
Submitted:
31 May 2024
Posted:
03 June 2024
You are already at the latest version
Abstract
This study aimed to explore the fermentation behavior of selected potential probiotic strains during sorghum fermentation, considering their microbial counts, pH levels, and antibacterial activity, their ability to break down starch. The fermentation processes were conducted in the presence and absence of glucose, and the lactic acid bacterial strains underwent controlled fermentation to assess microbial counts, amylase activity, pH, and lactic acid levels in the presence and absence of glucose. In addition, the inhibitory activity of these potential probiotic strains against pathogenic bacteria (Salmonella typhimurium, Escherichia coli, and Staph-ylococcus aureus) was examined through co-culturing. The results demonstrated that all nine Lactobacillus strains exhibited robust growth in both sugar and sugar-free fermentation experiments. Glucose supple-mentation significantly enhanced lactic acid yield which was in a range of 0.19-0.52 % compared to fer-mentation without glucose with a range of 0.03-0.29 %. The selected Lactobacillus strains effectively lowered the pH to less than 4 after 24 hours, producing substantial lactic acid. Notably, in the absence of glucose, only Lb. helveticus D7 and Lb. amylolyticus D12 achieved pH levels below 4 after 8 hours, producing the highest lactic acid amounts of 0.27 and 0.29 % after 24 hours, respectively. Amylase activity was detected on two strains D7 and D12. Furthermore, most of the tested Lactobacillus strains demonstrated complete inhibition of pathogen growth after 24 hours of co-culturing, suggesting their potential for enhancing the safety quality of sorghum-based fermented products.
Keywords:
Fermentation
; Sorghum
; Amylolytic LAB
1. Introduction
Lactic acid bacteria play a crucial role in many African fermented cereal-based products such as such as Ogi, Mageu, and Togwa by controlling foodborne pathogens. They are used to preserve food, increase the bioavailability of nutrients and functional properties of foods, [1,2,3]. Some LAB species have been well documented for their probiotic properties and have been used in many fermented products such Keffir, Kimchi, and Kombucha etc., [4]. LAB produces lactic acid as a major metabolic end-product of carbohydrate fermentation and thus exhibits an increased tolerance to acidity. These bacteria contribute to the organoleptic and textural profiles of many fermented foods, and they have been reported to confer health benefits when ingested in adequate amounts [5]. LAB has been extensively used in the fermentation of cereal-based foods in Africa (Kisra, Ogi, Acida and Merissa). The process of fermentation of carbohydrates by LAB involves the breakdown of six-carbon sugar glucose into two molecules of the three-carbon organic acid, pyruvic acid [6]. This process is coupled with the transfer of chemical energy to the synthesis of adenosine triphosphate (ATP) [7]. Depending on the availability of oxygen, pyruvate can either be oxidized through the tricarboxylic acid cycle or, in the absence of oxygen, to produce lactic acid as the major fermentation product [7,8]. Several different substrates (Milk, starches, oligosaccharides, glycerol, and other complex carbohydrates) are used for the fermentation of lactic acid by LAB. Therefore, the carbon source for microbial production of lactic acid can either be a sugar in its pure form, or it must be obtained from other substrates i.e. starch, during fermentation, [9].
Amylolytic or amylase producing LAB (ALAB) play a vital role in fermentation processes for efficient carbohydrate utilization during the fermentation of starch-based products. ALAB hydrolyzes the starch molecules into polymers composed of glucose units, and plays a role in flavor development, preservation, and nutritional enhancement of cereal-based products without the need to add sugar in the fermentation process. Lactic acid fermentation is linked to a decrease in pH with a simultaneous increase in acidity, as with lactic acid and other organic acids. This mechanism results in decreasing the survival time for pathogenic bacteria such as Escherichia coli, Staphylococcus aureus during food processing [10,11]. LAB have been incorporated into a cereal based product called togwa [12], and it has been demonstrated to significantly reduce enteropathogenesis in children under 5 years.
Sorghum is a staple food for many households and proves to be the grain of importance in Africa due to its high drought tolerance and low price [13]. Sorghum grains contain a substantial amount of carbohydrates, minerals, and vitamins. In Southern Africa, it is consumed in South Africa, Botswana, Zimbabwe and Lesotho throughout the year [10,13]. LAB isolated from fermented sorghum slurries (Ting) were shown to possess probiotic properties. Similar cereal based probiotic products includes togwa, busheri etc. Probiotic strains belonging to Lactiplantibacillus plantarum, are commonly used in the food industry, have frequently been isolated from traditional cereal-based fermentations and extensive research has highlighted its probiotic potential, showing that Lb. plantarum isolated from sourdough can establish itself in the human intestinal tract [14]. The exploration of cereal-based probiotic foods, leveraging the advantages of live cultures, emerges as a cost-effective and accessible solution with potential health benefits for diverse populations. Younger children could benefit from high energy, drinkable food products derived from starchy substrate (i.e. sorghum) fermented by amylolytic probiotic strains. These kinds of products could have an added assist in the alleviation of malnutrition and stunting in young children. Malnutrition and stunting are still a problem in most of the developing countries, including South Africa. However, the performance of these probiotics in the preparation of different substrates was not established. In the current study, Lactobacillus strains were examined to assess their fermentative capabilities, analyzing microbial counts, amylase activity, pH, and lactic acid production. This study aims to establish the amylolytic activity of probiotic ALAB isolated from Ting (fermented sorghum porridge) and their performance for application in the development of high energy food products suitable for young children.
2. Materials and Methods
2.1. Bacterial strains and Culture Conditions
4 probiotic strains were obtained from the Department of Biotechnology and Food Technology's culture collection (Tshwane University of Technology). The strains utilized in the current study had previously been tested for their probiotic potential in research conducted by [15]. The sequences of these strains were determined and deposited into the National Center for Biotechnology Information database (refer to supplementary Table S1 for list of strains and their sources). The Lactobacillus strains were initially cultivated in a propagation medium to allow them to acclimatize to the fermentation environment. The propagation medium was prepared as follows: MgSO4 (0.5 g/L), yeast extract (5 g/L), dextrose (10 g/L), NaCl (2.5 g/L), KH2PO4 (1.25 g/L), bacterial peptone (1 g/L), calcium carbonate, and sorghum (80 g/L) were carefully weighed and combined in 1 L of deionized water and placed in a cooking pot. The ingredients were heated for 15 minutes until the gelatinization of the sorghum slurry was observed. Subsequently, the slurry was dispensed into 250 mL Schott bottles and autoclaved at 121 º C for 15 minutes. The propagation medium was inoculated with the Probiotic strains listed in Table S1, and the samples were incubated at 37 º C in a water bath for 16 hours, to allow the pH of the medium to reach 3.5. This was done in duplicate and a volume of 25 mL of the propagated medium was used for each Lactobacillus strain for batch fermentation study.
2.2. Amylolytic Activity of LAB Strains
The 4 selected potential probiotic strains were grown on starch agar plates and starch was used as the only source of carbon. The assaying method was according to Bogale et al (2015). The medium is composed of corn starch (2%), bacterial peptone (0.5%), beef extract (0.5%), yeast extract (0.2%), sodium chloride (0.2%) and agar (1.5%). A volume of 30 µL overnight bacterial culture was inoculated at the center of the agar medium. The plates were left to dry out for 2 hours prior to aerobic incubation at 37 °C for 72 hours. Thereafter, the colonies were removed from agar medium using sterile blade and the plates were flooded with iodine for 30 minutes. The plates showing a zone of clearing were considered as positive for amylase enzyme and the experiment was repeated twice. Further work was undertaken to determine the viscosity of fermented sorghum porridge whereby sorghum flour 8% (w/v) was mixed with distilled water and cooked for 15 minutes at 94 °C to make porridge. The porridge was sterilized using an autoclave, cooled at room temperature, and thereafter inoculated with 4% (v/v) of bacteria strains. The porridges were incubated at 37 °C for 24 hours and their viscosities were measured using the Brookfield RV viscometer with spindles 4 and 6 at different rotation speeds. All viscosity measurements were done in thermostated conditions at 45 °C.
