
Submitted:
04 June 2024
Posted:
05 June 2024
You are already at the latest version
Abstract
The mystery phenomenon of fungal bioluminescence has always captivated human curiosity, although more attempts need to be made, particularly to bring the new species to the surface. In this study, we devoted considerable space to the taxonomy of these fungi while discussing their biogeography, evolution, bioluminescence mechanism, and potential ecological roles. We provide a detailed explanation of how these fungi produce light, including the role of the enzyme luciferase and luciferin. We also discussed their applications. Here, we presented an updated list of 122 bioluminescent fungi identified from five distinct evolutionary lineages viz. Armillaria, Eoscyphella, Lucentipes, Mycenoid, and Omphalotus worldwide, mainly in tropical and subtropical areas. The bioluminescent fungi descended from the last common ancestor of the mycenoid and the marasmioid clades of Agaricales, which have been maintained for at least 160 million years of evolution. We highlighted the need for further research to understand the ecological role of bioluminescent fungi. Applying bioluminescent fungi in various areas (e.g., environmental and medicinal) demonstrates their validity.
Keywords:
Biodiversity
; Bioluminescence
; Distribution
; Luciferin-luciferase
; Mushrooms
1. Introduction
Mother nature has created a remarkable phenomenon called bioluminescence. Bio (in Greek) stands for life, while lumen (in Latin) means light. Bioluminescence is the emission of light from living organisms, which is also a type of chemiluminescence that produces mild thermal radiation due to chemical reactions accompanied by energy stores, enzymes, substrates, and other molecules [1,2]. It is thought to represent the distribution of bioluminescent organisms throughout the Tree of Life across approximately 17 phyla and over 700 genera [3,4].
Paradoxically, bioluminescence occurs independently of at least 94 origins in the Tree of Life [5]. Although 40 distinct bioluminescent systems exist in nature, only 11 pairs of luciferin-luciferase have been thoroughly studied, understood, and practically applied [2,6]. However, more have yet to be identified.
Looking back at the historical records of bioluminescence, Harvey's early documents traced bioluminescent organisms back to ancient Greece and Rome [7,8]. Aristotle (384–322 BCE) stood out as a pioneer, making groundbreaking observations and recognizing the self-luminosity of these organisms. His detailed accounts included observations on dead fish, bioluminescent bacteria in fish and flesh, and the bioluminescence of fireflies and glow worms.
Advancing through time, Pliny the Elder's "Naturalis Historia" (23–79 CE) provided the first specific and comprehensive record of bioluminescent organisms. Despite lacking independent verification, beliefs in the existence of bioluminescent birds persisted for over a thousand years. The Dark Ages in Europe (500 CE) witnessed a scientific slowdown, but literature described ocean "phosphorescence" and mentioned the Chinese "candle fly."
Albertus Magnus, the first naturalist, brought the cataloging of bioluminescent species in the 12th Century for the first time. The Renaissance period saw a revival of learning, with reports of the "burning sea” and mysterious lights at sea by Christopher Columbus. Oviedo (1478–1557) documented bioluminescent organisms and Sir Frances Drake observed tropical fireflies. Conrad Gestner, in the 16th Century, contributed to bioluminescence studies.
The 17th Century emerged as a scientific revolution, marked by three intellectual movements. Philosophers like Sir Francis Bacon and René Descartes laid foundational principles, leading to a surge in interest in luminescent phenomena. Robert Boyle [9] championed experimental observation, emphasizing the importance of interrogating nature through experimentation.
Bacon's scientific method, focused on meticulous observation, persisted in biology for over two centuries. Descartes proposed a deductive theory of light, contributing to understanding bioluminescent systems. The French school further elaborated on various bioluminescent phenomena. Changing theories of light, from the wave theory to the corpuscular theory, marked the late 17th century.
Notable figures, including Kircher and Bartholin, contributed to bioluminescence studies. Robert Boyle, recognized as the ‘Father of Modern Chemistry,’ conducted experiments revealing critical insights into the effects of air on bioluminescence. His role as a founder of the Royal Society and pioneering work advanced our understanding of bioluminescent phenomena.
