Brunick, C.; Wu, H.; Luke, T.; Bausch, C.L.; et al. A fully human monoclonal antibody possesses antibody-dependent cellular cytotoxicity (ADCC) activity against the H1 subtype of influenza A virus by targeting a conserved epitope at the HA1 protomer interface. J Med Virol 2023, 95, e28901. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Pascua, P.N.Q.; Nguyen, H.T.; Chesnokov, A.; Champion, C.; Mishin, V.P.; Wentworth, D.E.; Gubareva, L.V. New insights into the neuraminidase-mediated hemagglutination activity of influenza A(H3N2) viruses. Antiviral Res 2023, 218, 105719. [Google Scholar] [CrossRef]
- Yu, J.; Sreenivasan, C.; Sheng, Z.; Zhai, S.L.; Wollman, J.W.; Luo, S.; Huang, C.; Gao, R.; Wang, Z.; Kaushik, R.S.; et al. A recombinant chimeric influenza virus vaccine expressing the consensus H3 hemagglutinin elicits broad hemagglutination inhibition antibodies against divergent swine H3N2 influenza viruses. Vaccine 2023, 41, 6318–6326. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Sun, C.; Gao, R.; Wang, H.; Liu, W.; Yu, K.; Zhou, G.; Zhao, B.; Yu, L. A Temperature-Dependent Translation Defect Caused by Internal Ribosome Entry Site Mutation Attenuates Foot-and-Mouth Disease Virus: Implications for Rational Vaccine Design. J Virol 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Yang, D.; Gao, R.; Liang, T.; Wang, H.; Zhou, G.; Yu, L. Modification of the internal ribosome entry site element impairs the growth of foot-and-mouth disease virus in porcine-derived cells. J Gen Virol 2016, 97, 901–911. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Sreenivasan, C.C.; Sheng, Z.; Hause, B.M.; Zhou, B.; Wentworth, D.E.; Clement, T.; Rausch, D.; Brunick, C.; Christopher-Hennings, J.; et al. Human Monoclonal Antibody Derived from Transchromosomic Cattle Neutralizes Multiple H1 Clades of Influenza A Virus by Recognizing a Novel Conformational Epitope in the Hemagglutinin Head Domain. J Virol 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Nogales, A.; Martinez-Sobrido, L. Reverse Genetics Approaches for the Development of Influenza Vaccines. Int J Mol Sci 2016, 18. [Google Scholar] [CrossRef] [PubMed]
- Govorkova, E.A.; Webby, R.J.; Humberd, J.; Seiler, J.P.; Webster, R.G. Immunization with reverse-genetics-produced H5N1 influenza vaccine protects ferrets against homologous and heterologous challenge. J Infect Dis 2006, 194, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Neumann, G.; Fujii, K.; Kino, Y.; Kawaoka, Y. An improved reverse genetics system for influenza A virus generation and its implications for vaccine production. Proc Natl Acad Sci U S A 2005, 102, 16825–16829. [Google Scholar] [CrossRef] [PubMed]
- Webby, R.J.; Perez, D.R.; Coleman, J.S.; Guan, Y.; Knight, J.H.; Govorkova, E.A.; McClain-Moss, L.R.; Peiris, J.S.; Rehg, J.E.; Tuomanen, E.I.; et al. Responsiveness to a pandemic alert: use of reverse genetics for rapid development of influenza vaccines. Lancet 2004, 363, 1099–1103. [Google Scholar] [CrossRef]
- Panickan, S.; Bhatia, S.; Bhat, S.; Bhandari, N.; Pateriya, A.K.; Kalaiyarasu, S.; Sood, R.; Tripathi, M. Reverse genetics based H5N2 vaccine provides clinical protection against H5N1, H5N8 and H9N2 avian influenza infection in chickens. Vaccine 2022, 40, 6998–7008. [Google Scholar] [CrossRef] [PubMed]
