
Submitted:
04 June 2024
Posted:
05 June 2024
You are already at the latest version
Abstract
Background: Commensal Neisseria species are part of the oropharyngeal microbiome and important for health, but also serve as a reservoir for antimicrobial resistance. Little is known about the prevalence of these species in the general population, how this varies by age and how antimicrobial susceptibility varies between species. Methods: We assessed the prevalence and antimicrobial susceptibility of commensal Neisseria species in the parents (n=38) and children (n=50) of 35 families in Belgium.Results: Various commensal Neisseria (n=5) could be isolated from the participants. Most abundant were N. subflava and N. mucosa. N. subflava was detected in 77 of 88 (87.5%) individuals and N. mucosa in 64 of 88 (72.7%). N. mucosa was more prevalent in children (41/50 [82%]) than parents (23/38 [60.5%]; P<0.05), while N. bacilliformis was more prevalent in parents (7/36 [19.4%]) than children (2/50 [4%]; P<0.05). N. bacilliformis showed high ceftriaxone minimum inhibitory concentrations (MICs; median MIC 0.5mg/L; IQR 0.38-0.75) and these high MICs explained the higher ceftriaxone MICs of all Neisseria isolates from the parents than the children. The median azithromycin MIC of all Neisseria isolates per individual was positively correlated with the median family MIC (rho=0.24; P=0.033). The same association was found for the analysis limited to N. subflava (rho=0.36; P=0.004) and N. mucosa (rho=0.30; P=0.027). Interpretation: The most abundant commensal Neisseria species found in this population were N. subflava and N. mucosa, of which the prevalence of N. mucosa varied by age. The prevalence of N. bacilliformis also varied by age and showed concerningly high ceftriaxone MICs which warrant further investigation. We found evidence of clustering of azithromycin, but not ceftriaxone, MIC by family.
Keywords:
Neisseria
; commensals
; intra-familial transmission
; oropharynx
; horizontal gene transfer
; Belgium
Introduction
Neisseria gonorrhoeae is becoming increasingly resistant to antimicrobials, including last-resort antimicrobials such as ceftriaxone and azithromycin [1,2]. Various types of evidence have established that much of this resistance has been acquired via horizontal gene transformation (HGT) from the non-pathogenic Neisseria species in the oropharynx [3,4,5]. The acquisition of sections of the penA, parC, gyrA, mtrCDE, rplB, rplD and rplV from non-pathogenic Neisseria has played an important role in the acquisition of penicillin, cephalosporin, macrolide, and/or fluoroquinolone resistance in N. meningitidis and N. gonorrhoeae [5,6,7]. These findings have led to calls for pheno- and genotypic surveillance of antimicrobial resistance in commensal Neisseria spp. as an early warning system of excessive antimicrobial consumption [8,9].
A systematic review of antimicrobial susceptibility of commensal Neisseria spp. found that minimum inhibitory concentrations (MICs) were increasing over time, but the findings were limited by the small number of studies available [10]. Most study samples were from populations attending STI clinics [11,12,13,14], from individuals that were colonized with Neisseria meningitidis [15] or employees sampled at their workplace [11,16]. Only one of these studies used a sample of the general population, and this study was limited to characterizing N. lactamica and N. meningitidis prevalence in children under 15 years [17].
A related problem is the paucity of studies describing the prevalence of the various oropharyngeal Neisseria spp. One of the largest studies used oropharyngeal swabs plated onto selective media (LBVT.SNR) and modified Thayer-Martin medium to characterize the prevalence of Neisseria spp. in 202 STI clinic attendees in Seattle in 1987 [14]. They found that 25% were colonized with N. mucosa, and 96% were colonized with N. perflava/ N.sicca (Table 1). The prevalence of N. meningitidis (29%) and N. cinerea (37%) was noted to be higher in men who have sex with men (MSM) than heterosexual men (7%/27%). A smaller study 10 years later involving 40 hospital staff in Madrid found that 93% of nasopharyngeal swab samples were culture positive for N. perflava/ N. sicca, 25% for N. mucosa, 20% for N. flava and 10% for N. cinerea. Only 2% to 5% were colonized by N. lactamica and N. meningitidis [16].
In 2019-2020, we assessed the Neisseria species distribution and antimicrobial susceptibility in 96 participants at our center in Belgium: 32 employees, 32 MSM who did not use antibiotics in the previous 6 months, and 32 MSM who did [11]. We used oral swabs plated onto Columbia Blood Agar and Modified Thayer-Martin Agar to characterize the distribution of specific Neisseria spp. and their individual colony MICs to azithromycin, ceftriaxone and ciprofloxacin. Oropharyngeal Neisseria were cultured and identified with MALDI-TOF–MS. Commensal Neisseria from employees, as well as MSM, were remarkably resistant. The commensal Neisseria isolated from MSM had significantly higher MICs for azithromycin (7.0 mg/L, IQR 3.0–280.2) and ciprofloxacin (0.250 mg/L, IQR 0.020–0.380) compared to those from the employees (3.0 mg/L, IQR 2.0–4.0, p < 0.0001; and 0.023 mg/L, IQR 0.012–0.064, p < 0.001, respectively). Susceptibility did not differ significantly according to recent antimicrobial exposure in MSM, suggesting that commensal Neisseria may be shared by activities such as kissing [18].
An oropharyngeal swab-based Neisseria survey of 207 MSM in Hanoi, Vietnam, from 2016 to 2017, detected at least one Neisseria spp. in all individuals: N. flavescens (47%), N. subflava (22%), N. perflava (11%), N. meningitidis (5%), N. macacae (5%), N. gonorrhoeae (3%), N. cinerea (3%), N. mucosa (3%) N. oralis (2%) and N. lactamica (0.5%) [12]. Swabs were cultured onto Gelose Chocolat PolyViteX VCAT3 and chocolate agar plates.
Most recently, a study in 2022 surveyed the distribution and antimicrobial susceptibility of commensal Neisseria spp. in MSM (n=108) and the general population in Italy (male adults visiting their GPs with a sore throat; n=87) [19]. Pharyngeal swabs were plated onto Columbia blood agar and modified Thayer-Martin agar plates. N. subflava (59.7%) and N. flavescens (28%) were the most frequently detected species. Other species included N. perflava (3%), N. macacae (3%) and N. mucosa (2%). This distribution was similar for the two populations except for N. mucosa, which was only detected in the general population. Antimicrobial susceptibilities per species and the variation between the MSM and the general population were very similar to the Belgian study [11].
