
Submitted:
31 May 2024
Posted:
04 June 2024
You are already at the latest version
Abstract
Depression is the most common chronic mental illness and is characterized by low mood, insomnia, and affective disorders. However, the pathological mechanisms of depression remain unclear. Numerous studies have suggested that the ghrelin/GHSR system may be involved in the pathophysiologic process of depression. Ghrelin plays a dual role in the experimental animals, increasing depressed behavior and decreasing anxiety. By combining several neuropeptides and traditional neurotransmitter systems to construct neural networks, it modifies signals connected to depression. The present review emphasizes the role of ghrelin in neuritogenesis, astrocyte protection, inflammatory factor production, and endocrine disruption in depression. Furthermore, ghrelin/GHSR can activate multiple signaling pathways to produce antidepressant effects, including cAMP/CREB/BDNF, PI3K/Akt, Jak2/STAT3, and p38-MAPK. Given the antidepressant effects of ghrelin/GHSR1a system, it is expected to become a potential therapeutic target for the treatment of depression.
Keywords:
GPCR
; Ghrelin
; GHSR
; Cell signaling
; Depression
1. Introduction
Growth hormone-releasing peptide (ghrelin) is a multifunctional 28-amino acid peptide hormone, which was originally discovered in rat stomach but has also been found in a variety of other organs including bowels, hypothalamus, pituitary, and hippocampus[1]. Ghrelin's serine 3 (Ser3) is n-octastylated, a modification that allows for the cycling of both the acylated and des-acylated forms of ghrelin[2]. Acylated ghrelin is able to cross the blood-brain barrier (BBB) and binds to the central growth hormone-releasing peptide receptor (GHSR), which are widely found in the central nervous system (CNS) and peripheral tissues and has two isoforms, GHSR1a and GHSR1b[3]. Acylated ghrelin mainly binds to GHSR1a and exerts a variety of physiological and behavioral modulatory effects, such as regulation of glucose homeostasis, regulation of metabolism and energy homeostasis, modulation of blood pressure, and renal protection[4,5,6]. GHSR1b contains 289 amino acids and is a splice variant and dominantly inactivated form of GHSR1a[7]. Due to the absence of the 6th and 7th transmembrane chains, it is unable to bind ghrelin and does not have signaling ability. There is much evidence that GHSR1b does not bind ghrelin, but it can heterodimerize with GHSR1a to interfere with GHSR1a function[8]. In an experiment to study, the expression of two GHSRs cloned from black seabream in HEK293 cells, GHSR1b was found to inhibit the signaling activity of GHSR1a-mediated elevation of intracellular Ca2+ concentration[9]. GHSR1a has also been shown to form mixed heterodimers with dopamine receptor subtype 2 (D2R)[10], serotonin 2c receptors (5-HT2cR)[11], orexin 1 receptor (OX1R)[12], and the dimers can fine-tune the activity of the GHSR1a receptor.
Depression is a prevalent severe chronic mental illness, which is characterized by appetite disorders, weight changes, bipolar disorder and insomnia[13,14,15]. The above mentioned disorders may be attributed to decreased size and function of the hippocampus and amygdala in the limbic brain regions[16]. With the rise in survival pressures in contemporary society and the dramatic increase in prevalence and incidence in different parts of the country, depression is emerging as a complex disorder present in all stages of the population, including the pregnant population, the adolescent population, and the elderly population[17,18]. Although the exact pathogenesis of the disorder remains obscure, a large body of data strongly suggests that stress, and genetic predisposition are collectively involved in the progression of the disorder [19]. The pathophysiologic processes of depression include inflammation in the brain, reduced neurogenesis, monoamine neurotransmitter changes, and endocrine abnormalities[20,21,22]. However, the interrelationships between these processes remain unclear. Currently, several classes of drugs are available for the treatment of major depressive disorder (MDD) [23], and most of them exert their antidepressant effects by enhancing monoaminergic function, such as typical monoamine oxidase inhibitors (MAOIs) and selective 5-hydroxytryptamine reuptake inhibitors (SSRIs)[24]. In addition, scientists are now working on research beyond typical monoamine targets and pathways to develop antidepressants with novel mechanisms of action[25].
However, the efficacy and severe side effects of these antidepressants remain major issues to be addressed[26], and there are no therapeutic candidates shown to completely eliminate disease progression. In line with ghrelin’s effects in the CNS, there is increasing evidence for an important role of ghrelin in depression[27,28,29]. The majority of research has elucidated the anti-depressive mechanisms of ghrelin. Conversely, few studies reported a detrimental effect as ghrelin itself induces depressive tendencies and patterns in animal models. Here, we have attempted to elucidate the aforementioned link between ghrelin and the onset as well as the treatment of depression, and to elucidate the mechanisms and pathways that may be involved in the development of ghrelin in depression. We made such an explanatory work by conducting a comprehensive review of the existing literature (reviews and research articles) through keyword searches.
2. Mechanisms of Ghrelin/GHSR System in Depressive Disorder
Numerous studies have shown that ghrelin/GHSR system inhibits key pathways and mechanisms in the development of depression that have been found to correlate with certain factors, transmitters, and cells in the body in a variety of models of depression, which we will describe in more detail in the following paragraphs.
2.1. Links between Monoamine Neurotransmitter Receptors and Ghrelin
The monoamine hypothesis, long considered to be the most common hypothesis of depression, is centred on the fact that the concentration of monoamines (5-hydroxytryptamine, norepinephrine, and dopamine) in the synaptic gap is reduced in depressive states[30]. The expression of these monoamine neurotransmitters may be involved in the neuropathological process of ghrelin. It was found that in neurons co-expressing D1R and GHSR, ghrelin can amplify dopamine signalling by activating GHSR to enhance its mediated downstream pathways[31]. And reduced mRNA expression of dopaminergic receptors in the amygdala and the dorsal nucleus of the middle suture was found in studies of GHSR1a-/- mice[32], suggesting a potential regulation of dopamine neuron production by the ghrelin/GHSR system. In addition to this, another study showed that the central 5-hydroxytryptamine system is a target of ghrelin[32], and increased mRNA expression of some 5-hydroxytryptamine receptors in the amygdala and the dorsal nucleus of the intermediate suture was found in mice acutely administered ghrelin centrally. Conversely, unlike the ghrelin/GHSR system described above, which regulates the expression of monoamine neurotransmitters, norepinephrine has been shown to stimulate ghrelin secretion in mouse cells[33], and depletion of catecholamine-secreting neurons can also inhibit fasting-induced ghrelin secretion[34]. In conclusion. There are many unknown connections between the monoamine system and the ghrelin/GHSR system, and more research is needed to determine whether these monoamine receptors can be used as targets in the treatment of depression.
2.2. The Ghrelin/GHSR System Mediates the Inflammatory Response to Depression
In the CNS, IL-1β stimulates microglia and astrocytes to produce other cytokines, such as IL-6 and TNF-α, to promote inflammation in the brain[35]. Furthermore, pro-inflammatory cytokines such as IL-1β and TNF-α have been reported to play an important role in the onset of depression[36], and an increase in pro-inflammatory cytokines contributes to the development of depression[20]. And IL-6 is the most persistently elevated cytokine in the blood of MDD patients, so it may serve as a predictive biomarker and a potential target for the treatment of depression in humans[37]. These experimental conclusions show that the role of pro-inflammatory cytokines of the CNS in the pathogenesis of depression cannot be ignored.
Serum ghrelin concentrations have been reported to increase in inflammatory bowel disease (IBD) and are associated with the degree of activity[38]. Similar reports have been made in animal model experiments: Serum ghrelin concentrations were higher in rats with severe pancreatitis[39]and arthritis[35], as well as acute colitis[40]. It can be seen that serum ghrelin concentration is positively correlated with inflammation. Nonetheless, an experiment is at odds with the conclusion just mentioned, it is believed that rheumatoid arthritis patients and arthritis rats have lower levels of serum ghrelin concentrations[41]. This discrepancy may be due to unpredictable reasons such as rat nutritional status or experimental conditions, making it difficult to analyze the effect of inflammation on serum ghrelin levels. However, it is undeniable that ghrelin levels are elevated in inflammation as demonstrated in most experiments. On the whole, the causal relationship between inflammation and changes in serum ghrelin concentrations requires further study, although the effects of various conditions cannot be ruled out. Pursuant to the existing data, there are many reports that elevated serum ghrelin concentrations are the result of inflammation. Interestingly, experiments have found that elevated ghrelin is involved in regulating inflammatory response, downregulating neutrophil transport and the number of pro-inflammatory cytokines, significantly reducing cerebral ischemic injury and improving neurobehavioral function[42]. The same study also reported that exogenous ghrelin inhibits endothelial cell production of IL-1, IL-6, and IL-8 by regulating the release of pro-inflammatory cytokines, which play an important role in the pathological process of depression[43]. In addition, GHSR and ghrelin are expressed in human T lymphocytes and monocytes, and ghrelin specifically inhibits the expression of pro-inflammatory cytokines via GHSR[44]. Also, two other investigations displayed decreased expression of pro-inflammatory cytokines such as TNF-α, IL-1β, and IL6 in rats with GHSR gene knockouts[45,46]. Thus, there is increasing evidence of the mediating role of the Ghrelin/GHSR system in regulating pro-inflammatory cytokine release events in depressive disorders.
On the other hand, we can note that inflammation reduces neuroplasticity by downregulating brain-derived neurotrophic factor (BDNF), which may be the basis of the pathophysiology of depression[47,48]. IL-6 has been shown to be a reliable positive predictor of BDNF in patients with melancholic MDD[49]. However, the correlation between inflammation and BDNF requires further study. And the Ghelin/GHSR system can also produce antidepressant effects through the regulation of BDNF, which we will elaborate below.
