
Submitted:
30 May 2024
Posted:
03 June 2024
You are already at the latest version
Abstract
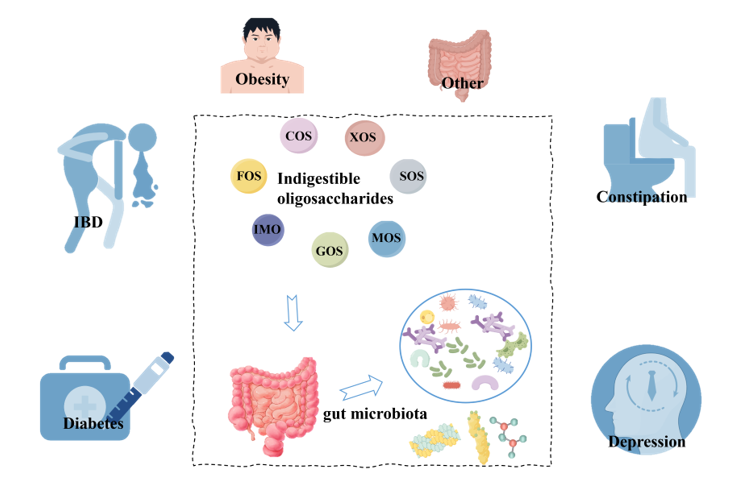
The gut has been a focus of chronic disease research. The gut microbiota produces metabolites that act as signaling molecules and substrates, closely influencing host health. Indigestible oli-gosaccharides (NDOs), as a common dietary fiber, play an important role in regulating the structure and function of gut microbiota. Their mechanism of action is mainly attributed to their carbon source as specific probiotics, producing related metabolites, and regulating the gut mi-crobial community. However, due to the selective utilization of oligosaccharides, some factors such as the types and structures of oligosaccharides have different impacts on the composition of microbial populations and the production of metabolites in the colon ecosystem. This review systematically describes the key factors influencing the selective utilization of oligosaccharides in microorganisms and elaborates how oligosaccharides affect the host's immune system, inflam-mation levels and energy metabolism by regulating microbial diversity and metabolic function, which in turn will affect the onset and progress of chronic diseases, especially diabetes, obesity, depression and intestinal inflammatory diseases, constipation. In this review, we re-examine the interaction mechanisms between gut microbiota and its associated metabolites and diseases, and explore new strategies for promoting human health and combating chronic diseases through di-etary interventions.
Keywords:
nondigestible oligosaccharides
; gut microbiota
; short-chain fatty acids
; constipation
; diabetes
; obesity
; depression
; inflammatory bowel disease
1. Introduction
Chronic diseases refer to diseases that have a slow onset, slow progression, long course, and usually cannot be completely cured. In the past few years, the role of gut microbiota in related diseases has been extensively studied, such as ulcerative colitis [1], irritable bowel syndrome [2], diabetes [3], Alzheimer's disease [4], non-alcoholic fatty liver disease [5], obesity [6]. In this case, the role of probiotics becomes particularly important. By regulating the balance of gut microbiota, it is possible to maintain the stability of the gut environment, enhance intestinal barrier function, reduce gut inflammation levels, and help regulate energy metabolism and insulin sensitivity [7,8,9]. For example, in obese individuals, the diversity of the gut microbiota is decreased, which is usually accompanied by changes in key species [6,10,11,12,13], like Firmicutes and Bacteroidetes [14,15]. Diabetes mellitus type 2 (T2DM) is characterized by an impaired gut microbiota [16,17], and Akkermansia and Enterobacter have been found to have a causal role in the onset and progression of T2DM [18]. Nonalcoholic fatty liver disease (NAFLD) is regarded as a typical metabolic disorder at the intersection of obesity, metabolic syndrome and T2DM [19], often with a proliferation of Enterobacteriaceae and Escherichia coli [20]. Dysbiosis of the gut microbiota has also been associated with inflammatory bowel disease (IBD), including encompassing three main phenotypes-Crohn disease (CD) and ulcerative colitis (UC) and IBD unclassified (IBDU) [21]. There are common microbial patterns among IBD patients, manifested at the phylum level as a loss of Firmicutes [22,23,24], an increase in Proteobacteria and potential pathogenic bacteria [25,26,27,28], At the genus level, Clostridium and Escherichia coli were significantly increased in IBD, whereas Phascolarctobacterium, Faecalibacterium, and Roseburia intestinalis were significantly decreased [27,29,30,31,32]. The gut-brain axis plays an important role in neurological diseases including Alzheimer's disease (AD), anxiety, and depression. The gut microbiota is linked to central nervous system in a variety of ways, including regulation of immune responses, formation of metabolites (e.g., short-chain fatty acid), and synthesis of neurotransmitters (e.g., serotonin and dopamine). Now some studies have shown that some microorganisms can regulate intestinal homeostasis and hinder the progression of neurological diseases [33,34,35,36,37,38,39,40,41]. In fact, diet is the main determinant that drives the growth of gut microbiota and regulates its interaction with the host. The lack of sufficient functional microbiota and accessible carbohydrates (MAC) is the main reason for the disappearance of important species that regulate intestinal homeostasis [42,43]. Indigestible oligosaccharides (NODs) are typical MAC that are easily broken down by Bacteroides and Lactobacillus containing metabolic enzymes. The use of oligosaccharide fermentation to improve intestinal health is a widespread and current focus [44,45]. The beneficial effects and promising prospects of NODs on the host by targeting the gut microbiota and microbial metabolites are now generally accepted [46,47,48,49,50], including improvement of bowel movements, suppression of appetite, enhancement of postprandial glycemic response and promotion of mineral absorption. This article describes the structure and sources of NODs, as well as the factors affecting the microbial fermentation of NODs, and then elucidates the important role of oligosaccharides in chronic diseases by regulating the gut microbiota.
2. Classification and Sources of Oligosaccharides
Oligosaccharides are a class of carbohydrates that consist of 2 to 10 monosaccharide molecules, typically consisting of two or more different monosaccharides that are linked together in various ways through glycosidic bonds [51], classified as digestible or indigestible [52]. In some dietary oligosaccharides, the anomeric C atom (C1 or C2) of the mono-saccharide units has a configuration that makes their glycosidic bounds nondigestible to the human digestive enzymes [52]. In addition to α-1,4 glycosidic bonds, the chemical bonds between monosaccharides in NDOs also contain α-1,6 glycosidic bonds or β-1,2 glycosidic bonds that cannot be degraded by human digestive enzymes. It belongs to prebiotics along with fiber polysaccharides, which are selectively fermented and cause specific changes in the composition and/or activity of gastrointestinal microbiota, thereby benefiting the well-being and health of the host [53,54].The source and structural characteristics of main functional oligosaccharides are listed in Table 1.
3. The impact of Different NODs on the Diversity of Gut Microbiota
Non digestible oligosaccharides (NODs) are mainly fermented by Bifidobacteria, Lactic acid bacteria (LAB), and other symbiotic bacteria, which typically contain various enzymes related to carbohydrate metabolism in the large intestine [64,65]. For example, LAB can degrade FOS and GOS by encoding β-FFase and β-galactosidase [66]. Different NODs often have many differences, such as source, degree of polymerization, glycosidic bonds and overall structural complexity (e.g., side chains) [67,68,69]. These differences result in a diverse microbiota for fermenting NDOs, manifested in varying degrees of beneficial bacterial proliferation, inhibition of harmful bacteria, and differences in acid and gas production [70].
3.1. Degree of Polymerization (DP)
In fact, NODs with different DP can be utilized by different strains. For example, DP3 oligofructose (FOS) promoted the growth of Bifidobacterium species in vitro while inhibiting the proliferation of Clostridia, and it showed superior selective stimulatory activity compared to other FOS [71]. In previous work, Gopal et al. [72] demonstrated that Bifidobacterium lactis DR10 consumes oligosaccharides (GOS) with higher DP, while Lactobacillus rhamnosus DR20 prefers to use galactose disaccharides. In general, oligosaccharides with higher DP are more resistant to digestion when passing through the upper digestive tract and are therefore more likely to reach the colon to be utilized by the gut microbiota. However, high DP may also inhibit selective fermentation by microorganisms. Zhao et al. [73] evaluated the prebiotic ability of Bamboo Shoot Shells Xylo-Oligosaccharide (XOS) with different DP, and showed that XOS with a high DP (>X5) led to an increase in the abundance of beneficial bacteria such as Phascolarctobacterium and Bacteroides, while low DP XOS with more X2-X3 components promoted the proliferation of other beneficial bacteria, including Lachnochlostridium. Immerzeel et al. [74] and Falck et al. [75] demonstrated that Bifidobacterium adolescentis and Lactobacillus acidophilus predominantly utilize the X2-X3 fractions. In fact, low DP NODs has a stronger ability to support the growth and proliferation of gut microbiota [76]. When high DP XOS exists, the microbial community consumes more energy to secrete xylanase, preferentially degrading high DP XOS to low DP XOS, thereby limiting the metabolic capacity of gut microbiota [77]. Another study on oligomeric dextran showed that the component with DP 3-4 (≥ 80%) was utilized by all probiotic strains, the portion with DP 5-6 was utilized by some strains, and the portion with DP ≥ 7 was only utilized by LAB and Bifidobacteria [78]. Similarly, isomaltose from DP 3-5 is more indigestible than DP 2, and although it has a higher chance of reaching the colon, its fermentation selectivity for beneficial probiotic species is lower [79,80,81].
3.2. Glycosidic Bond
Different structures of NODs have different effects on gut microbiome, including glycosidic bond types and connection modes. For example, α-(1→3) and α-(1→2)-linked glucose oligosaccharides (GlcOS) formed during isomaltooligosaccharides (IMOs) production promote the growth of Bifidobacterium spp. and Lactobacillus spp. [82], and their fermentation selectivity (against probiotic species in gut bacteria) is higher than that of only α-(1→6)-linked IMOs [83]. Zeng et al. [84] studied the effect of NODs with different disaccharide bonds on selected Lactobacillus, and most of the species had low growth rates on IMOs with α-(1→6), except L. fermentum FUA 3589, where β-(1→4)-linked fibrous disaccharides and β-(1→6)-linked gentian disaccharides were able to significantly promote the proliferation of L. brevis ATCC 8287, L. rhamnosus ATCC 53103, L. plantarum WCFS1, and L. gasseri ATCC 33323. Similarly, Sanz et al. [70] investigated the prebiotic index (PI) of disaccharide, which was higher for α-glucose disaccharides than for β-types, except for α, α-alginose. Djouzi Z et al. [85] compared the effects of β-oligofructose, β-oligogalactose and α-oligosaccharides (non-disaccharides) on the metabolism of gut microbiota, and compared to α-oligosaccharides, oligofructose and oligogalactose containing β-glycosidic bonds were the preferred substrates for the growth of Bifidobacteria, which have a great impact on microbial composition, which has been found in several studies [86,87]. More importantly, it has been reported that intake of GOS containing β-1, 3, β-1, 4 and β-1, 6 glycosidic bond showed better prebiotic effects in healthy volunteers [88]. In addition, Kittibunchakul et al. [89] concluded that the fermentation activity of Bifidobacterium and Lactobacillus as well as E. faecium on GOS (mainly containing β-1, 3 and β-1, 6 glycosidic bonds) showed better fermentative activity than those GOS which contain a single β-1, 4 glycosidic bond. This is similar to the conclusion of Cardelle Cobas [90], who found that Bifidobacteria, LAB and Streptococcus preferred galactose containing β-1, 6 glycosidic bond, but not β-1, 4 glycosidic bond. In conclusion, the growth ability of probiotics is related to the type of glycosidic bond. Therefore, it is important to understand the structure-activity relationship of NODs.
3.3. Linear and Branching Structures
The type, number and branching complexity of substituents in NODs will affect the microbial fermentation rate and composition. It has been reported that the presence of substituents on XOS can affect Bifidobacterium fermentation, resulting in different amounts of lactic acid [91], In addition, Kabel observed that the lactic acid content in unsubstituted XOS was higher than that in substituted XOS [92], that is, the XOS with linear structure was fermented by more gut microorganisms. When fermented in vitro with human feces, the fermentation speed of linear XOS and arabinose substituted XOS (AXOS) was faster than acetylated XOS (AXOS), and XOS containing 4-O-methylglucuronic acid group (GlcAmeXOS) was the slowest [93], which indicated that the presence of substituents delayed or completely hindered the fermentation. On the other hand, Bacteroidetes showed superior growth performance when XOS was used as the sole carbon source compared to AXOS [94]. Bacteroidetes was able to utilize AXOS and released ferulic acid [95,96,97]. The process of XOS degradation by Bacteroidetes involves extracellular degradation and debranching of the skeleton, followed by intracellular hydrolysis of the oligomers, which ultimately produces acetate, propionate, and butyrate as the major metabolic end products [98]. It has been proved that microbial degrading enzymes are not easy to contact NODs with high content of branches, resulting in the reduction of microbial fermentation rate [99]. In addition, the chain length and shape of NODs are equally important. For example, long-chain IMO has stronger resistance to the hydrolysis and degradation of the upper intestine, and is more effective than short-chain IMO in stimulating the growth of Bifidobacterium, Prevotella and Lactobacillus. Bifidobacteria have selective specificity in utilizing FOS with different chain lengths. Generally, short-chain FOS are preferentially fermented, followed by long-chain FOS [100]. Cyclic IMO have been shown to inhibit Streptococcus [80,101,102,103].
From the above viewpoints, the degree of polymerization, substituents, glycosidic bond types have a significant impact on the fermentation activity of probiotics. However, the reasons for the preference of probiotics for oligosaccharides are lack of in-depth investigation, and the metabolic mechanism of microorganisms for oligosaccharides with different structures needs further study.