In these studies, participants complained about the discomfort induced by the pharyngeal swabbing, which typically induces a gag reflex [20]. In addition, centres in low- and middle-income countries indicated that the MALDI TOF-MS and individual colony MICs would be costly and difficult to perform [20]. To circumvent these obstacles, we developed and validated a new surveillance protocol that involved the use of an oral rinse/gargle with water instead of the oro-pharyngeal swab to sample the oropharyngeal Neisseria spp. [20,21,22]. Dilutions of this oral rinse could then be plated onto LBVT.SNR plates with and without antimicrobials to establish the proportion of Neisseria spp. with resistance to each antimicrobial of interest without the necessity of MALDI-TOF-MS [20,21,22]. Except for a small pilot study, we have not directly compared this proportion method with the original method of ascertaining individual colony MICs [20].
In this study, we aimed to conduct this comparison. We also aimed to expand our knowledge of the epidemiology of commensal Neisseria spp. by assessing the prevalence of various commensal Neisseria spp. in a randomly selected group of parents and their children in Belgium. Our specific objectives were to 1) assess the prevalence of different species of oropharyngeal Neisseria spp. in adults and children, 2) assess the proportion of commensal Neisseria spp. per person with resistance to azithromycin, ceftriaxone and ciprofloxacin, 3) assess the azithromycin and ceftriaxone MIC distribution of commensal Neisseria spp. per species per person, 4) evaluate the correlation between the proportion resistance and the MIC distribution for azithromycin and ceftriaxone, and 5) assess if there is a correlation between the azithromycin and ceftriaxone MICs of commensal Neisseria between family members.
Methods
Survey Population
The study population consisted of 35 randomly selected families that were attending children’s sports events at a municipal sporting facility in Antwerp, Belgium. Recruitment took place over the course of two weekends in October 2023. Random adults with at least one child with them were approached by the study team and invited to participate if they met the following criteria: be part of a family where at least one child (aged 5 to 13) and one adult who is either the parent or a first degree relative and are living with the child are willing to participate. The participating child needed to be present with at least one of the parents. The child provided oral informed consent, and the parent provided written informed consent. The first 35 eligible families agreeing to participate were included in the study.
Data Collection and Sampling Procedure
All parental participants provided written informed consent prior to the collection of data and samples. Baseline characteristics were noted (including self-reported age, sex, time since last antibiotic use and omnivorous or vegetarian diet). The participants were instructed to rinse/gargle their mouths with 15 mL sterile water for 30s, after which they were collected in a sterile container [20]. Specifically, under direct observation, participants rinsed their mouths, followed by gargling, and then repeated the rinse and gargle one additional time for a total duration of 30 seconds. Immediately upon arrival at the laboratory (within 6h after sample collection), 1000µL of each sample was added to 1000 µL of skim milk with 30% glycerol and s stored at -80°C until further processing in batch.
Sample Processing
Culture, MIC Determination and Identification of Neisseria Species
One aliquot of each sample in skim milk was allowed to thaw completely at ambient temperature and vortexed vigorously before 100µL was plated on commensal Neisseria selective medium (LBVT.SNR- LB medium containing 1% Bacto-Tryptone, 0.5% yeast extract, 0.5% sodium chloride, 1.5% Bacto-Agar and 5.0 ml of neutral red indicator (0.3% [wt/vol]) per litre was added, and sterilized by autoclaving for 15 min at 121°C) with and without azithromycin (1mg/L, Sigma Aldrich, Steinheim am Albuch, Germany), ceftriaxone (0.125mg/L) and ciprofloxacin (0.06mg/L). Plates were incubated for up to 48h at 37°C in 5–7% CO2 incubator.
Bacterial isolates were obtained from LBVT.SNR plates without antibiotics. A maximum of four colonies with distinct morphologies were randomly selected from each plate and subcultured for further analysis. Species identification was performed using a MALDI Biotyper® Sirius IVD system equipped with the MBT Compass IVD/RUO software and library (Bruker Daltonics, Bremen, Germany). Briefly, bacterial isolates were prepared by smearing the growth of a single colony onto polished steel target plates, followed by overlaying with α-cyano-4-hydroxycinnamic acid (CHCA) matrix solution. Subsequently, the target plates were loaded into the MALDI-TOF MS instrument for spectra acquisition. Spectra were obtained in linear mode within a mass range of 2 to 20 kDa. The acquired spectra were compared against a comprehensive library module containing 12438 spectra. Identification results were evaluated based on recommended cut-off values of 1.7 and 2 for genus and species levels, respectively. Only isolates belonging to the genus Neisseria were included in further analyses.
Isolates identified as N. sicca and N. macacae were grouped into one category with N. mucosa, whereas isolates identified as N. perflava and N. flavescens were grouped into one category with N. subflava [23].
Antimicrobial Susceptibility Determination
MICs of the isolated Neisseria species to azithromycin and ceftriaxone were determined on GC chocolate agar plates (Becton Dickinson, Belgium) using ETEST® (BioMérieux Marcy-l’Étoile, France) incubated for 18-24h at 37°C and 5-7% CO2, and expressed in mg/L. The inoculum is prepared in PBS and the turbidity of the bacterial suspension is standardized to 0,5 McFarland using a densitometer (DEN 1B, BioSan, Latvia).
Proportion Resistant
The proportion of colonies of Neisseria spp. resistant to azithromycin, ceftriaxone and ciprofloxacin were determined by plating on LBVT.SNR. 100µl of the oral rinse samples in skimmilk were spread using a plate spinner (Petriturn-M), on respective plates with and without the addition of 1µg/ml azithromycin, 0.06µg/ml ciprofloxacin and 0.125µg/ml ceftriaxone (all Sigma Aldrich, Steinheim am Albuch, Germany). The concentration for ciprofloxacin and ceftriaxone chosen were the EUCAST breakpoints for N. gonorrhoeae. EUCAST has an ECOFF for N. gonorrhoeae for azithromycin of 1µg/ml. Plates were incubated up to 48 hrs at 37°C in a 5-7% CO2 incubator. The total number of colonies on the plates with and without antimicrobials were determined, and counts were taken from the plate with 20-200 colonies using a colony counter (Scan 300, Interscience, France).
Statistics
Neisseria prevalence: Prevalence was expressed as the proportion of participants from whom a certain species was isolated. Prevalence was compared between groups using Chi-square/Fischer’s exact test.
Neisseria species richness: Neisseria species richness was defined as the number of different Neisseria species detected per participant. Species richness was reported as median (interquartile range) and compared between groups using Kruskal-Wallis rank sum tests.
Antimicrobial susceptibility: To enable statistical testing, MICs above the maximum or below the minimum level of the ETEST strip were simplified as follows: azithromycin MIC > 256 mg/L was recorded as 256 mg/L; ceftriaxone MIC < 0·016 mg/L as 0·016 mg/L; and ciprofloxacin MIC > 32 mg/L as 32 mg/L. MICs were reported as median (interquartile range), for all Neisseria spp. combined and per individual species. Differences were compared between groups using Kruskal-Wallis rank sum tests. Bonferroni corrections were applied to adjust for multiple comparisons.