Expanding our perspective globally, evidence of bioluminescent species in eastern countries, particularly China and Japan, became apparent. Records from China's Tang and Liang dynasties traced bioluminescence back centuries. The first appearance of bioluminescent fungi in Japanese literature was in ancient tales in Japan’s Heian Period (6–12th century) [10]. The 19th century brought a renewed focus, with significant contributions by Dubois and Harvey, shedding light on the mechanisms of bioluminescence. Since the 20th century, more precise identifications and research have been carried out thanks to methodological and technological advancements. The blooming of molecular approaches over the past 20 years has brought significant changes in fungal taxonomy, and several databases have been launched to the public, e.g., Index Fungorum Online [11] and MycoBank [12]. Amsterdam Declaration on Fungal Nomenclature “One fungus = one name” [13], ITS designated as a universal barcode for fungi and NCBI RefSeq Targeted Loci project for ITS initiated [14,15], Obligate registration for valid publication of new fungal names [16]. Remarkably, the progression of molecular phylogeny has unveiled an unprecedented spectrum of fungal diversity. Incorporating culture-independent techniques, notably high-throughput amplicon sequencing, has substantially escalated the enumeration of fungal operational taxonomic units. Further, throughout the last two decades, numerous innovative taxa encompassing novel divisions, classes, orders, and families have been methodically established. Molecular phylogeny, in particular, has been instrumental in discerning myriad cryptic species, thereby advancing our comprehension of the intricate fungal panorama. Correspondingly, numbers of bioluminescent mushrooms are also showing up.
Today, bioluminescent fungi (e.g., Armillaria, Mycena, Roridomyces) [17,18,19] and bacteria (e.g., Photobacterium, Vibrio) [20,21] are the most recognized microbes, while animals such as fishes (e.g., Lanternfish) [22], and insects (e.g., Cheguevaria, Photuris) are also among the popularly studied groups [23,24]. Owing to the continuous identification of bioluminescent fungi creates enormous attention among the many research groups around the world and has led to the finding of many novel species [25,26], also becoming the driving aspect of this review.
Bioluminescent fungi, on the other word— glowing fungi can be recognized in nature due to emitting a yellowish-green light, which is visible at nightfall. Nevertheless, in the dense forest without sunshine, it could be a better place to see them with the naked eye at nighttime. At present, this mysterious group of organisms has been demolished and resolved the puzzle behind their peculiar nature, however, not completely. Balance work between the biochemists and the taxonomists never seems to be in line, with considerable weightage held by the taxonomists. In light of recent taxonomic studies, it is clear that many works have been attempted on the taxonomy and also the evolution of bioluminescent mushrooms; as a result, more than 40 bioluminescent mushroom species have been reported in the last decade [2,18,27,28,29,30].
In another way, as previously highlighted, the curiosity about these creatures is still there; for instance, scientists attempted to make mountains out of molehills to find out the perception of the bioluminescence in fungi. The main question is why this bioluminescence, and whether this is advantageous to them or not [10,31]. Outcomes are not yet proven; several hypotheses came to the battlefield [32,33,34]. One such is that bioluminescence in fungi is crucial in attracting diverse dispersal agents, particularly insects, which many fungi depend on for spore dispersal. The emitted light from bioluminescent fungi draws insects towards the fruiting bodies, effectively aiding in the dispersal of fungal spores as these insects carry them to new locations. Furthermore, certain bioluminescent fungi employ their light to dissuade potential predators. The intense glow serves as a warning, indicating the existence of toxins or an unpalatable nature, effectively discouraging herbivores from consuming them. This defensive strategy enhances the fungi's chances of survival and reproduction by preventing them from being consumed [35,36]. Nevertheless, this hypothesis lacks greater persuasiveness in species where bioluminescence emanates from only stipe and/or mycelium [36,37].
In contrast, others didn’t see this as anything more, and it has considered the process merely a metabolic by-product and has no ecological benefits [31,38]. This review delves into the recent upsurge of interest in these enigmatic life forms by expanding upon the extensive corpus of research about bioluminescent organisms and specifically emphasizing the burgeoning domain of bioluminescent fungi.
