
Submitted:
03 June 2024
Posted:
04 June 2024
You are already at the latest version
Abstract
Lipids have not traditionally been considered likely candidates for catalyzing reactions in biological systems. However, there is significant evidence that aggregates of amphiphilic compounds are capable of catalyzing reactions in synthetic organic chemistry. Here we demonstrate the potential for the hydrophobic region of a lipid bilayer to provide an environment suitable for catalysis by means of a lipozyme, or lipid aggregate capable of speeding up a chemical reaction. By bringing organic molecules into the non-polar or hydrophobic region of a lipid bilayer, reactions can be catalyzed by collections of small, nonpolar or amphiphilic molecules. We demonstrate this concept by the ester hydrolysis of calcein-AM to produce a fluorescent product, which is a widely used assay for esterase activity in cells. The reaction was first carried out in a two phase octanol-water system with the organic phase containing the cationic amphiphiles cetyltrimethylammonium bromide (CTAB) or octadecylamine. The octanol was then replaced with DOPC added to the water in the form of vesicles, where the reaction was also found to be carried out. The reaction was monitored using quantitative fluorescence which revealed catalytic turnover numbers on a scale of 10−4s−1 for each system, which is comparable to some slow enzymes. The reaction product was characterized by 1H-NMR measurements which were consistent with ester hydrolysis. The implications of thinking about lipid aggregates as catalytic entities are discussed in the context of biochemistry, pharmacology and synthetic biology.
Keywords:
catalysis
; partitioning
; lipid droplet
; vesicle
; heterogeneous
; biochemistry
; pharmacology
; synthetic biology
1. Introduction
Chemical reactions in biological systems form the basis of metabolism and signaling pathways, and are carried out by protein based catalysts known as enzymes. Enzymes are vital drug targets in treating major diseases due to their role in nearly all physiological and pathological processes.[1,2,3] Globular enzymes are essentially polymeric micelles, and there are also enzymes that are embedded within a lipid bilayer. Enzyme activity is carried out at a catalytic site where the substrate binds and is exposed to specific functional groups such as catalytic triads that directly participate in the chemical reaction.[4] The three dimensional structure of the enzyme orients these functional groups such that they can coordinate to lower the energy of transition states.[5] One difference between enzymes and traditional synthetic organic catalysts is that enzymes are capable of carrying out carrying out molecular recognition, which also depends on the three dimensional conformation of the enzyme. These processes have come to be understood in terms of an induced fit model.[6,7,8,9,10] Supramolecular catalysis is a field in organic chemistry inspired by enzymes that involves development of novel synthetic catalysts capable of molecular recognition[11,12] Within this domain, a key objective involves creating artificial host structures that can effectively control chemical reactivity and catalytic processes. Numerous host structures demonstrate supramolecular catalytic characteristics by developing their recognition abilities,[13] particularly when these attributes facilitate the stabilization of intermediate compounds or transition states.[14]
The emergence of effective catalysts for biochemical reactions is thought to be a crucial step in the origin and evolution of life.[15] While enzymes are prevalent in today’s cell biology, the discovery of catalytic RNA molecules, known as ribozymes, raised the idea that nucleic acids might have been fundamental to the beginnings of biocatalysts.[16] This is partly because RNA is crucial in the essential process of protein creation in all cells. The ’RNA world’ theory suggests that RNA once functioned as both the genetic code and the main biocatalyst in living organisms. As these early RNA-driven processes evolved, the need for more advanced enzymes with enhanced catalytic abilities likely prompted the shift to protein-based catalysis.[17] It’s reasonable that existing ribozymes in organisms, from bacteria to humans, are leftover traces from this hypothesized RNA-centric period, which implies that some tasks of contemporary enzymes were initially performed by ribozymes. [18]
In 2001, Lancet at al. introduced the concept of a “lipozyme” as a lipid aggregate that is capable of catalyzing chemical reactions in the context of the lipid world hypothesis for the origin of life.[19,20] Numerous authors have documented instances of chemical reactions that exhibit accelerated rates when specific lipid micelles or vesicles are present.[21,22] While most documented instances of micellar catalysis pertain to hydrolytic reactions, there have also been demonstrations of enhanced reaction rates in synthetic processes. This includes cases such as the synthesis of non-biological surfactants and the oligomerization of amino acids catalyzed by liposomes.[15,23]
While enzymes and synthetic supramolecular catalysts demonstrate molecular recognition through molecular binding events, another way of thinking about this goal is through selective partitioning. For instance, we’ve previously demonstrated partitioning-based molecular recognition in a two phase octanol/water system.[24] In that model system, the concentration of organic solutes dissolved in the organic phase were found capable of regulating charge-driven selective partitioning of two water soluble dyes into the organic phase with both kinetic and thermodynamic mechanisms.[25] Similarly, we used a combinatorial materials approach to identify synergistic extraction conditions for extraction of copper ions from an aqueous phase into an oleic acid organic phase. [26] Phospholipids in bilayer or monolayer form have also been shown to respond selectively to small molecule partitioning, with implications in understanding membrane function.[27,28]
Here we demonstrate the concept of a lipozyme in model systems including an octanol/water two phase system and in phospholipid vesicles. We demonstrate the possibility of a lipid-bilayer-based catalysis of ester hydroloysis, i.e. esterase activity using an established indicator. Esterases are a ubiquitous class of enzymes that typically hydrolize esters through a base catalyzed mechanism.[29,30] Calcein acetoxymethyl ester (Calcein-AM) is a widely used indicator of esterase activity in cells.[31] Calcein-AM is relatively hydrophobic and not fluorescent but is converted to the anionic fluorophore calcein when taken up by cells and exposed to intracellular esterases. We use this compound as an indicator of esterase activity of a phase-transfer based lipozyme model. This new way of thinking about lipozymes as catalytic entitities suggests the importance of considering local lipid composition of lipid droplets and bilayers.
2. Results
The chemical structures of the main compounds used here, and schematics of the two phase systems where the reactions are observed are shown in Figure 1. Calcein acetomethylester (calcein AM) is a colorless hydrophobic dye that contains several ester groups. Calcein is the expected fluorescent product when the ester groups on calcein AM are cleaved, for instance by intracellular esterases.[32] In particular, hydrolysis of the two esters on the xanthane backbone and opening of the lactone ring lead to aromatic conjugation associated with the fluorescent properties of calcein.[33] Octanol was chosen as the organic phase for bulk phase transfer catalysis because it has a polarity similar to that of the hydrophobic region of a lipid bilayer, and for this reason it is widely used as a standard for characterizing the partitioning of pharmaceuticals.[34] Octadecylamine (ODA) was found to function as an efficient catalyst when included in the organic phase of the two phase system. The decision to utilize ODA was primarily driven by its primary amine group, which can be expected to both facilitate the partitioning of the anionic product into the non-polar phase and participate in a base catalyzed reaction analagous to the histitidine residues in active sites of esterases.[30] DOPC (1,2-dioleoyl-sn-glycero-3-phosphocholine) vesicles were used to replace octanol component as a non-polar hydrophobic environment suitable for catalysis.
Figure 1b-left shows the schematic of reactions in the octanol/water two phase system. In this case we dissolved ODA in octanol and calcein AM in aqueous solution. The calcein AM molecules transfer from the aqueous phase to the organic or hydrophobic phase where they react in the presence of ODA, resulting in the formation of a fluorescent product such as calcein via ester hydrolysis. Figure 1b-right shows a schematic of reaction in the presence of a lipid bilayer. ODA was added to the DOPC prior to vesicle formation. When the calcein AM was added to water, it was observed to enter the lamellar lipid bilayer phase and react only in the presence of ODA to generate the fluorescent product.