Proportion versus MIC distribution: Spearman’s correlation was used to assess if there was an association between each individual’s median MIC of their Neisseria spp. and the proportion of Neisseria spp. that were resistant to that antimicrobial.
Evidence of Transmission of Commensal Neisseria spp. between Family Members
If we acquire further funding, we will conduct whole genome sequencing of all the isolates and use this data to evaluate evidence of intrafamilial transmission. In the interim, we use Spearman’s correlation between the median azithromycin/ceftriaxone MIC of all Neisseria isolates per individual and the median azithromycin/ceftriaxone Neisseria MIC of the rest of the individual’s family, i.e., calculated by excluding the MICs from the index individual from the family median. All statistical analyses were performed with Stata/MP V16.1 (StataCorp).
Ethics
Ethics approval was obtained from ITM’s Institutional Review Board (1574/22) and from the Ethics Committee of the University of Antwerp (3831).
Role of the Funding Source
This study was funded by a SOFI-B Grant: PRESTIP. The funder was not involved in any stage of the study.
Results
Survey Population Data
In the 35 families recruited, the median age of the 38 parents was 45 (IQR 35 – 55) years and 10 (IQR 8 – 14) years in the 50 children (Table 2). The median number of parents/children per family was one (IQR 1-1)/one (IQR 1-2). Fifty-one (58%) of the participants were male and only one was a vegetarian. All the participants reported consuming antibiotics in the past, but the vast majority (90.9%) reported that this was more than 6 months prior to the study. Two individuals reported antibiotic use 7 to 30 days prior and 6 individuals between one and 3 months prior.
Prevalence of Different Species of Oropharyngeal Neisseria spp. in Adults and Children
N. subflava was isolated in 77 of 88 (87.5%) individuals (Table 3; Figure 1). N. mucosa was also detected in the majority of participants (64/88 [72.7%]. N. elongata (24/88 [27.3%]), N. bacilliformis (9/88 [10.2%]) and N. oralis (4/88 [4.5%]) were detected in a lower proportion of individuals. Only N. mucosa and N. bacilliformis varied by age; N. mucosa was more prevalent in children (41/50 [82.0%]) than parents (23/38 [60.5%]; P<0.05) and N. bacilliformis was more prevalent in parents (7/36 [19.4%]) than children (2/50 [4%]; P<0.05). No other Neisseria species were identified.
Richness of Neisseria Species
The participants were colonized by a median of 2 Neisseria species (IQR 2-2; Figure 1). This did not differ between children and parents (both median 2; IQR 2-2).
Antimicrobial Susceptibility of Commensal Neisseria spp.
Proportion method: The median per person percent of Neisseria spp. isolates with resistance to azithromycin (37.4%; IQR 24-84%) and ciprofloxacin (23.7%; IQR 9-63%) was considerably higher than that for ceftriaxone (0.2%; IQR 0-2%; Table 3; Figure 2). Only in the case of ceftriaxone resistance was the prevalence higher in the parents (0.81%) than the children (0.07%; P<0.005).
MIC distribution method: In a similar vein, the individual colony MICs of the commensal Neisseria spp. were higher for azithromycin (median 3mg/L; IQR 1.5 - 4) than ceftriaxone (median 0.047mg/L; IQR 0.032-0.094; Table 4). The ceftriaxone MICs were also higher in the parents (0.064mg/L; IQR 0.032-0.094) than the children (0.047mg/L; IQR 0.032-0.064; P<0.05).
MIC distribution per species: The ceftriaxone MICs of N. elongata (0.064mg/L; IQR 0.047-0.125; P<0.0005) and N. bacilliformis (0.5mg/L; IQR 0.38-0.75; P<0.0005) were higher than those of N. subflava (0.047mg/L; IQR 0.023-0.064; Table 4). The azithromycin MICs of N. subflava (3mg/L; IQR 2-4) was lower than that of N. mucosa (4mg/L; IQR 3-8; P<0.005) but higher than those of N. elongata (0.5mg/L; IQR 0.38-0.75; P<0.005) and N. bacilliformis (2mg/L; IQR 1.5-2; P<0.005).
Correlation between MIC Distribution and Proportion Resistant Methods
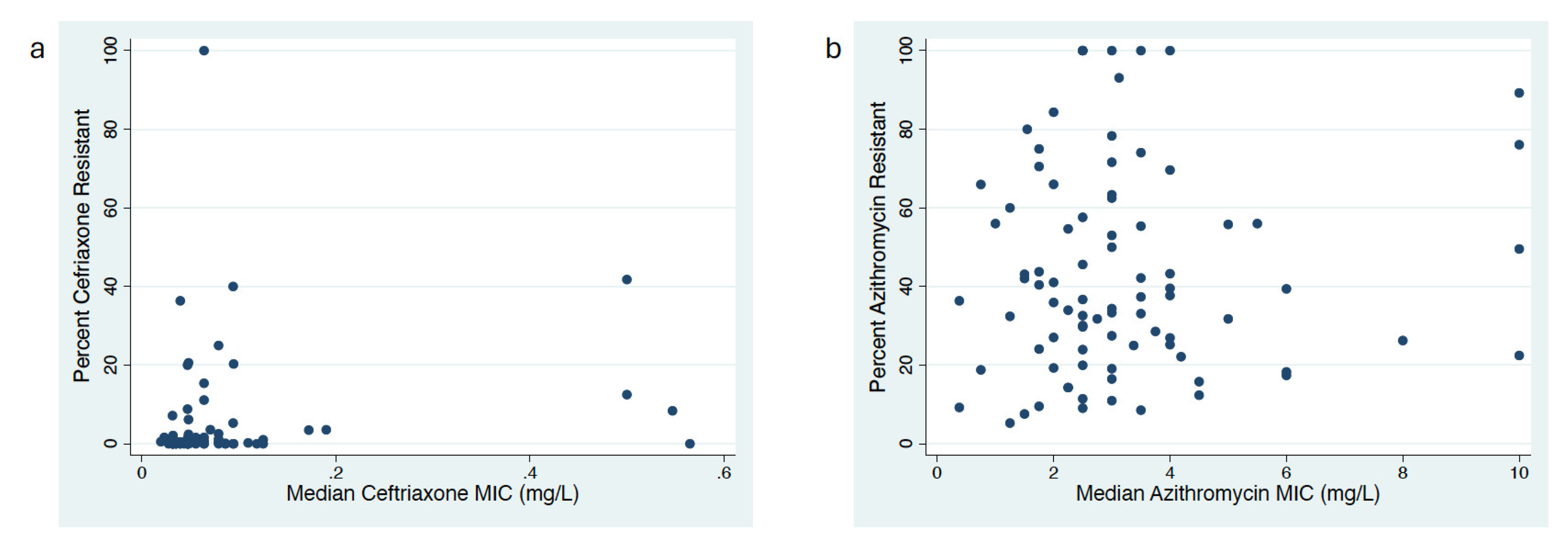
In the case of ceftriaxone (rho=0.27; P=0.012) but not azithromycin (rho=0.04; P=0.720), there was a significant correlation between each individual’s median MIC of their Neisseria spp. and the proportion of Neisseria spp. that were resistant to that antimicrobial (Figure 3).