4. The Impact of Different NODs on the Diversity of Gut Microbiota
4.1. T2DM
Diabetes is a chronic disease characterized by hyperglycemia, caused by absolute or relative insufficiency of insulin secretion and impaired utilization. At present, increasing dietary fiber intake is one of the recommendations for patients with T2DM [104]. In recent years, many studies have shown that, including in vivo and in vitro experiments, NODs can effectively improve diabetes by regulating gut microbiota to enrich or decrease specific flora. For example, in T2DM rats, the abundance of Firmicutes increased at the phylum level, while the abundance of Bacteroidetes, Actinobacteria, and Verrucomicrobia decreased [105]. At the genus level, Bifidobacterium, Roseburia, Faecalibacterium, Bacteroides, and Akkermansia decreased, while Desulfovibrio, Oscillibacter, Fusobacterium, Ruminococcus, and Blautia increased [106,107]. Weninger et al. [107] showed that FOS reduced the species α-diversity after 16 weeks of treatment in T2DM rat, that the relative abundance of Bifidobacterium increased and the relative abundance of Ruminococcus decreased significantly. This result was also found in the later study of FOS and GOS [108], with the difference that Phascolarctobacterium, Coprococcus and Oscillospira were also decreased.
In addition, NODs can regulate insulin sensitivity, blood glucose and lipid metabolism by influencing microorganisms to produce beneficial metabolites, such as short-chain fatty acids (SCFAs), amino acid metabolites, and bioactive polypeptides, thereby alleviating the onset and progression of diabetes. 1) SCFAs are currently the most studied metabolites. Gobinath et al. [109] demonstrated that the decrease of plasma cholesterol in diabetic rats fed XOS or FOS was due to the inhibition of cholesterol synthesis by increased propionate or altered bile acid metabolism. In addition, the reduction of butyrate producing bacteria is associated with impaired glucose control [17,110]. SCFAs act as signaling molecules to maintain host energy homeostasis by activating G protein-coupled receptors GPR41 and GPR43 to regulate insulin levels and inhibit hepatic gluconeogenesis [111]. It also promotes the secretion of glucagon-like peptide-1 (GLP-1) and peptide YY (PYY). GLP-1 participates in glucose homeostasis primarily by reducing blood glucose levels and improving insulin secretion and resistance. PYY, as a hormone mediating satiety, is secreted from endocrine L cells in the distal intestine after the body intakes enough food. SCFAs play a crucial role in the development of T2DM and can be used as markers of metabolic disorders or homeostasis [112]. 2) Amino acids and their metabolites: The intake of NODs can produce a variety of amino acid metabolites, which have an impact on blood glucose levels and insulin sensitivity. It has been reported that fecal metabolic profiling showed the content of glycogen amino acids such as proline, serine, and leucine increased after NODs intervention in T2DM rats [113]. This result was also confirmed by Chen et al. [114], who found that an increase in serine and glutamine can reduce the risk of T2DM progression. In addition, the expression of glucose transporter (GLUT4) was decreased in STZ induced diabetic rats, whereas Rice husk oligosaccharides (RH-XOS)-treated T2DM rats significantly regulated total GLUT4 protein expression [115].
In conclusion, NODs can influence glucose metabolism (especially glucose metabolism), lipid metabolism, protein and carbohydrate metabolism by regulating the gut microbiota and fecal metabolites [113,114]. Specifically, they can improve insulin sensitivity by regulating AMPK pathway and INS-R/IRS/Glut4 insulin signaling pathway to achieve the goal of treating T2DM [116,117]. On the other hand, hundreds of metabolites of gut microbiota can be fed back to the brain, which plays a key role in influencing many gastrointestinal processes through the autonomic nervous system and the hypothalamus-pituitary-adrenal axis [118] (Figure 1).
4.2. Obesity
Several studies have revealed the role of gut microbiota in host physiology, behavior and metabolism, providing new insights into the pathogenic mechanism of obesity.
Firstly, gut microbial dysbiosis is a common feature in obese patients and obese animal models. Studies have shown that prebiotic oligosaccharides, such as FOS, XOS, GOS, and COS, have an anti-obesity effect by regulating the gut microbiota[49]. Generally, obese individuals have a higher proportion of Firmicutes and Bacteroidetes. A study on obese mouse model showed that the administration of NODs increased the abundance of Bacteroidetes, decreased the abundance of Firmicutes, and changed more than100 bacterial taxa [119]. Long et al. [120] demonstrated that XOS diet reduced visceral fat in mice, which was associated with changes in gut microbiota. In addition, the increase in Proteobacteria containing gram negative lipopolysaccharide (LPS) is also a microbial signature of intestinal dysbacteriosis in mice fed a high-fat diet (HFD) [121]. HFD leads to a sustained increase in plasma LPS levels (i.e., metabolic endotoxemia), as well as a systemic inflammatory response, which play a promoting role in the development of insulin resistance in obese patients. In a study of obese women, Bifidobacterium and Faecalibacterium prausnitzii were increased after intervention with FOS, it was proved that the changes of gut microbiota were related to the changes of fat mass, serum LPS level and metabolism (hippuric acid, lactate, and plasma protein C) [122]. Similarly, Thiennimitr et al. [121] demonstrated that consumption of XOS reduced metabolic endotoxemia in obese mice. The results indicate that NODs can inhibit the growth of pro-inflammatory bacteria and promote the growth of beneficial bacteria, thereby helping to alleviate intestinal inflammation and reduce the risk of obesity. In summary, manipulation of the composition of the gut microbiota through NODs has been considered as a possible approach for the prevention and treatment of obesity.
The direct cause of obesity is that the rate of fat synthesis exceeds the rate of fat consumption. Therefore, blocking fat synthesis and increasing energy consumption is the most effective strategy to combat obesity. The impact of NODs on weight loss is mainly attributed to the production of SCFAs by the gut microbiota [123], as they play an important role in regulating food intake and energy metabolism [124]. Research has shown that GOS improves lipid metabolism in mice by regulating the synthesis of SCFAs [125]. Dietary supplementation of MOS can inhibit appetite and systemic insulin resistance in obese mice by remodeling gut microbial composition and enhancing the formation of SCFAs [126]. Chen et al. [127] found that XOS changed the gut microbiota and affected the levels of SCFAs. After removing the gut microbiota, SCFAs levels were significantly decreased, indicating that XOS indirectly affected the generation of SCFAs by regulating the metabolic activities of the microbiota. In addition, acetate is catalyzed to acetyl-CoA and butyrate is converted to butyryl-COA, both of which act as inhibitors of fatty acid synthesis by acetyl CoA carboxylase (ACC) to prevent obesity [127]. Butyrate is enriched by the intake of oligosaccharides, and regulating the ratio of butyrate to acetate in the gut microbiota is a potential strategy for the treatment of obesity. For example, specific enrichment of butyrate producing bacteria may be beneficial for obesity treatment.
NODs can regulate the synthesis and release of bile acids by affecting gut microbiota associated with bile acid (BA) metabolism, such as Clostridia, Bacteroidetes, Bacteroides, Lactobacillus, and Bifidobacterium [49]. The G-protein-coupled bile acid receptor (TGR5) is one of the key receptors for BA. Under the action of Clostridia, bile acids can be converted to deoxycholic acid (DCA) and lithocholic acid (LCA), which activate TGR5 and enhance the cyclic adenosine monophosphate/protein kinase A (cAMP-PKA) signaling pathway. Activation of TGR5 also promotes conversion of glucagon to GLP-1. Lun et al. [128] revealed the dynamic balance relationship between gut microbiota and bile acids.
The study found that after COS intervention, gut microbiota can promote the release of bile acid, and then activate TGR5, enhance brown fat thermogenesis and TGR5 dominated fatty acid oxidation signaling pathway, thus playing a role in weight loss and lipid reduction. Farnesoid X receptor (FXR) is another key receptor for BA, and several studies have shown that activation of FXR promote adipose tissue browning and reduce obesity in mice [129]. Mechanistically, activation of FXR induces the release of fibroblast growth factor 15 (FGF15) or FGF19, which reach hepatocytes through the portal vein, thereby inhibiting BA synthesis in the liver [130]. In addition, FGF19 promotes hepatic glycogen synthesis and inhibits hepatic gluconeogenesis. Other studies have shown that GOS significantly reduced the concentration of bile acids in the small intestine, improved glucose metabolism, and influenced lipid absorption by regulating lipid digestion [131,132]. Pectin oligosaccharides have also been shown to mediate cholesterol metabolism through the gut microbiota and their metabolites [133].
The gut microbiota can utilize NODs to generate nutritional metabolites that act as "messengers" for the bacterial community, influencing host energy homeostasis and regulating host metabolism, thus contributing to the fight against obesity (Figure 1).
4.3. Depression
NODs may influence the development of depression by regulating gut microbiota. For example, FOS inhibits depression-associated bacteria such as Anaerostipes, Lachnospiraceae incertae sedis, Oscillibacter, Proteobacteria, and Streptococcus, while effectively promoting the growth of beneficial bacteria with antidepressant properties [134,135]. The combination of FOS and GOS has been shown to reduce depressive symptoms [136]. At the microbial level, there is evidence that the combination of the two can prevent the change of the ratio of Actinomycetes and Proteobacteria, which is a typical feature of patients with major depression [137]. Chi et al. [134] conducted a comparative analysis of FOS and the standard antidepressant fluoxetine, noting that they had similar effects and that both FOS and fluoxetine promoted the growth of Dialister spp. A recent cohort study reported that Dialister was not found in depressed patients [138]. In other words, FOS and fluoxetine may improve depressive symptoms by regulating the presence of Dialister. In addition, Eubacterium [139], Lactobacillus [139,140] and Oxalobacter [141] are also considered beneficial. Overall, NODs showed antidepressant activity, and part of this effect was achieved by regulating of functional gut microbiota, although the exact mechanism in depressed patients requires further investigation.
The microbiota-gut-brain axis (MGBA) has been considered as a potential pathway for the treatment of neurodegenerative diseases [142] involving the autonomic nervous system (ANS), the enteric nervous system (ENS), and the hypothalamic-pituitary-adrenal axis (HPA). Gut microbes can affect the brain through a variety of pathways, including neural, immune, endocrine pathways and microbial metabolites, and even stimulate the vagal nerve endings of the gut through neurotransmitters produced by microbes themselves [143]. Neurotransmitters known to be affected by gut microbiota include SCFAs [144], γ-aminobutyric acid (GABA) [145], tryptophan (Trp) [146], and serotonin (5-HT) [147]. For example, valeric acid can alleviate intestinal injury, protect neurons, and regulate neurotransmitters [148]. And the increase of butyrate may directly affect the central nervous system [149,150,151]. This view was validated in a pig model, where oral butyrate was found to affect brain metabolism and hippocampal neurogenesis [152]. Therefore, NODs supplementation may inhibit the development of depression by enhancing the formation of SCFAs, including improving cognitive functioning and alleviating anxiety behaviors. Another important pathway is tryptophan → 5-hydroxytryptophan → serotonin. Gut microbiota plays an indispensable role in intestinal tryptophan metabolism. Tryptophan is the only precursor for the synthesis of the neurotransmitter 5-HT, which influences the development of depression [153]. Kelly et al. [154] found that transplantation of feces from depressed patients into recipient animals resulting in these recipient animals showing behavioral and physiological characteristics of depression, and the metabolic pathway of tryptophan was changed. Deng et al. [155] demonstrated that feruloylated oligosaccharides (FO) exhibited antidepressant and anxiolytic effects by regulating 5-HTP, suggesting that the gut microbiota and microbial metabolism were key mediators supporting FO to protect the nervous system. Similarly, Zhang et al. [156] revealed that Morinda officinalis oligosaccharide (MOO) regulates the 5-HT synthesis pathway (Trp → 5-HTP → 5-HT) in the gut microbiota, so that the elevated 5-HTP from the gut microbiota is absorbed into the blood, and then crosses the blood-brain barrier to improve the 5-HT level in the brain. These findings suggest that microbiota can regulate neurotransmitter levels to affect the development of depression.
Gut microbial dysbiosis may lead to physiological and behavioral disorders by inducing neuroinflammation in the central nervous system. NODs alters neural function by manipulating the gut microbiota. Currently, the exact mechanisms by which NODs play a role in depression are unknown, and it is difficult to definitively show how many or which key pathways are responsible for the pathophysiology of depression. However, it is still a very important research direction to regulate the gut microbiota and its metabolites to affect host metabolic pathways, thereby alleviating depressive symptoms (Figure 2).
4.4. Inflammatory Bowel Disease (IBD)
Inflammatory bowel disease (IBD) is a common non-specific chronic gastrointestinal inflammatory disease, including crohn's disease (CD) and ulcerative colitis (UC). The mechanisms by which oligosaccharides alleviate IBD by regulating gut microbiota mainly include: 1) enriching the diversity of the gut microbiota; 2) protecting the intestinal barrier; and 3) regulating T-cell differentiation and related signaling pathways (Figure 3). Recent studies have verified that breast milk oligosaccharides (HMOs) alleviated sodium disulfide (DSS)-induced ulcerative colitis by improving intestinal barrier function and regulating gut microbiota [48]. The gut microbiota of colitis mice treated with unsaturated alginate oligosaccharides (UAOS) was altered, with increased abundance of Firmicutes and Actinomycetes and decreased abundance of Bacteroidetes and the results showed that the protective effect of UAOS treatment was attributed to the maintenance of mucosal barrier function and suppression of immune injury by regulating gut microbiota [157].
Increasing evidence shows that functional oligosaccharides as natural active substances can effectively improve IBD [100,158]. 1) NODs can alleviate intestinal inflammation in animals by regulating the gut microbiota. Gut microbial dysbiosis will lead to ecological imbalance of intestinal mucosa, increase the production of pro-inflammatory cytokines and reduce the production of anti-inflammatory cytokines. It was found that the levels of Escherichia coli and Enterococcus faecalis in the feces of patients with IBD were significantly higher than those of normal people, while the abundance of Lactobacillus and Bifidobacterium were significantly lower [159]. In addition, FOS alleviated intestinal inflammation by promoting the growth of LAB [157]. SCFAs can be absorbed by intestinal epithelial cells (IEC) and inhibit the production of pro-inflammatory cytokines IL-8 and IL-6, thereby alleviating the inflammatory response of colonic epithelial cells. In conclusion, NDOS can alleviate the symptoms of IBD by reducing pro-inflammatory bacteria, increasing anti-inflammatory bacteria, and promoting the production of SCFAs [160].
