Submitted:
04 June 2024
Posted:
04 June 2024
You are already at the latest version
Abstract
Small peptides (SPs), ranging from 5 to 100 amino acids, play integral roles in plants due to their diverse functions. Despite their low abundance and small molecular weight, SPs intricately regulate critical aspects of plant life, including cell division, growth, differentiation, flowering, fruiting, maturation, and stress responses. As vital mediators of intercellular signaling, SPs have garnered significant attention in plant biology research. This comprehensive review delves into SPs' structure, classification, and identification, providing a detailed understanding of their significance. Additionally, we summarize recent findings on the biological functions and signaling pathways of prominent SPs that regulate plant growth and development. The review also offers a forward-looking perspective on future research directions in peptide signaling pathways.
Keywords:
Small peptides
; Peptide hormones
; Small secreted peptides
; Peptide-receptor module
; Plant growth and development
1. Introduction
Small peptides (SPs), consisting of 5 to 100 amino acids, are widely distributed in plants and animals and function as critical signaling molecules regulating growth, development, and stress responses (Hellens et al., 2016; Ong et al., 2022). Most SPs are encoded by precursor genes, with typical peptide precursors having conserved family-specific sequences at the C-terminus (Olsson et al., 2019). These precursors undergo proteolytic cleavage and post-translational modifications to yield mature, functional peptides (Yu et al., 2019b). Mature SPs are recognized by nearby cell membrane surface receptors, such as receptor kinases and receptor-like kinase families. Alternatively, SPs may undergo long-distance transport to bind to the extracellular domains of corresponding receptors (Takahashi et al., 2018; Datta et al., 2024). This binding event activates the intracellular protein kinase domains of the receptors, triggering downstream signaling cascades, such as MAPK signaling and transcription factors (Fletcher, 2020; Zhou et al., 2022; Yang et al., 2024).
SPs are essential regulators of plant growth and development, influencing cell proliferation in stems, roots, leaves, flowers, fruits, and seeds (Song et al., 2016; Zhang et al., 2018; Ogawa-Ohnishi et al., 2022). They also mediate plant resistance to environmental stresses such as drought, salt, pests, and diseases (Reichardt et al., 2020; Zhou et al., 2022; Xiao and Zhou, 2023). Numerous SPs have been identified in various crops beyond model plants like Arabidopsis and rice, with their homologous genes and structures showing high conservation across plant species (Olsson et al., 2019; Jeon et al., 2021).
This review provides an overview of the latest methods for identifying peptides, which involve screening all amino acid sequences and small open reading frames (sORFs) of a species to pinpoint candidate peptides. We then delve into SPs intricately linked to plant growth and development, focusing on peptide families such as phytosulfokine (PSK), plant peptide-containing sulfated tyrosine (PSY), casparian strip integrity factors (CIF), root growth factor (RGF), clavata/embryo surrounding region (CLE), epidermal patterning factor/epidermal patterning factor-like (EPF/EPFL), LURE, rapid alkalinization factor (RALF), C-terminally encoded peptide (CEP), and Inflorescence deficient in abscission (IDA). Using these small peptides as exemplars, the review explores their origins and characteristics, summarizing how they fine-tune plant growth and development by activating various downstream signal transduction pathways. Additionally, it offers a forward-looking perspective on future research directions in peptide signaling pathways, aiming to inspire further exploration and discovery in this dynamic field.
1.1. Classification & Identification of SPs
SPs, known as small molecule signaling peptides, exhibit remarkably conserved homologous gene structures and domains among plants (Ong et al., 2022). The first small plant peptide, systemin, was identified in Solanum lycopersicum and regulates the resistance response to wounds caused by pests and diseases (Ryan and Pearce, 1998). Since then, various plant SPs have been identified and annotated using classical biochemical and molecular methods. Additionally, many predictive tools and bioinformatics approaches for identifying SPs are emerging.
1.2. Classification Based on Origin
Most SPs typically have a molecular weight of around 25 amino acids. They can be categorized into two main groups based on their origin: precursor-derived and non-precursor-derived peptides (Figure 1; Datta et al., 2024). Precursor-derived peptides are processed from precursor proteins, often with a signal peptide at the N-terminus guiding their maturation into functional peptides (Ong et al., 2022; Datta et al., 2024). These peptides can be further classified into functional and non-functional precursors. For example, CAPE (CAP-derived peptide 1) and SUBPEP (Subtilase peptide) are functional precursors involved in immune defense mechanisms, while non-functional precursors lack a defined biological role, typically arising from extended preproproteins (Pearce et al., 2010; Chien et al., 2015; Xu et al., 2024). Non-functional precursors can be categorized into three groups based on their mature peptide characteristics (Figure 1).
1. Peptides incorporating posttranslational modifications (PTMs), such as proline hydroxylation, hydroxyproline arabinosylation, and tyrosine sulfation, which confer biological activity and chemical stability (Royek S et al., 2022). Examples include PAMP-induced Secreted Peptides (PIP), CEP, IDA, and CLE peptides.
2. Cysteine-rich peptides (CRPs) containing structural domains with 2-16 Cys residues influence peptide structure and activity via intramolecular disulfide bond formation (Ong et al., 2022; Tian et al., 2022). The EPF/EPFL peptide family is a notable example of regulating plant stomata development (Kosentka et al., 2019).
3. Non-cysteine-rich/non-PTM peptides play roles in plant defense responses. Examples include systemin (SYS), plant elicitor peptide (PEP), and plant natriuretic peptides (PNP) (Lee et al., 2020).
Nonprecursor-derived peptides are directly translated from sORFs without intermediate propeptides or additional processing steps (Datta et al., 2024). These peptides can be classified into three classes based on the genomic location of the sORF genes (Figure 1): (1) peptides encoded by upstream ORFs on the mRNA 5’ end, (2) peptides encoded within longer non-coding proteins in other transcript, and (3) peptides encoded by sORFs within primary transcripts of microRNAs (pri-miRNAs), In summary, SPs display diverse characteristics and functions in plants. Understanding the origin and classification of SPs provides insights into their regulatory roles in plant growth, development, and stress responses.
1.3. Based on the N-Terminal Sequence
Based on the variation in the N-terminal sequence of the peptide precursor protein, plant SPs can be categorized as non-secretory and secretory peptides (Xu et al., 2023; Datta et al., 2024). Non-secretory peptides can regulate intracellular processes and are released into the extracellular matrix from injured cells to activate plant defense responses directly (Jing et al., 2024). For example, PEP is a typical non-secretory peptide that triggers plant defense responses against pathogen (Zelman et al., 2023). Recent research has also revealed that PEP/REF1 functions as a systemin-independent local wound signal in tomato plants, predominantly orchestrating localized defense mechanisms and regenerative processes in response to injury (Yang et al., 2024). Additionally, it enhances the regenerative capacity of callus tissue (Lu et al., 2018; Yang et al., 2024).
In contrast, secretory peptides are synthesized intracellularly and transported to the extracellular space (Shen et al., 2022; Datta et al., 2024). They exert their effects by traversing the epidermis or xylem, facilitating intercellular signal transduction, and regulating the activities of adjacent cells in the extracellular milieu (Olsson et al., 2019; Shen et al., 2022). Peptides incorporating post-translational modifications (PTMs) and cysteine-rich peptides (CRPs) are classified as secreted peptides. Importantly, PTMs represent the most prevalent type of plant peptides (Xu et al., 2023). The precursor protein transforms into a mature peptide through electrostatic charge, hydrophilicity, and conformation modifications. The mature peptide then binds to receptors, activating downstream signal transduction pathways (Tian et al., 2022; Xu et al., 2023).
1.4. Identification Methods of SPs
Various functional plant SPs have been identified and annotated to regulate multiple aspects of plant growth (Hsiao and Yamada, 2020; Datta et al., 2024). Traditional biochemical and genetic approaches have been used to validate the biological functions of plant peptides. However, these methods are not ideal for high-efficiency peptide identification, often yielding only a few identified peptides (Molisso et al., 2022).
Currently, researchers have integrated novel peptidomics, bioinformatics screening, and biochemical and genetic validation methods for peptide identification, overcoming the high costs and technical difficulties associated with traditional biochemical analysis and circumventing obstacles arising from low gene abundance and redundancy (Wang et al., 2020; Ong et al., 2022). For instance, high-throughput analytical approaches that couple selective enrichment, fractionation, phenotype screening, and mass spectrometry identification provide an established framework for screening plant tissues for biologically relevant small secreted peptides (SSPs) (Wang et al., 2020; Hu et al., 2021). This method begins with selective enrichment strategies to isolate molecules of interest from complex crude extracts, commonly using techniques such as size exclusion ultrafiltration, gel-based separations, solvent extractions, and size exclusion chromatography (Figure 2A; Feng et al., 2023).
Next, further fractionation is performed based on physicochemical properties (e.g., polarity, hydrophobicity, stability, solubility). SSPs can be screened for bioactivity in cell-based or cell-free systems to evaluate functions such as antimicrobial activity or free radical scavenging (Feng et al., 2023). After detecting fractions with relevant bioactivity, high-throughput LC-MS/MS is used to sequence unknown SSPs (Figure 2A; Hu et al., 2021). However, mass spectrometry identification of SSPs faces challenges due to insufficient SSP representation in protein databases and unclear mechanisms of SSP maturation.
A recent bioinformatics approach for peptide identification involves screening for SSPs within the entire proteome of a given plant species. Initially, the focus is on the amino acid sequences of proteins. Proteins containing fewer than 250 amino acids are selected as potential secretory peptides, as they typically possess N-terminal signal peptides. Subsequently, signal peptide prediction is performed on these small peptides, eliminating those lacking N-terminal signal peptides. Since secretory peptides lack transmembrane domains, further prediction of transmembrane domains is employed on the remaining peptides, removing those exhibiting such structures. Finally, peptides with C-terminal K/HDLE sequences, which serve as endoplasmic reticulum docking proteins, are discarded, yielding the secretory peptide sequences for the given species (Figure 2B; Tian et al., 2022; Xu et al., 2023).
Notably, genes encoding SSPs are often overlooked during genome annotation due to their diminutive size (Ong et al., 2022). Therefore, a more comprehensive approach involves screening sORFs to identify SSPs (Figure 2B). This method first identifies non-coding sequences (NCDS) from the chromosomes using software tools like sORF finder. The subsequent steps are similar to the methods described earlier, wherein the presence or absence of an N-terminal signal peptide, the presence of transmembrane domains, and whether the sORF encodes an ER-docking protein are employed as distinguishing SSP characteristics for selecting potential sORFs within the NCDS (Figure 2B). It is worth noting that only a tiny fraction of sORFs can be categorized into known gene families, while most remain unclassifiable. This issue may be attributed to undiscovered peptide families and the lack of comprehensive sORF annotation, which could lead to false positives. Together, this represents a novel approach to plant peptide identification, highlighting the potential of bioinformatics in unraveling SSPs. This method offers a promising direction for future research, utilizing bioinformatics to identify and characterize secretory peptides, thereby advancing our understanding of their roles in plant biology.
1.5. Tyrosine-Sulfated Peptides
Sulfated peptides are plant hormone-like molecules known for their potent activity even at nanomolar concentrations. Four distinct sulfated peptides have been characterized in plants—PSK, PSY, CIF and RGF. These peptides mediate growth and development processes by activating specific signal transduction pathways.
1.6. PSK Peptides
PSK peptides, originating from 80 to 120 amino acid preproproteins, were first identified in asparagus suspension cell cultures as growth promoters (Matsubayashi and Sakagami, 1996; Kutschmar et al., 2008; Ding et al., 2023). They are widely distributed across plants, including angiosperms and gymnosperms, with several members in various species sharing significant sequence similarity and a conserved C-terminal domain (Sauter, 2015; Zhang et al., 2018; Geng et al., 2020; Kou et al., 2020). PSK maturation involves tyrosine sulfation and proteolytic cleavage, facilitated by plant-specific enzymes such as tyrosylprotein sulfotransferases (TPSTs) and subtilisin serine proteases (Komori et al., 2009; Stührwohldt et al., 2021).
PSK peptides play crucial roles in various aspects of plant growth (Kutschmar et al., 2008; Reichardt et al., 2020; Geng et al., 2020). Synthetic PSK-α peptide treatment significantly enhances root and hypocotyl elongation in Arabidopsis, mediated by the PSK RECEPTOR (PSKR) (Bendahmane et al., 2011; Wang et al., 2015). Loss-of-function mutants of PSKR and AtTPST, exhibit defects in root growth and cell size reduction, confirming the importance of PSKs in promoting growth (Matsubayashi et al., 2006; Bendahmane et al., 2011; Fang et al., 2024). BRI1-ASSOCIATED RECEPTOR KINASE 1 (BAK1)/SOMATIC EMBRYOGENESIS RECEPTOR-LIKE KINASEs (SERKs) act as co-receptors in the PSK signaling pathway, they are not directly involved in the binding of PSK (Li et al., 2002; Wang et al., 2015). Furthermore, PSK regulates plant reproductive development, promoting quorum sensing during pollen germination and pollen tube growth (Chen et al., 2000; Huang et al., 2014; Kou et al., 2020). Specific PSK expression patterns in soybean seeds and overexpression of PSK genes in wheat enhance seed and grain growth, respectively (Yu et al., 2019a; Geng et al., 2020).
PSK also modulates fruit ripening and nutrient accumulation, with exogenous PSK application promoting tomato fruit ripening via PSKR-dependent pathways (Figure 3A; Fang et al., 2024). Exogenous application of SlPSK5 enhances tomato fruit ripening, but this effect is lost in pskr1 mutants, indicating PSKR dependency (Fang et al., 2024). PSK also impedes the ubiquitination alteration of its receptor by PUB12/13, amplifying the stability of PSKR1 in tomatoes (Figure 3A; Hu et al., 2023). Activation of the SlPSK5-SlPSKR module phosphorylates the transcription factor DREBF2, boosting the expression of ripening-related genes (Figure 3A; Fang et al., 2024). A recent investigation elucidated that PSK orchestrates the equilibrium between growth and defense mechanisms in tomatoes by phosphorylating distinct amino acid residues of GLUTAMINE SYNTHETASE 2 (GS2) via the action of CALCIUM-DEPENDENT PROTEIN KINASE 28 (CPK28) (Figure 3A; Ding et al., 2023). In summary, PSK peptides are critical in root development, pollen germination, and fruit ripening processes.
