
Submitted:
04 June 2024
Posted:
05 June 2024
You are already at the latest version
Abstract
Mangrove ecosystems are one of the most productive on our planet but have declined globally by 30-50%. Many species rely on mangrove habitat and thus their conversion to aquaculture farms has led to a noticeable loss in commodities such as wild fish stocks. This study aimed to assess 1) the influence of the presence of aquaculture and 2) restoration projects on the ecosystems ability to provide resources. We collected data on mangrove vegetation (i.e., biomass, richness and abundance), soil nutrients (i.e. organic carbon, aluminium and nitrogen), crab abundance and fishing pressure in six sites. We set up 15 plots in each site and collected data between May and July 2023. Via generalised linear mixed models we found that the abundance and richness of crabs was significantly higher in aquaculture plots than in no aquaculture plots. Aquaculture plots had higher topsoil aluminium, topsoil and subsoil Nitrogen, and had lower topsoil Carbon than non-aquaculture sites. Restored sites had less Nitrogen in the topsoil than unmanaged sites. Biomass did not change between aquaculture, restored and unmanaged plots. We found a negative correlation between crab abundance and richness and mangrove DBH, suggesting that the species of crabs present preferred areas with propagules for feeding (e.g., Grapsidae crabs). The content of nitrogen in the subsoil was positively correlated with mangrove richness, diversity and height, suggesting the importance of nitrogen availability for mangrove growth. The content of aluminium in the subsoil was negatively correlated with the content of organic carbon both in topsoil and subsoil, suggesting the detrimental effect of aluminium on the carbon cycle. Fishing practices were observed in all sites during the data collection period. Even if most of the vegetation parameters were not significantly affected by presence of aquaculture or management for restoration, it is clear that important variables such as aluminium carbon, and nitrogen content in the soil and crab assemblages were highly variable.
Keywords:
biomass
; carbon
; aluminium
; nitrogen
; crab
; fishing pressure
; restoration
; soil
; management
1. Introduction
Mangroves are highly specialised plant species that provide significant ecological and economic value [1]. By definition, mangroves are subtropical to tropical trees or shrubs adapted to the extreme conditions posed by intertidal regions of brackish to coastal saline waters [2]. Here mangroves rely on highly specialised traits to deal with high salinity, anoxic soil, exposure, and tidal fluxes which can be very large [2,3,4]. Because of the traits required to occupy these zones, mangroves have an exceedingly high capacity for carbon capture and storage, through both biomass and soil [5,6,7]. Estimates suggest they exceed all other aquatic systems and tropical forests (estimates of 5x) and thus have a major role to play in managing global C [8,9]. Likewise, their specialised roots stabilise the shoreline by trapping sediment which reduces coastal erosion and buffers against storm surges [10,11,12]. Mangroves intrinsically have high biodiversity value given few other species of plant can occupy the areas where they thrive [13]. They support a rich diversity of terrestrial and marine life, that rely on mangroves for feeding, spawning or sheltering during some phase of their life cycle with many highly specialised to the habitat [1,2,14,15]. Their presence also enhances the diversity and biomass of communities in adjacent coral reefs and sea grasses [16]. While their socio-economic benefits are diverse extending from sequestering pollutants, providing timber products, medicinal products and food with 80% of fisheries (e.g., crabs, prawns, oysters and many fish) being linked to mangroves [17,18,19].
Despite the importance of mangroves, they are continuing to be lost at an alarming rate for a variety of reasons [20,21]. Globally it has been estimated that between 30-50% mangrove habitat has disappeared in half a century [9,20]. In some parts of the world up to 40% of mangrove species are threatened with extinction [22] and likewise the other organisms that rely on mangroves (e.g., [23]). The loss of mangroves has alarming ramifications for carbon emissions [24]. Stressors on mangrove survival include human-induced climate change from increased coastal storms intensity and sea levels [21]; while more direct human impacts include overexploitation of fisheries and deforestation for urban and agriculture development or aquaculture conversions [25,26].
Deforestation of mangrove for agriculture and aquaculture are particularly noticeable in South-East Asia. South-east Asia is a mangrove hotspot [13], Indonesia alone for instance hosts hold 22% of the worlds’ mangroves [27]. Across South-East Asia though approx. 30% of mangroves have been lost to aquaculture [28]. Some of the losses come from planting of palm oil and rice patties requiring felling of the mangroves which is predicted to increase [29]. But by far the major loss comes from conversion to aquacultural ponds. Estimates of up to 85% of former mangrove areas in Borneo and Indonesia have been converted to aquaculture ponds [30]. Aquaculture conversion involves creating walls of mangroves aligning a constructed pond or the pond has an isolated stand central to the pond [31]. It also requires wild seed of mangroves to ensure the stands which is ironically made difficult by the loss of wild populations of mangroves. Irrespective of the strategy used for aquaculture ultimately the stands of mangroves become fragmented and lose much of the ecosystem service value beyond cultivating a select group of commercially consumable species. This is concerning given it has been estimated that an intact mangrove can generate 70% higher output from goods and services than aquaculture ponds [32]. But simply ceasing aquaculture is unfeasible as it represents job opportunities, food security and substantial profit within primarily low socio-economic countries [26,28]. Therefore, there is a need to find more sustainable practices, and to incentivise the protection and restoration of mangroves to ensure mangrove populations are maintained and the essential ecosystem services are restored.
There is no question that globally there is a need to preserve, manage and restore mangrove populations but there are barriers to this plight. Local communities for instance still need to benefit from mangroves to incentivise their restoration [33]. Which needs to be managed in a sustainable way [25,34]. One of the many challenges here includes tracking the flux in goods and services relative to the mangrove health [25,35,36,37,38]. Indeed, it is difficult to measure the success of restoration if success is not well defined. The aim of this study then was to gain some initial insights into: 1) the influence of aquaculture and 2) restoration projects on mangrove sites. Targeted features were attributes reputed to be useful for detecting shifts in mangrove health:
- -
- Mangrove above and below biomass: a lack of biodiversity is typically linked with a loss of biomass [39]. Likewise, tree characteristics such as age and density have been found to directly correlate with biomass [40]. Evaluating the level of biomass, a forest contains is vital to ensure the correct forestry management practices are implemented [41].
- -
- Mangrove density. The density of the mangroves affects their ability to protect the coast from storm or wave damage [11]. Although mangroves attenuate wave action [12], the more dispersed they are the less of an impact they have. Density can also potentially impact wildlife relying on the mudflats, e.g., crabs.
- -
- Soil quality. Soil composition can provide an indication of mangrove ecosystems health [36]. Mangroves regulate carbon found within coastal soils through CO2 sequestration and biomass accumulation [42,43]. They can also nullify pollutants and denitrify waterways which are secured within the soils [44]. The presence of aluminium, however, can hinder propagule growth, and is often associated with aquaculture [45].
- -
- Crab assemblages. Crabs are intertwined with the growth and development of mangroves [37]. Ferreira et al. [46] suggest that monitoring the abundance of crabs could be used to measure the success of mangrove restoration. Crabs facilitate litter decomposition through leaf processing, burying of leaves and mixing of soil and decomposing bacteria through excavations [15,47,48,49]. Concurrently, they may also influence density and community structure through direct consumption of the propagules.
- -
- Fishing pressure. Pressure from fishing activities can directly impact mangrove health. Local communities rely on resources from mangroves but can be overexploited through intense use.
2. Materials and Methods
2.1. Site Selection and Descriptions
Six sites were investigated in this study (Figure 1). The sites were categorised as unmanaged or managed prior to surveying (Table 1). Designations were made based on local knowledge, historical evidence, and the current use of the land. All sites varied in their perceived mangrove ecosystem health and differing management. Sites were matched as best as possible, so managed and unmanaged sites were in the near vicinity. One of the couplets was selected to include a site currently under aquaculture operation. All transects were labelled as originating from either an unmanaged or managed site irrespective of whether there was within site variation in management.
2.2. Survey Design
At each site, five transects were placed 60m apart. On each transect, three 5m2 plots were measured using a reel marked out at every 5m (Figure 2). This was for ease of use in the field to ensure that the corners of the plots were the same throughout the surveys. The plots started at 5m, 42.5m, and 90m with a 37.5m gap in between the plots. Mangroves were classed as trees if they had leaves, this is due to the premise that propagules that were less than two months old had not produced leaves and therefore may not have established themselves.
2.3. Mangrove Vegetation
Every mangrove tree in each plot were identified to species and then their diameter at breast height (DBH) measured using a measuring tape. AGB and BGB were calculated for thirteen species. Estimates were calculated using either species-specific equations when available or a common allometric equation specific for mangroves (Table 2). This common allometric equation required wood density which was sourced from the Tree Functional Attributes and Ecological Database ([51]; Table 2). We further calculated mangrove richness via Shannon Index and abundance (as total number of individuals per plot) (Table S1).

Table 2.
List of mangrove species found across the study sites with their respective species specific AGB and BGB equations and the source of the equation. Common allometric equation was used for all other species. All equations required Diameter at breast height (D) and general equation required wood density (ρ).
Table 2.
List of mangrove species found across the study sites with their respective species specific AGB and BGB equations and the source of the equation. Common allometric equation was used for all other species. All equations required Diameter at breast height (D) and general equation required wood density (ρ).
| Species | Equation | Source |
|---|---|---|
| Above Ground Biomass | ||
| Avicennia marina | 0.1848D²˙³⁵²⁴ | [51] |
| Rhizophora apiculata | 0.38363D²˙²³⁴⁸ | [52] |
| Rhizophora mucronata | 0.128D²˙⁶ | [53] |
| Rhizophora stylosa | 0.105D²˙⁶⁸ | [54] |
| Sonneratia caseolaris | 0.04975D¹˙⁹⁴⁷⁴⁸ | [55] |
| All other species* | 0.251ρD²˙⁴⁶ | [56] |
| Below Ground Biomass | ||
| Avicennia marina | 1.28D¹˙¹⁷ | [57] |
| Rhizophora stylosa | 0.134D²˙⁴ | [58] |
| Sonneratia caseolaris | 0.0142D²˙¹²¹⁴⁶ | [55] |
| All other species | 0.199ρ0.899D²˙²² | [56] |
| * Wood density for species without an equation (ρ): Avicennia alba (0.72); Avicennia officinalis (0.72); Bruguiera gymnirosa (0.77); Ceriops decandra (0.78); Ceriops tagal (0.78); Lumnitzera racemose (0.87); Rhizophora apiculata (0.85); Rhizophora mucronate (0.82); Sonneratia alba (0.51); Xylocarpus granatum (0.70) | ||
2.5. Soil
Soil samples were taken from six plots per site (Figure 2). Cores were taken at the centre of the plots of topsoil (0-30cm) and subsoil (greater than 30cm). If the centre was occupied by a mangrove or crab burrow, the sample was taken immediately adjacent. The organic carbon, nitrogen and aluminium levels were analysed at Universitas Udayana. We calculated the organic carbon as % via the Walkley and Black method and Spectrophometry UV-VIS; nitrogen as % via the Kjeldhal method and titration; available aluminium as mg/kg via spark-atomic emission spectrometry [59] (Table S2).
