Submitted:
04 June 2024
Posted:
05 June 2024
You are already at the latest version
Abstract
Beneficial microbes are crucial for improving crop adaptation and growth under vari-ous stresses. They enhance nutrient uptake, improve plant immune responses, and help plants tolerate stresses like drought, salinity, and heat. The yield potential of any crop is significantly influenced by its associated microbiomes and their potential to im-prove growth under different stressful environments. Therefore, it’s crucial and excit-ing to understand the mechanisms of plant-microbe interactions. Maize (Zea mays L.) is one of the primary staple foods worldwide, in addition to wheat and rice. Maize is also an industrial crop globally, contributing 83% of its production for use in feed, starch, and biofuel industries. Maize requires significant nitrogen fertilization to achieve opti-mal growth and yield. Maize plants are highly susceptible to heat, salinity, and drought stresses and require innovative methods to mitigate the harmful effects of environmental stresses and reduce the use of chemical fertilizers. This review summarizes our current understanding of the beneficial interactions between maize plants and specific microbes. These beneficial microbes improve plant resilience to stress and in-crease productivity. For example, they regulate electron transport, downregulate cata-ase, and upregulate antioxidants. We also review the roles of plant growth promoting rhizobacteria (PGPR) enhancing stress tolerance in maize. Additionally, we explore the application of these microbes in maize production and identify major knowledge gaps that need to be addressed to utilize the potential of beneficial microbes fully.
Keywords:
abiotic and biotic stress
; beneficial microbes
; stress tolerance
; maize
; plant growth-promoting rhizobacteria (PGPR)
; AMF
; Mycorrhiza
1. Introduction
Maize (Zea mays L.), alongside wheat and rice, stands as one of the primary staple foods worldwide, boasting a global production of 1147.7 million metric tons in 2020 [1]. Maize has risen to prominence as an industrial crop on a global scale, with 83% of its production allocated for use in feed, starch, and biofuel industries. Among the 125 developing countries, approximately 75 consider maize a staple crop, contributing to 70% of the world's maize production originating from these nations [2]. Maize is a nitrogen-hungry crop requiring a significant amount of nitrogen fertilization to achieve optimal growth and yield, particularly during its vegetative and early reproductive stages, which are more sensitive to nitrogen requirements. Inadequate nitrogen supply during these phases limits plant development, reduces photosynthetic efficiency, and ultimately decreases the grain yield [3,4]. Moreover, maize crops are extremely susceptible to heat, salinity, and drought stresses. Global maize yield is projected to decline by 20-40% under a 2°C warming scenario and by 40-60% under a 4°C warming scenario [5]. The variability in global maize production between 1980 and 2013 can be attributed to heat stress and drought [5]. Salinity stress further exacerbates challenges by decreasing the germination rate in maize. It causes osmotic stress, inhibiting water uptake by seeds delaying germination [6]. In salinity stress, the accumulation of Na+ competes with K+, leading to inhibition of protein synthesis [7]. This stress causes ionic toxicity, reactive oxygen species (ROS) generation, and osmotic stress [8]. In addition to abiotic stresses, between 6% to 19% of maize production worldwide is lost annually due to damage caused by insects and other herbivores. The primary pests of maize are Leaf aphids (Rhopalosiphum maidis), thrips (Frankliniella williamsi) [9], fall armyworm (Spodoptera frugiperda), black cutworms (Agrotis ípsilon), cotton bollworm (Helicoverpa armígera), corn earworm (Helicoverpa zea) [10] stalk borer (Elasmopalpus lignosellus) and armyworm (Spodoptera spp.).
Plant microbiomes are microorganisms that live in and around plant, forming a complex microbial ecosystem and can play a significant role in plant growth and development. These microbiomes includes bacteria, fungi, nematodes, archaea and viruses that inhabits at different parts of plants. These parts include rhizosphere (soil surrounding roots), phyllosphere (above-ground parts like leaves and stems), endosphere (internal tissue) and spermosphere (seed surfaces) [11,12]. Beneficial plant-microbial interactions significantly affect plant’s growth and development and mitigate environmental stresses [13,14]. Plants are intimately associated with microbes for their growth and survival; they play a significant role in plant nutrient availability and uptake and plant stress tolerance [15]. Studies reported the presence of plant growth-promoting rhizobacteria from the native plant Ceanothus velutinus, which contains several rhizobacteria possessing plant growth-promoting traits such as the production of IAA, siderophore, protease, catalase, ability to fix nitrogen, and phosphate solubilization [16]. Moreover, inoculating native soil from the Ceanothus velutinus to the propagation mix enhanced the cutting propagation, and IAA-producing isolates from the rhizosphere promote Arabidopsis growth [17]. Thus, exploring plant microbiomes to improve the maize yield and help it withstand different biotic and abiotic stresses is crucial. This review focuses on the beneficial plant-microbe interactions in maize to enhance yield and mitigate environmental stresses.
2. Abiotic Stresses and Their Impact on Crop Productivity
To meet high consumptive demand of maize they are often grown in arid locations where maize may experience drought-related stress. Maize life cycle has various distinct growth stages, including seedling emergence and development, vegetative growth, flowering and pollination, grain filling, and maturation. Drought and high temperatures can negatively impact maize crops throughout these growth stages, with the most significant effects at vegetative and during grain filling stage and when plants reach the 8th leaf stage [18]. In regions where water is scarce during the growing season, maize production may decline by up to 15% [19]. In China's key maize-producing areas, approximately 60% of crops face water and heat stress, leading to a 30% reduction in annual yield [20]. However, the most alarming prospect is the future. With the ongoing climate change and global shifting weather patterns, water, and heat stress are projected to diminish the global maize supply by 15-20%, annually [18]. Elevated temperatures exceeding 35℃ can impede maize crop’s reproductive and vegetative growth, from seed germination to grain filling, the final stage [21]. Concurrently, when maize faces water and heat stress during its reproductive phases, it becomes even more vulnerable [22]. The impact of drought stress on maize includes reduced leaf area, low water-use efficiency, lesser nutrient uptake, decreased photosynthetic efficiency, and reduced biomass accumulation and lower productivity. Studies have shown that water stress during vegetative growth can diminish growth rate, decrease root system development, prolong the vegetative phase, and affect CO2 distribution. A brief period of water scarcity can lead to a 28-32% reduction in dry weight during vegetative growth and 66-93% during tasseling/ear formation [23]. Extended drought stress before flowering can decrease leaf size and internodal distance, delaying silk emergence and tasseling and resulting in a 15-25% overall yield decrease [24]. Additionally, even a few days of drought stress during pollination/fertilization can lead to abnormal embryo formation and fewer kernels per plant. Drought stress before and after pollination is associated with a significant decline in kernel set [24]. The primary photosynthetic activity of maize plants occurs in their five- or six-ear leaves, mainly contributing to plant biomass. However, drought stress can diminish the photosynthetic rate by reducing ear leaf size, and slowing crop growth [25].
Higher temperatures at reproductive stages, such as tasseling, pollination, and grain filling, can lower maize grain quality. A study by Izaurralde et al.,[26] suggests that increasing the mean seasonal temperature by 1°C can reduce the maize economic yield by 3-13%. The study by Hussain et al., [20] on two maize hybrids, Xida 319 and Xida 889, subjected to heat stress, observed reduced plant height, lowered biomass accumulation, and decreased yield. Increased heat stress reduces the efficiency of light utilization in maize plants leading to chlorophyll degradation. Additionally, the increased temperature during the anthesis stage of maize cultivation results in reduced growth [27]. Similarly, exposure of maize to heat stress during the 12-leaf stage reduces pollen production, germination rate, zeatin content, salicylic acid content, and tassel size [28].
Salt stress is among several abiotic stresses affecting maize growth and yield. Increased salt concentration reduces plant height and biomass due to high osmotic stress and ion toxicity [29]. This reduction in growth is followed by decreased stomatal conductance and photosynthetic pigments, disturbance in cytosolic enzyme activity, and impairment of carbon fixation enzymes [30,31]. In a research by Kaya et al., [32] applying a salt concentration of 100 mM NaCl during the reproductive phase of maize reduced kernel weight and yield by 8% and 25%, respectively. Similarly, a study by Katerji et al., [33] showed an 11.3% reduction in maize grain yield in clay soil subjected to salinity stress. The increase in salt concentration interferes with the maize plant’s ability to absorb nitrate ions due to antagonistic action between chloride and nitrate ions [34]. The effect of abiotic stress has been described in Figure 1B)
3. Biotic Stress and Crop Production
Abiotic stress is the major obstacle in attaining potential yield for maize production worldwide. In addition to abiotic stress biotic stress also threaten maize cultivation, often leading to substantial yield losses [35]. Diseases, insects and pests are the primary factors responsible for these losses, with pathogens such as fungi, bacteria, and viruses eliciting syndromes like ear/stalk/kernel rot, rough dwarf/wilt disease, and northern leaf blight/maize mosaic [36], which are the major diseases that reduce maize yield. The simultaneous occurrence of abiotic and biotic stresses exacerbates the situation, resulting in a remarkable reduction in global maize production. Studies indicate that over 50% reduction in yield occurs in major crops, including maize, due to abiotic stresses alone. In comparison, approximately 10% of maize productivity is lost annually to biotic stresses worldwide [37], and 22.5% of global maize loss is due to diseases and pests. The effect of abiotic stress has been described in Figure 1B.