1.7. PSY Peptides
PSY peptides were initially discovered in Arabidopsis suspension culture cells. In Arabidopsis, nine PSY genes encoding PSY precursor proteins with lengths ranging from 71 to 104 amino acids have been identified (Amano et al., 2007; Ogawa-Ohnishi et al., 2022). The mature PSY1 peptide comprises 18 amino acids, featuring sulfation modification on Tyr2 and hydroxylation modifications on Pro16 and Pro17 (Tost et al., 2021). Recent studies have revealed that PSY–PSYR signaling is a switch to mediate the trade-off between plant growth and stress response (Ogawa-Ohnishi et al., 2022). PSYR1, PSYR2, and PSYR3 act as redundant negative regulators of plant growth in the absence of the PSY peptide, while PSY peptides suppress PSYR signaling, promoting root growth (Figure 3B; Ogawa-Ohnishi et al., 2022). PSYR1 can intricately interact with and enhance the activity of AHA2 by phosphorylating its T881 residue. Activation of the plasma membrane H+-ATPase AHA2 by PSY1 occurs in a PSYR1-dependent manner, leading to extracellular acidification and promoting hypocotyl elongation (Figure 3B; Fuglsang et al., 2014; Pei et al., 2022). Thus, the growth-promoting function of PSY family peptides may represent a delicate balance between growth and stress resilience (Amano et al., 2007; Ogawa-Ohnishi et al., 2022). Overall, the interaction between PSY and PSYR is crucial in plant growth and development.
1.8. CIFs and TWS1
CIFs play a crucial role in regulating the integrity of the plant Casparian strip (Nakayama et al., 2017; Fujita, 2021)). In Arabidopsis, five CIF genes—CIF1, CIF2, CIF3, CIF4, and TWS1—encode post-translationally modified peptides. Their receptors, GASSHO 1 (GSO1) and GSO2, belong to the leucine-rich repeat receptor-like kinase family (Tsuwamoto et al., 2008). CIF1 and CIF2 share 21 conserved amino acids at the C-terminus and contain sulfation and hydroxylation sites crucial for binding to GSO1 (Doll et al., 2020; Okuda et al., 2020; Fujita, 2021). The structure of the GSO1-CIF2 complex reveals an interaction that stabilizes the receptor/peptide binding, forming the GSO pathway (Figure 3C; Doll et al., 2020; Royek et al., 2022). After sulfation by tyrosylprotein sulfotransferases (TPSTs) in the vascular bundle, CIF1 and CIF2 are transported to endodermal cells, where they bind to the receptor complex GSO1 and SERK family proteins (Figure 3C; Zhang et al., 2022). CIF transport halts upon Casparian strip completion, terminating the GSO pathway and lignin synthesis. Additionally, the GSO pathway regulates CASP structural domain formation and suberin synthesis in cork tissue (Doll et al., 2020; Hohmann et al., 2020; Okuda et al., 2020).
CIF3 and CIF4 are crucial in regulating pollen development; CIF3 and CIF4 precursor proteins require cleavage by the protease SBT5.4 (Truskina et al., 2022). SBT5.4 is expressed in pollen. Mature CIF3 or CIF4 molecules are detectable by GSO1/GSO2 receptors, thereby initiating the process of pollen wall development (Figure 3C; Truskina et al., 2022). TWS1, expressed in the embryo, exhibits weaker sequence similarity to the core region of CIF1 (Royek et al., 2022). Its precursor is directly processed at the C-terminus by the ALE1 peptidase, allowing it to cross the cuticle barrier and re-enter the embryo (Royek et al., 2022). It interacts with GSO1 and GSO2 receptors to establish cell-cell connections during embryonic epidermis formation (Figure 3C; Moussu et al., 2017). However, few studies have examined the processing and modification of CIF peptides in plants, indicating a need for further comprehensive research. However, few studies have examined the processing and modification of CIF peptides in plants, indicating a need for further comprehensive research.
1.9. RGF/GLV/CLEL Peptides
The RGF peptides, also known as GOLVEN (GLV) or CLE-like (CLEL) peptide family, consist of 13 amino acids and originate from precursor peptides of approximately 100 amino acids (Meng et al., 2012; Shinohara et al., 2021). These sulfated peptides typically feature the characteristic DY-motif, except for GLV9, and a highly conserved hydroxylated proline residue. Across plant species, RGFs exhibit a combination of conserved and non-conserved amino acids (Komori et al., 2009; Shinohara et al., 2021). Biologically active RGF peptides undergo tyrosine sulfation and proteolytic cleavage (Shinohara et al., 2021).
RGFs play a crucial role in root development by regulating the apical meristem. Recent studies have shown that in vitro treatments with GLVs and RGFs can significantly inhibit lateral root formation by impeding auxin accumulation during lateral root initiation (Jourquin et al., 2023). Specifically, RGF peptides modulate root apical meristem activity by regulating the expression of PLETHORA (PLT) proteins, which serve as master regulators of root formation (Figure 3D; Aida et al., 2004; Hsiao et al., 2024). Most RGF genes are expressed in quiescent center cells and adjacent cells in roots, leading to the anticipation of peptide diffusion in the meristematic region with a gradient (Matsuzaki et al., 2010; Ou et al., 2016). This gradient of RGF peptides delineates the gradient of PLT protein, modulating protein stability to ensure resilient root growth and development in dynamic environments (Shinohara et al., 2021). Researchers have found that the RGF and their receptor pair (RGF-RGFR) orchestrate the concentration gradient of the PLT protein to maintain root apical meristem activity (Figure 3D; Aida et al., 2004; Galinha et al., 2007).
In Arabidopsis, three RGF receptors (RGFRs), namely RGFR1, RGFR2, and RGFR3, have been identified in proximal meristematic tissues (Shinohara et al., 2016; Yamada et al., 2020). It has been documented that the receptor kinases RGI4/SKM2 and RGI5 are involved in RGF perception in conjunction with RGI1/RGFR1, RGI2/RGFR2, and RGI3/RGFR3 (Ou et al., 2016; Shinohara et al., 2016; Yamada et al., 2020). The quintuple mutant rgi1,2,3,4,5 shows a significant decrease in root apical meristem activity and complete insensitivity to RGF (Ou et al., 2016). Moreover, it has been confirmed that RGI4/SKM2, a paralog of RGI3/RGFR3, directly binds to RGF (Figure 3D; Song et al., 2016). Subsequent studies revealed that BAK1 and its paralogs, the SERKs, act as coreceptors of RGI1 for sensing RGF1 (Figure 3D; Song et al., 2016; Ou et al., 2022). Upon recognition by the RGI1-SERKS complex at the cellular periphery, the RGF1 signal is relayed to PLTs through a YODA–MKK4/5–MPK3/6 signaling cascade (Figure 3D; Lu et al., 2020; Shao et al., 2020). However, the direct interaction between RGI5 and RGF has not yet been demonstrated. These findings underscore the pivotal role of the RGF-RGFR module in root development by regulating the apical meristem.
1.10. CLE Peptides
CLE peptides, crucial plant signaling molecules, interact with membrane-bound receptors, modulating transcription factors and phytohormone pathways (Goad et al., 2016; Hake et al., 2019). Notably, specific CLE peptides like CLV3, TDIF, CLE40, and CLE45 play vital roles in plant development and response to stimuli (Willoughby and Nimchuk, 2021; Takahashi et al., 2018). Subsequent sections of this review will delve deeper into their regulatory roles and signaling mechanisms in plant growth and development.
1.11. CLV3
The CLAVATA3 (CLV3) peptide, crucial for cell fate determination in Arabidopsis’ shoot apical meristem, has two mature forms identified via mass spectrometry: a 12-amino acid peptide with hydroxyproline (Hyp) residues at positions 4 and 7, and a 13-amino acid arabinosylated form with a histidine at position 13 (Zeng et al., 2017; Okamoto et al., 2022). These mature CLE peptides are recognized by the CLV1/CLV2 receptors, which consist of a leucine-rich repeat receptor-like kinase (LRR-RLK) and a leucine-rich repeat receptor-like protein (LRR-RLP) lacking a kinase domain (Je et al., 2018; Lee et al., 2019). Additionally, receptors CORYNE (CRN) and RECEPTOR-LIKE PROTEIN KINASE 2 (RPK2) are involved in CLV3 signaling (Somssich et al., 2015). Recent studies show that CLV1, CLV2, and CRN form a receptor complex for CLV3 (Figure 4; Somssich et al., 2015).
RPK2, also known as TOADSTOOL 2 (TOAD2), mediates the CLV3 signaling cascade by associating with BAM1 (Han et al., 2020; Wang et al., 2022). The three BARELY ANY MERISTEM (BAM) receptor kinases, clustered within the same clade as CLV1, play a pivotal role in maintaining stem cell preservation along the peripheries of the shoot apical meristem (Somssich et al., 2015; Willoughby and Nimchuk 2021). BAM1 and BAM2 can bind to CLV3, functioning synergistically with CLV1 as supplementary receptors for CLV3. This interaction initiates a downstream phosphorylation cascade via mitogen-activated protein kinase (MAPK) pathways, with MPK3 and MPK6 phosphorylation ultimately regulating stem cell equilibrium (Figure 4; Li et al., 2019). The CLV3 signal is perceived by multiple receptor complexes—CLV1/CLV1 homomeric, CLV2/CRN heteromeric, and possibly RPK2 homomeric—negatively regulating WUSCHEL, a master regulator in plant growth signaling, thereby controlling the shoot apical meristem (Figure 4; Somssich et al., 2015; Wang et al., 2022).
1.12. TDIF
TDIF, initially derived from Zinnia elegans, regulates plant vascular meristem development (Ito et al., 2006). In Arabidopsis, the CLE41 and CLE44 genes share a 12-amino-acid sequence within their CLE domains, with CLE44 resembling TDIF but lacking glycosylation (Ohyama et al., 2008). Treatment with TDIF inhibits xylem cell differentiation while promoting procambial cell proliferation, highlighting its role in plant development (Hirakawa et al., 2008). The receptor for TDIF, TDR, belongs to the LRR-RLK XI subfamily and plays a pivotal role in maintaining vascular meristem (Morita et al., 2016; Smit et al., 2020; Yang et al., 2020). In a manner parallel to the CLV3 signaling pathways, the WUS family protein WUS-RELATED HOMEOBOX 4 (WOX4) acts as a mediator of TDIF signaling by facilitating the proliferation of procambial cells (Figure 4; Hirakawa et al., 2010). Downstream effectors of the TDIF pathway, glycogen synthase kinase 3 proteins (GSK3s), suppress xylem differentiation via the WOX4 pathway (Figure 4; Hirakawa et al., 2010). TDR activates GSK3s at the plasma membrane in a TDIF-dependent manner (Hirakawa et al., 2010). Inhibition of GSK3s induces xylem cell differentiation through the transcription factor BRI1-EMS SUPPRESSOR 1 (BES1) (Hirakawa et al., 2010; Zhang et al., 2016). Additionally, a NAC domain transcription factor, XVP, localizes on the plasma membrane, interacting with the TDIF co-receptor PXY-BAK1 to form a complex (Figure 4; Yang et al., 2020). XVP expression is localized in the cambium and is critical in regulating xylem differentiation and vascular patterning (Yang et al., 2020). Subsequent investigations elucidated that XVP governs xylem differentiation via the pivotal factor VASCULAR-RELATED NAC-DOMAIN6 (VDN6). Notably, the conservation of the CLE41/TDIF-TDR-WOX4 signaling pathway in wood-forming Populus species suggests its role as a universal regulator of vascular development (Figure 4; Yamaguchi et al., 2016).
1.13. CLE40
CLAVATA3/ESR-RELATED40 (CLE40), the closest homolog of CLV3 in Arabidopsis, regulates columella stem cells (CSCs) in the root meristem (Hobe et al., 2003; Zhu et al., 2021). CSCs, located at the root apices, perceive gravitational stimuli and have a unique cell wall composition that protects the root meristem (Hobe et al., 2003). CLE40 expression in CSCs controls cell division, as evidenced by cle40 mutants displaying excessive CSC layers (Stahl et al., 2013). Genetic studies indicate that CLE40 functions through the receptor kinases ARABIDOPSIS CRINKLY4 (ACR4) and CLAVATA1 (CLV1) to regulate WOX5 expression (Figure 4; Berckmans et al., 2020). ACR4, but not CLV1, is transcriptionally upregulated following ectopic CLE40 peptide treatment (Sarkar et al., 2007; Qin et al., 2019). ACR4 is also implicated in forming the epidermal cell layer and initiating lateral roots (Figure 4; De Smet et al., 2008). CLE40’s action is limited to cortex cells, whereas WOX5 expression in the quiescent center positively regulates stem cell production, suggesting a balance between CLE40 and WOX5 in regulating the stem cell population of the root meristem (Figure 4; Stahl et al., 2013; Richards et al., 2015; Berckmans et al., 2020).