2) Protecting the intestinal barrier. In animal models of IBD, alterations in tight junction proteins (e.g., Claudin1, Occludin, ZO-1) and adhesion junction proteins (e.g., E-cadherin, β-catenin) are usually accompanied by increased intestinal permeability and disruption of intestinal structure. In fact, it helps to restore the integrity of the mucosal barrier by regulating gut microbiota diversity [161]. For example, HMOs restored the expression of tight junction proteins and MUC-2, inhibited the production of LPS and LBP, and restored the integrity of the intestinal barrier in colitis mice [48]. SCFAs may provide energy for IEC, promote epithelial cell proliferation and differentiation, and enhance intestinal barrier function, which may be related to their regulation of the AP-1 signaling pathway [162]. In addition, konjac oligosaccharides increased the concentration of SCFAs in the mice colon, which could promote IL-18 and repair the integrity of IEC [163]. Essential aromatic amino acids (e.g., Trp) can be metabolized into indoles and their derivatives, and these metabolites can activate the aryl hydrocarbon receptor (AhR), which is closely associated with the pathogenesis of IBD [164]. Consistent with the findings of Xia et al. that three microbial metabolites of Trp alleviate IBD by improving tight junctions [165].
3) Regulation of immune homeostasis. Toll-like receptors (TLRs) in intestinal epithelial cells and immune cells recognize and respond to different microbial structural motifs. When pathogens invade, macrophages recognize pathogen-associated molecular patterns (PAMPs) through TLRs and secrete various proinflammatory cytokines (such as TNF-α, IL-1 β, IL-6, IL-12, etc.). TLR-4 plays a crucial role as a receptor for LPS in the pathogenesis of UC. The number of Tregs cells decreases in patients with IBD. SCFAs can induce the differentiation of T cells into Treg cells, promote the synthesis of the anti-inflammatory cytokine to regulate the immune response, and restore the balance of Th1/Th2 and Th17/Treg cells. A study showed that XOS were metabolized into SCFAs, and propionate and butyrate can promote the differentiation of CD4+ T cells into Treg cells, thereby slowing down inflammatory bowel disease [166]. Similarly, feruloylated oligosaccharides (FOs) promoted the percentage of Treg cells and the production of corresponding specific cytokines, thereby regulating the immune homeostasis of Th17/Treg cell in IBD mice [167]. In addition, gut microbiota can also stimulate B cells (Breg) to produce inhibitory cytokines (e.g., IL-10 and TGF-β) that affect other immune cells and immunoglobulins (e.g., sIgA). Nowadays, numerous studies have confirmed the effectiveness of oligosaccharides in improving IBD by modulating the gut microbiota, both in animal models and in human experiments [168].
4.5. Constipation
Intestinal motility is regulated by the interaction of the intestinal immune system, intestinal secretions, intestinal microbiota and their fermentation products, and impaired intestinal motility can lead to constipation [169]. The increase in the abundance of beneficial bacteria such as Bifidobacterium and Alistipes, as well as the decrease in colonic transport-associated bacteria (Oscillospira, Odoribacter), contributes to the relief of constipation [170]. NODs can significantly improve defecation frequency and shorten colonic transit time, with few side effects, such as FOS, IMO, and GOS [171]. In addition to the fact that NODs themselves occupy intestinal volume to increase fecal volume and promote intestinal motility, several studies have shown that NODs also have significant benefits in alleviating constipation in terms of microbiota remodeling [84,172,173,174,175]. Specifically, Lactic acid bacteria utilize FOS to improve fecal consistency and reduce defecation time [176]. IMO effectively improves fecal consistency and frequency of defecation of subjects, which may be attributed to increased abundance of some microorganisms after colonic fermentation [174,175]. Lotus seed oligosaccharides alleviate constipation by stimulating the growth of beneficial bacteria in the host intestine to promote intestinal motility and defecation [177]. In addition, Zhang et al. demonstrated that COS could improve constipation by regulating the production of intestinal metabolites. This regulatory effect is mainly achieved by reshaping the structure of the gut microbiota in constipated mice [178].
The intake of NODs increases the substrate for probiotic fermentation and promotes the production of SCFAs, which stimulate the growth of colonic epithelial cells, thereby restoring intestinal morphology and increasing intestinal motility to alleviate constipation [179]. Current research suggests that butyrate improves intestinal motility by affecting water and electrolyte metabolism. Similarly, COS relieves constipation by restoring deteriorated water-electrolyte metabolism, including AQP3/4 and ENaC-β/γ expression. Among them, AQP3/4 transports water from the intestinal lumen to the colonic epithelium, ENaC-β/γ mediates the uptake of Na⁺ into colonic epithelial cells [178,180].
In the current literature, the improvement of constipation by NODs may be related to the regulation of some metabolic pathways, including lipid metabolism (e.g., sphingolipid metabolism, glycerophospholipid metabolism and arachidonic acid metabolism), bile acid metabolism, and tryptophan metabolism [169,180]. Sphingolipid metabolism and glycerophospholipid metabolism have been proved to be associated with inflammatory responses to intestinal disorders. Inflammation will damage the integrity of intestinal epithelium, which reduces the secretion of neurotransmitters and hormones, affects gastrointestinal motility, and induces constipation [181]. Tryptophan generation has a significant accelerating effect on gastrointestinal motility. In this pathway, L-tryptophan, 5-HTP and other intermediate metabolites are substrates produced by neurotransmitters and neuromodulators, which can enhance the sensitivity of visceral nerves in the gastrointestinal tract and stimulate gastrointestinal motility. Several studies have shown that NODs can relieve constipation by activating the 5-HT signaling pathway [179,182]. In addition, it is equally important that bile acids regulate constipation through the metabolism of intestinal bacteria, such as Bacteroides [178].
In conclusion, NODs can effectively relieve constipation through a variety of mechanisms, such as promoting intestinal motility, enhancing intestinal barrier function, and regulating immune response (Figure 4). These findings indicate the potential value of NODs in the treatment of constipation, further supporting the direction of research to improve constipation by targeting the gut microbiota and its metabolic functions.
5. Conclusions and Perspectives
NODs, as prebiotics, show remarkable potential in alleviating a variety of human chronic diseases, such as diabetes, depression, constipation, colitis, and obesity, by modulating the gut microbiota to produce beneficial metabolites. However, the relationship between the precise structure of oligosaccharides and their fermentation state in the gut remains unclear, which limits their application.
In the future, it is necessary to study the interaction between the structure of NODs and gut microbiota, further explore the specific mechanism of NODs in different chronic diseases, and compare the effects of different oligosaccharide types. The efficacy and safety of NODs have been demonstrated in multi-center, large-scale human clinical trials. We can make better use of NODs to develop new dietary supplement and functional foods, to provide effective prevention and treatment strategies for chronic diseases.
Author Contributions
Meiyu Yuan: Data curation, Formal analysis, Methodology, Writing – original draft, review & editing. Zhongwei Zhang: Data curation, Formal analysis, Writing – original draft, review & editing. Tongying Liu: Formal analysis, Methodology, Writing – original draft. Hua Feng: Formal analysis, Methodology, review & editing. Kai Chen: Formal analysis, Data curation. Yuhuan Liu: Project administration, Resources, Supervision, Writing – original draft, review & editing.
Acknowledgments
This research was funded by the Natural Science Foundation of Chongqing City (No. CSTB2022NSCQ-MSX1634), the Research Project of the State Key Laboratory of Food Science and Resource of Nanchang University (SKLF-ZZB-202321), the Technology System of Pueraria Industry in Jiangxi Province (JXARS-16) and 'Leading the Charge with Open Competition' Project of Shangrao City of Jiangxi Province (K8000403T).
Conflicts of Interest
There are no conflicts of interest to declare.
References
- Costello, S.P.; Hughes, P.A.; Waters, O.; Bryant, R.V.; Vincent, A.D.; Blatchford, P.; Katsikeros, R.; Makanyanga, J.; Campaniello, M.A.; Mavrangelos, C.; Rosewarne, C.P.; Bickley, C.; Peters, C.; Schoeman, M.N.; Conlon, M.A.; Roberts-Thomson, I.C.; Andrews, J.M. Effect of Fecal Microbiota Transplantation on 8-Week Remission in Patients with Ulcerative Colitis a Randomized Clinical Trial. Jama-J Am Med Assoc. 2019, 321, 156–164. [Google Scholar] [CrossRef]
- El-Salhy, M.; Casen, C.; Valeur, J.; Hausken, T.; Hatlebakk, J.G. Responses to faecal microbiota transplantation in female and male patients with irritable bowel syndrome. World J. Gastroenterol. 2021, 27, 2219–2237. [Google Scholar] [CrossRef]
- He, L.; Chen, R.; Zhang, B.; Zhang, S.; Khan, B.A.; Zhu, D.; Wu, Z.; Xiao, C.; Chen, B.; Chen, F.; et al. Fecal microbiota transplantation treatment of autoimmune-mediated type 1 diabetes mellitus. Front. Immunol. 2022, 13, 930872. [Google Scholar] [CrossRef]
- Kim, M.S.; Kim, Y.; Choi, H.; Kim, W.; Park, S.; Lee, D.; Kim, D.K.; Kim, H.J.; Choi, H.; Hyun, D.W.; Lee, J.Y.; Choi, E.Y.; Lee, D.S.; Bae, J.W.; Mook-Jung, I. Transfer of a Healthy Microbiota Reduces Amyloid and Tau Pathology in an Alzheimer's Disease Animal Model. Gut 2020, 69, 283–294. [Google Scholar] [CrossRef]
- Xue, L.; Deng, Z.; Luo, W.; He, X.; Chen, Y. Effect of Fecal Microbiota Transplantation on Non-Alcoholic Fatty Liver Disease: A Randomized Clinical Trial. Front. Cell. Infect. Microbiol. 2022, 12, 759306. [Google Scholar] [CrossRef]
- Zhu, X.; Zhao, L.; Lei, L.; Zhu, Y.; Xu, J.; Liu, L. Fecal microbiota transplantation ameliorates abdominal obesity through inhibiting microbiota-mediated intestinal barrier damage and inflammation in mice. Microbiol. Res. 2024, 282, 127654. [Google Scholar] [CrossRef]
- de Vos, W.M.; Tilg, H.; Van Hul, M.; Cani, P.D. Gut microbiome and health: mechanistic insights. Gut 2022, 71, 1020–1032. [Google Scholar] [CrossRef]
- Krautkramer, K.A.; Fan, J.; Bäckhed, F. Gut microbial metabolites as multi-kingdom intermediates. Nat. Rev. Microbiol. 2020, 19, 77–94. [Google Scholar] [CrossRef]
- Chassaing, B.; Miles-Brown, J.; Pellizzon, M.; Ulman, E.; Ricci, M.; Zhang, L.; Patterson, A.D.; Vijay-Kumar, M.; Gewirtz, A.T.; Klurfeld, D.M.; et al. Lack of soluble fiber drives diet-induced adiposity in mice. Am. J. Physiol. Liver Physiol. 2015, 309, G528–G541. [Google Scholar] [CrossRef]
- Cotillard, A.; Kennedy, S.P.; Kong, L.C.; Prifti, E.; Pons, N.; Le Chatelier, E.; Almeida, M.; Quinquis, B.; Levenez, F.; Galleron, N.; et al. Dietary intervention impact on gut microbial gene richness. Nature 2013, 500, 585–588. [Google Scholar] [CrossRef]
- Zhu, J.; Ren, H.; Zhong, H.; Li, X.; Zou, Y.; Han, M.; Li, M.; Madsen, L.; Kristiansen, K.; Xiao, L. An Expanded Gene Catalog of Mouse Gut Metagenomes. mSphere 2021, 6. [Google Scholar] [CrossRef]