4. Mechanism of Abiotic Stress Tolerance in Maize
Plants developed various mechanisms to cope with various abiotic stresses, which are described below for three major abiotic stresses in maize.
4.1. Drought Stress
To cope with drought stress, maize plants have evolved various mechanisms broadly categorized into escape, avoidance, and tolerance strategies [38]. Drought escape refers to shortening a crop's life cycle to avoid drought stress, which is particularly crucial during reproductive growth stages. Traits like days to sowing, flowering, and maturity are genetically heritable, allowing for phenological adjustments in response to water availability [38]. Developing early maturing cultivars aids in evading terminal drought stress [39]. However, this strategy may reduce yields, as crop duration directly correlates with yield [40]. Through selection, crops adjust their growth period based on available moisture, completing their life cycle before drought onset. The maize plant tries to complete the reproductive stage before the drought becomes more prevalent. Maize, highly susceptible to drought, benefits significantly from this escape mechanism [41].
Drought avoidance in maize is assessed by measuring tissue water status, typically indicated by turgor water potential under drought stress conditions. Avoidance involves maintaining plant water status by reducing transpiration rates or increasing water uptake [42]. Various physiological and morphological traits are essential selection criteria for drought avoidance in maize, including leaf rolling, leaf firing, canopy temperature, stomatal closure, leaf attributes, and root traits [43]. Stomata regulate transpiration and gaseous exchange, governing photosynthesis and respiration. Plants reduce water loss by closing their stomata, preserving water status, and enhancing drought avoidance [40]. Drought tolerance for the combination of heat and drought stress involves maintaining growth and development through cellular and biochemical adaptations. Along with sustaining average physical growth, drought tolerance is also associated with yield stability under water-stressed conditions, a complex process in which crops have developed various natural mechanisms to adapt and tolerate drought stress [44]. These adaptations include accumulating compatible osmolytes like proline, glycine betaine, soluble sugars, and various inorganic ions (K+, Na+, Ca2+, Mg2+, Cl–, and NO3–) to support plant water status via osmotic adjustment [44,45].
Additionally, the enzymatic and non-enzymatic antioxidant systems, including superoxide dismutase (SOD), catalase (CAT), peroxidase (POD), and ascorbate peroxidase (APX), are activated to mitigate oxidative damage. Growth regulators like abscisic acid (ABA) also play a role [20,25]. Moreover, transcription factors (TFs) are activated to regulate gene expression sensitive to drought and high temperatures, while stress proteins like heat shock proteins (HSP), late embryogenesis abundant (LEA) proteins, and aquaporins assist in water movement under stress [41].
4.2. Maintaining Ion Homeostasis in Salinity Stress
An imbalance in the cellular ion exchange process causes salinity stress in the plant. Due to this ionic imbalance, Na+ influx and K+ efflux through various ion transporters in the cell membrane [46]. The excess concentration of Na+ increases oxidative stress by enhancing ROS (reactive oxygen species) production [47]. Consequently, cellular membranes become disrupted, leading to a breakdown in cell homeostasis. During salinity stress, genes and transcription factors regulating ion transports are activated which, helps alleviate ion toxicity in cells. These include plasma membrane protein (PMP), high sodium affinity transporter (HKT), Salt overly sensitive (SOS) pathway, and Na+/H+ exchangers (NHXs) [48]. ZmCIPK24a and ZmCBL4 plus ZmCBL8 act as SOS2 and SOS3 in maize [49]. During salinity stress, SOS3 senses the changes in the cytoplasmic Ca2+ level, which activates SOS2. The SOS2-SOS3 complex phosphorylates ZmSOS1, activating SOS1 and increasing root-to-soil Na+ efflux, enhancing salt tolerance [50]. The study identified QTL for K+ content (qKC3), which encodes ZmHKT2, a K+ transporter localized in the xylem parenchyma [51]. ZmHKT2 reduces shoot K+ content by retrieving K+ from xylem vessels. The mutants lacking ZmHKT2 have higher shoot K+ content and salt tolerance [52]. Decreasing the activity of ZmHKT2 is one viable strategy for developing salt tolerant maize varieties.
4.3. Osmotic Adjustment
Osmotic adjustment involves creating a water gradient to enhance water influx, thereby maintaining turgor by reducing osmotic potential. This adjustment aids in preserving tissue water status by minimizing the detrimental effects of drought through the accumulation of solutes in cellular cytoplasm and vacuoles. By sustaining turgor potential and supporting physiological processes, osmotic adjustment protects against drought-induced damage [53]. Relative water content is a crucial indicator for estimating drought tolerance in plants with closed stomata and reduced CO2 accumulation resulting from decreased relative water content under drought stress [42]. The sustainable regulation of photosynthetic rate and turgor potential ensures the translocation of photosynthetic assimilates to developing kernels [54]. Osmoprotectants, including nitrogenous compounds like proline, polyols, polyamines, and glycine betaine, as well as hydroxy compounds like polyhydric alcohols, sucrose, and oligosaccharides, play crucial roles in osmotic adjustment [55]. These compounds protect cellular proteins and membranes against dehydration effects and help maintain cellular integrity [56]. Glycine betaine, for instance, acts as an important osmoprotectant, safeguarding plants against various stresses such as drought, salinity, cold, and heat by protecting the photosynthetic apparatus and stabilizing cellular proteins [57]. Proline, another osmoprotectant, helps maintain water status, protect cellular membranes, and prevent protein denaturation under osmotic stress [57,58]. Soluble sugars, accumulated in response to drought stress, serve multiple roles in plant metabolism and protection, including acting as substrates for biosynthesis processes and protecting cellular organelles through vitrification [59]. Polyols, such as sorbitol, glycerol, and mannitol, form hydration spheres around macromolecules, safeguarding them from dehydration [60]. These mechanisms collectively contribute to plants' ability to tolerate drought stress and maintain essential physiological processes.
Antioxidants and plant growth regulators
Antioxidants are molecules that protect plants by scavenging reactive oxygen species, thus preventing oxidative damage. They form a defense shield against oxidative stress. Antioxidants can be enzymatic or non-enzymatic. Enzymatic antioxidants include catalase (CAT), superoxide dismutase (SOD), glutathione reductase (GR), ascorbate peroxidase (APX), peroxidase, and polyphenol oxidase. Non-enzymatic antioxidants include α-tocopherol, ascorbic acid, β-carotene, glutathione, and cysteine [61]. These components are essential in neutralizing reactive oxygen species and preserving plant health under oxidative stress conditions. Plant hormones, also known as plant growth regulators or phytohormones, play vital roles in governing the growth and development of plants, acting as signaling molecules that trigger cellular differentiation and function locally or are transported to distant targets.
In response to drought stress, plants undergo various adaptations, including maintaining endogenous hormonal balance [62]. Different plant growth regulators confer drought tolerance, including auxins, cytokinins, abscisic acid (ABA), gibberellins, salicylic acid, brassinosteroids, methyl jasmonate, polyamines, ethylene, and zeatin. These hormones interact to regulate plant responses, specific growth stages, tissues, and environmental conditions. For instance, auxins are involved in drought stress responses, with interactions observed between ethylene, cytokinins, and auxins affecting their biosynthesis [63,64]. The concentration of indole acetic acid (IAA) decreases in maize leaves under drought stress, while the accumulation of ABA increases, influencing hormonal balance. The IAA accumulation increases under moderate stress (13.4%) while decreases under severe drought stress (63.2%) in maize [25]. Salicylic acid helps maintain photosynthesis by retaining higher chlorophyll content under drought stress, contributing to drought tolerance [65]. ABA and ethylene regulate stomatal conductance, grain number, grain filling rate, and plant apex growth antagonistically, with cytokinin enhancing growth and development. ABA plays a crucial role as a stress hormone, modulating growth, development, and stress responses through a signaling pathway involving various components highly responsive to ABA. Average water availability does not induce ABA accumulation, and extremely severe drought reduces ABA accumulation due to the cessation of ABA precursors [66]. The antioxidant defense system detoxifies ROS and maintain redox homeostasis [67]. Overall plant hormonal balance is critical for various growth and development processes, with interactions between hormones playing a crucial role in plant adaptation to drought stress.
4.5. Polyamines and Their Roles in Biotic Stress in Maize
The detrimental effects of biotic stress on significant crop plants, maize, are not just a concern, but a pressing issue. The production of highly carcinogenic aflatoxins by Aspergillus flavus alone has led to a staggering loss of USD 686.6 million in maize cultivation in the U.S These statistics are not mere statistics but a stark reminder of the economic havoc wreaked by biotic stress. Other climate-dependent pathogens, such as Fusarium spp. and Ustilago maydis, further compound the issue [68,69].