1.14. CLE45
The interaction between CLE45 and BAM3 establishes a signaling module that regulates vascular development independently of the TDIF-TDR pathway (Yuan and Wang 2021). Synthetic CLE45 peptide inhibits root growth and protophloem differentiation in Arabidopsis (Kang and Hardtke, 2016). BAM3 acts as a receptor for CLE45, as the impact of synthetic CLE45 is mitigated in a bam3 mutant (Hazak et al., 2017). The class II LRR-RLK, known as CLE-RESISTANT RECEPTOR KINASE (CLERK), also termed CIK2, is indispensable for the complete perception of CLE45 and other CLE peptides active in early proto-phloem development (Anne et al., 2018). However, CLERK operates within a genetically distinct pathway from BAM3 and CLV2/CRN. Notably, MEMBRANE-ASSOCIATED KINASE REGULATOR 5 (MAKR5) emerges as a pivotal facilitator of CLE45 signaling, operating downstream of BAM3. CLE45 signaling recruits MAKR5 to the plasma membrane and promotes its accumulation in developing sieve elements (Figure 4; Kang and Hardtke, 2016). This signaling module interacts genetically with plasma membrane-localized proteins BREVIS RADIX (BRX) and OCTOPUS (OPS), which are critical for sieve-element (SE) cell fate control (Rodriguez et al., 2014; Anne et al., 2015). Deletion of BRX or OPS leads to gap cell development in the protophloem due to interference with interactions between CLE45 signaling receptors BAM3 and the CLV2/CRN complex. brx and ops mutants also exhibit abbreviated root characteristics and increased lateral root production from primary root branching zones (Anne et al., 2015; Rodriguez-Villalon et al., 2014). The CLE45-SKM1/SKM2 pathway facilitates intercellular communication between male and female cells, aiding pollen tube growth under heat stress and fertilization (Kang and Hardtke, 2016). While the regulatory role of CLE in root and pollen tube development is documented, the detailed molecular framework and specific signaling components governing these processes remain unclear.
In addition to the previously mentioned CLE peptides, various others exhibit diverse functionalities. For instance, peptides CLE1, 3, 4, and 7 suppress lateral root formation via the CLV1 receptor under nitrate deficiency (Araya et al., 2014). CLE18 and CLE26 are associated with root growth (Meng et al., 2012b; Czyzewicz et al., 2015). The CLE5 and CLE6 complexes influence B-type CLE and shoot development (Whitford et al., 2008). CLE8 regulates embryo and endosperm development (Xie et al., 2022), while CLE10 influences protoxylem vessel formation (Soyano et al., 2014). Overall, CLE peptides are crucial in various aspects of plant growth and development.
1.15. Other SPs
1.15.1. EPF/EPFL Peptides
The EPF/EPFL peptides are characterized by abundant cysteine residues and an N-terminal signal peptide (Kosentka et al., 2019). Upon proteolytic cleavage, mature EPF/EPFL peptides feature a conserved C-terminal region, typically containing six or eight cysteine residues that facilitate intramolecular disulfide bond formation (Tavormina et al., 2015; Bessho et al., 2016). EPF1 and EPF2, which share significant homology, interact with the ERL1 and ERL2 receptors, respectively (Hara et al., 2007). EPF/EPFL peptides are known to negatively regulate stomatal density negatively, thereby inhibiting stomatal development (Tavormina et al., 2015; Kosentka et al., 2019). Additionally, several Arabidopsis EPF members, including EPF1, EPF2, and EPFL9, regulate stomatal development by binding to a complex formed by TMM and ER receptor kinases (Figure 5A; Lin et al., 2017). EPFL9 (STOMAGEN) competes with EPF2 for ER receptor binding and fine-tuning stomatal development (Figure 5A; Yin et al., 2017; Lin et al., 2017). In contrast, CHALLAH (CHAL)/EPFL6 exhibits low expression in leaves and the stomatal lineage (Uchida et al., 2012). Although single chal mutants and triple mutants do not display any discernible phenotype in stomatal development, CLL2/EPFL4 and CHAL/EPFL6 are predominantly expressed in the endodermis of the inflorescence stem, where they regulate proper stem architecture (Figure 5A; Uchida et al., 2012; Kosentka et al., 2019). Consequently, CLL2/EPFL4 and CHAL/EPFL6 primarily act on inflorescence stem growth regulation (Kosentka et al., 2019). Recent research has elucidated that EPFL4-6 interacts with the ER-SERK receptor complex, activating the downstream MKK5/6-MPK6 cascade, thereby dynamically regulating cellular proliferation in stamen filaments (Figure 5A; He et al., 2023). Despite the incomplete understanding of EPF/EPFL-associated signaling pathways, research suggests that many EPF peptides exert a negative regulatory effect on stomatal density, thereby inhibiting stomatal development (Kosentka et al., 2019; He et al., 2023).
1.15.2. LURE Peptides
LUREs, the first identified pollen-tube attractants found in Torenia fournieri’s synergid cells, belong to the defensin-like cysteine-rich peptide family (Okuda et al., 2009). Among the sixteen cysteine-rich peptides isolated from these cells, TfCRP1 and TfCRP3, known as LURE1 and LURE2, act as pollen tube attractants (Maruyama and Higashiyama, 2016). Despite variations in their sequences, all LUREs share a conserved pattern of six cysteine residues for disulfide bond formation. However, comparisons across species, such as Torenia and Arabidopsis, reveal that the alignment of cysteine residues is the only conserved feature (Okuda et al., 2009; Maruyama and Higashiyama, 2016).
A recent study identified pollen-specific receptor kinase 6 (PRK6) as a crucial receptor for sensing the attractant peptide AtLURE1 (Figure 5B; Takeuchi and Higashiyama, 2016). It was observed that pollen tubes from prk loss-of-function mutants could not respond to AtLURE1 (Takeuchi and Higashiyama, 2016). PRK6, consisting of six leucine-rich repeats (LRRs), belongs to a subclade containing eight PRK family receptors (PRK1–8) in Arabidopsis (Takeuchi, 2021). Interestingly, studies on PRK family receptors in tomatoes and Arabidopsis have indicated their functional roles in regulating the efficiency of pollen tube growth (Yang et al., 2022). The results suggest that prk mutants (prk3 prk6 prk8 and prk1 prk3 prk6) demonstrate a dramatic decrease in pollen tube growth and fertility, while individual prk mutants do not exhibit such pronounced effects (Yang et al., 2022). Although the exact relationship with PRKs remains unclear, a separate group of pollen-expressed RLKs, including MALE DISCOVERER1 (MDIS1), MDIS2, MDIS1-INTERACTING RECEPTOR LIKE KINASE1 (MIK1), and MIK2, have also been implicated in the sensing of AtLURE1 (Figure 5B; Takeuchi, 2021; Yang et al., 2022). Even though the downstream signaling pathway of LUREs remains incompletely elucidated, we can still infer that PRK6, in conjunction with the multiple receptor components mentioned earlier, may serve as a pivotal player in regulating pollen tube growth and attraction by detecting extracellular ligands, such as AtLURE1.
1.15.3. RALF Peptides
RALF was initially discovered in tobacco plants for its ability to raise the pH of the culture medium quickly (Pearce et al., 2001). RALFs are widespread in various plant species, including dicots, monocots, and gymnosperms. In Arabidopsis, 40 RALF and RALF-like (RALFL) genes have been identified, showing diverse expression patterns in different organs and tissues (Pearce et al., 2001; Germain et al., 2005). RALF belongs to the cysteine-rich peptide family in plants and is characterized by four conserved cysteine residues forming two disulfide bonds, crucial for proper folding (Abarca et al., 2021). Upstream of the mature RALFs N-terminus are a pair of arginine residues, serving as cleavage sites for peptidases and coordinating immune responses (Datta et al., 2024). Downstream, the YISY motif is crucial for receptor binding (Matos et al., 2008). The C-terminal sequences GASYY and RCRR(S) of RALFs might stabilize the peptide structure or mediate interactions between peptides (Campbell and Turner, 2017).
Catharanthus roseus receptor-like kinase 1-like (CrRLK1L) proteins recognize RALFs, influencing plant growth (Xiao et al., 2019). For instance, the binding of RALF1 to FERONIA (FER), a member of the CrRLK1L family, suppresses lateral root growth, where BAK1 forms a receptor complex with FER (Figure 5C; Haruta et al., 2014; Stegmann et al., 2017). Additionally, RALFs regulate plant reproductive development. RALF4 and RALF19, expressed in pollen, interact with the ANXUR1/2 (ANX1/2)-BUPS1/2 (Buddha’s paper seal 1/2) complex, influencing pollen tube growth (Figure 5C; Ge et al., 2017). As the pollen tube approaches the embryo sac, RALF34 competes with RALF4 and RALF19 for binding to the ANX1/2-BUPS1/2 complex, leading to pollen tube rupture and sperm release, thereby completing double fertilization (Figure 5C; Franck et al., 2018; Galindo-Trigo et al., 2020). Moreover, LORELEI-like-GPI anchored proteins 2 and 3 (LLG2/3) serve as co-receptors alongside ANX1/2 and BUPS1/2, facilitating their function (Figure 5C; Ge et al., 2019). When the pollen tube grows towards the embryo sac, it was documented that RALF4/19 instigated the ANX1/2-BUPS1/2-LLG2/3-MARIS cascade, subsequently activating AtMLO1, 5, 9, and 15 (Figure 5C; Gao et al., 2023). This activation facilitated tip-focused Ca2+ influx, upholding pollen tube integrity (Figure 5C; Gao et al., 2023). Together, RALFs play crucial regulatory roles mediated through receptor interactions, impacting processes ranging from root development to reproductive success.
1.15.4. CEP Peptides
CEP peptides originate from proteins featuring an N-terminal secretion signal, a variable domain, and one or more CEP domains. These peptides, each spanning 15 amino acids, are reportedly essential in plant growth and development (Taleski et al., 2018; Ota et al., 2020;Taleski et al., 2023). For instance, AtCEP3 negatively regulates lateral root number in Arabidopsis, as demonstrated by increased lateral root density in cep3 mutant plants compared to wild-type under various nutrient-deficient and abiotic stress conditions (Imin et al., 2013). Additionally, CEPs are crucial in negatively regulating lateral root emergence in Medicago truncatula. Overexpression of MtCEP1 reduces lateral root number, while simultaneous knockdown of MtCEP1 and MtCEP2 increases lateral root number (Figure 5D; Imin et al., 2013). CEP5 regulates the auxin response in lateral root formation by enhancing AUX/indole-3-acetic acid (IAA) stability, thereby suppressing the expression of auxin-responsive genes (Figure 5D; Roberts et al., 2016).
In recent studies, overexpression of GhCEP46-D05 in cotton and Arabidopsis resulted in reduced plant height, fiber length, root length, and the length and width of mature seeds in transgenic lines (Figure 5D; Mei et al., 2024). Notably, CEP peptides impact plant development by specifically binding to CEP RECEPTORS 1/2 (CEPR1/CEPR2) kinases (Bellegarde et al., 2017; Chapman et al., 2019). The CEP-CEPR1 pathways inhibit lateral root growth in the presence of sucrose, other metabolizable sugars, and increased light intensity by reducing lateral root meristem size and the length of mature lateral root cells (Figure 5D; Chapman et al., 2019). Recent studies have revealed that CEP and cytokinin signaling mediate the action of CEP DOWNSTREAM (CEPD) glutaredoxins, thereby inhibiting primary root growth (Figure 5D; Taleski et al., 2023). Despite some literature outlining the functional roles of the CEP-CEPR signaling pathway in plant lateral root development, further elucidation is needed to understand the comprehensive regulatory network of this pathway in plant development (Chapman et al., 2019; Taleski et al., 2023).
1.15.5. IDA Peptides
IDA peptides, a class of small signaling proteins, are generated via the proteolytic cleavage of their precursor protein catalyzed by subtilases (Katharina et al., 2016; Stintzi and Schaller, 2022). They possess an N-terminal secretory signal peptide, guiding the protein to the extracellular milieu, while the C-terminal proline-rich extended motifs, known as the EPIP motif, comprised of 20 amino acids, play a pivotal role in inducing the abscission of floral organs post-pollination (Stenvik et al., 2008; Vie et al., 2017). Reportedly, IDA peptides regulate the shedding of cauline leaves in response to dehydration stress (Taylor et al., 2019). Under water deficit conditions, when leaves show signs of wilting, the bioactive IDA peptide ligand is perceived by a receptor complex composed of either the receptor kinases HAESA or HAESA-like 2 (HSL2) along with SERK coreceptors (Figure 5E; Stenvik et al., 2008; Taylor et al., 2019). This recognition event initiates a downstream signaling cascade involving MAPKs, ultimately governing the expression of cell-wall modifying and hydrolytic enzymes, particularly polygalacturonases and xyloglucan endotransglucosylase/hydrolases (Figure 5E; Stenvik et al., 2008; Taylor et al., 2019). These enzymes degrade the middle lamella, thereby inducing cell separation.
Furthermore, corroborating evidence confirms that IDA expression occurs shortly after abscission zone cells attain receptivity to abscission signals, signifying early loosening of cell walls (Vie et al., 2015). Moreover, overexpression of the IDA protein leads to the abscission of floral organs and the enlargement of abscission zone cells precisely at the sites where organs detach from the plant(Ying et al., 2016). These discoveries suggest that the IDA peptide likely orchestrates the initial relaxation of cell walls and subsequent detachment of abscission zone cells, possibly by regulating cell wall remodeling enzymes (Ying et al., 2016). The initial cell wall relaxation primarily occurs in the root cap and inflorescence, ultimately leading to their detachment (Geng et al., 2022; Wang et al., 2023). Furthermore, it has been observed that the overexpression of the IDA phenotype depends on the presence of the protein’s N-terminal secretion signal (Dong et al., 2022). No overexpression phenotypes are evident when IDA is overexpressed without a secretion signal (Dong et al., 2022). This finding emphasizes the vital role of IDA translocation to the apoplastic space in its functionality. In summary, IDA-mediated signaling pathways play essential roles in promoting plant organ abscission.