2.3. DNA Amplification of Amylase Gene of Probiotic Strains
Genomic DNA of the Probiotic isolates was extracted using a DNA Purification Kit (Epicenter, USA) by following manufacturer’s protocol. DNA amplification was done using thermal cycler (Eppendorf AG 22331 thermal cycler) by targeting the amylase gene, the primers used in the study were as follows amyA -1-F 5’- CGTCCAGTTGATGGTGGTAATG-3 and amyA -1-R 5’- CCAGGCCAAGCTGAAGTTATAG-3. The following volumes were measured to perform PCR: 12.5 μL Master mix (KAPA Biosystems), 1 μL forward primer and 1μL Reverse primer, 9.5 μL RNase nuclease free water and 1 μL DNA template to make a reaction volume of 25 μL. The PCR conditions were as follows: initial denaturation (94 °C for 15 minutes), 30 cycles of denaturation (94 °C for 30 seconds), annealing (63 °C for 30 seconds), extension (72 °C for 3 minutes) and final extension (72 °C for 7 minutes).
2.4. Sorghum Fermentation with LAB Strains
2.4.1. Fermentation Medium
The fermentation medium was prepared by combining cooked sorghum (60 g/L) and glucose (20 g/L), referred to as CSG (cooked sorghum and glucose). The second fermentation medium solely comprised of cooked sorghum (60 g/L) and was designated as CS.
2.4.2. Population Levels of Probiotic Strains during Fermentation
To determine the population levels of the Lactobacillus strains, 1 mL of each fermentation sample was withdrawn aseptically using a sterile 10 mL syringe. Tenfold serial dilutions were carried out, and 0.1 mL aliquots were evenly spread on MRS Agar plates (Merck). The plates were then incubated at 37 ºC and bacterial cell counts were recorded at 0, 4, 8, 16, and 24 hours.
2.4.3. pH
The pH of the samples fermented with the Lactobacillus strains was measured using a pH meter (Orion Star-A211, ThermoFisher Scientific, South Africa). The pH meter was calibrated using buffer solutions at pH 7 and pH 4. Before each analysis, the glass electrode was rinsed with sterile deionized water. The pH readings were recorded at initial and subsequently at 4, 8, 16, and 24 hours.
2.4.4. Titratable Acidity
Titratable acidity for each fermented slurry was determined as described by Nyanzi, (2007) and Molelekoa et al. (2018). A volume of 10 mL (diluted 1:10 using distilled water) of each sample was pipetted and titrated against 0.1 N NaOH to phenolphthalein end- point. Three drops of phenolphthalein indicator were added to each diluted sample solution. The concentration of Lactic acid was determined using the equation described below. All measurements were done in triplicate.
where:
N is the normality of the sodium hydroxide: 0.1 N
V is the volume of sodium hydroxide (mL) used to reach the titration endpoint
ME (milli-equivalent of Lactic acid): 0.09008
2.5. Antimicrobial Activity of LAB Strains Co-Cultured with Pathogenic Strains
Eleven LAB strains were used to determine the antimicrobial activity against indicator pathogenic microorganisms (Escherichia coli ATCC 8739, Staphylococcus aureus ATCC 6538 and Salmonella typhimurium ATCC 14028) during fermentation of sorghum gruels.
The inoculum was prepared by growing LAB strains overnight in 10 mL MRS Broth at 37 ºC. Sorghum flour (80 g/L) was mixed with distilled water which was supplemented with glucose. The mixture was cooked for 15 minutes at 94 ºC and sterilized using autoclave, thereafter the mixture was cooled at room temperature and the mixture was inoculated with 4% (v/v) of a specific LAB strain together with specific pathogenic (106 CFU/mL) strain and the mixture was allowed to ferment for 24 hours in a water bath. Mannitol salt agar was used to enumerate S. aureus, XLD agar for S. typhimurium and MacConkey agar for E. coli. The microbial counts of pathogenic strains were determined at 0, 4, 8 and 24 hours by employing spread plate technique. The plates were incubated for 24 hours at 37 ºC. This was done in triplicate.
2.6. Statistical Analysis
A two-way ANOVA (Analysis if variance) test was applied to the experimental data and the means were compared by Fisher test at P<0.05 using the software Statgraphics Centurion XVI version 16.1.11 (Statpoint, Warrenton, USA).
3. Results
3.1. Amylase Assay of Probiotic Strains
The amylase activity of the nine LAB strains isolated from household fermented sorghum slurries were grown on starch agar plates and starch was used as the sole source of carbon, the plates showing zone of clearing were considered positive for amylase enzyme. Supplementary Table S2 shows the results for amylase assay of the selected LAB strains. Only two strains D7 and D12 tested positive for amylase enzyme as compared to other strains.
Figure 1 shows the effect of amylolytic lactic bacteria on viscosity of sorghum porridges. The two tested Lactobacillus strains D7 and D12 were able to decrease the viscosity of the sorghum porridges during fermentation over 24 hours of incubation as shown in Figure S1 (refer to supplementary material). Strain D7 decreased the viscosity of fermented sorghum porridges from 2854 to 1209 cP after 24 hours and strain D12 decreased the viscosity from 2812 to 940 cP as compared paired to controls (K20 and T12). Strain D12 was the best strain in reducing the viscosity of the sorghum porridge with 940 cP after 24 hours of incubation as compared to strain D7. This implies that the effect of amylolytic Lactobacillus strains on viscosity could be ascribed to direct effects of amylase enzyme from amylolytic activity of LAB from the fermentative liquification of sorghum porridges.
The amylase gene was further confirmed through PCR amplification for Strains D7 and D17 which were successfully amplified. The results of PCR amplicons for the detected gene are shown in supplementary Figure S2.
3.2. Population Levels of Lactobacillus Strains during Fermentation in the Absence and Presence of Glucose
The results of two individual fermentation media, one containing glucose and the other without, are presented in supplementary material (Table S3 and Table S4). In the presence of sugar, all Lactobacillus strains exhibited strong growth, sustaining colony counts between 7 and 8 log10 cfu/mL following an 8-hour incubation period. Specifically, strains K20, and D7 which had shown colony counts within the range of 8.2 to 8.01 log10 cfu/mL. On the other hand, strains D12, and T12 exhibited mean colony counts within the range of 7.5 to 7.6 log10 cfu/mL. Significantly higher colony counts were observed at the end of the 24-hour fermentation period. Notably, the following strains exhibited the highest colony counts, ranging from 8.2 to 8.8 log10 cfu/mL: K20, T12, and D7.
Interestingly, the absence of glucose did not significantly impact the growth of the tested Lactobacillus strains (refer to supplementary Table S 2). At the 8-hour incubation period, strains K20, and T12 displayed the highest mean colony counts, ranging from 8.5 to 8.6 log10 cfu/mL. They were followed by strains D7, and D12 which had mean colony counts ranging from 7.2 to 7.4 log10 cfu/mL. After 24 hours of incubation, all the strains had the exhibited highest colony count ranging from 8.2 to 8.6 log10 cfu/mL. Remarkably, as illustrated in Figure S1, virtual liquefaction observations were made on Lactobacillus Strains D7 and D12 at the end of the fermentation, as they successfully liquefied the fermentation medium without addition of glucose.