2. Diversity and Distribution of Bioluminescent Fungi
A team of fungal experts recently assessed the fungal diversity in the world using four main academic pathways viz. scaling laws, fungus: plant ratios, actual versus previously known number of species, and DNA-based studies; according to them, there are likely to be 2–3 million species of fungi, with a best estimate of 2.5 million [39]. Nevertheless, the findings of those magnificent organisms are far behind; as of 2023, around 155,000 species only have been recorded and described by taxonomists, which is comparatively lower than other special types of fungi. For example, more than 800 genera of endophytic fungi [40] and 50,000 species of mycorrhizal fungi [41] have been recognized. Currently, over 2,500 species of novel fungi are named yearly; if this continues at the current rate, it will require 750–1,000 years to name the remaining unknown species [39]. Moving to our targeted group —bioluminescent fungi- to date, 122 species of bioluminescent fungi have been reported worldwide [2,18,29]. Table 1 presents a list of those identified bioluminescent fungi. The different parts of bioluminescence fungi species were compared; 37 species were reported that the fruiting body and mycelium both are bioluminescence (30.3%), 38 species are undetermined mycelium bioluminescence present (44.7% of known bioluminescence fruiting body), 36 species are only mycelium present bioluminescence, one species is undetermined fruiting body bioluminescence present, and 48 species are only fruiting body, among of them, 14 species don’t specify where they emit light from (Figure 1), while Figure 2 shows the global distribution of those all the bioluminescent fungi. Despite the regional study bias, according to the available findings, bioluminescent fungi are mainly documented in Asia, North America, and South America.
It is also worth mentioning that all bioluminescent fungi records belong to the Basidiomycota division, except for Xylaria hypoxylon (L.) Grev., which falls under the Ascomycota [58,73]. As shown in Figure 3, five lineages comprise all known species of bioluminescent fungi. They are part of the Mycenoid (92 species), Armillaria (13 species), Omphalotus (14 species), Lucentipes (two species), and Eoscyphella (one species) lineages [49].
Note that Mycena is the main genus that exhibits bioluminescence in fungi, and it is distributed worldwide [43,44,74]. Furthermore, accounting for the species level, for example, Mycena chlorophos, M. inclinata, and Neonothopanus nambi show a wide distribution worldwide [4,58,75]. While species like Favolaschia xtbgensis and Roridomyces viridiluminus show a restricted habitat, particularly in some places in Southwestern China (Figure 4) [36,50].
In line with this, Gerronema viridilucens was reported only in Brazil [76], and Neonothopanus gardneri from the states of Maranhão, Piauí, and Tocantins in Brazil [77]. Interestingly, some species like Pannellus stipticus are naturally found in different countries, however, they didn’t show bioluminescence in all the recorded places. For instance, P. stipticus shows the bioluminescence that grows in North America, though not in Eurasian [78]. In addition, Armillaria mellea, Mycena chlorophos, M. deeptha, Nothopanus eugrammus, Omphalotus olearius, O. olivascens, and Roridomyces cf. phyllostachydis have been reported from India [18,61,79,80], while Filoboletus manipularis, Mycena chlorophos, and Panellus luxfilamentus have been found in Sri Lanka [58].
In an early study, Desjardin et al. [44] presented a revised list of bioluminescent fungi with 64 species in their distribution. Two years after their publication, Desjardin et al. [58] reported seven new species of luminescent fungi. Later, Aravindakshan et al. [61] and Shih et al. [81] reported two additional novel species from India and the Taiwan Province of China, respectively. Besides, Chew et al. [55,58] disclosed 15 bioluminescent fungi from Peninsular Malaysia, where eight were reported for the first time ever. Mihail [43] detected bioluminescent mycelia of five Armillaria species for the first time.
In a most recent study, Terashima et al. [52] identified another eight new species of glowing mushrooms from Japan, thus bringing the country’s total reported number up to 25 [10]. Seven species of bioluminescent fungi were recorded from the cloud forests in Mexico, where six species have been identified as a new species of Mycena whereas M. globulispora made a new distribution record for the country [63]. Besides, several other new species have been reported from Taiwan provinces (M. jingyinga, M. luguensis, and M. venus), and Yunnan provinces of China (Favolaschia xtbgensis, and Roridomyces viridiluminus) and India (R. phyllostachydis) [18,29,36,50].
3. Evolution and Mechanisms of Bioluminescent Fungi
3.1. Evolution
Understanding the development of bioluminescence in fungi is a challenging and fascinating subject. Bioluminescence is most likely the result of ancient beginnings, convergent evolution, and potentially horizontal gene transfer [82]. Numerous fungal lineages have distinctive characteristics that have recently been investigated concerning the genetic and environmental factors that have influenced them [82]. Nonetheless, two main hypotheses have been proposed to explain the scattered phylogenetic distribution and the lesser occurrence of bioluminescence in fungi. The summary of those hypotheses can be explained as genes associated with bioluminescence shared by the common ancestor were missing in some branches and multiple convergent evolutions of bioluminescence in fungi [2]. Like other bioluminescent organisms, bioluminescent fungi have independent evolutionary occurrences, converging multiple times. Genomic analysis shows that this fragmented phylogenetic position may be another case.