2.3. The Ghrelin/GHSR System Promotes Neurogenesis in Depression
Numerous clinical studies have shown that depression is closely associated with decreased size and function of the hippocampus and amygdala in limbic brain regions, and that patients with depression have reduced hippocampal volume and decreased neurogenesis[50,51]. Where neurogenesis is closely related to the treatment of depression. Studies have found that ablation of hippocampal nerve genesis in mice impairs the efficacy of antidepressants[21]. In animal experiments, antidepressant drugs promote hippocampal neurogenesis. Similarly, alterations in adult hippocampal neurogenesis mediate the effects of antidepressants, and chronic administration of antidepressants enhances adult hippocampal neurogenesis [52].
The neuroprotective effects of ghrelin as an neuropeptide have been widely demonstrated. In rat cortical neuronal damage induced by hypoxia and hypoglycaemia[53], ghrelin inhibited the neuronal damage process and the protective effect disappeared after administration of a GHSR-specific inhibitor. In addition to this, a reduction in oligodendrocyte and neuronal apoptosis was found in an experiment in which ghrelin was administered after spinal cord injury in rats[54]. Most notably, the antidepressant effects exerted by ghrelin in vivo can be dependent on its neuroprotective effects. Published literature has displayed that ghrelin directly increases hippocampal neurogenesis in the treatment of depression[55]. Further studies have identified hippocampal neuroprotection as the primary mechanism by which stress-induced elevation of ghrelin protects the organism from stress-induced worsening of associated depression[56]. In addition to the protective effects of ghrelin on hippocampal neurons against depression, activation of catecholaminergic neurons has also been identified as a possible mechanism for contributing to the antidepressant effects of ghrelin[57]. Although numerous studies have demonstrated that ghrelin exerts its antidepressant effects through neuroprotection, the mechanisms involved are currently not well understood. It has been found that in the hippocampus, ghrelin is able to cross the BBB and bind to GHSR1a to improve cognitive function and enhance hippocampal neurogenesis[58], and to enhance LTP[59]. Ghrelin also directly induces proliferation and differentiation of adult neural progenitor cells in the hippocampal subgranular zone, and in the case of ghrelin receptor knockout mice (GHSR1a-/-) systemic administration, it was found that the number of progenitor cells was reduced compared to wild-type controls[55]. In addition to this, another report concluded that ghrelin provides neuroprotection through activation of AMPK and enhances clearance of damaged mitochondria[60], and it has also been demonstrated that ghrelin mediates neuroprotection through inhibition of glial cell activation and release of pro-inflammatory mediators[61]. In addition to the above mentioned related mechanisms, it has been experimentally confirmed that BDNF plays an important role in the pathophysiology of depression[62], and ghrelin exerts an antidepressant effect by regulating the relevant expression of BDNF is also a hot topic of research.
2.4. Regulation of Astrocyte Physiology by the Ghrelin/GHSR System
The above-mentioned neuronal physiology is closely related to the onset of depression. In addition to this, as non-neurons, the significant reduction in density and massive damage of astrocytes after the onset of depression has been confirmed by a number of experiments[63,64,65]. A study elucidated that astrocytes show dysfunction and a significant reduction in number in the medial prefrontal cortex (mPFC) region in depressed patients[66]. In addition, significant glial cell pathology is present in patients with MDD and cells with chronic unpredictable mild stress (CUMS) can be reversed by antidepressant pharmacological modulation that promotes glial cell Glu clearance[67]. Another investigations have detected that long-term administration of the antidepressant fluoxetine reversed the stress-induced decline in the number of hippocampal glial cells in tree shrews, and the relevance of structural plasticity of astrocytes in stress and therapeutic support with antidepressants has been proposed[65]. These all suggest that antidepressant-mediated changes in astrocytes may be key to their effects. Taken together, these suggest that astrocytes may play an active role in brain function and that there is some connection to the process of depression.
Traditionally, astrocytes have often been thought of as brain glue, a class of cells that only provide metabolic and functional support to neurons. However, with the discovery of various neurotransmitter receptors and channels on the astrocyte membrane, our understanding of the function of astrocytes in the nervous system has fundamentally changed[68]. A large number of studies have found that the most prominent neurotransmitter receptors expressed on astrocyte membranes are the family of GPCRs, including metabotropic glutamate receptors, adrenergic receptors, GABAergic receptors, cholinergic receptors, histaminic receptors, dopaminergic receptors, and neurotrophic factor receptors[69,70,71,72,73,74]. Upon activation of these receptors by neurotransmitters released from presynaptic neurons, astrocytes can undergo "gliotransduction"[75], releasing gliotransmitters to feedback on neuronal excitability and synaptic transmission[76,77]. Thus, astrocytes are now considered to be active participants in neuronal communication.
GHSR1a has been shown to be expressed in astrocytes in the arcuate nucleus of the hypothalamus[78]and in the dentate gyrus in the hippocampus[79], and that astrocytes respond to ghrelin and its analogues. Thus, we can make the hypothesis that glial cells may act as mediators of ghrelin activity in the brain. It is interesting to see whether ghrelin, as an important mediator in the development of depression, also plays a role in depression in some way linked to astrocytes. A study found that the astrocytoma cell line C6 could respond to GHRP-6 by upregulating GHSR1a levels, increasing activation of the PI3K/Akt pathway and increasing its own proliferation[80]. And this effect can be inhibited by D-Lys3-GHRP-6, an antagonist of GHSR1a[79], so ghrelin may exert neuroprotective effects by stimulating astrocyte proliferation through GHSR1a to increase the expression of the PI3K/Akt pathway. However, another study found that ghrelin did not show a promotive effect on astrocyte proliferation, and it found that ghrelin reversed the activation and accumulation of astrocytes in hippocampal neurodegeneration following hippocampal excitotoxicity injury induced by sea manate[61]. This difference may be related to the activation of astrocytes and the increase in the concentrated release of pro-inflammatory cytokines in inflammatory and related conditions and the triggering of neuroinflammation[81]. In conclusion, the specific mechanisms associated with ghrelin and astrocytes need to be investigated more thoroughly. ghrelin may have a neuroprotective role in the development of depression through the regulation of astrocyte physiological activity by the receptor GHSR1a.
2.5. The Role of the Ghrelin/GHSR System in Endocrine Disruption in Depression
Depression has long been recognised as having a correlation with endocrine disruption, with overactivity of the hypothalamic-pituitary-adrenal (HPA) axis being the most common[82]. In addition, the hypothalamic-pituitary-thyroid (HPT) axis and the hypothalamic-pituitary-gonadal (HPG) axis are also disturbed in depressed patients [83,84]. Therefore, changes in the levels of related hormones in the body can provide ideas for research on the treatment of depression. For example, testosterone levels are reduced in men suffering from depression[85], and the vulnerability of perimenopausal women to depression is associated with changes in oestrogen[86]. Then we can consider testosterone and oestrogen as targets for further research on antidepressants.
There is growing evidence that ghrelin is involved in the regulation of endocrine disruption in depression. Ghrelin has been reported to inhibit the secretion of luteinising hormone (LH) and thyroid stimulating hormone (TSH) in patients with major depression[87]. Ghrelin also affects the activity of the HPT axis by decreasing TSH and increasing free thyroxine in plasma[88]. In addition, the β1-adrenergic receptor blocker atenolol exacerbates depressive-like behaviours in chronic social defeat stress (CSDS) mice by attenuating the elevation of plasma acylgrowth factor-releasing peptide[89]. In conclusion, the relevant role of ghrelin in endocrine disruption in depressed patients requires more research.
3. Signalling Pathways Induced by Ghrelin/GHSR1a System in Depression
3.1. cAMP-CREB-BDNF Signaling Pathway
Neurodegenerative and neuropsychiatric disorders can be caused by an inadequate supply of neurotrophic factors[90]. Among them, BDNF, as a member of the neurotrophic protein family, plays an extremely important role in promoting neuronal growth, survival, differentiation and in synaptic transmission and enhancing central plasticity[91,92,93], and slows down depressive progression. Relatedly, ghrelin increases total BDNF mRNA expression in the mouse hippocampus and synthesises different kinds of BDNF mRNAs by acting on different promoters in rats of different ages[94]. Different BDNF transcripts exhibit different subcellular localisations that selectively shape the proximal and distal compartments of the cytosol or dendrites[95]and play an important role in increasing neuronal plasticity[96].
Impaired cAMP signalling occurs in patients with major depression[97]. In addition, in the hippocampus and prefrontal cortex of patients, the levels of BDNF, CREB and p-CREB are significantly reduced[98,99,100,101]and the levels of BDNF mRNA in peripheral monocytes of this population are also significantly reduced[102]. Interestingly, administration of ghrelin through the centre normalised hippocampal BDNF levels[103]. In addition, exogenous ghrelin can improve depressive behaviour by upregulating CREB signaling through activation of ghrelin receptors and cAMP/PKA signaling pathway, and increasing BDNF expression downstream[27]. Thus, ghrelin-induced increases in BDNF in the hippocampus involve activation of the GHSR1a/cAMP/PKA/CREB signalling pathway (Figure 1).
3.2. p38-MAPK Signaling Pathway
Several studies have demonstrated that p38-MAPK is activated in response to various stressful stimuli and is involved in the pathological process of depression [104,105]. p38-MAPK can be activated by interferon and lipopolysaccharide to up-regulate the expression of the depression-related gene IDO[106,107], and it can also exacerbate oesophageal cancer-associated depression by directly enhancing the expression of the IDO gene[108]. Therefore, the expression status of p38-MAPK pathway-related substances could be a powerful tool for depression monitoring, while p38-MAPK itself could be a target for antidepressant research. In addition, activation of the p38-MAPK pathway phosphorylates the glucocorticoid receptor (GR), and GR phosphorylation is associated with reduced glucocorticoid sensitivity[109], which may be closely related to glucocorticoid resistance exhibited by depressed patients[110]. In contrast, ghrelin treatment of rats activates GHSR1a and decreases p38-MAPK phosphorylation, which in turn increases GR levels[111]. Furthermore, no significant increase in phosphorylation of p38 by CSDS in vector-treated mice was observed after ghrelin treatment. Additionally, hippocampal GHSR-deficient mice showed higher levels of p38 phosphorylation than control mice, suggesting that ghrelin may also mediate antidepressant mechanisms by inhibiting the p38-MAPK signalling pathway in the hippocampus[112]. Interestingly, social failure stress produces depressive-like behaviour in wild-type mice, but selective deletion of p38-MAPK in serotonergic neurons of the nucleus dorsalis of the mouse middle suture protects the mice under stress induction[113]. This suggests that p38-MAPK has the ability to specifically regulate selected downstream targets, and the role played by this pathway in antidepressant disorders requires further investigation.