Polyamines (PA) has emerged as a beacon of hope in reducing biotic stress caused by various pathogens in maize. PAs play a crucial role in the production of H2O2, acting as both a defensive tool and a signaling molecule in response to biotic stress [69]. For example, spermine (Spm), a form of PA, functions as a signaling molecule in pathogen defense and plays a critical role in resistance against viral infections [70]. In the case of Ustilago maydis, a dimorphic host-specific fungus, it induces 'huitlacoche' or common smut in maize plants. The accumulation of H2O2 derived from polyamine oxidase plays a significant role in tumor formation caused by U. maydis in maize plants. The maize polyamine oxidases (zMPAOs) transcription factor was found to be downregulated in tumors. The symptoms of the disease were observed to reduce upon application of 1,8-diamino octane (1,8-DO), a potent polyamine oxidase inhibitor[69].
5. Beneficial Plant-Microbe Interactions in Maize
5.1. Arbuscular Mycorrhizal Symbiosis
Maize forms symbiotic associations with Arbuscular mycorrhizal (AM) fungi. This partnership, established via the mycorrhizal and root pathway, allows plants to efficiently uptake nutrients from the soil. In this symbiosis, the fungi and plants engage in a mutual exchange where the fungi provide mineral nutrients while the plant supplies carbon (C). Maize roots, in addition to beneficial relationships with microbes such as mycorrhizal fungi, play a crucial role in uptake of nutrients like phosphorus (P) and nitrogen (N). Maize root residues, a significant byproduct of this symbiosis, provide N for other plants in crop rotation, thereby improving agricultural productivity [71,72]. This exchange occurs via arbuscules inside root cortex cells, where AM fungi acquire 4–20% of the total photosynthetic carbon fixed by the plant through symbiotic relationships. The AM fungal hyphae then utilize this carbon to generate specialized exudates, which attract and establish a hyphosphere microbiome. This microbiome plays a crucial role in compensating for the fungi's incapacity to utilize organic nutrients directly. By secreting enzymes and fostering the mineralization of organic nutrient sources, the hyphosphere microbiome significantly increases nitrogen and phosphorus availability. This collaborative functionality within the holobiont substantially enhances nutrient accessibility for all interacting organisms, including plants, AM fungi, and hyphosphere bacteria (Figure 1C and D).
In maize, the colonization of maize roots by AM fungi begins early in the plant development phase, which peaks at the vegetative growth stages. Maize roots produce strigolactones (5-deoxy-strigol and Sorghumol), essential for establishing AM symbiosis [73,74]. These compounds act as chemoattractants and guide the fungal hyphae toward the root system [75]. Upon contact with strigolactones, AM fungi initiate signaling cascades that activate genes such as SYM and RAM1 involved in colonization. Like other plants such as carrots, maize roots form pre-penetration apparatuses (PPAs) at the root surface to facilitate penetration of fungal hyphae into root tissue. Upon penetration, the fungal hyphae initiate a series of molecular events, such as the production of chitin and lipochitooligosaccharides for recognition and signaling between plant and fungus [76]. The signal transduction also leads to activation of transcription factors and arbuscule formation. Fungal-derived proteins, such as Small Secreted Effector Proteins (SSEPs), are transported into the plant cell and are believed to play roles in arbuscule formation and function [77]. The nutrient exchange between fungus and maize plants occurs within the arbuscules. In addition to arbuscules, vesicles are formed within root cells, which act as storage structures for lipids, glycogen, and other metabolites.
Once the symbiosis between AM fungi and maize plants is established, AM fungi can increase the root volume, increasing the surface area for water absorption. D-myo-inositol-3-phosphate synthase (IPS) and the 14-3-3-like protein GF14 (14-3GF) are pivotal in facilitating signal communication between maize and AMF during drought stress. Co-expression of these two genes has been shown to enhance maize drought tolerance significantly [78]. Similarly, the AM fungi infection upregulates the expression level of NPF4.5 homologs, indicating higher nitrate uptake during symbiosis [78]. The ammonium transporter ZmAMT3;1 expressed in cortical cells of maize during AM fungi infection absorbs 68-70% of the transported nitrogen AM fungi to maize plants [79].
5.2. Nitrogen-Fixing Symbiosis with Rhizobia
Rhizobia, the widely distributed Gram-negative bacteria in soil, can enhance maize cultivation. Despite being primarily associated with legumes, these beneficial bacteria can promote growth and yield in maize through various mechanisms. While their efficiency with maize is generally lower than with legumes, the potential for improvement is promising [80].
The inoculation of the Azospirillum strain in maize roots increased the GA3 levels, thereby boosting root growth [81]. Similarly, the strains of Rhizobium (such as R. etli bv. Phaseoli and R. leguminosarum bv. trifolii) and Sinorhizobium sp. Have shown promising results in enhancing growth, increasing plant height, and improving grain yield in maize [82]. Numerous studies have reported on the nitrogen-fixing ability of Herbaspirillum seopedicae and Azospirillum spp. in maize. A study on two maize genotypes, Morgan 318 and Dekalb 4D-70, demonstrated a significant increase in grain yield and higher N accumulation with the inoculation of a mixture of Azospirillum spp. strains, a result comparable to the application of 100 Kg N ha-[83]. Another study unveiled the identification of a nitrogen-fixing association with the native variety of maize grown in nitrogen-depleted soils in Mexico. The microoxic environment for better nitrogen fixation is created by the mucilage tube surrounding the roots, which had a high abundance of proteobacteria. [84]. These symbiotic relationships are crucial in enhancing plant growth, higher nutrient acquisition, and crop yield, offering a hopeful outlook for the future of maize cultivation.
5.3. Agricultural Application of Stress-Tolerant Microbes
The use of stress-tolerant microbes shows a significant increase in the yield of maize plants. Maize plants inoculated with Piriformospora indica, an endophytic fungus grown under drought stress conditions, resulted in increased leaf area, SPAD value, higher root fresh and dry weight, and upregulation of antioxidants catalase and superoxide dismutase. Upregulating drought-related genes DREB2A, CBL1, ANAC072, and RD29A increased resistance to drought stress [85]. The Bacillus spp. PM31 also improved maize growth under salinity stress [86]. The microbes can be applied to enhance plant yield and improve soil health. Stress-tolerant microbes can replace 20-40% of chemical fertilizers while alleviating drought stress impact. Integrating stress-tolerant bacteria with other beneficial microbes, such as AM fungi, can increase stress tolerance in maize and other plants, offering more significant agricultural benefits. These microbes can be integrated into agronomic practices through various application strategies that contribute to sustainable agriculture (Table 1).
5.4. Microbe Mediated Induced Systemic Resistance (ISR) in Maize
Systemic Acquired Resistance (SAR) and Induced Systemic Resistance (ISR) are different mechanisms by which plants can develop systemic resistance against pathogens and diseases. SAR is a plant defense mechanism that protects plants against a broad spectrum of pathogens following an initial infection. SAR is induced by recognizing pathogen-associated molecular patterns (PAMPs) or effector molecules released by the pathogen [87]. ISR is a plant defense mechanism in which exposure to certain beneficial microorganisms, pathogens, or chemical compounds primes the plant's immune system to enhance its resistance against subsequent pathogen attacks. Unlike SAR, which is induced by direct pathogen infection, ISR is triggered by beneficial microbes or certain chemical compounds in the plant's environment [88].
ISR is a complex process that involves the activation of various defense responses within the plant, including the production of antimicrobial compounds, reinforcement of cell walls, and the activation of defense-related genes. ISR is triggered by non-pathogenic rhizobacteria, which colonizes the rhizosphere. The microbes prime the plant’s innate immune system, enhancing its defense response against subsequent pathogen and insect attacks [89]. Specific microorganisms, such as beneficial rhizobacteria, B. velezensis SQR9, and the fungus Trichoderma harzianum, play crucial roles in inducing ISR in maize against pathogens. B. velezensis SQR9 colonizes maize roots and activates defense signaling pathways. This colonization leads to the enrichment of phenylpropanoid biosynthesis, amino acid metabolism, and plant-pathogen interaction pathways in maize roots. The calcium signaling pathway is pivotal in SQR9-induced ISR, as inhibiting calcium signaling weakens the induced resistance [51]. Similarly, Trichoderma harzianum triggers ISR in maize against Curvularia leaf spot by releasing cellulases and cellobiose from roots. The cellobiose released from T. harzianum-colonized roots prompts the expression of defense-related genes (Opr7, Pr4, Aoc1, Erf1) in maize, thereby enhancing ISR against the pathogen [90]. ISR in maize involves jasmonic acid and ethylene signaling pathways mediated by the NPR1 protein.

Table 1.
