
Submitted:
04 June 2024
Posted:
05 June 2024
You are already at the latest version
Abstract
Antimicrobial resistance has been stated to be a global health problem. In Chile, the use of antibiotics should be declared by medical prescription, however, it is unknown what happens to the drugs once the treatment ends. One possibility is that these end up being disposed of in the trash or down the drain. In both scenarios, antibiotics could accumulate in the environment, stimulating the emergence of antimicrobial resistance mechanisms and their transfer between microorganisms. Unfortunately, sometimes wastewater ends up in bodies of water, due to the dragging of elements by rain, or by the presence of illegal water discharges. In this work, we use shotgun metagenomics to elucidate the functional and microbial composition of biohazard elements in the bay of Puerto Varas City, Chile. As expected, we found a high diversity in microbial communities, with bacterial elements described as human or animal pathogens. Also, we detect a diverse repertory of virulence and antibiotic-resistant genes (ARGs), related with macrolides, beta-lactams, and tetracyclines resistance, which are consistent with the families of antibiotics most used in Chile. Similar ARGs were identified in DNA mobile elements. In addition, we tested the AMR ability in 20 bacterial strains recovered from the Llanquihue lake. This is the first report of the presence of genomic elements that could constitute a health problem for the people who live around the Llanquihue Lake of Chile, considering the importance of the interconnection between environmental, animal, and human health, a concept known as One Health.
Keywords:
Antimicrobial resistance
; metagenomics
; DNA mobile elements
1. Introduction
The emergence and spread of antimicrobial resistance (AMR) is a major global health problem [1]. The World Health Organization (WHO) has classified it as a major threat to global public health [2]. AMR occurs when microbes, such as bacteria, fungi, viruses, and parasites, become resistant to the drugs used to treat them [3]. This makes it difficult or even impossible to treat infections caused by antimicrobial resistant microorganisms [4].
The increment in antimicrobial resistant mechanism and transmission of resistance genes from environmental bacteria to pathogenic strains is an imperative threatening to human and animal health which deserves the public and governmental attention [5]. AMR has economic consequences, its impacts include increased healthcare costs, lost productivity and overall economic burden [6]. Antimicrobial resistance can lead to increased costs of treating resistant bacterial infections. The economic burden goes beyond healthcare costs and includes reduced income due to prolonged illness and premature death, affecting both individuals and society [7]. In low- and middle-income countries, where infectious diseases are most prevalent, the failure of first-line antibiotics has led to increased mortality and costs. In addition, the economic impact of antimicrobial resistance affects not only humans, but also animals, leading to economic losses in animal husbandry and further straining economies [8].
There are many factors that contribute to the development and spread of AMR, including the overuse and misuse of antibiotics in human and animal clinical pharmacotherapy, the indiscriminate use of antibiotics in animal production as growth promoters, poor sanitation and hygiene of health care systems, and the improper disposal of waste contaminated with antibiotics and resistant bacteria [9]. One of the leading contributors to the dissemination of AMR is the presence of antimicrobial resistance genes (ARGs) in environmental reservoirs, such as lakes, rivers, and oceans [10]. Also, Mobile Genetic Elements (MGE) play a significant role in the transference and dispersion of Antimicrobial Resistance Genes (ARGs) among bacteria. They facilitate horizontal gene transfer (HGT) by various mechanisms, such as virions, conjugative systems, and plasmids. MGEs can carry ARGs and transfer them to other bacteria, contributing to the spread of antibiotic resistance [11]. Understanding the prevalence and distribution of ARGs and MGE in these aquatic ecosystems is crucial for developing strategies to mitigate their potential impact on public health. The resistome consists of all ARGs, including those circulating in both pathogenic and non-pathogenic bacteria [12], and aquatic environments has been already reported as reservoir of these ARGs elements [13,14].
Llanquihue Lake is the second largest lake in Chile, with a surface area of approximately 860 km2 and a maximum register depth of 317 m. The lake is situated in the Llanquihue Basin, which is bounded by the Andes Mountains to the east and the Coastal Range to the west. It is surrounded by several cities and towns, each with its own unique charm and attractions [15]. Puerto Varas, located on the eastern shore of the lake, is a popular tourist destination, with an urban population of around 26.000 inhabitants, based on the 2017 census of Chile. Given the significant anthropogenic activities and the growing concern regarding AMR, it becomes imperative to investigate the presence and characteristics of ARGs and MGE in the coastal waters of Llanquihue Lake. By investigating the presence of ARGs elements in coastal water recovered from Puerto Varas shore, this study aims to shed light on the prevalence, distribution, and potential implications of antimicrobial resistance in this specific aquatic environment. This research is essential for developing strategies to mitigate the spread of antimicrobial resistance and safeguard public health in the region.
In this work, we use shotgun metagenomic to perform a quick description of the microbial composition of Llanquihue lake in three points of Puerto Varas city shore, aiming to characterize bacterial communities present in the beach, and identify the existence and abundance of ARGs in the environmental DNA due to the important role of these biologic elements for human health. This research will help to understand the hazards associated with AMR transfer from environmental reservoirs to humans by assessing the ARGs presented, and identifying the MGE which may drive ARGs transference. This works constitutes an example of how metagenomics can be useful in the surveillance of microbiological risks at city scale.
2. Results
2.1. Composition of Bacterial Communities That Inhabit Llanquihue Lake
Shotgun metagenomic sequencing and bioinformatic analysis indicate a heterogeneous taxonomic composition of bacterial microorganism in the Llanquihue lake (Figure 1, Supplementary Figure 1 and Supplementary Table1), pointing the high diversity of bacterial microbial communities (Figure 2). Taxonomical assignation shows that the main phyla presented belong to Proteobacteria, Firmicutes, Bacteroidota, Actinobacteriota, and Verrucomicrobiota (Figure 2 and Supplementary Table 1). In the taxonomic data, we identify the presence of bacterial genera Brucella, Mycoplasma, Mycobacterium, Microcystis, Leptospira and Flavobacterium, which harbor bacterial species of interest in veterinary health [16]. In addition, we found the presence of bacterial genera belonging to the human intestinal microbiota such as Prevotella, Coprococcus, Bifidobacterium, Faecalibacterium, and Ruminococcus [17], in fact, Prevotella copri was one of the most abundant specie in the sample PV1.1. At specie level, we were able to identify a total of 3740 species (Supplementary Table 1), including environmental water related species such as Nanopelagicus abundans [18], Fonsibacter ubiquis [19], and Planktophila vernalis [20]. Interestingly, several taxonomical species related with skin and intestinal infections in humans were detected, mainly belonging to the genus Campylobacter, Clostridium, Escherichia, Mycobacterium, Salmonella, Shigella, Staphylococcus, Streptococcus, and Yersinia (Supplementary Figure 1). Although many of these taxonomic findings are repeated among the analyzed sites, the abundance of each of these taxonomies varies, indicating that microbial communities may be different regard its location (Figure 1).
2.2. Identification of AMR Genes Present on DNA Recovered from Llanquihue Lake
Our metagenomic analysis revealed a diverse array of antimicrobial resistance genes across the environmental DNA samples recovered from water. To report this, we group the different genes identified regarding their antimicrobial resistance family which belongs to (Figure 3). These included genes which confers resistance to antibiotics commonly used in human clinical treatments, such as β-lactams, chloramphenicol, and penicillin’s, as well as genes associated with resistance to antibiotics used in agriculture and veterinary medicine such as macrolides, tetracyclines and fluoroquinolones, such as the tet genes, which confer resistance to tetracyclines by encoding for efflux proteins, or by encoding ribosomal protection proteins or enzymes that chemically modify tetracycline [21]. Another’s remarkable families of genes with high presence were the bla family, which encode for the resistance to beta-lactam antibiotics [22,23], the dfr genes which encodes the trimethoprim resistant dihydrofolate reductase, initially founded in Escherichia coli, Salmonella enterica and Pasteurella multocida [24,25,26] and cat genes. This lately encodes for chloramphenicol acetyltransferase which inactivates chloramphenicol by addition of an acyl group [27]. Notably, the abundance and composition of ARGs varied spatially and temporally, with presence in areas impacted by anthropogenic activities, such as rainwater drainage and public beaches. Table 1 summarizes the antimicrobial family gene and the pharmaceutical drug example which confers resistance.