3.3. PI3K/Akt Signaling Pathway
Depression is closely related to neurogenic hypoplasia[114], and PI3K/Akt is thought to be an important signal for the proliferation of adult hippocampal progenitor cells [115]. Akt can exert its utility in controlling cellular proliferation by activating the phosphorylation of its downstream targets (GSK-3β, mTOR, and p70S6K), where GSK-3β is a pro-apoptotic protein whose activity plays an important role in neuropathology and psychiatric disorders [116]. β-catenin, as a transcription factor regulated by GSK-3β, undergoes nuclear translocation under conditions of GSK-3β inactivation [117], which is an indispensable step in its role in promoting cell survival. In addition, as downstream of PI3K/Akt, phosphorylation of mTOR and p70S6K also promotes the proliferation of neural stem cells [118]. Interestingly, ghrelin could induce hippocampal neural stem cell (NSC) proliferation by activating the PI3K/Akt signaling pathway by binding to GHSR1a, and the stimulatory effects of ghrelin on GSK-3β, mTOR and p70S6K phosphorylation were significantly inhibited by the treatment of GHSR1a-specific antagonist D-Lys-3-GHRP-6[119]. Furthermore, it has been demonstrated that ghrelin enhances the nuclear translocation of β-catenin, which in turn contributes to its anti-apoptotic effects[120]. Therefore, we may hypothesize that ghrelin may promote neuronal cell proliferation through activation of the PI3K/Akt pathway and subsequently play a role in the treatment of depression. Moreover, autophagy plays an important role in maintaining neuronal stem cells and adult neuronal plasticity, while ghrelin can stimulate autophagy by inhibiting PI3K/AKT/MTOR signaling pathway [121]. However, an experiment in a mouse model of corticosterone-induced depression found that overactive neuronal autophagy depleted BDNF and impaired adult hippocampal neurogenesis[92]. Therefore, it remains to be investigated whether ghrelin can regulate autophagy homeostasis in vivo through the PI3K/AKT/MTOR pathway and promote neurogenesis.
3.4. Jak2-STAT3 Signaling Pathway
The Jak2/STAT3 signaling pathway, like the aforementioned PI3K/Akt signaling pathway, has also been shown to play an important role in neuroprotection. Unlike single pathways that act independently, one study found that resveratrol may work together to exert neuronal protection by indirectly upregulating the PI3K/Akt/mTOR pathway through the activation of Jak2/STAT3 [122]. As shown in Figure 2, exposure of rat hippocampal NSCs to the Jak2/STAT3 inhibitor cucurbitacin I significantly blocked the proliferative effects of ghrelin on NSCs [119]. Thus, ghrelin would also be potent through activation of the Jak2/STAT3 pathway. Furthermore, this pathway not only plays an active role in neuroprotection and regeneration, its role in neuroinflammation has also been shown to be promoted. It has been found that inhibition of the Jak2/STAT3 pathway ameliorates neuroinflammation[123] and reduces neuronal senescence by suppressing the inflammatory response[124]. Since depression is closely related to decreased neuronal genesis and upregulation of inflammatory factors, and activation of the Jak2/STAT3 pathway was found to have completely opposite effects in these two aspects, whether this pathway can be a therapeutic target for depression needs to be investigated at a deeper level on the multifaceted mechanisms. The study of whether ghrelin regulates neuroinflammation when activating the Jak2/STAT3 pathway will be an important basis for comprehensively determining whether ghrelin can be used as a target for the treatment of depression.
4. Ghrelin/GHSR as a Therapeutic Target for Depressive Disorder
Many neuropeptides have been reported as targets for depression treatment. For example, nonselective glycopeptide receptor agonists (galnon) can exert antidepressant activity in preclinical models of depression[125,126]. VP antagonists have shown similar antidepressant behavior in preclinical studies[127,128]. Similarly, many studies have shown that ghrelin can be used as a powerful tool in the treatment of depression. It has been reported that ghrelin produces antidepressant effects in estrogen-deficient mice[103], and it may also counteract depressive symptoms caused by chronic stress[129]. In addition, many substances that exert antidepressant effects by modulating ghrelin/GHSR expression have been identified. For example, paeoniflorin (PF) significantly increased the expression of GHSR1a to mediate antidepressant effects[130]. The GHSR inhibitor JMV29282259 blocked saffronin-induced expression of neuroplasticity-related proteins[131]. Here, we briefly summarized the correlation between ghrelin/GHSR system and depression, as well as the research methodology, as shown in Table 1. Thus, ghrelin/GHSR has many potent functions in defense against depression-like symptoms. However, further studies are needed because of the unstable antidepressant behavioral effects of neuropeptides expressed in various tests and the two-sided nature of ghrelin's effects on depression in different models[132].
5. Conclusions and Future Direction
There is still ongoing debate about the role of ghrelin in depression. The majority of studies suggest that ghrelin has antidepressant effects, with few studies indicating depressogenic effects. Indeed, ghrelin/GHSR can exert antidepressant and neuroprotective effects by triggering multiple signaling pathways including cAMP/CREB/BDNF, PI3K/Akt, Jak2/STAT3, p38-MAPK et al. Moreover, GHSR1a can also form dimers with other GPCRs to exert antidepressant effects. So, ghrelin/GHSR system is becoming a new target for the treatment of depression. However, due to the two-sided claims of ghrelin's antidepressant potency from different experiments, ghrelin had no antidepressant effect on young rats, and the drug's neuropharmacology differed in adolescents and adults. Future experiments should therefore focus on coming to investigate the link between the pathogenesis of ghrelin in different depressive groups and the development of receptors biased drugs is also a good strategy. Moreover, more studies are necessary to determine the extent to which central and peripheral ghrelin signaling are functionally interconnected, as this understanding is crucial for the development of potential new ghrelin-based therapeutic agents. Finally, promising findings from animal studies necessitate further human-based research to ascertain the extent to which such results can be applied to human disorders.
Author Contributions
Writing-original draft preparation, Xingli Pan and Yuxin Gao; writing-review and editing, Jing Chen and Bingyuan Ji; supervision, Kaifu Guan. All authors have read and agreed to the published version of the manuscript.
Disclosure Statement
The authors declare that they have no conflicts of interest with the contents of this article.
Acknowledgments
This work was supported by the Natural Science Foundation of Shandong Province (ZR2020MH148) and the University Student Innovative Training Program of Jining Medical University (cx2023072z) .
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
BBB, blood-brain barrier; BDNF, brain-derived neurotrophic factor; CAO, coronary artery occlusion; CDK, cyclin-dependent kinases; CNS, central nervous system; CREB, cAMP response element-binding protein; CSDS, chronic social defeat stress ; CUMS, chronic unpredictable mild stress; DDR, disturbed diurnal rhythm; EPM, Elevated plus maze; FST, forced swimming test; GFAP, glial fibrillary acidic protein; GHSR, growth hormone-releasing peptide receptor; GSK-3β, Glycogen synthase kinase 3β; GR, glucocorticoid receptor; HFD, high-fat diet; HPA, hypothalamic-pituitary-adrenal; HPT, hypothalamic-pituitary-thyroid; HPG, hypothalamic-pituitary-gonadal; Iba-1, ionized calcium binding adapter molecule 1; LH, luteinising hormone; MDD, major depressive disorder; mPFC, medial prefrontal cortex; mTOR, mammalian target of rapamycin; NSCs, neural stem cells; OFT, open field test; OX1R, orexin 1 receptor; OB, Olfactory bulbectomy surgery; p70S6K, p70 ribosomal protein S6 kinase; RS, restraint stress; SIT, social interaction test; SPT, sucrose preference test; TST, tail suspension test.
