
Submitted:
31 May 2024
Posted:
31 May 2024
Read the latest preprint version here
Abstract
Listeria monocytogenes is an emerging pathogen responsible for serious foodborne disease, listeriosis. Commensal gut microbiota is the first line of defense against pathogen internalization. Gut microbiome can be modified by prebiotic substrates, which are among others frequently added to food products and dietary supplement. Prebiotics should selectively support the growth of beneficial microbes and thus improve host health. Nevertheless, little is known about their effect on growth of L. monocytogenes. The aim of this study was to evaluate the growth ability of four L. monocytogenes strains, representing the most common serotypes, on prebiotic oligosaccharides: beta(1,3)-D-glucan, inulin, fructooligosaccharides, galactooligosaccharides, lactulose, raffinose, stachyose, 2´-fucosyllactose and mixture of human milk oligosaccharides as a sole carbon source. The results showed that only beta(1,3)-D-glucan was metabolized by L. monocytogenes. Therefore, the health-safety of beta(1,3)-D-glucan in respect to listeriosis development should be furthermore studied.

Keywords:
Listeria monocytogenes
; prebiotics
; oligosaccharides
; beta(1
; 3)-D-glucan
; pathogen growth
1. Introduction
Listeria monocytogenes is ubiquitous gram-positive, facultative anaerobic, non-spore forming bacterium. This organism is opportunistic intracellular pathogen responsible for foodborne disease, listeriosis. Despite the fact, that the disease is relatively rare, it is characterized by severe symptoms and high mortality rate (it accounts for 20-30 % of the deaths of infected individuals worldwide). Ingestion of highly contaminated food (aprox. 109 cells) is in most cases associated with mild to severe febrile gastroenteritis. By contrast, in the case of high risks individuals such as children, elderly and immunocompromised people the disease is characterized by sepsis, meningitidis and meningoencephalitides [1,2,3]. Pregnant women are also highly endangered group because L. monocytogens traverse fetoplacental barrier and cause infection of fetus, resulting in fetal death, premature birth, or complications to pregnancy [4]. The infectious dose for these groups of people is as low as 100 to 1000 cells [1,2,3]. In general, listeriosis is the fifth most reported zoonosis in the EU. Despite strict hygiene precaution and regular food control, annually, around 2,000 people become infected in the EU. Around 50 % of cases are hospitalized and 10 % percent of those infected die, which is much more compared to other foodborne and zoonotic diseases [5]. L. monocytogenes can be subclassified in 13 known serotypes, from which 1/2a, 1/2b, 1/2c and 4b are the most common serotypes in food and are related with 95% of all listeriosis outbreaks. However, serotype 4b accounts for 50% of cases [3,6,7].
This disease can be treated by various antibiotics such as benzylpenicillin, ampicillin, erythromycin, meropenem, moxifloxacin, linezolid and trimethoprim-sulfamethoxazole (EUCAST, 2023). Nevertheless, not all individuals can be treated with antibiotics, especially pregnant women. In general, the best form of protection is prevention. One of the most important approaches of prevention is to avoid the consumption of food where these bacteria may be present, e.g., raw meat, soft ripening cheeses, uncooked fish products etc. [8]. However, due to L. monocytogenes ubiquitous dispersal, the risk of ingestion and disease outbreak cannot be fully excluded even in foods such as fresh vegetable or fruits [9,10].
Today, it is already known that the main barrier to the invasion of L. monocytogenes into the host's body is commensal bacteria, which are normally found in the digestive tract. Due to the spread of antibiotic resistant pathogenic strains, there are efforts to support the viability and metabolic activity of commensal microbiota in the digestive tract of the host. This effort can be supported by the administration of prebiotics in host nutrition, which can nourish the health-benefit bacteria. These bacteria can than occupy the adhesion sites on the intestinal mucosa, produce antimicrobial substances and thus prevent the colonization of digestive tract with pathogens and/or inhibit their growth [11,12,13].
Prebiotics are defined as substrates that are selectively utilized by host microorganisms conferring a health benefit. The most commonly used prebiotics in the food industry or in the pharmaceutical sector are mainly non-digestive nonstarch polysaccharides and oligosaccharides [14]. The most popular prebiotics are inulin, beta-glucans, fructooligosaccharides (FOS), galactooligosaccharides (GOS), xylooligosaccharides (XOS), isomaltooligosaccharides (IMO), raffinose series oligosaccharides (RSO) or lactulose. For infants, the best prebiotics are human milk oligosaccharides (HMO), which are found in breast milk [15,16]. Despite that prebiotics should only be metabolized by health-beneficial bacteria, it was found that even undesirable bacteria can grow on these substrates [17,18].
Because L. monocytogenes is able to live in different environments such as soil or vegetation as a saprophyte or in the cytosol of mammalian cells as an intracellular pathogen, it can use different substrates as sources of the main elements for its growth. Nutrient requirements are strain-specific and may vary depending on the strain origin [19]. This organism requires amino acids (leucin, isoleucine, valine, histidine, arginine, glutamine, and methionine) and organically bound sulphur for its growth, but it can also metabolize peptides or complex proteins [19,20]. In the absence of these substances in the environment, bacteria can use hexose-phosphate or glycerol as alternative energy sources. Bacteria use this mechanism especially in the role of an intracellular pathogen, when there is lack of amino acids in the cytosol [19]. The growth is improved by various saccharides especially by glucose. L. monocytogenes can ferment differently mono- and oligosaccharides, e.g., fructose, lactose, mannose, trehalose, cellobiose, L-rhamnose and maltose [21,22,23,24]. It is known that L. monocytogenes can also utilize some polysaccharides such as maltodextrin [22] and it has a functional chitinolytic system and an active lytic polysaccharide monooxygenase, the mechanisms associated with the ability to cleave chitin and cellulose [25,26].
To the best of our knowledge, there are no studies focused on the ability of L. monocytogenes to metabolize prebiotics. Therefore, the aim of this work was to investigate the growth properties of the most common serotypes of L. monocytogenes in media containing different prebiotics as the sole source of energy and to evaluate the safety of prebiotics oligosaccharides used in foodstuff and dietary supplements with respect to the possible support of listeria growth.