3.3. pH
Figure 2 illustrates the pH of the fermented sorghum medium in the absence of 10% glucose by selected LAB strains. It was evident that some strains failed to decrease the pH of the fermentation medium to the required level of pH 3 in the absence glucose. At a 4-hour interval there was already a slight decrease in pH for the tested strains. D12 had managed to decrease its pH to 3.93 as compared to other strains (D7, K20 and T12) which had a pH ranging from 4.0 - 4.4. At 8-hour incubation strain D12 further reduced its pH to 3.7, which was followed by D7, which decreased its pH from 4.1 to 3.9 and the remaining strains had minimal pH reduction. The two Lactobacillus strains D7 and D12 continued further to reduce the pH to 3.5 for both strains at the end of fermentation, followed by T12, with a pH 3.9. Strain K20 failed to reduce the pH of the fermented sorghum to below pH 4.
Figure 3 also illustrates the pH levels of Lactobacillus strains at various time intervals in the presence of added glucose. The addition of glucose to the fermentation medium provided a competitive advantage to the selected Lactobacillus strains. This advantage was particularly noticeable at the 4-hour mark, as all the strains (D12, D7, K20 and T12) had successfully reduced the pH below 4, the pH of the samples were in a range of 3.88 to 3.74. At the 8-hour period, all the Probiotic strains continued to further decrease the pH. By the end of the 24-hour fermentation period, all the strains had achieved a pH lower than 4.
3.4. Titratable Acidity
The lactic acid concentration (% w/v) for both fermentation batches, one with the addition of glucose and one without, is illustrated in Table S5 (supplementary). In this investigation, the fermentation batches that were inoculated with the Lactobacillus strains (K20, T12, D12 and D7) in the absence of glucose exhibited lactic acid concentrations ranging from 0.18% to 0.48% after a 24-hour period. Notably, the strains that demonstrated the highest production of lactic acid were D12 and D7, with lactic acid concentrations of 0.48% and 0.42%, respectively. Followed by strains K20 and T12 with lactic acid concentrations of 0.18 % and 0.24 %, respectively.
In contrast, when glucose was introduced, all the Lactobacillus strains demonstrated a competitive advantage, resulting in significantly higher lactic acid concentrations in comparison to fermentations without glucose. The strain that produced the highest lactic acid was T12, and D7, which had a percentage range of 0.67%, and 0.68%, respectively and this was followed by strain K20 and D12 which had a lactic acid percentage of 0.44 % and 0.55 %, respectively. Strains with the lowest lactic acid production included K20, J20, J22, and V, with concentrations ranging from 0.41% to 0.44%. The results suggest that the addition of glucose significantly increased lactic acid production compared to the batch without glucose. However, in the absence of glucose, strains (T12 and K20) faced challenges in achieving high lactic acid levels, with the notable exceptions of strains D7 and D12, which might contain the amylase gene.
3.5. Antibacterial Activity of LAB Strains against Pathogenic Strains in Sorghum Gruel
The inhibition results of the three pathogenic strains during the co-culturing assay with LAB are represented in Figure 4, Figure 5 and Figure 6. E. coli was inhibited by all the tested LAB strains, it was noted that the growth of the E. coli strain co-cultured with test LAB strains in sorghum flour was below the control experiment from 12 to 24 hours of incubation (Figure 4) meaning that there was a suppression of the E. coli strain. Total inhibition was observed with strains (K20, D12, and D7) after 24 hours of incubation as compared to strain T12 which inhibited E. coli growth from 7.6 Log CFU/mL to 3.14 Log CFU/mL.
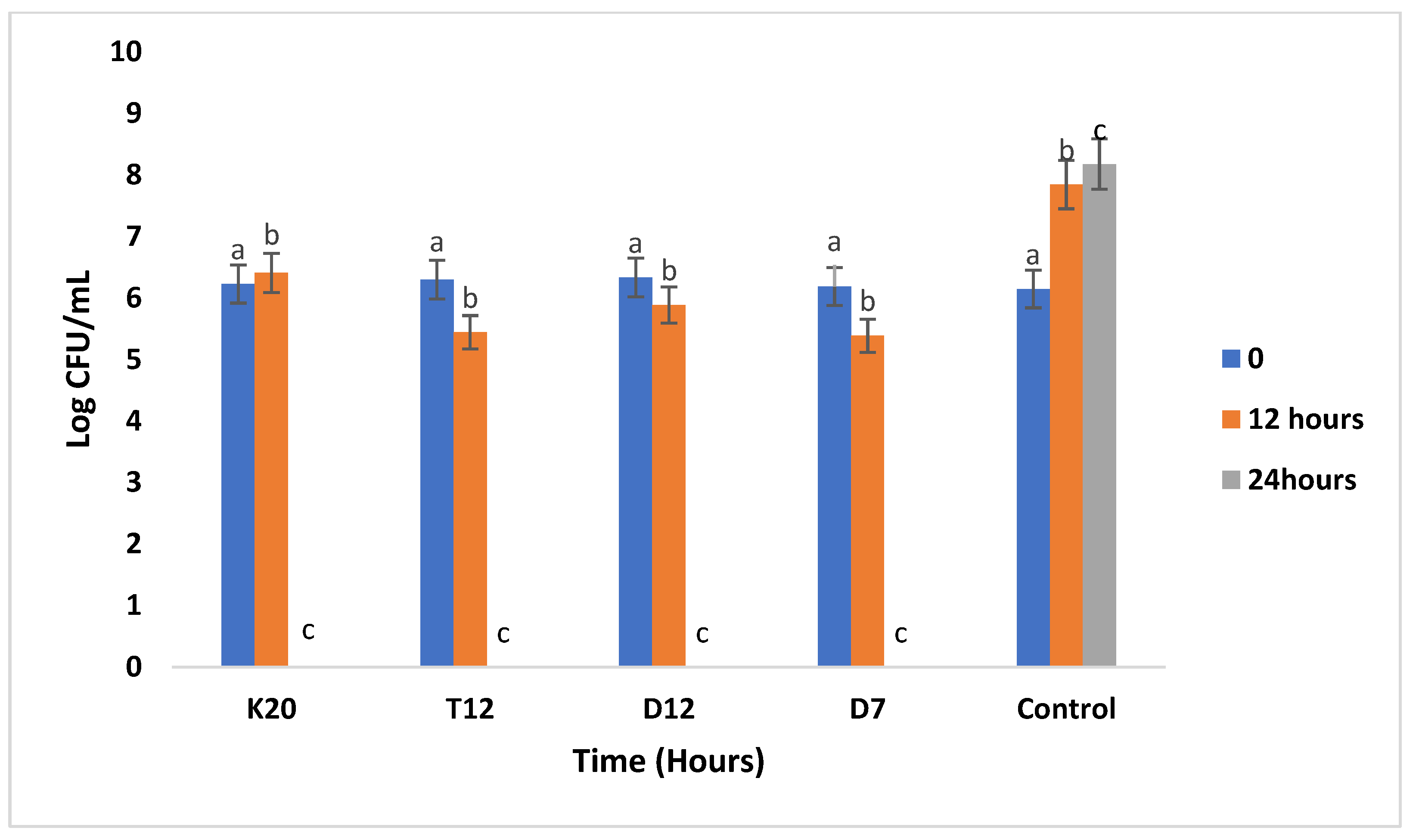
Figure 5 shows the inhibition pattern of S. aureus growth against individual LAB strains. After 12 hours of incubation there was slight inhibition of S. aureus by LAB strains (T12, D12, and D7). However, after 24 hours total inhibition was observed from the strains as compared to strain T12 which reduced the S. aureus growth from 6.6 to 2.3 Log CFU/mL.
Figure 6 below illustrates the antibacterial activity of LAB strains against S. typhimurium. All the tested LAB strains were able to inhibit S. typhimurium after 24 hours of incubation. Total inhibition of the pathogen was observed with LAB strains K20, T12, D12, and D7.