To observe the hydrolysis reaction, we first apply a flow cell setup as illustrated in Figure 2c. We spot the lipid droplets on the glass slide, then add a spacers and a second glass slide to form a flow cell. The aqueous calcein AM solution then flows into the cell chamber. Figure 2 shows the hydrolysis of calcein AM results using 60 mM ODA in octanol doped with 1 mol% of the fluorescently labeled phospholipid rhodamine PE (1,2-dioleoyl-sn-glycero-3-phosphoethanolamine-N-(lissamine rhodamine B sulfonyl) (ammonium salt) ) for visulazation of the organic phase. From Figure 2a, we notice that the green intensity increases after 60 minutes whereas the red intensity as an internal standard doesn’t change. This is evidence that the hydrolysis of calcein AM happens in the presense of ODA in octanol droplets. Control droplets of octanol without ODA did not show catalysis. Figure 2b,c shows the average green and green over red intensity along with time within the octanol droplet. Synthetic calcein was found to partition into octanol drops that containing ODA (Figure S1).
To quantitatively determine the concentration of the product generated after the hydrolysis of calcein AM, we calibrated the fluorescence using calcein. This calibration was then used to calculate the catalytic turnover of the lipozyme. The catalytic turnover refers to the quantity of substrate molecules that an enzyme molecule converts into product within a specific time. This value is determined by the maximum velocity of the enzyme-catalyzed reaction, which occurs when the enzyme is completely saturated with substrate. We perform a calibration curve to measure the fluorescence intensity of different concentrations of calcein in the aqueous solution as shown in Figure 2d. Then we calculate the catalytic turnover from the maximum reaction rate which is measured as the slope of the change in fluorescence intensity in the first 2 minutes of the reaction . Based on the calibration curve in Figure 2d, we convert this value to a reaction rate in units of change in concentration per time for 60 mM ODA. Finally, the catalytic turnover for ODA equals the max velocity divided by the concentration of ODA, which gives us . This value is comparable to that of a relatively slow enzyme.[35,36]
In order to test whether the reaction is catalyzed by the phase transfer agent or is possibly the result of aminolysis, we carried out the reaction using the quaternary ammonium salts cetrimonium bromide (CTAB) and cetrimonium chloride (CTACL) instead of ODA. These quanternary ammonium salts are not capable of participating in aminolysis. Figure 3 shows that octanol drops containing either of these two amonium salts significantly increase in fluorescence intensity within 30 minutes, indicating ester hydrolysis of calcein-AM. Again, octanol drops that do not contain the amines do not show any color change.Consequently, it is reasonable to conclude that lipid combinations are capable of catalyzing the ester hydrolysis reaction and the reaction is not the result of aminolysis. However, from this experiment, we can’t exclude the possibility that the CTAB or CTACL might transfer another catalyst from the PBS buffered aqueous phase into the organic phase.
Figure 4 shows the hydrolysis of calcein AM in surface supported lipid multilayer films composed of DOPC with 10 mol% ODA. We also included 1% Rhodamine PE into the DOPC as an internal standard. The lipid formulations were mixed in chloroform and spotted onto a glass slide, the chloroform was evaporated in a vacuum, and then a flow cell constructed by adding a top glass slide to form the flow cell. An aqueous solution of calcein AM was then added to the flow cell and the lipid multilayers were monitored by fluorescence microscopy. In Figure 4a, the red intensity of the TRITC filter remains relatively unchanged between t = 0 mins and t = 60 mins. However, the green intensity of the FITC filter shows an increase in the same time frame. Figure 4b clearly indicates that the intensity green/red rise over time, and the green intensity increase shown in Figure S2, indicating hydrolysis of the calcein AM. Phospholipid multilayers without ODA did not show any increase in fluorescence. We used the red intensity as a measure of droplet thickness[37] and plotted its relationship with the green intensity normalized to the thickness at t = 60 mins, as depicted in Figure 4d. This plot reveals a linear correlation between droplet thickness and green intensity, with a slope of 0.082, closely aligning with the 0.06 value shown in Figure 4b. It’s worth noting that our analysis is based on 40-pixel values within the droplets, leading to a minor variance between the slope and the actual green/red intensity at t = 60 mins due to the limited data points. This indicates that the phospoholipid DOPC is a suitable replacement for the octanol for this phase transfer catalysis reaction, with the fluorescent product partitioning into the phospholipid multilayer volume.
Its apparent that the fluorescence product partitions into the lipid multilayer, and in order to quantify the catalytic turnover in the lipid-based system we would like to distinguish partitionoing from the catalysis. We therefore carried out the experiment in vesicles using a bulk solution and measured fluorescence intensity in a plate reader. Multilamellar vesicle solutions were prepared as follows. We first dissolved different concentrations of ODA with DOPC and let the chloroform dry to leave a thin film of the product and evaporated any residual solvent in a vacuum chamber for 10 minutes. After that, the PBS buffer solution was added into the centrifuge tubes and vortexed then sonicated to make multilamellar vesicles. Calcein AM solution was then added to the vesicle solution and fluorescence was measured over time. Figure 5 shows the result of the kinetic of fluorescence intensity of calcein (10 µM) measured by a plate reader. From the data, we notice that the initial changing rate increases as the concentration of ODA increases. So, we calculated the maximum reaction rate by using a linear fitting for the first five data points. Figure 5b shows the relationship of ODA concentration with the maximum reaction rate. From Figure 5b, we noticed that the maximum reaction rate increases with the concentration of ODA in the linear region, and the equilibrium intensities for different concentrations of ODA reaching the same level, which demonstrates that the usage of biological molecule can catalyze the chemical reaction[38] in the vesicles system of calcein AM. We fit the curve in the linear region and get the catalytic turnover rate in Figure 5b based on the calibration curve in Figure 5c, and the catalytic turnover rate equals to , which is comparable to a relative slow enzyme.[39,40] However, it is likely the rate could be increased by optimizing the composition of the lipozyme in more heterogeneous systems. Figure 5d displays the fluorescence results for ODA vesicles with and without DOPC. The data reveal that the fluorescence intensity remains constant over time when DOPC is absent, indicating that the combination of ODA and DOPC in the vesicles is crucial for catalyzing the hydrolysis of calcein AM.