The Plant Growth Promoting Rhizobacteria (PGPR) enhancing stress tolerance in Maize.
| Host associated microbial strains | Effect/Mechanism of Stress tolerance | References |
| Microbial mediated beneficial drought stress tolerance | ||
|
Rhizobium (R. etli bv. Phaseoli, R. leguminosarum bv. Trifolii, Sinorhizobium sp |
Enhanced growth, increased plant height, improved grain yield |
[82] |
|
Herbaspirillum seopedicae Azospirillum sp |
Increased grain yield Higher N accumulation |
[83] |
| Piriformospora indica | Increased leaf area and SPAD value Increased root fresh and dry weight Decreased Malondialdehyde (MDA) accumulation Upregulation of antioxidants and drought related genes |
[85] |
| Pseudomonas putida | Forms viable biofilms around roots Increase soil holding capacity Improve soil structure |
[91] |
|
Pseudomonas aeruginosa Alcaligenes faecalis Proteus peneri |
Increase soil moisture content Enhances plant growth traits such as leaf area, shoot length, root length Downregulation of catalase, ascorbate peroxidase and glutathione peroxidase |
[92] |
|
Klebsiella variicola Pseudomonas fluorescens Raoultella planticola |
Increased levels of betaine, glycine and choline Improved plant growth |
[93] |
|
Burkholderia sp. Mitsuaria sp. |
Increased proline and phytohormone accumulation Higher antioxidant activity Decreased MDA content |
[94] |
| Megathyrsus maximus | Increased proline accumulation Decrease in MDA content Reduced glutathione reductase activity |
[95] |
|
Azospirillum brasilense Pseudomonas putida Sphingomonas |
Symcoms containing these microbes increases shoot dry weight, root dry weight and plant height | [96] |
| Azospirillum lipoferum | Increased proline, soluble sugar and amino acids accumulation Enhances shoot and root weight, root length |
[97,98] |
| Bacillus sp. | Increased proline accumulation Reduction in electrolyte leakage Decreased activity of antioxidants |
[99] |
|
Burkholderia phytofirmans Strain PsJN Enterobacter sp. FD17 |
Increased root and shoot biomass Higher chlorophyll content Increased leaf area and photosynthetic rate |
[100] |
| Rhizophagus irregularis | Increased hydraulic conductivity and water permeability coefficient Increased phosphorylation of Plasma membrane intrinsic proteins (PIPs) Increased photosynthetic activity |
[101] |
| B pumilus | Increased relative water content and osmotic potential Higher photosynthetic activity Increased ABA production |
[102] |
|
Azospirillum brasilense SP-7 Herbaspirillum seropedicae Z-152 |
Decreased expression of ZmVP14 | [103] |
| Microbial mediated beneficial saline stress tolerance | ||
| Bacillus sp. PM31 | Improved maize growth under salinity stress | [86] |
| Co-inoculation of Rhizophagus intraradices Massilia sp. RK4 |
Increased nutrient uptake Increased AMF root colonization Decreased leaf proline levels |
[104] |
|
Rhizobium sp. Pseudomonas sp. |
Enhanced proline production Decrease in electrolyte leakage Reduction in osmotic potential Selective K ions uptake |
[105] |
|
Pseudomonas fluorescens, P. syringae, P. chlororaphis Enterobacter aerogenes |
ACC-deaminase for increasing plant height, biomass, and cob yield Higher grain mass and straw yield Increased P and K uptake Higher K+/Na+ ratio |
[106] |
| Glomus mosseae | Enhanced soluble sugars accumulation Increased total organic acids, acetic acid, malic acid, oxalic acid, fumaric acid and citric acid accumulation Increased upregulation of osmoregulation process |
[107] |
| B. amyloliquefaciens SQR9 | Increased chlorophyll content Enhanced soluble sugar content Decreased level of Na+ Upregulation of RBCS, RBCL, H+-PPase, HKT1, NHX1, NHX2 and NHX3 |
[108] |
| Kocuria rhizophila Y1 | Increased photosynthetic capacity and relative water content Increased antioxidant levels Decreased level of Na+ |
[109] |
| Azotobacter chroococcum | Increased K+/Na+ ratio Higher chlorophyll content Increased proline concentration |
[91] |
| Microbial mediated beneficial heat stress tolerance | ||
|
Bacillus sp. AH-08, AH-67, AH-16 Pseudomonas sp. SH-29 |
Upregulation of heat shock proteins (HSP) Increased total chlorophyll, catalase, and peroxidase Enhances plant height, leaf area, root & shoot fresh and dry weight Decreased concentration of MDA |
[110] |
|
Rhizophagus intraradices Funneliformis mosseae F. geosporum |
Increased quantum efficiency of PSII Higher photosynthetic rate Increased plant height, leaf width and cob number |
[111] |
| Glomus etunicatum | Increased water content and leaf water potential Increased photosynthetic activity Higher stomatal conductance |
[112] |
| Glomus sp. | Regulation of electron transport through PSII Increased plant height and leaf width |
[113] |
6. Challenges and Future Perspectives
Abiotic and biotic stresses significantly influence the growth and development of maize plants. Salt stress disrupts water uptake and nutrient acquisition, while drought stress hinders photosynthetic activity, decreasing in maize yield. Despite their heat tolerance, prolonged exposure to temperatures exceeding 35°C is detrimental to crop growth and development, and exceeding 40°C during flowering and grain-filling season will reduce grain productivity.
Despite the known benefits of plant-microbe interactions such as arbuscular mycorrhizal (AM) fungi, rhizobia, as well as bacterial and fungal endophytes, there is still much to learn about the diversity of beneficial microbes present in maize rhizosphere and their specific functions. Understanding which microbes are most helpful under different growing conditions and soil types is crucial for optimizing microbial inoculants. The interactions between introduced beneficial microbes and native soil microbiota are complex and poorly understood. Competition, cooperation, and antagonistic interactions among microbes can influence their effectiveness in promoting plant growth. More research is needed to assess the long-term effects of microbial inoculation on soil health, microbial community dynamics, and crop productivity. We are responsible for developing sustainable management practices that integrate microbial interactions into existing agricultural systems.
7. Conclusions
The positive interactions between maize plants and beneficial microbes offer a promising solution for enhancing plant growth and nutrient absorption under challenging conditions. These interactions not only have the potential to bolster the environmental resilience of maize agriculture but also to promote sustainability. using microbes for defense against various stresses can significantly increase crop yield and productivity, leading to economic benefits for farmers and potentially reducing the need for chemical inputs, thus benefiting the environment.
Author Contributions
AK and SB conceived the concept. SB wrote the original draft. AK, AD, and SP edited and reviewed the article.
Funding
This research received no external funding. This work is the product of final assignment in Graduate level course in Plant Microbe Interaction.
Acknowledgments
In this section, you can acknowledge any support given which is not covered by the author contribution or funding sections. This may include administrative and technical support, or donations in kind (e.g., materials used for experiments).
Conflicts of Interest
“The authors declare no conflicts of interest.
References
- Chávez-Arias, C.C.; Ligarreto-Moreno, G.A.; Ramírez-Godoy, A.; Restrepo-Díaz, H. Maize Responses Challenged by Drought, Elevated Daytime Temperature and Arthropod Herbivory Stresses: A Physiological, Biochemical and Molecular View. Front Plant Sci 2021, 12, 702841. [Google Scholar] [CrossRef] [PubMed]
- Nyirenda, H.; Mwangomba, W.; Nyirenda, E.M. Delving into Possible Missing Links for Attainment of Food Security in Central Malawi: Farmers’ Perceptions and Long Term Dynamics in Maize (Zea Mays L.) Production. Heliyon 2021, 7, e07130. [Google Scholar] [CrossRef] [PubMed]
- Asibi, A.E.; Chai, Q.; A. Coulter, J. Mechanisms of Nitrogen Use in Maize. Agronomy 2019, 9. [CrossRef]
- Gheith, E.M.S.; El-Badry, O.Z.; Lamlom, S.F.; Ali, H.M.; Siddiqui, M.H.; Ghareeb, R.Y.; El-Sheikh, M.H.; Jebril, J.; Abdelsalam, N.R.; Kandil, E.E. Maize (Zea Mays L.) Productivity and Nitrogen Use Efficiency in Response to Nitrogen Application Levels and Time. Front Plant Sci 2022, 13, 941343. [Google Scholar] [CrossRef]
- Maitah, M.; Malec, K.; Maitah, K. Influence of Precipitation and Temperature on Maize Production in the Czech Republic from 2002 to 2019. Sci Rep 2021, 11, 10467. [Google Scholar] [CrossRef] [PubMed]
- AbdElgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R.; Asard, H.; Abuelsoud, W. High Salinity Induces Different Oxidative Stress and Antioxidant Responses in Maize Seedlings Organs. Front Plant Sci 2016, 7, 276. [Google Scholar] [CrossRef] [PubMed]
- Schachtman, D.; Liu, W. Molecular Pieces to the Puzzle of the Interaction between Potassium and Sodium Uptake in Plants. Trends Plant Sci 1999, 4, 281–287. [Google Scholar] [CrossRef] [PubMed]
- Chawla, S.; Jain, S.; Jain, V. Salinity Induced Oxidative Stress and Antioxidant System in Salt-Tolerant and Salt-Sensitive Cultivars of Rice (Oryza Sativa L.). J. Plant Biochem. Biotechnol. 2011, 22. [Google Scholar] [CrossRef]
- Corona, A.O. Insect Pests of Maize: A Guide for Field Identification.; 1987.