- Chambers, E.S.; Preston, T.; Frost, G.; Morrison, D.J. Role of Gut Microbiota-Generated Short-Chain Fatty Acids in Metabolic and Cardiovascular Health. Curr. Nutr. Rep. 2018, 7, 198–206. [Google Scholar] [CrossRef]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- Wu, D.-T.; Nie, X.-R.; Gan, R.-Y.; Guo, H.; Fu, Y.; Yuan, Q.; Zhang, Q.; Qin, W. In vitro digestion and fecal fermentation behaviors of a pectic polysaccharide from okra (Abelmoschus esculentus) and its impacts on human gut microbiota. Food Hydrocoll. 2020, 114, 106577. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An Obesity-Associated Gut Microbiome with Increased Capacity for Energy Harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I.; Bergström, G.; Behre, C.J.; Fagerberg, B.; Nielsen, J.; Bäckhed, F. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013, 498, 99–103. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Z.; Xu, X.; Jiang, H.; Cai, C.; Yu, G. Odd-numbered agaro-oligosaccharides alleviate type 2 diabetes mellitus and related colonic microbiota dysbiosis in mice. Carbohydr. Polym. 2020, 240, 116261. [Google Scholar] [CrossRef]
- Wu, H.; Tremaroli, V.; Schmidt, C.; Lundqvist, A.; Olsson, L.M.; Krämer, M.; Gummesson, A.; Perkins, R.; Bergström, G.; Bäckhed, F. The Gut Microbiota in Prediabetes and Diabetes: A Population-Based Cross-Sectional Study. Cell Metab. 2020, 32, 379–390. [Google Scholar] [CrossRef]
- Loomba, R.; Seguritan, V.; Li, W.; Long, T.; Klitgord, N.; Bhatt, A.; Dulai, P.S.; Caussy, C.; Bettencourt, R.; Highlander, S.K.; et al. Gut Microbiome-Based Metagenomic Signature for Non-invasive Detection of Advanced Fibrosis in Human Nonalcoholic Fatty Liver Disease. Cell Metab. 2017, 25, 1054–1062. [Google Scholar] [CrossRef]
- Villablanca, E.J.; Selin, K.; Hedin, C.R.H. Mechanisms of Mucosal Healing: Treating Inflammatory Bowel Disease without Immunosuppression? Nat Rev Gastro Hepat. 2022, 19, 493–507. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Arze, C.; Ananthakrishnan, A.N.; Schirmer, M.; Avila-Pacheco, J.; Poon, T.W.; Andrews, E.; Ajami, N.J.; Bonham, K.S.; Brislawn, C.J.; et al. Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature 2019, 569, 655–662. [Google Scholar] [CrossRef]
- Wan, T.; Wang, Y.; He, K.; Zhu, S. Microbial sensing in the intestine. Protein Cell 2023, 14, 824–860. [Google Scholar] [CrossRef]
- Rajca, S.; Grondin, V.; Louis, E.; Vernier-Massouille, G.; Grimaud, J.C.; Bouhnik, Y.; Laharie, D.; Dupas, J.L.; Pillant, H.; Picon, L.; Veyrac, M.; Flamant, M.; Savoye, G.; Jian, R.; Devos, M.; Paintaud, G.; Piver, E.; Allez, M.; Mary, J.Y.; Sokol, H.; Colombel, J.F.; Seksik, P. Alterations in the Intestinal Microbiome (Dysbiosis) as a Predictor of Relapse after Infliximab Withdrawal in Crohn's Disease. Inflamm Bowel Dis 2014, 20, 978–986. [Google Scholar]
- Zhang, W.; Lyu, M.; Bessman, N.J.; Xie, Z.; Arifuzzaman, M.; Yano, H.; Parkhurst, C.N.; Chu, C.; Zhou, L.; Putzel, G.G.; et al. Gut-innervating nociceptors regulate the intestinal microbiota to promote tissue protection. Cell 2022, 185, 4170–4189. [Google Scholar] [CrossRef]
- Ni, J.; Wu, G.D.; Albenberg, L.; Tomov, V.T. Gut Microbiota and IBD: Causation or Correlation? Nat Rev Gastro Hepat. 2017, 14, 573–584. [Google Scholar] [CrossRef]
- Imhann, F.; Vila, A.V.; Bonder, M.J.; Fu, J.; Gevers, D.; Visschedijk, M.C.; Spekhorst, L.M.; Alberts, R.; Franke, L.; van Dullemen, H.M.; et al. Interplay of host genetics and gut microbiota underlying the onset and clinical presentation of inflammatory bowel disease. Gut 2016, 67, 108–119. [Google Scholar] [CrossRef]
- Liu, L.; Li, M.; Yu, M.; Shen, M.; Wang, Q.; Yu, Y.; Xie, J. Natural polysaccharides exhibit anti-tumor activity by targeting gut microbiota. Int. J. Biol. Macromol. 2018, 121, 743–751. [Google Scholar] [CrossRef]
- Mutlu, E.; Mikolaitis, S.; Sedghi, S.; Chakradeo, P.S.; Engen, P.; Chlipala, G.; Green, S.; Keshavarzian, A. Mo1795 Dietary Treatment of Crohn's Disease: A Randomized, Placebo-Controlled, Double-Blinded Clinical Trial. Gastroenterology 2016, 150. [Google Scholar] [CrossRef]
- Morgan, X.C.; Tickle, T.L.; Sokol, H.; Gevers, D.; Devaney, K.L.; Ward, D.V.; A Reyes, J.; A Shah, S.; LeLeiko, N.; Snapper, S.B.; et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012, 13, R79–R79. [Google Scholar] [CrossRef]
- Gevers, D.; Kugathasan, S.; Denson, L.A.; Vázquez-Baeza, Y.; Van Treuren, W.; Ren, B.; Schwager, E.; Knights, D.; Song, S.J.; Yassour, M.; et al. The Treatment-Naive Microbiome in New-Onset Crohn’s Disease. Cell Host Microbe 2014, 15, 382–392. [Google Scholar] [CrossRef]
- Willing, B.P.; Dicksved, J.; Halfvarson, J.; Andersson, A.F.; Lucio, M.; Zheng, Z.; Järnerot, G.; Tysk, C.; Jansson, J.K.; Engstrand, L. A Pyrosequencing Study in Twins Shows That Gastrointestinal Microbial Profiles Vary With Inflammatory Bowel Disease Phenotypes. Gastroenterology 2010, 139, 1844–1854. [Google Scholar] [CrossRef]
- Sharma, R.; Padwad, Y. Probiotic bacteria as modulators of cellular senescence: emerging concepts and opportunities. Gut Microbes 2019, 11, 335–349. [Google Scholar] [CrossRef]
- Jayanama, K.; Theou, O. Effects of Probiotics and Prebiotics on Frailty and Ageing: A Narrative Review. Curr. Clin. Pharmacol. 2020, 15, 183–192. [Google Scholar] [CrossRef]
- Setbo, E.; Campbell, K.; O’cuiv, P.; Hubbard, R. Utility of Probiotics for Maintenance or Improvement of Health Status in Older People — A Scoping Review. J. Nutr. Heal. Aging 2019, 23, 364–372. [Google Scholar] [CrossRef]
- Wei, L.K.; Lye, H.S.; Lee, Y.T.; Ooi, S.Y.; Teh, L.K.; Lim, L.N. Modifying progression of aging and reducing the risk of neurodegenerative diseases by probiotics and synbiotics. Front. Biosci. 2018, 10, 344–351. [Google Scholar] [CrossRef]
- Maes, M.; Kubera, M.; Leunis, J.-C.; Berk, M. Increased IgA and IgM responses against gut commensals in chronic depression: Further evidence for increased bacterial translocation or leaky gut. J. Affect. Disord. 2012, 141, 55–62. [Google Scholar] [CrossRef]
- Simpson, C.A.; Diaz-Arteche, C.; Eliby, D.; Schwartz, O.S.; Simmons, J.G.; Cowan, C.S. The gut microbiota in anxiety and depression – A systematic review. Clin. Psychol. Rev. 2020, 83, 101943. [Google Scholar] [CrossRef]
- Jiang, H.-Y.; Zhang, X.; Yu, Z.-H.; Zhang, Z.; Deng, M.; Zhao, J.-H.; Ruan, B. Altered gut microbiota profile in patients with generalized anxiety disorder. J. Psychiatr. Res. 2018, 104, 130–136. [Google Scholar] [CrossRef]
- Madan, A.; Thompson, D.; Fowler, J.C.; Ajami, N.; Salas, R.; Frueh, B.; Bradshaw, M.; Weinstein, B.; Oldham, J.; Petrosino, J. The gut microbiota is associated with psychiatric symptom severity and treatment outcome among individuals with serious mental illness. J. Affect. Disord. 2020, 264, 98–106. [Google Scholar] [CrossRef]
- Naseribafrouei, A.; Hestad, K.; Avershina, E.; Sekelja, M.; Linløkken, A.; Wilson, R.; Rudi, K. Correlation between the human fecal microbiota and depression. Neurogastroenterol. Motil. 2014, 26, 1155–1162. [Google Scholar] [CrossRef]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The Impact of Dietary Fiber on Gut Microbiota in Host Health and Disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef]
- Sonnenburg, E.D.; Smits, S.A.; Tikhonov, M.; Higginbottom, S.K.; Wingreen, N.S.; Sonnenburg, J.L. Diet-induced extinctions in the gut microbiota compound over generations. Nature 2016, 529, 212–215. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, Y.; Lu, Y.; Hao, H.; Liu, J.; Huang, R. Structural features, interaction with the gut microbiota and anti-tumor activity of oligosaccharides. RSC Adv. 2020, 10, 16339–16348. [Google Scholar] [CrossRef]
- Zhang, N.; Jin, M.; Wang, K.; Zhang, Z.; Shah, N.P.; Wei, H. Functional oligosaccharide fermentation in the gut: Improving intestinal health and its determinant factors-A review. Carbohydr. Polym. 2022, 284, 119043. [Google Scholar] [CrossRef]
- Divyashri, G.; Karthik, P.; Murthy, T.P.K.; Priyadarshini, D.; Reddy, K.R.; Raghu, A.V.; Vaidyanathan, V.K. Non-digestible oligosaccharides-based prebiotics to ameliorate obesity: Overview of experimental evidence and future perspectives. Food Sci. Biotechnol. 2023, 32, 1993–2011. [Google Scholar] [CrossRef]
- Hu, M.; Li, M.; Li, C.; Miao, M.; Zhang, T. Effects of Human Milk Oligosaccharides in Infant Health Based on Gut Microbiota Alteration. J. Agric. Food Chem. 2023, 71, 994–1001. [Google Scholar] [CrossRef]
- Chen, X.; Hu, J.; Yang, J.; Yu, Q.; Chen, Y.; Shen, M.; Rong, L.; Xie, J. Human milk oligosaccharide 2′-fucosyllactose alleviates DSS-induced ulcerative colitis via improving intestinal barrier function and regulating gut microbiota. Food Biosci. 2024. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, J.; Tan, Q.; Deng, X.; Tsai, P.-J.; Chen, P.-H.; Ye, M.; Guo, J.; Su, Z. Nondigestible Oligosaccharides with Anti-Obesity Effects. J. Agric. Food Chem. 2019, 68, 4–16. [Google Scholar] [CrossRef]
- Duffuler, P.; Bhullar, K.S.; Wu, J. Targeting gut microbiota in osteoporosis: impact of the microbial based functional food ingredients. Food Sci. Hum. Wellness 2024, 13, 1–15. [Google Scholar] [CrossRef]
- Navarro, D.M.D.L.; Abelilla, J.J.; Stein, H.H. Structures and characteristics of carbohydrates in diets fed to pigs: a review. J. Anim. Sci. Biotechnol. 2019, 10, 1–17. [Google Scholar] [CrossRef]
- Mussatto, S.I.; Mancilha, I.M. Non-digestible oligosaccharides: A review. Carbohydr. Polym. 2007, 68, 587–597. [Google Scholar] [CrossRef]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods 2019, 8, 92. [Google Scholar] [CrossRef]
- Roberfroid, M. Prebiotics: The Concept Revisited1, J. Nutr. 2007, 137, 830S–837S. [Google Scholar] [CrossRef]
- de Sousa, V.M.C.; dos Santos, E.F.; Sgarbieri, V.C. The Importance of Prebiotics in Functional Foods and Clinical Practice. Food Nutr. Sci. 2011, 02, 133–144. [Google Scholar] [CrossRef]
- Martins, G.N.; Ureta, M.M.; Tymczyszyn, E.E.; Castilho, P.C.; Gomez-Zavaglia, A. Technological Aspects of the Production of Fructo and Galacto-Oligosaccharides. Enzymatic Synthesis and Hydrolysis. Front. Nutr. 2019, 6, 78. [Google Scholar] [CrossRef]
- Zhao, C.; Wu, Y.; Liu, X.; Liu, B.; Cao, H.; Yu, H.; Sarker, S.D.; Nahar, L.; Xiao, J. Functional properties, structural studies and chemo-enzymatic synthesis of oligosaccharides. Trends Food Sci. Technol. 2017, 66, 135–145. [Google Scholar] [CrossRef]
- Al-Sheraji, S.H.; Ismail, A.; Manap, M.Y.; Mustafa, S.; Yusof, R.M.; Hassan, F.A. Prebiotics as functional foods: A review. J. Funct. Foods 2013, 5, 1542–1553. [Google Scholar] [CrossRef]
- Precup, G.; Venus, J.; Heiermann, M.; Schneider, R.; Pop, I.D.; Vodnar, D.C. Chemical and Enzymatic Synthesis of Biobased Xylo-Oligosaccharides and Fermentable Sugars from Wheat Straw for Food Applications. Polymers 2022, 14, 1336. [Google Scholar] [CrossRef]
- Aachary, A.A.; Prapulla, S.G. Xylooligosaccharides (XOS) as an Emerging Prebiotic: Microbial Synthesis, Utilization, Structural Characterization, Bioactive Properties, and Applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 2–16. [Google Scholar] [CrossRef]
- Nopvichai, C.; Charoenwongpaiboon, T.; Luengluepunya, N.; Ito, K.; Muanprasat, C.; Pichyangkura, R. Production and purification of mannan oligosaccharide with epithelial tight junction enhancing activity. PeerJ 2019, 7, e7206. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Y. Characterization of Galacto-Oligosaccharides Using High-Performance Anion Exchange Chromatography-Tandem Mass Spectrometry. J Sep Sci. 2021, 44, 2221–2233. [Google Scholar] [CrossRef]
- Nguyen, T.H.P.; Le, N.A.T.; Tran, P.T.; Du Bui, D.; Nguyen, Q.H. Preparation of water-soluble chitosan oligosaccharides by oxidative hydrolysis of chitosan powder with hydrogen peroxide. Heliyon 2023, 9, e19565. [Google Scholar] [CrossRef]