2.6. Crabs
A 1m2 quadrat was marked out in six plots per site (Figure 2). This was done in an area along the transect where the entire plot was visible. This was to ensure that no crabs were missed and that mangrove roots were not obstructing the view of the plot. All members of the team moved away from the area and a 3-minute adjustment period was given. This was decided as the crabs were previously observed to come out of hiding after a few minutes after any stimulus was removed. The crab species and abundance were observed within the quadrat for 10 minutes to avoid double counting of any individuals. The member of the team conducting this sat 1m away from the plot and remained motionless for the 10-minute period. Furthermore, where the team member sat was chosen to avoid any visual stimulation such as shadows, influencing the crabs’ behaviours or choice to emerge from their burrows. We used morphospecies and identified the genus when possible (Table S2).
2.7. Fishing Pressure
Fishing activity was recorded adhoc at each site during the surveying period ranging from 7 hrs 29 mins to 20 hrs 20 mins. We recorded any boats netting, line fishing or returning to the mangrove site. The various fishing pressures observed were categorised based on [19]. Fishing pressure, even just local fishing can have a large impact on the fish species richness, the size, and the abundance in which they are found within the mangroves [60]. In addition to this, if the fishing methods are destructive or are continuously happening then the impact and damage caused is expected to increase over time [60,61].
2.8. Data Analysis
We tested whether the variables indicating resource availability and health of mangrove ecosystems were different between considering management and presence of aquaculture. In particular, we considered vegetation parameters (AGB, BGB, species richness), soil parameters (Aluminium, organic carbon, Nitrogen), and crab assemblages (richness, abundance, diversity) as dependent variables. We considered the combined effect of habitat management (managed vs unmanaged) and aquaculture (presence vs absence of aquaculture) as fixed factor. We used the nested term (1|Region/Site) as random effect to account for spatial closeness of plots. We used the function “glmmTMB” to run generalised linear mixed models and tested the family distributions offered by the corresponding package [62]. We then checked the model residuals plotted via the package “DHARMa” to decide on the family fit [63]. To check for potential links between the variables of investigation, we ran Pearson correlations via “rcorr” function and plotted them via the “corrplot” function [64]. For this, we considered only the 36 plots in which we collected all the data. We used p=0.05 as threshold for significance, and p=0.1 as threshold to define trends towards significance.
3. Results
3.1. Mangrove Vegetation
Between the six sites we found 13 species of mangroves, with site 1 having the highest richness. Sites 5 and 6 had a few large trees while the other sites, especially site 1, were dominated by tree saplings (Table 2).
Table 2.
Vegetation parameters by site.
| Variablea | Site 1 | Site 2 | Site 3 | Site 4 | Site 5 | Site 6 | Total |
|---|---|---|---|---|---|---|---|
| Richness | 12 | 8 | 7 | 6 | 6 | 6 | 13 |
| N trees | 1747 | 459 | 892 | 457 | 22 | 56 | 3633 |
| Mean DBH | 9.3 | 15.7 | 18.1 | 18.5 | 109.0 | 36.7 | 34.5 |
| Mean height | 2.5 | 4.2 | 4.5 | 4.1 | 6.0 | 4.9 | 4.4 |
| Mean diversity | 0.9 | 0.8 | 1.2 | 1.0 | 0.4 | 0.6 | 0.8 |
a Mean DBH, height and diversity are calculated based on 15 plots per site.
3.2. Soil
Aquaculture plots had higher topsoil Al, topsoil and subsoil N, and had lower topsoil C than non-aquaculture sites. Restored sites had less N than unmanaged sites (Figure 4). In particular, Al in topsoil tended to be higher in aquaculture restored plots than in non-aquaculture unmanaged plots. The aquaculture unmanaged plots had a higher nitrogen content in the topsoil than non-aquaculture sites (both unmanaged and restored). The aquaculture plots (both unmanaged and restored) had a higher nitrogen content in the subsoil than non-aquaculture sites (both unmanaged and restored).
3.3. Crabs
We found 18 species of crabs overall among the six sites, with site 1 hosting the highest richness and abundance, site 2 hosting the highest diversity, site 4 hosting the lowest richness and diversity, and site 6 hosting the lowest abundance (Table 3).
The abundance and richness of crabs was significantly higher in aquaculture sites than in no aquaculture sites, in particular the unmanaged aquaculture sites had higher abundance and richness than non-aquaculture sites (either unmanaged or restored) (Figure 5).
3.4. Correlations between Variables
Apart from the correlations that were expected between richness, abundance and diversity, we found the following correlations that are worth highlighting (Figure 6). The mean DBH of mangroves in plots was negatively correlated with crab abundance and richness, and the mean height of mangroves in plots was negatively correlated with crab abundance. The content of nitrogen in the subsoil was positively correlated with mangrove richness, diversity and height. The content of aluminium in the subsoil was negatively correlated with the content of organic carbon both in topsoil and subsoil.
3.5. Fishing Pressure
Fishing practices were observed in all sites during the data collection periods (Figure 7). Boats would sell their fish at a fish market near mangrove sites, thus were classed as offshore commercial fisheries. Inshore mixed species fisheries included people standing onshore or waist-deep in the ocean using a rod-and-line fishing method. Other fishing practices were also observed including the use of gill nets, cast in a circular motion by people standing in hip-deep shallow water who then reduced the net area to entrap the fish, as well as people casting small gill nets or using the rod-and-line method from a small boat. The inshore mollusks and crustacean fishing included mangrove crab traps, digging for mollusks and crustaceans when the tide was lower, and sifting the water to collect shrimps.
4. Discussion
From our investigation, we found that vegetation parameters were not affected by presence of aquaculture or restoration management, although we found site-specific variability. Crab richness and abundance were higher in aquaculture sites, and plots with smaller trees were preferred. Soil characteristics (carbon, nitrogen and aluminium) were highly impacted by presence of aquaculture and presence of restoration management. In addition, nitrogen in the subsoil was positively correlated with mangrove richness, diversity and mean height, and aluminium in the subsoil was linked with a decrease in organic carbon in both subsoil and topsoil. There was no difference between the AGB and BGB throughout the sites that were surveyed. Site 1 with the non-restored aquaculture was the sites with the least amount of biomass. This could have been due to the lack of natural recruitment able to take place due to the typography changes created by the aquaculture establishment, there-fore limiting the access for seeds or propagules to secure themselves. Therefore, the active restoration of the mangroves has been successful in increasing the biomass of the ecosystem. Studies suggest that this allows for increased resources in regard to carbon sequestration and a healthier ecosystem [65].
It has been reported that the higher the biomass of a forest the more potential for carbon sequestration it contains, and that biomass data can be used to calculate carbon storage [6]. Organisations such as the IUCN are driving for Blue Carbon to be introduced to REDD+ [66], which may be able to provide short-term funding for conservation projects involved in the restoration of mangrove ecosystems. The short-term funding could be provided by governments or private businesses involved in offsetting carbon emissions [66,67]. This provides stakeholder motivation to support the agreement governments have made to reach an equilibrium between human-made emissions and the absorption of these by 2100 [68]. Even though this may not be applicable on a global scale, for countries such as Indonesia, blue carbon could become a large source of carbon storage to help meet objectives set by the Paris Agreement [68,69].
The soil samples collected showed that there were increased levels of Al and N compared to that of non-aquaculture sites. This is mostly likely due to the human dis-turbances, such as aquaculture, disrupting the natural levels of N within the ecosystem [21]. This is also supported by other research showing that the levels of N increase in mangrove ecosystems where there are increase human influence [37]. This suggests that mangrove ecosystems that have been had aquaculture present have potentially influenced the nitrifying bacteria in the ecosystem that has the ability to break down and release N [21]. Even though N is essential for growth the increased levels can create an unbalanced ecosystem. However, areas that have been restored showed decreased levels of N, compared to that of the unmanaged sites. This could be because the activities required for restoration to occur are hindering the amount of N that is within the ecosystem. Furthermore, areas that are undergoing restoration or are exposed to any human disturbances could potentially alter ecosystem resources provided by the mangrove [70]. Whilst the restoration may be influencing the N content observed, the benefit to restoring these ecosystems should outweigh the impact of increasing the N levels during any restoration activities.
The Al topsoil of the restored aquaculture site has higher levels than that of the unmanaged site. This could be due to the restored site having more active aquaculture occurring than that of the unmanaged site, therefore increasing the level of Al observed. Whilst this was true for the aquaculture site, overall the influence of aquaculture shows an increase in the amount of Al, thus suggesting that any aquaculture present increases the amount of Al released into the ecosystem [71]. This can be harmful to the growth of propagules, therefore influencing the amount of growth occurring within the ecosystem, although there has also been stated that mangroves are able to withstand aluminium and this does not influence their growth as much as the varying environmental factors [72,73]. This is supported by the little variance in the biomass across the sites, therefore, it could been argued that metals found in mangroves are not necessarily influenced by aquaculture but through the composition of the soil itself [74]. The results gathered from this study contend this due to the increased levels of Al observed in the aquaculture sites. This could be explained however, by the positioning of these sites being set further inland compared to the others, therefore the soil composition would be different. Even though this is true the other sites show similarities between the Al in the subsoils, suggesting that the soil composition differences may not be the sole contributing factor to the increase in Al in the topsoil.
The most frequently observed family of crabs, throughout all ecosystems, was Grapsidae, which typically predate upon propagules as their main source of food [75]. As such, increased propagule availability could also increase the abundance of crabs from this family, given the propensity of these species to feed on the propagules. In particular, smaller propagules have been shown to be at greater risk of predation from crabs [76]. On the other hand, since it can take up to six years for a mangrove to produce propagules [77], areas where there are predominantly younger mangroves, are likely to have fewer propagules. For example, mangrove ecosystems that are being restored from a completely degraded state could see fewer Grapsidae species of crabs when conducting these surveys at the beginning of the restoration. It should be noted, however, that the average age of individual trees present is not the only factor that could influence propagule abundance. For example, mangrove ecosystems that are degraded could also produce fewer propagules due to a change in the coastal environment [35], and could therefore see fewer crabs.
Sites 3 and 4 are located within the GITA NADA MPA which has an area of 210km² [78]. At both of these sites, rod and line fishing methods were observed. When looking at this area on the Marine Conservation Institute [78] database of MAPs, the fishing level of protection for this area is classed as “Less Protected/Unknown”. From local knowledge, the area where the mangrove ecosystems that were surveyed are located, is classed as a rod and line or sustenance only fishing zone. This could also explain why there was an increase in the number of fish sightings at these sites, as the zone implemented minimises the impact that fishing has on the fish assemblages in the area [79]. Sites 1 and 2 proved to be interesting with the aquaculture observations. The utilisation of the mangroves in this way was seen in both sites however, site 1 was found to have more additional fishing pressure. This is most likely due to the protection offered by the local department of forestry at site 2. They are conducting the restoration on site 2 and are allowing locals to use the site for aquaculture, and so may be restricting fishing access. Additionally, the aquaculture permitted appears to be minimal based on the observations made at the site during this study.