- Widstrom, N.W. The Role of Insects and Other Plant Pests in Aflatoxin Contamination of Corn, Cotton, and Peanuts—A Review. Journal of Environmental Quality 1979, 8, 5–11. [Google Scholar] [CrossRef]
- Shelake, R.M.; Pramanik, D.; Kim, J.-Y. Exploration of Plant-Microbe Interactions for Sustainable Agriculture in CRISPR Era. Microorganisms 2019, 7, 269. [Google Scholar] [CrossRef]
- Rochefort, A.; Simonin, M.; Marais, C.; Guillerm-Erckelboudt, A.-Y.; Barret, M.; Sarniguet, A. Transmission of Seed and Soil Microbiota to Seedling. mSystems 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.; Khadka, R.; Doni, F.; Uphoff, N. Benefits to Plant Health and Productivity From Enhancing Plant Microbial Symbionts. Front. Plant Sci. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Vocciante, M.; Grifoni, M.; Fusini, D.; Petruzzelli, G.; Franchi, E. The Role of Plant Growth-Promoting Rhizobacteria (PGPR) in Mitigating Plant’s Environmental Stresses. Applied Sciences 2022, 12, 1231. [Google Scholar] [CrossRef]
- Zilber-Rosenberg, I.; Rosenberg, E. Role of Microorganisms in the Evolution of Animals and Plants: The Hologenome Theory of Evolution. FEMS Microbiol Rev 2008, 32, 723–735. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, J.; Singh, V.; Hewitt, K.; Kaundal, A. Exploration of the Rhizosphere Microbiome of Native Plant Ceanothus Velutinus – an Excellent Resource of Plant Growth-Promoting Bacteria. Front Plant Sci 2022, 13, 979069. [Google Scholar] [CrossRef] [PubMed]
- Ganesh, J.; Hewitt, K.; Devkota, A.R.; Wilson, T.; Kaundal, A. IAA-Producing Plant Growth Promoting Rhizobacteria from Ceanothus Velutinus Enhance Cutting Propagation Efficiency and Arabidopsis Biomass. Front. Plant Sci. 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-H.; Lee, B.-M. Effects of Climate Change and Drought Tolerance on Maize Growth. Plants (Basel) 2023, 12, 3548. [Google Scholar] [CrossRef] [PubMed]
- Ziyomo, C.; Bernardo, R. Drought Tolerance in Maize: Indirect Selection through Secondary Traits versus Genomewide Selection. Crop Science 2013, 53, 1269–1275. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive Effects of Drought and Heat Stresses on Morpho-Physiological Attributes, Yield, Nutrient Uptake and Oxidative Status in Maize Hybrids. Sci Rep 2019, 9, 3890. [Google Scholar] [CrossRef] [PubMed]
- Hatfield, J. Increased Temperatures Have Dramatic Effects on Growth and Grain Yield of Three Maize Hybrids. Agricultural & Environmental Letters 2016, 1, 1–5. [Google Scholar] [CrossRef]
- Boehlein, S.K.; Liu, P.; Webster, A.; Ribeiro, C.; Suzuki, M.; Wu, S.; Guan, J.-C.; Stewart, J.D.; Tracy, W.F.; Settles, A.M.; et al. Effects of Long-Term Exposure to Elevated Temperature on Zea Mays Endosperm Development during Grain Fill. The Plant Journal 2019, 99, 23–40. [Google Scholar] [CrossRef] [PubMed]
- Cakir, R. Effect of Water Stress at Different Development Stages on Vegetative and Reproductive Growth of Corn. Field Crops Research 2004, 89, 1–16. [Google Scholar] [CrossRef]
- Hütsch, B.W.; Faust, F.; Jung, S.; Schubert, S. Drought Stress during Maize Flowering May Cause Kernel Abortion by Inhibition of Plasma Membrane H+-ATPase Activity. Journal of Plant Nutrition and Soil Science 2024. [Google Scholar] [CrossRef]
- Aslam, M.; Maqbool, M.A.; Cengiz, R. Drought Stress in Maize (Zea Mays L.): Effects, Resistance Mechanism, Global Achievements and Biological Strategies for Improvement; 2015; ISBN 978-3-319-25442-5.
- Izaurralde, R.C.; Thomson, A.M.; Morgan, J.A.; Fay, P.A.; Polley, H.W.; Hatfield, J.L. Climate Impacts on Agriculture: Implications for Forage and Rangeland Production. Agronomy Journal 2011, 103, 371–381. [Google Scholar] [CrossRef]
- Gabaldón-Leal, C.; Webber, H.; Otegui, M.E.; Slafer, G.A.; Ordóñez, R.A.; Gaiser, T.; Lorite, I.J.; Ruiz-Ramos, M.; Ewert, F. Modelling the Impact of Heat Stress on Maize Yield Formation. Field Crops Research 2016, 198, 226–237. [Google Scholar] [CrossRef]
- Sun, J.; Wang, H.; Ren, H.; Zhao, B.; Zhang, J.; Ren, B.; Liu, P. Maize (Zea Mays L.) Responses to Heat Stress: Mechanisms That Disrupt the Development and Hormone Balance of Tassels and Pollen. Journal of Agronomy and Crop Science 2023, 209, 502–516. [Google Scholar] [CrossRef]
- Singh, V.; Krause, M.; Sandhu, D.; Sekhon, R.S.; Kaundal, A. Salinity Stress Tolerance Prediction for Biomass-Related Traits in Maize (Zea Mays L.) Using Genome-Wide Markers. The Plant Genome 2023, 16, e20385. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Hussain, M.; Wakeel, A.; Siddique, K.H.M. Salt Stress in Maize: Effects, Resistance Mechanisms, and Management. A Review. Agron. Sustain. Dev. 2015, 35, 461–481. [Google Scholar] [CrossRef]
- Gong, X.; Chao, L.; Zhou, M.; Hong, M.; Luo, L.; Wang, L.; Ying, W.; Jingwei, C.; Songjie, G.; Fashui, H. Oxidative Damages of Maize Seedlings Caused by Exposure to a Combination of Potassium Deficiency and Salt Stress. Plant Soil 2011, 340, 443–452. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Dikilitas, M.; Tuna, A.L. Alleviation of Salt Stress-Induced Adverse Effects on Maize Plants by Exogenous Application of Indoleacetic Acid (IAA) and Inorganic Nutrients - A Field Trial. Australian Journal of Crop Science 2013. [Google Scholar]
- Katerji, N.; van Hoorn, J.W.; Hamdy, A.; Karam, F.; Mastrorilli, M. Effect of Salinity on Water Stress, Growth, and Yield of Maize and Sunflower. Agricultural Water Management 1996, 30, 237–249. [Google Scholar] [CrossRef]
- Shahzad, M.; Witzel, K.; Zörb, C.; Mühling, K.H. Growth-Related Changes in Subcellular Ion Patterns in Maize Leaves (Zea Mays L.) under Salt Stress. Journal of Agronomy and Crop Science 2012, 198, 46–56. [Google Scholar] [CrossRef]
- Njeru, F.; Wambua, A.; Muge, E.; Haesaert, G.; Gettemans, J.; Misinzo, G. Major Biotic Stresses Affecting Maize Production in Kenya and Their Implications for Food Security. PeerJ 2023, 11, e15685. [Google Scholar] [CrossRef] [PubMed]
- Lodha, T.D.; Hembram, P.; Nitile Tep, J.B. Proteomics: A Successful Approach to Understand the Molecular Mechanism of Plant-Pathogen Interaction. 2013, 2013. [Google Scholar] [CrossRef]
- Gong, F.; Yang, L.; Tai, F.; Hu, X.; Wang, W. “Omics” of Maize Stress Response for Sustainable Food Production: Opportunities and Challenges. OMICS 2014, 18, 714–732. [Google Scholar] [CrossRef] [PubMed]
- Arshad, M.; Ranamukhaarachchi, S.L.; Ahmad, S.; Nawaz, R.; Qayyum, M.M.N.; Razaq, A.; Faiz, F. Variability and Correlation of Selected Soil Attributes and Maize Yield Influenced by Tillage Systems in Mountainous Agroecosystem. Journal of Soil and Water Conservation 2022, 77, 466–481. [Google Scholar] [CrossRef]
- Kumar, J.; Abbo, S. Genetics of Flowering Time in Chickpea and Its Bearing on Productivity in Semiarid Environments. Advances in Agronomy 2001, 72, 107–138. [Google Scholar] [CrossRef]
- Turner, N.; Wright, G.; Siddique, K. Adaptation of Grain Legumes (Pulses) to Water-Limited Environments. Advances in Agronomy, Vol 71 2001, 71, 193–231. [Google Scholar] [CrossRef]
- Khan, S.; Anwar, S.; Ashraf, M.Y.; Khaliq, B.; Sun, M.; Hussain, S.; Gao, Z.; Noor, H.; Alam, S. Mechanisms and Adaptation Strategies to Improve Heat Tolerance in Rice. A Review. Plants 2019, 8, 508. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants (Basel) 2021, 10, 259. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, P.; Zou, Y.-N.; Wu, Q.-S.; Kuča, K. Effects of Mycorrhizal Fungi on Root-Hair Growth and Hormone Levels of Taproot and Lateral Roots in Trifoliate Orange under Drought Stress. Archives of Agronomy and Soil Science 2019, 65, 1316–1330. [Google Scholar] [CrossRef]
- Blum, A. Osmotic Adjustment Is a Prime Drought Stress Adaptive Engine in Support of Plant Production. Plant Cell Environ 2017, 40, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Lamaoui, M.; Jemo, M.; Datla, R.; Bekkaoui, F. Heat and Drought Stresses in Crops and Approaches for Their Mitigation. Front. Chem. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- Sandhu, D.; Kaundal, A. Dynamics of Salt Tolerance: Molecular Perspectives. In Biotechnologies of Crop Improvement, Volume 3: Genomic Approaches; Gosal, S.S., Wani, S.H., Eds.; Springer International Publishing: Cham, 2018; pp. 25–40 ISBN 978-3-319-94746-4.