References
- A.D. Howard, S.D. Feighner, D.F. Cully, J.P. Arena, P.A. Liberator, C.I. Rosenblum, M. Hamelin, D.L. Hreniuk, O.C. Palyha, J. Anderson, P.S. Paress, C. Diaz, M. Chou, K.K. Liu, K.K. McKee, S.S. Pong, L.Y. Chaung, A. Elbrecht, M. Dashkevicz, R. Heavens, M. Rigby, D.J. Sirinathsinghji, D.C. Dean, D.G. Melillo, A.A. Patchett, R. Nargund, P.R. Griffin, J.A. DeMartino, S.K. Gupta, J.M. Schaeffer, R.G. Smith, L.H. Van der Ploeg, A receptor in pituitary and hypothalamus that functions in growth hormone release, Science, 273 (1996) 974-977. [CrossRef]
- Kojima, M.; Kangawa, K. Ghrelin: Structure and Function. Physiological Reviews 2005, 85, 495-522. [CrossRef]
- Albarrán-Zeckler, R.G.; Smith, R.G. The Ghrelin Receptors (GHS-R1a and GHS-R1b). In The Ghrelin System, 2013; pp. 5-15. [CrossRef]
- Sun, Y.; Asnicar, M.; Smith, R.G. Central and Peripheral Roles of Ghrelin on Glucose Homeostasis. Neuroendocrinology 2007, 86, 215-228. [CrossRef]
- Gortan Cappellari, G.; Barazzoni, R. Ghrelin forms in the modulation of energy balance and metabolism. Eating and Weight Disorders - Studies on Anorexia, Bulimia and Obesity 2018, 24, 997-1013. [CrossRef]
- Vinci, M.C.; Fujimura, K.; Wakino, S.; Minakuchi, H.; Hasegawa, K.; Hosoya, K.; Komatsu, M.; Kaneko, Y.; Shinozuka, K.; Washida, N.; et al. Ghrelin Protects against Renal Damages Induced by Angiotensin-II via an Antioxidative Stress Mechanism in Mice. PLoS ONE 2014, 9. [CrossRef]
- Laviano, A.; Molfino, A.; Rianda, S.; Rossi Fanelli, F. The growth hormone secretagogue receptor (Ghs-R). Curr Pharm Des 2012, 18, 4749-4754. [CrossRef]
- Leung, P.-K.; Chow, K.B.S.; Lau, P.-N.; Chu, K.-M.; Chan, C.-B.; Cheng, C.H.K.; Wise, H. The truncated ghrelin receptor polypeptide (GHS-R1b) acts as a dominant-negative mutant of the ghrelin receptor. Cellular Signalling 2007, 19, 1011-1022. [CrossRef]
- Chan, C.-B.; Leung, P.-K.; Wise, H.; Cheng, C.H.K. Signal transduction mechanism of the seabream growth hormone secretagogue receptor. FEBS Letters 2004, 577, 147-153. [CrossRef]
- Ting-Ting, T.; Ming-Xia, B.; Mei-Ning, D.; Xiao-Yi, Z.; Ling, C.; Xue, X.; Qian, J.; Xi, C.; Chun-Ling, Y.; Xi-Xun, D.; et al. Quinpirole ameliorates nigral dopaminergic neuron damage in Parkinson's disease mouse model through activating GHS-R1a/D(2)R heterodimers. Acta Pharmacol Sin 2023, 44. [CrossRef]
- Xiaoli, L.; Xia, L.; Xinyou, Z.; Huaiyu, Y.; Lijun, S.; Minmin, H.; Xiaoqi, C.; Mingxuan, Z.; Katrina, W.-G.; Tiantian, J.; et al. Olanzapine attenuates 5-HT2cR and GHSR1a interaction to increase orexigenic hypothalamic NPY: Implications for neuronal molecular mechanism of metabolic side effects of antipsychotics. Behav Brain Res 2024, 463. [CrossRef]
- Xue, Q.; Bai, B.; Ji, B.; Chen, X.; Wang, C.; Wang, P.; Yang, C.; Zhang, R.; Jiang, Y.; Pan, Y.; et al. Ghrelin Through GHSR1a and OX1R Heterodimers Reveals a Gαs-cAMP-cAMP Response Element Binding Protein Signaling Pathway in Vitro. Front Mol Neurosci 2018, 11, 245. [CrossRef]
- Baxter, L.C. Appetite Changes in Depression. American Journal of Psychiatry 2016, 173, 317-318. [CrossRef]
- Crouse, J.J.; Carpenter, J.S.; Song, Y.J.C.; Hockey, S.J.; Naismith, S.L.; Grunstein, R.R.; Scott, E.M.; Merikangas, K.R.; Scott, J.; Hickie, I.B. Circadian rhythm sleep–wake disturbances and depression in young people: implications for prevention and early intervention. The Lancet Psychiatry 2021, 8, 813-823. [CrossRef]
- Rakel, R.E. Depression. Prim Care 1999, 26, 211-224. [CrossRef]
- Duman, R.S.; Voleti, B. Signaling pathways underlying the pathophysiology and treatment of depression: novel mechanisms for rapid-acting agents. Trends in Neurosciences 2012, 35, 47-56. [CrossRef]
- Hauenstein, E.J. Depression in adolescence. J Obstet Gynecol Neonatal Nurs 2003, 32, 239-248. [CrossRef]
- Alexopoulos, G.S. Depression in the elderly. The Lancet 2005, 365, 1961-1970. [CrossRef]
- Park, C.; Rosenblat, J.D.; Brietzke, E.; Pan, Z.; Lee, Y.; Cao, B.; Zuckerman, H.; Kalantarova, A.; McIntyre, R.S. Stress, epigenetics and depression: A systematic review. Neuroscience & Biobehavioral Reviews 2019, 102, 139-152. [CrossRef]
- Felger, J.C.; Lotrich, F.E. Inflammatory cytokines in depression: Neurobiological mechanisms and therapeutic implications. Neuroscience 2013, 246, 199-229. [CrossRef]
- MacQueen, G.; Frodl, T. The hippocampus in major depression: evidence for the convergence of the bench and bedside in psychiatric research? Molecular Psychiatry 2010, 16, 252-264. [CrossRef]
- Salvat-Pujol, N.; Labad, J.; Urretavizcaya, M.; de Arriba-Arnau, A.; Segalàs, C.; Real, E.; Ferrer, A.; Crespo, J.M.; Jiménez-Murcia, S.; Soriano-Mas, C.; et al. Hypothalamic-pituitary-adrenal axis activity and cognition in major depression: The role of remission status. Psychoneuroendocrinology 2017, 76, 38-48. [CrossRef]
- Nierenberg, A.A. Current perspectives on the diagnosis and treatment of major depressive disorder. Am J Manag Care 2001, 7, S353-366.
- Pruckner, N.; Holthoff-Detto, V. Antidepressant pharmacotherapy in old-age depression—a review and clinical approach. European Journal of Clinical Pharmacology 2017, 73, 661-667. [CrossRef]
- Gonda, X.; Dome, P.; Neill, J.C.; Tarazi, F.I. Novel antidepressant drugs: Beyond monoamine targets. CNS Spectrums 2021, 28, 6-15. [CrossRef]
- Hashimoto, K.; Naudet, F.; Maria, A.S.; Falissard, B. Antidepressant Response in Major Depressive Disorder: A Meta-Regression Comparison of Randomized Controlled Trials and Observational Studies. PLoS ONE 2011, 6. [CrossRef]
- Li, Y.H.; Qing-Xiu, L.; Wang, J.S.; Xiang, H.; Zhang, R.F.; Huang, C.Q. Ghrelin improves cognition via activation of the cAMP- CREB signalling pathway in depressed male C57BL/6J mice. Int J Neurosci 2023, 133, 1233-1241. [CrossRef]
- Bianconi, S.; Poretti, M.B.; Rodríguez, P.; Maestri, G.; Rodríguez, P.E.; de Barioglio, S.R.; Schiöth, H.B.; Carlini, V.P. Ghrelin restores memory impairment following olfactory bulbectomy in mice by activating hippocampal NMDA1 and MAPK1 gene expression. Behav Brain Res 2021, 410, 113341. [CrossRef]
- Guo, L.; Niu, M.; Yang, J.; Li, L.; Liu, S.; Sun, Y.; Zhou, Z.; Zhou, Y. GHS-R1a Deficiency Alleviates Depression-Related Behaviors After Chronic Social Defeat Stress. Front Neurosci 2019, 13, 364. [CrossRef]
- Hirschfeld, R.M. History and evolution of the monoamine hypothesis of depression. J Clin Psychiatry 2000, 61 Suppl 6, 4-6.
- Jiang, H.; Betancourt, L.; Smith, R.G. Ghrelin Amplifies Dopamine Signaling by Cross Talk Involving Formation of Growth Hormone Secretagogue Receptor/Dopamine Receptor Subtype 1 Heterodimers. Molecular Endocrinology 2006, 20, 1772-1785. [CrossRef]
- Hansson, C.; Alvarez-Crespo, M.; Taube, M.; Skibicka, K.P.; Schmidt, L.; Karlsson-Lindahl, L.; Egecioglu, E.; Nissbrandt, H.; Dickson, S.L. Influence of ghrelin on the central serotonergic signaling system in mice. Neuropharmacology 2014, 79, 498-505. [CrossRef]
- Sakata, I.; Gong, Z.; Ikenoya, C.; Takemi, S.; Sakai, T. The study of ghrelin secretion and acyl-modification using mice and ghrelinoma cell lines. Endocr J 2017, 64, S27-s29. [CrossRef]
- Zhao, T.J.; Sakata, I.; Li, R.L.; Liang, G.; Richardson, J.A.; Brown, M.S.; Goldstein, J.L.; Zigman, J.M. Ghrelin secretion stimulated by {beta}1-adrenergic receptors in cultured ghrelinoma cells and in fasted mice. Proc Natl Acad Sci U S A 2010, 107, 15868-15873. [CrossRef]
- Granado, M.; Priego, T.; Martín, A.I.; Villanúa, M.Á.; López-Calderón, A. Anti-inflammatory effect of the ghrelin agonist growth hormone-releasing peptide-2 (GHRP-2) in arthritic rats. American Journal of Physiology-Endocrinology and Metabolism 2005, 288, E486-E492. [CrossRef]
- Schiepers, O.J.G.; Wichers, M.C.; Maes, M. Cytokines and major depression. Progress in Neuro-Psychopharmacology and Biological Psychiatry 2005, 29, 201-217. [CrossRef]