- Neumann, G. Influenza Reverse Genetics-Historical Perspective. Cold Spring Harb Perspect Med 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Tian, G.; Zeng, X.; Li, Y.; Shi, J.; Chen, H. Protective efficacy of the H5 inactivated vaccine against different highly pathogenic H5N1 avian influenza viruses isolated in China and Vietnam. Avian Dis 2010, 54, 287–289. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.; Zhang, Q.; Gu, C.; Shi, J.; Deng, G.; Ma, S.; Liu, J.; Chen, P.; Guan, Y.; Jiang, Y.; et al. A live attenuated vaccine prevents replication and transmission of H7N9 virus in mammals. Sci Rep 2015, 5, 11233. [Google Scholar] [CrossRef]
- Kozlov, M. US will vaccinate birds against avian flu for first time - what researchers think. Nature 2023, 618, 220–221. [Google Scholar] [CrossRef] [PubMed]
- Swayne, D.E.; Spackman, E.; Pantin-Jackwood, M. Success factors for avian influenza vaccine use in poultry and potential impact at the wild bird-agricultural interface. Ecohealth 2014, 11, 94–108. [Google Scholar] [CrossRef] [PubMed]
- Abdelwhab, E.M.; Hafez, H.M. An overview of the epidemic of highly pathogenic H5N1 avian influenza virus in Egypt: epidemiology and control challenges. Epidemiol Infect 2011, 139, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Desvaux, S.; Grosbois, V.; Pham, T.T.; Dao, D.T.; Nguyen, T.D.; Fenwick, S.; Roger, F.; Ellis, T.; Peyre, M. Evaluation of the vaccination efficacy against H5N1 in domestic poultry in the Red River Delta in Vietnam. Epidemiol Infect 2013, 141, 776–788. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zeng, X.; Cui, P.; Yan, C.; Chen, H. Alarming situation of emerging H5 and H7 avian influenza and effective control strategies. Emerg Microbes Infect 2023, 12, 2155072. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Bai, X.; Li, M.; Zeng, X.; Xu, J.; Li, P.; Wang, M.; Song, X.; Zhao, Z.; Tian, G.; et al. Highly Pathogenic Avian Influenza Virus (H5N1) Clade 2.3.4.4b Introduced by Wild Birds, China, 2021. Emerg Infect Dis 2023, 29, 1367–1375. [Google Scholar] [CrossRef]
- Swayne, D.E. Principles for vaccine protection in chickens and domestic waterfowl against avian influenza: emphasis on Asian H5N1 high pathogenicity avian influenza. Ann N Y Acad Sci 2006, 1081, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Fact Sheet: USDA Continues Partner Engagement to Mitigate Highly Pathogenic Avian Influenza for 2023 Season. Availabe online: https://www.usda.gov/media/press-releases/2023/04/14/fact-sheet-usda-continues-partner-engagement-mitigate-highly (accessed on April 27, 2024).
- Prevention and Antiviral Treatment of Bird Flu Viruses in People. Availabe online: https://www.cdc.gov/flu/avianflu/prevention.htm#anchor_1647619820462 (accessed on April 27, 2024).
- Wan, X.F.; Ferguson, L.; Oliva, J.; Rubrum, A.; Eckard, L.; Zhang, X.; Woolums, A.R.; Lion, A.; Meyer, G.; Murakami, S.; et al. Limited Cross-Protection Provided by Prior Infection Contributes to High Prevalence of Influenza D Viruses in Cattle. J Virol 2020, 94. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Current status of HPAI H5N1 virus spread in the U.S. dairy cow herds. As of May 16,2024, HPAI H5N1 virus was tested in dairy cow herds in nine states (shown in aqua color), 49 herds in total in the U.S.. The number of herds affected in each state was shown. Data was obtained from USDA website (https://www.aphis.usda.gov/livestock-poultry-disease/avian/avian-influenza/hpai-detections/livestock).
Figure 1.
Current status of HPAI H5N1 virus spread in the U.S. dairy cow herds. As of May 16,2024, HPAI H5N1 virus was tested in dairy cow herds in nine states (shown in aqua color), 49 herds in total in the U.S.. The number of herds affected in each state was shown. Data was obtained from USDA website (https://www.aphis.usda.gov/livestock-poultry-disease/avian/avian-influenza/hpai-detections/livestock).