Predictors of Antimicrobial Susceptibility
Our mixed-effect linear regression models confirmed that the ceftriaxone MICs of N. bacilliformis were significantly independently higher than those of N. subflava (Table 5). In contrast, the azithromycin MICs of N. subflava were higher than those of N. elongata but lower than those of N. mucosa.
In the model limited to N. subflava, the ceftriaxone (but not the azithromycin) MICs were independently associated with time since receipt of the last antibiotic (Table 5).
Association of MICs of Commensal Neisseria spp. between Family Members
The median azithromycin MIC of all Neisseria isolates per individual was positively correlated with the median family MIC (rho=0.24; P=0.033). The same association was found for the analysis limited to N. subflava (rho=0.36; P=0.004) and N. mucosa (rho=0.30; P=0.027). These associations for ceftriaxone were not significant.
Discussion
Differences in Prevalence of Neisseria spp. by Age Group
Similar to previous studies, we found near universal colonization with commensal Neisseria species in both age groups [11,14,20,24]. N. subflava was equally prevalent in the parents and children, but N. mucosa was slightly more prevalent in children (82%) than their parents (60.5%). Conversely, N. bacilliformis was more prevalent in the parents (19.4%) than in the children (4%).
The high prevalence of N. subflava is very similar to that seen in previous studies [14,16], but the prevalence of N. mucosa (60 to 82%) was more than double that of the previous studies reviewed in the introduction [14,16]. In our previous survey in Belgium, the prevalence of N. mucosa was 25% in the employees and 9% in the MSM [11]. In a recent study from Italy, N. mucosa and N. macacae were detected in 2% and 3% of the general population, respectively [19]. One possible explanation for the higher prevalence detected in the current study is that it is the first of these studies to use an oral rinse/gargle instead of a swab for sampling. Another explanation is the different culture media used -LBVT.SNR in this and the previous pilot study [20] compared to blood agar and modified Thayer-Martin in the recent Italian study [19].
There is evidence that different Neisseria spp. localize to different parts of the oropharynx. An analysis of sequence data from the Human Microbiome Project found that samples from the tongue were enriched with N. subflava, whereas N. mucosa, N. sicca, and N. macacae predominated in the gingival plaque [25]. The oral rinse/gargle may be more likely to sample bacteria from the teeth and gingival plaques than oro-pharyngeal swabs, which could account for the high prevalence of N. mucosa found in our study. This hypothesis is not, however, supported by our previous small study that aimed to compare the same oral rinse and swab sampling techniques [20]. This study in 10 individuals found that N. subflava was detected from all 10 individuals by both techniques and N. mucosa in 7 individuals in the swab sample and 6 individuals in the rinse samples. The rinse samples did, however, detect slightly more N. elongata and N. bacilliformis isolates than the swab samples.
Variations of Antimicrobial Susceptibility between Neisseria spp.
We found significant differences in azithromycin and ceftriaxone susceptibilities between Neisseria species. The azithromycin MICs of N. subflava were higher than those of N. elongata but lower than those of N. mucosa. The ceftriaxone MICs of N. bacilliformis were significantly independently higher than those of N. subflava. These differences persisted after controlling for potential confounders. These findings are to the best of our knowledge, novel. The only previous studies we could find with comparable data were our previous survey in Belgium [11] and the recent survey in Italy [19]. In the Belgian study, we found 96 isolates of N. subflava, 14 N. mucosa, three N. elongata and one N. bacilliformis. These small sample sizes did not warrant formal statistical comparisons of MICs between species, but in general, the same pattern was observed as the current study. The N. subflava azithromycin MICs were numerically higher than those of N. elongata but lower than those of N. mucosa, and the one isolate of N. bacilliformis had a markedly higher ceftriaxone MIC (1.5mg/L) than the highest MIC of N. subflava (0.064mg/L). The Italian study found likewise that N. mucosa had the highest azithromycin (and ciprofloxacin) MICs [19]. A systematic review of antimicrobial susceptibility in commensal Neisseria spp. was only able to find sufficient data to compare the susceptibilities of N. lactamica with N. gonorrhoeae and N. meningitidis [10]. This study found that MICs for azithromycin, penicillin, ceftriaxone, and ciprofloxacin were typically highest in N. lactamica. The study did not include any N. bacilliformis. Three previous reports of N. bacilliformis endocarditis have noted high third generation cephalosporin MICs (ceftriaxone 0.25mg/L in 2 cases in 2009 and 2011 [26,27] and cefotaxime 1.5mg/L in 2023 [28]). The original description of N. bacilliformis that described 8 isolates from clinical cases with invasive disease noted that the cefepime MICs were between 3 and 12mg/L [29]. The single isolate of N. bacilliformis in the Centers for Disease Control and Prevention (CDC) Antibiotic Resistance Isolate Bank, has a ceftriaxone MIC of 16mg/L [30]. The genetic basis for these high cephalosporin MICs in N. bacilliformis has not been established. Whole genome sequencing of the CDC Resistance Isolate Bank isolate revealed only 2 resistance associated mutations – porB A121D and penA I312M [30].
Horizontal gene transfer (HGT) of the penA gene from commensal Neisseria has played a crucial role in the emergence of cephalosporin resistance in N. gonorrhoeae [31]. A recent analysis of 35,513 Neisseria isolates from 15 different Neisseria species found that N. subflava and N. cinerea were the most common source of recombinant sequences in N. gonorrhoeae penA [31]. This and previous analyses did not, however, include any isolates from N. bacilliformis [31,32]. It will be important to evaluate the genotypic determinants of the high ceftriaxone MICs in N. bacilliformis and include N. bacilliformis in future studies of cephalosporin resistance in N. gonorrhoeae.
The highest azithromycin MIC in Neisseria isolates was 16mg/L. We have previously found that a high proportion of MSM in our setting are colonized with N. subflava’s with azithromycin MICs of >256mg/L [11,33] and that this is due to the acquisition of the ribosomal protecting msr(D) gene from streptococci [33]. The absence of these high MIC isolates in this study suggests that this gene has not spread widely in the general population.