- Hodes, G.E.; Ménard, C.; Russo, S.J. Integrating Interleukin-6 into depression diagnosis and treatment. Neurobiology of Stress 2016, 4, 15-22. [CrossRef]
- Karmiris, K.; Koutroubakis, I.E.; Xidakis, C.; Polychronaki, M.; Voudouri, T.; Kouroumalis, E.A. Circulating levels of leptin, adiponectin, resistin, and ghrelin in inflammatory bowel disease. Inflamm Bowel Dis 2006, 12, 100-105. [CrossRef]
- Kerem, M.; Bedirli, A.; Pasaoglu, H.; Unsal, C.; Yilmaz, T.U.; Ofluoglu, E.; Sahin, T.T. Role of Ghrelin and Leptin in Predicting the Severity of Acute Pancreatitis. Digestive Diseases and Sciences 2007, 52, 950-955. [CrossRef]
- Zhao, D.; Zhan, Y.; Zeng, H.; Moyer, M.P.; Mantzoros, C.S.; Pothoulakis, C. Ghrelin stimulates interleukin-8 gene expression through protein kinase C-mediated NF-κB pathway in human colonic epithelial cells. Journal of Cellular Biochemistry 2005, 97, 1317-1327. [CrossRef]
- Otero, M. Chronic inflammation modulates ghrelin levels in humans and rats. Rheumatology 2003, 43, 306-310. [CrossRef]
- Cheyuo, C.; Wu, R.; Zhou, M.; Jacob, A.; Coppa, G.; Wang, P. Ghrelin Suppresses Inflammation and Neuronal Nitric Oxide Synthase in Focal Cerebral Ischemia Via the Vagus Nerve. Shock 2011, 35, 258-265. [CrossRef]
- Waseem, T.; Duxbury, M.; Ito, H.; Ashley, S.W.; Robinson, M.K. Exogenous ghrelin modulates release of pro-inflammatory and anti-inflammatory cytokines in LPS-stimulated macrophages through distinct signaling pathways. Surgery 2008, 143, 334-342. [CrossRef]
- Dixit, V.D.; Schaffer, E.M.; Pyle, R.S.; Collins, G.D.; Sakthivel, S.K.; Palaniappan, R.; Lillard, J.W.; Taub, D.D. Ghrelin inhibits leptin- and activation-induced proinflammatory cytokine expression by human monocytes and T cells. Journal of Clinical Investigation 2004, 114, 57-66. [CrossRef]
- Lin, L.; Lee, J.H.; Buras, E.D.; Yu, K.; Wang, R.; Smith, C.W.; Wu, H.; Sheikh-Hamad, D.; Sun, Y. Ghrelin receptor regulates adipose tissue inflammation in aging. Aging (Albany NY) 2016, 8, 178-191. [CrossRef]
- Guo, L.; Niu, M.; Yang, J.; Li, L.; Liu, S.; Sun, Y.; Zhou, Z.; Zhou, Y. GHS-R1a Deficiency Alleviates Depression-Related Behaviors After Chronic Social Defeat Stress. Frontiers in Neuroscience 2019, 13. [CrossRef]
- Krishnan, V.; Nestler, E.J. The molecular neurobiology of depression. Nature 2008, 455, 894-902. [CrossRef]
- Schmidt, H.D.; Shelton, R.C.; Duman, R.S. Functional Biomarkers of Depression: Diagnosis, Treatment, and Pathophysiology. Neuropsychopharmacology 2011, 36, 2375-2394. [CrossRef]
- Patas, K.; Penninx, B.W.J.H.; Bus, B.A.A.; Vogelzangs, N.; Molendijk, M.L.; Elzinga, B.M.; Bosker, F.J.; Oude Voshaar, R.C. Association between serum brain-derived neurotrophic factor and plasma interleukin-6 in major depressive disorder with melancholic features. Brain, Behavior, and Immunity 2014, 36, 71-79. [CrossRef]
- Berman, R.M.; Cappiello, A.; Anand, A.; Oren, D.A.; Heninger, G.R.; Charney, D.S.; Krystal, J.H. Antidepressant effects of ketamine in depressed patients. Biol Psychiatry 2000, 47, 351-354. [CrossRef]
- Zarate, C.A., Jr.; Singh, J.B.; Carlson, P.J.; Brutsche, N.E.; Ameli, R.; Luckenbaugh, D.A.; Charney, D.S.; Manji, H.K. A randomized trial of an N-methyl-D-aspartate antagonist in treatment-resistant major depression. Arch Gen Psychiatry 2006, 63, 856-864. [CrossRef]
- Li, N.; Lee, B.; Liu, R.-J.; Banasr, M.; Dwyer, J.M.; Iwata, M.; Li, X.-Y.; Aghajanian, G.; Duman, R.S. mTOR-Dependent Synapse Formation Underlies the Rapid Antidepressant Effects of NMDA Antagonists. Science 2010, 329, 959-964. [CrossRef]
- Park, S.; Kim, H.; Lee, D.; Ju, S.; Seo, S.; Lee, D.H.; Kim, E.; Chung, H. Ghrelin Inhibits Apoptosis in Hypothalamic Neuronal Cells during Oxygen-Glucose Deprivation. Endocrinology 2007, 148, 148-159. [CrossRef]
- Lee, J.Y.; Chung, H.; Yoo, Y.S.; Oh, Y.J.; Oh, T.H.; Park, S.; Yune, T.Y. Inhibition of Apoptotic Cell Death by Ghrelin Improves Functional Recovery after Spinal Cord Injury. Endocrinology 2010, 151, 3815-3826. [CrossRef]
- Li, E.; Chung, H.; Kim, Y.; Kim, D.H.; Ryu, J.H.; Sato, T.; Kojima, M.; Park, S. Ghrelin directly stimulates adult hippocampal neurogenesis: implications for learning and memory. Endocrine Journal 2013, 60, 781-789. [CrossRef]
- Walker, A.K.; Rivera, P.D.; Wang, Q.; Chuang, J.C.; Tran, S.; Osborne-Lawrence, S.; Estill, S.J.; Starwalt, R.; Huntington, P.; Morlock, L.; et al. The P7C3 class of neuroprotective compounds exerts antidepressant efficacy in mice by increasing hippocampal neurogenesis. Molecular Psychiatry 2014, 20, 500-508. [CrossRef]
- Chuang, J.-C.; Perello, M.; Sakata, I.; Osborne-Lawrence, S.; Savitt, J.M.; Lutter, M.; Zigman, J.M. Ghrelin mediates stress-induced food-reward behavior in mice. Journal of Clinical Investigation 2011, 121, 2684-2692. [CrossRef]
- Cuellar, J.N.; Isokawa, M. Ghrelin-induced activation of cAMP signal transduction and its negative regulation by endocannabinoids in the hippocampus. Neuropharmacology 2011, 60, 842-851. [CrossRef]
- Diano, S.; Farr, S.A.; Benoit, S.C.; McNay, E.C.; da Silva, I.; Horvath, B.; Gaskin, F.S.; Nonaka, N.; Jaeger, L.B.; Banks, W.A.; et al. Ghrelin controls hippocampal spine synapse density and memory performance. Nature Neuroscience 2006, 9, 381-388. [CrossRef]
- Bayliss, J.A.; Andrews, Z.B. Ghrelin is neuroprotective in Parkinson’s disease: molecular mechanisms of metabolic neuroprotection. Therapeutic Advances in Endocrinology and Metabolism 2013, 4, 25-36. [CrossRef]
- Lee, J.; Lim, E.; Kim, Y.; Li, E.; Park, S. Ghrelin attenuates kainic acid-induced neuronal cell death in the mouse hippocampus. Journal of Endocrinology 2010, 205, 263-270. [CrossRef]
- Fornaro, M.; Escelsior, A.; Rocchi, G.; Conio, B.; Magioncalda, P.; Marozzi, V.; Presta, A.; Sterlini, B.; Contini, P.; Amore, M.; et al. BDNF plasma levels variations in major depressed patients receiving duloxetine. Neurological Sciences 2014, 36, 729-734. [CrossRef]
- Banasr, M.; Duman, R.S. Glial Loss in the Prefrontal Cortex Is Sufficient to Induce Depressive-like Behaviors. Biological Psychiatry 2008, 64, 863-870. [CrossRef]
- Cotter, D.; Mackay, D.; Landau, S.; Kerwin, R.; Everall, I. Reduced glial cell density and neuronal size in the anterior cingulate cortex in major depressive disorder. Arch Gen Psychiatry 2001, 58, 545-553. [CrossRef]
- Czéh, B.; Simon, M.; Schmelting, B.; Hiemke, C.; Fuchs, E. Astroglial Plasticity in the Hippocampus is Affected by Chronic Psychosocial Stress and Concomitant Fluoxetine Treatment. Neuropsychopharmacology 2005, 31, 1616-1626. [CrossRef]
- Choudary, P.V.; Molnar, M.; Evans, S.J.; Tomita, H.; Li, J.Z.; Vawter, M.P.; Myers, R.M.; Bunney, W.E., Jr.; Akil, H.; Watson, S.J.; et al. Altered cortical glutamatergic and GABAergic signal transmission with glial involvement in depression. Proc Natl Acad Sci U S A 2005, 102, 15653-15658. [CrossRef]
- Banasr, M.; Chowdhury, G.M.I.; Terwilliger, R.; Newton, S.S.; Duman, R.S.; Behar, K.L.; Sanacora, G. Glial pathology in an animal model of depression: reversal of stress-induced cellular, metabolic and behavioral deficits by the glutamate-modulating drug riluzole. Molecular Psychiatry 2008, 15, 501-511. [CrossRef]
- Volterra, A.; Meldolesi, J. Astrocytes, from brain glue to communication elements: the revolution continues. Nature Reviews Neuroscience 2005, 6, 626-640. [CrossRef]
- Murphy, S.; Pearce, B. Functional receptors for neurotransmitters on astroglial cells. Neuroscience 1987, 22, 381-394. [CrossRef]
- Porter, J.T.; McCarthy, K.D. Astrocytic neurotransmitter receptors in situ and in vivo. Prog Neurobiol 1997, 51, 439-455. [CrossRef]
- Rose, C.R.; Blum, R.; Pichler, B.; Lepier, A.; Kafitz, K.W.; Konnerth, A. Truncated TrkB-T1 mediates neurotrophin-evoked calcium signalling in glia cells. Nature 2003, 426, 74-78. [CrossRef]