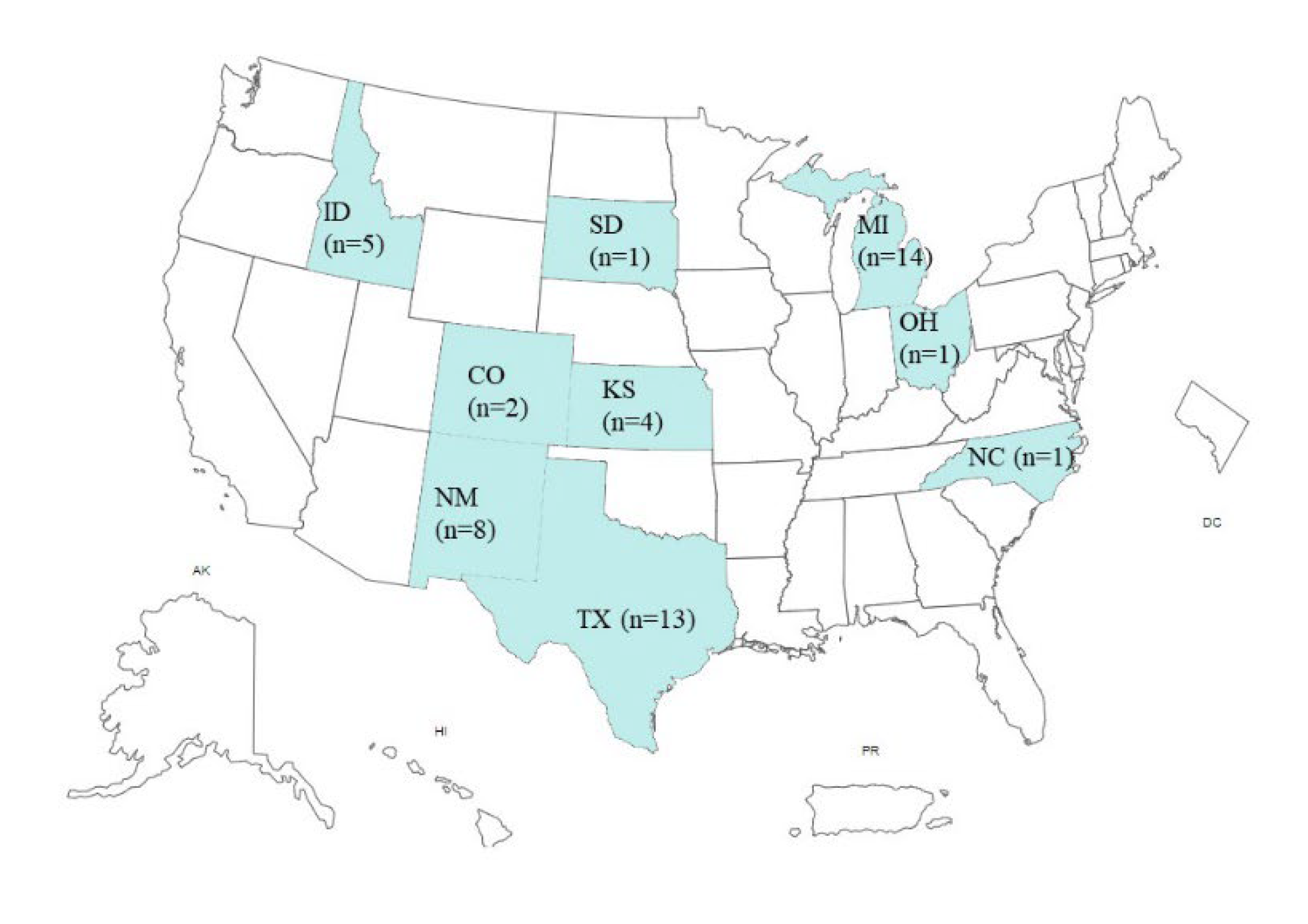
Figure 2.
Interspecies transmission and host range of HPAI H5N1. Wild aquatic birds are the reservoir hosts for type A avian influenza viruses. Wild birds, wild mammals, poultry, and domestic/pet animals were affected by HPAI H5N1. The bold dark blue arrows denote frequent spillover events, while the grey arrows denote sporadic/infrequent spillover. Sustained human-to-human transmission has not been reported. Vector images were downloaded from Freepik (www.freepik.com) and Microsoft Office 365 icons.
Figure 2.
Interspecies transmission and host range of HPAI H5N1. Wild aquatic birds are the reservoir hosts for type A avian influenza viruses. Wild birds, wild mammals, poultry, and domestic/pet animals were affected by HPAI H5N1. The bold dark blue arrows denote frequent spillover events, while the grey arrows denote sporadic/infrequent spillover. Sustained human-to-human transmission has not been reported. Vector images were downloaded from Freepik (www.freepik.com) and Microsoft Office 365 icons.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