- Milani, C.; Lugli, G.A.; Duranti, S.; Turroni, F.; Mancabelli, L.; Ferrario, C.; Mangifesta, M.; Hevia, A.; Viappiani, A.; Scholz, M.; et al. Bifidobacteria exhibit social behavior through carbohydrate resource sharing in the gut. Sci. Rep. 2015, 5, 15782–15782. [Google Scholar] [CrossRef]
- Zuniga, M.; Yebra, M.J.; Monedero, V. Complex Oligosaccharide Utilization Pathways in Lactobacillus. Curr. Issues Mol. Biol. 2020, 40, 49–80. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef]
- Sarbini, S.R.; Rastall, R.A. Prebiotics: Metabolism, Structure, and Function. Funct. Food Rev. 2011, 3, 93–106. [Google Scholar]
- Singh, R.D.; Muir, J.; Arora, A. Concentration of xylooligosaccharides with a low degree of polymerization using membranes and their effect on bacterial fermentation. Biofuels, Bioprod. Biorefining 2020, 15, 61–73. [Google Scholar] [CrossRef]
- Huang, C.; Yu, Y.; Li, Z.; Yan, B.; Pei, W.; Wu, H. The preparation technology and application of xylo-oligosaccharide as prebiotics in different fields: A review. Front. Nutr. 2022, 9, 996811. [Google Scholar] [CrossRef]
- Sanz, M.L.; Gibson, G.R.; Rastall, R.A. Influence of Disaccharide Structure on Prebiotic Selectivity in Vitro. J. Agric. Food Chem. 2005, 53, 5192–5199. [Google Scholar] [CrossRef]
- Suzuki, N.; Aiba, Y.; Takeda, H.; Fukumori, Y.; Koga, Y. Superiority of 1-kestose, the Smallest Fructo-oligosaccharide, to a Synthetic Mixture of Fructo-oligosaccharides in the Selective Stimulating Activity on Bifidobacteria. Biosci. Microflora 2006, 25, 109–116. [Google Scholar] [CrossRef]
- Böger, M.; van Leeuwen, S.S.; van Bueren, A.L.; Dijkhuizen, L. Structural Identity of Galactooligosaccharide Molecules Selectively Utilized by Single Cultures of Probiotic Bacterial Strains. J Agr Food Chem. 2019, 67, 13969–13977. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, Q.; Wang, T.; Su, Y.; Huang, C.; Lai, C.; Yong, Q. Evaluation of prebiotic ability of xylo-oligosaccharide fractions with different polymerization degrees from bamboo shoot shells. Food Bioprod. Process. 2024, 143, 202–211. [Google Scholar] [CrossRef]
- Immerzeel, P.; Falck, P.; Galbe, M.; Adlercreutz, P.; Karlsson, E.N.; Stålbrand, H. Extraction of water-soluble xylan from wheat bran and utilization of enzymatically produced xylooligosaccharides by Lactobacillus, Bifidobacterium and Weissella spp. LWT 2014, 56, 321–327. [Google Scholar] [CrossRef]
- Falck, P.; Precha-Atsawanan, S.; Grey, C.; Immerzeel, P.; Stålbrand, H.; Adlercreutz, P.; Karlsson, E.N. Correction to Xylooligosaccharides from Hardwood and Cereal Xylans Produced by a Thermostable Xylanase as Carbon Sources for Lactobacillus brevis and Bifidobacterium adolescentis. J. Agric. Food Chem. 2013, 61, 12744–12744. [Google Scholar] [CrossRef]
- Wang, J.; Sun, B.; Cao, Y.; Wang, C. In vitro fermentation of xylooligosaccharides from wheat bran insoluble dietary fiber by Bifidobacteria. Carbohydr. Polym. 2010, 82, 419–423. [Google Scholar] [CrossRef]
- Ryan, M.P.; Pembroke, J.T. Brevundimonas Spp: Emerging Global Opportunistic Pathogens. Virulence 2018, 9, 480–493. [Google Scholar] [CrossRef]
- Grimoud, J.; Durand, H.; Courtin, C.; Monsan, P.; Ouarné, F.; Theodorou, V.; Roques, C. In vitro screening of probiotic lactic acid bacteria and prebiotic glucooligosaccharides to select effective synbiotics. Anaerobe 2010, 16, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, T.; Kohmoto, T.; Kikuchi, H.; Shiota, M.; Iino, H.; Mitsuoka, T. Effects of Isomaltooligosaccharides with Different Degrees of Polymerization on Human Fecal Bifidobactcria. Biosci. Biotechnol. Biochem. 1994, 58, 2288–2290. [Google Scholar] [CrossRef]
- Sanz, M.L.; Côté, G.L.; Gibson, G.R.; Rastall, R.A. Influence of Glycosidic Linkages and Molecular Weight on the Fermentation of Maltose-Based Oligosaccharides by Human Gut Bacteria. J. Agric. Food Chem. 2006, 54, 9779–9784. [Google Scholar] [CrossRef]
- Wu, Q.; Pi, X.; Liu, W.; Chen, H.; Yin, Y.; Yu, H.D.; Wang, X.; Zhu, L. Fermentation properties of isomaltooligosaccharides are affected by human fecal enterotypes. Anaerobe 2017, 48, 206–214. [Google Scholar] [CrossRef]
- Wiater, A.; Waśko, A.; Adamczyk, P.; Gustaw, K.; Pleszczyńska, M.; Wlizło, K.; Skowronek, M.; Tomczyk, M.; Szczodrak, J. Prebiotic Potential of Oligosaccharides Obtained by Acid Hydrolysis of α-(1→3)-Glucan from Laetiporus sulphureus: A Pilot Study. Molecules 2020, 25, 5542. [Google Scholar] [CrossRef]
- Zeng, M.; Li, N.; Astmann, T.; Oh, J.-H.; van Pijkeren, J.-P.; Pan, X. Facile and efficient chemical synthesis of gluco-oligosaccharides (GlcOS) with diverse glycosidic linkages as potential prebiotics to promote the growth of probiotic bacteria. Food Res. Int. 2023, 165, 112436. [Google Scholar] [CrossRef]
- Zeng, M.J.; Oh, J.H.; van Pijkeren, J.P.; Pan, X.J. Selective Utilization of Gluco-Oligosaccharides by Lactobacilli: A Mechanism Study Revealing the Impact of Glycosidic Linkages and Degree of Polymerization on Their Utilization. Int J Food Sci. 2024, 89, 523–539. [Google Scholar] [CrossRef]
- Djouzi, Z.; Andlueux, C. Compared effects of three oligosaccharides on metabolism of intestinal microflora in rats inoculated with a human faecal flora. Br. J. Nutr. 1997, 78, 313–324. [Google Scholar] [CrossRef]
- Gibson, G.R.; Beatty, E.R.; Wang, X.; Cummings, J.H. Selective stimulation of bifidobacteria in the human colon by oligofructose and inulin. Gastroenterology 1995, 108, 975–982. [Google Scholar] [CrossRef]
- Ito, M.; Deguchi, Y.; Matsumoto, K.; Kimura, M.; Onodera, N.; Yajima, T. Influence of Galactooligosaccharides on the Human Fecal Microflora. J. Nutr. Sci. Vitaminol. 1993, 39, 635–640. [Google Scholar] [CrossRef]
- Goh, Y.J.; Klaenhammer, T.R. Genetic Mechanisms of Prebiotic Oligosaccharide Metabolism in Probiotic Microbes. Annu Rev Food Sci T. 2015, 6, 137–56. [Google Scholar] [CrossRef]
- Kittibunchakul, S.; Maischberger, T.; Domig, K.J.; Kneifel, W.; Nguyen, H.M.; Haltrich, D.; Nguyen, H. Fermentability of a Novel Galacto-Oligosaccharide Mixture by Lactobacillus Spp. And Bifidobacterium Spp. Molecules 2018, 23, 3352. [Google Scholar] [CrossRef]
- Cardelle-Cobas, A.; Corzo, N.; Olano, A.; Peláez, C.; Requena, T.; Ávila, M. Galactooligosaccharides derived from lactose and lactulose: Influence of structure on Lactobacillus, Streptococcus and Bifidobacterium growth. Int. J. Food Microbiol. 2011, 149, 81–87. [Google Scholar] [CrossRef]
- Palframan, R.J.; Gibson, G.R.; A Rastall, R. Carbohydrate preferences of Bifidobacterium species isolated from the human gut. . 2003, 4, 71–5. [Google Scholar]
- Kabel, M.A.; Kortenoeven, L.; Schols, H.A.; Voragen, A.G.J. In Vitro Fermentability of Differently Substituted Xylo-oligosaccharides. J. Agric. Food Chem. 2002, 50, 6205–6210. [Google Scholar] [CrossRef] [PubMed]
- Ho, A.L.; Kosik, O.; Lovegrove, A.; Charalampopoulos, D.; Rastall, R.A. In vitro fermentability of xylo-oligosaccharide and xylo-polysaccharide fractions with different molecular weights by human faecal bacteria. Carbohydr. Polym. 2018, 179, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Mendis, M.; Martens, E.C.; Simsek, S.S. How Fine Structural Differences of Xylooligosaccharides and Arabinoxylooligosaccharides Regulate Differential Growth of Bacteroides Species. J. Agric. Food Chem. 2018, 66, 8398–8405. [Google Scholar] [CrossRef]
- Pereira, G.V.; Abdel-Hamid, A.M.; Dutta, S.; D’alessandro-Gabazza, C.N.; Wefers, D.; Farris, J.A.; Bajaj, S.; Wawrzak, Z.; Atomi, H.; Mackie, R.I.; et al. Degradation of complex arabinoxylans by human colonic Bacteroidetes. Nat. Commun. 2021, 12, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Yasuma, T.; Toda, M.; Abdel-Hamid, A.M.; D’alessandro-Gabazza, C.; Kobayashi, T.; Nishihama, K.; D’alessandro, V.F.; Pereira, G.V.; Mackie, R.I.; Gabazza, E.C.; et al. Degradation Products of Complex Arabinoxylans by Bacteroides intestinalis Enhance the Host Immune Response. Microorganisms 2021, 9, 1126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhong, Y.; Dong, D.; Zheng, Z.; Hu, J. Gut microbial utilization of xylan and its implication in gut homeostasis and metabolic response. Carbohydr. Polym. 2022, 286, 119271. [Google Scholar] [CrossRef]
- Chassard, C.; Goumy, V.; Leclerc, M.; Del'Homme, C.; Bernalier-Donadille, A. Characterization of the xylan-degrading microbial community from human faeces. FEMS Microbiol. Ecol. 2007, 61, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Reuhs, B.L.; Cantu-Jungles, T.M.; Tuncil, Y.E.; Kaur, A.; Terekhov, A.; Martens, E.C.; Hamaker, B.R. Corn arabinoxylan has a repeating structure of subunits of high branch complexity with slow gut microbiota fermentation. Carbohydr. Polym. 2022, 289, 119435. [Google Scholar] [CrossRef]
- Yang, X.; Zeng, D.; Li, C.; Yu, W.; Xie, G.; Zhang, Y.; Lu, W. Therapeutic potential and mechanism of functional oligosaccharides in inflammatory bowel disease: a review. Food Sci. Hum. Wellness 2023, 12, 2135–2150. [Google Scholar] [CrossRef]
- Kobayashi, M.; Funane, K.; Oguma, T. Inhibition of Dextran and Mutan Synthesis by Cycloisomaltooligosaccharides. Biosci. Biotechnol. Biochem. 1995, 59, 1861–1865. [Google Scholar] [CrossRef]
- Sorndech, W.; Na Nakorn, K.; Tongta, S.; Blennow, A. Isomalto-oligosaccharides: Recent insights in production technology and their use for food and medical applications. LWT 2018, 95, 135–142. [Google Scholar] [CrossRef]
- Kaplan, H.; Hutkins, R.W. Fermentation of Fructooligosaccharides by Lactic Acid Bacteria and Bifidobacteria. Appl. Environ. Microbiol. 2000, 66, 2682–2684. [Google Scholar] [CrossRef]
- Qiang, X.; YongLie, C.; QianBing, W. Health benefit application of functional oligosaccharides. Carbohydr. Polym. 2009, 77, 435–441. [Google Scholar] [CrossRef]
- Song, Y.; Wu, M.-S.; Tao, G.; Lu, M.-W.; Lin, J.; Huang, J.-Q. Feruloylated oligosaccharides and ferulic acid alter gut microbiome to alleviate diabetic syndrome. Food Res. Int. 2020, 137, 109410. [Google Scholar] [CrossRef]
- Jana, U.K.; Kango, N.; Pletschke, B. Hemicellulose-Derived Oligosaccharides: Emerging Prebiotics in Disease Alleviation. Front. Nutr. 2021, 8. [Google Scholar] [CrossRef]
- Weninger, S.N.; Ding, A.; Browne, E.N.; Frost, M.L.; Schiro, G.; Laubitz, D.; Duca, F.A. Longitudinal Characterization of the Gut Microbiota in the Diabetic ZDSD Rat Model and Therapeutic Potential of Oligofructose. Metabolites 2023, 13, 660. [Google Scholar] [CrossRef]
- Liu, F.; Li, P.; Chen, M.; Luo, Y.; Prabhakar, M.; Zheng, H.; He, Y.; Qi, Q.; Long, H.; Zhang, Y.; et al. Fructooligosaccharide (FOS) and Galactooligosaccharide (GOS) Increase Bifidobacterium but Reduce Butyrate Producing Bacteria with Adverse Glycemic Metabolism in healthy young population. Sci. Rep. 2017, 7, 11789. [Google Scholar] [CrossRef]
- Gobinath, D.; Madhu, A.N.; Prashant, G.; Srinivasan, K.; Prapulla, S.G. Beneficial effect of xylo-oligosaccharides and fructo-oligosaccharides in streptozotocin-induced diabetic rats. Br. J. Nutr. 2010, 104, 40–47. [Google Scholar] [CrossRef]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef]
- Blaak, E.E.; Canfora, E.E.; Theis, S.; Frost, G.; Groen, A.K.; Mithieux, G.; Nauta, A.; Scott, K.; Stahl, B.; Van Harsselaar, J.; et al. Short chain fatty acids in human gut and metabolic health. Benef. Microbes 2020, 11, 411–455. [Google Scholar] [CrossRef]
- Hu, J.; Lin, S.; Zheng, B.; Cheung, P.C.K. Short-chain fatty acids in control of energy metabolism. Crit. Rev. Food Sci. Nutr. 2017, 58, 1243–1249. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Z.; Li, J.; Liu, W.; Warda, M.; Cui, B.; El-Aty, A.M.A. Oligosaccharides derived from Lycium barbarum ameliorate glycolipid metabolism and modulate the gut microbiota community and the faecal metabolites in a type 2 diabetes mouse model: metabolomic bioinformatic analysis. Food Funct. 2022, 13, 5416–5429. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, W.; Ni, X.; Farag, M.A.; Capanoglu, E.; Zhao, C. Regulatory mechanisms of the green alga Ulva lactuca oligosaccharide via the metabolomics and gut microbiome in diabetic mice. Curr. Res. Food Sci. 2022, 5, 1127–1139. [Google Scholar] [CrossRef]