2. Materials and Methods
2.1. Isolation, Identification, and Characterization of Strains
Listeria strains were isolated from foodstuffs purchased in the market network of the Czech Republic and from a smear from the work surface in the food processing plants according to ISO 11290-1:2017 standard. Subsequently, presumptive L. monocytogenes colonies were taken, aseptically transferred into the Brain-heart infusion broth (Oxoid, UK) and cultivated at 37 °C for 24 hours. Freshly grown isolates were checked for purity using a phase-contrast microscope (Nikon Eclipse E200LED MV RS, Japan) and identified by MALDI-TOF mass spectrometry (Bruker Daltonik GmbH, Germany) as described in Salmonová et al. [27] to the genus level. Listeria spp. were than identify by sequencing of 16S rDNA. Briefly, genomic DNA was isolated from fresh overnight cultures using the PrepMan Ultra kit (Applied Biosystems, USA) according to the manufacturer's instructions. Amplification of 16S rRNA gene was performed using fD1 [5' AGAGTTTGATCCTGGCTCAG 3'] and rP2 [5' ACGGCTACCTTGTTACGACTT 3'] primers [28] under the conditions described in Salmonová et al., [27]. The amplified PCR products were verified by electrophoresis on a 1% agarose gel, purified with E.Z.N.A.® Cycle Pure kit (Omega Bio-tek, USA) and sequenced by Sanger method (GATC Biotech, Germany). Sequences data were analyzed using the ClustalX package in BioEdit [29,30], and compared with sequences available in the GenBank nucleotide database in NCBI (https://blast.ncbi.nlm.nih.gov/Blast.cgi) and EZ Biocloud (https://www.ezbiocloud.net). The sequences were deposited in GenBank via the BankIt program at the NCBI website (https://www.ncbi.nlm.nih.gov/WebSub/?tool=genbank).
For confirmed L. monocytogenes, serotype was determined by slide agglutination method using commercial antisera according to the manufacturer's instructions (Merck Group, Germany). Furthermore, the pathogenic potential was studied (as follows). First the hemolytic activity was detected. Cultures were spread on the Columbia blood agar (Oxoid, UK) supplemented with 5% sheep blood (v/v) and incubated for 24 h at 37 °C. Next, the presence of internalin B was detected by the PCR method using primers lmo2821-F [5' TGTAACCCCGCTTACACAGTT 3'] and lmo2821-R [5' TTACGGCTGGATTGTCTGTG 3'] [31].
Four hemolytic and internalin B positive strains of different origin and serotype were selected for substrate preference testing. Strain LM1 (serotype 4b, Access. No. OR725603) was isolated from salami, LM11 (serotype 1/2b, Access. No. OR725605) from a smear from the work surface of the food industry, LM56 (serotype 1/2a, Access. No. OR725613) from raw beef meat and LM79 (serotype 1/2c, Access. No. OR725613) from vegie minced meat.
2.2. Tested Prebiotics and Cultivation Media Composition
The growth characteristics were determined in vitro in basal medium containing 5 g/L of tryptone (Oxoid, UK) and 9 g/L NaCl, which was prepared anaerobically by Hungate technique [32]. For testing, the most commonly used prebiotics for human and livestock nutrition were selected in form of food supplements: beta(1,3)-D-glucan (purity of 100% yeast beta-glucans of which > 86.6% beta(1,3)-D-glucan, Brainway Inc., USA), inulin (purity > 90%, Frutafit, Hages, SK), fructooligosaccharides (purity of 95%, Nutri-Extract, CZ), galactooligosaccharides (purity of 95%, Nutri-Extract, CZ) and 2´-fucosyllactose (purity > 94%, RAW´s, CZ). The less common prebiotics: lactulose (purity > 95%, Sigma-Aldich, CZ), raffinose (purity > 99%, Sigma-Aldich, CZ) and stachyose (purity > 98%, Sigma-Aldich, CZ) were also tested. All supplements were used as received without further purification. Finally, ability of listeria strains to grow on mixture of human milk oligosaccharides isolated from human milk according Rockova et al. [33] was studied. Concentrated stock solutions of each substrate (20 g/L) were filter-sterilized using syringe PES filters with a 0.22 μm pore size membrane (Rotilabo®, Germany). Vials with 9 mL of basal medium were supplemented with solution of each substrate to obtain a final concentration of 2 g/L. Due to the partial solubility of beta(1,3)-D-glucan in water and its heat resistance [34,35], this prebiotic was added directly to the basal medium and sterilized by autoclaving at 121 °C for 15 min. An amount of residual or released glucose after autoclaving was verified spectrophotometrically using Glucose Test (Supelco, Canada) and measured with Reflectoquant® RQflex 10 (Sigma-Aldich, Germany). The resulting amount of glucose in medium with beta(1,3)-D-glucan after autoclaving was as low as 2 mg/L.
As a positive control, glucose at the concentration of 2 g/L was used as a sole carbohydrate source. Due to the incomplete purity and residual content of undefined monosaccharides in tested substrates, control with 0,1 g/L of glucose was also included (residual control). The medium without any carbohydrate source was tested as a negative control (basal medium). The control media were sterilized by autoclaving.
2.3 Testing of Bacterial Growth
Fresh overnight cultures grown under aerobic condition at 37 °C in Brain-heart infusion broth (Oxoid, UK), were checked for purity and washed twice with sterile saline. Media were inoculated by bacteria suspension to final density of 3 x 106 CFU/mL and cultivated anaerobically at 37 °C for 24 h. All experiments were prepared in triplicates. Bacterial growth was determined spectrophotometrically at 620 nm using spectrometer UV/VIS SPEKOL 1300 (Analytik Jena GmbH, Germany), and calculated as a difference in optical density (OD) at the time of inoculation and after 24 hours of cultivation. Since the medium with beta(1,3)-D-glucan showed high density (OD=0,4), bacterial counts were evaluated by the agar plate count technique [36] using BHI agar, after 24 h cultivation under aerobic conditions at 37 °C. The growth of L. monocytogenes strains for beta(1,3)-D-glucan was analyzed by subtracting initial values of log of colony-forming units (CFU) per 1 mL from the final values.
The pH values of cultivation media were measured using HI98100 pH tester Checker® Plus (Hanna instruments, USA) at the time of inoculation and after 24 hours of cultivation.
Growth curves were constructed for strains showing growth (significant changes) on tested carbohydrates. Optical density (CFU/mL for beta(1,3)-D-glucan) as described above were measured in one-hour intervals during bacterial growth.
The specific growth rates were calculated for individual samples according to Rada et al. (2008) [17] using formula:
where x and x0 were optical densities or CFU/mL measured within the exponential growth phase at time t and t0, respectively.
μ = (ln x – ln x0)/(t – t0)
2.4 Statistical Analyses
All data were evaluated by Multiple range comparison with Multiple range test in program Statgraphics Centurion XV Version 15.02.06 (Statgraphics technologies, Inc., USA).
3. Results
3.1. Utilization of Water-Soluble Prebiotics
The results of listeria strains growth on prebiotics soluble in water are summarized in Table 1. The growth of all tested strains in positive control (2 g/L glucose) was significantly (P < 0.05) higher than on the tested carbohydrates. Slightly increased bacterial densities, which were significantly (P < 0.05) higher than in the negative control (no saccharides), were observed on inulin, fructooligosaccharides, galactooligosaccharides and in the residual control (0,1 g/L glucose). Other prebiotics were not utilised, and the optical density values of all strains were similar to the negative control. The strain specific utilization was found for inulin, galactooligosaccharides, stachyose, 2´-fucosyllactose and mixture of human milk oligosaccharides. Listeria growth in the negative and residual control was also strain specific.