2. Conclusions and Perspectives
Small peptides, consisting of 5-100 amino acids, are crucial signaling molecules in plants, exhibiting diverse processing, folding, and modifications (Zhang and Zhang, 2022). Advances in sequencing, informatics, molecular biology, genetics, and biochemistry have made it possible to identify and understand these peptides’ functions (Feng et al., 2023). These peptides regulate key plant processes such as cell proliferation, tissue differentiation, organ formation, reproductive development, maturation, and senescence, and they are vital for responding to environmental stresses. Despite progress, challenges remain due to their short length and low expression levels. Recent advancements in genomics and transcriptomics have expanded research from model plants like Arabidopsis and rice to economically important crops, leading to the identification and characterization of more functional peptides (Holzwart et al., 2018; Huang et al., 2016).
In conclusion, small plant peptides are a promising emerging research area. Simplified, accurate, and rapid methods for identifying and verifying their functions are urgently needed. Future developments could enable the quantitative production of plant peptide hormones to regulate growth, enhance stress resistance, improve yields, and address challenges in eco-agriculture. Continued innovation in this field could revolutionize agriculture and significantly contribute to sustainable food production.
Author Contributions
S.Q.L and F.X. authored the draft manuscript and created the illustrations. F.X. designed the figures and edited the manuscript. All authors contributed to the discussion and approved the final version for publication. No conflicts of interest were declared.
Acknowledgments
We regret that space limitations prevent us from citing specific authors’ works in this review. This study received support from the Youth Science Foundation of Xinjiang Uygur Autonomous Region (Grant 2022D01C94 to F.X.), and the National Natural Science Foundation of China (Grant 32260074 to F.X.).
References
- Abarca A, Franck CM, Zipfel C (2021) Family-wide evaluation of RAPID ALKALINIZATION FACTOR peptides. Plant Physiol 187 (2):996-1010.
- Aida M, Beis D, Heidstra R, Willemsen V, Blilou I, Galinha C, Nussaume L, Noh Y-S, Amasino R, Scheres B (2004) The PLETHORA Genes Mediate Patterning of the Arabidopsis Root Stem Cell Niche. Cell 119 (1):109-120.
- Amano Y, Tsubouchi H, Shinohara H, Ogawa M, Matsubayashi Y (2007) Tyrosine-sulfated glycopeptide involved in cellular proliferation and expansion in Arabidopsis. Proc Natl Acad Sci USA 104 (46):18333-18338.
- Anne P, Amiguet-Vercher A, Brandt B, Kalmbach L, Geldner N, Hothorn M, Hardtke CS (2018) CLERK is a novel receptor kinase required for sensing of root-active CLE peptides in Arabidopsis. Development 145 (10):dev162354.
- Anne P, Azzopardi M, Gissot L, Beaubiat S, Hématy K, Palauqui JC (2015) OCTOPUS Negatively Regulates BIN2 to Control Phloem Differentiation in Arabidopsis thaliana. Curr Biol 25 (19):2584-2590.
- Araya T, Miyamoto M, Wibowo J, Suzuki A, Kojima S, Tsuchiya YN, Sawa S, Fukuda H, von Wirén N, Takahashi H (2014) CLE-CLAVATA1 peptide-receptor signaling module regulates the expansion of plant root systems in a nitrogen-dependent manner. Proc Natl Acad Sci USA 111 (5):2029-2034.
- Bellegarde F, Gojon A, Martin A (2017) Signals and players in the transcriptional regulation of root responses by local and systemic N signaling in Arabidopsis thaliana. J Exp Bot 68 (10):2553-2565.
- Bendahmane M, Stührwohldt N, Dahlke RI, Steffens B, Johnson A, Sauter M (2011) Phytosulfokine-α Controls Hypocotyl Length and Cell Expansion in Arabidopsis thaliana through Phytosulfokine Receptor 1. PLoS One, 6(6), e21054.
- Berckmans B, Kirschner G, Gerlitz N, Stadler R, Simon R (2020) CLE40 Signaling Regulates Root Stem Cell Fate. Plant Physiol 182 (4):1776-1792.
- Bessho-Uehara K, Wang DR, Furuta T, Minami A, Nagai K, Gamuyao R, Asano K, Angeles-Shim RB, Shimizu Y, Ayano M, Komeda N, Doi K, Miura K, Toda Y, Kinoshita T, Okuda S, Higashiyama T, Nomoto M, Tada Y, Shinohara H, Matsubayashi Y, Greenberg A, Wu J, Yasui H, Yoshimura A, Mori H, McCouch SR, Ashikari M (2016) Loss of function at RAE2, a previously unidentified EPFL, is required for awnlessness in cultivated Asian rice. Proc Natl Acad Sci USA 113 (32):8969-8974.
- Campbell L, Turner SR (2017) A Comprehensive Analysis of RALF Proteins in Green Plants Suggests There Are Two Distinct Functional Groups. Front Plant Sci 8: 240798.
- Chapman K, Taleski M, Ogilvie HA, Imin N, Djordjevic MA (2019) CEP-CEPR1 signalling inhibits the sucrose-dependent enhancement of lateral root growth. J Exp Bot 70 (15):3955-3967.
- Chen YF, Matsubayashi Y, Sakagami Y (2000) Peptide growth factor phytosulfokine-alpha contributes to the pollen population effect. Planta 211 (5):752-755.
- Chien P-S, Nam HG, Chen Y-R (2015) A salt-regulated peptide derived from the CAP superfamily protein negatively regulates salt-stress tolerance in Arabidopsis. J Exp Bot 66 (17):5301-5313.
- Czyzewicz N, Shi CL, Vu LD, Van De Cotte B, Hodgman C, Butenko MA, De Smet I (2015) Modulation of Arabidopsis and monocot root architecture by CLAVATA3/EMBRYO SURROUNDING REGION 26 peptide. J Exp Bot 66 (17):5229-5243.
- Datta T, Kumar RS, Sinha H, Trivedi PK (2024) Small but mighty: Peptides regulating abiotic stress responses in plants. Plant Cell Environ 47 (4):1207-1223.
- De Smet I, Vassileva V, De Rybel B, Levesque MP, Grunewald W, Van Damme D, Van Noorden G, Naudts M, Van Isterdael G, De Clercq R, Wang JY, Meuli N, Vanneste S, Friml J, Hilson P, Jürgens G, Ingram GC, Inzé D, Benfey PN, Beeckman T (2008) Receptor-like kinase ACR4 restricts formative cell divisions in the Arabidopsis root. Science 322 (5901):594-597.
- Ding S, Lv J, Hu Z, Wang J, Wang P, Yu J, Foyer CH, Shi K (2023) Phytosulfokine peptide optimizes plant growth and defense via glutamine synthetase GS2 phosphorylation in tomato. EMBO J 42 (6):e111858.
- Doll NM, Royek S, Fujita S, Okuda S, Chamot S, Stintzi A, Widiez T, Hothorn M, Schaller A, Geldner N, Ingram G (2020) A two-way molecular dialogue between embryo and endosperm is required for seed development. Science 367 (6476):431-435.
- Dong Q, Wang G, Iqbal A, Muhammad N, Wang X, Gui H, Zhang H, Kayoumu M, Li X, Zhang X (2022) Identification and expression analysis of the NPF genes in cotton. Int J Mol Sci 23 (22):14262.
- Fang H, Zuo J, Ma Q, Zhang X, Xu Y, Ding S, Wang J, Luo Q, Li Y, Wu C, Lv J, Yu J, Shi K (2024) Phytosulfokine promotes fruit ripening and quality via phosphorylation of transcription factor DREB2F in tomato. Plant Physiol 194 (4):2739-2754.
- Feng YZ, Zhu QF, Xue J, Chen P, Yu Y (2023) Shining in the dark: the big world of small peptides in plants. aBIOTECH 4 (3):238-256.
- Fletcher JC (2020) Recent Advances in Arabidopsis CLE Peptide Signaling. Trends Plant Sci 25 (10):1005-1016.
- Franck CM, Westermann J, Bürssner S, Lentz R, Lituiev DS, Boisson-Dernier A (2018) The Protein Phosphatases ATUNIS1 and ATUNIS2 Regulate Cell Wall Integrity in Tip-Growing Cells. Plant Cell 30 (8):1906-1923.
- Fuglsang AT, Kristensen A, Cuin TA, Schulze WX, Persson J, Thuesen KH, Ytting CK, Oehlenschlæger CB, Mahmood K, Sondergaard TE, Shabala S, Palmgren MG (2014) Receptor kinase-mediated control of primary active proton pumping at the plasma membrane. Plant J 80 (6):951-964.
- Fujita S (2021) CASPARIAN STRIP INTEGRITY FACTOR (CIF) family peptides - regulator of plant extracellular barriers. Peptides 143:170599.
- Galindo-Trigo S, Blanco-Touriñán N, DeFalco TA, Wells ES, Gray JE, Zipfel C, Smith LM (2020) CrRLK1L receptor-like kinases HERK1 and ANJEA are female determinants of pollen tube reception. EMBO Rep 21 (2):e48466.
- Galinha C, Hofhuis H, Luijten M, Willemsen V, Blilou I, Heidstra R, Scheres B (2007) PLETHORA proteins as dose-dependent master regulators of Arabidopsis root development. Nature 449 (7165):1053-1057.
- Gao Q, Wang C, Xi Y, Shao Q, Hou C, Li L, Luan S (2023) RALF signaling pathway activates MLO calcium channels to maintain pollen tube integrity. Cell Res 33 (1):71-79.
- Ge Z, Bergonci T, Zhao Y, Zou Y, Du S, Liu M-C, Luo X, Ruan H, García-Valencia LE, Zhong S, Hou S, Huang Q, Lai L, Moura DS, Gu H, Dong J, Wu H-M, Dresselhaus T, Xiao J, Cheung AY, Qu L-J (2017) Arabidopsis pollen tube integrity and sperm release are regulated by RALF-mediated signaling. Science 358 (6370):1596-1600.
- Ge Z, Zhao Y, Liu MC, Zhou LZ, Wang L, Zhong S, Hou S, Jiang J, Liu T, Huang Q, Xiao J, Gu H, Wu HM, Dong J, Dresselhaus T, Cheung AY, Qu LJ (2019) LLG2/3 Are Co-receptors in BUPS/ANX-RALF Signaling to Regulate Arabidopsis Pollen Tube Integrity. Curr Biol 29 (19):3256-3265.e3255.
- Geng R, Shan Y, Li L, Shi CL, Zhang W, Wang J, Sarwar R, Xue YX, Li YL, Zhu KM, Wang Z, Xu LZ, Aalen RB, Tan XL (2022) CRISPR-mediated BnaIDA editing prevents silique shattering, floral organ abscission, and spreading of Sclerotinia sclerotiorum in Brassica napus. Plant Commun 3 (6):100452.
- Geng Y, Jian C, Xu W, Liu H, Hao C, Hou J, Liu H, Zhang X, Li T (2020) miR164-targeted TaPSK5 encodes a phytosulfokine precursor that regulates root growth and yield traits in common wheat (Triticum aestivum L.). Plant Mol Biol 104 (6):615-628.
- Germain H, Chevalier E, Caron S, Matton DP (2005) Characterization of five RALF-like genes from Solanum chacoense provides support for a developmental role in plants. Planta 220 (3):447-454.
- Goad DM, Zhu C, Kellogg EA (2016) Comprehensive identification and clustering of CLV3/ESR-related (De Smet et al.,) genes in plants finds groups with potentially shared function. New Phytol 216 (2):605-616.
- Hake S, Hirakawa Y, Uchida N, Yamaguchi YL, Tabata R, Ishida S, Ishizaki K, Nishihama R, Kohchi T, Sawa S, Bowman JL (2019) Control of proliferation in the haploid meristem by CLE peptide signaling in Marchantia polymorpha. PLOS Genetics 15 (3).
- Han H, Liu X, Zhou Y (2020) Transcriptional circuits in control of shoot stem cell homeostasis. Curr Opin Plant Biol 53:50-56.
- Hara K, Kajita R, Torii KU, Bergmann DC, Kakimoto T (2007) The secretory peptide gene EPF1 enforces the stomatal one-cell-spacing rule. Genes Dev 21 (14):1720-1725.
- Haruta M, Sabat G, Stecker K, Minkoff BB, Sussman MR (2014) A Peptide Hormone and Its Receptor Protein Kinase Regulate Plant Cell Expansion. Science 343 (6169):408-411.
- Hazak O, Brandt B, Cattaneo P, Santiago J, Rodriguez-Villalon A, Hothorn M, Hardtke CS (2017) Perception of root-active CLE peptides requires CORYNE function in the phloem vasculature. EMBO Rep 18 (8):1367-1381.
- He L, Wu L, Li J (2024) Sulfated peptides and their receptors: Key regulators of plant development and stress adaptation. Plant Commun:100918.
- He Y, He X, Wang X, Hao M, Gao J, Wang Y, Yang ZN, Meng X (2023) An EPFL peptide signaling pathway promotes stamen elongation via enhancing filament cell proliferation to ensure successful self-pollination in Arabidopsis thaliana. New Phytol 238 (3):1045-1058.
- Hellens RP, Brown CM, Chisnall MAW, Waterhouse PM, Macknight RC (2016) The Emerging World of Small ORFs. Trends Plant Sci 21 (4):317-328.
- Hirakawa Y, Kondo Y, Fukuda H (2010) TDIF peptide signaling regulates vascular stem cell proliferation via the WOX4 homeobox gene in Arabidopsis. Plant Cell 22 (8):2618-2629.
- Hirakawa Y, Shinohara H, Kondo Y, Inoue A, Nakanomyo I, Ogawa M, Sawa S, Ohashi-Ito K, Matsubayashi Y, Fukuda H (2008) Non-cell-autonomous control of vascular stem cell fate by a CLE peptide/receptor system. Proc Natl Acad Sci USA 105 (39):15208-15213.
- Hobe M, Müller R, Grünewald M, Brand U, Simon R (2003) Loss of CLE40, a protein functionally equivalent to the stem cell restricting signal CLV3, enhances root waving in Arabidopsis. Dev Genes Evol 213 (8):371-381.