- Khat-Udomkiri, N.; Toejing, P.; Sirilun, S.; Chaiyasut, C.; Lailerd, N. Antihyperglycemic effect of rice husk derived xylooligosaccharides in high-fat diet and low-dose streptozotocin-induced type 2 diabetic rat model. Food Sci. Nutr. 2019, 8, 428–444. [Google Scholar] [CrossRef]
- Bao, S.; Wang, X.; Cho, S.B.; Wu, Y.-L.; Wei, C.; Han, S.; Bao, L.; Wu, Q.; Ao, W.; Nan, J.-X. Agriophyllum Oligosaccharides Ameliorate Diabetic Insulin Resistance Through INS-R/IRS/Glut4-Mediated Insulin Pathway in db/db Mice and MIN6 Cells. Front. Pharmacol. 2021, 12. [Google Scholar] [CrossRef]
- Zhu, D.; Yan, Q.; Li, Y.; Liu, J.; Liu, H.; Jiang, Z. Effect of Konjac Mannan Oligosaccharides on Glucose Homeostasis via the Improvement of Insulin and Leptin Resistance In Vitro and In Vivo. Nutrients 2019, 11, 1705. [Google Scholar] [CrossRef]
- Gupta, A.; Osadchiy, V.; Mayer, E.A. Brain-Gut-Microbiome Interactions in Obesity and Food Addiction. Nat Rev Gastro Hepat. 2020, 17, 655–672. [Google Scholar] [CrossRef]
- Everard, A.; Lazarevic, V.; Derrien, M.; Girard, M.; Muccioli, G.G.; Neyrinck, A.M.; Possemiers, S.; Van Holle, A.; François, P.; de Vos, W.M.; et al. Responses of Gut Microbiota and Glucose and Lipid Metabolism to Prebiotics in Genetic Obese and Diet-Induced Leptin-Resistant Mice. Diabetes 2011, 60, 2775–2786. [Google Scholar] [CrossRef]
- Long, J.; Yang, J.; Henning, S.M.; Woo, S.L.; Hsu, M.; Chan, B.; Heber, D.; Li, Z. Xylooligosaccharide supplementation decreases visceral fat accumulation and modulates cecum microbiome in mice. J. Funct. Foods 2018, 52, 138–146. [Google Scholar] [CrossRef]
- Thiennimitr, P.; Yasom, S.; Tunapong, W.; Chunchai, T.; Wanchai, K.; Pongchaidecha, A.; Lungkaphin, A.; Sirilun, S.; Chaiyasut, C.; Chattipakorn, N.; et al. Lactobacillus paracasei HII01, xylooligosaccharides, and synbiotics reduce gut disturbance in obese rats. Nutrition 2018, 54, 40–47. [Google Scholar] [CrossRef]
- Dewulf, E.M.; Cani, P.D.; Claus, S.P.; Fuentes, S.; Puylaert, P.G.; Neyrinck, A.M.; Bindels, L.B.; de Vos, W.M.; Gibson, G.R.; Thissen, J.-P.; et al. Insight into the prebiotic concept: lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women. Gut 2012, 62, 1112–1121. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, J.; Xie, F.; He, H.; Johnston, L.J.; Dai, X.; Wu, C.; Ma, X. Dietary fiber-derived short-chain fatty acids: A potential therapeutic target to alleviate obesity-related nonalcoholic fatty liver disease. Obes. Rev. 2021, 22, e13316. [Google Scholar] [CrossRef]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef]
- Kong, S.; Huang, X.; Cao, H.; Bai, Y.; Che, Q.; Nie, H.; Su, Z. Anti-obesity effects of galacto-oligosaccharides in obese rats. Eur. J. Pharmacol. 2021, 917, 174728. [Google Scholar] [CrossRef]
- Liu, Q.; Xi, Y.; Wang, Q.; Liu, J.; Li, P.; Meng, X.; Liu, K.; Chen, W.; Liu, X.; Liu, Z. Mannan Oligosaccharide Attenuates Cognitive and Behavioral Disorders in the 5xfad Alzheimer's Disease Mouse Model Via Regulating the Gut Microbiota-Brain Axis. Brain Behav Immun. 2021, 95, 330–343. [Google Scholar] [CrossRef]
- Chen, K.; Hu, M.; Tang, M.; Gao, C.; Wang, H.; Man, S.; Lu, F. Oligosaccharide and short-chain fatty acid: A double-edged sword in obese mice by regulating food intake and fat synthesis. Food Res. Int. 2022, 159, 111619. [Google Scholar] [CrossRef]
- Lun, W.; Zhou, J.; Bai, Y.; Che, Q.; Cao, H.; Guo, J.; Su, Z. Chitosan oligosaccharide activates brown adipose tissue by modulating the gut microbiota and bile acid pathways based on faecal microbiota transplantation. J. Funct. Foods 2023, 108. [Google Scholar] [CrossRef]
- Fang, S.; Suh, J.M.; Reilly, S.M.; Yu, E.; Osborn, O.; Lackey, D.; Yoshihara, E.; Perino, A.; Jacinto, S.; Lukasheva, Y.; et al. Intestinal FXR agonism promotes adipose tissue browning and reduces obesity and insulin resistance. Nat. Med. 2015, 21, 159–165. [Google Scholar] [CrossRef]
- Shapiro, H.; Kolodziejczyk, A.A.; Halstuch, D.; Elinav, E. Bile acids in glucose metabolism in health and disease. J. Exp. Med. 2018, 215, 383–396. [Google Scholar] [CrossRef]
- Calderon, G.; McRae, A.; Rievaj, J.; Davis, J.; Zandvakili, I.; Linker-Nord, S.; Burton, D.; Roberts, G.; Reimann, F.; Gedulin, B.; et al. Ileo-colonic delivery of conjugated bile acids improves glucose homeostasis via colonic GLP-1-producing enteroendocrine cells in human obesity and diabetes. EBioMedicine 2020, 55, 102759. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Feng, S.; Liu, A.B.; Wang, H.; Zeng, X.; Yang, C.S. Protective effects of α-galacto-oligosaccharides against a high-fat/western-style diet-induced metabolic abnormalities in mice. Food Funct. 2019, 10, 3660–3670. [Google Scholar] [CrossRef]
- Hu, H.; Zhang, S.; Liu, F.; Zhang, P.; Muhammad, Z.; Pan, S. Role of the Gut Microbiota and Their Metabolites in Modulating the Cholesterol-Lowering Effects of Citrus Pectin Oligosaccharides in C57BL/6 Mice. J. Agric. Food Chem. 2019, 67, 11922–11930. [Google Scholar] [CrossRef]
- Chi, L.; Khan, I.; Lin, Z.; Zhang, J.; Lee, M.S.; Leong, W.; Hsiao, W.W.; Zheng, Y. Fructo-oligosaccharides from Morinda officinalis remodeled gut microbiota and alleviated depression features in a stress rat model. Phytomedicine 2019, 67, 153157. [Google Scholar] [CrossRef] [PubMed]
- Burokas, A.; Arboleya, S.; Moloney, R.D.; Peterson, V.L.; Murphy, K.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Targeting the Microbiota-Gut-Brain Axis: Prebiotics Have Anxiolytic and Antidepressant-like Effects and Reverse the Impact of Chronic Stress in Mice. Biol. Psychiatry 2017, 82, 472–487. [Google Scholar] [CrossRef] [PubMed]
- Varesi, A.; Campagnoli, L.I.M.; Chirumbolo, S.; Candiano, B.; Carrara, A.; Ricevuti, G.; Esposito, C.; Pascale, A. The brain-gut-microbiota interplay in depression: A key to design innovative therapeutic approaches. Pharmacol. Res. 2023, 192, 106799. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Ling, Z.; Zhang, Y.; Mao, H.; Ma, Z.; Yin, Y.; Wang, W.; Tang, W.; Tan, Z.; Shi, J.; et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav. Immun. 2015, 48, 186–194. [Google Scholar] [CrossRef]
- Yong, J.; Lu, C.; Huang, S.; Wu, X. Chemical Components Isolated from the Roots of Morinda officinalis. Chem. Nat. Compd. 2015, 51, 548–549. [Google Scholar] [CrossRef]
- Barcenilla, A.; Pryde, S.E.; Martin, J.C.; Duncan, S.H.; Stewart, C.S.; Henderson, C.; Flint, H.J. Phylogenetic Relationships of Butyrate-Producing Bacteria from the Human Gut. Appl. Environ. Microbiol. 2000, 66, 1654–1661. [Google Scholar] [CrossRef]
- Kunze, W.A.; Mao, Y.; Wang, B.; Huizinga, J.D.; Ma, X.; Forsythe, P.; Bienenstock, J. Lactobacillus reuteri enhances excitability of colonic AH neurons by inhibiting calcium-dependent potassium channel opening. J. Cell. Mol. Med. 2009, 13, 2261–2270. [Google Scholar] [CrossRef]
- Li, X.; Ellis, M.L.; Knight, J. Oxalobacter formigenes Colonization and Oxalate Dynamics in a Mouse Model. Appl. Environ. Microbiol. 2015, 81, 5048–5054. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Han, Y.; Du, J.; Liu, R.; Jin, K.; Yi, W. Microbiota-gut-brain axis and the central nervous system. Oncotarget 2017, 8, 53829–53838. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Baxter, N.T.; Schmidt, A.W.; Venkataraman, A.; Kim, K.S.; Waldron, C.; Schmidt, T.M. Dynamics of Human Gut Microbiota and Short-Chain Fatty Acids in Response to Dietary Interventions with Three Fermentable Fibers. mBio 2019, 10, e02566–18. [Google Scholar] [CrossRef] [PubMed]
- Barrett, E.; Ross, R.P.; O’Toole, P.W.; Fitzgerald, G.F.; Stanton, C. Gamma-Aminobutyric acid production by culturable bacteria from the human intestine. J. Appl. Microbiol. 2012, 113, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Pi, Y.; Mu, C.; Farzi, A.; Liu, Z.; Zhu, W. Increasing carbohydrate availability in the hindgut promotes hypothalamic neurotransmitter synthesis: aromatic amino acids linking the microbiota–brain axis. J. Neurochem. 2019, 149, 641–659. [Google Scholar] [CrossRef] [PubMed]
- Hata, T.; Asano, Y.; Yoshihara, K.; Kimura-Todani, T.; Miyata, N.; Zhang, X.-T.; Takakura, S.; Aiba, Y.; Koga, Y.; Sudo, N. Regulation of gut luminal serotonin by commensal microbiota in mice. PLOS ONE 2017, 12, e0180745–e0180745. [Google Scholar] [CrossRef] [PubMed]
- Min, Q.X.; Wang, Y.; Jin, T.C.; Zhu, L.; Wu, X.Y.; Li, Y.K.; Wang, Y.J.; Xu, N. Analysis of Intestinal Short-Chain Fatty Acid Metabolism Profile after Probiotics and Glp-1 Treatment for Type 2 Diabetes Mellitus. Front Endocrinol. 2022, 13, 892127. [Google Scholar] [CrossRef] [PubMed]
- Donohoe, D.R.; Garge, N.; Zhang, X.; Sun, W.; O'Connell, T.M.; Bunger, M.K.; Bultman, S.J. The Microbiome and Butyrate Regulate Energy Metabolism and Autophagy in the Mammalian Colon. Cell Metab. 2011, 13, 517–526. [Google Scholar] [CrossRef]
- Wolever TM, S.; Chiasson, J.L. Acarbose Raises Serum Butyrate in Human Subjects Withimpaired Glucose Tolerance. Brit J Nutr. 2000, 84, 57–61. [Google Scholar] [CrossRef]
- Al-Khafaji, A.H.; Jepsen, S.D.; Christensen, K.R.; Vigsnæs, L.K. The potential of human milk oligosaccharides to impact the microbiota-gut-brain axis through modulation of the gut microbiota. J. Funct. Foods 2020, 74, 104176. [Google Scholar] [CrossRef]
- Val-Laillet, D.; Guérin, S.; Coquery, N.; Nogret, I.; Formal, M.; Romé, V.; Le Normand, L.; Meurice, P.; Randuineau, G.; Guilloteau, P.; et al. Oral sodium butyrate impacts brain metabolism and hippocampal neurogenesis, with limited effects on gut anatomy and function in pigs. FASEB J. 2018, 32, 2160–2171. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Cheng, L.; Zeng, X.; Zhang, X.; Liu, Y.; Wu, Z.; Weng, P. The intervention of unique plant polysaccharides - Dietary fiber on depression from the gut-brain axis. Int. J. Biol. Macromol. 2020, 170, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.R.; Borre, Y.; O’Brien, C.; Patterson, E.; El Aidy, S.; Deane, J.; Kennedy, P.J.; Beers, S.; Scott, K.; Moloney, G.; et al. Transferring the blues: Depression-associated gut microbiota induces neurobehavioural changes in the rat. J. Psychiatr. Res. 2016, 82, 109–118. [Google Scholar] [CrossRef]
- Deng, L.; Zhou, X.; Tao, G.; Hao, W.; Wang, L.; Lan, Z.; Song, Y.; Wu, M.; Huang, J.-Q. Ferulic acid and feruloylated oligosaccharides alleviate anxiety and depression symptom via regulating gut microbiome and microbial metabolism. Food Res. Int. 2022, 162, 111887. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-W.; Gao, C.-S.; Zhang, H.; Yang, J.; Wang, Y.-P.; Pan, L.-B.; Yu, H.; He, C.-Y.; Luo, H.-B.; Zhao, Z.-X.; et al. Morinda officinalis oligosaccharides increase serotonin in the brain and ameliorate depression via promoting 5-hydroxytryptophan production in the gut microbiota. Acta Pharm. Sin. B 2022, 12, 3298–3312. [Google Scholar] [CrossRef] [PubMed]
- He, N.; Yang, Y.; Wang, H.; Liu, N.; Yang, Z.; Li, S. Unsaturated alginate oligosaccharides (UAOS) protects against dextran sulfate sodium-induced colitis associated with regulation of gut microbiota. J. Funct. Foods 2021, 83, 104536. [Google Scholar] [CrossRef]
- Tester, R.F.; Al-Ghazzewi, F.H. Beneficial health characteristics of native and hydrolysed konjac (Amorphophallus konjac) glucomannan. J. Sci. Food Agric. 2016, 96, 3283–3291. [Google Scholar] [CrossRef]
- Zhong, Y.; Zheng, C.; Zheng, J.-. -H.; Xu, S.-.-C. The relationship between intestinal flora changes and osteoporosis in rats with inflammatory bowel disease and the improvement effect of probiotics. 2020, 24, 5697–5702.