3.2. Utilization of beta(1,3)-D-glucan
L. monocytogenes growth on beta(1,3)-D-glucan are summarized in Table 2. Bacterial cell counts in media containing beta(1,3)-D-glucan were in line with counts determined in positive control. The growth in negative control was significantly (P < 0.05) lower than on bought above-mentioned media.
Bacterial growth was accompanied by a decrease in the pH of the culture medium. The initial pH value was 6.9 in all batches. In positive control and beta(1,3)-D-glucan batches, the pH decreased to 5.7. In the inulin experiments, pH decreased to 6.3, and in the residual control, fructooligosaccharides and galactooligosaccharides to pH 6.6. Values of pH did not change in media with saccharides, where bacterial growth was not detected. This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
3.3. Growth CURVES and rates
Growth curves were constructed and specific growth rates were calculated for positive control, residual control, beta(1,3)-D-glucan, inulin, fructooligosaccharides and galactooligosaccharides. Growth curves during the cultivation of listeria on water-soluble prebiotics are shown in Figure 1. The specific growth rates are summarized in Table 3. Although the same specific growth rates of L. monocytogenes on prebiotics and controls was found, the maximal optical density value was significantly higher in positive control than in prebiotic case. The growth curves show that for bacteria cultured on medium with residual control, inulin, fructooligosaccharides and galactooligosaccharides, the exponential phase last around 3 hours and growth was terminated at OD values between 0.028 and 0.047. Longer exponential phase (8 hours) was observed during the cultivation in a positive control, when the growth terminated at OD 0.128. As in the case of bacterial growth ability, the strain-specific growth rates were found for residual control, inulin, galactooligosaccharides and at this case also for fructooligosaccharides.
The growth curves of bacteria cultivated in media with positive control and beta(1,3)-D-glucan are presented in Figure 2, and the calculated specific growth rates are shown in Table 4. No statistically significant difference (P < 0.05) was found for the specific growth rates obtain for these two media, although the growth curve shapes were different with a maximal count of 3.50 x 108 CFU/mL after 9 hours of cultivation in positive control and 2.8 x 108 CFU/mL after 8 hours of cultivation in media with beta(1,3)-D-glucan.
4. Discussion
The consumption of prebiotics saccharides should selectively stimulate the growth of health-promoting (probiotic) bacteria, suppress occurrence of pathogenic bacteria and conferring benefit(s) upon host health [14,18]. However, it has been noted that some prebiotics can be utilized by undesirable bacteria as well [17,18]. Listeria monocytogenes as a foodborne pathogen, can also occur in the digestive tract, where it can use various nutrients for its growth, including several specific saccharides [19,20]. The ability to utilise different saccharides by bacteria depends on the bacteria's genetic predisposition [19,37,38]. To the best of our knowledge, there are no studies focused on the ability of L. monocytogenes to metabolize prebiotics. Therefore, the aim of this study was to evaluate the growth of the most common listeria serotypes on several prebiotics’ supplements with a purity of 90-100 %, depending on the product.
The results showed that lactulose, raffinose, stachyose, 2´-fucosyllactose and mixture of human milk oligosaccharides were unable to promote L. monocytogenes growth and even residual substrates presented in tested supplements did not affect the results. A slight increase in cell numbers was observed for supplements containing inulin, fructooligosaccharides and galactooligosaccharides. Inulin and fructooligosaccharides are prebiotics composed of linear chains of fructose units, linked by β-(2,1) glycosidic bonds, terminated by a non-reducing sucrose end. The difference between these two prebiotics is in the length of chain, whereas fructooligosaccharides comprise of 3 – 7 fructose units, inulin is mainly built form 10 – 60 units [12,39]. L. monocytogenes is capable to utilize fructose [24], but it seems that it is not able to cleave the β-(1,2) glycosidic bond that connects individual fructose units. Galactooligosaccharides are composed of 2 – 8 galactose units linked together with β-(1,6) glycosidic bonds and one molecule of terminal glucose connected by β-(1,3) or β-(1,4) glycosidic bond [40]. This prebiotic saccharide does not promote growth of L. monocytogenes, because galactose is not among substrates that provide a source of carbon for listeria growth [23]. The bacterial growth and specific growth rate on these prebiotic supplements were comparable to the growth in residue control. Therefore, we assume that the growth of L. monocytogenes strains was probably caused by the presence of residual monosaccharides in the prebiotic supplements, which constituted 5 % of fructooligosaccharides and galactooligosaccharides supplements and ≤ 10 % of inulin supplement. Higher growth on inulin than on fructooligosaccharides and galactooligosaccharides could be affected by higher content of monosaccharides. Strain-specific growth of listeria on certain substrates are known [19,38], which was also shown in our study. Although a statistically significant strain specificity growth was observed, no comparable growth of L. monocytogenes strains as in the positive control was found.
Beta(1,3)-D-glucan was the only prebiotic supplement, on which the comparable growth of L. monocytogenes strains as on the positive control was found. Beta(1,3)-D-glucan is a prebiotic polysaccharide belonging to the group of beta-glucans composed only from monomers of glucose that are linked by β-(1,3) glycosidic bonds [41,42]. They are generally considered effective and widely applied prebiotics, which are used for their ability to protect the body against various pathogens, cancer, high LDL cholesterol level and their antioxidant effects, which overall supports the host's health [43]. Beta-glucans are major structural components of plant and fungi and include the major biopolymer cellulose. The structure of cellulose is similar to beta(1,3)-D-glucan, but glucose units are linked by β-(1,4) glycosidic bonds [44]. It has been demonstrated that L. monocytogenes is able to utilize cellulose [25,26]. Therefore, it could be assumed that L. monocytogenes is capable to cleave beta-glucans as well. Beta-glucans used in this study were of S. cerevisiae origin, and in addition to β-(1,3)-glucans also contain less than 15% of β-(1,6)-glucans [43]. Since the results confirmed this presumption, it could be expected, that L. monocytogenes can clave both β-(1,3) and β-(1,6) bonds, and metabolize released glucose, which is the main source of carbon in listeria metabolism. Regarding the growth curve, there were observed some differences between the course of L. monocytogenes growth on beta(1,3)-D-glucan and the positive control. Slightly lower cell counts on beta(1,3)-D-glucan could be easily explained by the fact that part of the energy, that could have been used for reproduction, was spent on the substrate cleavage. It could be also result of gradual or incomplete cleavage. The decrease in the number of viable cells on beta(1,3)-D-glucan was slower compared to the positive control, probably due to the gradual release of glucose from the beta-glucan chain.
There are various natural sources of beta-glucans, e.g., yeast, barley, bacteria, seaweed, fungi or dahlia tuber. They differ not only in origin, but also in glycosidic bond position as well as in their other characteristic such as solubility in water or alkalis. Except the β-(1,3)- also β-(1,4)- or β-(1,6)- glucans are found in supplements [45,46,47]. The differences between the growths of Bifidobacterium sp. depending on the source of beta-glucans were demonstrated [47]. Thus, we can hypothesize that the L. monocytogenes ability to metabolize these prebiotics also depends on the source.