Variations in Ceftriaxone MIC by Species and Species by Age Group Explain Variation in MIC by Age Group
The higher ceftriaxone MIC found in Neisseria isolates from adults can be parsimoniously explained by the higher prevalence of N. bacilliformis and its higher ceftriaxone MICs. This is evident from the linear-regression model where N. bacilliformis but not age is a significant predictor of ceftriaxone MIC (Table 4).
Transmission of Neisseria spp.
We plan to sequence the Neisseria spp. in this study which should give us an indication of possible transmission between individuals. A previous study based on metagenomic sequencing of oropharyngeal bacteria of sexual partners found some evidence that oral Neisseria species were shared between partners via kissing [18]. The current study found that the azithromycin but not the ceftriaxone MICs of commensal Neisseria spp. were positively associated at a family level. This is the first evidence that we are aware of that phenotypic Neisseria antimicrobial susceptibility may be transmitted between family members, as has been noted with numerous other genera [34]. Alternatively, this association could be explained by the family members joint exposure to another risk factor for AMR. If this association is due to transmitted resistance, then this may help to explain previous study findings such as that from our previous survey in Belgium which found higher azithromycin and ciprofloxacin MICs in commensal Neisseria in MSM than the employees, but no difference between the MSM who had and had not taken antimicrobials in the prior 6 months [11]. If there is transmission of resistant commensal Neisseria spp. following receipt of antimicrobials then this may explain why the association between antimicrobial exposure and resistance is stronger at the population than the individual level [11,35].
Proportion versus MIC Distribution to Assess AMR
We compared the proportion and MIC distribution methods to assess AMR of commensal Neisseria spp. We found a large variation in the percent with azithromycin resistance (median 37% [IQR 24-84%]) and a low percent resistant for ceftriaxone (median 0.2% [IQR 0-2.4%]. There was less variation in the MIC distribution of both azithromycin (median 3mg/L [IQR 1.5-4]) and ceftriaxone (median 0.047mg/L [0.032-0.094]) for commensal Neisseria spp. Surprisingly, we found that only in the case of ceftriaxone there was a significant positive association between the two methods. These findings together with those from other studies suggest that the proportion resistant method may not be an adequate replacement to the MIC distribution method for the surveillance of AMR in commensal Neisseria spp. [36].
This study has a number of limitations, including the small sample size and the fact that the samples may not be representative of the general Belgian population. All the samples were obtained from a single site on two days in a single season. It is unlikely that this sampling strategy would generate a sample representative of the Belgian population. There is some evidence that gonococcal azithromycin susceptibility varies by season [37]. The same may be true for commensal Neisseria species [37]. Our samples were obtained via oral rinsing and not swabs. Although this sampling method is preferred by study participants and physicians [22] and is being increasingly used [20,21,22], there may be differences in the relative proportion of Neisseria species obtained via this method compared to oropharyngeal swabs.
These limitations notwithstanding; this study was the first to characterise the prevalence and antimicrobial susceptibilities of commensal Neisseria spp. in both parents and their children. Previous studies have noted high and increasing azithromycin, ciprofloxacin and ceftriaxone MICs of commensal Neisseria spp. in adult populations. For the first time, we have been able to establish that the MICs of Neisseria spp. in children were very similar to that of their parents. The small differences in MIC could largely be explained by differences in the prevalence of Neisseria species. Our finding of clustering of azithromycin MICs by family builds on previous evidence to suggest that commensal Neisseria spp. are shared. If resistant strains can be transmitted, this may help explain why certain studies have found little or no association between antimicrobial consumption and resistance at an individual level but an association at a population level [11,21,35,38]. These studies have been in high antimicrobial consumption populations such as MSM taking HIV preexposure prophylaxis (PrEP), where the consumption of macrolides may exceed a threshold for macrolide resistance induction by up to seven-fold [11,21,39,40]. The consumption of macrolides in the general population in Belgium is approximately 50% greater than this threshold [40]. Likewise, the consumption of all antibiotics in the general population in Belgium is approximately double that of a lower antibiotic consumption country such as the Netherlands [41]. This relatively high consumption of antimicrobials is a likely cause of the relatively high MICs of commensal Neisseria spp. in our study. If we accept the utility of using antimicrobial susceptibility of commensal Neisseria spp. as early warning system of excessive antimicrobial consumption, then our findings build on other types of evidence to motivate for enhanced antimicrobial stewardship in the general population of Belgium [8].
Data Sharing
All deidentified data are available as a Supplement to this manuscript. Additional related documents such as the study protocol, laboratory analysis plan, and informed consent form, can be obtained from the corresponding author.
References
- Mitjà O, Padovese V, Folch C, Rossoni I, Marks M, i Arias MAR, et al. Epidemiology and determinants of reemerging bacterial sexually transmitted infections (STIs) and emerging STIs in Europe. The Lancet Regional Health–Europe. 2023, 34.
- Wi T, Lahra MM, Ndowa F, Bala M, Dillon J-AR, Ramon-Pardo, P, et al. Antimicrobial resistance in Neisseria gonorrhoeae: global surveillance and a call for international collaborative action. PLoS medicine. 2017, 14, e1002344. [Google Scholar]
- Hanao M, Aoki K, Ishii Y, Shimuta K, Ohnishi M, Tateda, K. Molecular characterization of Neisseria gonorrhoeae isolates collected through a national surveillance programme in Japan, 2013: evidence of the emergence of a ceftriaxone-resistant strain from a ceftriaxone-susceptible lineage. Journal of Antimicrobial Chemotherapy. 2021, 76, 1769–75. [Google Scholar] [CrossRef] [PubMed]
- Zapun, A., Morlot, C., Taha MK. Resistance to β-Lactams in Due to Chromosomally Encoded Penicillin-Binding Proteins. Antibiotics-Basel. 2016, 5(4). doi: ARTN 35 10.3390/antibiotics5040035. PubMed PMID: WOS:000391620900004. [CrossRef]
- Wadsworth CB, Arnold BJ, Sater MRA, Grad YH. Azithromycin Resistance through Interspecific Acquisition of an Epistasis-Dependent Efflux Pump Component and Transcriptional Regulator in. Mbio. 2018, 9(4). doi: ARTN e01419-18 10.1128/mBio.01419-18. PubMed PMID: WOS:000443884300078. [CrossRef]
- Chen ML, Zhang, C., Zhang, X., Chen, M. Meningococcal Quinolone Resistance Originated from Several Commensal Species. Antimicrobial Agents and Chemotherapy. 2020, 64(2). doi: ARTN e01494-19 10.1128/AAC.01494-19. PubMed PMID: WOS:000509748200005. [CrossRef]
- Manoharan-Basil SS, Laumen JGE, Van Dijck C, De Block T, De Baetselier, I., Kenyon, C. Evidence of Horizontal Gene Transfer of 50S Ribosomal Genes rplB, rplD, and rplY in Neisseria gonorrhoeae. Frontiers in Microbiology. 2021, 12, 1263. [Google Scholar]
- Goytia, M., Wadsworth CB. Canary in the Coal Mine: How Resistance Surveillance in Commensals Could Help Curb the Spread of AMR in Pathogenic Neisseria. Mbio. 2022, 13, e01991–e22. [Google Scholar]
- Kenyon C, Laumen J, Manoharan-Basil, S. Choosing new therapies for gonorrhoea: we need to consider the impact on the pan-Neisseria genome: a viewpoint. Antibiotics-Basel. 2021, 10(5). [CrossRef]
- Vanbaelen T, Van Dijck C, Laumen J, Gonzalez N, De Baetselier I, Manoharan-Basil SS, et al. Global epidemiology of antimicrobial resistance in commensal Neisseria species: a systematic review. International Journal of Medical Microbiology. 2022, 312(3). [CrossRef]
- Laumen JGE, Manoharan-Basil SS, Abdellati S, De Baetselier I, Van Dijck, C., Martiny, D., et al. Antimicrobial susceptibility of commensal Neisseria in the general population and men who have sex with men in Belgium. Scientific Reports. 2022. [CrossRef]
- Dong HV, Pham LQ, Nguyen HT, Nguyen MXB, Nguyen TV, May, F., et al. Decreased Cephalosporin Susceptibility of Oropharyngeal Neisseria Species in Antibiotic-Using Men-who-have-sex-with-men of Hanoi, Vietnam. Clin Infect Dis. 2019. Epub 2019/05/03. [CrossRef] [PubMed]
- Furuya R, Onoye Y, Kanayama A, Saika T, Iyoda T, Tatewaki M, et al. Antimicrobial resistance in clinical isolates of Neisseria subflava from the oral cavities of a Japanese population. J Infect Chemother. 2007, 13, 302–4. [Google Scholar] [CrossRef] [PubMed]
- Knapp J, Hook 3rd E. Prevalence and persistence of Neisseria cinerea and other Neisseria spp. in adults. Journal of clinical microbiology. 1988, 26, 896–900.