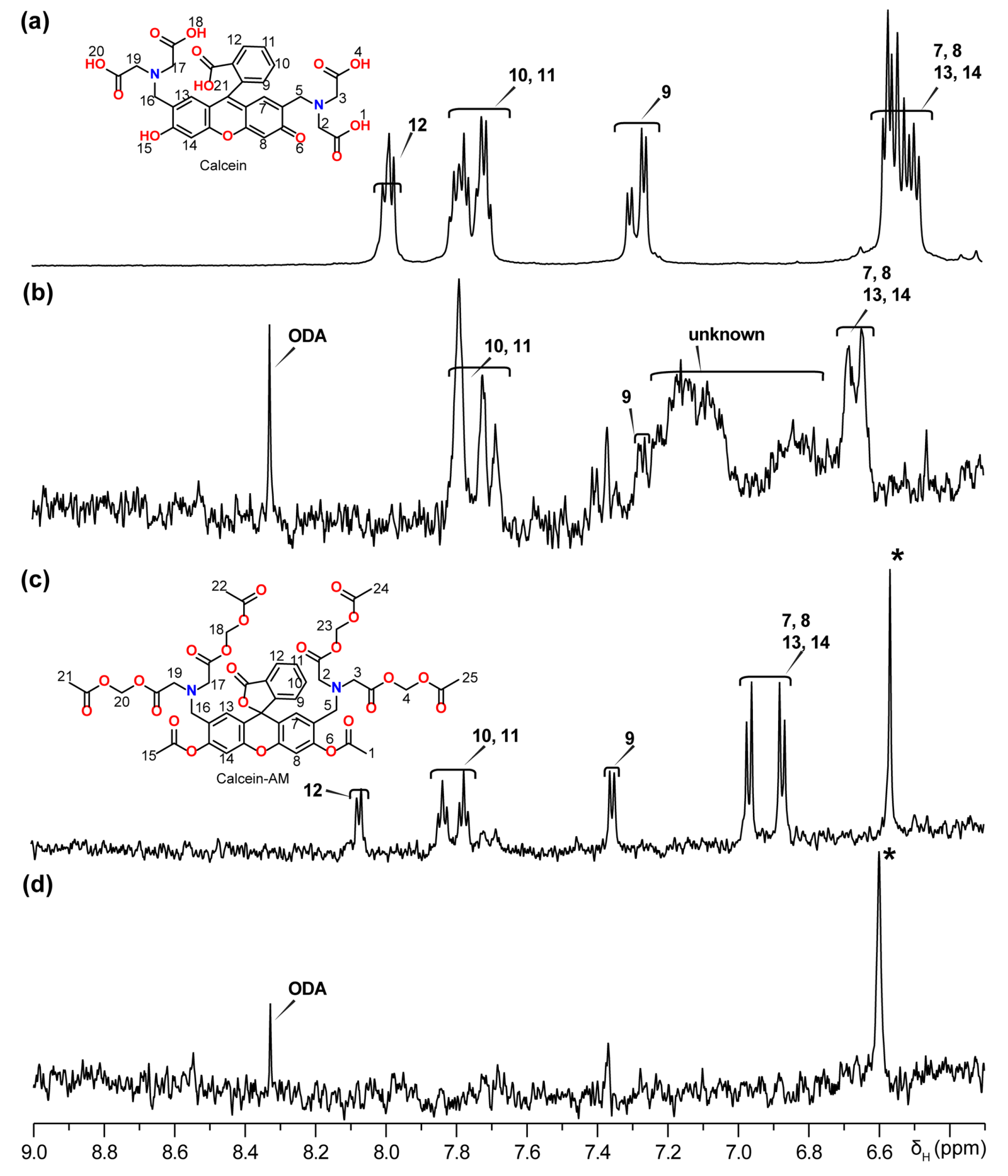
H-NMR experiments were used to further characterize the ester hydrolysis reaction. H NMR spectra are shown in Figure S3. Since the concentration of lipids used were much higher than calcein-AM, we focused our analysis only in the aromatic region that is devoid of any peaks from lipids. Figure 6 shows the aromatic regions of H-NMR spectra of synthetic calcein as a positive control (Figure 6a), the reaction product after incubating calcein AM in DOPC/ODA vesicles (Figure 6b), pure calcein-AM as a negative control (Figure 6c), and DOPC/ODA vesicles without calcein-AM (Figure 6d). Table S1 summarizes the chemical shift values measured for assignment of aromatic peaks based on reported NMR spectra for eosin Y, a similar molecule,[41] and 2D COSY & TOCSY NMR assignment (Figure S4 and S5). In order to detect signals from calcein-AM and hydrolyzed compounds, higher concentrations (100-500 µM) and volumes (600 µL) of the calcein-AM solutions were needed in the NMR samples than that used in previous experiments. Due to the high cost of calcein-AM we had to use a lower concentration (100 µM) which resulted in lower signal-to-noise ratio spectra for solutions requiring this compound. Interestingly, the DOPC + ODA with calcein-AM sample exhibit H NMR peaks corresponding to both the reactant (i.e. calcein-AM) and product (i.e. similar to calcein) in addition to peaks from DOPC and ODA. Peaks 7, 8, 13, and 14 appear in different locations in calcein and calcein-AM, which is understandable due to the different aromatic nature of the xanthene core. In the reaction product We also identified peaks 9, 10, 11 corresponding to both calcein and calcein AM with significant overlap. It is noticeable that peak 12 does not appear with the same chemical shift in reaction product and a precise assignment of peak 12 was not possible in the reaction product due to peak shift and peak overlap. This shift or overlap may be explained as the calcein produced from reaction likely in equilibrium with lactone ring close-open structures, or possibly due to the ODA containing a positively charged amino group which could serve as a counter-ion to neutralize the carboxylic acid group.[33] The observed broad H peak(s) in the aromatic region of the DOPC + ODA and calcein-AM sample is likely due to presence of more than one type of “calcein-like” molecules. Although the mechanism of the reaction remains elusive, based on the H-NMR results and fluorescence data, we conclude that the reaction product contains a product with a calcein-like aromatic region (Figure 6b). The fluorescence generated by the product can be used as an indicator of the hydrolysis.
We aimed to assess whether the amino group alone drives the catalysis of calcein AM hydrolysis. To this end, we conducted experiments using solutions of calcein AM with ethylamine, as both ethylamine and octadecylamine possess the same amino group. Figure S6 presents the results from the plate reader for 10 M calcein AM with 66% liquid ethylamine, and pure calcein AM as a control. It was observed that the calcein AM with ethylamine did not catalyze the reaction. This lack of activity might be attributed to ethylamine being in liquid form at room temperature, unlike solid octadecylamine. This, along with the observation that ODA without an organic phase or lipid bilayer also does not catalyze the reaction indicates that a non-polar environment is necessary for the reaction to occur.
3. Discussion
We observed the hydrolysis of calcein AM in aqueous form within an octanol-water phase system, as well as in lipid multilayer and vesicle solutions in the presence of the cationic amphiphiles octadecylamine or CTAB. The catalytic turnover numbers determined by quantiative fluorescence measurements were found to be on a scale of for each system which is comparable to some slow enzymes. Although it is worth noting that the fluorescence intensities were calibrated in aqueous solution, while the brightness of the fluorescent product may be sensitive to the non-polar environment, the values obtained were found to be reproducible in both the octanol droplet and the vesicle-based systtems. These values therefore suggest that lipid aggregates or lipozymes have the potential to catalyze chemical reactions at comparable rates to some enzymes. As the cationic amphiphiles by themselves proved ineffective in catalyzing the reaction, the presence of a non-polar or hydrophobic environment appears crucial to the reaction mechanism.
The idea of organic aggregates speeding up chemical reactions is well established in synthetic organic chemistry. For example, phase-transfer catalysis is a valuable method used extensively in various chemical fields. This technique facilitates reactions between reagents that are in different phases and might not readily interact due to their separation. By introducing a phase transfer agent, one of the reagents is moved to a position where it can efficiently react with another.[42,43,44] CTAB (cetyltrimethylammonium bromide)[45] and ODA (octadecylamine)[46,47] have been pivotal as early phase transfer catalysts in chemical processes.[48] ODA, for instance, has been effectively used to transfer platinum nanoparticles from water to organic solvents by forming a hydrophobic layer around the nanoparticles, enabling their dispersion in nonpolar environments. This technique underscores the significant role of such catalysts in both academic research and industrial applications.[49,50] However, in our system, the amine or ammonium groups are introduced into the organic phase. We can’t exclude the possibility that they may be bringing a counterion such as chlorine, bromide, or other ion from the aqueous phase that may participate in the reaction. However, as synthetic calcein was observed to preferentially partition into the ODA containing octanol phase, it seems likely that the cationic amphiphiles may stabilize the product in the hydrophobic environment by neutralizing the charge of the carboxylic acids on the hydrolysed product. The precise mechanism of the reaction described here, however remains undetermined.