- Huang, G.; Ye, L.; Du, G.; Huang, Y.; Wu, Y.; Ge, S.; Yang, Z.; Zhu, G. Effects of curcumin plus Soy oligosaccharides on intestinal flora of rats with ulcerative colitis. Cell. Mol. Biol. 2017, 63, 20–25. [Google Scholar] [CrossRef]
- Li, Y.; Wu, L.; Yong, Y.; Niu, X.; Gao, Y.; Zhou, Q.; Xie, H.; Liu, X.; Li, Y.; Yu, Z.; El-Aty, A.M.A.; Ju, X. Enhancing Gut Barrier Integrity: Upregulation of Tight Junction Proteins by Chitosan Oligosaccharide through the ERK1/2 Signaling Pathway. Nutrition 2024, 124, 112428. [Google Scholar] [CrossRef] [PubMed]
- Venegas, D.P.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front Immunol. 2019, 10, 00277. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.Y.; Liu, X.J.; Hao, J.Y. Gut microbiota in ulcerative colitis: insights on pathogenesis and treatment. J. Dig. Dis. 2020, 21, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Liu, X.; Li, Z.; Ren, J.; Liu, X. The effects of microbiota-targeted approaches in inflammatory bowel disease: probiotics, probiotic foods, and prebiotics. Curr. Opin. Food Sci. 2023, 49. [Google Scholar] [CrossRef]
- Scott, S.A.; Fu, J.; Chang, P.V. Microbial tryptophan metabolites regulate gut barrier function via the aryl hydrocarbon receptor. Proc. Natl. Acad. Sci. 2020, 117, 19376–19387. [Google Scholar] [CrossRef] [PubMed]
- Youssef, I.M.; Khalil, H.A.; Shakoori, A.M.; Bagadood, R.M.; Alyahyawi, A.Y.; Alhazzaa, R.A.; Fakiha, K.G.; Nasr, S.; Abo-Samra, M.A.; Hassan, M.S.; El Halim, H.S.A.; El-Hack, M.E.A.; Jaremko, M.; Al-Nemi, R.; Youssef, K.M. Immune Response, Hematological Traits, Biochemical Blood Parameters, and Histological Status of Laying Hens Influenced by Dietary Chitosan- Oligosaccharides. Poultry Sci. 2023, 102, 102834. [Google Scholar] [CrossRef]
- Xia, X.; Zhu, L.; Lei, Z.; Song, Y.; Tang, F.; Yin, Z.; Wang, J.; Huang, J. Feruloylated Oligosaccharides Alleviate Dextran Sulfate Sodium-Induced Colitis in Vivo. J. Agric. Food Chem. 2019, 67, 9522–9531. [Google Scholar] [CrossRef] [PubMed]
- Suwannaporn, P.; Thepwong, K.; Tester, R.; Al-Ghazzewi, F.; Piggott, J.; Shen, N.; Chen, Z.; Chen, F.; Yang, J.; Zhang, D.; et al. Tolerance and nutritional therapy of dietary fibre from konjac glucomannan hydrolysates for patients with inflammatory bowel disease (IBD). Bioact. Carbohydrates Diet. Fibre 2013, 2, 93–98. [Google Scholar] [CrossRef]
- Mayer, E.A.; Savidge, T.; Shulman, R.J. Brain–Gut Microbiome Interactions and Functional Bowel Disorders. Gastroenterology 2014, 146, 1500–1512. [Google Scholar] [CrossRef]
- Parthasarathy, G.; Chen, J.; Chen, X.; Chia, N.; O'Connor, H.M.; Wolf, P.G.; Gaskins, H.R.; Bharucha, A.E. Relationship Between Microbiota of the Colonic Mucosa vs Feces and Symptoms, Colonic Transit, and Methane Production in Female Patients With Chronic Constipation. Gastroenterology 2016, 150, 367–379. [Google Scholar] [CrossRef]
- Kwon, J.I.; Park, Y.; Noh, D.O.; Suh, H.J.; Han, S.H. Complex-oligosaccharide composed of galacto-oligosaccharide and lactulose ameliorates loperamide-induced constipation in rats. Food Sci. Biotechnol. 2018, 27, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Pan, M.; Li, D.; Yin, Y.; Jiang, T.; Fang, S.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Metagenomic insights into the effects of oligosaccharides on the microbial composition of cecal contents in constipated mice. J. Funct. Foods 2017, 38, 486–496. [Google Scholar] [CrossRef]
- Vandeputte, D.; Falony, G.; Vieira-Silva, S.; Tito, R.Y.; Joossens, M.; Raes, J. Stool consistency is strongly associated with gut microbiota richness and composition, enterotypes and bacterial growth rates. Gut 2016, 65, PAGE. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-L.; Lu, Y.-H.; Lin, J.-J.; Ko, L.-Y. Effects of Isomalto-Oligosaccharides on Bowel Functions and Indicators of Nutritional Status in Constipated Elderly Men. J. Am. Coll. Nutr. 2001, 20, 44–49. [Google Scholar] [CrossRef] [PubMed]
- Yen, C.-H.; Tseng, Y.-H.; Kuo, Y.-W.; Lee, M.-C.; Chen, H.-L. Long-term supplementation of isomalto-oligosaccharides improved colonic microflora profile, bowel function, and blood cholesterol levels in constipated elderly people—A placebo-controlled, diet-controlled trial. Nutrition 2011, 27, 445–450. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zheng, J.; Jiang, N.; Sun, G.; Bao, X.; Kong, M.; Cheng, X.; Lin, A.; Liu, H. Modulation of gut microbiota and intestinal metabolites by lactulose improves loperamide-induced constipation in mice. Eur. J. Pharm. Sci. 2020, 158, 105676. [Google Scholar] [CrossRef]
- Su, H.; Chen, J.; Miao, S.; Deng, K.; Liu, J.; Zeng, S.; Zheng, B.; Lu, X. Lotus seed oligosaccharides at various dosages with prebiotic activity regulate gut microbiota and relieve constipation in mice. Food Chem. Toxicol. 2019, 134, 110838. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Yang, H.; Zheng, J.; Jiang, N.; Sun, G.; Bao, X.; Lin, A.; Liu, H. Chitosan oligosaccharides attenuate loperamide-induced constipation through regulation of gut microbiota in mice. Carbohydr. Polym. 2020, 253, 117218. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, S.; Yan, Q.; Li, Y.; Jiang, Z. Effect of Konjac mannan oligosaccharides on diphenoxylate-induced constipation in mice. J. Funct. Foods 2019, 57, 399–407. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, B.; Sun, G.; Zheng, J.; Hu, H.; Yang, H.; Cheng, X.; Lin, A.; Liu, H. Plasma metabolomic profiles reveal regulatory effect of chitosan oligosaccharides on loperamide-induced constipation in mice. J. Pharm. Biomed. Anal. 2022, 211, 114590. [Google Scholar] [CrossRef]
- Yang, Z.; Ye, S.; Xu, Z.; Su, H.; Tian, X.; Han, B.; Shen, B.; Liao, Q.; Xie, Z.; Hong, Y. Dietary synbiotic ameliorates constipation through the modulation of gut microbiota and its metabolic function. Food Res. Int. 2021, 147, 110569. [Google Scholar] [CrossRef] [PubMed]
- Kendig, D.M.; Grider, J.R. Serotonin and colonic motility. Neurogastroenterol. Motil. 2015, 27, 899–905. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The effect of NODs on T2DM and Obesity by regulating gut microbiota composition and metabolites. 1) SCFAs: SCFAs can promote the release of PYY and GLP-1 from intestinal endocrine L cells, increase energy consumption of adipose tissue, reduce lipid accumulation and inflammation. In the liver, SCFAs affect the generation of fat and glucose. In the pancreas, SCFAs promote insulin secretion. SCFAs can transmit neural signals to maintain homeostasis of systemic functions. 2) Amino acids: amino acids are converted by intestinal bacteria into other bioactive metabolites, such as indole, which can stimulate L cells to secrete GLP-1, thereby promoting insulin secretion and inhibiting glucagon secretion. 3) Bile acids (BAs): BAs is converted to secondary bile acids through uncoupling, dehydroxylation and epimerization by intestinal bacteria. BAs act on two receptors (FXR and TGR5) to regulate glucose homeostasis. In addition, the activation of FXR by BAs promotes the release of FGF15/19, which promotes hepatic glycogen synthesis and reduces BAs synthesis and gluconeogenesis in the liver.
Figure 1.
The effect of NODs on T2DM and Obesity by regulating gut microbiota composition and metabolites. 1) SCFAs: SCFAs can promote the release of PYY and GLP-1 from intestinal endocrine L cells, increase energy consumption of adipose tissue, reduce lipid accumulation and inflammation. In the liver, SCFAs affect the generation of fat and glucose. In the pancreas, SCFAs promote insulin secretion. SCFAs can transmit neural signals to maintain homeostasis of systemic functions. 2) Amino acids: amino acids are converted by intestinal bacteria into other bioactive metabolites, such as indole, which can stimulate L cells to secrete GLP-1, thereby promoting insulin secretion and inhibiting glucagon secretion. 3) Bile acids (BAs): BAs is converted to secondary bile acids through uncoupling, dehydroxylation and epimerization by intestinal bacteria. BAs act on two receptors (FXR and TGR5) to regulate glucose homeostasis. In addition, the activation of FXR by BAs promotes the release of FGF15/19, which promotes hepatic glycogen synthesis and reduces BAs synthesis and gluconeogenesis in the liver.
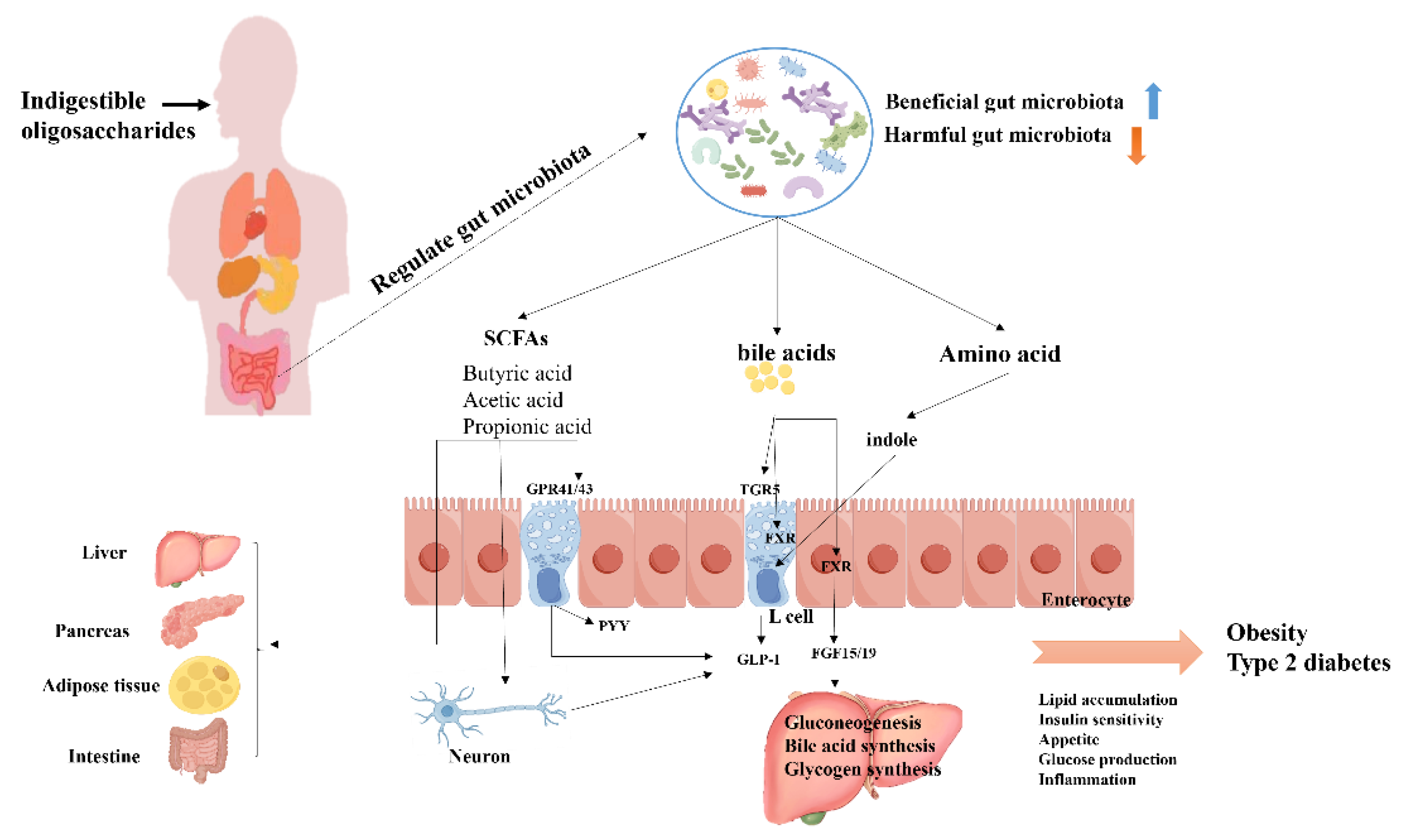
Figure 2.
Gut microbiota and the brain communicate in both directions through the immune system, intestinal endocrine signals, neurotransmitters, amino acids, BAs, SCFAs, the hypothalamic-pituitary-adrenal axis, etc.
Figure 2.
Gut microbiota and the brain communicate in both directions through the immune system, intestinal endocrine signals, neurotransmitters, amino acids, BAs, SCFAs, the hypothalamic-pituitary-adrenal axis, etc.