Oliveira et al. [17] revealed that the origin of fungal bioluminescence could be attributed to a single evolution ancestry, in which the evidence was demonstrated by cross-reactions between the luciferins and luciferases of distant lineages to yield light successfully.
Some recent studies conducted by Kotlobay et al. [28], and Ke et al. [82] concluded that the bioluminescence in fungi was descended from the last common ancestor of the mycenoid and the marasmioid clades of Agaricales that have been maintained for at least 160 million years of evolution. In recent surveys, Ke and colleagues echoed that view [37].
Ke et al. [82] investigated the evolutionary dynamics of the luciferase cluster. The study's results explained that the ancestral luciferase cluster carries the genes luciferase (Luz), hispidin-3-hydroxylase (H3H), cytochrome P450 (CYP450), hispidin synthase (HispS), and caffeylpyruvate hydrolase (CPH) on the same chromosome. This involves the recycling of oxyluciferin, except for the Omphalotus lineage.
Further studies revealed that gene clusters frequently undergo either deletions or retention due to differences in genomic plasticity, which explains the often loss of bioluminescence property of Mycenaean fungi [2,82,83]. The conservation of the gene cluster during the process of evolution signifies that, unlike other groups of bioluminescent organisms, bioluminescence evolves once in fungi with Luz, H3H, and HispS genes generated through gene duplications [28]. Besides, the species phylogenetic tree and reconstructed phylogenetic trees of Luz, H3H, and HispS genes of the family Agaricaceae reveal the evolution of bioluminescent cascades in fungi. The primary Luz enzyme formed through a gene duplication at the base of Agaricales, followed by the duplication of H3H and HipS a few million years later [28].
3.2. Mechanisms
As previously mentioned, bioluminescent fungi, including mushrooms, have been discovered worldwide in a wide range of terrestrial environments; nevertheless, fungal bioluminescence remains the least studied [84]. In general, bioluminescence occurs through the chemical oxidation of the luciferin, catalyzed by the luciferase enzyme in the presence of oxygen [85,86]. The molecular oxygen reacts with luciferin, forming a high-energy intermediary whose decomposition emits sufficient energy to generate the emitter “oxyluciferin” in the singlet, which is electronically excited. This excited metabolite's fluorescence property results in the emission of visible light used in nature for illumination [2,30,87].
First, it is interesting to understand how this bioluminescence mechanism has been revealed throughout history. In an early study, Dubois [88] used an in vitro luciferin/luciferase system with a mixture of heated substrate and cold enzyme-water extracts to demonstrate the first light emission experiment. In his experiment, he utilized extracts (cold and hot) from the light-emitting organs of the beetle Pyrophorus noctilucus. In this experimental setup, the cold extract process contained a heat-labile enzyme needed to emit light named luciferase. The hot extract was the thermo-stable fraction that was named luciferin. Further, it was determined that the luminescence of the mixture formed with two extracts resulted from the substrate/enzyme reaction [77].
In a study that supported Dubois [88], Airth and McElroy [89] used an in vitro setup made up of cold and hot extracts from bioluminescent fungi to confirm the role and nature of an enzymatic reaction. Later experiments by Airth and Foerster [90] explained that adding DPNH (the obsolete name for NADH) or NADPH to the cold and hot extracts activates light emission. Besides, the proteinaceous cold extracts could be separated into two fractions, a pellet (insoluble) and supernatant (soluble), by ultracentrifugation, which is necessary for the light emission in luminous fungi. Thus, essential enzymes in each fraction for light generation postulated a two-step mechanism of enzymatic reaction [90]. Note that basidiomycetous fungi emit a green light with maximum intensity in the 520–530 nm range [44]. Returning to Airth and Foerster's [90] study, they proposed the following two-step mechanism for fungal bioluminescence.