Regardless of whether the L. monocytogenes is able to utilize these prebiotics or not, effects against pathogens, such as supporting the individual's immunity, growth of beneficial microbes and reduced adherence ability of pathogens in general, have been reported for these substances [48,49,50,51,52]. However, the results are sometimes ambiguous or contradictory. For example, Ebersbach et al. [53] found that inulin, which L. monocytogenes is not able to use as source of carbon, decreased the resistance of the host body against the L. monocytogenes. On the contrary, despite that L. monocytogenes strains are able to utilize beta(1,3)-D-glucan, it has been found that these prebiotics stimulate immune system in mice or guinea pig and effectively inhibit pathogens proliferation, reduce adhesion to the digestive tract wall and suppress L. monocytogenes infection [54,55,56]. In addition beta(1,3)-D-glucan are metabolized by commensal and probiotic intestinal bacteria which leads to competition for substrate. Moreover, short-chain fatty acids (SCFA), which are formed through microbial metabolite cross-feeding, suppress listeria growth and support the body's disease defense. The health-promoting bacteria also produce various substances that inhibit L. monocytogenes, such as bacteriocins [57,58,59,60,61,62,63,64]. The fact that L. monocytogenes is able to utilize beta(1,3)-D-glucan is therefore not alarming. Compared to the growth of previously studied Lactobacillus, Bifidobacterium and commensal Clostridium strains in the presence of selected prebiotics, the growth of L. monocytogenes was lower [17,47,65]. This is most likely affected by the medium composition. The media used for probiotics/commensals testing are composed for fastidious bacteria and contained complex nutritional mixture except the energy source. Some of these nutrients can L. monocytogenes utilize for its growth even without the presence of saccharides, therefore nutrient-poor medium is required for this organism [17,47,65].
Finally, the question how else prebiotics can affect metabolism and behavior of this pathogen because different saccharides have been found to affect the pathogenic potential [66,67,68,69], biofilm formation potential [69] and resistance to antimicrobial substances [21], heat or acids [69], arises. To our best knowledge, there are no studies focusing on these topics for prebiotics used in our study.
Authors should discuss the results and how they can be interpreted from the perspective of previous studies and of the working hypotheses. The findings and their implications should be discussed in the broadest context possible. Future research directions may also be highlighted.
5. Conclusions
Our study attempted to evaluates the growth ability of listeria on different prebiotics. The results showed that inulin, fructooligosaccharides, galactooligosaccharides, lactulose, raffinose, stachyose, 2´-fucosyllactose and mixture of human milk oligosaccharides are not used by L. monocytogenes as a nutrient and source of energy. Therefore, these prebiotics can be considered as health-safe with respect to the possible support of listeria growth. Beta(1,3)-D-glucan was the only prebiotic utilized by L. monocytogenes, thus this substrate does not meet the general definition of prebiotics. Even though the results showed that beta(1,3)-D-glucan support the growth of L. monocytogenes, available studies highlight suppressive effects on listeriosis, probably due to the function complexity. Therefore, the health-safety of beta(1,3)-D-glucan with respect to listeriosis development should be furthermore studied.
Author Contributions
Conceptualization, Kodešová, T. and Šubrtová Salmonová, H..; methodology, Kodešová, T.; Šubrtová Salmonová, H. and Musilová Š.; validation, Kodešová, T.; formal analysis, Kodešová, T.; investigation, Kodešová, T.; Mašlejová, A., Horváthová, K.; resources, Vlková, E.; writing— original draft, Kodešová, T.; writing— review & editing, Šubrtová Salmonová, H., Vlková, E.; visualization, Kodešová, T.; supervision, Šubrtová Salmonová. H.; funding acquisition, Vlková, E. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the NutRisk Centre, Ministry of Education, Youth and Sports of the Czech Republic [CZ.02.1.01/0.0/0.0/16_019/0000845, 2019-2023]; and METROFOOD-CZ research infrastructure project [MEYS Grant No: LM2023064] including access to its facilities.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Farber, J.M.; Peterkin, P.I. Listeria monocytogenes, a food-borne pathogen. Microbiol Rev 1991, 55, 476–511. [Google Scholar] [CrossRef]
- Shamloo, E.; Hosseini, H.; Moghadam, A.Z.; Larsen, H.M.; Haslberger, A.; Alebouyeh, M. Importance of Listeria monocytogenes in food safety: a review of its prevalence, detection, and antibiotic resistance. Iran J Vet Res 2019, 20, 241–254. [Google Scholar]
- Bhunia, A.K. Foodborne microbial pathogens, mechanisms and pathogenesis, 2nd ed.; Springer: Switzerland, 2018; p. 365. ISBN 978-1-4939-7347-7. [Google Scholar]
- Azari, S.; Johnson, L.J.; Webb, A.; Kozlowski, S.M.; Zhang, X.; Rood, K.; Amer, A.; Seveau, S. Hofbauer cells spread Listeria monocytogenes among placental cells and undergo pro-inflammatory reprogramming while retaining production of tolerogenic factors. mBio 2021, 12. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union one health 2020 zoonoses report. EFSA Journal 2021, 19, 6971. [Google Scholar] [CrossRef]
- Das, S.; Surendran, P.K.; Thampuran, N. Detection and differentiation of Listeria monocytogenes and Listeria innocua by multiplex PCR. Fishery Technology 2010, 47, 91–94. [Google Scholar]
- Nadon, C.A.; Woodward, D.L.; Young, C.; Rodgers, F.G.; Wiedmann, M. Correlations between molecular subtyping and serotyping of Listeria monocytogenes. J Clin Microbiol 2001, 39, 2704–2707. [Google Scholar] [CrossRef]
- Bortolussi, R. Public Health - Listeriosis: a primer. Can Med Assoc J 2008, 179, 795–797. [Google Scholar] [CrossRef]
- Miceli, A.; Settanni, L. Influence of agronomic practices and pre-harvest conditions on the attachment and development of Listeria monocytogenes in vegetables. Annals of Microbiology 2019, 69, 185–199. [Google Scholar] [CrossRef]
- Simonetti, T.; Peter, K.; Chen, Y.; Jin, Q.; Zhang, G.; LaBorde, L.F.; Macarisin, D. Prevalence and distribution of Listeria monocytogenes in three commercial tree fruit packinghouses. Front Microbiol 2021, 12, 652708. [Google Scholar] [CrossRef]