- Hohmann U, Ramakrishna P, Wang K, Lorenzo-Orts L, Nicolet J, Henschen A, Barberon M, Bayer M, Hothorn M (2020) Constitutive Activation of Leucine-Rich Repeat Receptor Kinase Signaling Pathways by BAK1-INTERACTING RECEPTOR-LIKE KINASE3 Chimera. Plant Cell 32 (10):3311-3323.
- Holzwart E, Huerta AI, Glöckner N, Garnelo Gómez B, Wanke F, Augustin S, Askani JC, Schürholz AK, Harter K, Wolf S (2018) BRI1 controls vascular cell fate in the Arabidopsis root through RLP44 and phytosulfokine signaling. Proc Natl Acad Sci USA 115 (46):11838-11843.
- Hsiao Y C, Shiue S Y, Yen M R(2024). RGF1 controls PLT2 protein stability through ROS-dependent regulation of a cysteine residue in root meristem development. bioRxiv, 2024: 2024.04. 08.588570.
- Hsiao YC, Yamada M (2020) The Roles of Peptide Hormones and Their Receptors during Plant Root Development. Genes, 12(1), 22.
- Hu X-L, Lu H, Hassan MM, Zhang J, Yuan G, Abraham PE, Shrestha HK, Villalobos Solis MI, Chen J-G, Tschaplinski TJ (2021) Advances and perspectives in discovery and functional analysis of small secreted proteins in plants. Hortic, 8(1), 130.
- Hu Z, Fang H, Zhu C, Gu S, Ding S, Yu J, Shi K (2023) Ubiquitylation of PHYTOSULFOKINE RECEPTOR 1 modulates the defense response in tomato. Plant Physiol 192 (3):2507-2522.
- Huang CH, Sun R, Hu Y, Zeng L, Zhang N, Cai L, Zhang Q, Koch MA, Al-Shehbaz I, Edger PP, Pires JC, Tan DY, Zhong Y, Ma H (2016) Resolution of Brassicaceae Phylogeny Using Nuclear Genes Uncovers Nested Radiations and Supports Convergent Morphological Evolution. Mol Biol Evol 33 (2):394-412.
- Huang WJ, Liu HK, McCormick S, Tang WH (2014) Tomato Pistil Factor STIG1 Promotes in Vivo Pollen Tube Growth by Binding to Phosphatidylinositol 3-Phosphate and the Extracellular Domain of the Pollen Receptor Kinase LePRK2. Plant Cell 26 (6):2505-2523.
- Imin N, Mohd-Radzman NA, Ogilvie HA, Djordjevic MA (2013) The peptide-encoding CEP1 gene modulates lateral root and nodule numbers in Medicago truncatula. J Exp Bot 64 (17):5395-5409.
- Ito Y, Nakanomyo I, Motose H, Iwamoto K, Sawa S, Dohmae N, Fukuda H (2006) Dodeca-CLE peptides as suppressors of plant stem cell differentiation. Science 313 (5788):842-845.
- Je BI, Xu F, Wu Q, Liu L, Meeley R, Gallagher JP, Corcilius L, Payne RJ, Bartlett ME, Jackson D (2018) The CLAVATA receptor FASCIATED EAR2 responds to distinct CLE peptides by signaling through two downstream effectors. Elife, 7, e35673.
- Jeon BW, Kim MJ, Pandey SK, Oh E, Seo PJ, Kim J (2021) Recent advances in peptide signaling during Arabidopsis root development. J Exp Bot 72(8):2889-2902.
- Jing Y, Zhao F, Lai K, Sun F, Sun C, Zou X, Xu M, Fu A, Sharifi R, Chen J (2024) Plant elicitor Peptides regulate root hair development in Arabidopsis. Frontiers in Plant Sci 15:1336129.
- Jones DS, John A, VanDerMolen KR, Nimchuk ZL (2021) CLAVATA Signaling Ensures Reproductive Development in Plants across Thermal Environments. Curr Biol 31 (1):220-227.e225.
- Jourquin J, Fernandez AI, Wang Q, Xu K, Chen J, Šimura J, Ljung K, Vanneste S, Beeckman T (2023) GOLVEN peptides regulate lateral root spacing as part of a negative feedback loop on the establishment of auxin maxima. J Exp Bot 74 (14):4031-4049.
- Katharina Schardon (2016) Precursor processing for plant peptide hormone maturation by subtilisin-like serine proteinases. Science 354(6319), 1594–1597.
- Kang YH, Hardtke CS (2016) Arabidopsis MAKR5 is a positive effector of BAM3-dependent CLE45 signaling. EMBO Rep 17 (8):1145-1154.
- Komori R, Amano Y, Ogawa-Ohnishi M, Matsubayashi Y (2009) Identification of tyrosylprotein sulfotransferase in Arabidopsis. Proc Natl Acad Sci USA 106 (35):15067-15072.
- Kosentka PZ, Overholt A, Maradiaga R, Mitoubsi O, Shpak ED (2019) EPFL Signals in the Boundary Region of the SAM Restrict Its Size and Promote Leaf Initiation. Plant Physiol 179 (1):265-279.
- Kou X, Liu Q, Sun Y, (2020) The peptide PbrPSK2 from phytosulfokine family induces reactive oxygen species (ROS) production to regulate pear pollen tube growth [J]. Frontiers in Plant Sci 11: 601993.
- Kutschmar A, Rzewuski G, Stührwohldt N, Beemster GTS, Inzé D, Sauter M (2008) PSK-α promotes root growth in Arabidopsis. New Phytol 181 (4):820-831.
- Lee H, Jun YS, Cha O-K, Sheen J (2019) Mitogen-activated protein kinases MPK3 and MPK6 are required for stem cell maintenance in the Arabidopsis shoot apical meristem. Plant Cell Rep 38 (3):311-319.
- Lee KP, Liu K, Kim EY, Medina-Puche L, Dong H, Duan J, Li M, Dogra V, Li Y, Lv R, Li Z, Lozano-Duran R, Kim C (2020) PLANT NATRIURETIC PEPTIDE A and Its Putative Receptor PNP-R2 Antagonize Salicylic Acid-Mediated Signaling and Cell Death. Plant Cell 32 (7):2237-2250.
- Li J, Wen J, Lease K.A, Doke J.T, Tax F.E, Walker, J.C (2002). BAK1, an Arabidopsis LRR receptor-like protein kinase, interacts with BRI1 and modulates brassinosteroid signaling. Cell 110:213–222.
- Li Z, Liu D, Xia Y, Li Z, Niu N, Ma S, Wang J, Song Y, Zhang G (2019) Identification and Functional Analysis of the CLAVATA3/EMBRYO SURROUNDING REGION. Int J Mol Sci 20(17), 4319.
- Lin G, Zhang L, Han Z, Yang X, Liu W, Li E, Chang J, Qi Y, Shpak ED, Chai J (2017) A receptor-like protein acts as a specificity switch for the regulation of stomatal development. Genes Dev 31 (9):927-938.
- Lu X, Shi H, Ou Y, Cui Y, Chang J, Peng L, Gou X, He K, Li J. RGF1-RGI1(2020) a Peptide-Receptor Complex, Regulates Arabidopsis Root Meristem Development via a MAPK Signaling Cascade. Mol Plant 13(11):1594-1607.
- Lu Q, Li H, Hong Y, Zhang G, Wen S, Li X, Zhou G, Li S, Liu H, Liu H, Liu Z, Varshney RK, Chen X, Liang X (2018) Genome Sequencing and Analysis of the Peanut B-Genome Progenitor (Arachis ipaensis). Front Plant Sci 9:604.
- Maruyama D, Higashiyama T (2016) The end of temptation: the elimination of persistent synergid cell identity. Curr Opin Plant Biol 34:122-126.
- Matos JL, Fiori CS, Silva-Filho MC, Moura DS (2008) A conserved dibasic site is essential for correct processing of the peptide hormone AtRALF1 in Arabidopsis thaliana. FEBS Lett 582 (23-24):3343-3347.
- Matsubayashi Y, Ogawa M, Kihara H, Niwa M, Sakagami Y (2006) Disruption and overexpression of Arabidopsis phytosulfokine receptor gene affects cellular longevity and potential for growth. Plant Physiol 142 (1):45-53.
- Matsubayashi Y, Sakagami Y (1996) Phytosulfokine, sulfated peptides that induce the proliferation of single mesophyll cells of Asparagus officinalis L. Proc Natl Acad Sci USA 93 (15):7623-7627.
- Matsuzaki Y, Ogawa-Ohnishi M, Mori A, Matsubayashi Y (2010) Secreted peptide signals required for maintenance of root stem cell niche in Arabidopsis. Science 329 (5995):1065-1067.
- Mei Z, Li B, Zhu S, Li Y, Yao J, Pan J, Zhang Y, Chen W (2024) A Genome-Wide Analysis of the CEP Gene Family in Cotton and a Functional Study of GhCEP46-D05 in Plant Development. Int J Mol Sci 25 (8).
- Meng L, Buchanan BB, Feldman LJ, Luan S (2012) CLE-like (CLEL) peptides control the pattern of root growth and lateral root development in Arabidopsis. Proc Natl Acad Sci USA 109 (5):1760-1765.
- Morita J, Kato K, Nakane T, Kondo Y, Fukuda H, Nishimasu H, Ishitani R, Nureki O (2016) Crystal structure of the plant receptor-like kinase TDR in complex with the TDIF peptide. Nat Commun 7:12383.
- Moussu S, Doll NM, Chamot S, Brocard L, Creff A, Fourquin C, Widiez T, Nimchuk ZL, Ingram G (2017) ZHOUPI and KERBEROS Mediate Embryo/Endosperm Separation by Promoting the Formation of an Extracuticular Sheath at the Embryo Surface. Plant Cell 29 (7):1642-1656.
- Ogawa-Ohnishi M, Yamashita T, Kakita M, Nakayama T, Ohkubo Y, Hayashi Y, Yamashita Y, Nomura T, Noda S, Shinohara H, Matsubayashi Y (2022) Peptide ligand-mediated trade-off between plant growth and stress response. Science 378 (6616):175-180.
- Ohyama K, Ogawa M, Matsubayashi Y (2008) Identification of a biologically active, small, secreted peptide in Arabidopsis by in silico gene screening, followed by LC-MS-based structure analysis. Plant J 55 (1):152-160.
- Okamoto S, Kawasaki A, Makino Y, Ishida T, Sawa S (2022) Long-distance translocation of CLAVATA3/ESR-related 2 peptide and its positive effect on roots sucrose status. Plant Physiol 189 (4):2357-2367.
- Okuda S, Fujita S, Moretti A, Hohmann U, Doblas VG, Ma Y, Pfister A, Brandt B, Geldner N, Hothorn M (2020) Molecular mechanism for the recognition of sequence-divergent CIF peptides by the plant receptor kinases GSO1/SGN3 and GSO2. Proc Natl Acad Sci USA 117 (5):2693-2703.
- Okuda S, Tsutsui H, Shiina K, Sprunck S, Takeuchi H, Yui R, Kasahara RD, Hamamura Y, Mizukami A, Susaki D, Kawano N, Sakakibara T, Namiki S, Itoh K, Otsuka K, Matsuzaki M, Nozaki H, Kuroiwa T, Nakano A, Kanaoka MM, Dresselhaus T, Sasaki N, Higashiyama T (2009) Defensin-like polypeptide LUREs are pollen tube attractants secreted from synergid cells. Nature 458 (7236):357-361.
- Olsson V, Joos L, Zhu S, Gevaert K, Butenko MA, De Smet I (2019) Look Closely, the Beautiful May Be Small: Precursor-Derived Peptides in Plants. Annu Rev Plant Biol 70:153-186.
- Ong SN, Tan BC, Al-Idrus A, Teo CH (2022) Small open reading frames in plant research: from prediction to functional characterization. 3 Biotech 12 (3):76.
- Ota R, Ohkubo Y, Yamashita Y, Ogawa-Ohnishi M, Matsubayashi Y (2020) Shoot-to-root mobile CEPD-like 2 integrates shoot nitrogen status to systemically regulate nitrate uptake in Arabidopsis. Nat Commun 11 (1):641.
- Ou Y, Lu X, Zi Q, Xun Q, Zhang J, Wu Y, Shi H, Wei Z, Zhao B, Zhang X, He K, Gou X, Li C, Li J (2016) RGF1 INSENSITIVE 1 to 5, a group of LRR receptor-like kinases, are essential for the perception of root meristem growth factor 1 in Arabidopsis thaliana. Cell Res 26 (6):686-698.
- Ou Y, Tao B, Wu Y, Cai Z, Li H, Li M, He K, Gou X, Li J (2022) Essential roles of SERKs in the ROOT MERISTEM GROWTH FACTOR-mediated signaling pathway. Plant Physiol 189 (1):165-177.
- Pearce G, Moura DS, Stratmann J, Ryan CA (2001) RALF, a 5-kDa ubiquitous polypeptide in plants, arrests root growth and development. Proc Natl. Acad. Sci. USA, 98, 12 843–12 847.
- Pearce G, Munske G, Yamaguchi Y, Ryan CA (2010) Structure-activity studies of GmSubPep, a soybean peptide defense signal derived from an extracellular protease. Peptides 31 (12):2159-2164.
- Pei D, Hua D, Deng J, Wang Z, Gong Z (2022). Phosphorylation of the plasma membrane H+-ATPase AHA2 by BAK1 is required for ABA-induced stomatal closure in Arabidopsis. Plant Cell 34(7), 2708-2729.
- Qin Y, Yang L, Sun Z, Wang X, Wang Y, Zhang J, Rehman AU, Chen Z, Qi J, Wang B, Song C, Yang S, Gong Z (2019) Redox-Mediated Endocytosis of a Receptor-Like Kinase during Distal Stem Cell Differentiation Depends on Its Tumor Necrosis Factor Receptor Domain. Plant Physiol 181 (3):1075-1095.