This was not the same for the restored site 5, where numerous fishing techniques were observed. Given that there appeared to be very little presence from the organisations running the restoration program at that site, it is highly probable that the site is not being effectively monitored which may have led to an increase in fishing pressure. Additionally, the survey at this site only took a total of ten hours and ten minutes, so this only provides a snapshot of the fishing pressures suggesting that it could be greater than was observed. On the other hand, the short survey time also means that any monitoring that is in place for this site could have been missed. Additionally, site 6 appeared to have little to no regulations regarding any of the fishing vessels seen or the fishing methods being used. Monitoring fishing pressure is vital to ensure the productivity of the mangrove ecosystems and to ensure that local people are able to benefit from them, not only now, but in the future as well [80]. Both site 5 and 6 support a large fishing industry for the local communities surrounding them [81]. As such, both sites should be continually monitored with data being collected not only on the health of the mangroves but also regarding the fishing pressure and catch per unit effort. This data could not only provide vital information on the fish diversity and richness of the species that are occupying the mangroves but could also provide insight into that of the areas outside of the MPA or protected ecosystem that are benefitting from a spill-over effect [82].
5. Conclusions
In conclusion, we found that presence of aquaculture and management does not necessarily impact vegetation structure in mangrove ecosystems, but there are more hidden mechanisms that should be considered. First, aluminium content is high in aquaculture sites, and this also determined a negative flow of organic carbon available. Second, nitrogen content, which is positively linked with mangrove growth, was higher in aquaculture sites but this can be due to the presence of nitrifying bacteria in the ecosystem that has the ability to break down and release N. The presence of restoration projects decreased available nitrogen in the ecosystem, potentially suggesting that the activities required for restoration to occur are hindering the amount of N that is available within the ecosystem. Crab richness and abundance were higher in aquaculture sites and in plots with small trees, suggesting that the species of crabs present preferred areas with propagules for feeding. Finally, on a worrying note we found fishing pressure in all the sites surveyed during our assessment, and we suggest that the sustainability of fishing should be evaluated.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: dataset used on mangrove vegetation parameters; Table S2: dataset used on soil, crabs and linked mangrove vegetation.
Author Contributions
Conceptualization, C.H. and M.C.; methodology, C.H. and M.C.; software, C.H. and M.C.; validation, C.H. and M.C.; formal analysis, C.H. and M.C.; investigation, C.H.; resources, C.H., I.N.Y.P, K.M. and M.C.; data curation, C.H.; writing—original draft preparation, C.H., M.W.B. and M.C.; writing—review and editing, J.C., I.N.Y.P. and K.M.; visualization, C.H. and M.C.; supervision, M.C. and M.W.B.; project administration, M.C. and K.M.; funding acquisition, C.H., J.C. and M.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Oxford Brookes University HLS Developing Potential Research Excellence Award and by the Royal Geographical Society Geographical Fieldwork Grant.
Data Availability Statement
Data is contained within the article or supplementary material.
Acknowledgments
We thank Indonesia’s National Research and Innovation Agency in Jakarta (BRIN) for authorizing this study.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Appendix A
Table A1.
Results of the Generalised Linear Mixed Models.
| Response variable | Predictor | Estimate | Std Error | Z value | P Value |
|---|---|---|---|---|---|
| Crab abundance | Intercept | 3.09 | 0.35 | 8.93** | <0.001 |
| No aquaculture vs aquaculture | -1.30 | 0.49 | -2.59** | 0.010 | |
| Managed vs unmanaged | -0.72 | 0.53 | -1.36 | 0.174 | |
| Interaction effect | 0.85 | 0.73 | 1.16 | 0.246 | |
| Crab richness | Intercept | 1.51 | 0.29 | 5.27** | <0.001 |
| No aquaculture vs aquaculture | -1.21 | 0.45 | -2.69** | 0.007 | |
| Managed vs unmanaged | -0.70 | 0.48 | -1.46 | 0.146 | |
| Interaction effect | 0.80 | 0.68 | 1.18 | 0.239 | |
| Crab diversity | Intercept | -0.58 | 0.56 | -1.04 | 0.297 |
| No aquaculture vs aquaculture | -1.13 | 0.83 | -1.37 | 0.171 | |
| Managed vs unmanaged | -0.36 | 0.85 | -0.43 | 0.668 | |
| Interaction effect | -0.01 | 1.26 | -0.01 | 0.994 | |
| Mangrove abundance | Intercept | 4.16 | 2.02 | 2.06* | 0.039 |
| No aquaculture vs aquaculture | -1.13 | 2.02 | -0.56 | 0.575 | |
| Managed vs unmanaged | -0.10 | 0.62 | -0.16 | 0.874 | |
| Interaction effect | -0.26 | 0.80 | -0.33 | 0.744 | |
| Mangrove AGB | Intercept | 5.91 | 0.39 | 15.15** | <0.001 |
| No aquaculture vs aquaculture | 0.71 | 0.47 | 1.53 | 0.126 | |
| Managed vs unmanaged | 0.45 | 0.54 | 0.83 | 0.409 | |
| Interaction effect | -0.67 | 0.65 | -1.02 | 0.306 | |
| Mangrove BGB | Intercept | 5.64 | 0.36 | 15.48** | <0.001 |
| No aquaculture vs aquaculture | 0.04 | 0.45 | 0.10 | 0.925 | |
| Managed vs unmanaged | 0.17 | 0.51 | 0.33 | 0.742 | |
| Interaction effect | -0.02 | 0.62 | -0.03 | 0.979 | |
| Mangrove richness | Intercept | 1.18 | 1.22 | 0.94 | 0.335 |
| No aquaculture vs aquaculture | -0.79 | 1.23 | -0.64 | 0.522 | |
| Managed vs unmanaged | -0.23 | 0.33 | -0.68 | 0.500 | |
| Interaction effect | 0.09 | 0.41 | 0.23 | 0.822 | |
| Subsoil Al | Intercept | 6.95 | 0.37 | 19.01** | <0.001 |
| No aquaculture vs aquaculture | -0.31 | 0.57 | -0.54 | 0.589 | |
| Managed vs unmanaged | 0.49 | 0.47 | 1.04 | 0.297 | |
| Interaction effect | 0.21 | 0.60 | 0.34 | 0.731 | |
| Subsoil C | Intercept | 2.20 | 0.49 | 4.48** | <0.001 |
| No aquaculture vs aquaculture | 0.95 | 0.60 | 1.57 | 0.116 | |
| Managed vs unmanaged | 0.59 | 0.70 | 0.84 | 0.400 | |
| Interaction effect | -0.64 | 0.85 | -0.76 | 0.450 | |
| Subsoil N | Intercept | -2.26 | 0.17 | -13.53** | <0.001 |
| No aquaculture vs aquaculture | -0.85 | 0.23 | -3.64** | <0.001 | |
| Managed vs unmanaged | 0.08 | 0.23 | 0.37 | 0.715 | |
| Interaction effect | -0.04 | 0.32 | -0.12 | 0.902 | |
| Topsoil Al | Intercept | 7.30 | 0.42 | 17.48** | <0.001 |
| No aquaculture vs aquaculture | -1.17 | 0.60 | -1.93t | 0.054 | |
| Managed vs unmanaged | 0.24 | 0.57 | 0.42 | 0.672 | |
| Interaction effect | 0.84 | 0.78 | 1.08 | 0.280 | |
| Topsoil C | Intercept | 2.07 | 0.48 | 4.35** | <0.001 |
| No aquaculture vs aquaculture | 1.01 | 0.58 | 1.73t | 0.084 | |
| Managed vs unmanaged | 0.38 | 0.67 | 0.56 | 0.573 | |
| Interaction effect | -0.58 | 0.82 | -0.71 | 0.480 | |
| Topsoil N | Intercept | -2.07 | 0.17 | -11.97** | <0.001 |
| No aquaculture vs aquaculture | -0.93 | 0.24 | -3.82** | <0.001 | |
| Managed vs unmanaged | -0.47 | 0.27 | -1.75t | 0.080 | |
| Interaction effect | 0.61 | 0.36 | 1.72t | 0.086 |
tp<0.1; *p<0.05; **p<0.01.
Table A2.
Contrast ratios (means and SE) and correspondent Z values and p values for the interaction effect between no aquaculture (NoA) vs aquaculture (Aqu) and managed (Man) vs unmanaged (Unm).
Table A2.
Contrast ratios (means and SE) and correspondent Z values and p values for the interaction effect between no aquaculture (NoA) vs aquaculture (Aqu) and managed (Man) vs unmanaged (Unm).