4. Discussion
Lactic acid fermentation remains the preferred method for food processing due to its ability to preserve foods, enhance shelf life, and improve flavor [16]. Cereals fermented with LAB have been proven as a good vehicle for delivering beneficial bacteria. The applicability of probiotic strains in functional food development requires to survive processing conditions and low pH derived from lactic acid production,[17,18]. In this study sorghum was chosen as a vehicle for the selected probiotic strains and as a climate smart crop and is also a cheaper commodity for many Southern African households. The selected 4 probiotics were subjected to controlled fermentation to assess amylase activity, microbial counts, pH, and lactic acid levels in the absence and presence of glucose. Furthermore, the selected strains were also tested for inhibitory activity against pathogenic bacteria.
There were differences between the two fermentation batches with regards to pH values which made it clear that in the presence of sugar the tested Lactobacillus strains were all able to acidify the fermentation medium to pH levels below 4. This was attributed to the readily available sugar (glucose) which can be easily used by Lactobacillus strains in the fermentation medium because of the observed accelerated growth. In the absence of glucose, it was seen that most of the Lactobacillus strains tested were struggling to reduce the pH to below 4. Only three strains D12, D7 and T12 were able to reduce their pH to below 3. Strains of D12 and D7 had the lowest pH of 3.5 as compared to T12 with a pH of 3.9. It could be possible that the amylase gene detected in our earlier study from the two strains, D12 and D7 had a direct role during fermentation by breaking down starch into simple sugars which can be easily utilized by LAB [19]. Hence, the two strains D12 and D7 were able to reduce the pH to below 4 after 8 hours of incubation as compared to other strains. Detection of amylase activity in LAB has been demonstrated by several researchers ([20,21]. The amylase enzyme in LAB has been reported to play a direct role on starch molecules by converting starch into simpler monomers which aid liquification of starch-based products [22]. Addition of amylase producing LAB in fermentation of cereal based products will aid in increasing the fluidity of porridges and increase energy density of cereals such as maize, sorghum and millets for infants. This study is in line with the work conducted by [22], on the amylolytic effect of LAB in cereal based fermentation.
Furthermore, a decrease in pH is a result of Lactobacillus strains being able to convert a simple sugar (glucose) into lactic acid (C6H12O6 → 2C3H6O3), which resulted in a low pH of the fermentation medium. The result of the present study is in agreement with the work conducted by [23,24], who reported a significant pH decrease in the fermentation of sorghum and maize sorghum flours. [24] also reported production lactic acid and other organic acids which included acetic acid, formic acid, and succinic acid in the fermentation of sorghum flour by Lactobacillus species. A reduction in pH and increase in LAB followed a trend reported by [25]. Moreover, a study conducted by [26], has demonstrated that the addition of glucose to the fermentation medium has a direct link to higher organic acid production which is in line with our current study. Lactic acid values recorded in this study were in line with other studies [27,28].
Taking into consideration the addition of sugar on product quality and development plays a significant role due on the growth and metabolic activity of the Lactobacillus strains and it may yield products with better quality attributes such as improved taste, texture, and shelf life. However, in regions where sugar is expensive and difficult to procure. It becomes crucial to explore alternatives such as using amylolytic Lactobacillus strains that can break complex starches in simpler sugar which can used by Lactobacillus strain to produces similar attributes as in fermentation conducted sugar with addition of sugar. In the current study Lactobacillus strain D12 and D7 performed better in fermentation without added sugar and is in line with a study conducted by [19] who reported higher lactic acid production using amylolytic Limosilactobacillus fermentum 04BBA19. Furthermore amylolytic Lactobacillus have been reported by other studies from different tropical starchy fermented foods such as cassava and sweet potato or grains as maize [29]
Antibacterial activity of 4 probiotic strains (K20, D7, D12 and, 12) were tested on clinically important food pathogens namely E. coli ATCC 8739, S. aureus ATCC 6538 and S. typhimurium ATCC 14028 by co-culturing with the selected probiotic strains in the fermentation medium. The LAB strains that were able to completely inhibit all the three indicator pathogens were D12, and D7. Strains K20 only inhibited S. typhimurium and E. coli completely, strain T12 (S. typhimurium and S. aureus). The inhibition of the three pathogens is attributed to the production of fermentation byproducts such as lactic acid, acetic acid, and small portions of propionic acid. These metabolites synergistically work to inhibit the growth of many target pathogens by making the environment unfavorable for most pathogens by reducing the pH to acidic levels [26,30,31].
Escherichia coli is classified as gram-negative bacteria within the Enterobacteriaceae family, that colonizes the human gastrointestinal tract but has the potential to induce intestinal or extraintestinal infections [32]. These infections cause severe invasive diseases, including bacteremia and sepsis. Notably, E. coli stands out as the most common cause of bacteremia in both low and high-income settings, surpassing other pathogens like Staphylococcus aureus and Streptococcus pneumoniae. Additionally, E. coli is a principal causative agent of neonatal meningitis [15,33]. Therefore, consumption of fermented sorghum beverage may also aid in therapeutic management of various clinical infections and foodborne diseases attributed to E. coli, S. typhimurium and S. aureus.
It can be deduced from the observations made in Figure 4, Figure 5, and Figure 6 that the use of one strain in the preparation of fermented porridge/products will not be effective in inhibiting enteropathogens [34]. The use of combing probiotic strains for the preparation of fermented beverages would help to prevent disease causing foodborne pathogens such as E. coli, S. aureus and S. typhimurium. Therefore, combing strains that showed total inhibition with the strains that showed little to moderate inhibition will ensure the safety of fermented sorghum beverage. The findings of the current study are in line with the work conducted by [34,35]. The study conducted by [35], reported antibacterial activity of Lb. plantarum which completely inhibited the E. coli D1288 within 8 hours of incubation during the fermentation of Ghanaian maize dough. Several studies conducted on fermented cereal-based porridge (maize dough and motoho and ting) have demonstrated antimicrobial action against a variety of diarrhoeal pathogens [10,36].
5. Conclusions
This study focused on assessing the performance of probiotic strains in a sorghum-based medium. During fermentation, all selected Lactobacillus strains demonstrated robust growth, maintaining levels above 6 log10 cfu/mL after 24 hours, regardless of the presence or absence of sugar. However, fermenting without the addition of glucose resulted in a limited reduction in pH, ranging from 4.0 to 4.4 at the end of the 24-hour incubation. Notably, strains D7 and D12 with amylolytic activity achieved a faster reduction, reaching pH 3 within 8 to 24 hours. Regarding lactic acid production, strains D7 and D12 exhibited higher levels, measuring 0.42 and 0.48 units, respectively, as determined by titratable acidity. With regards to antibacterial activity, strains D12 and D7 successfully inhibited E. coli, S. aureus and S. typhimurium to undetectable levels after 24 hours, while strains T12 and K20 did not inhibit all the three indicator pathogens to undetectable levels after the same duration. These findings suggest the potential for employing combined probiotic strains in the future development of sorghum-based beverages for impoverished communities who cannot afford probiotic supplements.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Figure S1: title; Table S1: title; Video S1: title.
Author Contributions
R.S.M. and T.M.L. conceived and designed the experiments; R.S.M. performed the experiments; R.S.M. and T.M.L. analyzed the data; R.S.M prepared the first manuscript draft. T.M.L. reviewed and edited the final manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
All relevant data generated in this study are available in this manuscript and a supplemented data.
Acknowledgments
We wish to express our appreciation to NRF and Tshwane University of Technology (TUT) for funding this research.
Conflicts of Interest
The authors declare that there are no conflicts of interest.