- Reichardt S, Piepho H-P, Stintzi A, Schaller A (2020) Peptide signaling for drought-induced tomato flower drop. Science 367 (6485):1482-1485.
- Richards S, Wink RH, Simon R (2015) Mathematical modelling of WOX5- and CLE40-mediated columella stem cell homeostasis in Arabidopsis. J Exp Bot 66 (17):5375-5384.
- Roberts I, Smith S, Stes E, De Rybel B, Staes A, van de Cotte B, Njo MF, Dedeyne L, Demol H, Lavenus J, Audenaert D, Gevaert K, Beeckman T, De Smet I (2016) CEP5 and XIP1/CEPR1 regulate lateral root initiation in Arabidopsis. J Exp Bot 67 (16):4889-4899.
- Rodriguez-Villalon A, Gujas B, Kang YH, Breda AS, Cattaneo P, Depuydt S, Hardtke CS (2014) Molecular genetic framework for protophloem formation. Proc Natl Acad Sci USA 111 (31):11551-11556.
- Royek S, Bayer M, Pfannstiel J, Pleiss J, Ingram G, Stintzi A, Schaller A (2022) Processing of a plant peptide hormone precursor facilitated by posttranslational tyrosine sulfation. Proc Natl Acad Sci USA 119 (16):e2201195119.
- Ryan CA, Pearce G (1998) Systemin: a polypeptide signal for plant defensive genes. Annu Rev Cell Dev Biol 14:1-17.
- Rzemieniewski J, Leicher H, Lee H K(2022) CEP signaling coordinates plant immunity with nitrogen status. bioRxiv, 2022-12.
- Sarkar AK, Luijten M, Miyashima S, Lenhard M, Hashimoto T, Nakajima K, Scheres B, Heidstra R, Laux T (2007) Conserved factors regulate signalling in Arabidopsis thaliana shoot and root stem cell organizers. Nature 446 (7137):811-814.
- Sauter M (2015) Phytosulfokine peptide signalling. J Exp Bot 66 (17):5161-5169.
- Shao Y, Yu X, Xu X, Li Y, Yuan W, Xu Y, Mao C, Zhang S, Xu J(2020). The YDA-MKK4/MKK5-MPK3/MPK6 Cascade Functions Downstream of the RGF1-RGI Ligand-Receptor Pair in Regulating Mitotic Activity in Root Apical Meristem. Mol Plant.13(11):1608-1623.
- Shen W, Zhang X, Liu J, Tao K, Li C, Xiao S, Zhang W, Li JF (2022) Plant elicitor peptide signalling confers rice resistance to piercing-sucking insect herbivores and pathogens. Plant Biotechnol J 20 (5):991-1005.
- Shinohara H (2021) Root meristem growth factor RGF, a sulfated peptide hormone in plants. Peptides 142, 170556.
- Shinohara H, Mori A, Yasue N, Sumida K, Matsubayashi Y (2016) Identification of three LRR-RKs involved in perception of root meristem growth factor in Arabidopsis. Proc Natl Acad Sci USA 113 (14):3897-3902.
- Smit ME, McGregor SR, Sun H, Gough C, Bågman AM, Soyars CL, Kroon JT, Gaudinier A, Williams CJ, Yang X, Nimchuk ZL, Weijers D, Turner SR, Brady SM, Etchells JP (2020) A PXY-Mediated Transcriptional Network Integrates Signaling Mechanisms to Control Vascular Development in Arabidopsis. Plant Cell 32 (2):319-335.
- Somssich M, Ma Q, Weidtkamp-Peters S, Stahl Y, Felekyan S, Bleckmann A, Seidel CA, Simon R (2015) Real-time dynamics of peptide ligand-dependent receptor complex formation in planta. Sci Signal 8(388), ra76-ra76.
- Song W, Liu L, Wang J, Wu Z, Zhang H, Tang J, Lin G, Wang Y, Wen X, Li W, Han Z, Guo H, Chai J (2016) Signature motif-guided identification of receptors for peptide hormones essential for root meristem growth. Cell Res 26 (6):674-685.
- Soyano T, Hirakawa H, Sato S, Hayashi M, Kawaguchi M (2014) Nodule Inception creates a long-distance negative feedback loop involved in homeostatic regulation of nodule organ production. Proc Natl Acad Sci USA 111 (40):14607-14612.
- Stahl Y, Grabowski S, Bleckmann A, Kühnemuth R, Weidtkamp-Peters S, Pinto KG, Kirschner GK, Schmid JB, Wink RH, Hülsewede A, Felekyan S, Seidel CA, Simon R (2013) Moderation of Arabidopsis root stemness by CLAVATA1 and ARABIDOPSIS CRINKLY4 receptor kinase complexes. Curr Biol 23 (5):362-371.
- Stegmann M, Monaghan J, Smakowska-Luzan E, Rovenich H, Lehner A, Holton N, Belkhadir Y, Zipfel C (2017) The receptor kinase FER is a RALF-regulated scaffold controlling plant immune signaling. Science 355 (6322):287-289.
- Stenvik GE, Tandstad NM, Guo Y, Shi CL, Kristiansen W, Holmgren A, Clark SE, Aalen RB, Butenko MA (2008) The EPIP peptide of INFLORESCENCE DEFICIENT IN ABSCISSION is sufficient to induce abscission in arabidopsis through the receptor-like kinases HAESA and HAESA-LIKE2. Plant Cell 20 (7):1805-1817.
- Stintzi A, Schaller A (2022) Biogenesis of post-translationally modified peptide signals for plant reproductive development. Curr Opin Plant Biol 69:102274.
- Stührwohldt N, Bühler E, Sauter M, Schaller A, Bozhkov P (2021) Phytosulfokine (PSK) precursor processing by subtilase SBT3.8 and PSK signaling improve drought stress tolerance in Arabidopsis. J Exp Bot 72 (9):3427-3440.
- Takahashi, F., Suzuki, T., Osakabe, Y. (2018). A small peptide modulates stomatal control via abscisic acid in long-distance signalling. Nature 556, 235–238.
- Takeuchi H (2021) The role of diverse LURE-type cysteine-rich peptides as signaling molecules in plant reproduction. Peptides 142:170572.
- Takeuchi H, Higashiyama T (2016) Tip-localized receptors control pollen tube growth and LURE sensing in Arabidopsis. Nature 531 (7593):245-248.
- Taleski M, Chapman K, Novák O, Schmülling T, Frank M, Djordjevic MA (2023) CEP peptide and cytokinin pathways converge on CEPD glutaredoxins to inhibit root growth. Nature Communications 14 (1):1683.
- Taleski M, Imin N, Djordjevic MA (2018) CEP peptide hormones: key players in orchestrating nitrogen-demand signalling, root nodulation, and lateral root development. J Exp Bot 69 (8):1829-1836.
- Tavormina P, De Coninck B, Nikonorova N, De Smet I, Cammue BP (2015) The Plant Peptidome: An Expanding Repertoire of Structural Features and Biological Functions. Plant Cell 27 (8):2095-2118.
- Taylor I, Baer J, Calcutt R, Walker JC (2019) Hypermorphic SERK1 Mutations Function via a SOBIR1 Pathway to Activate Floral Abscission Signaling. Plant Physiol 180 (2):1219-1229.
- Tian D, Xie Q, Deng Z, Xue J, Li W, Zhang Z, Dai Y, Zheng B, Lu T, De Smet I, Guo Y (2022) Small secreted peptides encoded on the wheat (triticum aestivum L.) genome and their potential roles in stress responses. Front Plant Sci 13:1000297.
- Tost AS, Kristensen A, Olsen LI, Axelsen KB, Fuglsang AT (2021) The PSY Peptide Family-Expression, Modification and Physiological Implications. Genes, 12(2), 218.
- Truskina J, Brück S, Stintzi A, Boeuf S, Doll NM, Fujita S, Geldner N, Schaller A, Ingram GC (2022) A peptide-mediated, multilateral molecular dialogue for the coordination of pollen wall formation. Proc Natl Acad Sci USA 119 (22):e2201446119.
- Tsuwamoto R, Fukuoka H, Takahata Y (2008) GASSHO1 and GASSHO2 encoding a putative leucine-rich repeat transmembrane-type receptor kinase are essential for the normal development of the epidermal surface in Arabidopsis embryos. The Plant Journal 54 (1):30-42.
- Uchida N, Lee JS, Horst RJ, Lai HH, Kajita R, Kakimoto T, Tasaka M, Torii KU (2012) Regulation of inflorescence architecture by intertissue layer ligand-receptor communication between endodermis and phloem. Proc Natl Acad Sci USA 109 (16):6337-6342.
- Vie AK, Najafi J, Liu B, Winge P, Butenko MA, Hornslien KS, Kumpf R, Aalen RB, Bones AM, Brembu T (2015) The IDA/IDA-LIKE and PIP/PIP-LIKE gene families in Arabidopsis: phylogenetic relationship, expression patterns, and transcriptional effect of the PIPL3 peptide. J Exp Bot 66 (17):5351-5365.
- Vie AK, Najafi J, Winge P, Cattan E, Wrzaczek M, Kangasjärvi J, Miller G, Brembu T, Bones AM (2017) The IDA-LIKE peptides IDL6 and IDL7 are negative modulators of stress responses in Arabidopsis thaliana. J Exp Bot 68 (13):3557-3571.
- Wang J, Li H, Han Z, Zhang H, Wang T, Lin G, Chang J, Yang W, Chai J (2015) Allosteric receptor activation by the plant peptide hormone phytosulfokine. Nature 525 (7568):265-268.
- Wang P, Yao S, Kosami K.I, Guo T, Li J, Zhang Y, Fukao Y, Kaneko-Kawano T, Zhang H, She Y.M, Wang P, Xing W, Hanada K, Liu R, and Kawano Y. (2020). Identification of endogenous small peptides involved in rice immunity through transcriptomics- and proteomics-based screening. Plant Biotechnol J 18, 415-428.
- Wang P, Wu T, Jiang C, Huang B, Li Z (2023) Brt9SIDA/IDALs as peptide signals mediate diverse biological pathways in plants. Plant Science 330:111642.
- Wang Y, Shirakawa M, Ito T (2022) Dynamic Changes in Reactive Oxygen Species in the Shoot Apex Contribute to Stem Cell Death in Arabidopsis thaliana. Int J Mol Sci 23 (7).
- Whitford R, Fernandez A, De Groodt R, Ortega E, Hilson P (2008) Plant CLE peptides from two distinct functional classes synergistically induce division of vascular cells. Proc Natl Acad Sci USA 105 (47):18625-18630.
- Willoughby AC, Nimchuk ZL (2021) WOX going on: CLE peptides in plant development. Curr Opin Plant Biol 63:102056.
- Xiao F, Zhou H (2023) Plant salt response: Perception, signaling, and tolerance. Front Plant Sci 13:1053699.
- Xiao Y, Stegmann M, Han Z, DeFalco TA, Parys K, Xu L, Belkhadir Y, Zipfel C, Chai J (2019) Mechanisms of RALF peptide perception by a heterotypic receptor complex. Nature 572 (7768):270-274.
- Xie M, Zhao C, Song M, Xiang Y, Tong C (2022) Genome-wide identification and comparative analysis of CLE family in rapeseed and its diploid progenitors. Front Plant Sci 13:998082.
- Xu C, Xiang L, Huang W, Zhang X, Mao C, Wu S, Li T, Wang S, Wang S(2024). Unraveling a Small Secreted Peptide SUBPEP3 That Positively Regulates Salt-Stress Tolerance in Pyrus betulifolia. Int J Mol Sci.25(9):4612.
- Xu K, Tian D, Wang T, Zhang A, Elsadek MAY, Liu W, Chen L, Guo Y (2023) Small secreted peptides (SSPs) in tomato and their potential roles in drought stress response. Mol Hortic 3 (1):17.
- Yamaguchi YL, Ishida T, Sawa S (2016) CLE peptides and their signaling pathways in plant development. J Exp Bot 67 (16):4813-4826.
- Yamada M, Han X, Benfey P.N (2020). RGF1 controls root meristem size through ROS signalling. Nature 577, 85–88.
- Yang JH, Lee KH, Du Q, Yang S, Yuan B, Qi L, Wang H (2020) A membrane-associated NAC domain transcription factor XVP interacts with TDIF co-receptor and regulates vascular meristem activity. New Phytol 226 (1):59-74.
- Yang W, Zhai H, Wu F, Deng L, Chao Y, Meng X, Chen Q, Liu C, Bie X, Sun C, Yu Y, Zhang X, Zhang X, Chang Z, Xue M, Zhao Y, Meng X, Li B, Zhang X, Zhang D, Zhao X, Gao C, Li J, Li C (2024) Peptide REF1 is a local wound signal promoting plant regeneration. Cell, S0092-8674(24)00466-5.
- Yang Y, Niu Y, Chen T, Zhang H, Zhang J, Qian D, Bi M, Fan Y, An L, Xiang Y (2022) The phospholipid flippase ALA3 regulates pollen tube growth and guidance in Arabidopsis. Plant Cell 34 (10):3718-3736.
- Yin X, Biswal AK, Dionora J, Perdigon KM, Balahadia CP, Mazumdar S, Chater C, Lin H-C, Coe RA, Kretzschmar T, Gray JE, Quick PW, Bandyopadhyay A (2017) CRISPR-Cas9 and CRISPR-Cpf1 mediated targeting of a stomatal developmental gene EPFL9 in rice. Plant Cell Rep 36 (5):745-757.
- Ying P, Li C, Liu X, Xia R, Zhao M, Li J (2016) Identification and molecular characterization of an IDA-like gene from litchi, LcIDL1, whose ectopic expression promotes floral organ abscission in Arabidopsis. Scientific Reports 6 (1):37135.
- Yu L, Liu Y, Zeng S, Yan J, Wang E, Luo L (2019a) Expression of a novel PSK-encoding gene from soybean improves seed growth and yield in transgenic plants. Planta 249 (4):1239-1250.