- Zhang, J.M.; Wang, H.K.; Ye, C.Q.; Ge, W.; Chen, Y.; Jiang, Z.L.; Wu, C.P.; Poo, M.M.; Duan, S. ATP released by astrocytes mediates glutamatergic activity-dependent heterosynaptic suppression. Neuron 2003, 40, 971-982. [CrossRef]
- Matyash, V.; Filippov, V.; Mohrhagen, K.; Kettenmann, H. Nitric Oxide Signals Parallel Fiber Activity to Bergmann Glial Cells in the Mouse Cerebellar Slice. Molecular and Cellular Neuroscience 2001, 18, 664-670. [CrossRef]
- Araque, A.; Martín, E.D.; Perea, G.; Arellano, J.I.; Buño, W. Synaptically released acetylcholine evokes Ca2+ elevations in astrocytes in hippocampal slices. J Neurosci 2002, 22, 2443-2450. [CrossRef]
- Bezzi, P.; Volterra, A. A neuron-glia signalling network in the active brain. Curr Opin Neurobiol 2001, 11, 387-394. [CrossRef]
- Halassa, M.M.; Haydon, P.G. Integrated Brain Circuits: Astrocytic Networks Modulate Neuronal Activity and Behavior. Annual Review of Physiology 2010, 72, 335-355. [CrossRef]
- Araque, A.; Parpura, V.; Sanzgiri, R.P.; Haydon, P.G. Tripartite synapses: glia, the unacknowledged partner. Trends Neurosci 1999, 22, 208-215. [CrossRef]
- Fuente-Martín, E.; García-Cáceres, C.; Argente-Arizón, P.; Díaz, F.; Granado, M.; Freire-Regatillo, A.; Castro-González, D.; Ceballos, M.L.; Frago, L.M.; Dickson, S.L.; et al. Ghrelin Regulates Glucose and Glutamate Transporters in Hypothalamic Astrocytes. Scientific Reports 2016, 6. [CrossRef]
- Baquedano, E.; Chowen, J.A.; Argente, J.; Frago, L.M. Differential effects of GH and GH-releasing peptide-6 on astrocytes. Journal of Endocrinology 2013, 218, 263-274. [CrossRef]
- Dixit, V.D.; Weeraratna, A.T.; Yang, H.; Bertak, D.; Cooper-Jenkins, A.; Riggins, G.J.; Eberhart, C.G.; Taub, D.D. Ghrelin and the Growth Hormone Secretagogue Receptor Constitute a Novel Autocrine Pathway in Astrocytoma Motility. Journal of Biological Chemistry 2006, 281, 16681-16690. [CrossRef]
- Fischer, R.; Maier, O. Interrelation of Oxidative Stress and Inflammation in Neurodegenerative Disease: Role of TNF. Oxidative Medicine and Cellular Longevity 2015, 2015, 1-18. [CrossRef]
- Pariante, C.M.; Lightman, S.L. The HPA axis in major depression: classical theories and new developments. Trends in Neurosciences 2008, 31, 464-468. [CrossRef]
- Vandoolaeghe, E.; Maes, M.; Vandevyvere, J.; Neels, H. Hypothalamic-pituitary-thyroid-axis function in treatment resistant depression. J Affect Disord 1997, 43, 143-150. [CrossRef]
- Bartalena, L.; Placidi, G.F.; Martino, E.; Falcone, M.; Pellegrini, L.; Dell'Osso, L.; Pacchiarotti, A.; Pinchera, A. Nocturnal serum thyrotropin (TSH) surge and the TSH response to TSH-releasing hormone: dissociated behavior in untreated depressives. J Clin Endocrinol Metab 1990, 71, 650-655. [CrossRef]
- Fischer, S.; Ehlert, U.; Amiel Castro, R. Hormones of the hypothalamic-pituitary-gonadal (HPG) axis in male depressive disorders – A systematic review and meta-analysis. Frontiers in Neuroendocrinology 2019, 55. [CrossRef]
- Han, Y.; Gu, S.; Li, Y.; Qian, X.; Wang, F.; Huang, J.H. Neuroendocrine pathogenesis of perimenopausal depression. Frontiers in Psychiatry 2023, 14. [CrossRef]
- Kluge, M.; Schmidt, D.; Uhr, M.; Steiger, A. Ghrelin suppresses nocturnal secretion of luteinizing hormone (LH) and thyroid stimulating hormone (TSH) in patients with major depression. Journal of Psychiatric Research 2013, 47, 1236-1239. [CrossRef]
- Kluge, M.; Riedl, S.; Uhr, M.; Schmidt, D.; Zhang, X.; Yassouridis, A.; Steiger, A. Ghrelin affects the hypothalamus–pituitary–thyroid axis in humans by increasing free thyroxine and decreasing TSH in plasma. European Journal of Endocrinology 2010, 162, 1059-1065. [CrossRef]
- Gupta, D.; Chuang, J.-C.; Mani, B.K.; Shankar, K.; Rodriguez, J.A.; Osborne-Lawrence, S.; Metzger, N.P.; Zigman, J.M. β1-adrenergic receptors mediate plasma acyl-ghrelin elevation and depressive-like behavior induced by chronic psychosocial stress. Neuropsychopharmacology 2019, 44, 1319-1327. [CrossRef]
- Zuccato, C.; Cattaneo, E. Brain-derived neurotrophic factor in neurodegenerative diseases. Nature Reviews Neurology 2009, 5, 311-322. [CrossRef]
- Keifer, J. Regulation of AMPAR trafficking in synaptic plasticity by BDNF and the impact of neurodegenerative disease. J Neurosci Res 2022, 100, 979-991. [CrossRef]
- Zhang, K.; Wang, F.; Zhai, M.; He, M.; Hu, Y.; Feng, L.; Li, Y.; Yang, J.; Wu, C. Hyperactive neuronal autophagy depletes BDNF and impairs adult hippocampal neurogenesis in a corticosterone-induced mouse model of depression. Theranostics 2023, 13, 1059-1075. [CrossRef]
- Liu, S.; Tao, G.; Zhou, C.; Wang, Q.; Wang, W.; Fei, X. Ketamine inhibits neuronal differentiation by regulating brain-derived neurotrophic factor (BDNF) signaling. Toxicol In Vitro 2021, 72, 105091. [CrossRef]
- Perea Vega, M.L.; Sanchez, M.S.; Fernández, G.; Paglini, M.G.; Martin, M.; de Barioglio, S.R. Ghrelin treatment leads to dendritic spine remodeling in hippocampal neurons and increases the expression of specific BDNF-mRNA species. Neurobiology of Learning and Memory 2021, 179. [CrossRef]
- Baj, G.; Leone, E.; Chao, M.V.; Tongiorgi, E. Spatial segregation of BDNF transcripts enables BDNF to differentially shape distinct dendritic compartments. Proc Natl Acad Sci U S A 2011, 108, 16813-16818. [CrossRef]
- Aid, T.; Kazantseva, A.; Piirsoo, M.; Palm, K.; Timmusk, T. Mouse and rat BDNF gene structure and expression revisited. J Neurosci Res 2007, 85, 525-535. [CrossRef]
- Duman, R.S.; Heninger, G.R.; Nestler, E.J. A molecular and cellular theory of depression. Arch Gen Psychiatry 1997, 54, 597-606. [CrossRef]
- Banerjee, R.; Ghosh, A.K.; Ghosh, B.; Bhattacharyya, S.; Mondal, A.C. Decreased mRNA and Protein Expression of BDNF, NGF, and their Receptors in the Hippocampus from Suicide: An Analysis in Human Postmortem Brain. Clin Med Insights Pathol 2013, 6, 1-11. [CrossRef]
- Paska, A.V.; Zupanc, T.; Pregelj, P. The role of brain-derived neurotrophic factor in the pathophysiology of suicidal behavior. Psychiatr Danub 2013, 25 Suppl 2, S341-344.
- Dowlatshahi, D.; MacQueen, G.M.; Wang, J.F.; Young, L.T. Increased temporal cortex CREB concentrations and antidepressant treatment in major depression. Lancet 1998, 352, 1754-1755. [CrossRef]
- Nakagawa, S.; Kim, J.E.; Lee, R.; Malberg, J.E.; Chen, J.; Steffen, C.; Zhang, Y.J.; Nestler, E.J.; Duman, R.S. Regulation of neurogenesis in adult mouse hippocampus by cAMP and the cAMP response element-binding protein. J Neurosci 2002, 22, 3673-3682. [CrossRef]
- Lee, B.H.; Kim, Y.K. BDNF mRNA expression of peripheral blood mononuclear cells was decreased in depressive patients who had or had not recently attempted suicide. J Affect Disord 2010, 125, 369-373. [CrossRef]
- Fan, J.; Li, B.J.; Wang, X.F.; Zhong, L.L.; Cui, R.J. Ghrelin produces antidepressant-like effect in the estrogen deficient mice. Oncotarget 2017, 8, 58964-58973. [CrossRef]
- Lizama, C.; Lagos, C.F.; Lagos-Cabré, R.; Cantuarias, L.; Rivera, F.; Huenchuñir, P.; Pérez-Acle, T.; Carrión, F.; Moreno, R.D. Calpain inhibitors prevent p38 MAPK activation and germ cell apoptosis after heat stress in pubertal rat testes. J Cell Physiol 2009, 221, 296-305. [CrossRef]
- Peng, Z.; Wang, H.; Zhang, R.; Chen, Y.; Xue, F.; Nie, H.; Chen, Y.; Wu, D.; Wang, Y.; Wang, H.; et al. Gastrodin ameliorates anxiety-like behaviors and inhibits IL-1beta level and p38 MAPK phosphorylation of hippocampus in the rat model of posttraumatic stress disorder. Physiol Res 2013, 62, 537-545. [CrossRef]
- Zoga, M.; Oulis, P.; Chatzipanagiotou, S.; Masdrakis, V.G.; Pliatsika, P.; Boufidou, F.; Foteli, S.; Soldatos, C.R.; Nikolaou, C.; Papageorgiou, C. Indoleamine 2,3-dioxygenase and immune changes under antidepressive treatment in major depression in females. In Vivo 2014, 28, 633-638.