References
- M. C. Setta, A. Matemu, and E. R. Mbega. Potential of probiotics from fermented cereal-based beverages in improving health of poor people in Africa. J. Food Sci. Technol. 2020, 57, 3935–3946. [Google Scholar] [CrossRef] [PubMed]
- R. Nyanzi, P. J. Jooste, and E. M. Buys. Invited review: Probiotic yogurt quality criteria, regulatory framework, clinical evidence, and analytical aspects. J. Dairy Sci. 2021, 104, 1–19. [Google Scholar] [CrossRef] [PubMed]
- S. J. Rhee, J. E. Lee, and C. H. Lee. Importance of lactic acid bacteria in Asian fermented foods. Microb. Cell Fact. 2011, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- V. Jhamb and P. Swaminathan. Role and importance of lactic acid bacteria in different Indian fermented foods. Biologia (Bratisl). 2023, 78, 3609–3623. [Google Scholar] [CrossRef]
- E. Mani-López, N. Ramírez-Corona, and A. López-Malo. Advances in probiotic incorporation into cereal-based baked foods: Strategies, viability, and effects–A review. Appl. Food Res. 2023, 3, 100330. [Google Scholar] [CrossRef]
- H. Tang, W. Huang, and Y. F. Yao. The metabolites of lactic acid bacteria: classification, biosynthesis and modulation of gut microbiota. Microb. Cell 2023, 10, 49–62. [Google Scholar] [CrossRef] [PubMed]
- B. H. Lunelli, R. R. Andrade, D. I. P. Atala, M. R. W. MacIel, F. M. Filho, and R. M. I. Filho. Production of lactic acid from sucrose: Strain selection, fermentation, and kinetic modeling. Appl. Biochem. Biotechnol. 2010, 161, 227–237. [Google Scholar] [CrossRef] [PubMed]
- R. P. John, K. M. Nampoothiri, and A. Pandey. Fermentative production of lactic acid from biomass: An overview on process developments and future perspectives. Appl. Microbiol. Biotechnol. 2007, 74, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Y. J. Wee, J. N. Kim, and H. W. Ryu. Biotechnological production of lactic acid and its recent applications. Food Technol. Biotechnol. 2006, 44, 163–172. [Google Scholar]
- E. Madoroba, E. T. Steenkamp, J. Theron, I. Scheirlinck, T. E. Cloete, and G. Huys. Diversity and dynamics of bacterial populations during spontaneous sorghum fermentations used to produce ting, a South African food. Syst. Appl. Microbiol. 2011, 34, 227–234. [Google Scholar] [CrossRef] [PubMed]
- B. Szala, Z. Paluszak, and I. Motyl. Antagonistic Effect of Lactic Acid Bacteria on Salmonella Senftenberg in Mixed Cultures. 2012, 21, 1399–1403. [Google Scholar]
- J. K. Mugula, S. A. M. Nnko, J. A. Narvhus, and T. Sørhaug. Microbiological and fermentation characteristics of togwa, a Tanzanian fermented food. Int. J. Food Microbiol. 2003, 80, 187–199. [Google Scholar] [CrossRef] [PubMed]
- 13. O. A. Adebo. African sorghum-based fermented foods: Past, current and future prospects. Nutrients 2020, 12. [CrossRef] [PubMed]
- M. Fidanza, P. Panigrahi, and T. R. Kollmann. Lactiplantibacillus plantarum–Nomad and Ideal Probiotic. Front. Microbiol. 2021, 12, 1–13. [Google Scholar] [CrossRef] [PubMed]
- S. M. Rapoo, P. 15. S. M. Rapoo, P. Budeli, and M. L. Thaoge. Recovery of Potential Starter Cultures and Probiotics from Fermented Sorghum (Ting) Slurries. Microorganisms 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- K. Y. Kim, O. D. Ekpe, H. Lee, and J. Oh. Perfluoroalkyl substances and pharmaceuticals removal in full-scale drinking water treatment plants. J. Hazard. Mater. 2020, 400, 123235. [Google Scholar] [CrossRef]
- X. Zhang et al.. Saudi Journal of Biological Sciences Probiotic characteristics of Lactobacillus strains isolated from cheese and their antibacterial properties against gastrointestinal tract pathogens. Saudi J. Biol. Sci. 2020, 27, 3505–3513. [Google Scholar] [CrossRef] [PubMed]
- G. Xia. Characteristics of Probiotic Preparations and Their Applications. 2022.
- 19. B. T. Fossi and F. Tavea. Application of Amylolytic Lactobacillus fermentum 04BBA19 in Fermentation for Simultaneous Production of Thermostable a -Amylase and Lactic Acid. Lact. Acid Bact. – R D Food, Heal. Livest. Purp. 2013, 633–58.
- G. Reddy, M. Altaf, B. J. Naveena, M. Venkateshwar, and E. V. Kumar. Amylolytic bacterial lactic acid fermentation - A review. Biotechnol. Adv. 2008, 26, 22–34. [Google Scholar] [CrossRef] [PubMed]
- T. Padmavathi, R. Bhargavi, P. R. Priyanka, N. R. Niranjan, and P. V. Pavitra. Screening of potential probiotic lactic acid bacteria and production of amylase and its partial purification. J. Genet. Eng. Biotechnol. 2018, 16, 357–362. [Google Scholar] [CrossRef] [PubMed]
- L. Ferry, J. Hort, J. R. Mitchell, S. Lagarrigue, and B. V. Pamies. Effect of amylase activity on starch paste viscosity and its implications for flavor perception. J. Texture Stud. 2004, 35, 511–524. [Google Scholar] [CrossRef]
- N. M. K. Yousif et al.. Diversity of lactic acid bacteria from Hussuwa, a traditional African fermented sorghum food. Food Microbiol. 2010, 27, 757–768. [Google Scholar] [CrossRef] [PubMed]
- F. S. Ibrahim, E. E. Babiker, N. E. Yousif, and A. H. El Tinay. Effect of fermentation on biochemical and sensory characteristics of sorghum flour supplemented with whey protein. Food Chem. 2005, 92, 285–292. [Google Scholar] [CrossRef]
- E. Nwachukwu, O. K. Achi, and I. O. Ijeoma. Lactic acid bacteria in fermentation of cereals for the production of indigenous Nigerian foods. African J. Food Sci. Technol. 2010, 1, 21–026. [Google Scholar]
- T. B. J. Molelekoa, T. Regnier, L. S. Da Silva, and W. A. Augustyn. Potential of marula (Sclerocarya birrea subsp. caffra) waste for the production of vinegar through surface and submerged fermentation. S. Afr. J. Sci. 2018, 114, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hassani, M. Zarnkow, and T. Becker. Optimisation of fermentation conditions for probiotication of sorghum wort by Lactobacillus acidophilus LA5. Int. J. Food Sci. Technol. 2015, 50, 2271–2279. [Google Scholar] [CrossRef]
- M. Salari, S. H. Razavi, and S. M. T. Gharibzahedi. Characterising the synbiotic beverages based on barley and malt flours fermented by Lactobacillus delbrueckii and paracasei strains. Qual. Assur. Saf. Crop. Foods 2015, 7, 355–361. [Google Scholar] [CrossRef]
- L. T. Songré-Ouattara, C. Mouquet-Rivier, C. Icard-Vernière, C. Humblot, B. Diawara, and J. P. Guyot. Enzyme activities of lactic acid bacteria from a pearl millet fermented gruel (ben-saalga) of functional interest in nutrition. Int. J. Food Microbiol. 2008, 128, 395–400. [Google Scholar] [CrossRef] [PubMed]
- F. Dejene, B. Regasa Dadi, and D. Tadesse. In Vitro Antagonistic Effect of Lactic Acid Bacteria Isolated from Fermented Beverage and Finfish on Pathogenic and Foodborne Pathogenic Microorganism in Ethiopia. Int. J. Microbiol. 2021, 2021. [Google Scholar] [CrossRef] [PubMed]
- J. A. Vázquez, M. P. González, and M. A. Murado. Effects of lactic acid bacteria cultures on pathogenic microbiota from fish. Aquaculture 2005, 245, 149–161. [Google Scholar] [CrossRef]
- Z. Akhondali, M. Dianat, and M. Radan. The antimicrobial activity of probiotic bacteria Escherichia coli isolated from different natural sources against hemorrhagic E. coli O157:H7. Electron Physician 2015, 7, 971–976. [Google Scholar]
- C. de M. Santos, F. F. Santos, R. M. Silva, and T. A. T. Gomes. Diversity of Hybrid- and Hetero-Pathogenic Escherichia coli and Their Potential Implication in More Severe Diseases. Front. Cell. Infect. Microbiol. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- R. Nyanzi, M. D. Awouafack, P. Steenkamp, P. J. Jooste, and J. N. Eloff. Anticandidal activity of cell extracts from 13 probiotic Lactobacillus strains and characterisation of lactic acid and a novel fatty acid derivative from one strain. Food Chem. 2014, 164, 470–475. [CrossRef] [PubMed]
- E. S. Mante, E. Sakyi-Dawson, and W. K. Amoa-Awua. Antimicrobial interactions of microbial species involved in the fermentation of cassava dough into agbelima with particular reference to the inhibitory effect of lactic acid bacteria on enteric pathogens. Int. J. Food Microbiol. 2003, 89, 41–50. [CrossRef] [PubMed]
- L. C. Peyer, E. Zannini, and E. K. Arendt. Lactic acid bacteria as sensory biomodulators for fermented cereal-based beverages. Trends Food Sci. Technol. 2016, 54, 17–25. [CrossRef]
Figure 1.