- Amin, I.; Rasool, S.; Mir, M.A.; Wani, W.; Masoodi, K.Z.; Ahmad, P. Ion Homeostasis for Salinity Tolerance in Plants: A Molecular Approach. Physiol Plant 2021, 171, 578–594. [Google Scholar] [CrossRef] [PubMed]
- Basu, S.; Kumar, A.; Benazir, I.; Kumar, G. Reassessing the Role of Ion Homeostasis for Improving Salinity Tolerance in Crop Plants. Physiologia Plantarum 2021, 171, 502–519. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, J.; Wang, Y.; Liang, X.; Zhang, M.; Lu, M.; Guo, Y.; Qin, F.; Jiang, C. The Classical SOS Pathway Confers Natural Variation of Salt Tolerance in Maize. New Phytologist 2022, 236, 479–494. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, Y. Elucidating the Molecular Mechanisms Mediating Plant Salt-Stress Responses. New Phytologist 2018, 217, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Wang, Y.; Gui, C.; Nguvo, K.J.; Ma, L.; Wang, Q.; Shen, Q.; Zhang, R.; Gao, X. Beneficial Rhizobacterium Triggers Induced Systemic Resistance of Maize to Gibberella Stalk Rot via Calcium Signaling. Mol Plant Microbe Interact 2023, 36, 516–528. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Horie, T.; Xue, S.; Leung, H.-Y.; Katsuhara, M.; Brodsky, D.E.; Wu, Y.; Schroeder, J.I. Differential Sodium and Potassium Transport Selectivities of the Rice OsHKT2;1 and OsHKT2;2 Transporters in Plant Cells. Plant Physiology 2009, 152, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Zivcak, M.; Brestic, M.; Sytar, O. Osmotic Adjustment and Plant Adaptation to Drought Stress. In Drought Stress Tolerance in Plants, Vol 1: Physiology and Biochemistry; Hossain, M.A., Wani, S.H., Bhattacharjee, S., Burritt, D.J., Tran, L.-S.P., Eds.; Springer International Publishing: Cham, 2016; pp. 105–143. ISBN 978-3-319-28899-4. [Google Scholar]
- Subbarao, G.V.; Nam, N.H.; Chauhan, Y.S.; Johansen, C. Osmotic Adjustment, Water Relations and Carbohydrate Remobilization in Pigeonpea under Water Deficits. Journal of Plant Physiology 2000, 157, 651–659. [Google Scholar] [CrossRef]
- Ghosh, U.K.; Islam, M.N.; Siddiqui, M.N.; Khan, M.A.R. Understanding the Roles of Osmolytes for Acclimatizing Plants to Changing Environment: A Review of Potential Mechanism. Plant Signal Behav 2021, 16, 1913306. [Google Scholar] [CrossRef] [PubMed]
- Yancey, P.H. Compatible and Counteracting Solutes: Protecting Cells from the Dead Sea to the Deep Sea. Science Progress 2004, 87, 1–24. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of Glycine Betaine and Proline in Improving Plant Abiotic Stress Resistance. Environmental and Experimental Botany 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Yoshiba, Y.; Kiyosue, T.; Nakashima, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Regulation of Levels of Proline as an Osmolyte in Plants under Water Stress. Plant Cell Physiol 1997, 38, 1095–1102. [Google Scholar] [CrossRef]
- Rosa, M.; Prado, C.; Podazza, G.; Interdonato, R.; González, J.A.; Hilal, M.; Prado, F.E. Soluble Sugars—Metabolism, Sensing and Abiotic Stress. Plant Signal Behav 2009, 4, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.-F.; Ji, L.; Zhang, L.; Dong, X.-Y.; Sun, Y. Molecular Basis for Polyol-Induced Protein Stability Revealed by Molecular Dynamics Simulations. J Chem Phys 2010, 132, 225103. [Google Scholar] [CrossRef]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent Developments in Enzymatic Antioxidant Defence Mechanism in Plants with Special Reference to Abiotic Stress. Biology (Basel) 2021, 10, 267. [CrossRef] [PubMed]
- Wang, C.; Yang, A.; Yin, H.; Zhang, J. Influence of Water Stress on Endogenous Hormone Contents and Cell Damage of Maize Seedlings. J Integr Plant Biol 2008, 50, 427–434. [Google Scholar] [CrossRef]
- Jones, B.; Gunnerås, S.A.; Petersson, S.V.; Tarkowski, P.; Graham, N.; May, S.; Dolezal, K.; Sandberg, G.; Ljung, K. Cytokinin Regulation of Auxin Synthesis in Arabidopsis Involves a Homeostatic Feedback Loop Regulated via Auxin and Cytokinin Signal Transduction. Plant Cell 2010, 22, 2956–2969. [Google Scholar] [CrossRef]
- Weiss, D.; Ori, N. Mechanisms of Cross Talk between Gibberellin and Other Hormones. Plant Physiol 2007, 144, 1240–1246. [Google Scholar] [CrossRef]
- Rao, S.R.; Qayyum, A.; Razzaq, A.; Ahmad, M.; Mahmood, I.; Sher, A. Role of Foliar Application of Salicylic Acid and L-Tryptophan in Drought Tolerance of Maize. J. Anim. Plant Sci. 2012, 22, 768–772. [Google Scholar]
- Shen, Y.-Y.; Wang, X.-F.; Wu, F.-Q.; Du, S.-Y.; Cao, Z.; Shang, Y.; Wang, X.-L.; Peng, C.-C.; Yu, X.-C.; Zhu, S.-Y.; et al. The Mg-Chelatase H Subunit Is an Abscisic Acid Receptor. Nature 2006, 443, 823–826. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants (Basel) 2020, 9, 681. [Google Scholar] [CrossRef] [PubMed]
- Hung, H.-Y.; Shannon, L.M.; Tian, F.; Bradbury, P.J.; Chen, C.; Flint-Garcia, S.A.; McMullen, M.D.; Ware, D.; Buckler, E.S.; Doebley, J.F.; et al. ZmCCT and the Genetic Basis of Day-Length Adaptation Underlying the Postdomestication Spread of Maize. Proceedings of the National Academy of Sciences 2012, 109, E1913–E1921. [Google Scholar] [CrossRef] [PubMed]
- Jasso-Robles, F.I.; Jiménez-Bremont, J.F.; Becerra-Flora, A.; Juárez-Montiel, M.; Gonzalez, M.E.; Pieckenstain, F.L.; García De La Cruz, R.F.; Rodríguez-Kessler, M. Inhibition of Polyamine Oxidase Activity Affects Tumor Development during the Maize-Ustilago Maydis Interaction. Plant Physiology and Biochemistry 2016, 102, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Berberich, T.; Miyazaki, A.; Seo, S.; Ohashi, Y.; Kusano, T. Spermine Signalling in Tobacco: Activation of Mitogen-Activated Protein Kinases by Spermine Is Mediated through Mitochondrial Dysfunction. Plant J 2003, 36, 820–829. [Google Scholar] [CrossRef] [PubMed]
- Herridge, D.F.; Peoples, M.B.; Boddey, R.M. Global Inputs of Biological Nitrogen Fixation in Agricultural Systems. Plant Soil 2008, 311, 1–18. [Google Scholar] [CrossRef]
- Peoples, M.B.; Brockwell, J.; Herridge, D.F.; Rochester, I.J.; Alves, B.J.R.; Urquiaga, S.; Boddey, R.M.; Dakora, F.D.; Bhattarai, S.; Maskey, S.L.; et al. The Contributions of Nitrogen-Fixing Crop Legumes to the Productivity of Agricultural Systems. Symbiosis 2009, 48, 1–17. [Google Scholar] [CrossRef]
- Akiyama, K.; Hayashi, H. Strigolactones: Chemical Signals for Fungal Symbionts and Parasitic Weeds in Plant Roots. Ann Bot 2006, 97, 925–931. [Google Scholar] [CrossRef]