Considering that the number of possible lipid mixtures in a cell scales in a way comparable to nucleic acid or protein sequence information, [19,24] it is likely that these rates could be significantly increased through combinatorial formulation.[26] As lipid formulations have been shown to be capable of partitioning-based molecular recognition,[25,27,28] lipid composition may play a larger role in catalysis than simple bilayer compartmentalization. This suggests that there may be value in looking for reactions that may take place in hydrophobic environments in vivo, and to consider the role of local lipid composition and heterogeneity in metabolism and signaling.[51] For example Coenzyme Q [52] and vitamin[53] K are examples of lipids with functional groups that directly participate in well characterized enzymatic reactions, and are therefore good candidates for the identification of lipozymes in vivo.
Lipozymes have several implications for pharmaceuticals. First, if they could be identified in vivo may serve as innovative drug targets. As the majority of FDA approved drugs are lipophilic, it is possible that some of them may already function by affecting local lipid composition. Second, as lipids are widely used as for drug delivery and formulation,[54] the idea that certain formulation may catalyze reactions could be used for therapeutic benefit. For instance, lipozymes could be designed to mimic the specificity and efficiency of traditional enzymes but in non-aqueous environments. This opens up possibilities for developing new drug delivery systems where the drug release can be controlled catalytically within lipid-based carriers.[55]
Finally, as the idea of a lipozyme was conceived in the context of origin of life research, the possibility of lipid-based catalysis has implications for synthetic biology. Efforts to modify organisms for technological applications or build new life-like systems from a bottom up approach can benefit from non-polymeric catalysis. It is still somewhat of a mystery why cells invest so many resources into generating highly diverse lipids, with approximately 5% of existing organisms’ genes being used to generate the lipidome.[56] The presence of lipozymes could justify this investment. Phase separated lipid based systems such as micelles, vesicles and droplets of highly heterogeneous composition are promising systems for the emergence of biological phenomena.
4. Methods
4.1. Materials
Calcein acetoxymethyl ester (calcein AM) was purchased from VWR (4591070). ODA (L15458) was purchased from Alfa Asear Chemical (Ward Hill, MA). CTAB (H5882) and calcein (154071) were purchased from Millipore-Sigma (St. Louis, USA). DOPC (850375) and Rhodamine PE (810150) were purchased from Avanti polar lipids. 1-Octanol (222920) was purchased from Beantown Chemicals. Chloroform (for HPLC, >= 99.9%) was purchased from Sigma-Aldrich (650498).
4.2. Vesicle Preparation
Multilamellar vesicle solutions were prepared as follows. 1 mM stock solutions of calcein AM were prepared. These solutions were diluted 100x with PBS buffer to result in 10 M solutions. DOPC and ODA solutions in chloroform were prepared and the chloroform was evaporated in a vacuum at 10 mbar for 5 minutes. We maintained a fixed quantity of DOPC molecules and adjusted the volume of ODA to alter the molecular ratio between DOPC and ODA for testing the reaction with calcein AM.
4.3. Fluorescence Microscopy
Fluorescence microscopy was done using a Ti-E inverted microscope (Nikon Instruments, Melville, NY, USA) to measure the optical response of the droplets. A 4x objective lens was used for magnification. The exposure time was set to 1 second for both the TRITC filter (excitation at 544 nm, emission at 570 nm) and the FITC filter (excitation at 490 nm, emission at 515 nm) to measure the fluorescence intensity of the droplets. The lamp intensity was set to 16 on the ND scale, corresponding to a transmittance of 6.25%.
4.4. Fluorescence Characterization
Fluorescence was measured using a Molecular Devices plate reader integrated with the SoftMax Pro software. The lipid vesicle solution was put in the 96 well plate. We use the kinetics function to measure the absorbance and fluorescence of the lipid vesicles for more than 10 hours. We select a medium gain setting to measure the relative fluorescence intensity of the solutions. If the intensity reaches its maximum, we switch to a low gain setting and perform another measurement. The fluorescence function is measured with an excitation wavelength of 480 nm and emission wavelength at 520 nm. The absorbance measurement was done using a scanning function of wavelength from 230 nm to 1000 nm with step 10 nm.
4.5. Flow Cell Setup
The droplet of mixtures were spotted onto the substrate. After that, we use polydimethylsiloxane (PDMS) as the spacer and cover the top with a glass slide. The aqueous solution flowed into the cell, and we recorded the optical response of the droplet arrays using the fluorescence microscope.
4.6. NMR
DOPC and ODA vesicle solutions were prepared as described previously and exposed to calcein AM. Once the reaction was completed, sample were lyophilized overnight to remove any residual water and were redissolved in deuterated dimethyl sulfoxide (DMSO-d6). Control calcein AM, calcein and DOPC/ODA samples were directly dissolved in DMSO. NMR was experiments were carried out on a 600 MHz NMR spectrometer equipped with a broadband BBI probe. Two-dimensional COSY and TOCSY spectra were recorded using standard pulse programs available in the NMR spectrometer’s library with 16 scans and 128 or 256 complex points. A relaxation delay of 2 s was used throughout 2D experiments. DMSO solvent peak was used as an internal reference for H chemical shifts. All NMR data were processed with Topspin 4.3.0 software (from Bruker).
Author Contributions
S. Liu and conducted most experiments under the guidance of D.V.W. and S.L.; T.B. characterized the partitioning behavior of calcein. K.K. carried out the NMR experiments under the supervision of A.R.; S.L. conceived of the study and directed the research. All authors contributed to the writing of the manuscript.
Acknowledgments
S. Lenhert thanks Doron Lancet for the inspiration and critical discussion. S. Liu, and S. Lenhert thank Jack Saltiel, Edwin Hilinski, Sumesh Krishnan, and Mohammad Khizr for the helpful discussion regarding the organic chemistry. S. Liu, TB, and S. Lenhert thank Huanhuan Zhou and Vincent Tocci for the discussion about the experiment and manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Supuran, C.T.; Winum, J.Y. Introduction to zinc enzymes as drug targets; Wiley, Hoboken, 2009.
- Robertson, J.G. Mechanistic basis of enzyme-targeted drugs. Biochemistry 2005, 44, 5561–5571. [Google Scholar] [CrossRef] [PubMed]
- Copeland, R.A.; Harpel, M.R.; Tummino, P.J. Targeting enzyme inhibitors in drug discovery. Expert opinion on therapeutic targets 2007, 11, 967–978. [Google Scholar] [CrossRef] [PubMed]
- Dodson, G.; Wlodawer, A. Catalytic triads and their relatives. Trends in biochemical sciences 1998, 23, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Schramm, V.L. Transition states and transition state analogue interactions with enzymes. Accounts of Chemical Research 2015, 48, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Koshland Jr, D.E. Application of a theory of enzyme specificity to protein synthesis. Proceedings of the National Academy of Sciences 1958, 44, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Koshland Jr, D.E. The key–lock theory and the induced fit theory. Angewandte Chemie International Edition in English 1995, 33, 2375–2378. [Google Scholar] [CrossRef]
- Rebek Jr, J. Molecular recognition with model systems. Angewandte Chemie International Edition in English 1990, 29, 245–255. [Google Scholar] [CrossRef]
- Bosshard, H.R. Molecular recognition by induced fit: how fit is the concept? Physiology 2001, 16, 171–173. [Google Scholar] [CrossRef] [PubMed]
- Boehr, D.D.; Nussinov, R.; Wright, P.E. The role of dynamic conformational ensembles in biomolecular recognition. Nature chemical biology 2009, 5, 789–796. [Google Scholar] [CrossRef]
- Ballester, P.; Scarso, A. Supramolecular Aspects in Catalysis, 2019.
- Wei, H.; Wang, E. Nanomaterials with enzyme-like characteristics (nanozymes): next-generation artificial enzymes. Chemical Society Reviews 2013, 42, 6060–6093. [Google Scholar] [CrossRef]
- Steed, J.W.; Atwood, J.L. Supramolecular chemistry; John Wiley & Sons, 2022.