| Variable | AquUnm/ NoAUnm |
AquUnm/ AquMan |
AquUnm/ NoAMan |
NoAUnm/ AquMan |
NoAUnm/ NoAMan |
AquMan/ NoAMan |
|---|---|---|---|---|---|---|
| Crab abundance | 3.60 ± 1.78 Z=2.6*, p=0.047 |
2.05 ± 1.08 Z=1.4, p=0.524 |
3.14 ± 1.53 Z=2.4t, p=0.087 |
0.57 ± 0.31 Z=-1.0, p=0.737 |
0.87 ± 0.45 Z=-0.3, p=0.994 |
1.53 ± 0.84 Z=0.8, p=0.862 |
| Crab richness | 3.35 ± 1.50 Z=2.7*, p=0.035 |
2.01 ± 0.96 Z=1.5, p=0.465 |
3.01 ± 1.33 Z=2.5t, p=0.059 |
0.60 ± 0.31 Z=-1.0, p=0.758 |
0.90 ± 0.44 Z=-0.2, p=0.996 |
1.50 ± 0.77 Z=0.8, p=0.859 |
| Crab diversity | 3.10 ± 2.56 Z=1.4, p=0.520 |
1.44 ± 1.23 Z=0.4, p=0.974 |
4.50 ± 4.05 Z=1.7, p=0.337 |
0.47 ± 0.41 Z=-0.9, p=0.823 |
1.45 ± 1.35 Z=0.4, p=0.978 |
3.13 ± 2.98 Z=1.2, p=0.630 |
| Mangrove abundance |
3.11 ± 6.27 Z=0.6, p=0.944 |
1.10 ± 0.69 Z=0.2, p=0.999 |
4.45 ± 9.23 Z=0.7, p=0.889 |
0.36 ± 0.58 Z=-0.6, p=0.920 |
1.43 ± 0.71 Z=0.7, p=0.888 |
4.03 ± 6.80 Z=0.8, p=0.842 |
| Mangrove AGB | 0.49 ± 0.23 Z=-1.5, p=0.420 |
0.64 ± 0.35 Z=-0.8, p=0.842 |
0.61 ± 0.29 Z=-1.0, p=0.722 |
1.31 ± 0.59 Z=0.6, p=0.935 |
1.25 ± 0.46 Z=0.6, p=0.931 |
0.96 ± 0.44 Z=-0.1, p=0.999 |
| Mangrove BGB | 0.96 ± 0.43 Z=-0.1, p=0.999 |
0.85 ± 0.43 Z=-0.3, p=0.988 |
0.82 ± 0.37 Z=-0.4, p=0.972 |
0.88 ± 0.39 Z=-0.3, p=0.992 |
0.85 ± 0.31 Z=-0.4, p=0.975 |
0.98 ± 0.43 Z=-0.1, p=1.000 |
| Mangrove richness | 2.20 ± 2.71 Z=0.6, p=0.919 |
1.25 ± 0.42 Z=0.7, p=0.907 |
2.51 ± 3.15 Z=0.7, p=0.884 |
0.57 ± 0.56 Z=-0.6, p=0.941 |
1.14 ± 0.28 Z=0.5, p=0.949 |
2.00 ± 2.03 Z=0.7, p=0.902 |
| Subsoil Al | 1.36 ± 0.77 Z=0.5, p=0.995 |
0.61 ± 0.29 Z=-1.0, p=0.880 |
0.68 ± 0.34 Z=-0.8, p=0.968 |
0.45 ± 0.25 Z=-1.4, p=0.634 |
0.50 ± 0.19 Z=-1.9, p=0.326 |
1.11 ± 0.54 Z=0.2, p=1.000 |
| Subsoil C | 0.71 ± 0.16 Z=-1.6, p=0.454 |
0.80 ± 0.24 Z=-0.8, p=0.741 |
0.73 ± 0.16 Z=-1.5, 0.454 |
1.13 ± 0.23 Z=0.6, p=0.742 |
1.02 ± 0.22 Z=0.1, p=0.907 |
0.90 ± 0.24 Z=-0.5, p=0.742 |
| Subsoil N | 2.33 ± 0.54 Z=3.6**, p=0.002 |
0.92 ± 0.21 Z=-0.4, p=0.983 |
2.23 ± 0.51 Z=3.47**, p=0.003 |
0.40 ± 0.09 Z=-4.1**, p<0.001 |
0.96 ± 0.22 Z=-0.2, p=0.997 |
2.42 ± 0.55 Z=3.9**, p<0.001 |
| Topsoil Al | 3.21 ± 1.94 Z=1.9, p=0.281 |
0.79 ± 0.45 Z=-0.4, p=0.999 |
1.09 ± 0.56 Z=0.2, p=1.000 |
0.25 ± 0.14 Z=-2.4t, p=0.090 |
0.34 ± 0.18 Z=-2.0, p=0.226 |
1.34 ± 0.68 Z=0.66, p=0.986 |
| Topsoil C | 0.67 ± 0.19 Z=-1.7, p=0.535 |
0.85 ± 0.20 Z=-0.6, p=0.673 |
0.72 ± 0.18 Z=-1.4, p=0.535 |
1.26 ± 0.21 Z=1.1, p=0.580 |
1.07 ± 0.14 Z=0.4, p=0.673 |
0.85 ± 0.20 Z=-0.7, p=0.673 |
| Topsoil N | 2.54 ± 0.62 Z=3.8**, p<0.001 |
1.60 ± 0.43 Z=1.7, p=0.122 |
2.20 ± 0.52 Z=3.32**, p=0.003 |
0.63 ± 0.17 Z=-1.7, p=0.122 |
0.87 ± 0.20 Z=-0.6, p=0.538 |
1.38 ± 0.36 Z=1.2, p=0.264 |
tp<0.1; *p<0.05; **p<0.01.
References
- Biswas, P.L.; Biswas, S.R. Mangrove Forests: Ecology, Management, and Threats. In Life on Land, Encyclopedia of the UN Sustainable Development Goals; Leal Filho, W., Azul, A.M., Brandli, L., Özuyar, P.G., Wall, T., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–14. [Google Scholar]
- Kathiresan, K.; Bingham, B.L. Biology of Mangroves and Mangrove Ecosystems. Adv. Mar. Biol. 2001, 40, 81–251. [Google Scholar]
- Krauss, K.W.; Ball, M.C. On the halophytic nature of mangroves. Trees-Struct. Funct. 2013, 27, 7–11. [Google Scholar] [CrossRef]
- Naidoo, T.; Rajkaran, A.; Sershen. Impacts of plastic debris on biota and implications for human health: A South African perspective. S. Afr. J. Sci. 2020, 116, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Suratman, M.N. Carbon Sequestration Potential of Mangroves in Southeast Asia. Manag. For. Ecosyst. Chall. Clim. Chang. 2008. [Google Scholar]
- Kauffman, J.C.H.; Thomas, G.C.; Kathleen, A.D.; Daniel, C.; Donato, D. Ecosystem carbon stocks of micronesian mangrove forests. Wetlands 2011, 31, 343–352. [Google Scholar] [CrossRef]
- Ferraz, A.; Saatchi, S.S.; Xu, L.; Hagen, S.C.; Chave, J.; Yu, Y.; Meyer, V.; Garcia, M.; Silva, C.A.; Roswintiart, O.; et al. Carbon storage potential in degraded forests of Kalimantan, Indonesia. Environ. Res. Lett. 2018, 13, 095001. [Google Scholar] [CrossRef]
- Donato, D.C.; Kauffman, J.B.; Murdiyarso, D.; Kurnianto, S.; Stidham, M.; Kanninen, M. Mangroves among the most carbon-rich forests in the tropics. Nat. Geosci. 2011, 4, 293–297. [Google Scholar] [CrossRef]
- Chatting, M.; Al-Maslamani, I.; Walton, M.; Skov, M.W.; Kennedy, H.; Husrevoglu, Y.S.; Vay, L.L. Future mangrove carbon storage under climate change and deforestation. Front. Mar. Sci. 2022, 9, 781876. [Google Scholar] [CrossRef]
- Sánchez-Núñez, D.A.; Bernal, G.; Pineda, J.E.M. The relative role of mangroves on wave erosion mitigation and sediment properties. Estuaries Coasts 2019, 42, 2124–2138. [Google Scholar] [CrossRef]
- Kamil, E.A.; Takaijudin, H.; Hashim, A.M. Mangroves As Coastal Bio-Shield: A Review of Mangroves Performance in Wave Attenuation. Civ. Eng. J. 2021, 7, 1964–1981. [Google Scholar] [CrossRef]
- Lee, W.K.; Tay, S.H.X.; Ooi, S.K.; Friess, D.A. Potential short wave attenuation function of disturbed mangroves. Estuar. Coast. Shelf Sci. 2021, 248, 106747. [Google Scholar] [CrossRef]
- Giesen, W.; Wulffraat, S.; Zieren, M.; Scholten, L. Mangrove Guidebook for Southeast Asia; UN Food and Agricultural Organization: Phnom Penh, Combodi, 2007. [Google Scholar]
- Faridah-Hanum, I.; Kudus, K.A.; Saari, N.S. Plant Diversity and Biomass of Marudu Bay Mangroves in Malaysia. Pakistan J. Bot. 2012, 44, 151–156. [Google Scholar]
- Twilley, R.R.; Snedaker, S.C.; Yaez-Arancibia, A.; Medina, E. Biodiversity and ecosystem processes in tropical estuaries: perspectives of mangrove ecosystems. Scope-scientific committee on problems of the Environment International Council of Scientific Unions 1996, 55, 327–370. [Google Scholar]
- Mumby, P.J.; Edwards, A.J.; Rias-Gonzalez, J.E.; Lindeman, K.C.; Blackwell, P.G.; Gall, A.; Gorczynska, M.I.; Harborne, A.R.; Pescod, C.L.; Renken, H.; et al. Mangroves enhance the biomass of coral reef fish communities in the Caribbean. Nature 2004, 427, 533–536. [Google Scholar] [CrossRef]
- Satyanarayana, B.; Quispe-Zuniga, M.R.; Hugé, J.; Sulong, I.; Mohd-Lokman, H.; Dahdouh-Guebas, F. Mangroves Fueling Livelihoods: A Socio-Economic Stakeholder Analysis of the Charcoal and Pole Production Systems in the World’s Longest Managed Mangrove Forest. Front. Ecol. Evol. 2021, 9, 621721. [Google Scholar] [CrossRef]
- Aziz, A.A.; Dargusch, P.; Phinn, S.; Ward, A. Using REDD+ to balance timber production with conservation objectives in a mangrove forest in Malaysia. Ecol. Econ. 2015, 120, 108–116. [Google Scholar] [CrossRef]
- Hutchison, J.; Spalding, M.; zu Ermgassen, P. The Role of Mangroves in Fisheries Enhancement; The Nature Conservancy: Arlington County, VA, USA; Wetlands International: Wageningen, The Netherlands, 2014; pp. 1–54. [Google Scholar]
- Alongi, D.M.D.M. Present State and Future of the World’s Mangrove Forests. Environ. Conserv. 2002, 29, 331–349. [Google Scholar] [CrossRef]
- Alongi, D.M. Climate Change and Mangroves. In Mangroves: Biodiversity, Livelihoods and Conservation; Springer Nature: Singapore, 2022; pp. 175–198. [Google Scholar]
- Polidoro, B.A.; Carpenter, K.E.; Collins, L.; Duke, N.C.; Ellison, A.M.; Ellison, J.C.; Farnsworth, E.J.; Fernando, E.S.; Kathiresan, K.; Koedam, N.E.; et al. The Loss of Species: Mangrove Extinction Risk and Geographic Areas of Global Concern. PLoS ONE 2010, 5, e10095. [Google Scholar] [CrossRef]
- Luther, D.A.; Greenberg, R. Mangroves: A Global Perspective on the Evolution and Conservation of Their Terrestrial Vertebrates. BioScience 2009, 59, 602–612. [Google Scholar] [CrossRef]
- Adame, M.F.; Connolly, R.M.; Turschwell, M.P.; Lovelock, C.E.; Fatoyinbo, T.; Lagomasino, D.; Goldberg, L.A.; Holdorf, J.; Friess, D.A.; Sasmito, S.D.; et al. Future carbon emissions from global mangrove forest loss. Glob. Chang. Biol. 2021, 27, 2856–2866. [Google Scholar] [CrossRef]
- Arifanti, V.B.; Sidik, F.; Mulyanto, B.; Susilowati, A.; Wahyuni, T.; Subarno; Yulianti; Yuniarti, N.; Aminah, A.; Suita, E.; et al. Challenges and Strategies for Sustainable Mangrove Management in Indonesia: A Review. Forests 2022, 13, 695. [Google Scholar] [CrossRef]
- Luom, T.T.; Phong, N.T.; Smithers, S.; Van Tai, T. Protected mangrove forests and aquaculture development for livelihoods. Ocean Coast. Manag. 2021, 205, 105553. [Google Scholar] [CrossRef]
- Miteva, D.A.; Murray, B.C.; Pattanayak, S.K. Do protected areas reduce blue carbon emissions? A quasi-experimental evaluation of mangroves in Indonesia. Ecol. Econ. 2015, 119, 127–135. [Google Scholar] [CrossRef]
- Tengku Hashim, T.M.Z.; Engku Ariff, E.A.R.; Suratman, M.N. Aquaculture in mangroves. Mangroves: Ecology, Biodiversity and Management, 2021, pp. 419–438.