Viscosity analysis of fermented sorghum porridges with strains K20, T12, D7 and D12 over 24 hours. Bars with the same letter are not significantly (P>0.05) different. Values reported are means of at least three independent measures.
Figure 1.
Viscosity analysis of fermented sorghum porridges with strains K20, T12, D7 and D12 over 24 hours. Bars with the same letter are not significantly (P>0.05) different. Values reported are means of at least three independent measures.
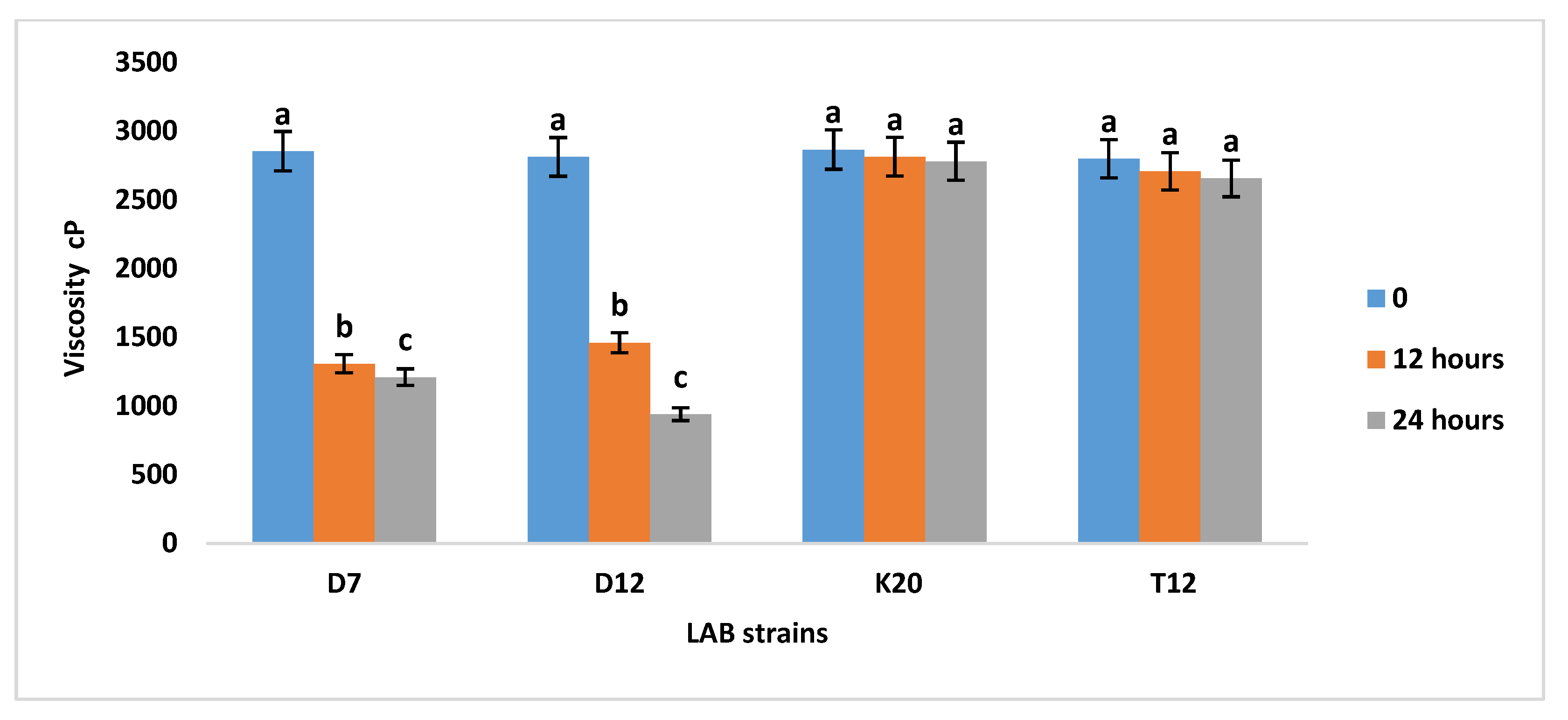
Figure 2.
The pH for sorghum gruel fermented with LAB strains in the absence of 10% glucose. pH levels were monitored hourly throughout the fermentation process. Values reported are means of at least three independent measures. Bars with the same letter are not significantly (P>0.05) different. values reported are means of at least three independent measures.
Figure 2.
The pH for sorghum gruel fermented with LAB strains in the absence of 10% glucose. pH levels were monitored hourly throughout the fermentation process. Values reported are means of at least three independent measures. Bars with the same letter are not significantly (P>0.05) different. values reported are means of at least three independent measures.
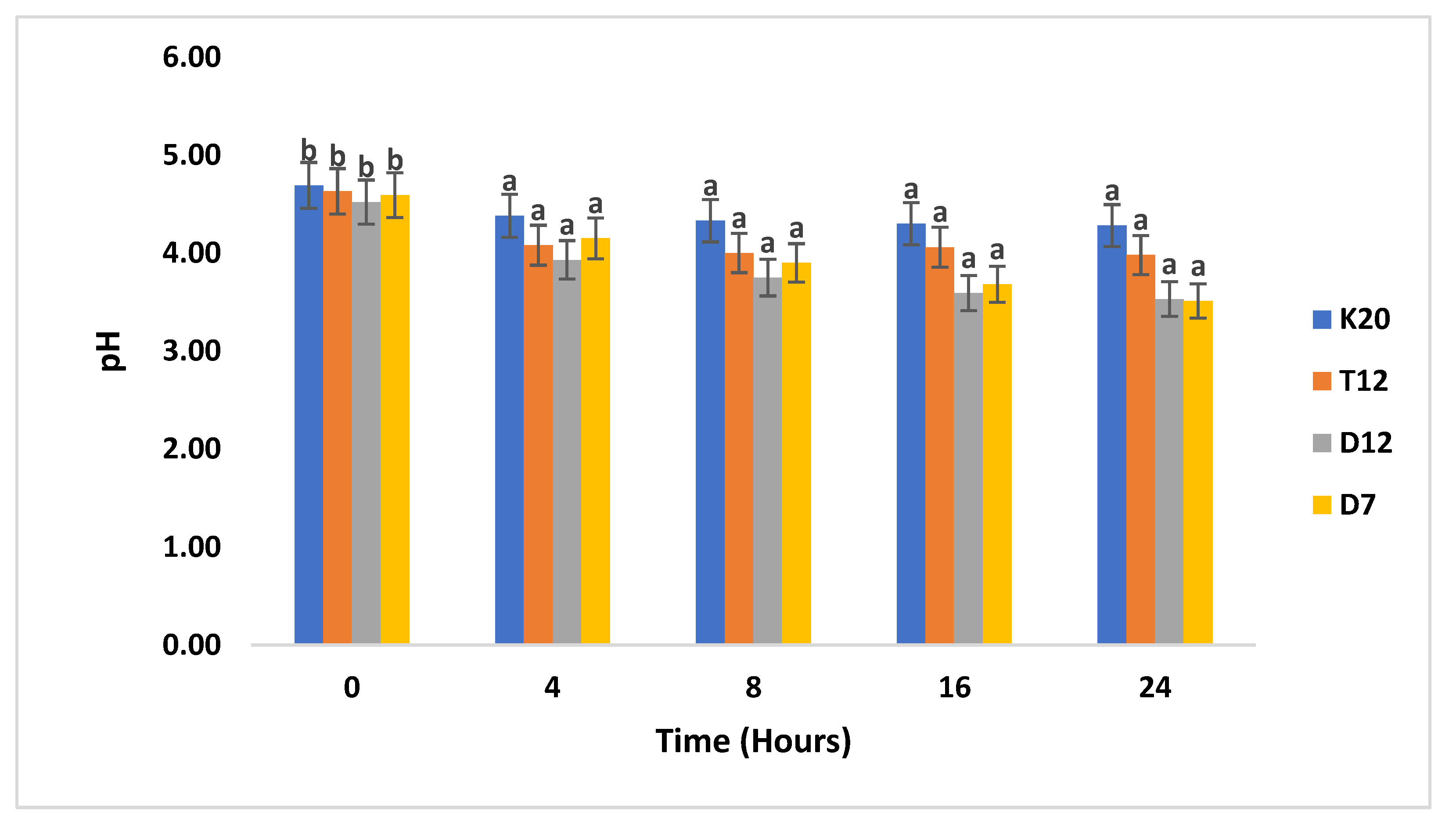
Figure 3.