- Zhou, W.; Dantzer, R.; Budac, D.P.; Walker, A.K.; Mao-Ying, Q.L.; Lee, A.W.; Heijnen, C.J.; Kavelaars, A. Peripheral indoleamine 2,3-dioxygenase 1 is required for comorbid depression-like behavior but does not contribute to neuropathic pain in mice. Brain Behav Immun 2015, 46, 147-153. [CrossRef]
- Cheng, Y.; Qiao, Z.; Dang, C.; Zhou, B.; Li, S.; Zhang, W.; Jiang, J.; Song, Y.; Zhang, J.; Diao, D. p38 predicts depression and poor outcome in esophageal cancer. Oncol Lett 2017, 14, 7241-7249. [CrossRef]
- Irusen, E.; Matthews, J.G.; Takahashi, A.; Barnes, P.J.; Chung, K.F.; Adcock, I.M. p38 Mitogen-activated protein kinase-induced glucocorticoid receptor phosphorylation reduces its activity: role in steroid-insensitive asthma. J Allergy Clin Immunol 2002, 109, 649-657. [CrossRef]
- Holsboer, F. The corticosteroid receptor hypothesis of depression. Neuropsychopharmacology 2000, 23, 477-501. [CrossRef]
- Liu, C.; Huang, J.; Li, H.; Yang, Z.; Zeng, Y.; Liu, J.; Hao, Y.; Li, R. Ghrelin accelerates wound healing through GHS-R1a-mediated MAPK-NF-κB/GR signaling pathways in combined radiation and burn injury in rats. Sci Rep 2016, 6, 27499. [CrossRef]
- Han, Q.Q.; Huang, H.J.; Wang, Y.L.; Yang, L.; Pilot, A.; Zhu, X.C.; Yu, R.; Wang, J.; Chen, X.R.; Liu, Q.; et al. Ghrelin exhibited antidepressant and anxiolytic effect via the p38-MAPK signaling pathway in hippocampus. Prog Neuropsychopharmacol Biol Psychiatry 2019, 93, 11-20. [CrossRef]
- Bruchas, M.R.; Schindler, A.G.; Shankar, H.; Messinger, D.I.; Miyatake, M.; Land, B.B.; Lemos, J.C.; Hagan, C.E.; Neumaier, J.F.; Quintana, A.; et al. Selective p38α MAPK deletion in serotonergic neurons produces stress resilience in models of depression and addiction. Neuron 2011, 71, 498-511. [CrossRef]
- Schoenfeld, T.J.; Cameron, H.A. Adult neurogenesis and mental illness. Neuropsychopharmacology 2015, 40, 113-128. [CrossRef]
- Peltier, J.; O'Neill, A.; Schaffer, D.V. PI3K/Akt and CREB regulate adult neural hippocampal progenitor proliferation and differentiation. Dev Neurobiol 2007, 67, 1348-1361. [CrossRef]
- Doble, B.W.; Woodgett, J.R. GSK-3: tricks of the trade for a multi-tasking kinase. Journal of Cell Science 2003, 116, 1175-1186. [CrossRef]
- Hart, M.J.; de los Santos, R.; Albert, I.N.; Rubinfeld, B.; Polakis, P. Downregulation of beta-catenin by human Axin and its association with the APC tumor suppressor, beta-catenin and GSK3 beta. Curr Biol 1998, 8, 573-581. [CrossRef]
- Ryu, J.K.; Choi, H.B.; Hatori, K.; Heisel, R.L.; Pelech, S.L.; McLarnon, J.G.; Kim, S.U. Adenosine triphosphate induces proliferation of human neural stem cells: Role of calcium and p70 ribosomal protein S6 kinase. J Neurosci Res 2003, 72, 352-362. [CrossRef]
- Chung, H.; Li, E.; Kim, Y.; Kim, S.; Park, S. Multiple signaling pathways mediate ghrelin-induced proliferation of hippocampal neural stem cells. Journal of Endocrinology 2013, 218, 49-59. [CrossRef]
- Chung, H.; Seo, S.; Moon, M.; Park, S. Phosphatidylinositol-3-kinase/Akt/glycogen synthase kinase-3β and ERK1/2 pathways mediate protective effects of acylated and unacylated ghrelin against oxygen–glucose deprivation-induced apoptosis in primary rat cortical neuronal cells. Journal of Endocrinology 2008, 198, 511-521. [CrossRef]
- Ferreira-Marques, M.; Carvalho, A.; Cavadas, C.; Aveleira, C.A. PI3K/AKT/MTOR and ERK1/2-MAPK signaling pathways are involved in autophagy stimulation induced by caloric restriction or caloric restriction mimetics in cortical neurons. Aging (Albany NY) 2021, 13, 7872-7882. [CrossRef]
- Hou, Y.; Wang, K.; Wan, W.; Cheng, Y.; Pu, X.; Ye, X. Resveratrol provides neuroprotection by regulating the JAK2/STAT3/PI3K/AKT/mTOR pathway after stroke in rats. Genes Dis 2018, 5, 245-255. [CrossRef]
- Zhu, H.; Jian, Z.; Zhong, Y.; Ye, Y.; Zhang, Y.; Hu, X.; Pu, B.; Gu, L.; Xiong, X. Janus Kinase Inhibition Ameliorates Ischemic Stroke Injury and Neuroinflammation Through Reducing NLRP3 Inflammasome Activation via JAK2/STAT3 Pathway Inhibition. Front Immunol 2021, 12, 714943. [CrossRef]
- Zhang, W.; Xu, M.; Chen, F.; Su, Y.; Yu, M.; Xing, L.; Chang, Y.; Yan, T. Targeting the JAK2-STAT3 pathway to inhibit cGAS-STING activation improves neuronal senescence after ischemic stroke. Exp Neurol 2023, 368, 114474. [CrossRef]
- Ring, R.H.; Malberg, J.E.; Potestio, L.; Ping, J.; Boikess, S.; Luo, B.; Schechter, L.E.; Rizzo, S.; Rahman, Z.; Rosenzweig-Lipson, S. Anxiolytic-like activity of oxytocin in male mice: behavioral and autonomic evidence, therapeutic implications. Psychopharmacology (Berl) 2006, 185, 218-225. [CrossRef]
- Weissman, M.M.; Klerman, G.L. Sex differences and the epidemiology of depression. Arch Gen Psychiatry 1977, 34, 98-111. [CrossRef]
- Bleickardt, C.J.; Mullins, D.E.; Macsweeney, C.P.; Werner, B.J.; Pond, A.J.; Guzzi, M.F.; Martin, F.D.; Varty, G.B.; Hodgson, R.A. Characterization of the V1a antagonist, JNJ-17308616, in rodent models of anxiety-like behavior. Psychopharmacology (Berl) 2009, 202, 711-718. [CrossRef]
- Serradeil-Le Gal, C.; Wagnon, J., 3rd; Tonnerre, B.; Roux, R.; Garcia, G.; Griebel, G.; Aulombard, A. An overview of SSR149415, a selective nonpeptide vasopressin V(1b) receptor antagonist for the treatment of stress-related disorders. CNS Drug Rev 2005, 11, 53-68. [CrossRef]
- Lutter, M.; Sakata, I.; Osborne-Lawrence, S.; Rovinsky, S.A.; Anderson, J.G.; Jung, S.; Birnbaum, S.; Yanagisawa, M.; Elmquist, J.K.; Nestler, E.J.; et al. The orexigenic hormone ghrelin defends against depressive symptoms of chronic stress. Nat Neurosci 2008, 11, 752-753. [CrossRef]
- Zhang, Y.; Zhu, M.Z.; Qin, X.H.; Zeng, Y.N.; Zhu, X.H. The Ghrelin/Growth Hormone Secretagogue Receptor System Is Involved in the Rapid and Sustained Antidepressant-Like Effect of Paeoniflorin. Front Neurosci 2021, 15, 631424. [CrossRef]
- Wu, R.; Xiao, D.; Shan, X.; Dong, Y.; Tao, W.W. Rapid and Prolonged Antidepressant-like Effect of Crocin Is Associated with GHSR-Mediated Hippocampal Plasticity-related Proteins in Mice Exposed to Prenatal Stress. ACS chemical neuroscience 2020, 11, 1159-1170. [CrossRef]
- Rana, T.; Behl, T.; Sehgal, A.; Singh, S.; Sharma, N.; Abdeen, A.; Ibrahim, S.F.; Mani, V.; Iqbal, M.S.; Bhatia, S.; et al. Exploring the role of neuropeptides in depression and anxiety. Prog Neuropsychopharmacol Biol Psychiatry 2022, 114, 110478. [CrossRef]
- Jackson, T.M.; Ostrowski, T.D.; Middlemas, D.S. Intracerebroventricular Ghrelin Administration Increases Depressive-Like Behavior in Male Juvenile Rats. Front Behav Neurosci 2019, 13, 77. [CrossRef]
- Hansson, C.; Haage, D.; Taube, M.; Egecioglu, E.; Salomé, N.; Dickson, S.L. Central administration of ghrelin alters emotional responses in rats: behavioural, electrophysiological and molecular evidence. Neuroscience 2011, 180, 201-211. [CrossRef]
- Jensen, M.; Ratner, C.; Rudenko, O.; Christiansen, S.H.; Skov, L.J.; Hundahl, C.; Woldbye, D.P.; Holst, B. Anxiolytic-Like Effects of Increased Ghrelin Receptor Signaling in the Amygdala. The international journal of neuropsychopharmacology 2016, 19. [CrossRef]
- Mahbod, P.; Smith, E.P.; Fitzgerald, M.E.; Morano, R.L.; Packard, B.A.; Ghosal, S.; Scheimann, J.R.; Perez-Tilve, D.; Herman, J.P.; Tong, J. Desacyl Ghrelin Decreases Anxiety-like Behavior in Male Mice. Endocrinology 2018, 159, 388-399. [CrossRef]
- Pawar, G.R.; Agrawal, Y.O.; Nakhate, K.T.; Patil, C.R.; Sharma, C.; Ojha, S.; Mahajan, U.B.; Goyal, S.N. Ghrelin alleviates depression-like behaviour in rats subjected to high-fat diet and diurnal rhythm disturbance. Am J Transl Res 2022, 14, 7098-7108.
- Sun, N.; Mei, Y.; Hu, Z.; Xing, W.; Lv, K.; Hu, N.; Zhang, T.; Wang, D. Ghrelin attenuates depressive-like behavior, heart failure, and neuroinflammation in postmyocardial infarction rat model. Eur J Pharmacol 2021, 901, 174096. [CrossRef]
- Huang, H.J.; Chen, X.R.; Han, Q.Q.; Wang, J.; Pilot, A.; Yu, R.; Liu, Q.; Li, B.; Wu, G.C.; Wang, Y.Q.; et al. The protective effects of Ghrelin/GHSR on hippocampal neurogenesis in CUMS mice. Neuropharmacology 2019, 155, 31-43. [CrossRef]
Figure 1.