- Richards, D.R.; Friess, D.A. Rates and drivers of mangrove deforestation in Southeast Asia, 2000–2012. Proc. Natl. Acad. Sci. USA 2016, 113, 344–349. [Google Scholar] [CrossRef]
- Valiela, I.; Bowen, J.L.; York, J.K. Mangrove Forests: One of the World’s Threatened Major Tropical Environments. Bioscience 2001, 51, 807–815. [Google Scholar] [CrossRef]
- McSherry, M.; Davis, R.P.; Andradi-Brown, D.A.; Ahmadia, G.N.; Van Kempen, M.; Brian, S.W. Integrated mangrove aquaculture: The sustainable choice for mangroves and aquaculture? Front. For. Glob. Chang. 2023, 6, 1094306. [Google Scholar] [CrossRef]
- Balmford, A.; Bruner, A.; Cooper, P.; Costanza, R.; Farber, S.; Green, R.E.; Jenkins, M.; Jefferiss, P.; Jessamy, V.; Madden, J.; et al. Economic Reasons for Conserving Wild Nature. Science 2002, 297, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Aheto, D.W.; Kankam, S.; Okyere, I.; Mensah, E.; Osman, A.; Jonah, F.E.; Mensah, J.C. Community-Based Mangrove Forest Management: Implications for Local Livelihoods and Coastal Resource Conservation along the Volta Estuary Catchment Area of Ghana. Ocean Coast. Manag. 2016, 127, 43–54. [Google Scholar] [CrossRef]
- Mesta, P.; Setturu, B.; Chandran, M.D.S.; Ramachandra, T.V. Inventorying, Mapping and Monitoring of Mangroves towards Sustainable Management of West Coast, India. Geophys. Remote Sens. 2014, 3, 3–9. [Google Scholar]
- Field, C.D. Rehabilitation of mangrove ecosystems, a review. Mar. Pollut. Bull. 1998, 37, 383–392. [Google Scholar] [CrossRef]
- Ibrahim, F.H.; Yusoff, F.M.; Fitrianto, A.; Nuruddin, A.A.; Gandaseca, S.; Samdin, Z.; Kamarudin, N.; Nurhidayu, S.; Kassim, M.R.; Hakeem, K.R.; Ibrahim, S. How to develop a comprehensive Mangrove Quality Index? MethodsX 2019, 6, 1591–1599. [Google Scholar] [CrossRef]
- Faridah-Hanum, I.; Yusoff, F.M.; Fitrianto, A.; Ainuddin, N.A.; Gandaseca, S.; Zaiton, S.; Norizah, K.; Nurhidayu, S.; Roslan, M.K.; Hakeem, K.R.; et al. Development of a comprehensive mangrove quality index (MQI) in Matang Mangrove; Assessing mangrove ecosystem health. Ecol. Indic. 2019, 102, 103–117. [Google Scholar] [CrossRef]
- Biswas, S.R.; Mallik, A.U.; Choudhury, J.K.; Nishat, A. A unified framework for the restored of Southeast Asian mangroves—Bridging ecology, society and economics. Wetl. Ecol. Manag. 2009, 17, 365–383. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nat. Cell Biol. 2012, 486, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Zaman, M.R.; Rahman, M.S.; Ahmed, S.; Zuidema, P.A. What drives carbon stocks in a mangrove forest? The role of stand structure, species diversity and functional traits. Estuar. Coast. Shelf Sci 2023, 295, 108556. [Google Scholar] [CrossRef]
- Duncanson, L.; Armston, J.; Disney, M.; Avitabile, V.; Barbier, N.; Calders, K.; Carter, S.; Chave, J.; Herold, M.; Crowther, T.W.; et al. The importance of consistent global forest aboveground biomass product validation. Surv. Geophys. 2019, 40, 979–999. [Google Scholar] [CrossRef] [PubMed]
- Mcleod, E.; Chmura, G.L.; Bouillon, S.; Salm, R.; Björk, M.; Duarte, C.M.; Lovelock, C.E.; Schlesinger, W.H.; Silliman, B.R. A Blueprint for Blue Carbon: Toward an Improved Understanding of the Role of Vegetated Coastal Habitats in Sequestering CO2. Front. Ecol. Environ. 2011, 9, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Inoue, T. Carbon sequestration in mangroves. Blue Carbon in Shallow Coastal Ecosystems. Carbon Dynamics, Policy, and Implementation, 2019, pp.73-99.
- Valiela, I.; Cole, M.L. Comparative evidence that saltmarshes and mangroves may protect sea grass meadow from land-derived nitrogen loads. Ecosystems 2002, 5, 92–102. [Google Scholar] [CrossRef]
- Ma, L.; Yang, S. Growth and physiological response of Kandelia obovata and Bruguiera sexangula seedlings to aluminum stress. Environ. Sci. Pollut. Res. 2022, 29, 43251–43266. [Google Scholar] [CrossRef]
- Ferreira, A.C.; Ganade, G.; de Attayde, J.L. Restoration versus natural regeneration in a neotropical mangrove: Effects on plant biomass and crab communities. Ocean Coast. Manag. 2015, 110, 38–45. [Google Scholar] [CrossRef]
- Aschenbroich, A.; Michaud, E.; Stieglitz, T.; Fromard, F.; Gardel, A.; Tavares, M.; Thouzeau, G. Brachyuran crab community structure and associated sediment reworking activities in pioneer and young mangroves of French Guiana, South America. Estuar. Coast. Shelf Sci. 2016, 182, 60–71. [Google Scholar] [CrossRef]
- Kristensen, E.; Penha-Lopes, G.; Delefosse, M.; Valdemarsen, T.; Quintana, C.O.; Banta, G.T. What is bioturbation? The need for a precise definition for fauna in aquatic sciences. Mar. Ecol. Prog. Ser. 2012, 446, 285–302. [Google Scholar] [CrossRef]
- Agusto, L.E.; Sara, F.; Jimenez, P.J.; Quadros, A.; Cannicci, S. Structural characteristics of crab burrows in Hong Kong mangrove forests and their role in ecosystem engineering. Estuar. Coast. Shelf Sci. 2020, 248, 106973. [Google Scholar] [CrossRef]
- Tobias, A.; Umali, A.G.A.; Malabrigo, P.; Galang, M.A. Mangrove Forest Inventory and Estimation of Carbon Storage and Sedimentation in Pagbilao. Wealth Account. Valuat. Ecosyst. Serv. 2017, 1–95. [Google Scholar]
- Dharmawan, I.W.S.; Siregar, C.A. Karbon tanah dan pendugaan karbon tegakan. Jurnal Penelitian Hutan Dan Konservasi Alam 2008, 5, 317–328. [Google Scholar] [CrossRef]
- Vinh, T.V.; Marchand, C.; Linh, T.V.K.; Vinh, D.D.; Allenbach, M. Allometric models to estimate above-ground biomass and carbon stocks in Rhizophora apiculata tropical managed mangrove forests (Southern Viet Nam). For. Ecol. Manag. 2019, 434, 131–141. [Google Scholar] [CrossRef]
- Fromard, F.; Puig, H.; Mougin, E.; Marty, G.; Betoulle, J.L.; Cadamuro, L. Structure, above-ground biomass and dynamics of mangrove ecosystems: New data from French Guiana. Oecologia 1998, 115, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Clough, B.F.; Scott, K. Allometric relationships for estimating above-ground biomass in six mangrove species. For. Ecol. Manag. 1989, 27, 117–127. [Google Scholar] [CrossRef]
- Hanh, N.T.H.; Tinh, P.H.; Tuan, M.S. Allometry and biomass accounting for mangroves Kandelia obovata Sheue, Liu & Yong and Sonneratia caseolaris (L.) Engler planted in coastal zone of Red River Delta, Vietnam. Int. J. Dev. Res 2016, 6, 7804–7808. [Google Scholar]
- Komiyama, A.; Poungparn, S.; Kato, S. Common allometric equations for estimating the tree weight of mangroves. J. Trop. Ecol. 2005, 21, 471–477. [Google Scholar] [CrossRef]
- Komiyama, A.; Ong, J.E.; Poungparn, S. Allometry, biomass, and productivity of mangrove forests: A review. Aquat. Bot. 2008, 89, 128–137. [Google Scholar] [CrossRef]
- Gevana, D.; Im, S. Allometric models for Rhizophora stylosa Griff. in dense monoculture plantation in the Philippines. Malaysian For. 2016, 79, 39–53. [Google Scholar]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H.; Soltanpour, P.N.; Tabatabai, M.A.; Johnston, C.T.; Sumner, M.E. Methods of Soil Analysis, Part 3–Chemical Methods; Soil Science Society of America Inc.: Madison, WI, USA, 1996. [Google Scholar]
- Reis-Filho, J.A.; Harvey, E.S.; Giarrizzo, T. Impacts of small-scale fisheries on mangrove fish assemblages. ICES J. Mar. Sci. 2018, 76, 153–164. [Google Scholar] [CrossRef]
- Tahiluddin, A.; Sarri̇, J. An Overview of Destructive Fishing in the Philippines. Acta Nat. Sci 2022, 3, 116–125. [Google Scholar] [CrossRef]
- Brooks, M.E.; Kristensen, K.; Van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Machler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modelling. R J 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Hartig, F. DHARMa: residual diagnostics for hierarchical (multi-level/mixed) regression models. R Packag version 020, 2018.
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J. Package ‘corrplot’. Statistician 2017, 56, e24. [Google Scholar]
- Zhila, H.; Mahmood, H.; Rozainah, M.Z. Biodiversity and biomass of a natural and degraded mangrove forest of Peninsular Malaysia. Environ. Earth Sci 2015, 71, 4629–4635. [Google Scholar] [CrossRef]
- Hilmi, N.; Chami, R.; Sutherland, M.D.; Hall-Spencer, J.M.; Lebleu, L.; Benitez, M.B.; Levin, L.A. The role of blue carbon in climate change mitigation and carbon stock conservation. Front. Clim. 2021, 3, 710546. [Google Scholar] [CrossRef]
- Herrera-Silveira, J.A.; Pech-Cardenas, M.A.; Morales-Ojeda, S.M.; Cinco-Castro, S.; Camacho-Rico, A.; Sosa, J.P.C.; Mendoza-Martinez, J.E.; Pech-Poot, E.Y.; Montero, J.; Teutli-Hernandez, C. Blue carbon of Mexico, carbon stocks and fluxes: a systematic review. PeerJ 2020, 8, e8790. [Google Scholar] [CrossRef]
- NFCCC C. (2015). Adoption of the Paris Agreement. I:proposal by the president (Draft Decision). https://unfccc.int/resource/docs/2015/cop21/eng/l09r01.pdf.