In the first step, Luciferin acts as an electron acceptor in the hot extract. Here, reductase presents in the supernatant, while luciferase resides in the pellet. During the first step, a dark reaction occurs between NAD(P)H, luciferin, and the soluble enzyme in the supernatant. In the second step, the reduced substrate luciferin reacts with molecular oxygen, catalyzed by the enzyme (luciferase) in the re-suspended pellet that produces visible light [90].
Those initial findings led to understanding the chemistry behind this scenario; however, along with the development of technologies, more questions were raised, such as the in-depth aspects of the specific roles of the enzymes in bioluminescence [44,91,92]. Oliveira and Stevani [92] attempted to find the answer using an enzyme-mediated reaction by mixing a hot extract containing heat-stable substrate/luciferin with a cold extract containing the enzyme luciferase. Later on, it was demonstrated that combinations of substrates/enzyme extract from mycelium of different bioluminescent species (Armillaria mellea, Gerronema viridilucens, Mycena luxperpetua, and Neonothopanus gardneri), and these results strongly suggest that all known bioluminescent fungi share similar types of luciferins/ luciferases in bioluminescent systems [17]. Purtov et al. [93] identified the structure of fungal luciferin and its precursor as 3-hydroxyhispidin and hispidin in extracts from four diverse genera of bioluminescent fungi viz. Armillaria borealis, Mycena citricolor, Neonothopanus nambi, and Panellus stipticus. Kaskova et al. [27] conducted an in-depth study of the mechanisms of fungal bioluminescence and color modulation and reported the structure of fungal oxyluciferin to investigate the mechanism of fungal bioluminescence. Hispidin is produced through the enzymatic activity of HispS. Subsequently, the resulting hispidin undergoes hydroxylation mediated by H3H, leading to the formation of 3-hydroxyhispidin, also known as fungal luciferin. Then, it is oxidized by O2, generating a high-energy intermediary that decomposes in CO2 and the excited oxyluciferin. Light emission produces the ground-state oxyluciferin and hydrolyzes enzymatically into caffeic acid [27].
Kotlobay et al. [28] identified the fungal Luz and three other key enzymes HispS, H3H, and CPH in Neonothopanus nambi that jointly form the biosynthetic cycle of the fungal luciferin from caffeic acid. Fungal luciferin can be biosynthesized and recycled within this proposed mechanism. Caffeic acid is transformed to hispidin owing to HispS activity and hydroxylated by H3H, producing 3-hydroxyhispidin fungal luciferin. The luciferase adds molecular oxygen, producing an endoperoxide (a high-energy intermediate) through decomposition that produces oxyluciferin (caffeylpyruvate) and light. Oxyluciferin can be recycled to caffeic acid by CPH [28].
In a study, Wang and Liu [94] revealed cross-reactions among four lineages of luminescent fungi indicating that they share a common bioluminescent mechanism and described the bioluminescent process at the molecular level and the electronic state by using multireference and density functional theory calculations. The findings revealed that fungal bioluminescence began with the cycloaddition of O2 to luciferin and that formed a high-energy intermediate called α-pyrone endoperoxide. This oxygenation can be explained by a charge transfer followed by a spin inversion mechanism. The high-energy intermediate thermolysis produces S1-Oxyluciferin (S1-singlet excited state) via a zwitterion intermediate. De-excitation of S1-Oxyluciferin can be a light emitter [94]. Nevertheless, according to Ke et al. [82], the complete cluster of genes involved in the bioluminescence process is still unknown to science.
In fungi, the genes that code for the enzymes that produce secondary metabolites are frequently grouped together in the fungi genome [95]. In most bioluminescent fungi (e.g., Armillaria fuscipes, A. mellea, A. ostoyae, A. gallica, Mycena citricolor, M. chlorophos, Neonothopanus nambi, N. gardneri, Omphalotus olearius, and Panellus stipticus) it has been demonstrated that aforesaid genes are generally found to be located adjacent to each other forming a cluster [28]. Rokas et al. [96] also revealed that genes in the primary and secondary metabolic pathways of fungi are often physically connected on fungal chromosomes, creating metabolic gene clusters, thus hypothesized that this might be the reason for the formation of enzymes in the bioluminescent cascade, as this thought to be conserved among bioluminescent fungi. In addition to the four key genes (Luz, H3H, HispH, and CPH) coding for four enzymes, CYP450 is inside the cluster in all bioluminescent Armillaria and Mycena genomes [82]. Thus, there is a controversy about the involvement of other enzymes or regulators in the bioluminescence process. On the other hand, few studies reported the possible involvement of other genes in fungi bioluminescence [97]. In another study, Oliveira et al. [98] investigated the circadian rhythm in Neonothopanus gardneri and found that bioluminescence of the mycelium is controlled by a temperature-compensated circadian clock and the result of cycles in content/activity between the luciferase, reductase, and luciferin that comprise within the bioluminescent system. Ke et al. [82] determined the regulation of bioluminescence in Mycena kentingensis during its development and identified 57 genes bearing expressions correlated to Luz, H3H, and HispS, agreeing with the afore-discussed bioluminescence mechanism presented by Kotlobay et al. [28].