- Raynal, M.; Ballester, P.; Vidal-Ferran, A.; Van Leeuwen, P.W. Supramolecular catalysis. Part 2: artificial enzyme mimics. Chemical Society Reviews 2014, 43, 1734–1787. [Google Scholar] [CrossRef] [PubMed]
- Kahana, A.; Lancet, D. Self-reproducing catalytic micelles as nanoscopic protocell precursors. Nature Reviews Chemistry 2021, 5, 870–878. [Google Scholar] [CrossRef] [PubMed]
- Doherty, E.A.; Doudna, J.A. Ribozyme structures and mechanisms. Annual review of biochemistry 2000, 69, 597–615. [Google Scholar] [CrossRef] [PubMed]
- Doudna, J.A.; Lorsch, J.R. Ribozyme catalysis: not different, just worse. Nature structural & molecular biology 2005, 12, 395–402. [Google Scholar]
- Chen, X.; Li, N.; Ellington, A.D. Ribozyme catalysis of metabolism in the RNA world. Chemistry & biodiversity 2007, 4, 633–655. [Google Scholar]
- Segré, D.; Ben-Eli, D.; Deamer, D.W.; Lancet, D. The lipid world. Origins of Life and Evolution of the Biosphere 2001, 31, 119–145. [Google Scholar] [CrossRef] [PubMed]
- Lancet, D.; Segrè, D.; Kahana, A. Twenty years of “lipid world”: a fertile partnership with David Deamer. Life 2019, 9, 77. [Google Scholar] [CrossRef] [PubMed]
- Fendler, J.H. Interactions and reactions in reversed micellar systems. Accounts of Chemical Research 1976, 9, 153–161. [Google Scholar] [CrossRef]
- Cuccovia, I.M.; Quina, F.H.; Chaimovich, H. A remarkable enhancement of the rate of ester thiolysis by synthetic amphiphile vesicles. Tetrahedron 1982, 38, 917–920. [Google Scholar] [CrossRef]
- Kust, P.R.; Rathman, J.F. Synthesis of surfactants by micellar autocatalysis: N, N-dimethyldodecylamine N-oxide. Langmuir 1995, 11, 3007–3012. [Google Scholar] [CrossRef]
- Bell, T.N.; Feng, K.; Calvin, G.; Van Winkle, D.H.; Lenhert, S. Organic composomes as supramolecular aptamers. ACS omega 2020, 5, 27393–27400. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Shiel, E.; Bell, T.; Lin, S.; Lenhert, S. Kinetic Mechanism of Surfactant-Based Molecular Recognition: Selective Permeability across an Oil–Water Interface Regulated by Supramolecular Aggregates. The Journal of Physical Chemistry B 2023, 127, 10201–10214. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wei, A.T.; Wang, H.; Van Winkle, D.; Lenhert, S. Combinatorial mixtures of organic solutes for improved liquid/liquid extraction of ions. Soft Matter 2023, 19, 6903–6910. [Google Scholar] [CrossRef] [PubMed]
- Lowry, T.W.; Kusi-Appiah, A.E.; Fadool, D.A.; Lenhert, S. Odor discrimination by lipid membranes. Membranes 2023, 13, 151. [Google Scholar] [CrossRef] [PubMed]
- Yotsumoto, M.; Matsuo, M.; Kitahata, H.; Nakanishi, S.; Denda, M.; Nagayama, M.; Nakata, S. Phospholipid Molecular Layer that Enhances Distinction of Odors Based on Artificial Sniffing. ACS sensors 2023, 8, 4494–4503. [Google Scholar] [CrossRef] [PubMed]
- Rafeeq, H.; Hussain, A.; Shabbir, S.; Ali, S.; Bilal, M.; Sher, F.; Iqbal, H.M. Esterases as emerging biocatalysts: Mechanistic insights, genomic and metagenomic, immobilization, and biotechnological applications. Biotechnology and applied biochemistry 2022, 69, 2176–2194. [Google Scholar] [CrossRef] [PubMed]
- Barzkar, N.; Sohail, M.; Tamadoni Jahromi, S.; Gozari, M.; Poormozaffar, S.; Nahavandi, R.; Hafezieh, M. Marine bacterial esterases: Emerging biocatalysts for industrial applications. Applied Biochemistry and Biotechnology 2021, 193, 1187–1214. [Google Scholar] [CrossRef] [PubMed]
- Bratosin, D.; Mitrofan, L.; Palii, C.; Estaquier, J.; Montreuil, J. Novel fluorescence assay using calcein-AM for the determination of human erythrocyte viability and aging. Cytometry Part A: the journal of the International Society for Analytical Cytology 2005, 66, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Uggeri, J.; Gatti, R.; Belletti, S.; Scandroglio, R.; Corradini, R.; Rotoli, B.M.; Orlandini, G. Calcein-AM is a detector of intracellular oxidative activity. Histochemistry and cell biology 2000, 122, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Klonis, N.; Sawyer, W.H. Spectral properties of the prototropic forms of fluorescein in aqueous solution. Journal of fluorescence 1996, 6, 147–157. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Advanced drug delivery reviews 1997, 23, 3–25. [Google Scholar] [CrossRef]
- Boudart, M. Turnover rates in heterogeneous catalysis. Chemical reviews 1995, 95, 661–666. [Google Scholar] [CrossRef]
- Zhou, X.; Xu, W.; Liu, G.; Panda, D.; Chen, P. Size-dependent catalytic activity and dynamics of gold nanoparticles at the single-molecule level. Journal of the American Chemical Society 2010, 132, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Nafday, O.A.; Lenhert, S. High-throughput optical quality control of lipid multilayers fabricated by dip-pen nanolithography. Nanotechnology 2011, 22, 225301. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.E.; MacMillan, D.W. Synergistic catalysis: a powerful synthetic strategy for new reaction development. Chemical science 2012, 3, 633–658. [Google Scholar] [CrossRef] [PubMed]
- Costentin, C.; Drouet, S.; Robert, M.; Saveant, J.M. Turnover numbers, turnover frequencies, and overpotential in molecular catalysis of electrochemical reactions. Cyclic voltammetry and preparative-scale electrolysis. Journal of the American Chemical Society 2012, 134, 11235–11242. [Google Scholar] [CrossRef] [PubMed]
- Pegis, M.L.; McKeown, B.A.; Kumar, N.; Lang, K.; Wasylenko, D.J.; Zhang, X.P.; Raugei, S.; Mayer, J.M. Homogenous electrocatalytic oxygen reduction rates correlate with reaction overpotential in acidic organic solutions. ACS Central Science 2016, 2, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Okuom, M.O.; Wilson, M.V.; Jackson, A.; Holmes, A.E.; et al. Intermolecular Interactions between Eosin Y and Caffeine Using 1 H-NMR Spectroscopy. International journal of spectroscopy 2013, 2013. [Google Scholar] [CrossRef]
- Herriott, A.W.; Picker, D. Phase transfer catalysis. Evaluation of catalysis. Journal of the American chemical Society 1975, 97, 2345–2349. [Google Scholar] [CrossRef]
- Starks, C.M.; Halper, M. Phase-transfer catalysis: fundamentals, applications, and industrial perspectives; Springer Science & Business Media, 2012.