- Taillardat, P.; Friess, D.A.; Lupascu, M. Mangrove blue carbon strategies for climate change mitigation are most effective at the national scale. Biol. Lett. 2018, 14, 20180251. [Google Scholar] [CrossRef]
- Reis, C.R.G.; Nardoto, G.B.; Oliveira, R.S. Global overview on nitrogen dynamics in mangroves and consequences of increasing nitrogen availability for these systems. Plant Soil 2017, 410, 1–19. [Google Scholar] [CrossRef]
- Karstens, S.; Lukas, M.C. Contested aquaculture development in the protected mangrove forests of the Kapuas estuary, West Kalimantan. Geoöko 2014, 35, 78–121. [Google Scholar]
- Reef, R.; Feller, I.C.; Lovelock, C.E. Nutrition of mangroves. Tree Physiol 2010, 30, 1148–1160. [Google Scholar] [CrossRef]
- Oxmann, J.F.; Pham, Q.H.; Schwendenmann, L.; Stellman, J.M.; Lara, R.J. Mangrove reforestation in Vietnam: the effect of sediment physicochemical properties on nutrient cycling. Plant Soil 2010, 326, 225–241. [Google Scholar] [CrossRef]
- Tam, N.F.Y.; Yao, M. Normalisation and heavy metal contamination in mangrove sediments. SciTotal Environ 2019, 216, 33–39. [Google Scholar] [CrossRef]
- Parman, R.P.; Kamarudin, N.; Ibrahim, F.H.; Nuruddin, A.A.; Omar, H.; Abdul Wahab, Z. Geostatistical Analysis of Mangrove Ecosystem Health: Mapping and Modelling of Sampling Uncertainty Using Kriging. Forests 2022, 13, 1185. [Google Scholar] [CrossRef]
- Clarke, P.J.; Kerrigan, R.A. The effects of seed predators on the recruitment of mangroves. J Ecol 2002, 90, 728–736. [Google Scholar] [CrossRef]
- Imbert, D.; Rousteau, A.; Scherrer, P. Ecology of mangrove growth and recovery in the Lesser Antilles: State of knowledge and basis for restoration projects. Restor Ecol 2000, 8, 230–236. [Google Scholar] [CrossRef]
- Marine Conservation Institute. (2023) Global Marine Fishing Protection, Available at: Global Marine Protection | Marine Protection Atlas (mpatlas.org) (Accessed on 24 May 2024).
- Grorud-Colvert, K.; Sullivan-Stack, J.; Roberts, C.; Constant, V.; Horta e Costa, B.; Pike, E.P.; Kingston, N.; Laffoley, D.; Sala, E.; Claudet, J.; et al. The MPA Guide: A framework to achieve global goals for the ocean. Science 2021, 373, 1215. [Google Scholar] [CrossRef]
- Russ, G.R.; Alcala, A.C.; Maypa, A.P.; Calumpong, H.P.; White, A.T. Marine reserve benefits local fisheries. Ecol. Appl. 2004, 14, 597–606. [Google Scholar] [CrossRef]
- Gokkon, B. (2019). ‘Bali Mangrove Bay is Now a Conservation Zone, Nixing Reclamation Plan. Retrieved August 06, 2020’. Available at: https://news.mongabay.com/2019/10/bali-benoa-bay-mangrovesconservation-reclamation/ (Accessed: 6 May 2024).
- Gell, F.R.; Roberts, C.M. Benefits beyond boundaries: The fishery effects of marine reserves. Trends Ecol Evol 2003, 18, 448–455. [Google Scholar] [CrossRef]
Figure 1.
Location of the study sites in Bali and Lombok, Indonesia.
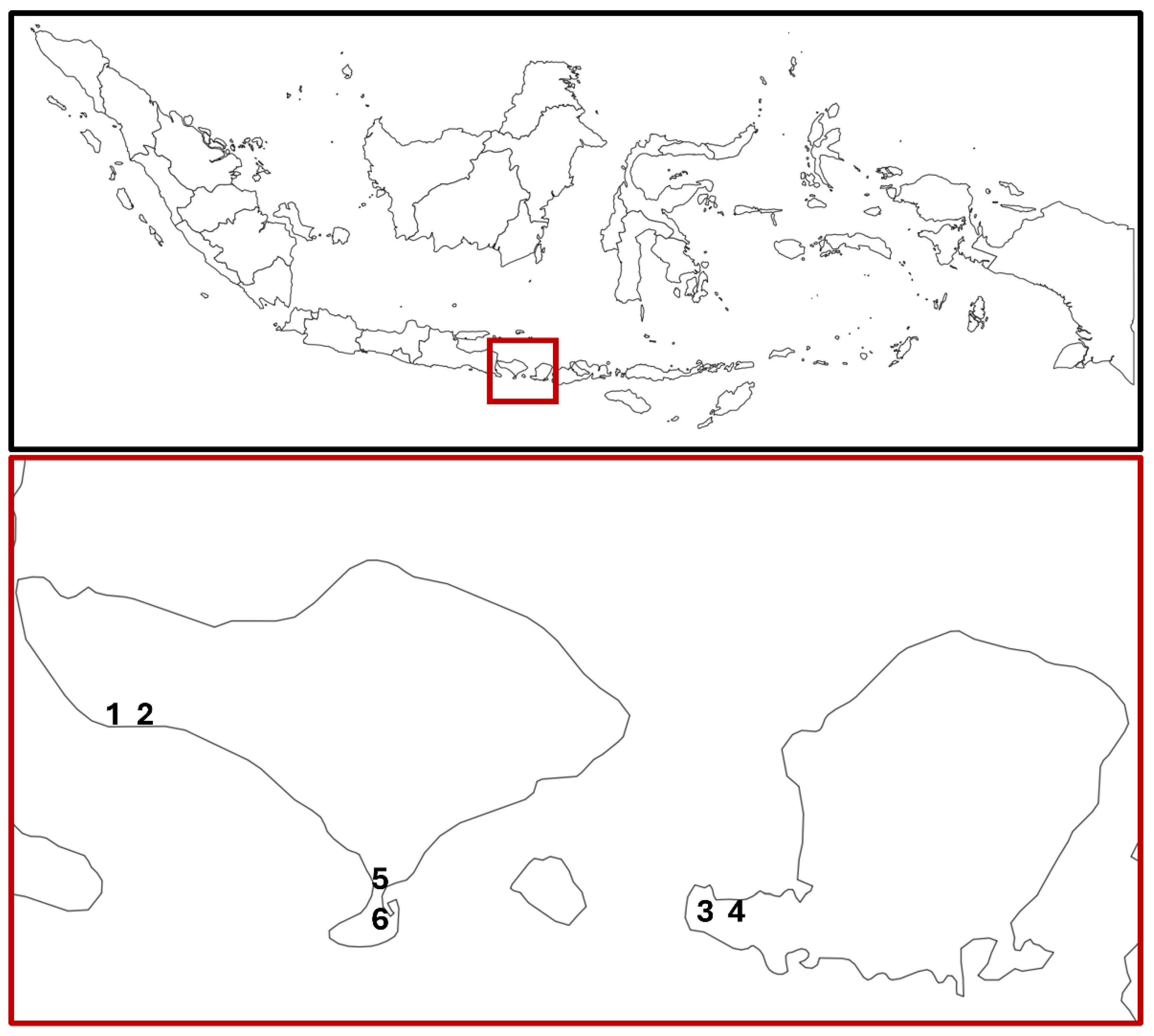
Figure 2.
Layout of the transects and plots to conduct the surveys at each site. The red plots show where the crab surveys were conducted and soil samples were taken. Vegetation data were collected in all plots.
Figure 2.
Layout of the transects and plots to conduct the surveys at each site. The red plots show where the crab surveys were conducted and soil samples were taken. Vegetation data were collected in all plots.
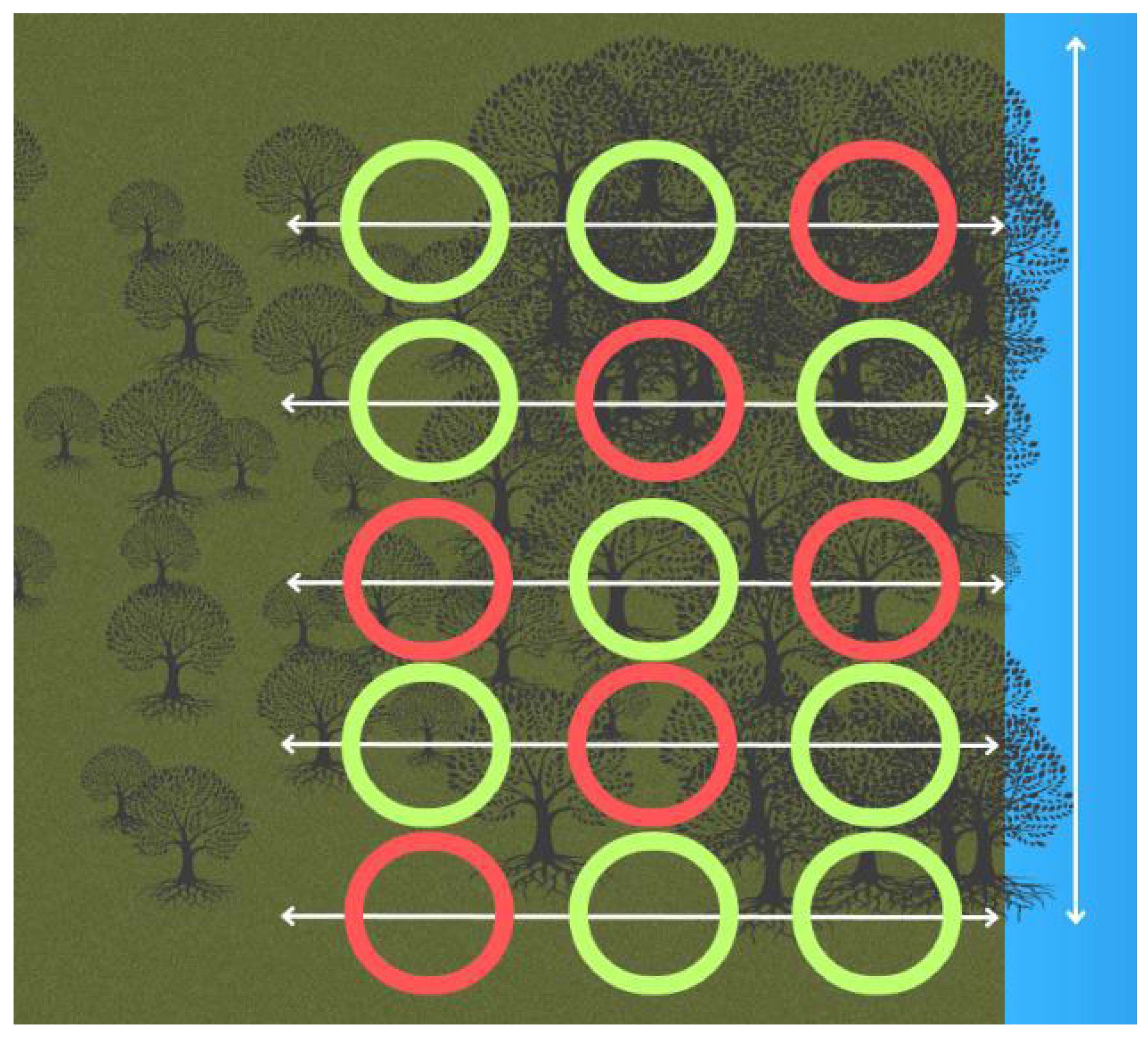
Figure 3.