- Khan, R.; Petersen, F.C.; Shekhar, S. Commensal bacteria: an emerging player in defense against respiratory pathogens. Front Immunol 2019, 10, 1203. [Google Scholar] [CrossRef]
- Gibson, G.R.; Probert, H.M.; Van Loo, J.; Rastall, R.A.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: updating the concept of prebiotics. Nutr Res Rev 2004, 17, 259–275. [Google Scholar] [CrossRef]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: metabolic and health benefits. Br. J. Nutr. 2010, 104 Suppl. 2, 1–63. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: the International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat Rev Gastroenterol Hepatol 2017, 14, 491–502. [Google Scholar] [CrossRef]
- Charalampopoulos, D.; Rastall, R.A. Prebiotics in foods. Curr Opin Biotechnol 2012, 23, 187–191. [Google Scholar] [CrossRef]
- Musilova, S.; Rada, V.; Vlkova, E.; Bunesova, V. Beneficial effects of human milk oligosaccharides on gut microbiota. Benef Microbes 2014, 5, 273–283. [Google Scholar] [CrossRef]
- Rada, V.; Nevoral, J.; Trojanová, I.; Tománková, E.; Šmehilová, M.; Killer, J. Growth of infant faecal bifidobacteria and clostridia on prebiotic oligosaccharides in in vitro conditions. Anaerobe 2008, 14, 205–208. [Google Scholar] [CrossRef]
- Bunešová, V.; Vlková, E.; Rada, V.; Kňazovická, V.; Ročková, Š.; Geigerová, M.; Božik, M. Growth of infant fecal bacteria on commercial prebiotics. Folia Microbiol 2012, 57, 273–275. [Google Scholar] [CrossRef]
- Sauer, J.-D.; Herskovits, A.A.; O’Riordan, M.X.D. Metabolism of the Gram-positive bacterial pathogen Listeria monocytogenes. Microbiol Spectr 2019, 7, 1–12. [Google Scholar] [CrossRef]
- Friedman, M.E.; Roessler, W.G. Growth of Listeria monocytogenes in defined media. J Bacteriol 1961, 82, 528–533. [Google Scholar] [CrossRef]
- Balay, D.R.; Gänzle, M.G.; McMullen, L.M. The effect of carbohydrates and bacteriocins on the growth kinetics and resistance of Listeria monocytogenes. Front Microbiol 2018, 9, 347. [Google Scholar] [CrossRef]
- Gopal, S.; Berg, D.; Hagen, N.; Schriefer, E.M.; Stoll, R.; Goebel, W.; Kreft, J. Maltose and maltodextrin utilization by Listeria monocytogenes depend on an inducible ABC transporter which is repressed by glucose. PLoS One 2010, 5, 10349. [Google Scholar] [CrossRef]
- Pine, L.; Malcolm, G.B.; Brooks, J.B.; Daneshvar, M.I. Physiological studies on the growth and utilization of sugars by Listeria species. Can J Microbiol 1989, 35, 245–254. [Google Scholar] [CrossRef]
- Schardt, J.; Jones, G.; Müller-Herbst, S.; Schauer, K.; D’Orazio, S.E.F.; Fuchs, T.M. Comparison between Listeria sensu stricto and Listeria sensu lato strains identifies novel determinants involved in infection. Sci Rep 2017, 7, 17821. [Google Scholar] [CrossRef]
- Paspaliari, D.K.; Loose, J.S.M.; Larsen, M.H.; Vaaje-Kolstad, G. Listeria monocytogenes has a functional chitinolytic system and an active lytic polysaccharide monooxygenase. FEBS J 2015, 282, 921–936. [Google Scholar] [CrossRef]
- Bae, D.; Seo, K.S.; Zhang, T.; Wang, C. Characterization of a potential Listeria monocytogenes virulence factor associated with attachment to fresh produce. Appl Environ Microbiol 2013, 79, 6855–6861. [Google Scholar] [CrossRef]
- Salmonová, H.; Salmonová, S.; Killer, J.; Bunešová, V.; Bunešová, B.; Geigerová, M.; Geigerová, G.; Vlková, E.; Vlková, V. Cultivable bacteria from Pectinatella magnifica and the surrounding water in South Bohemia indicate potential new Gammaproteobacterial, Betaproteobacterial and Firmicutes taxa. FEMS Microbiol Lett 2018, 365. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J Bacteriol 1991, 173, 697–703. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X Windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 1997, 25, 4876–4882. [Google Scholar] [CrossRef]
- Hall, T. BioEdit: an important software for molecular biology. GERF Bull Biosci 2011, 2, 60–61. [Google Scholar]
- Liu, D.; Ainsworth, A.J.; Austin, F.W.; Lawrence, M.L. Characterization of virulent and avirulent Listeria monocytogenes strains by PCR amplification of putative transcriptional regulator and internalin genes. J Med Microbiol 2003, 52, 1065–1070. [Google Scholar] [CrossRef]
- Hungate, R.E. A roll tube method for cultivation of strict anaerobes. In Methods in Microbiology. 1st ed.; Norris, J.R.; Ribbons, D.W., Eds.; Academic Press, San Diego, US, 1969; Volume 3, pp. 117–132. [CrossRef]
- Rockova, S.; Nevoral, J.; Rada, V.; Marsik, P.; Sklenar, J.; Hinkova, A.; Vlkova, E.; Marounek, M. factors affecting the growth of bifidobacteria in human milk. Int Dairy J 2011, 21, 504–508. [Google Scholar] [CrossRef]
- Bai, Y.P.; Zhou, H.M.; Zhu, K.R.; Li, Q. Effect of thermal processing on the molecular, structural, and antioxidant characteristics of highland barley β-glucan. Carbohydr Polym 2021, 271, 118416. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhou, H.M.; Huang, Z.H.; Zhao, R.Y. Different aggregation states of barley β-glucan molecules affects their solution behavior: a comparative analysis. Food Hydrocoll 2020, 101, 105543. [Google Scholar] [CrossRef]
- Jones, G.S.; D’Orazio, S.E.F. Listeria monocytogenes: cultivation and laboratory. Curr Protoc Microbiol 2013, 31, 9B.2.1–9B.2.7. [Google Scholar] [CrossRef]
- Durica-Mitic, S.; Göpel, Y.; Görke, B. Carbohydrate utilization in bacteria: making the most out of sugars with the help of small regulatory RNAs. Microbiol Spectr 2018, 6. [Google Scholar] [CrossRef]
- Gahan, C.G.M.; Hill, C. Listeria monocytogenes: survival and adaptation in the gastrointestinal tract. Front Cell Infect Microbiol 2014, 5, 1–7. [Google Scholar] [CrossRef]
- Lockyer, S.; Stanner, S. Prebiotics – an added benefit of some fibre types. Nutr Bull 2019, 44, 74–91. [Google Scholar] [CrossRef]
- Ibrahim, O.O. Functional oligosaccharides: chemicals structure, manufacturing, health benefits, applications and regulations. J Food Chem Nanotechnol 2018, 4, 65–76. [Google Scholar] [CrossRef]
- Mudgil, D. The interaction between insoluble and soluble fiber. In Dietary fiber for the prevention of cardiovascular disease: fiber’s interaction between gut micoflora, sugar metabolism, weight control and cardiovascular health., 1st ed.; Samaan, R.A., Ed.; Academic Press: San Diego, US, 2017; pp. 35–59. [Google Scholar] [CrossRef]
- Stone, B.A. Chemistry of β-glucans. In Chemistry, biochemistry, and biology of 1-3 beta glucans and related polysaccharides, 1st ed.; Bacic, A., Fincher, G.B., Stone, B.A., Eds.; Academic Press: San Diego, CA, USA, 2009; pp. 5–46. [Google Scholar] [CrossRef]