- Ooi, T.; Maruoka, K. Recent advances in asymmetric phase-transfer catalysis. Angewandte Chemie International Edition 2007, 46, 4222–4266. [Google Scholar] [CrossRef]
- Godha, A.K.; Thiruvengadam, J.; Abhilash, V.; Balgi, P.; Narayanareddy, A.; Vignesh, K.; Gadakh, A.V.; Sathiyanarayanan, A.; Ganesh, S. Environmentally benign nucleophilic substitution reaction of arylalkyl halides in water using CTAB as the inverse phase transfer catalyst. New Journal of Chemistry 2019, 43, 16041–16045. [Google Scholar] [CrossRef]
- Kumar, A.; Joshi, H.; Pasricha, R.; Mandale, A.; Sastry, M. Phase transfer of silver nanoparticles from aqueous to organic solutions using fatty amine molecules. Journal of colloid and interface science 2003, 264, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Wikander, K.; Petit, C.; Holmberg, K.; Pileni, M.P. Size control and growth process of alkylamine-stabilized platinum nanocrystals: a comparison between the phase transfer and reverse micelles methods. Langmuir 2006, 22, 4863–4868. [Google Scholar] [CrossRef] [PubMed]
- Starks, C.M. Phase-transfer catalysis. I. Heterogeneous reactions involving anion transfer by quaternary ammonium and phosphonium salts. Journal of the American Chemical Society 1971, 93, 195–199. [Google Scholar] [CrossRef]
- Kumar, A.; Joshi, H.M.; Mandale, A.B.; Srivastava, R.; Adyanthaya, S.D.; Pasricha, R.; Sastry, M. Phase transfer of platinum nanoparticles from aqueous to organic solutions using fatty amine molecules. Journal of Chemical Sciences 2004, 116, 293–300. [Google Scholar] [CrossRef]
- Xu, D.Q.; Pan, Z.W. Phase-transfer catalysis of a new cationic gemini surfactant with ester groups for nucleophilic substitution reaction. Chinese Chemical Letters 2014, 25, 1169–1173. [Google Scholar] [CrossRef]
- Tan, S.; Zhang, X.; Kong, W.; Yang, X.L.; Molnár, G.; Vondráková, Z.; Filepová, R.; Petrášek, J.; Friml, J.; Xue, H.W. The lipid code-dependent phosphoswitch PDK1–D6PK activates PIN-mediated auxin efflux in Arabidopsis. Nature Plants 2020, 6, 556–569. [Google Scholar] [CrossRef]
- Turunen, M.; Olsson, J.; Dallner, G. Metabolism and function of coenzyme Q. Biochimica et Biophysica Acta (BBA)-Biomembranes 2004, 1660, 171–199. [Google Scholar] [CrossRef]
- Stafford, D. The vitamin K cycle. Journal of Thrombosis and Haemostasis 2005, 3, 1873–1878. [Google Scholar] [CrossRef]
- Lokugamage, M.P.; Sago, C.D.; Gan, Z.; Krupczak, B.R.; Dahlman, J.E. Constrained nanoparticles deliver siRNA and sgRNA to T cells in vivo without targeting ligands. Advanced Materials 2019, 31, 1902251. [Google Scholar] [CrossRef]
- Lian, T.; Ho, R.J. Trends and developments in liposome drug delivery systems. Journal of pharmaceutical sciences 2001, 90, 667–680. [Google Scholar] [CrossRef] [PubMed]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: where they are and how they behave. Nature reviews Molecular cell biology 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Schematic showing the materials used in this study and the setups to demonstrate the hydrolysis of calcein AM. (a) Chemical structure of calcein AM, calcein, octadecylamine (ODA), dioleoylphosphatidylcholine (DOPC), and octanol. (b) Schematics of ester hydrolysis of calcein AM in octanol-water (left) and in lipid vesicle. (c) Schematic illustration of a flow cell setup used to observe the reaction.
Figure 1.
Schematic showing the materials used in this study and the setups to demonstrate the hydrolysis of calcein AM. (a) Chemical structure of calcein AM, calcein, octadecylamine (ODA), dioleoylphosphatidylcholine (DOPC), and octanol. (b) Schematics of ester hydrolysis of calcein AM in octanol-water (left) and in lipid vesicle. (c) Schematic illustration of a flow cell setup used to observe the reaction.
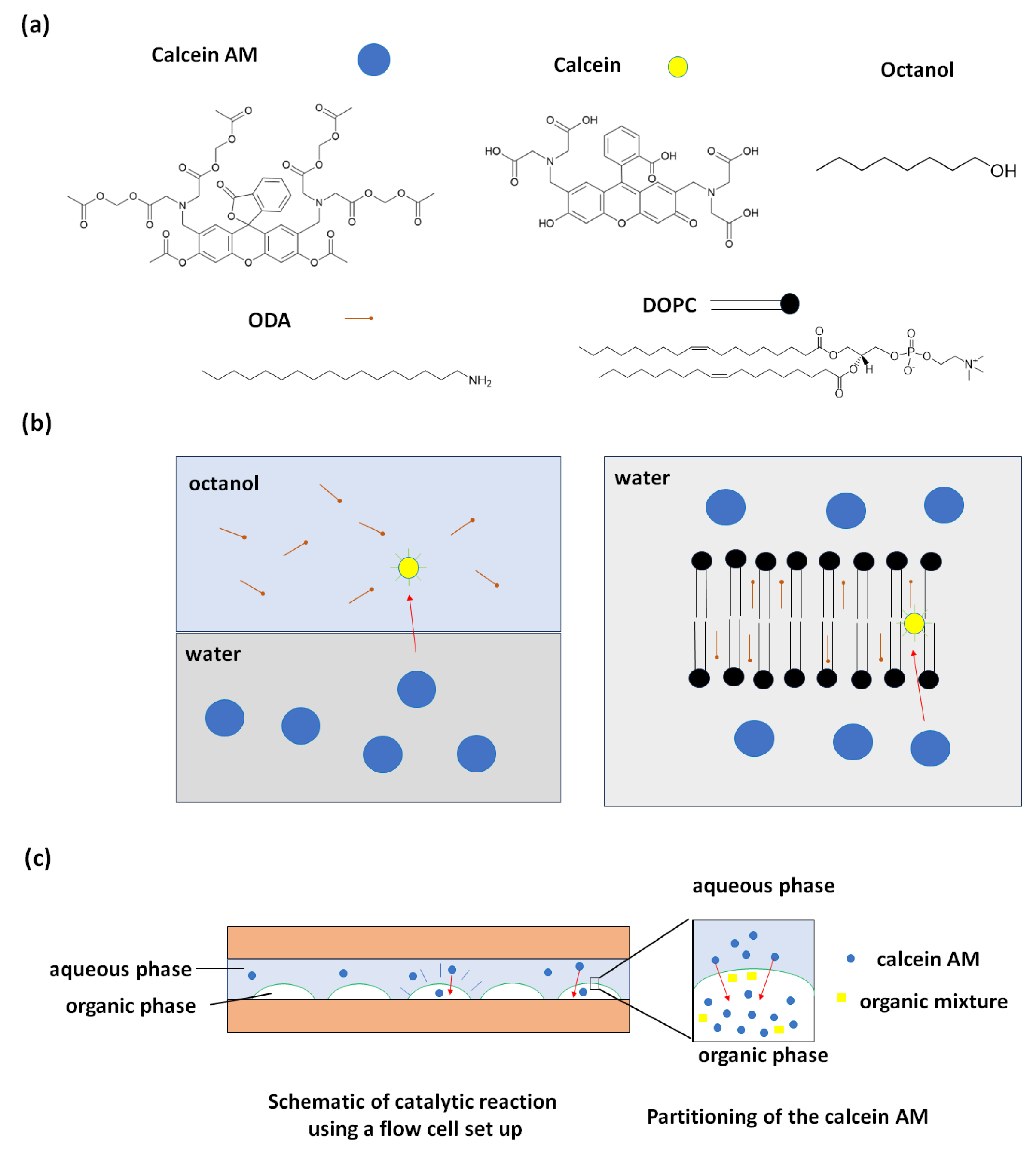
Figure 2.