4. Importance of Bioluminescent Fungi In Ecology
The ecological importance of bioluminescent fungi is quite debatable as there is a clear dearth of consensus among researchers; suggestions are regarding active spore dispersers, elimination of undesirable phototrophic fungivores, and signal generation for aposematism [4]. Initially, Sivinski [99] proposed the use of bioluminescence in fungi as a warning signal to deter nocturnal fungivores or an attractant for fungivore predators, though Sivinski’s theories challenge other hypotheses built on the attraction of animals towards bioluminescent fungi since fungivores may attract to the aroma/odor of fruiting bodies rather than the property of bioluminescence. Therefore, electroretinography must be used to clarify whether invertebrates attract bioluminescent fungi due to the light emitted by fungi; however, such studies are still on the bench [66].
Another trust is the capability of bioluminescent fungi to attract insects at night and use them for spore dispersal mechanisms. The studies conducted with Neonothopanus gardneri revealed that the bioluminescence mechanism of the fungus is regulated by the circadian rhythm, as the bioluminescent system consists of cycles in the content/activity of luciferase, reductase, and luciferin, yielding the observed results [4,98]. This conclusion is further strengthened as researchers found the beetles bathe with fruiting bodies of N. gardneri [100]. In a recent study, Karunarathna et al. [18] explained that members of Roridomyces inhabit humid environments co-evolved with some insects that aid in spore dispersal. Bechara [35] investigated the attraction of insects such as ants, beetles, flies, wasps, and bugs to the green light emitted by mushrooms than non-bioluminescent ones and concluded that the nocturnal transfer of spores was facilitated by insects in deep forests with minimal wind but higher levels of humidity which enhance spore germination. A small breeze is enough to carry out the light fungi spores; thus, dispersal in deep forests through the air is also possible. Nonetheless, winds are typically infrequent where N. gardneri is situated at the base of the forest canopy, whereas the dispersal through the air is highly challengeable; thus, it is assumed that fungi seek the help of insects to carry their spores.
In contrast to all the above, according to Weinstein et al. [31], there is nothing to wonder about the role behind the bioluminescence. Weinstein et al. [31] investigated a ghost fungus (Omphalotus nidiformis) and showed that bioluminescence is just an incidental by-product of metabolism rather than conferring any selective advantage; however, this study speculates that the role of bioluminescence may differ among evolutionary lineages of fungi and/or with growth environment attributes that may affect spore dispersals, such as wind and insect abundance.
5. Application of Fungal Bioluminescence
In the aspect of fungal bioluminescence, as previously highlighted, folk stories and historical reports show that different tribes or local people, particularly in India and Indonesia, use glowing mushrooms to find their way through the dense forests [100,101,102]. Nevertheless, Aboriginal people in Australia considered glowing mushrooms related to the spirit [103]. Today, bioluminescence has revolutionized plant biology and inspired experiments and research in biochemistry, cell biology, evolution, and photochemistry. Bioluminescence is also applied in scientific research, including several aspects such as biological sensors in the environmental monitoring, effectors, hygiene control, preservation of artworks, gene assays, the detection of protein-protein interactions, bioluminescence-based imaging and photodynamic therapy, neuron treatments, and high-throughput screening in drug discovery [6,104,105]. Interestingly, now, scientists are looking for ways to switch to green light instead of light generated through electricity [100]. Khakhar et al. [87] demonstrated that the fungal bioluminescence pathway can generate bioluminescence across various tissues in various plants without external substrate addition. Mitiouchkina et al. [30] engineered tobacco plants with a fungal bioluminescence system that converts caffeic acid (present in all plants) into luciferin and reports self-sustained luminescence visible to the naked eye. It has also been reported that fungal bioluminescence can be used for environmental pollution bioassays and auto-luminescent plants [2]. Mitiouchkina et al. [30] showed that it is possible to insert light-emitting genes from fungi into tobacco plants through genetic engineering, resulting in long-lasting illumination.