Highlighting the antidepressant potential of ghrelin, which exerts neuroprotective effects by promoting neuronal proliferation and neurotrophic factor production in the brain. Ghrelin can inhibit apoptosis, reduce inflammation, increase astrocyte production, and significantly inhibit the process of depression. ghrelin interacts with GHS-R1a to increase the proliferation of hippocampal NSCs by increasing the expression of the Jak2/STAT3 signaling pathway, which also indirectly upregulates the PI3K/Akt/mTOR pathway to jointly exert neuronal protective effects. In addition, ghrelin binding to its receptor can also increase the expression of PI3K/Akt signaling pathway, play a pro-neural cell proliferation role by promoting the phosphorylation of downstream molecules, mTOR, p70S6K, and also inactivate GSK-3β by promoting GSK-3β phosphorylation, which can further increase the nuclear translocation of β-catenin, thus decreasing the cellular apoptosis, playing a role in promoting cell survival. The activation of this pathway also promotes the proliferation of astrocytes in the brain and the communication of neurons in the brain. Moreover, ghrelin treatment can inhibit the p38-MAPK signaling pathway, increase GR expression, and inhibit glucocorticoid resistance in depressed patients. Additionally, ghrelin increases CREB phosphorylation by promoting the expression of cAMP/CREB signaling pathway, thus regulating BDNF transcription and eventually exerting neuroprotective effects to inhibit depression. Apart from this, ghrelin elevates its involvement in the regulation of inflammatory response, down-regulates neutrophil transport and the amount of pro-inflammatory cytokine, thus reducing inflammation, and also increases the expression of BDNF and improves neurobehavioral function. Subtraction symbol indicates inhibitory/suppressing action, while addition symbol indicates stimulatory action. The P labeled in the circle represents substrate phosphorylation.↑, increasing; ↓, decreasing.
Figure 1.
Highlighting the antidepressant potential of ghrelin, which exerts neuroprotective effects by promoting neuronal proliferation and neurotrophic factor production in the brain. Ghrelin can inhibit apoptosis, reduce inflammation, increase astrocyte production, and significantly inhibit the process of depression. ghrelin interacts with GHS-R1a to increase the proliferation of hippocampal NSCs by increasing the expression of the Jak2/STAT3 signaling pathway, which also indirectly upregulates the PI3K/Akt/mTOR pathway to jointly exert neuronal protective effects. In addition, ghrelin binding to its receptor can also increase the expression of PI3K/Akt signaling pathway, play a pro-neural cell proliferation role by promoting the phosphorylation of downstream molecules, mTOR, p70S6K, and also inactivate GSK-3β by promoting GSK-3β phosphorylation, which can further increase the nuclear translocation of β-catenin, thus decreasing the cellular apoptosis, playing a role in promoting cell survival. The activation of this pathway also promotes the proliferation of astrocytes in the brain and the communication of neurons in the brain. Moreover, ghrelin treatment can inhibit the p38-MAPK signaling pathway, increase GR expression, and inhibit glucocorticoid resistance in depressed patients. Additionally, ghrelin increases CREB phosphorylation by promoting the expression of cAMP/CREB signaling pathway, thus regulating BDNF transcription and eventually exerting neuroprotective effects to inhibit depression. Apart from this, ghrelin elevates its involvement in the regulation of inflammatory response, down-regulates neutrophil transport and the amount of pro-inflammatory cytokine, thus reducing inflammation, and also increases the expression of BDNF and improves neurobehavioral function. Subtraction symbol indicates inhibitory/suppressing action, while addition symbol indicates stimulatory action. The P labeled in the circle represents substrate phosphorylation.↑, increasing; ↓, decreasing.
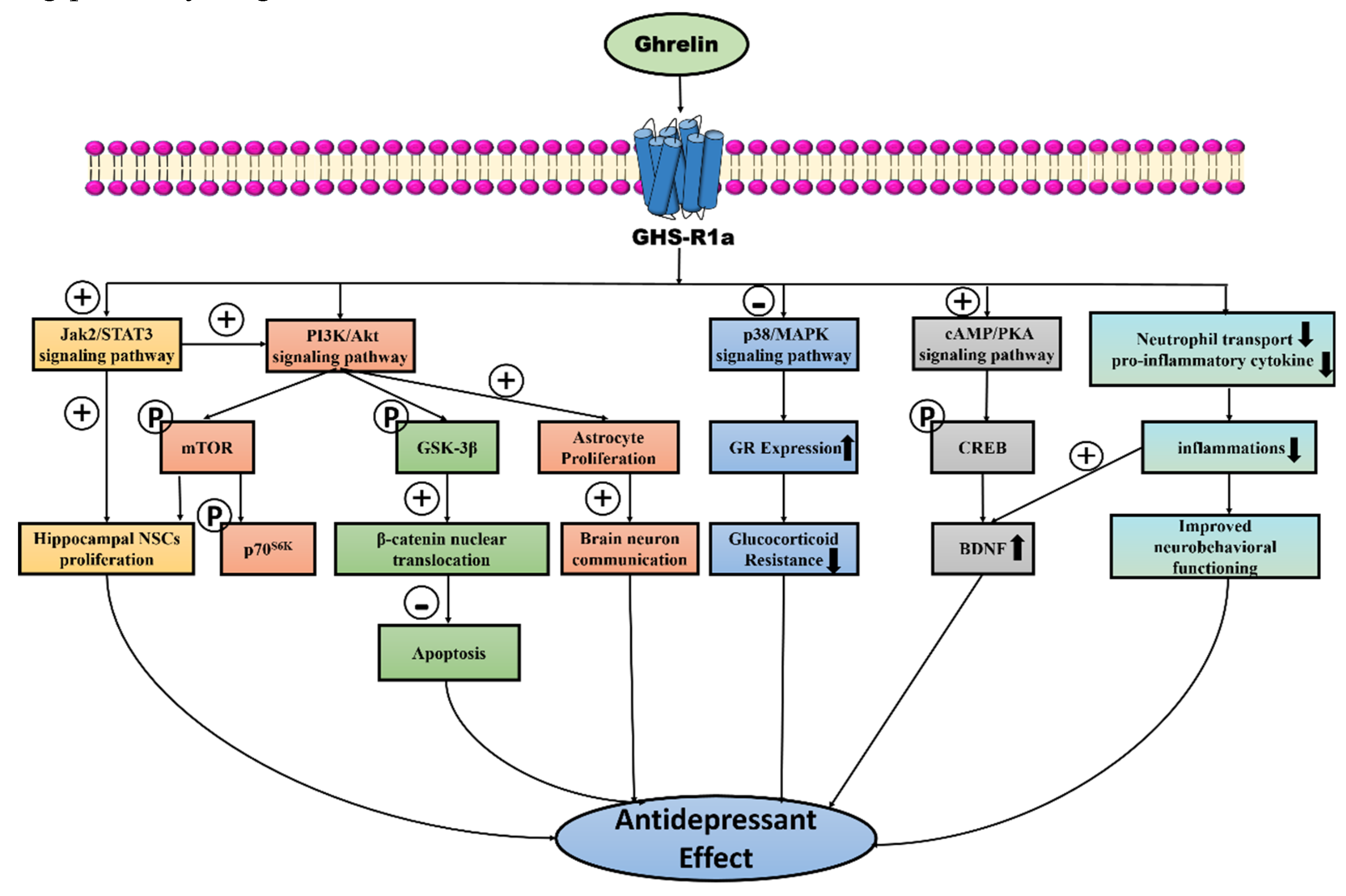

Table 1.
Effect of ghrelin/GHSR system on MDD in rodents.
| Animal and Stress paradigm | Behavioral test | Intervention | Signal molecules | Effects | Reference |
| Mice CSDS | FST, TST | GHSR1a knock-out | BDNF↓, IL-6↑ | Pro-depression effect | [29] |
| Rat | FST, TST | i.c.v injection of ghrelin | HPA↑ | Immobility time↑ in TST | [133,134] |
| Male C57BL/J6 mice, RS | TST OFT FST |
rAAV-Mediated Overexpression of GHSR1a | c-Fos↑ | Antidepressant-like effect | [135] |
| Male mice, Restraint stress | EPM | Ghrelin KO | pERK↓ | Decreases Anxiety-like Behavior |
[136] |
| C57BL/J6 mice, Prenatal stress | OFT,TST FST,SPT |
i.p. injection of crocin | PI3K/Akt↑ mTOR↑ |
Antidepressant-like effect | [131] |
| Male SD rats, HFD and DDR | FST OFT EPM |
Intra-VTA administration of ghrelin | TNF-α↓ IL-1β↓ IL-6↓ |
Alleviates depression-like behaviour | [137] |
| Male SD rats, CAO | SPT,OFT EPM |
subcutaneously injection of ghrelin | Iba-1↓ GFAP↓ |
Attenuates depressive-like behavior | [138] |
| Male C57BL/J6 mice, CSDS | SIT, FST,OFT EPM |
Intrahippocampal ghrelin infusions, AAV-siRNA of GHSR1a |
p38-MAPK↓ | Antidepressant effect | [112] |
| Male C57BL/J6 mice | TST FST |
Lateral ventricle injection of ghrelin | CREB↑ BDNF↑ |
Improves cognition Antidepressant-like effects |
[27] [103] |
| Adult female mice,OB | OFT TST |
Ghrelin into the hippocampus | MAPK↑ CaMKIIa↑ |
Antidepressant-like effects | [28] |
| Male C57BL/J6 mice,CUMS | OFT EPM FST |
Intraperitoneally (i.p.) injection of ghrelin, GHSR knockdown | PI3K/Akt ↑ CDK2↑ CyclinD1↑ |
Spine density↑ Proliferation of hippocampal NSCs Neurogenesis↑ |
[139] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