- Yu Z, Xu Y, Liu L, Guo Y, Yuan X, Man X, Liu C, Yang G, Huang J, Yan K, Zheng C, Wu C, Zhang S (2019b) The Importance of Conserved Serine for C-Terminally Encoded Peptides Function Exertion in Apple. Int J Mol Sci, 20(3), 775.
- Yuan B, Wang H (2021) Peptide signaling pathways regulate plant vascular development. Front Plant Sci 12:719606.
- Zelman AK, Berkowitz GA(2023). Plant Elicitor Peptide (Pep) Signaling and Pathogen Defense in Tomato. Plants 12(15):2856.
- Zeng J, Dong Z, Wu H, Tian Z, Zhao Z (2017) Redox regulation of plant stem cell fate.EMBO J 36(19): 2844–2855.
- Zhang H, Hu Z, Lei C, Zheng C, Wang J, Shao S, Li X, Xia X, Cai X, Zhou J, Zhou Y, Yu J, Foyer CH, Shi K (2018) A Plant Phytosulfokine Peptide Initiates Auxin-Dependent Immunity through Cytosolic Ca2+ Signaling in Tomato. Plant Cell 30 (3):652-667.
- Zhang H, Li X, Wang W, Li H, Cui Y, Zhu Y, Kui H, Yi J, Li J, Gou X (2022) SERKs regulate embryonic cuticle integrity through the TWS1-GSO1/2 signaling pathway in Arabidopsis. New Phytol 233 (1):313-328.
- Zhang H, Lin X, Han Z, Qu LJ, Chai J (2016) Crystal structure of PXY-TDIF complex reveals a conserved recognition mechanism among CLE peptide-receptor pairs. Cell Res 26 (5):543-555.
- Zhang M, Zhang S (2022) Mitogen-activated protein kinase cascades in plant signaling. J Integr Plant Biol 64 (2):301-341.
- Zhou H, Xia F, Zheng Y, Liu G, Zhuang Y, Wang Z, Zhang Y, He J, Fu C, and Lin H.(2022). PAMP-INDUCED SECRETED PEPTIDE 3 modulates salt tolerance through RECEPTOR-LIKE KINASE 7 in plants. Plant Cell 34, 927-944.
- Zhu Y, Hu C, Cui Y, Zeng L, Li S, Zhu M, Gou, X. (2021). Conserved and differentiated functions of CIK receptor kinases in modulating stem cell signaling in Arabidopsis. Mol Plant 14(7): 1119-1134.
Figure 1.
Classification of Small Peptides in Plants. Small peptides (SPs) in plants fall into two categories: precursor-derived and nonprecursor-derived. Precursor-derived peptides can be functional or nonfunctional; functional ones exhibit their role post-transport, while nonfunctional ones may undergo posttranslational modifications and are categorized as Cys-rich or non-Cys-rich/non-PTM. Nonprecursor-derived peptides are encoded by small open reading frames (sORFs) containing fewer than 100 amino acids, typically found in (a) the 5′ upstream region of the main open reading frame (ORF), (b) transcripts not exceeding 100 amino acids, or (c) primary transcripts of microRNAs (pri-miRNAs).
Figure 1.
Classification of Small Peptides in Plants. Small peptides (SPs) in plants fall into two categories: precursor-derived and nonprecursor-derived. Precursor-derived peptides can be functional or nonfunctional; functional ones exhibit their role post-transport, while nonfunctional ones may undergo posttranslational modifications and are categorized as Cys-rich or non-Cys-rich/non-PTM. Nonprecursor-derived peptides are encoded by small open reading frames (sORFs) containing fewer than 100 amino acids, typically found in (a) the 5′ upstream region of the main open reading frame (ORF), (b) transcripts not exceeding 100 amino acids, or (c) primary transcripts of microRNAs (pri-miRNAs).
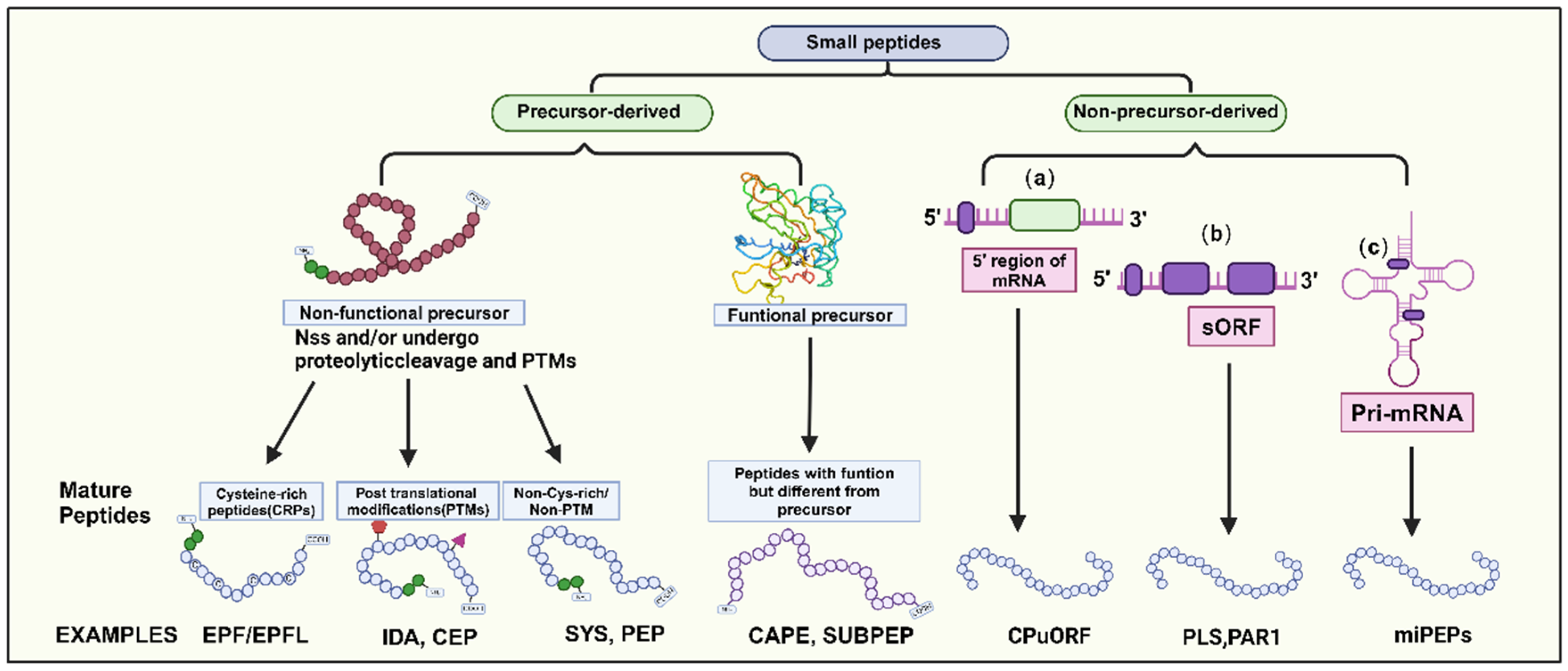
Figure 2.
Identification of Small Peptides in Plants. (A) The workflow includes four steps: extracting and enriching the low molecular weight (MW) fraction of the secreted proteome, fractionating or isolating low MW fractions, testing SSP bioactivity, and analyzing SSP fraction libraries using high-resolution/high-mass accuracy LC-MS/MS with de novo search strategies. (B) Recent bioinformatics methods for peptide identification follow a comprehensive approach. Initially, proteins are predicted based on size criteria, with SignalP predicting N-terminal signal sequences (NSS) and TMMHMM identifying and excluding transmembrane domains. ER domains are targeted to eliminate proteins associated with endoplasmic reticulum docking. This process identifies Putative SPs for further validation. Similarly, A similar strategy is applied to identify Noncoding Small Secreted Peptides (NCDS), initially identifying potential SSP-encoding small open reading frames (sORFs) and validating them using similar predictive tools.
Figure 2.
Identification of Small Peptides in Plants. (A) The workflow includes four steps: extracting and enriching the low molecular weight (MW) fraction of the secreted proteome, fractionating or isolating low MW fractions, testing SSP bioactivity, and analyzing SSP fraction libraries using high-resolution/high-mass accuracy LC-MS/MS with de novo search strategies. (B) Recent bioinformatics methods for peptide identification follow a comprehensive approach. Initially, proteins are predicted based on size criteria, with SignalP predicting N-terminal signal sequences (NSS) and TMMHMM identifying and excluding transmembrane domains. ER domains are targeted to eliminate proteins associated with endoplasmic reticulum docking. This process identifies Putative SPs for further validation. Similarly, A similar strategy is applied to identify Noncoding Small Secreted Peptides (NCDS), initially identifying potential SSP-encoding small open reading frames (sORFs) and validating them using similar predictive tools.
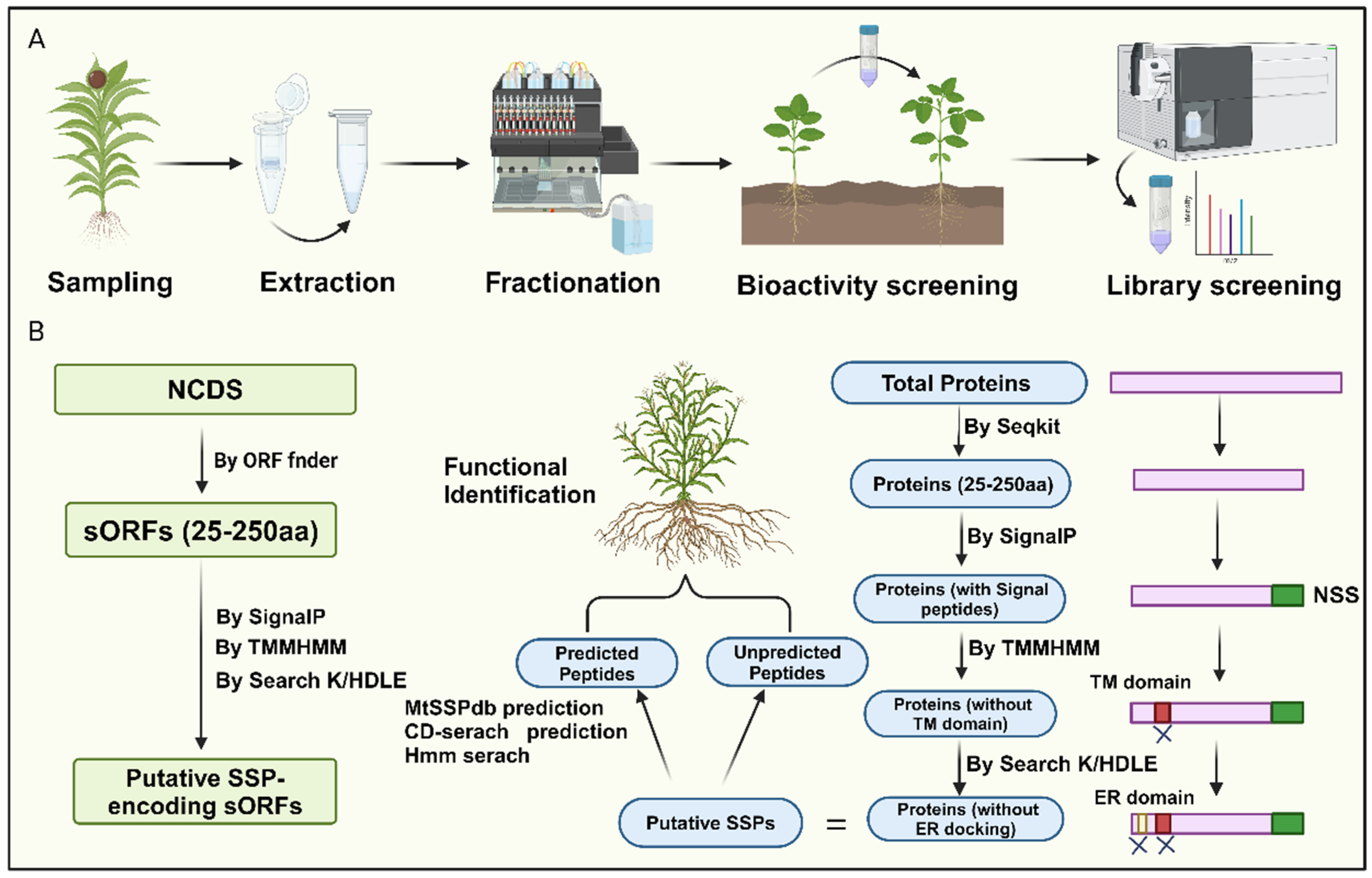
Figure 3.
The signaling pathways of four sulfated peptides. (A) The PSK signaling pathway regulates plant growth, development, and fruit ripening. (B) The interaction between PSYs and PSYR governs the regulation of root elongation. (C) CIFs interact with the CSOs-BAK1 co-receptor to modulate the formation of the plant’s Casparian strip, pollen wall development, and embryonic cuticle formation. (D) The RGF signaling pathway modulates root meristem development and elongation.
Figure 3.
The signaling pathways of four sulfated peptides. (A) The PSK signaling pathway regulates plant growth, development, and fruit ripening. (B) The interaction between PSYs and PSYR governs the regulation of root elongation. (C) CIFs interact with the CSOs-BAK1 co-receptor to modulate the formation of the plant’s Casparian strip, pollen wall development, and embryonic cuticle formation. (D) The RGF signaling pathway modulates root meristem development and elongation.