Environmental bioassays can be performed using fungi's natural bioluminescent enzyme reaction [77]. Eukaryotic bioluminescent fungi are a more suitable research organism for soil toxicology than luminescent bacteria; although the mechanism of toxicity and its specific impact on the fungal bioluminescence response is not yet fully understood, the uncoupling of oxidative phosphorylation and the depolarization of mitochondrial membranes by toxic compounds can be possible pathways, it would possibly indirectly have a way on the NADH availability that involved the bioluminescent reaction [77]. Additionally, it is still feasible to use organisms in a terrestrial setting, including bioluminescent fungi, without further engineering marker-labeling [2], even if known uses include marker-labeled bacteria and marine bioluminescent Vibrio species [106,107]. Since 2002, toxicity tests have been developed using bioluminescent fungi, particularly with Armillaria mellea and Mycena citricolor. These studies were based on globular mycelia grown on liquid media with varying concentrations of heavy metals (copper and zinc) or chemical compounds (chlorophenol). Several bioassays with bioluminescent basidiomycetes have been developed using Pannellus stipticus and Omphalotus olearius [77]. More recently, the toxicity of the other bioluminescent lineages of G. viridilucens and N. gardneri, was assessed [108,109]
On the one hand, reconstruction of the fungal bioluminescent pathway in the organism, making it autonomous luminescence, is beneficial to detect the status of its various growth stages. It could also facilitate the development of the next generation of organic architecture, modified light-emitting plants into buildings, and urban infrastructure [28]. On the other hand, fungal bioluminescence can be indicated in agriculture when crops need water or nutrients. Due to this autonomous bioluminescence, plants can warn early about illnesses and pest attacks that could harm harvests. Furthermore, bioluminescence paves the path for eco-friendly house/street lighting, health applications, and food industries [100,101,102]. The alterations of these technologies will drive massive growth in bioluminescence in the coming future [74].
6. Conclusions
To date, 122 bioluminescent fungi have been taxonomically classified, and the complete diversity of fungi has yet to be assessed owing to the continuous identification of new species. All known bioluminescent fungi mainly belong to the group of Basidiomycota and are scattered among four phylogenetic lineages (Armillaria, Lucentipes, Mycenoid, and Omphalotus), and Eoscyphella lineage represents a whole new fifth lineage of bioluminescent mushrooms. Even though species differ, all species share a common bioluminescent mechanism. Fungi possess a simple bioluminescent system, with luciferin having only two enzymatic steps from a known metabolic pathway. Gene expression in the bioluminescence process is yet to be studied. Understanding the detailed mechanism of catalytic oxidation of luciferin by luciferase in fungal bioluminescence would be an excellent ease for further development of its applications. Perceiving the lessons from nature, some limited devices and technologies have been developed from bioluminescent systems. Further, recent findings on fungal bioluminescence expand the existing knowledge and applications such as pollution detection, bioassays in ecosystems, and in vivo gene expression detection of reporter genes. Moreover, the fungal bioluminescent system could bring bioluminescent technologies beyond being used as scientific research tools. Also, this technology will enable the development of a new dimension based on light emission.
7. Future Directions
Future investigations into bioluminescent fungi have great promise for various sectors, including biotechnology, synthetic biology, ecology, agriculture, art, medicine, and education. These fungi provide innovative solutions such as bioindicators for environmental monitoring, sustainable lighting, and novel medical imaging techniques. By modifying metabolic pathways, synthetic biology advancements may create new bioluminescent animals and chemicals. While ecological research might improve our knowledge of fungal biodiversity and ecosystem dynamics, to increase the number of discovered bioluminescent fungi and investigate their signification in biological, agricultural applications can involve crop health monitoring and pest management. These fungi's inherent luminosity inspires creative and innovative designs, such as luminous gardens and live art pieces. Research in pharmaceuticals may provide novel therapeutic molecules and diagnostic aids. Furthermore, bioluminescent fungi offer captivating methods to improve biology teaching and public comprehension of science. However, reaching these fantastic creatures' full potential will require ongoing studies and the invaluable collaboration of scientists, researchers, educators, and professionals in various fields.