- Kim, K.S.; Yun, H.S. Production of soluble β-glucan from the cell wall of Saccharomyces cerevisiae. Enzyme Microb Technol 2006, 39, 496–500. [Google Scholar] [CrossRef]
- Synytsya, A.; Novak, M. Structural analysis of glucans. Ann Transl Med 2014, 2, 17. [Google Scholar] [CrossRef]
- Jayachandran, M.; Chen, J.; Chung, S.S.M.; Xu, B. A critical review on the impacts of β-glucans on gut microbiota and human health. J Nutr Biochem 2018, 61, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Shokri, H.; Asadi, F.; Khosravi, A.R.; Shokriy, H.; Khosraviy, A.R. Isolation of β -glucan from the cell wall of Saccharomyces cerevisiae. Nat Pro Res 2009, 22, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Cheung, P.C.K. Fermentation of β-glucans derived from different sources by bifidobacteria: evaluation of their bifidogenic effect. J Agric Food Chem 2011, 59, 5986–5992. [Google Scholar] [CrossRef]
- Buddington, K.K.; Donahoo, J.B.; Buddington, R.K. Dietary oligofructose and inulin protect mice from enteric and systemic pathogens and tumor inducers. J Nutr 2002, 132, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Zheng, J.; Lin, A.; Xia, H.; Zhang, Z.; Gao, Q.; Lv, W.; Liu, H. A review: roles of carbohydrates in human diseases through regulation of imbalanced intestinal microbiota. J Funct Foods 2020, 74, 104197. [Google Scholar] [CrossRef]
- Chen, P.; Reiter, T.; Huang, B.; Kong, N.; Weimer, B.C. Prebiotic oligosaccharides potentiate host protective responses against L. monocytogenes infection. Pathogens 2017, 6, 68. [Google Scholar] [CrossRef] [PubMed]
- Karakan, T.; Tuohy, K.M.; Janssen-van Solingen, G. Low-dose lactulose as a prebiotic for improved gut health and enhanced mineral absorption. Front Nutr 2021, 8, 672925. [Google Scholar] [CrossRef] [PubMed]
- Sangwan, V.; Tomar, S.K.; Ali, B.; Singh, R.R.B.; Singh, A.K. Galactooligosaccharides reduce infection caused by Listeria monocytogenes and modulate IgG and IgA levels in mice. Int Dairy J 2015, 41, 58–63. [Google Scholar] [CrossRef]
- Ebersbach, T.; Jørgensen, J.B.; Heegaard, P.M.; Lahtinen, S.J.; Ouwehand, A.C.; Poulsen, M.; Frøkiær, H.; Licht, T.R. Certain dietary carbohydrates promote Listeria infection in a guinea pig model, while others prevent it. Int J Food Microbiol 2010, 140, 218–224. [Google Scholar] [CrossRef]
- Kupfahl, C.; Geginat, G.; Hof, H. Lentinan has a stimulatory effect on innate and adaptive immunity against murine Listeria monocytogenes infection. Int Immunopharmacol 2006, 6, 686–696. [Google Scholar] [CrossRef]
- Li, W.; Yajima, T.; Saito, K.; Nishimura, H.; Fushimi, T.; Ohshima, Y.; Tsukamoto, Y.; Yoshikai, Y. Immunostimulating properties of intragastrically administered Acetobacter-derived soluble branched (1,4)-β-d-glucans decrease murine susceptibility to Listeria monocytogenes. Infect Immun 2004, 72, 7005–7011. [Google Scholar] [CrossRef] [PubMed]
- Torello, C.O.; De Souza Queiroz, J.; Oliveira, S.C.; Queiroz, M.L.S. Immunohematopoietic modulation by oral β-1,3-glucan in mice infected with Listeria monocytogenes. Int Immunopharmacol 2010, 10, 1573–1579. [Google Scholar] [CrossRef] [PubMed]
- Shao, X.; Fang, K.; Medina, D.; Wan, J.; Lee, J.L.; Hong, S.H. The probiotic Leuconostoc mesenteroides inhibits Listeria monocytogenes biofilm formation. J Food Saf 2020, 40, e12750. [Google Scholar] [CrossRef]
- Corr, S.C.; Gahan, C.G.M.; Hill, C. Impact of selected Lactobacillus and Bifidobacterium species on Listeria monocytogenes infection and the mucosal immune response. FEMS Immunol Med Microbiol 2007, 50, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Amézquita, A.; Brashears, M.M. Competitive inhibition of Listeria monocytogenes in ready-to-eat meat products by lactic acid bacteria. J Food Prot 2002, 65, 316–325. [Google Scholar] [CrossRef] [PubMed]
- García, M.J.; Ruíz, F.; Asurmendi, P.; Pascual, L.; Barberis, L. Searching potential candidates for development of protective cultures: evaluation of two Lactobacillus strains to reduce Listeria monocytogenes in artificially contaminated milk. J Food Saf 2020, 40, e12723. [Google Scholar] [CrossRef]
- Hascoët, A.S.; Ripolles-avila, C.; Cervantes-huamán, B.R.H.; Rodríguez-jerez, J.J. In vitro preformed biofilms of Bacillus safensis inhibit the adhesion and subsequent development of Listeria monocytogenes on stainless-steel surfaces. Biomolecules 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Tran, T.D.; Del Cid, C.; Hnasko, R.; Gorski, L.; McGarvey, J.A. Bacillus amyloliquefaciens ALB65 inhibits the growth of Listeria monocytogenes on Cantaloupe melons. Appl Environ Microbiol 2020, 87, 1–10. [Google Scholar] [CrossRef]
- Alonso, V.P.P.; Harada, A.M.M.; Kabuki, D.Y. competitive and/or cooperative interactions of Listeria monocytogenes with Bacillus cereus in dual-species biofilm formation. Front Microbiol 2020, 11, 177. [Google Scholar] [CrossRef]
- da Silva Sabo, S.; Converti, A.; Todorov, S.D.; Domínguez, J.M.; de Souza Oliveira, R.P. effect of inulin on growth and bacteriocin production by Lactobacillus plantarum in stationary and shaken cultures. Int J Food Sci Technol 2015, 50, 864–870. [Google Scholar] [CrossRef]
- Kunová, G.; Rada, V.; Lisová, I.; Ročková, Š.; Vlková, E. In vitro fermentability of prebiotic oligosaccharides by lactobacilli. Czech J. Food Sci 2011, 29, 49–54. [Google Scholar] [CrossRef]
- Jaradat, Z.W.; Bhunia, A.K. Glucose and nutrient concentrations affect the expression of a 104-kilodalton listeria adhesion protein in Listeria monocytogenes. Appl Environ Microbiol 2002, 68, 4876–4883. [Google Scholar] [CrossRef] [PubMed]
- Aké, F.M.D.; Joyet, P.; Deutscher, J.; Milohanic, E. mutational analysis of glucose transport regulation and glucose-mediated virulence gene repression in Listeria monocytogenes. Mol Microbiol 2011, 81, 274–293. [Google Scholar] [CrossRef] [PubMed]
- Park, S.F.; Kroll, R.G. Expression of listeriolysin and phosphatidylinositol-specific phospholipase c is repressed by the plant-derived molecule cellobiose in Listeria monocytogenes. Mol Microbiol 1993, 8, 653–661. [Google Scholar] [CrossRef]
- Crespo Tapia, N.; Dorey, A.L.; Gahan, C.G.M.; den Besten, H.M.W.; O’Byrne, C.P.; Abee, T. different carbon sources result in differential activation of sigma B and stress resistance in Listeria monocytogenes. Int J Food Microbiol 2020, 320, 108504. [Google Scholar] [CrossRef]
Figure 1.