- Chen ML, Zhang, C., Zhang, X., Chen, M. Meningococcal Quinolone Resistance Originated from Several Commensal Neisseria Species. Antimicrobial Agents and Chemotherapy. 2020, 64(2). doi: ARTN e01494-19. 10.1128/AAC.01494-19. PubMed PMID: WOS:000509748200005. [CrossRef]
- Sáez Nieto JA, Marcos, C. , Vindel, A. Multicolonization of human nasopharynx due to Neisseria spp. International Microbiology. 1998, 1, 59–63.
- Shen YF, Chen ML. Prevalence, sequence type, and quinolone resistance of Neisseria lactamica carried in children younger than 15 years in Shanghai, China. Journal of Infection. 2020, 80, 61–8. https://doi.org/10.1016/j.jinf.2019.08.020. PubMed PMID: WOS:000507459600008. [CrossRef]
- Van Dijck C, Laumen JG, Manoharan-Basil SS, Kenyon, C. Commensal neisseria are shared between sexual partners: implications for gonococcal and meningococcal antimicrobial resistance. Pathogens. 2020, 9, 228. [Google Scholar] [CrossRef] [PubMed]
- Gaspari V, Djusse ME, Morselli S, Rapparini L, Foschi, C., Ambretti, S., et al. Non-pathogenic Neisseria species of the oropharynx as a reservoir of antimicrobial resistance: a cross-sectional study. Frontiers in Cellular and Infection Microbiology. 2023, 13.
- Laumen JGE, Abdellati S, Van Dijck C, Martiny D, De Baetselier I, Manoharan-Basil SS, et al. A Novel Method to Assess Antimicrobial Susceptibility in Commensal Oropharyngeal Neisseria—A Pilot Study. Antibiotics. 2022, 11, 100. [Google Scholar]
- Vanbaelen T, Eric F, Christophe VD, Achilleas T, Laumen JGE, Sheeba, M.-BS, et al., editors. Effect on the Resistome of Dual vs Monotherapy for the Treatment of Neisseria gonorrhoeae: Results From a Randomized Controlled Trial (ResistAZM Trial). Open Forum Infectious Diseases; 2023: Oxford University Press US.
- Vanbaelen, T. Self-sampling with oral rinse to detect oropharyngeal Neisseria gonorrhoeae among men who have sex with men: results from an exploratory study in Belgium (the SSONG study). Sex Transm Infec. 2024.
- Bennett JS, Jolley KA, Earle SG, Corton C, Bentley SD, Parkhill, J, et al. A genomic approach to bacterial taxonomy: an examination and proposed reclassification of species within the genus Neisseria. Microbiology. 2012, 158, 1570–80. [Google Scholar] [CrossRef] [PubMed]
- Nieto JAS, Marcos, C. , Vindel, A. Multicolonization of human nasopharynx due to Neisseria spp. International Microbiology. 1998, 1, 59–63.
- Donati C, Zolfo M, Albanese D, Tin Truong D, Asnicar F, Iebba V, et al. Uncovering oral Neisseria tropism and persistence using metagenomic sequencing. Nature microbiology. 2016, 1, 1–9.
- Masliah-Planchon J, Breton G, Jarlier V, Simon A, Benveniste O, Herson, S. , et al. Endocarditis due to Neisseria bacilliformis in a patient with a bicuspid aortic valve. Journal of clinical microbiology. 2009, 47, 1973–5.
- Abandeh FI, Balada-Llasat J-M, Pancholi P, Risaliti CM, Maher WE, Bazan JA. A rare case of Neisseria bacilliformis native valve endocarditis. Diagnostic microbiology and infectious disease. 2012, 73, 378–9.
- Gavalda M, Vilchez H, Martin M, Ruiz, E. , Ribas, M., Riera, M. Endocarditis caused by Neisseria bacilliformis: a case report and review of literature. IDCases. 2023, 31, e01725.
- Han XY, Hong, T. , Falsen, E. Neisseria bacilliformis sp. nov. isolated from human infections. Journal of clinical microbiology. 2006, 44, 474–9.