Total biomass (ABG: Above Ground Biomass; BGB: Below Ground Biomass) in mangrove sites.
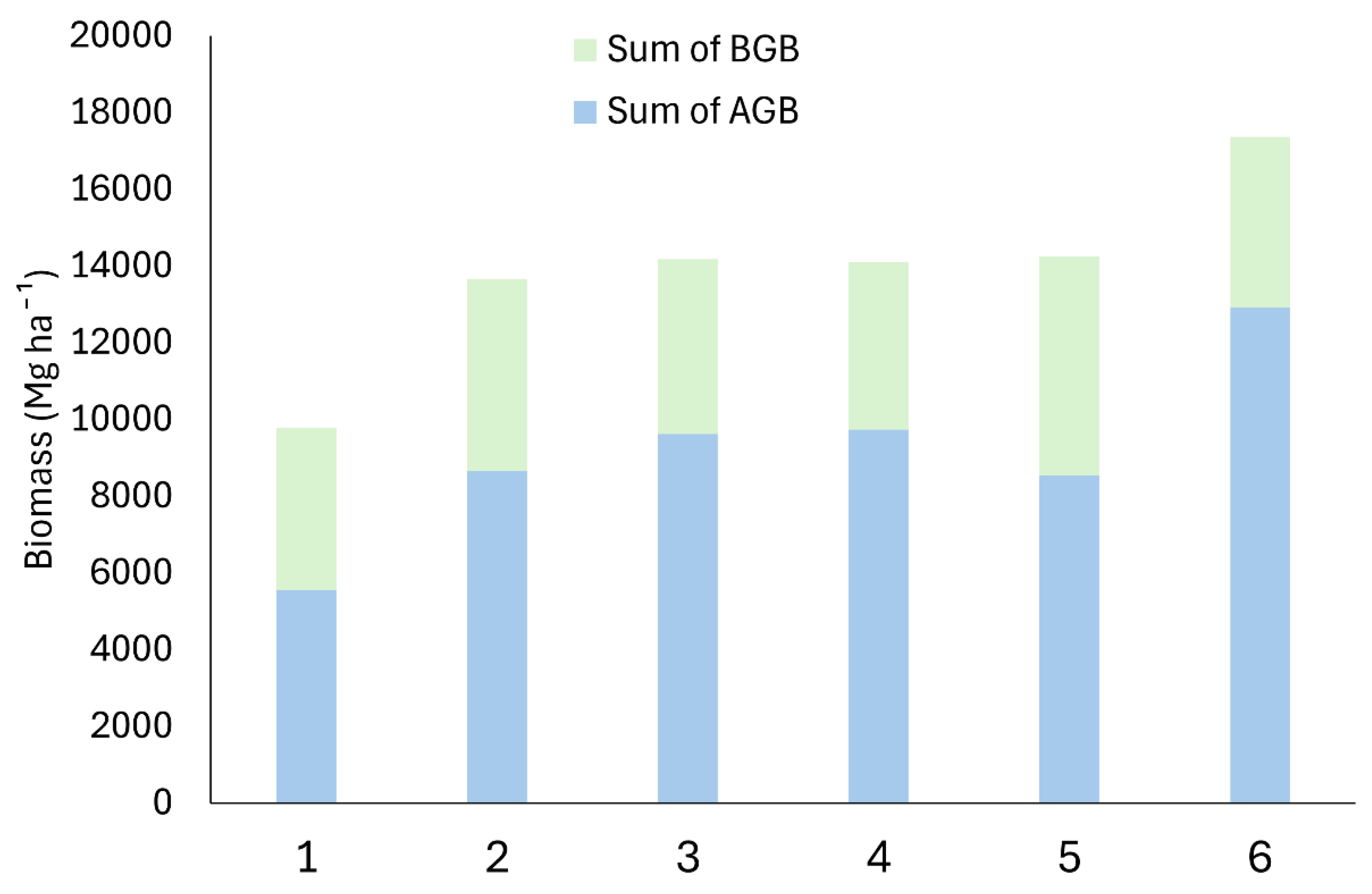
Figure 4.
Content of Aluminium (mg/kg), organic carbon (%) and Nitrogen (%) in topsoil (A) and subsoil (B) samples. Values are estimated marginal means and 95% confidence intervals.
Figure 4.
Content of Aluminium (mg/kg), organic carbon (%) and Nitrogen (%) in topsoil (A) and subsoil (B) samples. Values are estimated marginal means and 95% confidence intervals.
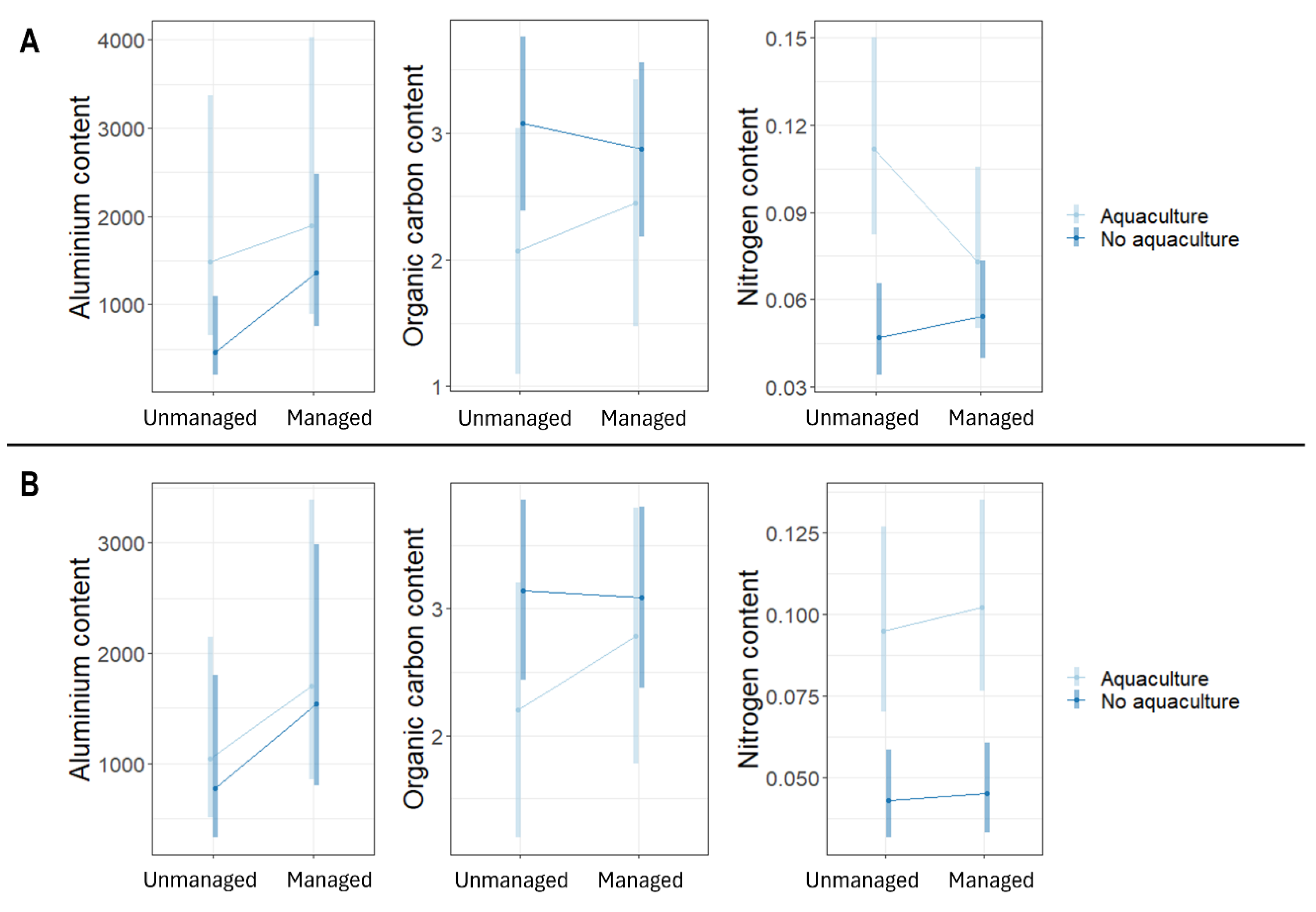
Figure 5.
Estimated marginal means and 95% confidence intervals of crab abundance (A) and richness (B) in 5m2 plots in mangroves based on generalized linear mixed models.
Figure 5.
Estimated marginal means and 95% confidence intervals of crab abundance (A) and richness (B) in 5m2 plots in mangroves based on generalized linear mixed models.
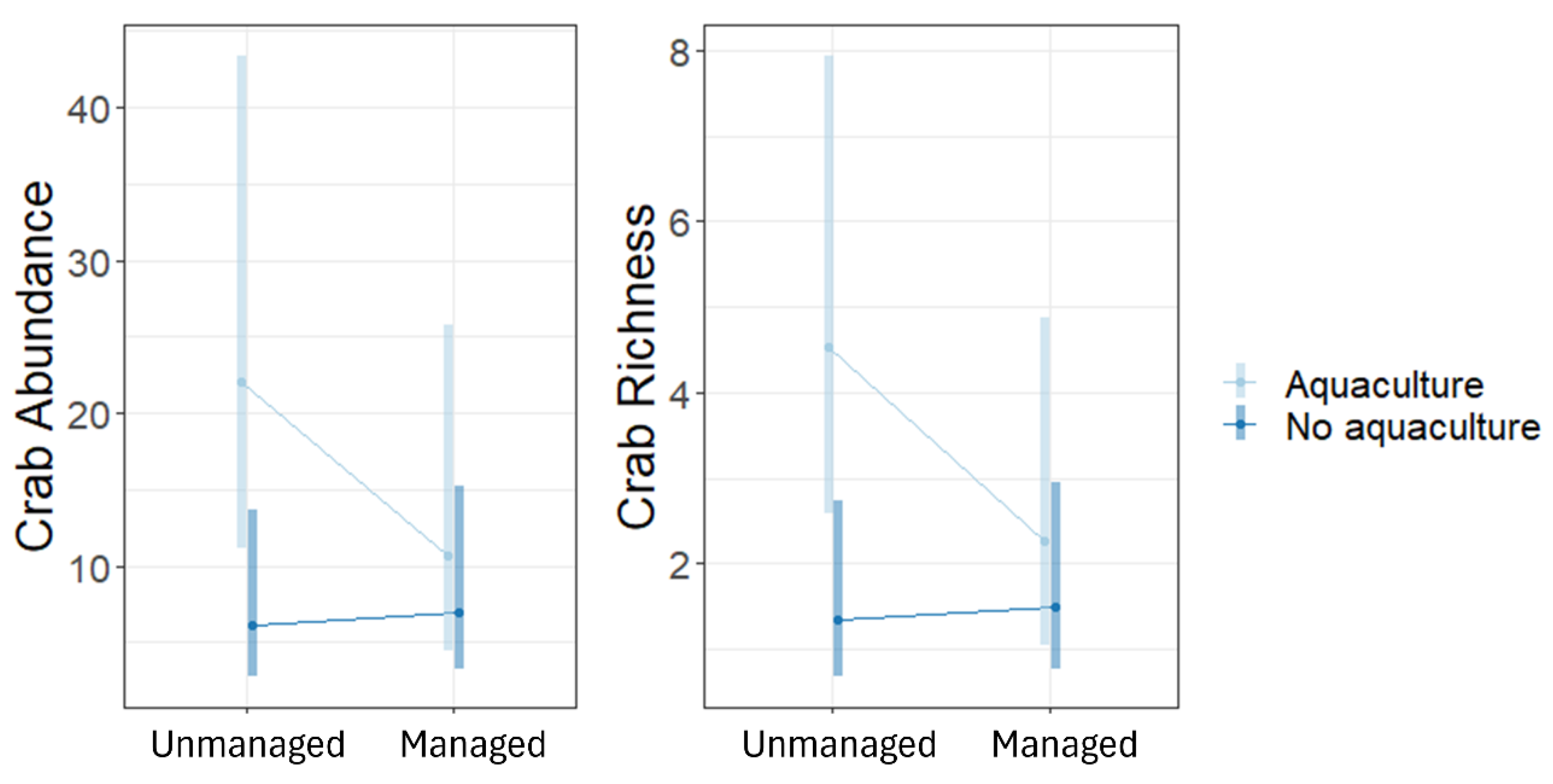
Figure 6.