The growth curves of Listeria monocytogenes strains cultivated on prebiotics (2 g/L) soluble in water. All data for individual strains are average from triplicates. Positive control = 2 g/L glucose, Residual control = 0,1 g/L glucose. OD = optical density. A: Data show average of 4 strains – LM1 (serotype 4b), LM11 (serotype 1/2b), LM 56 (serotype 1/2a), LM 79 (serotype 1/2c); B: The growth of individual strains in positive control (2 g/L glucose); C: The growth of individual strains in residual control (0,1 g/L glucose). D: The growth of individual strains on inulin (2 g/L); E: The growth of individual strains on fructooligosaccharides (2 g/L); F: The growth of individual strains on galactooligosaccharides (2 g/L).
Figure 1.
The growth curves of Listeria monocytogenes strains cultivated on prebiotics (2 g/L) soluble in water. All data for individual strains are average from triplicates. Positive control = 2 g/L glucose, Residual control = 0,1 g/L glucose. OD = optical density. A: Data show average of 4 strains – LM1 (serotype 4b), LM11 (serotype 1/2b), LM 56 (serotype 1/2a), LM 79 (serotype 1/2c); B: The growth of individual strains in positive control (2 g/L glucose); C: The growth of individual strains in residual control (0,1 g/L glucose). D: The growth of individual strains on inulin (2 g/L); E: The growth of individual strains on fructooligosaccharides (2 g/L); F: The growth of individual strains on galactooligosaccharides (2 g/L).

Figure 2.
The growth curves of Listeria monocytogenes strains on beta(1,3)-D-glucan and positive control. All data for individual strains are average from triplicates. CFU – colony forming units. A: Data show mean of 4 strains – LM1 (serotype 4b), LM11 (serotype 1/2b), LM 56 (serotype 1/2a), LM 79 (serotype 1/2c); B: The growth of individual strains in positive control (2 g/L glucose); C: The growth of individual strains on beta(1,3)-D-glucan (2 g/L).
Figure 2.
The growth curves of Listeria monocytogenes strains on beta(1,3)-D-glucan and positive control. All data for individual strains are average from triplicates. CFU – colony forming units. A: Data show mean of 4 strains – LM1 (serotype 4b), LM11 (serotype 1/2b), LM 56 (serotype 1/2a), LM 79 (serotype 1/2c); B: The growth of individual strains in positive control (2 g/L glucose); C: The growth of individual strains on beta(1,3)-D-glucan (2 g/L).
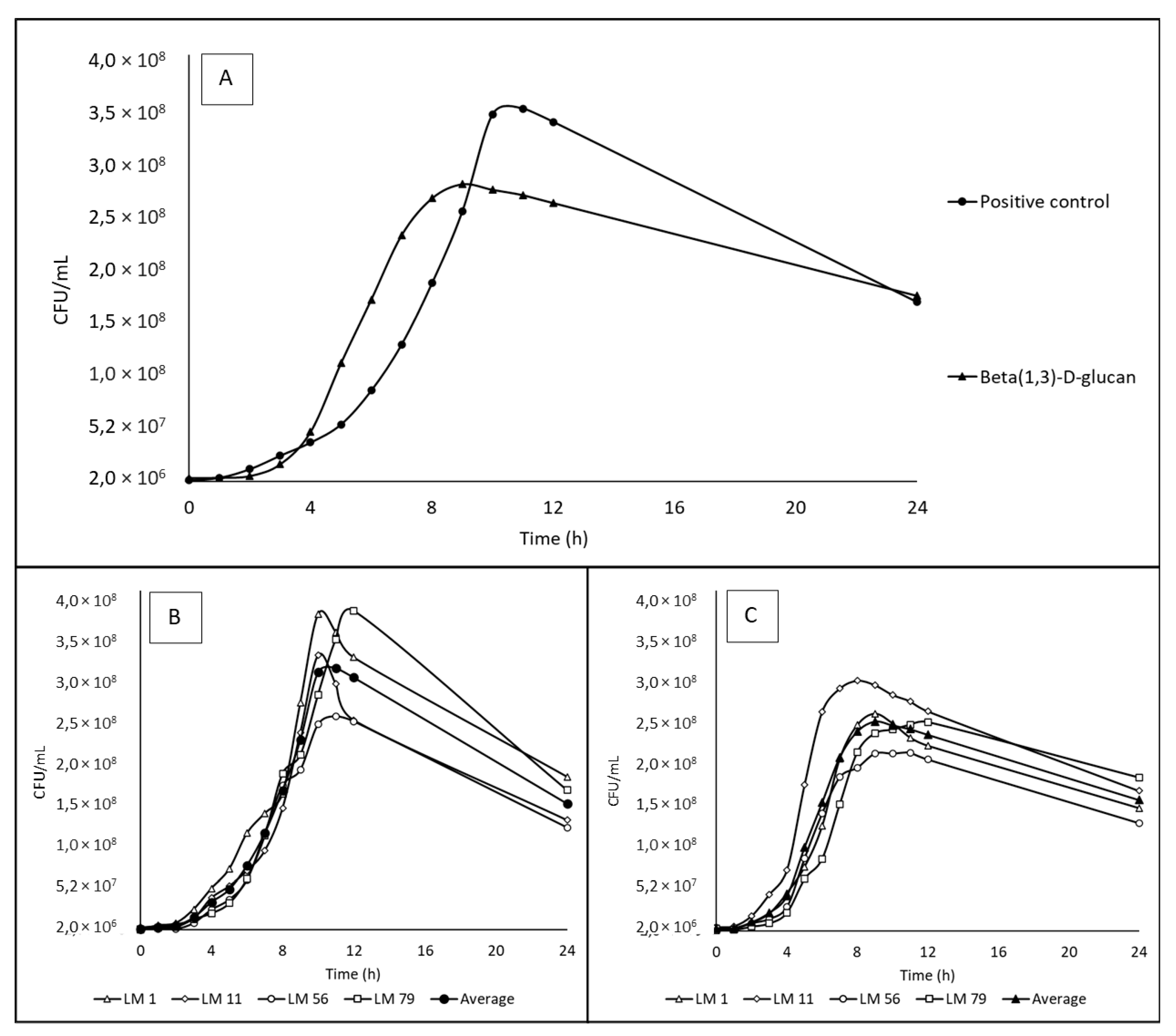

Table 1.