The pH for sorghum gruels fermented with LAB strains in the presence of 10% glucose. pH levels were monitored hourly throughout the fermentation process. Values reported are means of at least three independent measures. Bars with the same letter are not significantly (P>0.05) different. values reported are means of at least three independent measures.
Figure 3.
The pH for sorghum gruels fermented with LAB strains in the presence of 10% glucose. pH levels were monitored hourly throughout the fermentation process. Values reported are means of at least three independent measures. Bars with the same letter are not significantly (P>0.05) different. values reported are means of at least three independent measures.
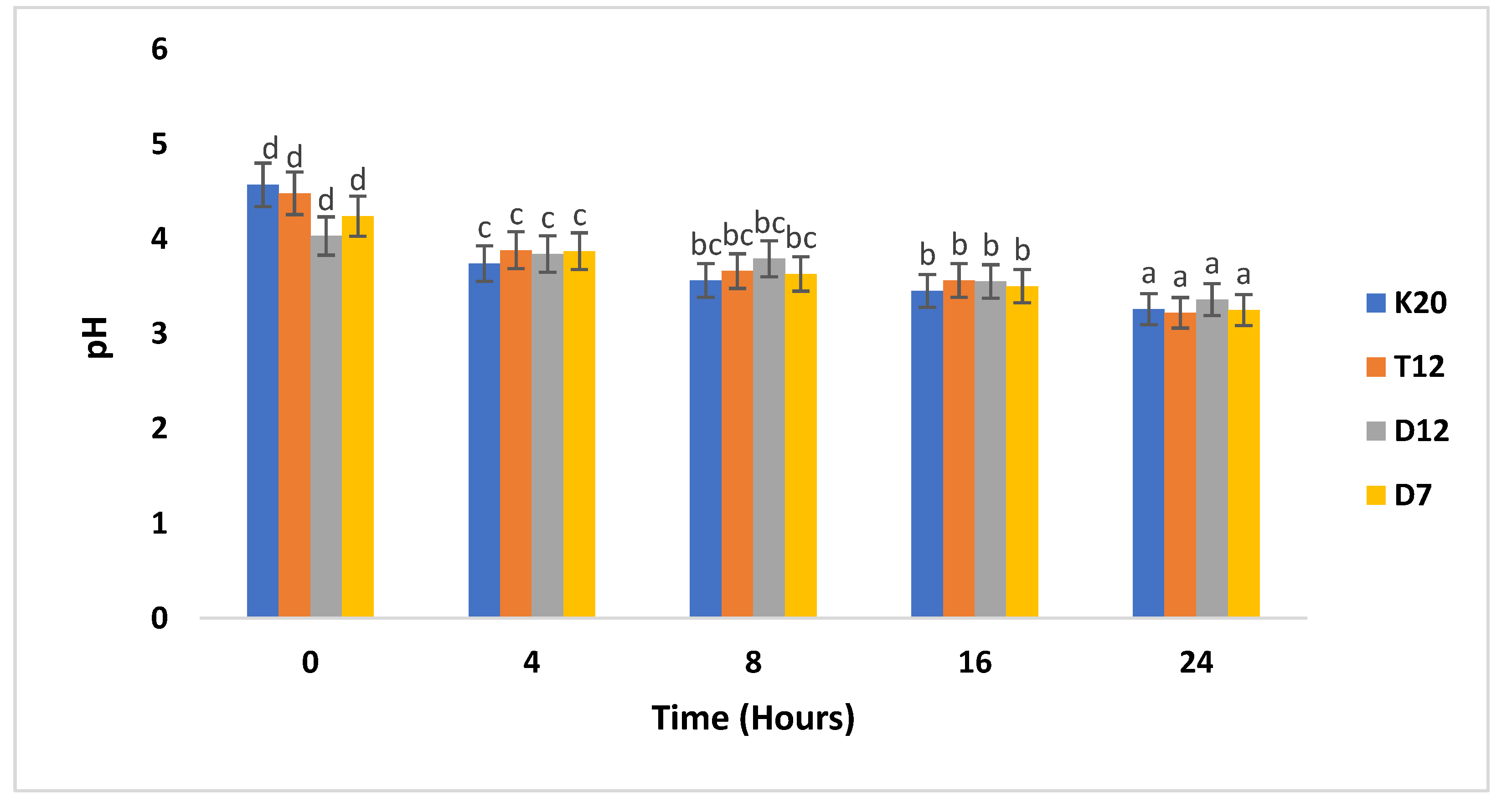
Figure 4.
The pH for sorghum gruels fermented with LAB strains in the presence of 10% glucose. pH levels were monitored hourly throughout the fermentation process. Values reported are means of at least three independent measures. Bars with the same letter are not significantly (P>0.05) different. values reported are means of at least three independent measures.
Figure 4.
The pH for sorghum gruels fermented with LAB strains in the presence of 10% glucose. pH levels were monitored hourly throughout the fermentation process. Values reported are means of at least three independent measures. Bars with the same letter are not significantly (P>0.05) different. values reported are means of at least three independent measures.