2.3. Mobile Genetic Elements Carrying AMR Genes Are Related with Microbial Species of Health Interest
To further completion of the characterization of ARGs present in the Llanquihue lake, we also screened metagenomic assemblies to explore the presence of MGE. The results showed the presence of mobile elements belonging mainly to the MOBP1 group, classification based on their relaxase gene [35], which included elements identified in bacterial host belonging to genus Clostridiales, Clostridioides, Aeromonas, Vibrio, Enterococcus, Escherichia, Bacteroides, Parabacteroides, Klebsiella, and Proteus (Supplementary Table 2). However, a significant portion of the mobile elements were not classified according to a MOB group, such as pR997, pSX2_LC6, pRIVM_C010068_1, pAFAEC, and pMMCAT_PdisCL06T03, identified originally in host such us Proteus mirabilis, Shewanella sp., Enterobacter hormaechei, Aliarcobacter faecis, and Parabacteroides distasonis, respectively. Furthermore, we identified an overlap between the original host reported for MGE identified in our metagenomic data and the presence of harmful microbial species (Supplementary Figure 1 and Supplementary Table 2).
Because MGE plays a significant role in the evolution and adaptation of organisms by facilitating genetic diversity and horizontal gene transfer, we wondered what kind of genes are being carried by the plasmid sequences identified. For this purpose, we explore MGE sequences (Supplementary File 1), to look for the specific presence of ARGs in its genomic code. As a general trend, we found ARGs which confers resistance for erythromycin, azithromycin, lincomycin, doxycycline, tetracycline, amoxicillin, ampicillin, encodes mostly by the genes mph(E), msr(D), mef(A), erm(B), erm(F), lnu(C), tet(M), tet(W), tet(C), tet(O), blaSHV-12, and ant(6), respectively. These ARGs were carried in the MGE sequences (Supplementary File 1) identified initially in bacteria belonging to species such as Lactococcus garvieae, Enterococcus faecalis, Acinetobacter sp., Escherichia coli, and Shewanella sp.
2.4. Antimicrobial Susceptibility Test of Microbial Isolates Do Not Show the Presence of Antimicrobial Resistance Patterns
A total of 14 different isolates of enterobacteria were recovered and cultured from water samples. A total of 3 isolates belonging to Citrobacter spp., 3 of Enterobacter spp., 6 of E. coli, and 1 of Rahnella aquatilis, were identified by 16S rRNA PCR, Sanger sequencing, and BLAST. Six antibiotics were tested on all isolated bacteria, the inhibition halos obtained ranged from 15 to 49 mm. No resistant bacterial populations according to CLSI classification could be detected (Table 2).
3. Discussion
Currently, studies with a One Health perspective are an urgent need due to threats associated with AMR. It is crucial to understand that this phenomenon is ubiquitous, so research must be conducted in humans, animals and the environment. Detection of a high abundance of ARGs in different environments corresponds to one of the first steps required to counteract this phenomenon. Nevertheless, as cultivable bacteria only represent a small fraction of the whole microbiota within a specific environment [37,38], the AR monitoring mostly depends on studios in total DNA extracts [39]. Studies related to ARGs in a variety of environmental compartments has been supported by molecular biology-based methods and sequencing methods [40,41,42,43,44]. Here, we studied the presence of ARGs in Llanquihue lake through metagenome sequencing. As metagenomics is a non-targeted method for detection and quantifying taxonomic and functional genetic diversity in each environment; these strategies allow to infer about occurrence and proportions of a variety of groups within a complex microbial community [45]. In addition, metagenomics is one of the most attractive tools to explore natural environments due to the large amount of information that can be obtained [46,47,48].
We could evidence the presence of cfxA6 and a cfxA2 genes in lake Llanquihue, related to the expression of class A beta lactamases, which have both cephalosporins and penicillins as substrates. In China, the presence of these genes related to this family has been detected also in different water bodies [49]. These genes were also found in wastewater from a treatment plant in Poland using metagenomics techniques [50]. In this study, we have found genes associated with the bla family: blaFAR-1, blaOXA-490 and blaOXA-491 and blaTEM-102 and -104, these genes are also associated with the expression of beta-lactamases in different pathogenic bacteria. Regarding blaFAR-1, blaOXA-490 and -491, we did not find reports of their presence in water bodies, but we did find reports of their presence in blaTEM-102 and -104. These genes have been reported in different bodies of water, and these studies mention the potential risk to human health of these genes being found in the environment [51,52,53].
Interestingly, we were also able to demonstrate the presence of different genes associated with the mcr gene family (mcr-3, -4, -5 and -7). These genes are related to colistin resistance, a highly relevant drug in the treatment of infections complicated by multiresistant gram-negative bacteria [54,55,56]. Some studies demonstrate the presence of these genes in water bodies around the world [57,58]. This gene’s presence in a lake where recreational activities are done constitutes a risk for the population. Other of the most abundant gene families found in Lake Llanquihue were genes associated with resistance to tetracyclines such as tet(37), tet(A), tet(C), tet(O), tet(Q) and tet(W). Several studies have shown the presence of these resistance genes in aquatic environments [59,60,61,62]. For example, tet(37) gene family, has been reported at the environmental level in an anthropogenically stressed estuary on the northwest coast of Portugal [63]. In particular, the presence of these genes poses risks for productive activities associated with aquaculture. This is because one of the most widely used antibiotics in freshwater production cycles is oxytetracycline [64]. The potential expression of these genes in pathogenic bacteria affecting farmed fish could cause the ineffectiveness of these treatments. In summary, the presence of these genes in DNA isolated from water reservoirs highlights the widespread distribution of antimicrobial resistance determinants in the environment. Additionally, our analysis unveiled the presence of MGE, such as plasmids associated with ARGs, highlighting the dynamic nature of AMR in aquatic ecosystems and its potential dissemination.
One limitation of our study was to isolate and characterize bacterial strains with antimicrobial resistance phenomena. As detection of the presence of ARGs from data obtained by sequencing does not necessarily imply the expression of such genes in each microbial community, further empirical tests are required for describing putative expression of resistant phenotypes [65]. To aboard this, we performed susceptibility assays in 20 microbial isolates obtained from Llanquihue lake. Although in our microbiologic results the presence of multiresistant strains was not detected as was expected regard to the metagenomic results presented here, these results might not be entirely representative of the occurrence of multiresistant strains in the environment. ARGs detection through metagenome sequencing gives information about both, culturable and unculturable bacteria, thus classic microbiological techniques for culture and susceptibility assays from microbial isolated have limitations that should be considered.
Proper watershed management has important positive effects on the mitigation of human health risks associated with the presence of ARG in the environment [66]. The implementation of effective public politics on water management, such as regulating and monitoring the discharges of domestic, industrial and hospital wastewater into water bodies, can help to avoid ARG transference and pharmaceutical dispersion [67,68], for example, reducing the use of agricultural antibiotics close to water bodies [69] or limiting the presence of aquaculture activities in fresh water [70], would contribute to reducing the load of pharmaceutical pollutants in water systems. Additionally, constant monitoring of water quality would contribute significantly to decision making in promoting sustainable practices in the watershed [71]. Moreover, evidence-based decision-making on water quality, supported by monitoring data, can be crucial for delivering safe drinking water, optimizing water quality, and managing water resources effectively [72,73]. By minimizing water pollution, selective pressure on harmful microorganisms that inhabit the aquatic environment would be reduced, diminishing the spread of antimicrobial resistance and virulence genomic elements [74,75]. This comprehensive approach would not only protect the health of local communities by safeguarding the purity of the water resource but would also contribute to the preservation of antibiotic effectiveness and sustainable public health management in the long term [76].