- Fiore MA, Raisman JC, Wong NRH, Hudson AO, Wadsworth CB. Exploration of the Neisseria Resistome Reveals Resistance Mechanisms in Commensals That May Be Acquired by N. gonorrhoeae through Horizontal Gene Transfer. Antibiotics-Basel. 2020, 9(10). doi: ARTN 656. 10.3390/antibiotics9100656. PubMed PMID: WOS:000584161000001. [CrossRef]
- Unitt A, Maiden M, Harrison, O. Characterizing the diversity and commensal origins of penA mosaicism in the genus Neisseria. Microbial Genomics. 2024, 10, 001209. [Google Scholar]
- Spratt BG, Bowler LD, Zhang, Q.-Y., Zhou, J., Smith JM. Role of interspecies transfer of chromosomal genes in the evolution of penicillin resistance in pathogenic and commensal Neisseria species. Journal of molecular evolution. 1992, 34, 115–25. [Google Scholar]
- de Block T, Laumen JGE, Van Dijck C, Abdellati S, De Baetselier I, Manoharan-Basil SS, et al. WGS of Commensal Neisseria Reveals Acquisition of a New Ribosomal Protection Protein (MsrD) as a Possible Explanation for High Level Azithromycin Resistance in Belgium. Pathogens. 2021, 10(3). Epub 2021/04/04. https://doi.org/10.3390/pathogens10030384. PubMed PMID: 33806962, PubMed Central PMCID: PMCPMC8005064. [CrossRef] [PubMed]
- Matsumoto K, Hohashi N, Sugishita, C. A study on the transmission of MRSA among the family members including clients of visiting nurse and related infection control. [Nihon Koshu Eisei Zasshi] Japanese Journal of Public Health. 2001, 48, 190–9. [Google Scholar]
- Vanbaelen T, Laumen J, Van Dijck C, De Block T, Manoharan-Basil SS, Kenyon, C. Lack of Association between Antimicrobial Consumption and Antimicrobial Resistance in a HIV Preexposure Prophylaxis Population: A Cross-Sectional Study. Antibiotics. 2024, 13, 188. [Google Scholar]
- Vanbaelen T, Manoharan-Basil S, Kenyon, C. 45 Years of Tetracycline Post Exposure Prophylaxis for STIs and the Risk of Tetracycline Resistance: A Systematic Review and Meta-Analysis. BMC infectious diseases. 2024.
- Olesen SW, Torrone EA, Papp JR, Kirkcaldy RD, Lipsitch, M. , Grad YH. Azithromycin Susceptibility Among Neisseria gonorrhoeae Isolates and Seasonal Macrolide Use. J Infect Dis. 2019, 219, 619–23. [CrossRef] [PubMed] [PubMed Central]
- Kenyon, C. Commentary: Non-pathogenic Neisseria species of the oropharynx as a reservoir of antimicrobial resistance: a cross-sectional study. Frontiers in Cellular and Infection Microbiology. 2024, 13, 1343608. [Google Scholar] [CrossRef] [PubMed]
- Kenyon, C. Dual azithromycin/ceftriaxone therapy for gonorrhea in PrEP cohorts results in levels of macrolide consumption that exceed resistance thresholds by up to 7-fold. The Journal of Infectious Diseases. 2021, 224, 1623–4. [Google Scholar] [CrossRef] [PubMed]
- Kenyon C, Manoharan-Basil SS, Van Dijck, C. Is there a resistance threshold for macrolide consumption? Positive evidence from an ecological analysis of resistance data from Streptococcus pneumoniae, Treponema pallidum, and Mycoplasma genitalium. Microbial Drug Resistance. 2021, 27, 1079–86. [Google Scholar] [CrossRef]
- Bruyndonckx R, Adriaenssens N, Versporten A, Hens N, Monnet DL, Molenberghs, G., et al. Consumption of antibiotics in the community, European union/European economic area, 1997–2017. Journal of Antimicrobial Chemotherapy. 2021, 76, ii7–ii13. [Google Scholar] [CrossRef]
Figure 1.
Lasagna plot of Neisseria species identity of up to four isolates of Neisseria spp. per individual and grouped by family. Each column provides the species identity of up to 4 isolates of Neisseria spp. per individual. The labels on X-axis are unique identifiers for each individual and family. The first two characters specify the family number and last two characters specify if it is the first child/parent (K1/P1), second child/parent (K2/P2) etc. An empty space is used to depict no Neisseria spp. isolated.
Figure 1.
Lasagna plot of Neisseria species identity of up to four isolates of Neisseria spp. per individual and grouped by family. Each column provides the species identity of up to 4 isolates of Neisseria spp. per individual. The labels on X-axis are unique identifiers for each individual and family. The first two characters specify the family number and last two characters specify if it is the first child/parent (K1/P1), second child/parent (K2/P2) etc. An empty space is used to depict no Neisseria spp. isolated.
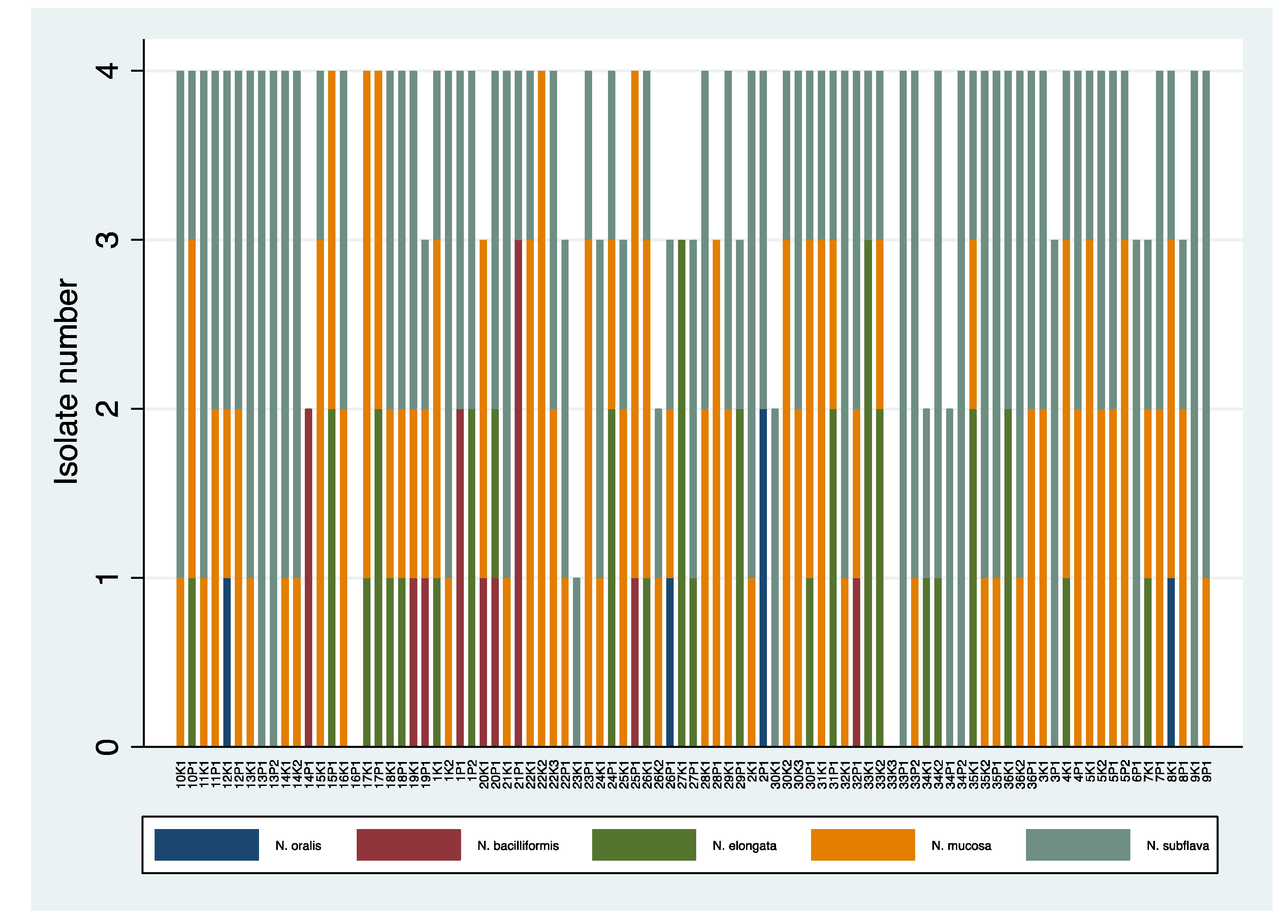
Figure 2.