- Lucido, A.; Andrade, F.; Basallo, O.; Eleiwa, A.; Marin-Sanguino, A.; Vilaprinyo, E.; Sorribas, A.; Alves, R. Modeling the Effects of Strigolactone Levels on Maize Root System Architecture. Front. Plant Sci. 2024, 14. [Google Scholar] [CrossRef]
- Guan, J.C.; Koch, K.E.; Suzuki, M.; Wu, S.; Latshaw, S.; Petruff, T.; Goulet, C.; Klee, H.J.; McCarty, D.R. Diverse Roles of Strigolactone Signaling in Maize Architecture and the Uncoupling of a Branching-Specific Subnetwork. Plant Physiol 2012, 160, 1303–1317. [Google Scholar] [CrossRef] [PubMed]
- Genre, A.; Chabaud, M.; Timmers, T.; Bonfante, P.; Barker, D.G. Arbuscular Mycorrhizal Fungi Elicit a Novel Intracellular Apparatus in Medicago Truncatula Root Epidermal Cells before Infection. Plant Cell 2005, 17, 3489–3499. [Google Scholar] [CrossRef] [PubMed]
- Mortier, E.; Mounier, A.; Kreplak, J.; Martin-Laurent, F.; Recorbet, G.; Lamotte, O. Evidence That a Common Arbuscular Mycorrhizal Network Alleviates Phosphate Shortage in Interconnected Walnut Sapling and Maize Plants. Front. Plant Sci. 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Liu, M.; Wang, Z.; Li, J.; Liu, K.; Huang, D. The Role of Arbuscular Mycorrhizal Symbiosis in Plant Abiotic Stress. Front Microbiol 2024, 14, 1323881. [Google Scholar] [CrossRef] [PubMed]
- Hui, J.; An, X.; Li, Z.; Neuhäuser, B.; Ludewig, U.; Wu, X.; Schulze, W.; Chen, F.; Feng, G.; Lambers, H.; et al. The Mycorrhiza-Specific Ammonium Transporter ZmAMT3;1 Mediates Mycorrhiza-Dependent Nitrogen Uptake in Maize Roots. The Plant Cell 2022, 34. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Meng, L.; Yin, T.; Li, Y.; Zhang, Y.; Li, S. Changes in Soil Rhizobia Diversity and Their Effects on the Symbiotic Efficiency of Soybean Intercropped with Maize. Agronomy 2023, 13, 997. [Google Scholar] [CrossRef]
- Lucas, T.M.R.; Carlos, H.C.; Fabio, L.C.M.; Gustavo, V.M. Azospirillum Spp. Potential for Maize Growth and Yield. Afr. J. Biotechnol. 2018, 17, 574–585. [Google Scholar] [CrossRef]
- Hayat, R.; Ahmed, I.; Sheirdil, R.A. An Overview of Plant Growth Promoting Rhizobacteria (PGPR) for Sustainable Agriculture. In Crop Production for Agricultural Improvement; Ashraf, M., Öztürk, M., Ahmad, M.S.A., Aksoy, A., Eds.; Springer Netherlands: Dordrecht, 2012; pp. 557–579. ISBN 978-94-007-4115-7. [Google Scholar]
- Garcia de Salamone, I.E.; Döbereiner, J.; Urquiaga, S.; Boddey, R.M. Biological Nitrogen Fixation in Azospirillum Strain-Maize Genotype Associations as Evaluated by the 15N Isotope Dilution Technique. Biol Fertil Soils 1996, 23, 249–256. [Google Scholar] [CrossRef]
- Deynze, A.V.; Zamora, P.; Delaux, P.-M.; Heitmann, C.; Jayaraman, D.; Rajasekar, S.; Graham, D.; Maeda, J.; Gibson, D.; Schwartz, K.D.; et al. Nitrogen Fixation in a Landrace of Maize Is Supported by a Mucilage-Associated Diazotrophic Microbiota. PLOS Biology 2018, 16, e2006352. [Google Scholar] [CrossRef]
- Xu, L.; Wang, A.; Wang, J.; Wei, Q.; Zhang, W. Piriformospora Indica Confers Drought Tolerance on Zea Mays L. through Enhancement of Antioxidant Activity and Expression of Drought-Related Genes. The Crop Journal 2017, 5, 251–258. [Google Scholar] [CrossRef]
- Ali, B.; Hafeez, A.; Afridi, M.S.; Javed, M.A.; Sumaira; Suleman, F.; Nadeem, M.; Ali, S.; Alwahibi, M.S.; Elshikh, M.S.; et al. Bacterial-Mediated Salinity Stress Tolerance in Maize (Zea Mays L.): A Fortunate Way toward Sustainable Agriculture. ACS Omega 2023, 8, 20471–20487. [Google Scholar] [CrossRef] [PubMed]
- Ryals, J.; Neuenschwander, U.; Willits, M.; Molina, A.; Steiner, H.; Hunt, M. Systemic Acquired Resistance. Plant Cell 1996, 8, 1809–1819. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Gui, Y.; Li, Z.; Jiang, C.; Guo, J.; Niu, D. Induced Systemic Resistance for Improving Plant Immunity by Beneficial Microbes. Plants (Basel) 2022, 11, 386. [Google Scholar] [CrossRef] [PubMed]
- Bakker, P.A.H.M.; Doornbos, R.F.; Zamioudis, C.; Berendsen, R.L.; Pieterse, C.M.J. Induced Systemic Resistance and the Rhizosphere Microbiome. Plant Pathol J 2013, 29, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, K.; Fan, L.; Fu, K.; Yu, C.; Wang, M.; Xia, H.; Sun, J.; Li, Y.; Chen, J. Cellulase from Trichoderma Harzianum Interacts with Roots and Triggers Induced Systemic Resistance to Foliar Disease in Maize. Sci Rep 2016, 6, 35543. [Google Scholar] [CrossRef] [PubMed]
- Rojas-Tapias, D.; Moreno-Galván, A.; Pardo-Díaz, S.; Obando, M.; Rivera, D.; Bonilla, R. Effect of Inoculation with Plant Growth-Promoting Bacteria (PGPB) on Amelioration of Saline Stress in Maize (Zea Mays). Applied Soil Ecology 2012, 61, 264–272. [Google Scholar] [CrossRef]
- Naseem, H.; Bano, A. Role of Plant Growth-Promoting Rhizobacteria and Their Exopolysaccharide in Drought Tolerance of Maize. Journal of Plant Interactions 2014, 9, 689–701. [Google Scholar] [CrossRef]
- Gou, W.; Tian, L.; Ruan, Z.; Zheng, P.; Chen, F.; Zhang, L.; Cui, Z.; Zheng, P.; Li, Z.; Gao, M.; et al. Accumulation of Choline and Glycinebetaine and Drought Stress Tolerance Induced in Maize (Zea Mays) by Three Plant Growth Promoting Rhizobacteria (PGPR) Strains. Pakistan Journal of Botany 2015, 47, 581–586. [Google Scholar]
- Huang, X.-F.; Zhou, D.; Lapsansky, E.; Reardon, K.; Guo, J.; Andales, M.; Vivanco, J.; Manter, D. Mitsuaria Sp. and Burkholderia Sp. from Arabidopsis Rhizosphere Enhance Drought Tolerance in Arabidopsis Thaliana and Maize (Zea Mays L.). Plant and Soil 2017, 419. [Google Scholar] [CrossRef]
- Moreno-Galván, A.; Cortés-Patiño, S.; Romero-Perdomo, F.; Uribe-Vélez, D.; Bashan, Y.; Bonilla, R. Proline Accumulation and Glutathione Reductase Activity Induced by Drought-Tolerant Rhizobacteria as Potential Mechanisms to Alleviate Drought Stress in Guinea Grass. Applied Soil Ecology 2020, 147, 103367. [Google Scholar] [CrossRef]
- Molina-Romero, D.; Baez, A.; Quintero-Hernández, V.; Castañeda-Lucio, M.; Fuentes-Ramírez, L.E.; Bustillos-Cristales, M. del R.; Rodríguez-Andrade, O.; Morales-García, Y.E.; Munive, A.; Muñoz-Rojas, J. Compatible Bacterial Mixture, Tolerant to Desiccation, Improves Maize Plant Growth. PLoS One 2017, 12, e0187913. [Google Scholar] [CrossRef] [PubMed]
- Bano, Q.; Ilyas, N.; Bano, A.; Zafar, N.; Akram, A.; Hassan, F.U. Effect of Azospirillum Innoculation on Maize (Zea Mays L.) Under Drought Stress. Pakistan Journal of Botany 45, 13–20.