4. Materials and Methods
Sample Collection and Microbial Isolation
We collected 10 water samples from the coastal shore of Puerto Varas city (Supplementary Figure 2). 3 liters for each sample site were collected using sterile 1 L glass bottles and preserved with icepack until they were processed within the same day they were taken. A total of 3 liters of water were filtered through mixed cellulose ester (MCE) membranes of 0.22 µm pore size (Merck-Millipore #GSWP04700, USA), using a glass filter system pumped with negative pressure. Filters were stored in RNA Later (Sigma-Aldrich #R0901, USA), until DNA extraction as described below. In parallel, 1 mL of water was streaking on BHI, TSA, EMB, Miller-Hinton, and McConkey agar plates, and cultivated at 25° C for 24 h. Cell colonies obtained were passed 2 times to facilitate its purification in agar plates and then, Gram stain was used to check the purity of the isolated bacteria. Isolated microorganisms were observed under microscopy and stored in sterile glycerol 10% v/v at -80 °C.
DNA Purification and Metagenomic Sequencing
The stored MCE filters were used for DNA extraction employing AccuPrep Genomic DNA Extraction Kit (Bioneer #K-3032, Korea), following manufacturer instructions. Briefly, filters were resuspended in 500 µl DNA Extraction buffer and stirred to release microbial cells. An enzymatic digestion with 20 µl of lysozyme (20 mg/mL) and 20 µl of proteinase K (20 mg/mL) were used to disrupt microbial cells. The suspension was incubated for 1 h at 37 °C and then for 1 h at 55 °C. After enzymatic digestion, we followed the steps provided by the manufacturer for bacterial DNA extraction. The quality of obtained the DNA was checked by 1% agarose gel electrophoresis, while DNA quantity was measured by absorbance, and the ratios 260/280 nm were calculated to assess the purity of DNA obtained. Before DNA sequencing, we test the amplification capacity of DNA using 16S bacterial universal PCR. A total of 1 µg of DNA was sent to Novogene (USA) genomic service for shotgun metagenomic sequencing. DNA was sequenced by paired-end (2x150 bp) reads using the Illumina NovaSeq 6000 platform with an output of 6 GB per sample.
Metagenomic Data Analysis and Identification of AMR Genes
Raw data obtained from the sequencing provider was initially inspected with FastQC (https://www.bioinformatics.babraham.ac.uk/projects/fastqc), and then reads were filtered and trimmed using Trimmomatic [77] using the following parameters: LEADING:20, TRAILING:20, SLIDINGWINDOW:5:20, AVGQUAL:20, and MINLEN:90, followed by the application of Bowtie2 to screen out the contaminant DNA sequences from human and viruses [78]. The paired-end files were merged using the script provided in the Microbiome Helper v2.3 pipeline [79] and metagenomic data were procesed to obtain metagenomics de novo assembly using MegaHit [80] and the quality of conting obtained were inspected using Quast [81]. The taxonomic profiling was obtained at specie level using Kraken2 [82], keeping the taxonomic assignation with over 500 hits by sample, while antimicrobial resistance genes were inspected using ABRicate [83], utilizing the Resfinder [84] databases. Mobile genetic elements were retrieved using plaSquid [85]. The fasta files obtained from plaSquid were used to look for the presence of AMR genes carried in the mobile elements using ABRicate as described above. Data obtained was imported to R statistical language [86] for further analysis and representation using phyloseq [87] and ggplot2 [88] packages.
Antimicrobial Susceptibility Assay
Antimicrobial susceptibility testing of 14 isolates was performed using the disk diffusion method described by Hudzicki, 2009 [89]. Mueller-Hinton I agar (DIFCO) was employed to evaluate bacterial susceptibility to six antibiotics drugs: ampicillin (2 µg), imipenem (10 µg), norfloxacin (10 µg), gentamicin (30 µg), vancomycin (5 µg), erythromycin (15 µg), tetracycline (30 μg) and trimethoprim-sulfamethoxazole (1.25–23.75 µg). Zone inhibition diameters were interpreted according to CLSI breakpoint tables [90]. All studies were done in triplicate. The halo measurements were expressed as the average of the measurements plus the standard deviation. Eschericha coli ATCCÓ 25922 was used as a quality control strain.
5. Conclusions
Our study provides valuable insights into the prevalence, diversity, and nature of antimicrobial resistance genes presented in environmental water recovered from a lake system enclosed beside a city. By elucidating the dynamics of ARG and virulence genes dissemination, we contribute to the collective efforts aimed at combatting the occurrence of resistance phenoms and preserving the efficacy of antimicrobial agents for future generations. Research on the identification of antimicrobial resistance and virulence genes in environmental water highlights the urgent need for standardized monitoring methods to address the global public health threat posed by antibiotic resistance. Understanding the presence, diversity, and transmission pathways of resistance genes in water environments is essential for developing effective strategies to mitigate the spread of antimicrobial resistance between microbial species and the generation of antimicrobial multidrug resistant microorganism.
Author Contributions
Conceptualization, J.C.S. and D.A.M.; Methodology, J.C.S. and D.A.M; Validation J.C.S., D.A.M., M.C.T and P.P; Formal Analysis, D.A.M. and M.G.; Investigation, J.C.S., C.O., J.S.M, D.C, N.D.R; Resources, J.C.S. and D.A.M; Data Curation, D.A.M. and M.G; Writing – Original Draft Preparation, J.C.S. and D.A.M; Writing – Review & Editing, J.C.S., D.A.M., M.C.T, D.C. and P.P Visualization, J.C.S., C.O, J.S.M. and D.A.M.; Supervision, J.C.S. and D.A.M; Project Administration, C.O. and D.A.M.;.; Funding Acquisition, J.C.S. and D.A.M..
Funding
This research was funded by ANID [FOVI#220211] (DAM) and by the Universidad San Sebastián by grants VRID_FAPPE21-07 and VRID_INTER23/02 (DAM). Also, this work was supported by Fondos de Innovación para la Competitividad [#BIP40018207-0] del Gobierno Regional de Los Lagos. We appreciate the support from the National Laboratory for High-Performance Computing (NLHPC) of Chile, for facilitating their infrastructure for data analysis. JCS thanks to the VRID of Universidad San Sebastián for funding VRID_DocI22/06.
Data Availability Statement
The raw data produced from DNA sequencing in this study was deposited at the ENA-EMBL database under the accession number PRJEB76156. Metadata and metagenomic tables obtained from bioinformatics analysis can be found in Supplementary Files.
Acknowledgments
We are grateful for the support of the Gobierno Regional Los Lagos, Chile, as well as the support of the Agencia Nacional de Investigación y Desarrollo of Chile. This work would not have been possible without the support of the Vicerrectoría de Investigación y Doctorados and the Vicerrectoría de Vinculación con el Medio of Universidad San Sebastián. Finally, we are grateful for the collaboration of the Universidad Nacional de Río Negro of Argentina.
Conflicts of Interest
The authors declare no conflicts of interest
References
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial Resistance: A Global Multifaceted Phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef]
- World Health Organization (WHO) Antimicrobial Resistance Available online:. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 27 May 2024).