Histograms of the percent of colonies of Neisseria species per individual that were resistant to (a) azithromycin, (b) ciprofloxacin and (c) ceftriaxone. The Y axis provides the percent of individuals with each percent of isolates resistant to each antimicrobial. For example, in (c), over 80% of individuals had 0% ceftriaxone resistance in their Neisseria spp, whereas one individual had 100% resistance.
Figure 2.
Histograms of the percent of colonies of Neisseria species per individual that were resistant to (a) azithromycin, (b) ciprofloxacin and (c) ceftriaxone. The Y axis provides the percent of individuals with each percent of isolates resistant to each antimicrobial. For example, in (c), over 80% of individuals had 0% ceftriaxone resistance in their Neisseria spp, whereas one individual had 100% resistance.
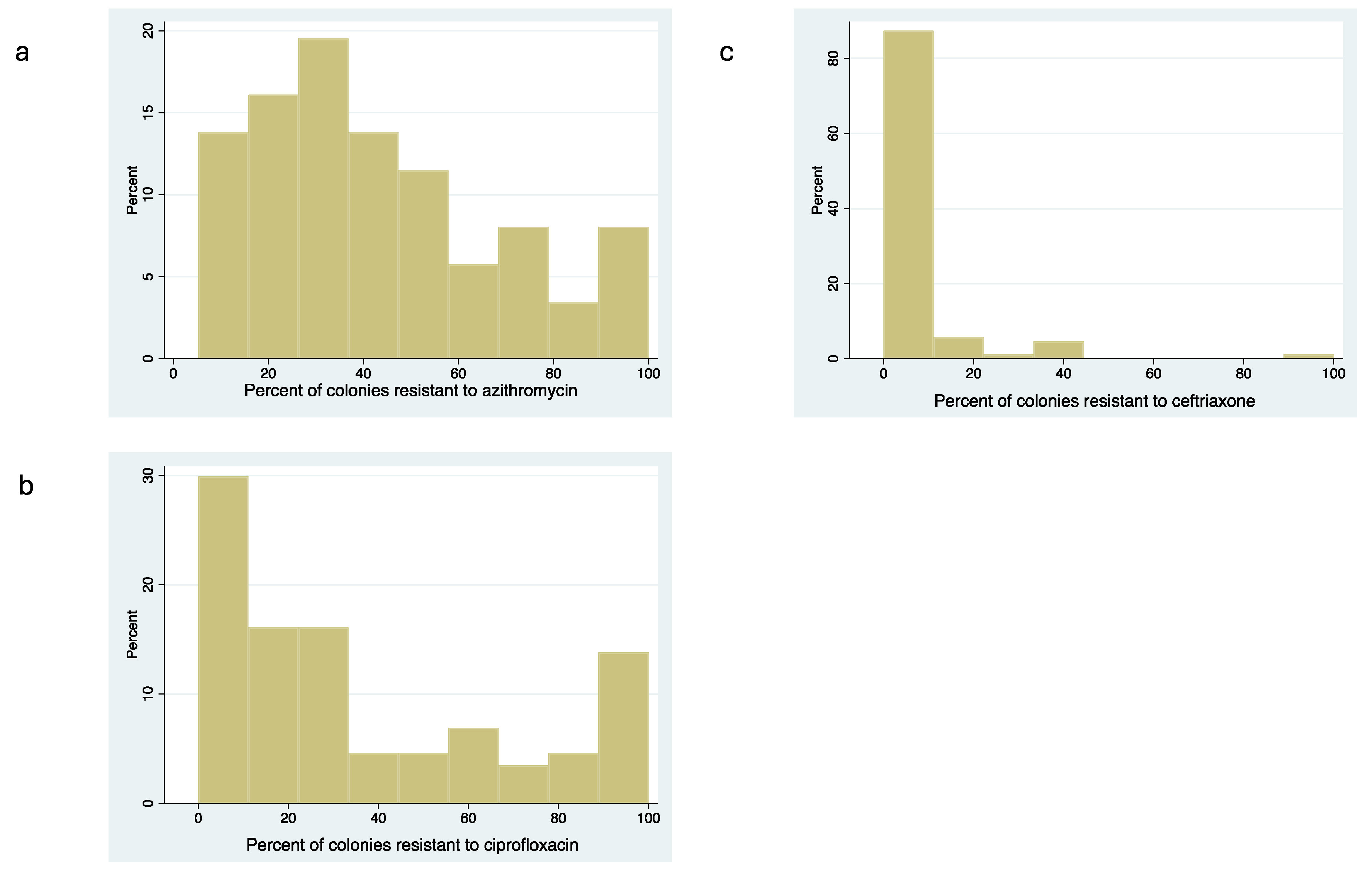
Figure 3.
Scatter plots of the association between the proportion and MIC distribution methods for assessing antimicrobial resistance to a) ceftriaxone or b) azithromycin. Each dot represents the results from a single participant. The Y-axis represents the percent of colonies from that participant that were resistant to ceftriaxone or azithromycin. The X-axis details the median ceftriaxone or azithromycin MIC of that participant (calculated from up to 4 colonies of Neisseria spp. randomly selected from each individual’s sample).
Figure 3.
Scatter plots of the association between the proportion and MIC distribution methods for assessing antimicrobial resistance to a) ceftriaxone or b) azithromycin. Each dot represents the results from a single participant. The Y-axis represents the percent of colonies from that participant that were resistant to ceftriaxone or azithromycin. The X-axis details the median ceftriaxone or azithromycin MIC of that participant (calculated from up to 4 colonies of Neisseria spp. randomly selected from each individual’s sample).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