Octanol droplets containing ODA become fluorescent when exposed to aqueous Calcein-AM (a) Images of ODA/octanol droplets at t = 0 mins and t = 60 mins under red, green filters and merged channels. (b) A plot of average green intensity along with time within the droplets. (c) A plot of average green over red along with time within the droplet (d) Calibration of calcein fluorescence in water detected on a fluorescence microscope. Top: images of different concertation of aqueous calcein. Bottom: a plot of average of fluorescence intensity of 16 pixels in the center of the droplet.
Figure 2.
Octanol droplets containing ODA become fluorescent when exposed to aqueous Calcein-AM (a) Images of ODA/octanol droplets at t = 0 mins and t = 60 mins under red, green filters and merged channels. (b) A plot of average green intensity along with time within the droplets. (c) A plot of average green over red along with time within the droplet (d) Calibration of calcein fluorescence in water detected on a fluorescence microscope. Top: images of different concertation of aqueous calcein. Bottom: a plot of average of fluorescence intensity of 16 pixels in the center of the droplet.
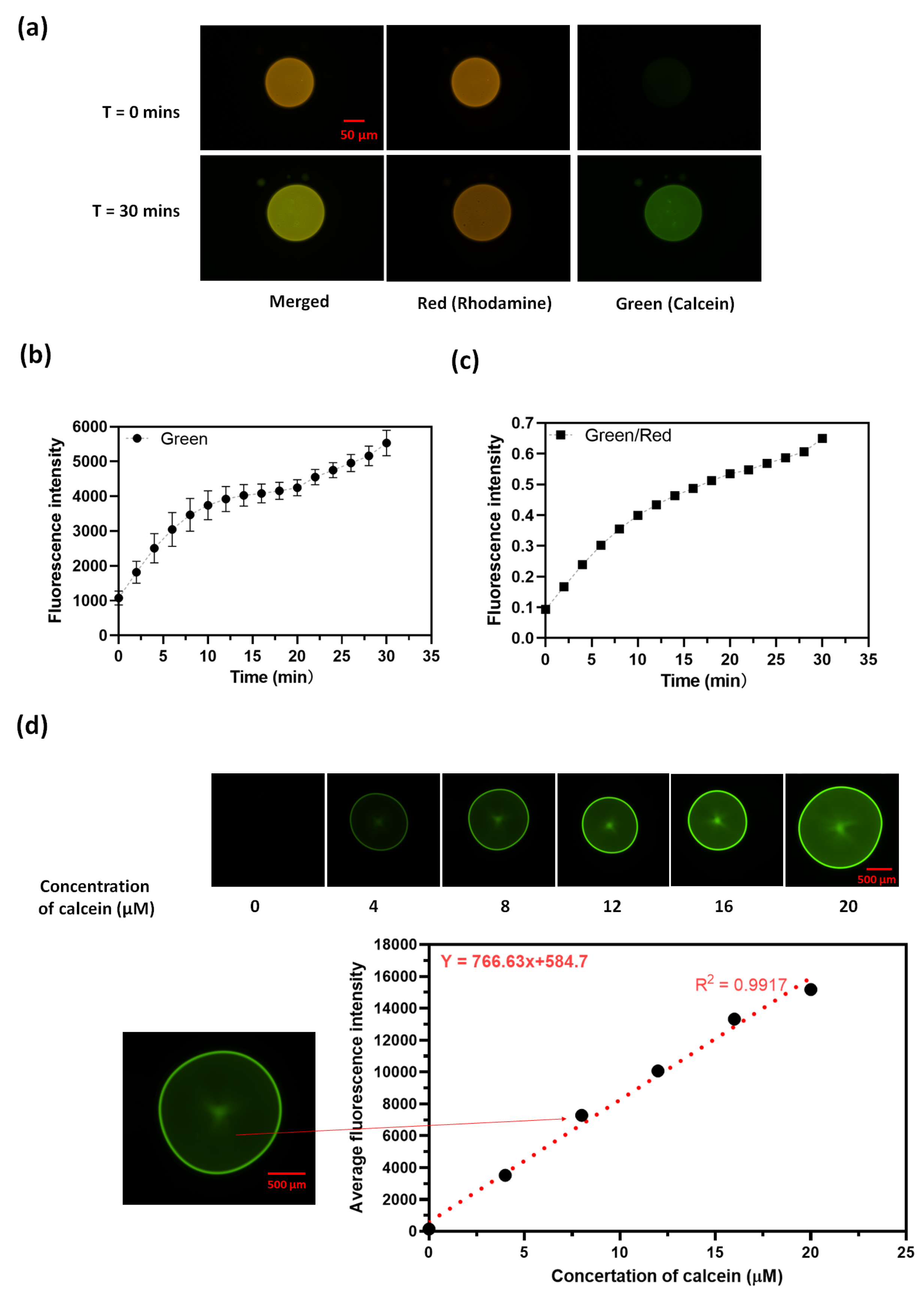
Figure 3.
Octanol droplets containing CTAB or CTACL become fluorescent when exposed to aqueous calcein AM (a) Images of CTAB and CTACL/octanol droplets at t = 0 mins and t = 60 mins under green filters. (b) A plot of average green intensity along with time within the droplets.
Figure 3.
Octanol droplets containing CTAB or CTACL become fluorescent when exposed to aqueous calcein AM (a) Images of CTAB and CTACL/octanol droplets at t = 0 mins and t = 60 mins under green filters. (b) A plot of average green intensity along with time within the droplets.
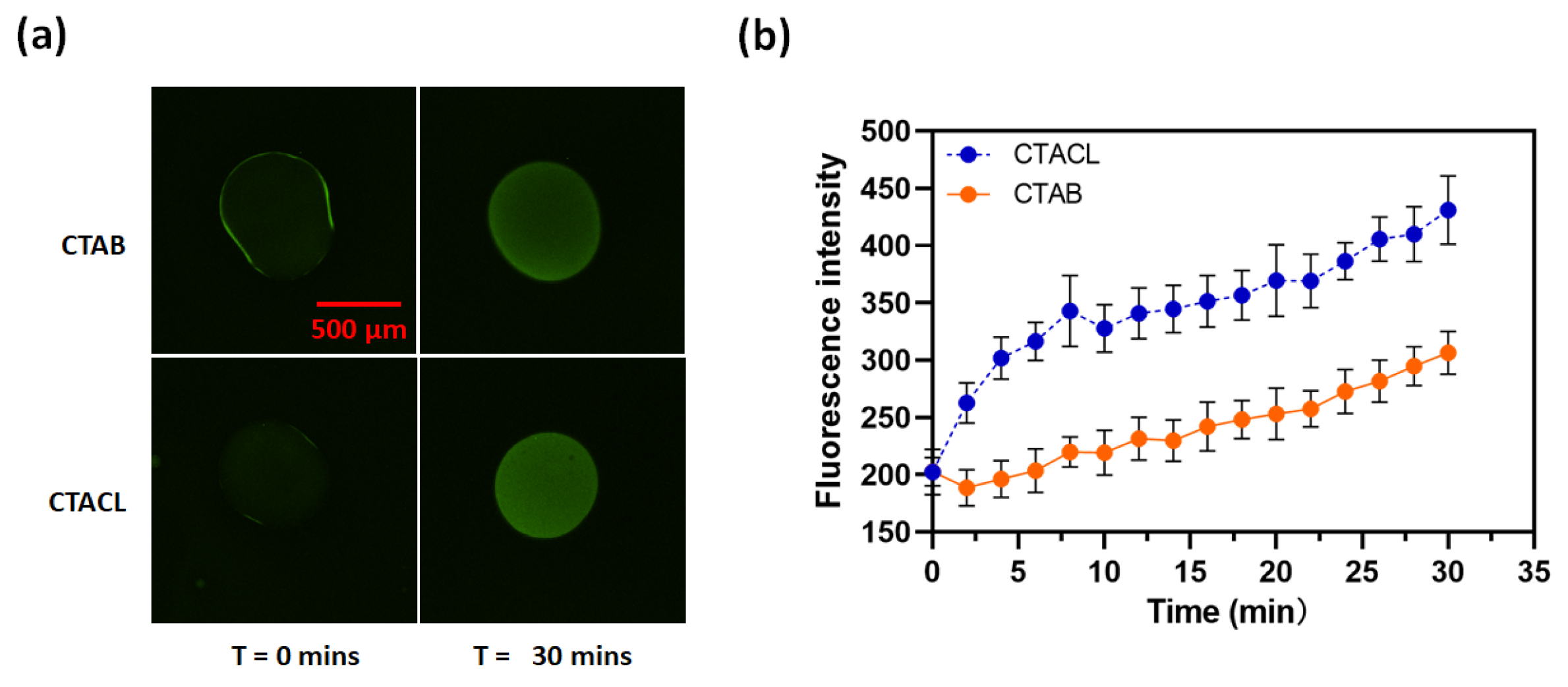
Figure 4.