- Uddin, T.M.; Chakraborty, A.J.; Khusro, A.; Zidan, B.M.R.M.; Mitra, S.; Emran, T.B.; Dhama, K.; Ripon, M.K.H.; Gajdács, M.; Sahibzada, M.U.K.; et al. Antibiotic Resistance in Microbes: History, Mechanisms, Therapeutic Strategies and Future Prospects. J. Infect. Public Health 2021, 14, 1750–1766. [Google Scholar] [CrossRef]
- Mancuso, G.; Midiri, A.; Gerace, E.; Biondo, C. Bacterial Antibiotic Resistance: The Most Critical Pathogens. Pathogens 2021, 10, 1310. [Google Scholar] [CrossRef]
- Salam, M.A.; Al-Amin, M.Y.; Salam, M.T.; Pawar, J.S.; Akhter, N.; Rabaan, A.A.; Alqumber, M.A.A. Antimicrobial Resistance: A Growing Serious Threat for Global Public Health. Healthcare (Basel) 2023, 11, 1946. [Google Scholar] [CrossRef] [PubMed]
- National Academies of Sciences, Engineering, and Medicine; Health and Medicine Division; Board on Population Health and Public Health Practice; Committee on the Long-Term Health and Economic Effects of Antimicrobial Resistance; Palmer, G.H.; Buckley, G.J. <italic>The Health and Economic Burden of Resistance</italic>; National Academies Press: Washington, D.C., DC, 2021;
- World Bank Group <italic>Drug-Resistant Infections: A Threat to Our Economic Future</italic>; World Bank Group, 2017;
- McGowan, J.E. , Jr Economic Impact of Antimicrobial Resistance. Emerg. Infect. Dis. 2001, 7, 286–292. [Google Scholar] [CrossRef]
- Fletcher, S. Understanding the Contribution of Environmental Factors in the Spread of Antimicrobial Resistance. Environ. Health Prev. Med. 2015, 20, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Huijbers, P.M.C.; Blaak, H.; de Jong, M.C.M.; Graat, E.A.M.; Vandenbroucke-Grauls, C.M.J.E.; de Roda Husman, A.M. Role of the Environment in the Transmission of Antimicrobial Resistance to Humans: A Review. Environ. Sci. Technol. 2015, 49, 11993–12004. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.H.K.; Aarestrup, F.M.; Petersen, T.N. Importance of Mobile Genetic Elements for Dissemination of Antimicrobial Resistance in Metagenomic Sewage Samples across the World. PLoS One 2023, 18, e0293169. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. Antibiotic Resistance in the Environment: A Link to the Clinic? Curr. Opin. Microbiol. 2010, 13, 589–594. [Google Scholar] [CrossRef]
- Zhang, X.-X.; Zhang, T.; Fang, H.H.P. Antibiotic Resistance Genes in Water Environment. Appl. Microbiol. Biotechnol. 2009, 82, 397–414. [Google Scholar] [CrossRef]
- Baquero, F.; Martínez, J.-L.; Cantón, R. Antibiotics and Antibiotic Resistance in Water Environments. Curr. Opin. Biotechnol. 2008, 19, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Kies, F.; De Los Rios-Escalante, P.; Matemilola, S.; Elegbede, I. Water Bodies Quality Assessment and Trophic Gradient Monitoring of the Llanquihue Lake-Maullin River in Chile from Years 1999-2014 Available online:. Available online: https://www.jmaterenvironsci.com/Document/vol12/vol12_N7/JMES-2021-12076-Kies.pdf (accessed on 27 May 2024).
- Quinn, P.J.; Markey, B.K.; Leonard, F.C.; Hartigan, P.; Fanning, S.; Fitzpatrick, E.S. Veterinary Microbiology and Microbial Disease; John Wiley & Sons, 2011; ISBN 9781405158237.
- Requena, T.; Velasco, M. The Human Microbiome in Sickness and in Health. Rev. Clin. Esp. (Barc.) 2021, 221, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Farkas, M.; Szoboszlay, S.; Vörös, L.; Lovász, Z.; Méhes, N.; Mátyás, K.; Sebők, R.; Kaszab, E.; Háhn, J.; Tóth, G.; et al. Bacterial Community Dynamics along a River-Wetland-Lake System. Water (Basel) 2022, 14, 3519. [Google Scholar] [CrossRef]
- Henson, M.W.; Lanclos, V.C.; Faircloth, B.C.; Thrash, J.C. Cultivation and Genomics of the First Freshwater SAR11 (LD12) Isolate. ISME J. 2018, 12, 1846–1860. [Google Scholar] [CrossRef] [PubMed]
- Neuenschwander, S.M.; Ghai, R.; Pernthaler, J.; Salcher, M.M. Microdiversification in Genome-Streamlined Ubiquitous Freshwater Actinobacteria. ISME J. 2018, 12, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Grossman, T.H. Tetracycline Antibiotics and Resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025387. [Google Scholar] [CrossRef]
- Ranjbar, R.; Sami, M. Genetic Investigation of Beta-Lactam Associated Antibiotic Resistance among Escherichia Coli Strains Isolated from Water Sources. Open Microbiol. J. 2017, 11, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Worthington, R.J.; Melander, C. Overcoming Resistance to β-Lactam Antibiotics. J. Org. Chem. 2013, 78, 4207–4213. [Google Scholar] [CrossRef] [PubMed]
- Wüthrich, D.; Brilhante, M.; Hausherr, A.; Becker, J.; Meylan, M.; Perreten, V. A Novel Trimethoprim Resistance Gene, DfrA36, Characterized from Escherichia Coli from Calves. mSphere 2019, 4. [Google Scholar] [CrossRef]
- Murphy, B.P.; O’Mahony, R.; Buckley, J.F.; Shine, P.; Fidelma Boyd, E.; Gilroy, D.; Fanning, S. Investigation of a Global Collection of NontyphoidalSalmonellaof Various Serotypes Cultured between 1953 and 2004 for the Presence of Class 1 Integrons. FEMS Microbiol. Lett. 2007, 266, 170–176. [Google Scholar] [CrossRef]
- Kehrenberg, C.; Schwarz, S. DfrA20, a Novel Trimethoprim Resistance Gene from Pasteurella Multocida. Antimicrob. Agents Chemother. 2005, 49, 414–417. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; Kehrenberg, C.; Doublet, B.; Cloeckaert, A. Molecular Basis of Bacterial Resistance to Chloramphenicol and Florfenicol. FEMS Microbiol. Rev. 2004, 28, 519–542. [Google Scholar] [CrossRef] [PubMed]
- Madinier, I.; Fosse, T.; Giudicelli, J.; Labia, R. Cloning and Biochemical Characterization of a Class A β-Lactamase from Prevotella Intermedia. Antimicrob. Agents Chemother. 2001, 45, 2386–2389. [Google Scholar] [CrossRef] [PubMed]
- Karray, F.; Darbon, E.; Oestreicher, N.; Dominguez, H.; Tuphile, K.; Gagnat, J.; Blondelet-Rouault, M.-H.; Gerbaud, C.; Pernodet, J.-L. Organization of the Biosynthetic Gene Cluster for the Macrolide Antibiotic Spiramycin in Streptomyces Ambofaciens. Microbiology 2007, 153, 4111–4122. [Google Scholar] [CrossRef] [PubMed]
- Lüthje, P.; von Köckritz-Blickwede, M.; Schwarz, S. Identification and Characterization of Nine Novel Types of Small Staphylococcal Plasmids Carrying the Lincosamide Nucleotidyltransferase Gene Lnu(A). J. Antimicrob. Chemother. 2007, 59, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Del Grosso, M.; Iannelli, F.; Messina, C.; Santagati, M.; Petrosillo, N.; Stefani, S.; Pozzi, G.; Pantosti, A. Macrolide Efflux Genes Mef (A) and Mef (E) Are Carried by Different Genetic Elements in Streptococcus Pneumoniae. J. Clin. Microbiol. 2002, 40, 774–778. [Google Scholar] [CrossRef] [PubMed]
- Iannelli, F.; Santoro, F.; Santagati, M.; Docquier, J.-D.; Lazzeri, E.; Pastore, G.; Cassone, M.; Oggioni, M.R.; Rossolini, G.M.; Stefani, S.; et al. Type M Resistance to Macrolides Is Due to a Two-Gene Efflux Transport System of the ATP-Binding Cassette (ABC) Superfamily. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Bharatham, N.; Bhowmik, P.; Aoki, M.; Okada, U.; Sharma, S.; Yamashita, E.; Shanbhag, A.P.; Rajagopal, S.; Thomas, T.; Sarma, M.; et al. Structure and Function Relationship of OqxB Efflux Pump from Klebsiella Pneumoniae. Nat. Commun. 2021, 12. [Google Scholar] [CrossRef] [PubMed]
- Allignet, J.; Loncle, V.; Simenel, C.; Delepierre, M.; El Solh, N. Sequence of a Staphylococcal Gene, Vat, Encoding an Acetyltransferase Inactivating the A-Type Compounds of Virginiamycin-like Antibiotics. Gene 1993, 130, 91–98. [Google Scholar] [CrossRef]
- Garcillán-Barcia, M.P.; Redondo-Salvo, S.; de la Cruz, F. Plasmid Classifications. Plasmid 2023, 126, 102684. [Google Scholar] [CrossRef]
- Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard-Tenth Edition Available online:. Available online: https://clsi.org/media/1632/m07a10_sample.pdf (accessed on 15 October 2023).