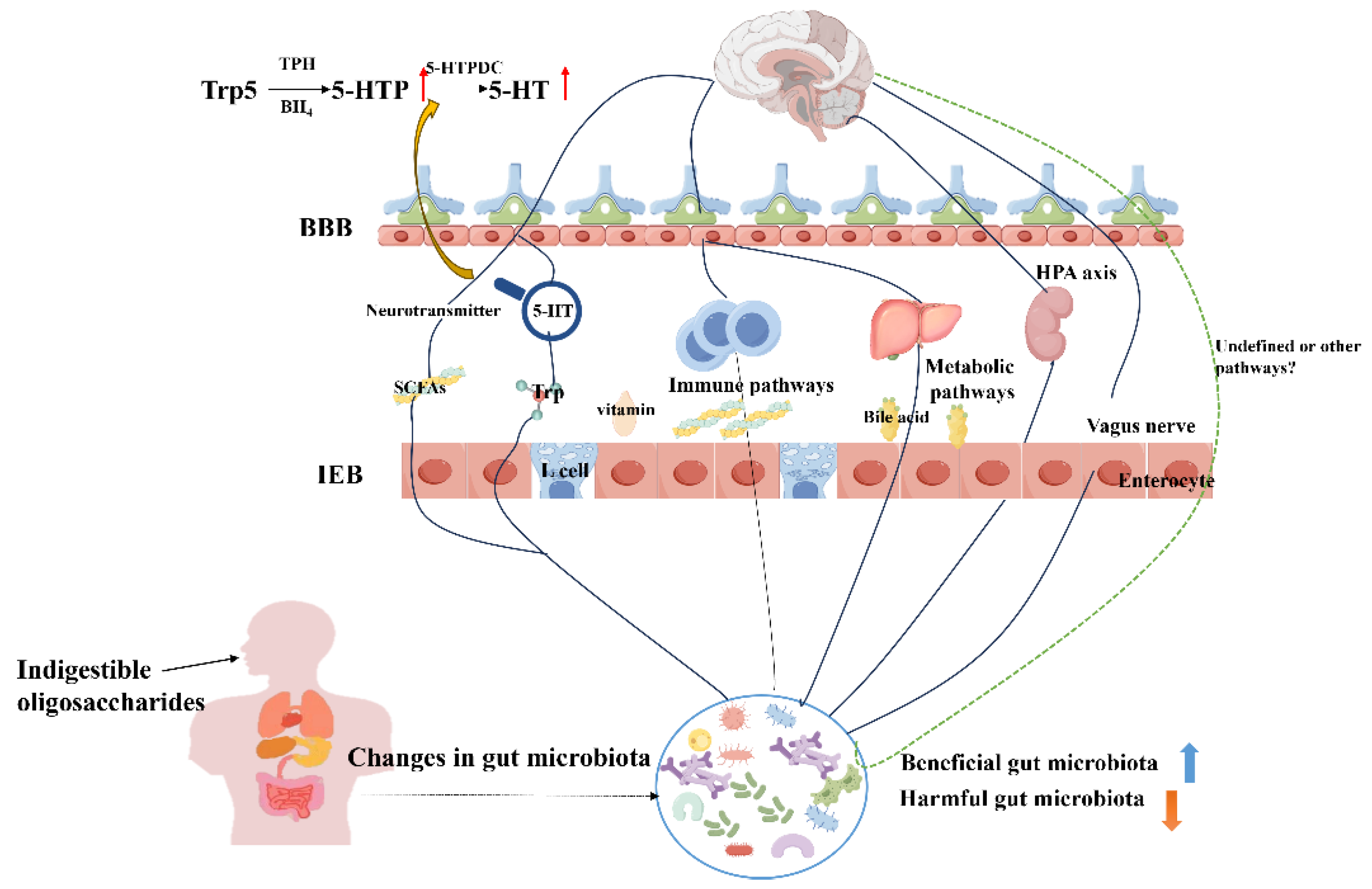
Figure 3.
NODs alleviate IBD progression by regulating the gut microbiota. 1) NODs promote the proliferation of beneficial bacteria and corrects intestinal dysbiosis in IBD patients. 2) SCFAs can improve the barrier function and reduce the permeability of intestinal barrier by affecting the maturation of intestinal epithelial cells (IECs) and regulating the expression of tight junction proteins (Claudin1, Occludin, ZO-1). SCFAs also affect the interaction between immune cells (T cells, B cells), regulate the expression of pro-inflammatory/anti-inflammatory cytokines, and reduce intestinal inflammation. 3) Microbial metabolites of Trp regulate the expression of adhesion junction proteins (E-cadherin, β-catenin) and maintain the integrity of the intestinal barrier.
Figure 3.
NODs alleviate IBD progression by regulating the gut microbiota. 1) NODs promote the proliferation of beneficial bacteria and corrects intestinal dysbiosis in IBD patients. 2) SCFAs can improve the barrier function and reduce the permeability of intestinal barrier by affecting the maturation of intestinal epithelial cells (IECs) and regulating the expression of tight junction proteins (Claudin1, Occludin, ZO-1). SCFAs also affect the interaction between immune cells (T cells, B cells), regulate the expression of pro-inflammatory/anti-inflammatory cytokines, and reduce intestinal inflammation. 3) Microbial metabolites of Trp regulate the expression of adhesion junction proteins (E-cadherin, β-catenin) and maintain the integrity of the intestinal barrier.
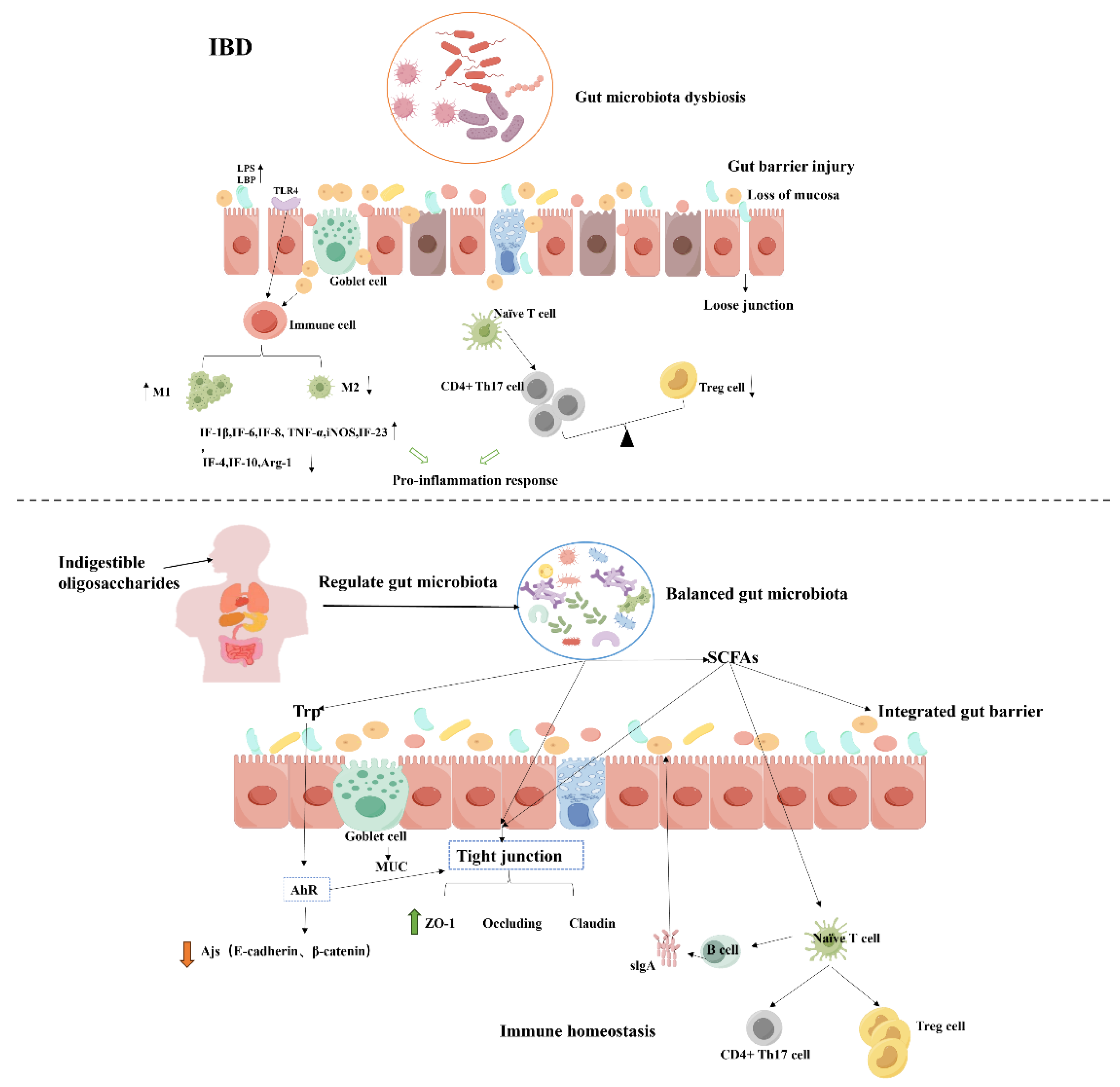
Figure 4.
Schematic diagram of the mechanism of action of NODs in relieving constipation.
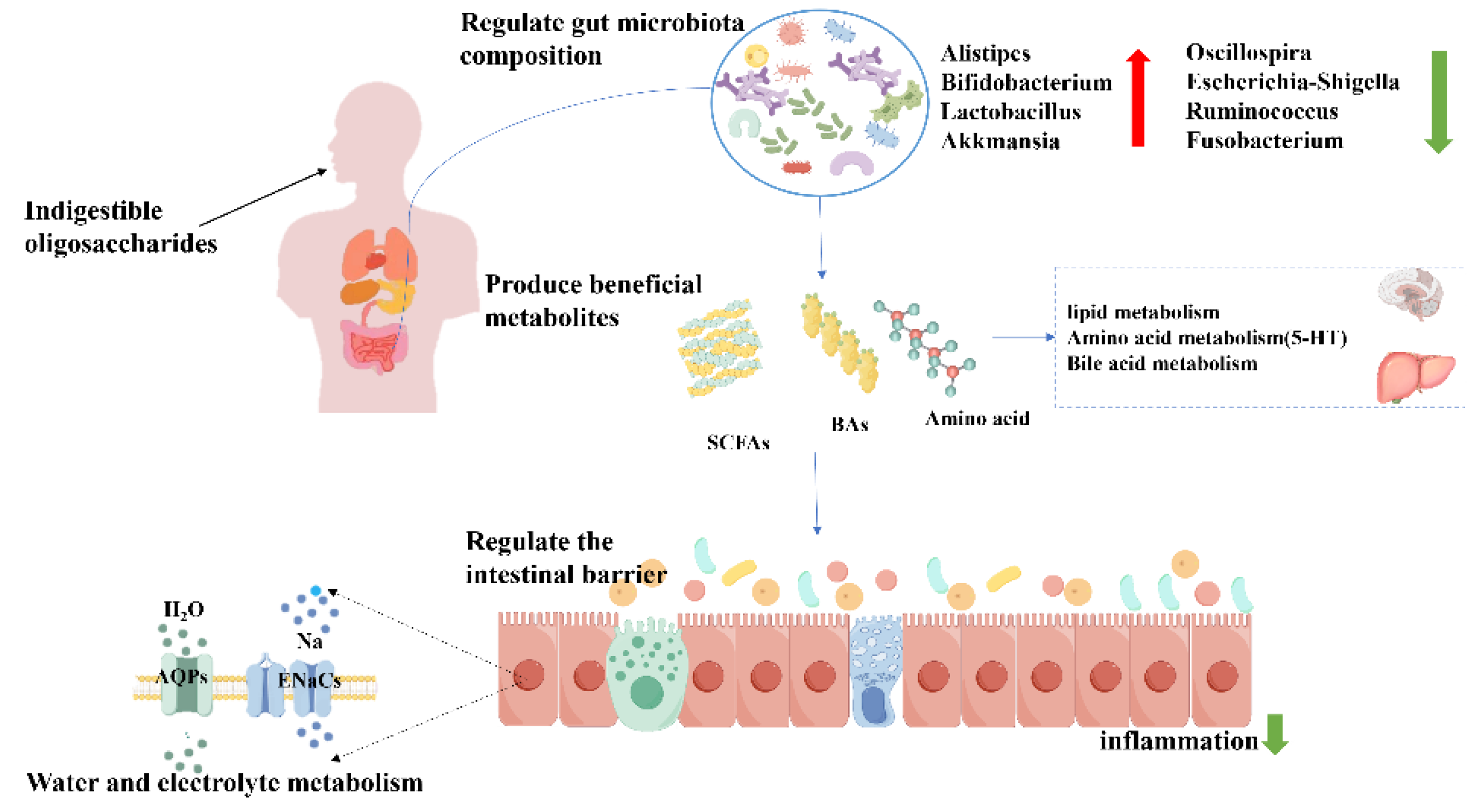

Table 1.
Basic information of the main nondigestible oligosaccharides (NDOs).
| Name | Glucosidic linkage |
Monosaccharides | DP | Preparation | Natural source |
Reference |
| FOS | β-1,2 |
Fructose Glucose |
2-5 | Enzyme degradation、 Microbial Fermentation |
Onions, chicory, Wheat |
[55] [56] |
| IMO | α-1,3 α-1,4 α-1,6 |
Glucose | 2-5 | Enzyme synthesis、 Enzymatic hydrolysis、 Enzymatic conversion |
Starch, Corn milk, Wheat bran |
[57] [58] |
| XOS | β-1,4 | Xylose | 2-7 | Enzymatic conversion | hoots, corncob, wheat straw |
[59] [60] |
| MOS | α-1,2 α-1,6 |
Mannose Glucose |
2-10 | Enzymatic hydrolysis | Carrageenan Konjaku Flour |
[61] [57] |
| GOS | β-1,4 β-1,6 |
Galactose Glucose |
3-6 | Enzymatic conversion | Lactose | [55] [62] |
|
SOS |
α-1,2 α-1,6 |
Galactose, Glucose Fructose |
2-5 |
Extracted |
Soybean |
[57] |
| COS | β-1,4 | D-glucosamine | — |
Enzymatic hydrolysis | Shell of crustaceans | [63] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