Correlation matrix of the variables taken into consideration for the investigation of mangrove ecosystems in Bali and Lombok. Only significant (p<0.05) correlation coefficients are shown.
Figure 6.
Correlation matrix of the variables taken into consideration for the investigation of mangrove ecosystems in Bali and Lombok. Only significant (p<0.05) correlation coefficients are shown.
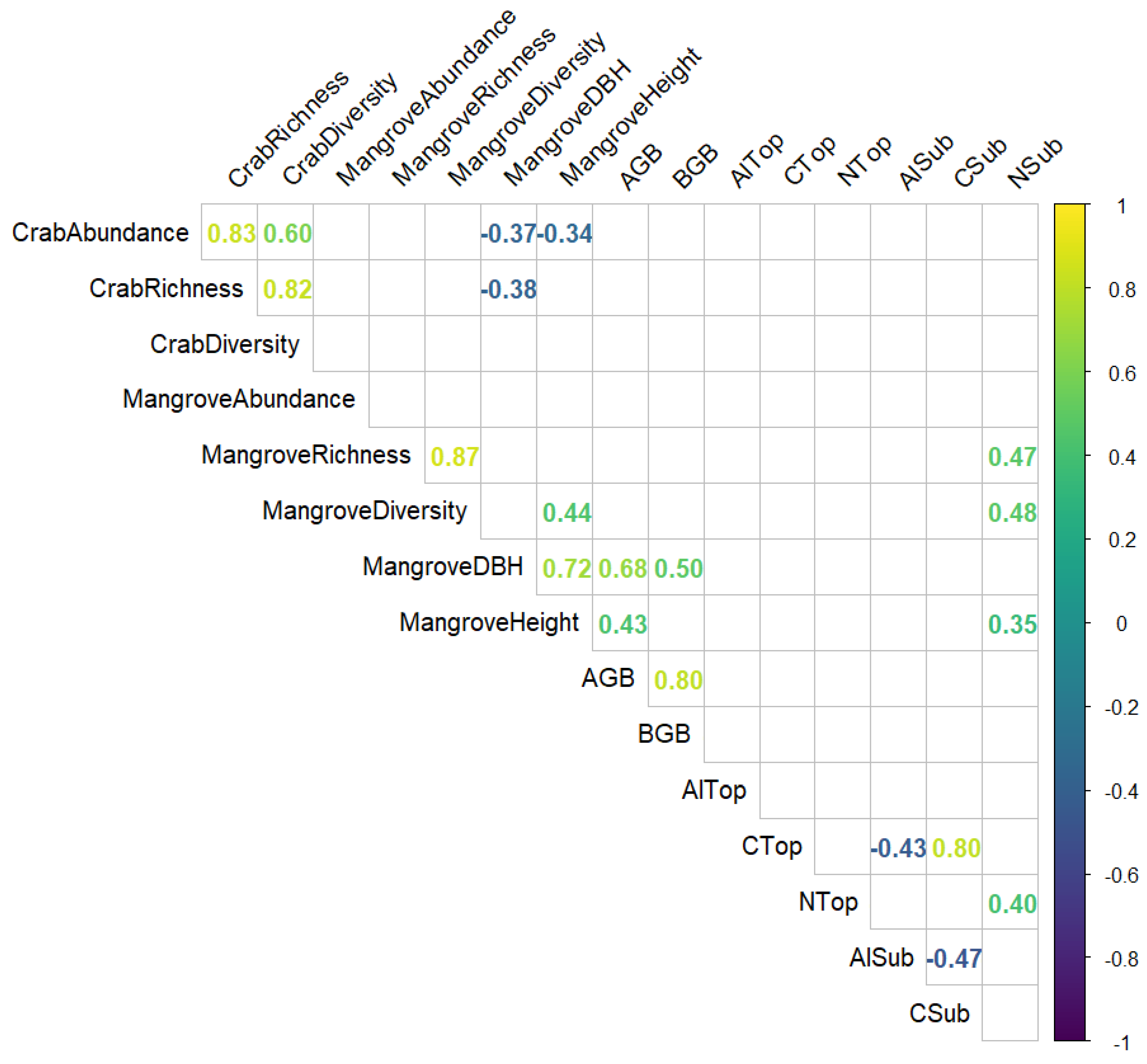
Figure 7.
Fishing pressure events recorded during the observation times.
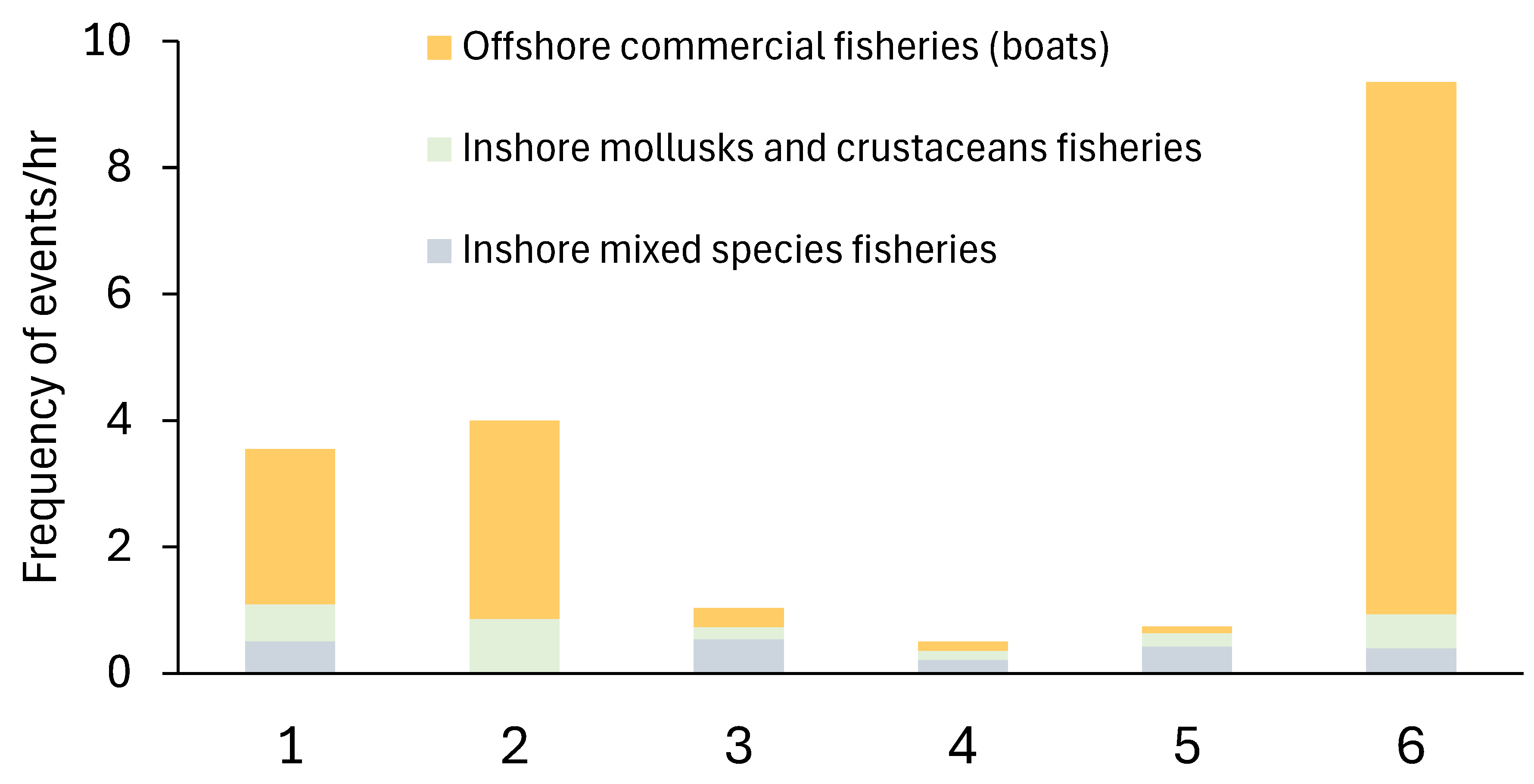
Table 1.
Description and designation of the study areas.
| Site | Designation | Use |
|---|---|---|
| Site 1 | Unmanaged | Current aquaculture site which has been unmanaged for more than 10 years, with local communities extracting resources (shrimps and fishes) |
| Site 2 | Managed | Current aquaculture site which has been managed and restored over the last 20 years, and still used as an aquaculture farm for mangrove mud crabs, shrimp, and oysters. |
| Site 3 | Managed | Under restoration since 2021 from the NGO SORCE. This site has been subjected to deforestation of the mangrohasto create a road through the landward area of the mangrove forest. |
| Site 4 | Unmanaged | Site selected for future restoration by SORCE but was unmanaged at the time of the surveys. This site has seen more natural recruitment in these areas than that of Site 3, with many young Ceriops species in the areas that could be classed as mudflats |
| Site 5 | Managed | A tourist attraction with wooden walkways built within the mangrove forest, however this has been closed off to the public for several years and needs permission to be entered. There is evidence that restoration was conducted recently and is on-going. |
| Site 6 | Unmanaged | Used by local fisherman to moor boats and fish along the mudflats. This site is unmanaged and has been historically deforested for economic development |
Table 3.
Crab assemblages in the six sites.
| Variablea | Site 1 | Site 2 | Site 3 | Site 4 | Site 5 | Site 6 | Total |
|---|---|---|---|---|---|---|---|
| Richness | 14 | 8 | 8 | 3 | 5 | 6 | 18 |
| N individuals | 123 | 65 | 26 | 46 | 63 | 22 | 345 |
| Diversity | 1.66 | 1.80 | 1.45 | 0.79 | 1.18 | 1.52 | 1.96 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