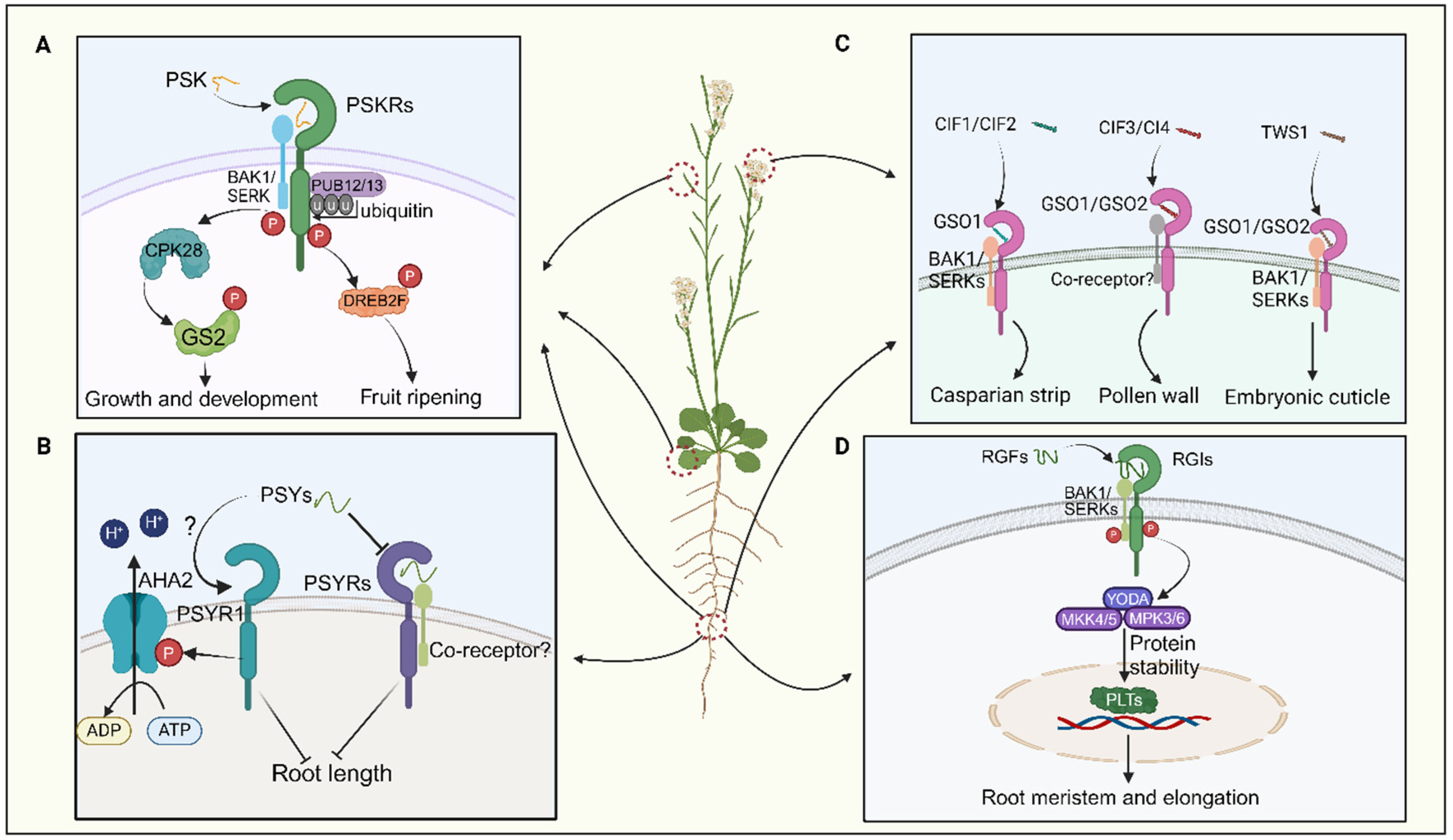
Figure 4.
The processes modulated by CLE signaling pathways and their impact on cells. CLEs bind to receptors, including CLV2, CLV3, BAMs, TDR, CIK2, and ACR4, triggering recruitment of co-receptors CRN, RPK2, and BAK1 to the complex. These initial interactions initiate downstream signaling cascades regulating SAM homeostasis, xylem differentiation, cambial cell division, protophloem differentiation, columella stem cells, and lateral root development. Arrows and bars represent positive and negative regulation, while solid and dashed lines denote direct and indirect regulation.
Figure 4.
The processes modulated by CLE signaling pathways and their impact on cells. CLEs bind to receptors, including CLV2, CLV3, BAMs, TDR, CIK2, and ACR4, triggering recruitment of co-receptors CRN, RPK2, and BAK1 to the complex. These initial interactions initiate downstream signaling cascades regulating SAM homeostasis, xylem differentiation, cambial cell division, protophloem differentiation, columella stem cells, and lateral root development. Arrows and bars represent positive and negative regulation, while solid and dashed lines denote direct and indirect regulation.
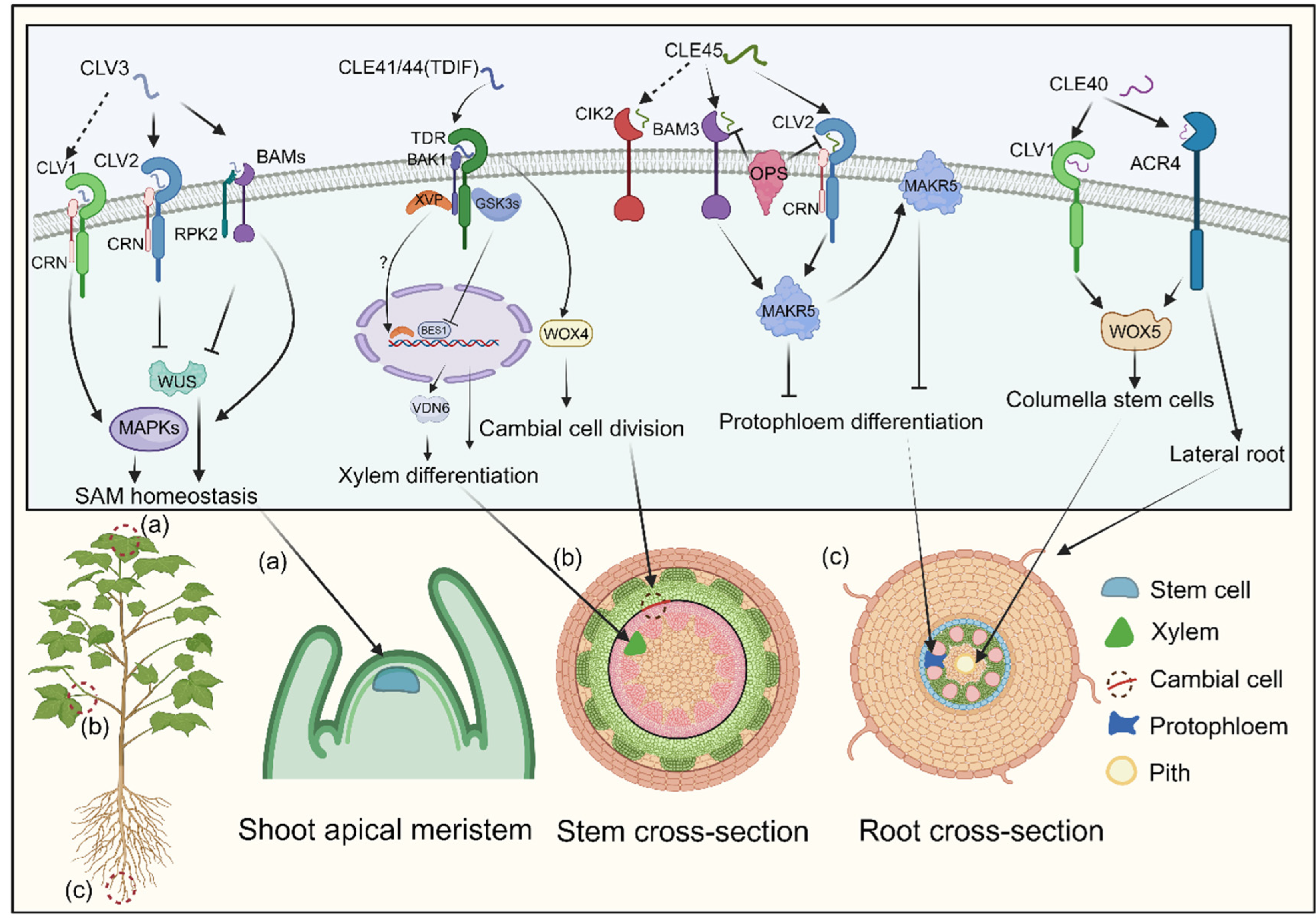
Figure 5.
The signaling transduction pathways and functional implications of EPF/EPFL, CEP, RALF, LURE, and IDA/IDL. (A) EPFs modulate stomatal development by binding to the ER-TMM co-receptor and regulate stamen filament proliferation by interacting downstream with the ER-SERK co-receptor. (B) The binding of LURE1 to its receptor promotes the growth of pollen tubes. (C) Activation of downstream signaling pathways by the binding of RALFs to their receptors regulates the development of pollen tubes and the elongation of root cells. (D) AtCEP3 and AtCEP5 interact with CEPRs to initiate signaling cascades, thereby modulating lateral root growth. GhCEP46 binds to CEPRs to inhibit fiber length and plant height. (E) IDA/IDL’s binding with the HSL2-SERK co-receptor triggers downstream signal transduction, facilitating root cap and inflorescence detachment.
Figure 5.
The signaling transduction pathways and functional implications of EPF/EPFL, CEP, RALF, LURE, and IDA/IDL. (A) EPFs modulate stomatal development by binding to the ER-TMM co-receptor and regulate stamen filament proliferation by interacting downstream with the ER-SERK co-receptor. (B) The binding of LURE1 to its receptor promotes the growth of pollen tubes. (C) Activation of downstream signaling pathways by the binding of RALFs to their receptors regulates the development of pollen tubes and the elongation of root cells. (D) AtCEP3 and AtCEP5 interact with CEPRs to initiate signaling cascades, thereby modulating lateral root growth. GhCEP46 binds to CEPRs to inhibit fiber length and plant height. (E) IDA/IDL’s binding with the HSL2-SERK co-receptor triggers downstream signal transduction, facilitating root cap and inflorescence detachment.
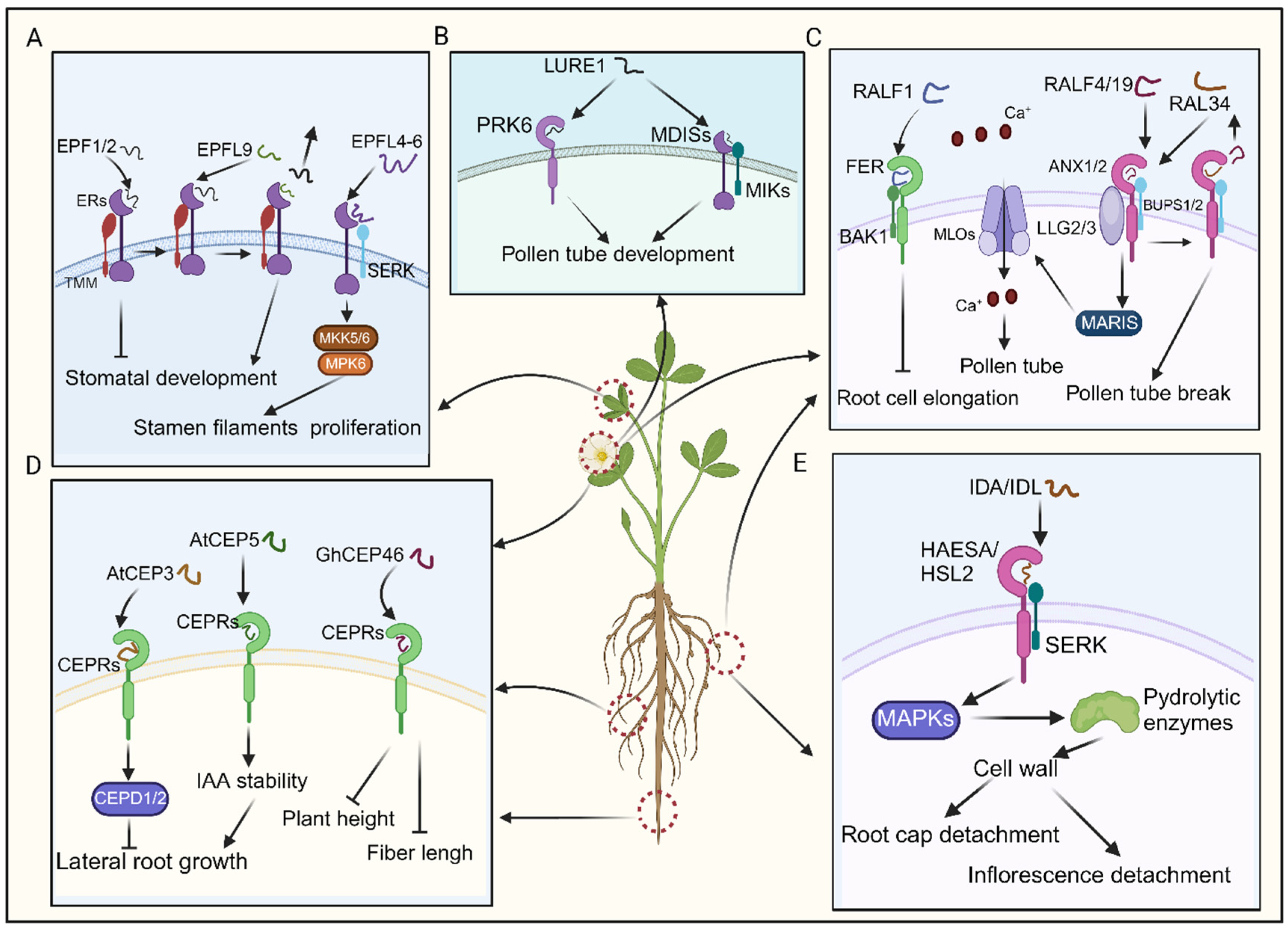
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