- Amann, R.I.; Ludwig, W.; Schleifer, K.H. Phylogenetic Identification and in Situ Detection of Individual Microbial Cells without Cultivation. Microbiol. Rev. 1995, 59, 143–169. [Google Scholar] [CrossRef] [PubMed]
- Vaz-Moreira, I.; Egas, C.; Nunes, O.C.; Manaia, C.M. Bacterial Diversity from the Source to the Tap: A Comparative Study Based on 16S RRNA Gene-DGGE and Culture-Dependent Methods. FEMS Microbiol. Ecol. 2013, 83, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Miłobedzka, A.; Ferreira, C.; Vaz-Moreira, I.; Calderón-Franco, D.; Gorecki, A.; Purkrtova, S.; Bartacek, J.; Dziewit, L.; Singleton, C.M.; Nielsen, P.H.; et al. Monitoring Antibiotic Resistance Genes in Wastewater Environments: The Challenges of Filling a Gap in the One-Health Cycle. J. Hazard. Mater. 2022, 424, 127407. [Google Scholar] [CrossRef] [PubMed]
- Gandra, S.; Alvarez-Uria, G.; Turner, P.; Joshi, J.; Limmathurotsakul, D.; van Doorn, H.R. Antimicrobial Resistance Surveillance in Low- and Middle-Income Countries: Progress and Challenges in Eight South Asian and Southeast Asian Countries. Clin. Microbiol. Rev. 2020, 33. [Google Scholar] [CrossRef] [PubMed]
- George, A. Antimicrobial Resistance (AMR) in the Food Chain: Trade, One Health and Codex. Trop. Med. Infect. Dis. 2019, 4, 54. [Google Scholar] [CrossRef] [PubMed]
- Hendriksen, R.S.; Bortolaia, V.; Tate, H.; Tyson, G.H.; Aarestrup, F.M.; McDermott, P.F. Using Genomics to Track Global Antimicrobial Resistance. Front. Public Health 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Munk, P.; Knudsen, B.E.; Lukjancenko, O.; Duarte, A.S.R.; Van Gompel, L.; Luiken, R.E.C.; Smit, L.A.M.; Schmitt, H.; Garcia, A.D.; Hansen, R.B.; et al. Abundance and Diversity of the Faecal Resistome in Slaughter Pigs and Broilers in Nine European Countries. Nat. Microbiol. 2018, 3, 898–908. [Google Scholar] [CrossRef] [PubMed]
- Waseem, H.; Williams, M.R.; Stedtfeld, T.; Chai, B.; Stedtfeld, R.D.; Cole, J.R.; Tiedje, J.M.; Hashsham, S.A. Virulence Factor Activity Relationships (VFARs): A Bioinformatics Perspective. Environ. Sci. Process. Impacts 2017, 19, 247–260. [Google Scholar] [CrossRef]
- <i>The New Science of, Metagenomics</i>; National Academies Press: Washington, D.C. 45. The New Science of Metagenomics, W: Press, 2007. [Google Scholar]
- Che, Y.; Xia, Y.; Liu, L.; Li, A.-D.; Yang, Y.; Zhang, T. Mobile Antibiotic Resistome in Wastewater Treatment Plants Revealed by Nanopore Metagenomic Sequencing. Microbiome 2019, 7. [Google Scholar] [CrossRef]
- Hiraoka, S.; Machiyama, A.; Ijichi, M.; Inoue, K.; Oshima, K.; Hattori, M.; Yoshizawa, S.; Kogure, K.; Iwasaki, W. Genomic and Metagenomic Analysis of Microbes in a Soil Environment Affected by the 2011 Great East Japan Earthquake Tsunami. BMC Genomics 2016, 17. [Google Scholar] [CrossRef]
- Huson, D.H.; Albrecht, B.; Bağcı, C.; Bessarab, I.; Górska, A.; Jolic, D.; Williams, R.B.H. MEGAN-LR: New Algorithms Allow Accurate Binning and Easy Interactive Exploration of Metagenomic Long Reads and Contigs. Biol. Direct 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y. , Liu, R., and Shang, R GLOBMAP SWF: A Global Annual Surface Water Cover Frequency Dataset during 2000–2020. Earth Syst. Sci. Data 2020, 4505–4523. [Google Scholar]
- Zieliński, W.; Hubeny, J.; Buta-Hubeny, M.; Rolbiecki, D.; Harnisz, M.; Paukszto, Ł.; Korzeniewska, E. Metagenomics Analysis of Probable Transmission of Determinants of Antibiotic Resistance from Wastewater to the Environment - A Case Study. Sci. Total Environ. 2022, 827, 154354. [Google Scholar] [CrossRef]
- Berruti, I.; Nahim-Granados, S.; Abeledo-Lameiro, M.J.; Oller, I.; Polo-López, M.I. Peroxymonosulfate/Solar Process for Urban Wastewater Purification at a Pilot Plant Scale: A Techno-Economic Assessment. Sci. Total Environ. 2023, 881, 163407. [Google Scholar] [CrossRef]
- Islam, M.S.; Rahman, A.M.M.T.; Hassan, J.; Rahman, M.T. Extended-Spectrum Beta-Lactamase in Escherichia Coli Isolated from Humans, Animals, and Environments in Bangladesh: A One Health Perspective Systematic Review and Meta-Analysis. One Health 2023, 16, 100526. [Google Scholar] [CrossRef] [PubMed]
- Yoneda, I.; Rozanah, U.N.; Nishiyama, M.; Mith, H.; Watanabe, T. Detection and Genetic Analysis of Escherichia Coli from Tonle Sap Lake and Its Tributaries in Cambodia: Spatial Distribution, Seasonal Variation, Pathogenicity, and Antimicrobial Resistance. Environ. Pollut. 2022, 315, 120406. [Google Scholar] [CrossRef]
- Sorlí, L.; Luque, S.; Li, J.; Campillo, N.; Danés, M.; Montero, M.; Segura, C.; Grau, S.; Horcajada, J.P. Colistin for the Treatment of Urinary Tract Infections Caused by Extremely Drug-Resistant Pseudomonas Aeruginosa: Dose Is Critical. J. Infect. 2019, 79, 253–261. [Google Scholar] [CrossRef]
- Tanita, M.T.; Carrilho, C.M.D. de M.; Garcia, J.P.; Festti, J.; Cardoso, L.T.Q.; Grion, C.M.C. Parenteral Colistin for the Treatment of Severe Infections: A Single Center Experience. Rev Bras Ter Intensiva 2013, 25, 297–305. [Google Scholar] [CrossRef]
- Falagas, M.E.; Kasiakou, S.K. Colistin: The Revival of Polymyxins for the Management of Multidrug-Resistant Gram-Negative Bacterial Infections. Clin. Infect. Dis. 2005, 40, 1333–1341. [Google Scholar] [CrossRef] [PubMed]
- Snyman, Y.; Whitelaw, A.C.; Barnes, J.M.; Maloba, M.R.B.; Newton-Foot, M. Characterisation of Mobile Colistin Resistance Genes (Mcr-3 and Mcr-5) in River and Storm Water in Regions of the Western Cape of South Africa. Antimicrob. Resist. Infect. Control 2021, 10, 96. [Google Scholar] [CrossRef]