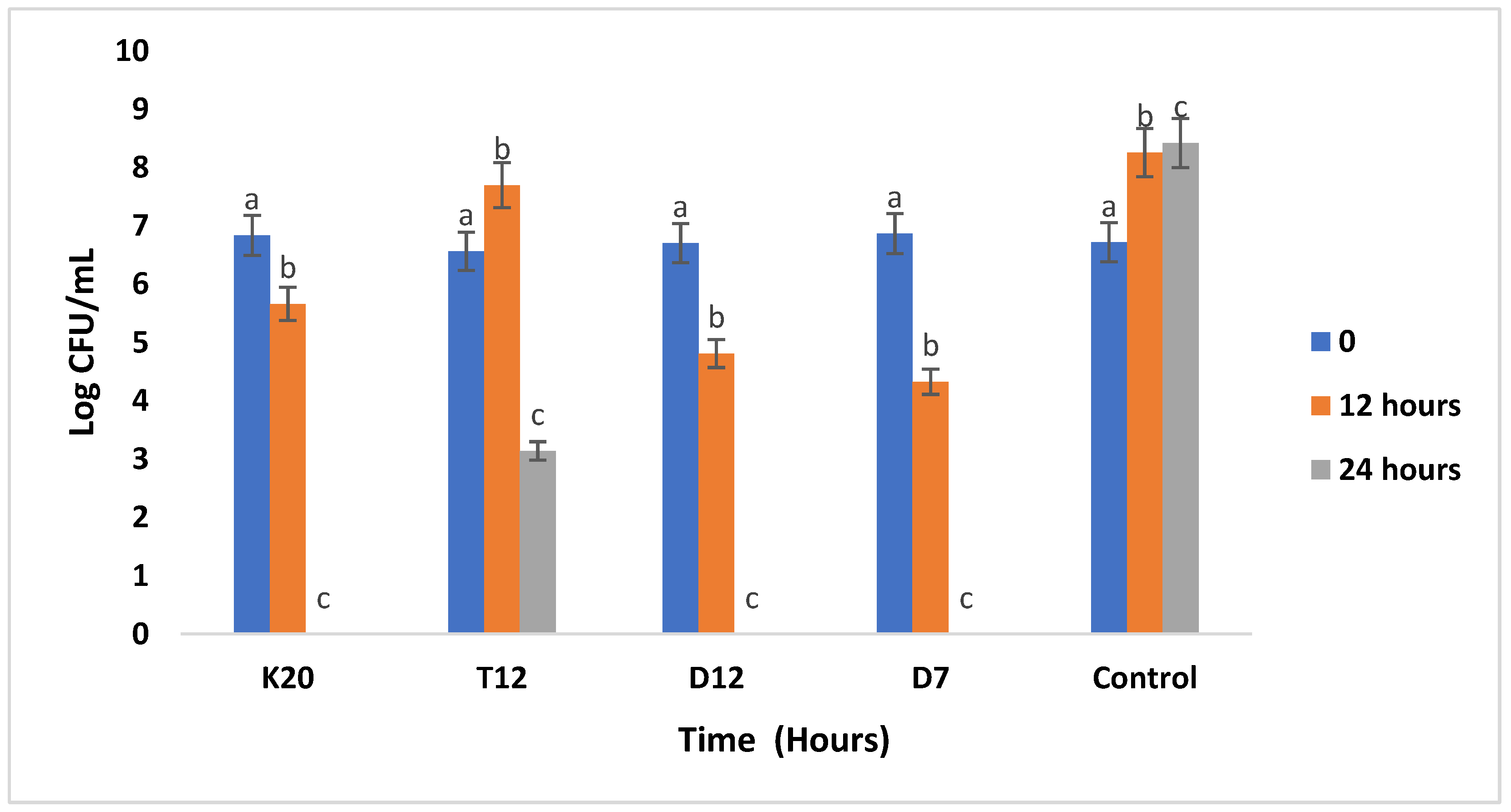
Figure 5.
Antibacterial activity of the 4 tested LAB strains against Staphylococcus aureus ATCC 6538 in sorghum gruel fermentation. Bars with the same letter are not significantly different (P>0.05). Values reported are by means of three independent measures.
Figure 5.
Antibacterial activity of the 4 tested LAB strains against Staphylococcus aureus ATCC 6538 in sorghum gruel fermentation. Bars with the same letter are not significantly different (P>0.05). Values reported are by means of three independent measures.
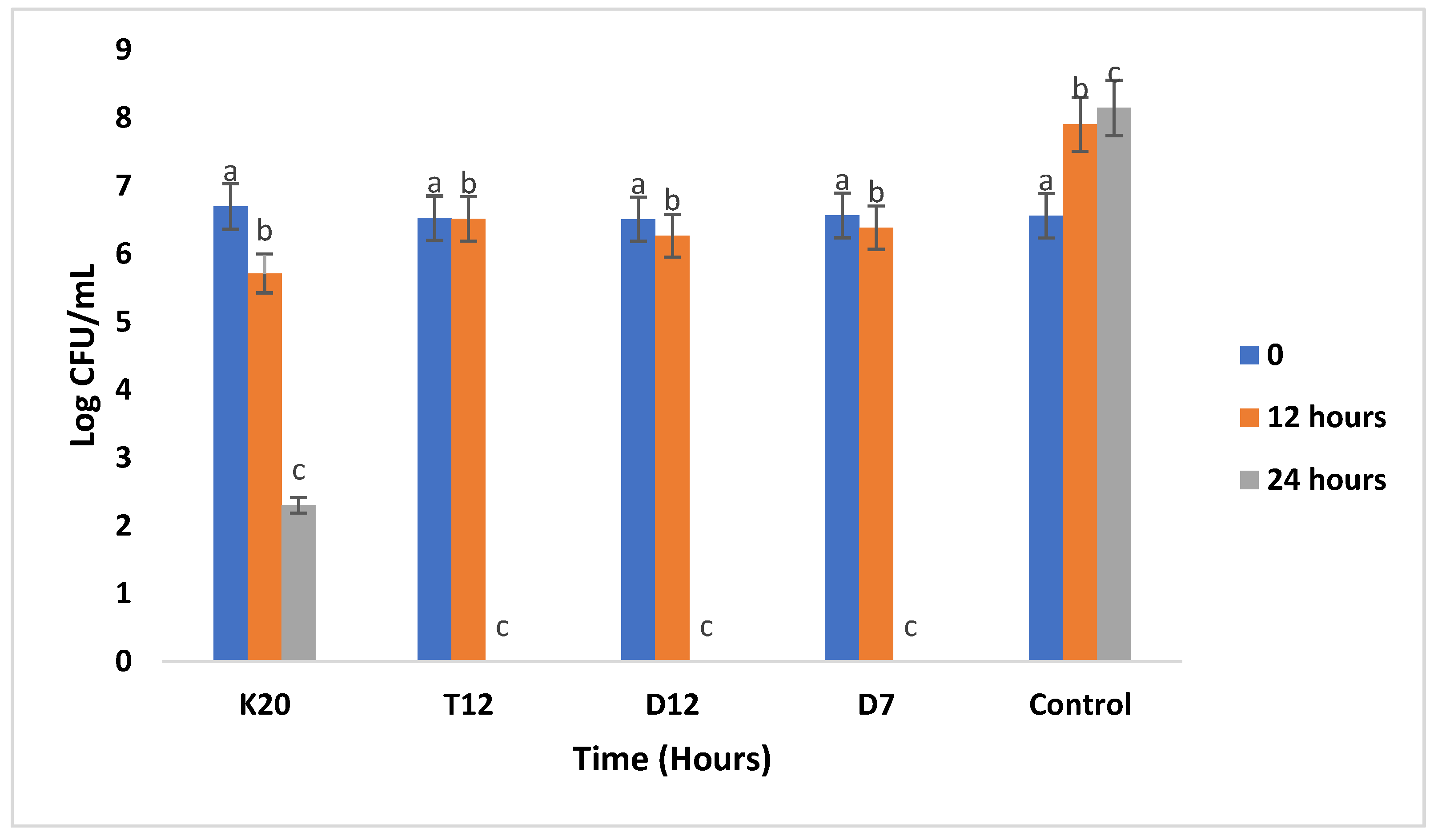
Figure 6.
Antibacterial activity of the 4 tested LAB strains against E.coli ATCC 8739 in sorghum gruel fermentation. Bars with the same letter are not significantly (P>0.05) different. Values reported are means of at least three independent measures.
Figure 6.
Antibacterial activity of the 4 tested LAB strains against E.coli ATCC 8739 in sorghum gruel fermentation. Bars with the same letter are not significantly (P>0.05) different. Values reported are means of at least three independent measures.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