Phospholipid multilayers containing octadecylamine fluoresce when exposed to aqueous calcein AM. (a) Images of ODA/DOPC droplets at t = 0 mins and t = 60 mins under red, green filters and merged channels. (b) A plot of average green/red intensity along with time within the droplets. (c) A plot of fluorescence intensity with thickness within the droplet.
Figure 4.
Phospholipid multilayers containing octadecylamine fluoresce when exposed to aqueous calcein AM. (a) Images of ODA/DOPC droplets at t = 0 mins and t = 60 mins under red, green filters and merged channels. (b) A plot of average green/red intensity along with time within the droplets. (c) A plot of fluorescence intensity with thickness within the droplet.
Figure 5.
Lipid vesicle solution-based ester hydrolysis. (a) Fluorescence intensity along with time for different concentrations of octadecylamine in DOPC vesicles. (b) Calculated max fluorescence changing rate with different concentration of octadecylamine to get the catalytic turnover number with linear fitting. (c) Calibration of different concentration of calcein in the 96 well plate measured with plate reader.(d) Fluorescence intensity along with time for ODA vesicles solution with/without DOPC.
Figure 5.
Lipid vesicle solution-based ester hydrolysis. (a) Fluorescence intensity along with time for different concentrations of octadecylamine in DOPC vesicles. (b) Calculated max fluorescence changing rate with different concentration of octadecylamine to get the catalytic turnover number with linear fitting. (c) Calibration of different concentration of calcein in the 96 well plate measured with plate reader.(d) Fluorescence intensity along with time for ODA vesicles solution with/without DOPC.
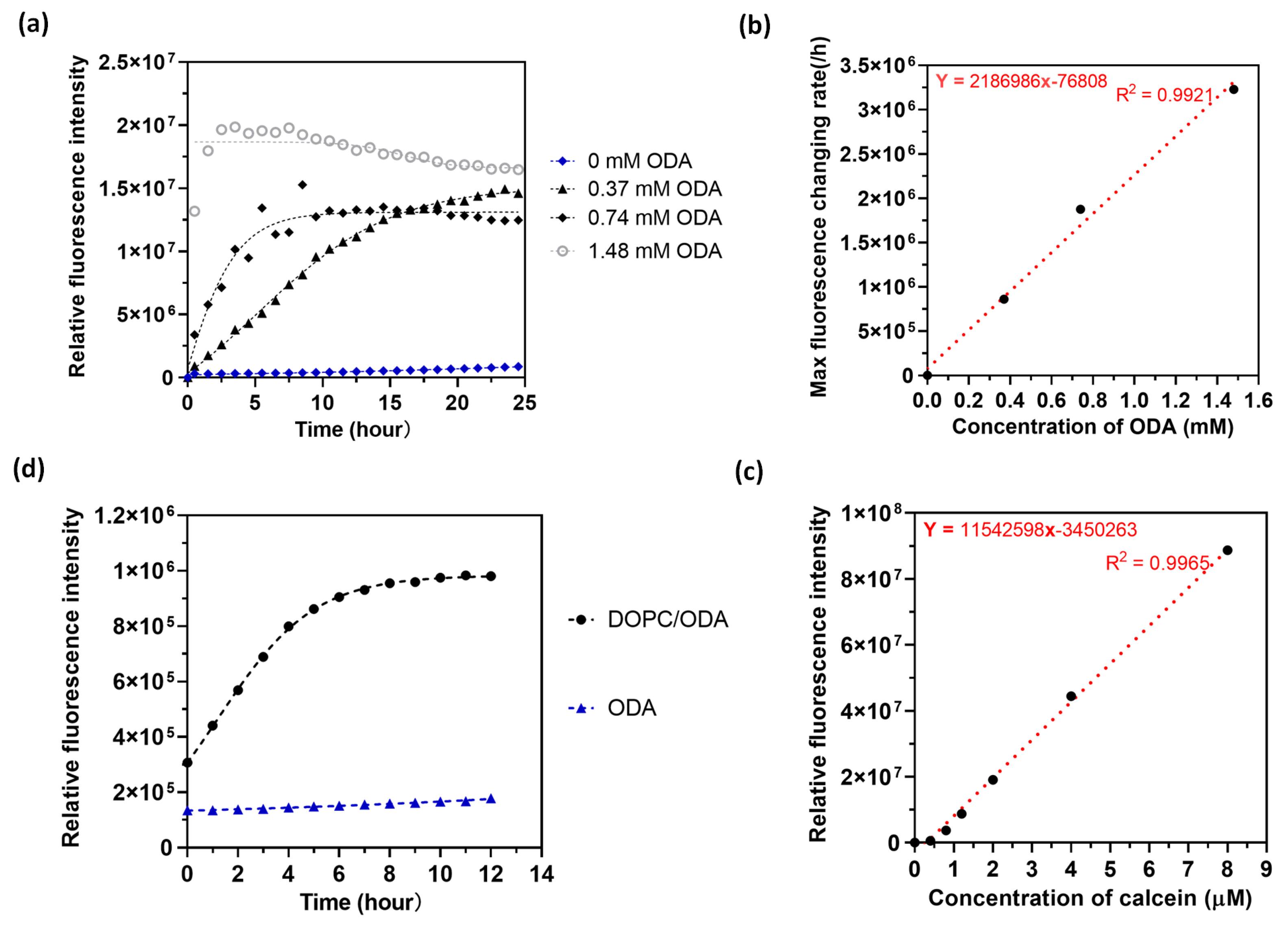
Figure 6.
H NMR spectra and assignments of calcein molecules: (a) 500 µM calcein-A, (b) 100 µM calcein-AM incubated with DOPC and ODA, (c) 100 µM calcein-AM and (d) DOPC and ODA. All samples were dissolved in deuterated DMSO-d6 solvent and experiments were performed at 25 °C using a 600 MHz NMR spectrometer (full spectra are provided in Figure S3). *impurities
Figure 6.
H NMR spectra and assignments of calcein molecules: (a) 500 µM calcein-A, (b) 100 µM calcein-AM incubated with DOPC and ODA, (c) 100 µM calcein-AM and (d) DOPC and ODA. All samples were dissolved in deuterated DMSO-d6 solvent and experiments were performed at 25 °C using a 600 MHz NMR spectrometer (full spectra are provided in Figure S3). *impurities
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