- Gomi, R.; Matsuda, T.; Yamamoto, M.; Tanaka, M.; Ichiyama, S.; Yoneda, M.; Matsumura, Y. Molecular Characterization of a Multidrug-Resistant IncF Plasmid Carrying Mcr-3.1 in an Escherichia Coli Sequence Type 393 Strain of Wastewater Origin. Int. J. Antimicrob. Agents 2019, 54, 524–526. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhang, Y.; Yang, M.; Tian, Z.; Ren, L.; Zhang, S. Abundance and Distribution of Tetracycline Resistance Genes and Mobile Elements in an Oxytetracycline Production Wastewater Treatment System. Environ. Sci. Technol. 2012, 46, 7551–7557. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; He, X.; Bu, Y.; Shi, P.; Miao, Y.; Zhou, H.; Shan, Z.; Zhang, X.-X. Environmental Fate of Tetracycline Resistance Genes Originating from Swine Feedlots in River Water. J. Environ. Sci. Health B 2014, 49, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Engemann, C.A.; Keen, P.L.; Knapp, C.W.; Hall, K.J.; Graham, D.W. Fate of Tetracycline Resistance Genes in Aquatic Systems: Migration from the Water Column to Peripheral Biofilms. Environ. Sci. Technol. 2008, 42, 5131–5136. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, G.; Huys, G.; Swings, J.; McGann, P.; Hiney, M.; Smith, P.; Pickup, R.W. Distribution of Oxytetracycline Resistance Plasmids between Aeromonads in Hospital and Aquaculture Environments: Implication of Tn1721 in Dissemination of the Tetracycline Resistance Determinant Tet A. Appl. Environ. Microbiol. 2000, 66, 3883–3890. [Google Scholar] [CrossRef] [PubMed]
- Henriques, I.S.; Fonseca, F.; Alves, A.; Saavedra, M.J.; Correia, A. Tetracycline-Resistance Genes in Gram-Negative Isolates from Estuarine Waters. Lett. Appl. Microbiol. 2008, 47, 526–533. [Google Scholar] [CrossRef]
- Semestre-Año, P. INFORME SOBRE USO DE ANTIMICROBIANOS EN LA SALMONICULTURA NACIONAL Available online:. Available online: http://www.sernapesca.cl/sites/default/files/informe_sobre_el_uso_de_antimicrobianos_en_la_salmonicultura_nacional_-_primer_semestre_-_ano_2022.pdf (accessed on 27 May 2024).
- Ben Maamar, S.; Glawe, A.J.; Brown, T.K.; Hellgeth, N.; Hu, J.; Wang, J.-P.; Huttenhower, C.; Hartmann, E.M. Mobilizable Antibiotic Resistance Genes Are Present in Dust Microbial Communities. PLoS Pathog. 2020, 16, e1008211. [Google Scholar] [CrossRef]
- Pruden, A.; Larsson, D.G.J.; Amézquita, A.; Collignon, P.; Brandt, K.K.; Graham, D.W.; Lazorchak, J.M.; Suzuki, S.; Silley, P.; Snape, J.R.; et al. Management Options for Reducing the Release of Antibiotics and Antibiotic Resistance Genes to the Environment. Environ. Health Perspect. 2013, 121, 878–885. [Google Scholar] [CrossRef]
- Katusiime, J.; Schütt, B. Integrated Water Resources Management Approaches to Improve Water Resources Governance. Water (Basel) 2020, 12, 3424. [Google Scholar] [CrossRef]
- Kovalova, L.; Siegrist, H.; Singer, H.; Wittmer, A.; McArdell, C.S. Hospital Wastewater Treatment by Membrane Bioreactor: Performance and Efficiency for Organic Micropollutant Elimination. Environ. Sci. Technol. 2012, 46, 1536–1545. [Google Scholar] [CrossRef]
- Manyi-Loh, C.; Mamphweli, S.; Meyer, E.; Okoh, A. Antibiotic Use in Agriculture and Its Consequential Resistance in Environmental Sources: Potential Public Health Implications. Molecules 2018, 23, 795. [Google Scholar] [CrossRef] [PubMed]
- Millanao, A.R.; Barrientos-Schaffeld, C.; Siegel-Tike, C.D.; Tomova, A.; Ivanova, L.; Godfrey, H.P.; Dölz, H.J.; Buschmann, A.H.; Cabello, F.C. Resistencia a los antimicrobianos en Chile y el paradigma de Una Salud: manejando los riesgos para la salud pública humana y animal resultante del uso de antimicrobianos en la acuicultura del salmón y en medicina. Rev. Chilena Infectol. 2018, 35. [Google Scholar] [CrossRef] [PubMed]
- Hammerum, A.M.; Heuer, O.E.; Emborg, H.-D.; Bagger-Skjøt, L.; Jensen, V.F.; Rogues, A.-M.; Skov, R.L.; Agersø, Y.; Brandt, C.T.; Seyfarth, A.M.; et al. Danish Integrated Antimicrobial Resistance Monitoring and Research Program. Emerg. Infect. Dis. 2007, 13, 1633–1639. [Google Scholar] [CrossRef] [PubMed]
- Amjad, U.Q.; Dalcanale, F.; Kayser, G.; Bentley, P.; Bartram, J. Evidence-Based Decision-Making on Water Quality in Domestic Water Supply in Malawi, Ecuador, and Brazil. Water Policy 2018, 20, 530–545. [Google Scholar] [CrossRef] [PubMed]
- Zulkifli, C.Z.; Garfan, S.; Talal, M.; Alamoodi, A.H.; Alamleh, A.; Ahmaro, I.Y.Y.; Sulaiman, S.; Ibrahim, A.B.; Zaidan, B.B.; Ismail, A.R.; et al. IoT-Based Water Monitoring Systems: A Systematic Review. Water (Basel) 2022, 14, 3621. [Google Scholar] [CrossRef]
- Saini, P.; Bandsode, V.; Singh, A.; Mendem, S.K.; Semmler, T.; Alam, M.; Ahmed, N. Genomic Insights into Virulence, Antimicrobial Resistance, and Adaptation Acumen of Escherichia Coli Isolated from an Urban Environment. MBio 2024, 15. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Cortés, G.; Arenas-Hernández, M.M.P.; Ballesteros-Monrreal, M.G.; Rocha-Gracia, R. del C.; Barrios-Villa, E. Editorial: Epidemiology of Antimicrobial Resistance and Virulence Factors of Emerging and Re-Emerging Bacteria. Front. Cell. Infect. Microbiol. 2024, 14. [Google Scholar] [CrossRef] [PubMed]
- Nijsingh, N.; Munthe, C.; Larsson, D.G.J. Managing Pollution from Antibiotics Manufacturing: Charting Actors, Incentives and Disincentives. Environ. Health 2019, 18. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast Gapped-Read Alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Comeau, A.M.; Douglas, G.M.; Langille, M.G.I. Microbiome Helper: A Custom and Streamlined Workflow for Microbiome Research. mSystems 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Liu, C.-M.; Luo, R.; Sadakane, K.; Lam, T.-W. MEGAHIT: An Ultra-Fast Single-Node Solution for Large and Complex Metagenomics Assembly via Succinct de Bruijn Graph. Bioinformatics 2015, 31, 1674–1676. [Google Scholar] [CrossRef] [PubMed]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef] [PubMed]
- Wood, D.E.; Lu, J.; Langmead, B. Improved Metagenomic Analysis with Kraken 2. Genome Biol. 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Abricate: :Mag_right: Mass Screening of Contigs for Antimicrobial and Virulence Genes; Github;
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of Acquired Antimicrobial Resistance Genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Giménez, M.; Ferrés, I.; Iraola, G. Improved Detection and Classification of Plasmids from Circularized and Fragmented Assemblies. bioRxiv 2022.