Listeria monocytogenes growth on selected prebiotic saccharides soluble in water.
| Strain code | LM1 | LM11 | LM56 | LM79 | Average ± SD | |
|---|---|---|---|---|---|---|
| Serotype | 4b | 1/2b | 1/2a | 1/2c | - | |
| Positive control (2 g/L glucose) | 0.156 ± 0.036Aa | 0.140 ± 0.027Aa | 0.127 ± 0.032Aa | 0.130 ± 0.030Aa | 0.138 ± 0.032A | |
| Residual control (0,1 g/L glucose) | 0.028 ± 0.002Ba | 0.029 ± 0.001BCa | 0.028 ± 0.001BCDab | 0.026 ± 0.001BCb | 0.028 ± 0.002BC | |
| Negative control (no saccharides) | 0.016 ± 0.015Ba | -0.006 ± 0.026Db | 0.008 ± 0.008Deab | 0.005 ± 0.005Dab | 0.006 ± 0.017EF | |
| Inulin | 0.027 ± 0.006Bb | 0.028 ± 0.004BCb | 0.047 ± 0.013Ba | 0.043 ± 0.005Ba | 0.036 ± 0.012B | |
| Fructooligosaccharides | 0.026 ± 0.031Ba | 0.029 ± 0.008Bca | 0.034 ± 0.011Bca | 0.016 ± 0.005Cda | 0.027 ± 0.016BCD | |
| Galactooligosaccharides | 0.008 ± 0.018Bb | 0.046 ± 0.006Ba | 0.018 ± 0.002CDEb | 0.011 ± 0.007CDb | 0.021 ± 0.018BCD | |
| Lactulose | 0.014 ± 0.040Ba | 0.010 ± 0.014Cda | -0.001 ± 0.005Ea | -0.005 ± 0.001Da | -0.001 ± 0.010F | |
| Raffinose | 0.004 ± 0.006Ba | 0.006 ± 0,002Cda | 0.004 ± 0.006DEa | -0.003 ± 0.007Da | 0.003 ± 0.006EF | |
| Stachyose | 0.032 ± 0.005Ba | 0.011 ± 0.009CDbc | 0.015 ± 0.005CDEb | 0.003 ± 0.005CDc | 0.015 ± 0.012CDE | |
| 2´-fucosyllactose | 0.019 ± 0.002Ba | 0.013 ± 0.003Cdab | 0.015 ± 0.004CDEab | 0.011 ± 0.006CDb | 0.014 ± 0.005CDE | |
| Mixture of HMOs | 0.028 ± 0.014Ba | 0.010 ± 0.007Cdab | 0.016 ± 0.007CDEa | -0.006 ± 0.010Db | 0.012 ± 0.015DEF | |
Data are expressed as differences in OD at the time of inoculation and after 24 hours of cultivation measured at 620 nm. All data for individual strains are average from triplicates ± SD. OD – optical density. SD – standard deviation. HMOs – Human milk oligosaccharides. ABCDEFdata with different superscripts differ (P < 0.05) in column. abcdata with different superscripts differ (P < 0.05) in line.
Table 2.
Listeria monocytogenes growth on beta(1,3)-D-glucan.
| Strain code | LM1 | LM11 | LM56 | LM79 | Average ± SD |
|---|---|---|---|---|---|
| Serotype | 4b | 1/2b | 1/2a | 1/2c | - |
| Positive control (2 g/L glucose) | 1.56 ± 0.72Aa | 1.2 ± 0.04Aba | 0.95 ± 0.01Aa | 1.24 ± 0.15Aa | 1.24 ± 0.39A |
| Negative control (no saccharides) | 0.33 ± 0.06Ba | 0.49 ± 0.12Ba | 0.16 ± 0.02Ba | 0.24 ± 0.11Ba | 0.31 ± 0.15B |
| Beta(1-3)-D-glucan | 1.05 ± 0.09Aba | 1.43 ± 0.67Aa | 0.96 ± 0.07Aa | 1.48 ± 0.72Aa | 1.23 ± 0.48A |
Data are expressed as differences in log CFU/mL at the time of inoculation and after 24 hours of cultivation. All data for individual strains are average from triplicates ± SD. CFU – colony forming units. SD – standard deviation. ABdata with different superscripts differ (P < 0.05) in column. abcdata with different superscripts differ (P < 0.05) in line.
Table 3.
Listeria monocytogenes specific growth rates on prebiotic saccharides soluble in water.
| Strain code | LM1 | LM11 | LM56 | LM79 | Average ± SD |
|---|---|---|---|---|---|
| Serotype | 4b | 1/2b | 1/2a | 1/2c | - |
| Positive control (2 g/L glucose) | 0.42 ± 0.21Aa | 0.63 ± 0.04Aa | 0.49 ± 0.02Ba | 0.64 ± 0.10Aa | 0.55 ± 0.14A |
| Residual control (0,1 g/L glucose) | 0.48 ± 0.07Aab | 0.32 ± 0.03Bb | 0.40 ± 0.08Bab | 0.55 ± 0.20ABCa | 0.44 ± 0.14A |
| Inulin | 0.35 ± 0.02ABc | 0.63 ± 0.12Aab | 0.68 ± 0.03Aa | 0.51 ± 0.04ABb | 0.54 ± 0.11A |
| Fructooligosaccharides | 0.38 ± 0.10ABb | 0.67 ± 0.12Aa | 0.70 ± 0.09Aa | 0.47 ± 0.09BCb | 0.55 ± 0.20A |
| Galactooligosaccharides | 0.19 ± 0.05Bb | 0.66 ± 0.03Aa | 0.73 ± 0.18Aa | 0.28 ± 0.13Cab | 0.46 ± 0.19A |
All data for individual strains average from triplicates ± SD. SD - standard deviation. ABCDdata with different superscripts differ (P < 0.05) in column. abcdata with different superscripts differ (P < 0.05) in line.
Table 4.
Listeria monocytogenes specific growth rates on beta(1,3)-D-glucan.
| Strain code | LM1 | LM11 | LM56 | LM79 | Average ± SD |
|---|---|---|---|---|---|
| Serotype | 4b | 1/2b | 1/2a | 1/2c | - |
| Positive control (2 g/L glucose) | 0.53 ± 0.01 | 0.53 ± 0.01 | 0.52 ± 0.01 | 0.47 ± 0.16 | 0.51 ± 0.07 |
| Beta(1,3)-D-glucan | 0.53 ± 0.01 | 0.54 ± 0.01 | 0.53 ± 0.01 | 0.52 ± 0.01 | 0.53 ± 0.01 |
All data are average from triplicates ± SD. SD - standard deviation. No different superscripts differ (P < 0.05) was found.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