- Cohen, A.; Travaglia, C.; Bottini, R.; Piccoli, P. Participation of Abscisic Acid and Gibberellins Produced by Endophytic Azospirillum in the Alleviation of Drought Effects in Maize. Botany 2009, 87, 455–462. [Google Scholar] [CrossRef]
- Vardharajula, S.; Shaik, Z.A.; Grover, M.; Reddy, G.; Venkateswarlu, B. Drought-Tolerant Plant Growth Promoting Bacillus Spp.: Effect on Growth, Osmolytes, and Antioxidant Status of Maize under Drought Stress. Journal of Plant Interactions 2011, 6, 1–14. [Google Scholar] [CrossRef]
- Naveed, M.; Mitter, B.; Reichenauer, T.G.; Wieczorek, K.; Sessitsch, A. Increased Drought Stress Resilience of Maize through Endophytic Colonization by Burkholderia Phytofirmans PsJN and Enterobacter Sp. FD17. Environmental and Experimental Botany 2014, 97, 30–39. [Google Scholar] [CrossRef]
- Quiroga, G.; Erice, G.; Ding, L.; Chaumont, F.; Aroca, R.; Ruiz-Lozano, J.M. The Arbuscular Mycorrhizal Symbiosis Regulates Aquaporins Activity and Improves Root Cell Water Permeability in Maize Plants Subjected to Water Stress. Plant, Cell & Environment 2019, 42, 2274–2290. [Google Scholar] [CrossRef]
- Yasmin, H.; Nosheen, A.; Naz, R.; Bano, A.; Keyani, R. L-Tryptophan-Assisted PGPR-Mediated Induction of Drought Tolerance in Maize (Zea Mays L.). Journal of Plant Interactions 2017, 12, 567–578. [Google Scholar] [CrossRef]
- Curá, J.A.; Franz, D.R.; Filosofía, J.E.; Balestrasse, K.B.; Burgueño, L.E. Inoculation with Azospirillum Sp. and Herbaspirillum Sp. Bacteria Increases the Tolerance of Maize to Drought Stress. Microorganisms 2017, 5, 41. [Google Scholar] [CrossRef]
- Krishnamoorthy, R.; Kim, K.; Subramanian, P.; Senthilkumar, M.; Anandham, R.; Sa, T. Arbuscular Mycorrhizal Fungi and Associated Bacteria Isolated from Salt-Affected Soil Enhances the Tolerance of Maize to Salinity in Coastal Reclamation Soil. Agriculture, Ecosystems & Environment 2016, 231, 233–239. [Google Scholar] [CrossRef]
- Bano, A.; Fatima, M. Salt Tolerance in Zea Mays (L). Following Inoculation with Rhizobium and Pseudomonas. Biol Fertil Soils 2009, 45, 405–413. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Zahir, Z.A.; Naveed, M.; Arshad, M. Rhizobacteria Containing ACC-Deaminase Confer Salt Tolerance in Maize Grown on Salt-Affected Fields. Can J Microbiol 2009, 55, 1302–1309. [Google Scholar] [CrossRef]
- Sheng, M.; Tang, M.; Zhang, F.; Huang, Y. Influence of Arbuscular Mycorrhiza on Organic Solutes in Maize Leaves under Salt Stress. Mycorrhiza 2011, 21, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, Y.; Wu, G.; Kimani, V.; Shen, Q.; Zhang, N.; Zhang, R. Induced Maize Salt Tolerance by Rhizosphere Inoculation of Bacillus Amyloliquefaciens SQR9. Physiologia Plantarum 2016, 158. n/a-n/a. [Google Scholar] [CrossRef]
- Li, X.; Sun, P.; Zhang, Y.; Jin, C.; Guan, C. A Novel PGPR Strain Kocuria Rhizophila Y1 Enhances Salt Stress Tolerance in Maize by Regulating Phytohormone Levels, Nutrient Acquisition, Redox Potential, Ion Homeostasis, Photosynthetic Capacity and Stress-Responsive Genes Expression. Environmental and Experimental Botany 2020, 174, 104023. [Google Scholar] [CrossRef]
- Ahmad, M.; Imtiaz, M.; Nawaz, M.S.; Mubeen, F.; Sarwar, Y.; Hayat, M.; Asif, M.; Naqvi, R.Z.; Ahmad, M.; Imran, A. Thermotolerant PGPR Consortium B3P Modulates Physio-Biochemical and Molecular Machinery for Enhanced Heat Tolerance in Maize during Early Vegetative Growth. Annals of Microbiology 2023, 73, 34. [Google Scholar] [CrossRef]
- Mathur, S.; Agnihotri, R.; Sharma, M.P.; Reddy, V.R.; Jajoo, A. Effect of High-Temperature Stress on Plant Physiological Traits and Mycorrhizal Symbiosis in Maize Plants. Journal of Fungi 2021, 7, 867. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Song, F.-B.; Xu, H.-W. Arbuscular Mycorrhizae Improves Low Temperature Stress in Maize via Alterations in Host Water Status and Photosynthesis. Plant and Soil 2010, 331, 129–137. [Google Scholar] [CrossRef]
- Mathur, S.; Jajoo, A. Arbuscular Mycorrhizal Fungi Protects Maize Plants from High Temperature Stress by Regulating Photosystem II Heterogeneity. Industrial Crops and Products 2020, 143, 111934. [Google Scholar] [CrossRef]
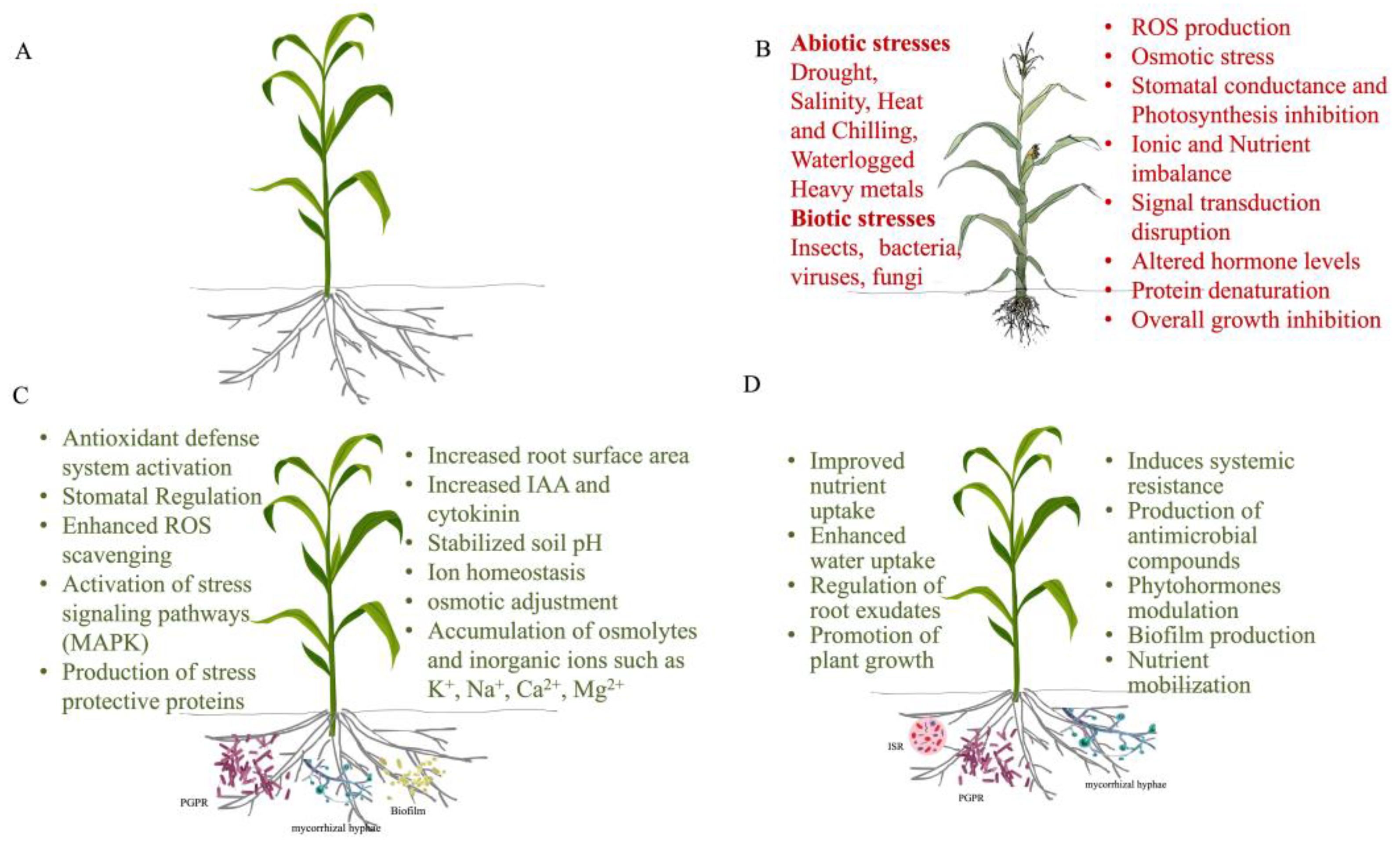
Figure 1.
An overview of maize plants showing (A) a healthy maize plant, (B) a maize plant affected by abiotic and biotic stresses, (C) mechanisms of abiotic stress tolerance including osmotic adjustment, antioxidant activity, and stomatal regulation, and (D) mechanisms of biotic stress tolerance such as activation of pathogenesis-related proteins and structural barriers.
Figure 1.
An overview of maize plants showing (A) a healthy maize plant, (B) a maize plant affected by abiotic and biotic stresses, (C) mechanisms of abiotic stress tolerance including osmotic adjustment, antioxidant activity, and stomatal regulation, and (D) mechanisms of biotic stress tolerance such as activation of pathogenesis-related proteins and structural barriers.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