- R Core Team (2021) R A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna. - References - Scientific Research Publishing Available online:. Available online: https://www.scirp.org/reference/referencespapers?referenceid=3131254 (accessed on 27 May 2024).
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS One 2013, 8, e61217. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2; Springer International Publishing: Cham, 2016; ISBN 9783319242750. [Google Scholar]
- Hudzicki, J. Kirby-Bauer Disk Diffusion Susceptibility Test Protocol Available online:. Available online: https://asm.org/getattachment/2594ce26-bd44-47f6-8287-0657aa9185ad/Kirby-Bauer-Disk-Diffusion-Susceptibility-Test-Protocol-pdf.pdf (accessed on 27 May 2024).
- PATEL, J.B. , COCKERILL, F.R., & BRADFORD, P.A. PERFORMANCE STANDARDS FOR ANTIMICROBIAL SUSCEPTIBILITY TESTING: TWENTY-FIFTH INFORMATIONAL SUPPLEMENT Clinical & Laboratory Standards Institute, 2015.
Figure 1.
Taxonomy abundance at species level represented as stacked bar plot of each sample site of Puerto Varas shore. Three points of the Llanquihue lake beach were sampled (PV1, PV2, PV3) and one point (EC) 200 m far from the coast was sampled, at surface (SUP) and 5 meters submerged from the water column (5m). The coloration pattern of each bar depicts the percentage of abundance of each taxonomy, showing the microbial community structure.
Figure 1.
Taxonomy abundance at species level represented as stacked bar plot of each sample site of Puerto Varas shore. Three points of the Llanquihue lake beach were sampled (PV1, PV2, PV3) and one point (EC) 200 m far from the coast was sampled, at surface (SUP) and 5 meters submerged from the water column (5m). The coloration pattern of each bar depicts the percentage of abundance of each taxonomy, showing the microbial community structure.
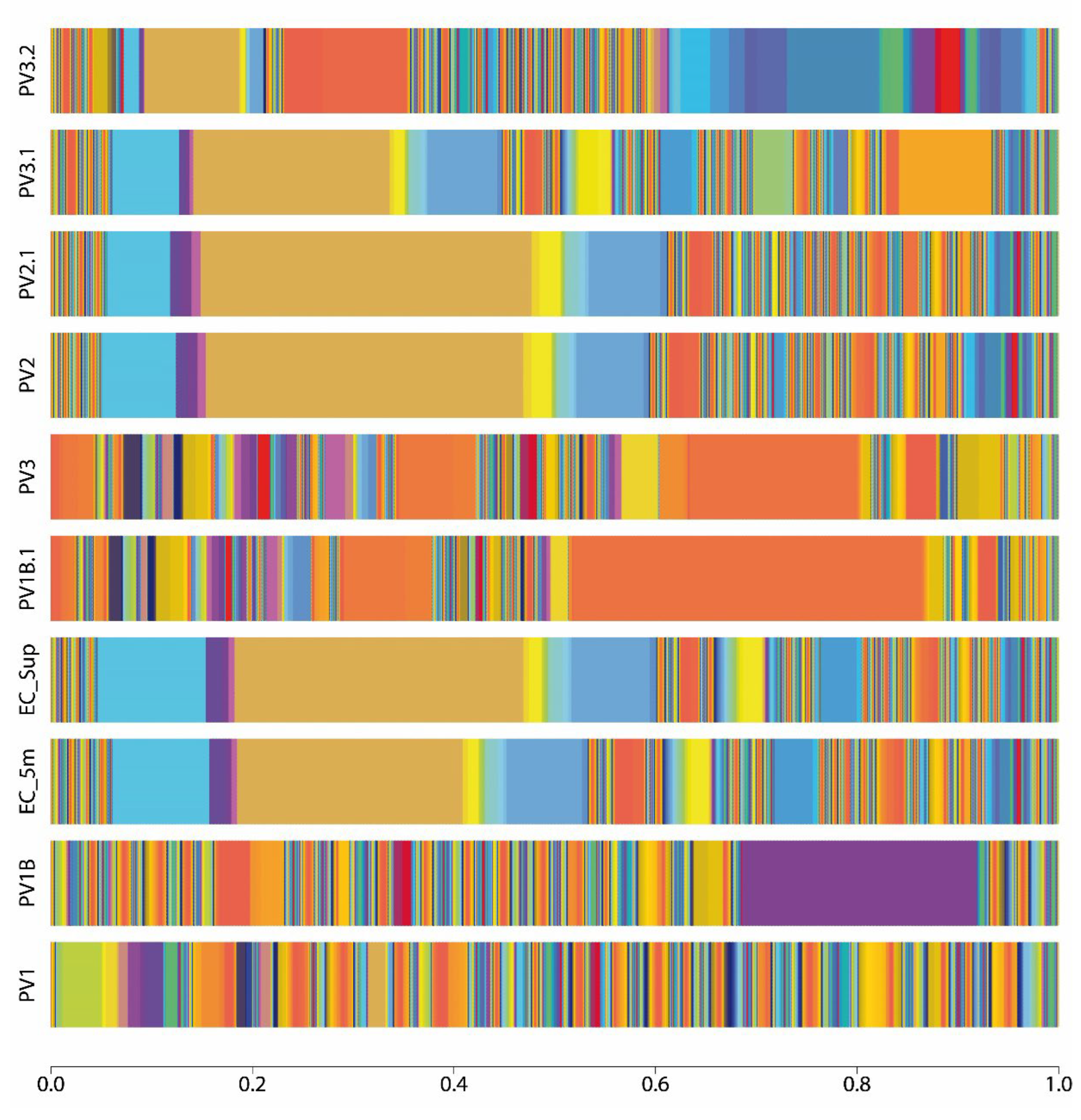
Figure 2.
Phylogenetic trees reveal the intricate web of relationships among microbial phyla. Major phyla such as Verrumicrobiota, Fusobacteria, Proteobacteria, Firmicutes, Actinobacteria, and Bacteroidetes emerge as prominent branches, highlighting their importance in processes such as fermentation and natural decomposition.
Figure 2.
Phylogenetic trees reveal the intricate web of relationships among microbial phyla. Major phyla such as Verrumicrobiota, Fusobacteria, Proteobacteria, Firmicutes, Actinobacteria, and Bacteroidetes emerge as prominent branches, highlighting their importance in processes such as fermentation and natural decomposition.

Figure 3.
AMR genes identified on environmental DNA recovered from Llanquihue lake. The heatmap represents the genes identified grouped according to their antimicrobial resistance family which belongs to.
Figure 3.
AMR genes identified on environmental DNA recovered from Llanquihue lake. The heatmap represents the genes identified grouped according to their antimicrobial resistance family which belongs to.
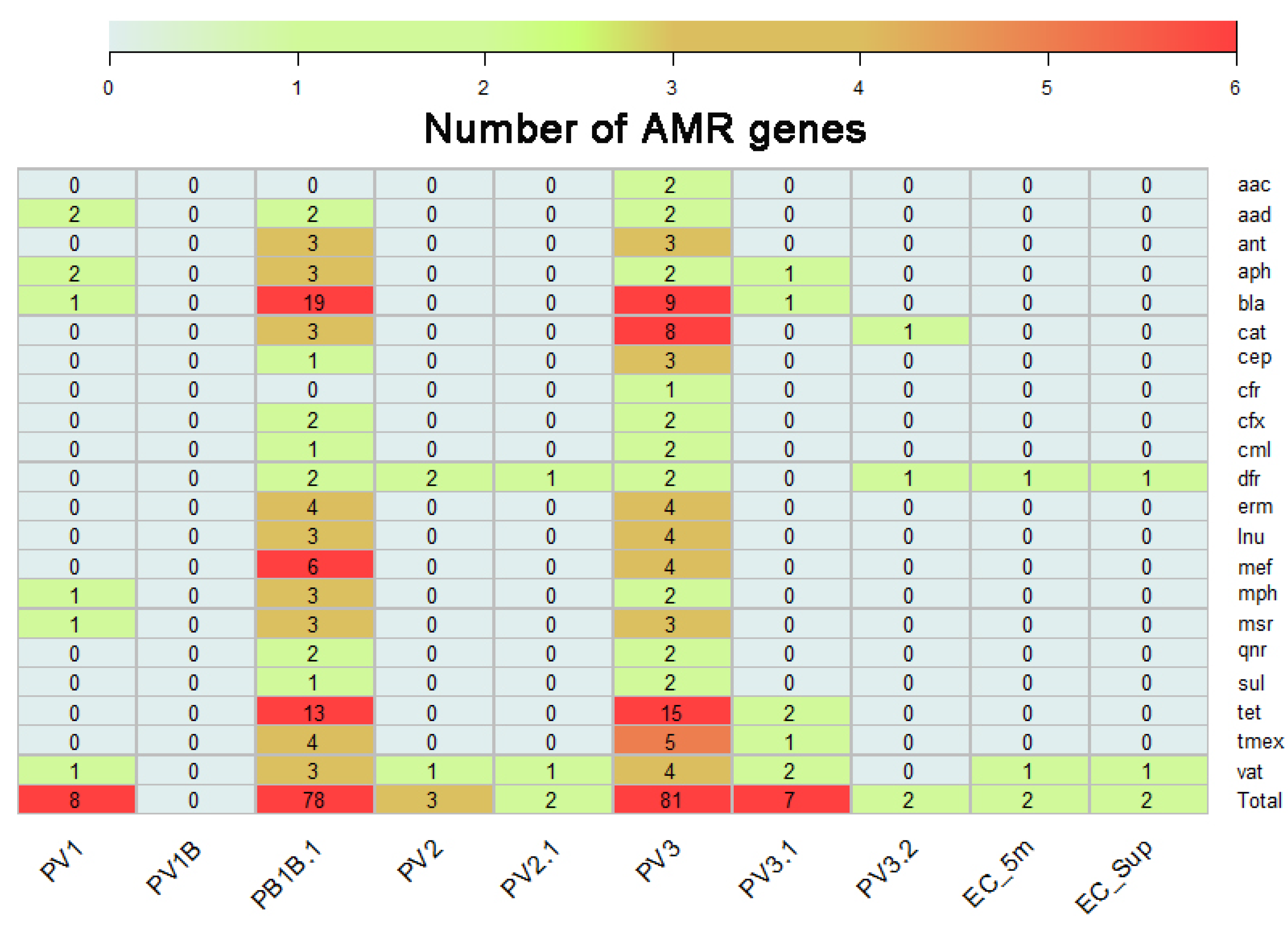

Table 1.
Summary of antibiotic resistance genes found and associated resistance mechanisms.
| Family gen | Antibiotic family | Drug example | Resistance mechanism | References |
|---|---|---|---|---|
| bla | Betalactamics | Imipenem | Antibiotic Inactivation | [22] |
| cat | Phenicols | Chloramphenicol | Antibiotic Inactivation | [27] |
| cfx | Cefamycins | Cefoxitin | Antibiotic Inactivation | [28] |
| dfr | Diaminopyridines | Trimetoprim | Target modification | [24] |
| erm | Macrolides | Erythromycin | Target modification | [29] |
| Inu | Lincosamides | Clindamicin | Antibiotic Inactivation | [30] |
| mef | Macrolides | Erythromycin | Efflux Pump | [31] |
| msr | Macrolides | Erythromycin | Efflux Pump | [32] |
| oqx | Multi-Drug | Multi-Drug | Efflux Pump | [33] |
| tet | Tetracyclines | Doxicycline | Efflux Pump, Target Modification, Antibiotic Inactivation | [21] |
| vat | Streptogramins | Virginiamycin | Antibiotic Inactivation | [34] |
Table 2.
Susceptibility studies on bacteria isolated from Llanquihue lake.
| nº | Species | Antibiotic drug tested | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| cefotaxime | Ampicillin / sulbactam |
Sulfamethoxosazole /trimethoprim |
gentamicin | ciprofloxacin | imipenem | ||||||||
| IZD(mm) | Int | IZD(mm) | Int | IZD(mm) | Int | IZD(mm) | Int | IZD(mm) | Int | IZD(mm) | Int | ||
| 23 | Citrobacter freundii | 36,3±0,6 | S | 19,7±0,6 | S | 26,0±1,0 | S | 19,3±0,6 | S | 39,0±1,0 | S | 28,3±1,5 | S |
| 55 | Citrobacter gillenii | 36,3±1,5 | S | 32,0±1,0 | S | 24,3±0,6 | S | 24,0±1,0 | S | 44,3±1,2 | S | 27,3±0,6 | S |
| 62 | Citrobacter gillenii | 32,7±1,2 | S | 38,0±2,0 | S | 22,7±0,6 | S | 20,7±0,6 | S | 50,0±1,0 | S | 37,0±1,7 | S |
| 2 | Enterobacter absuriae | 34,7±2,9 | S | 29,7±0,6 | S | 33,3±0,6 | S | 19,3±1,5 | S | 36,3±0,6 | S | 27,3±0,6 | S |
| 14 | Enterobacter cloacae | 35,7±1,2 | S | 30,0±0,0 | S | 32,7±1,2 | S | 19,7±1,2 | S | 40,3±0,6 | S | 31,0±1,0 | S |
| 39 | Enterobacter ludwigii | 34,0±1,7 | S | 32,3±1,2 | S | 31,3±0,6 | S | 23,3±0,6 | S | 47,3±0,6 | S | 33,0±1,0 | S |
| 41 | Enterobacter ludwigii | 35,0±2,0 | S | 34,7±0,6 | S | 35,7±0,6 | S | 24,0±0,0 | S | 49,0±1,0 | S | 35,3±1,2 | S |
| 21 | Escherichia coli | 36,7±0,6 | S | 19,7±0,6 | S | 24,7±0,6 | S | 24,3±0,6 | S | 34,0±0,0 | S | 31,0±1,0 | S |
| 22 | Escherichia coli | 35,3±06 | S | 20,3±1,2 | S | 26,3±0,6 | S | 20,3±0,6 | S | 41,3±1,2 | S | 31,3±1,5 | S |
| 26 | Escherichia coli | 37,3±0,6 | S | 23,3±0,6 | S | 28,3±2,1 | S | 22,0±1,0 | S | 38,3±0,0 | S | 29,3±1,5 | S |
| 27 | Escherichia coli | 34,0±1,0 | S | 20,0±0,0 | S | 27,0±1,0 | S | 19,7±2,1 | S | 35,3±0,6 | S | 31,3±1,5 | S |
| 28 | Escherichia coli | 33,0±0,0 | S | 21,3±1,5 | S | 27,3±0,6 | S | 23,7±0,6 | S | 41,0±1,0 | S | 32,3±1,5 | S |
| 42 | Escherichia coli | 36,7±1,5 | S | 20,3±1,2 | S | 24,7±0,6 | S | 19,0±1,0 | S | 38,3±0,6 | S | 30,3±2,3 | S |
| 3 | Rahnella aquatilis | 25,3±3,1 | S | 18,7±1,2 | S | 21,3±1,5 | S | 15,3±0,6 | S | 25,3±0,6 | S | 23,3±0,6 | S |
| ATCC 25922 | Escherichia coli | 31,3±0,6 | ✓ | 20,7±0,6 | ✓ | 24,7±0,6 | ✓ | 24,3±0,6 | ✓ | 45,0±0,0 | ✓ | 34,7±0,6 | ✓ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