Author Contributions
SCK and ST conceptualized the topic; WHL and AKHP wrote the first draft of the manuscript; MCAG, NMP, ANE, BMP, and DMK helped with the literature searches and drafted the chemistry part of the manuscript; TSN prepared the map; ST, SCS, DQD, AKHP, AT, NS, SL and SCK edited the manuscript; ST and SCK supervised the work; DQD, NS, SL, SCS, and SCK provided funding for the research. All authors have read, edited, and approved the final manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 32260004, 31760013, and 32171655), the West Light Talent Program of the Chinese Academy of Sciences (Grant No. E1XB011B01), and Chiang Mai University and Kaomai Lanna Res., Thailand (Grant No. R66IN00132). Samantha C. Karunarathna, Dong-Qin Dai, and Saowaluck Tibpromma thank the High-Level Talent Recruitment Plan of Yunnan Province ("High-End Foreign Expert" and “Young Talents” Programs) for their support.
Data Availability Statement
The data that support this study are available at request.
Acknowledgments
The authors thank the Key Laboratory of Yunnan Provincial Department of Education of the Deep-Time Evolution on Biodiversity from the Origin of the Pearl River for the support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pandey, A.; Sharon, M. Bioluminescent organisms. BAOJ Chem. 2017, 3, e029. [Google Scholar]
- Ke, H.M.; Tsai, I.J. Understanding and using fungal bioluminescence-Recent progress and future perspectives. In: Kümmerer, K., Liu, Z. (Eds), Curr. Opin. Green. Sustain. Chem. 2022, 33, 100570. [Google Scholar] [CrossRef]
- Waldenmaier, H.E.; Oliveira, A.G.; Stevani, C.V. Thoughts on the diversity of onvergent evolution of bioluminescence on earth. Int. J. Astrobiol. 2012, 11, 335–343. [Google Scholar] [CrossRef]
- Vieira, M.B.B.; Oliveira, I.C.; de Oliveira, M.D.D.A.; da Costa Júnior, J.S.; dos Santos, T.D.J.A.; Feitosa, C.M.; Rai, M.; Lima, N.M.; da Costa Silva, D. A review on bioluminescent fungus Neonothopanus gardneri. Res., Soc. Dev. 2022, 11, e16811528009. [Google Scholar] [CrossRef]
- Lau, E.S.; Oakley, T.H. Multi-level convergence of complex traits and the evolution of bioluminescence. Biol. Rev. Camb. Philos. Soc. 2020, 96, 673–691. [Google Scholar] [CrossRef]
- Syed, A.J.; Anderson, J.C. Applications of bioluminescence in biotechnology and beyond. Chem. Soc. Rev. 2021, 50, 5668–5705. [Google Scholar] [CrossRef] [PubMed]
- Harvey, E.N. Bioluminescence. Academic Press, New York, 1952; 1–649.
- Harvey, E.N. A history of luminescence: from the earliest times until 1900. The American Philosophical Society, Philadelphia, 1957; 1–692. [CrossRef]
- Boyle, R. Observations and tryals about the resemblances and differences between a burning coal and shining wood. Phil. Trans. 1667, 2, 605–612. [Google Scholar] [CrossRef]
- Oba, Y.; Hosaka, K. The luminous fungi of Japan. J. Fungi 2023, 9, e615. [Google Scholar] [CrossRef]
- Kirk, P.M. World catalogue of 340K fungal names on-line. Mycol. Res. 2000, 104, 516–517. [Google Scholar]
- Crous, P.W.; Gams, W.; Stalpers, J.A.; Robert, V.; Stegehuis, G. MycoBank: an online initiative to launch mycology into the 21st century. Stud. Mycol. 2004, 50, 19–22. [Google Scholar]
- Taylor, J.W. One Fungus = One Name: DNA and fungal nomenclature twenty years after PCR. IMA Fungus 2011, 2, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A. ; Wen Chen Fungal Barcoding Consortium Fungal Barcoding Consortium Author List; Bolchacova, E.; Voigt, K.; Crous, P.W.; et al. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Robbertse, B.; Robert, V.; Vu, D.; Cardinali, G.; Irinyi, L.; Meyer, W.; Nilsson, R.H.; Hughes, K.; Miller, A.N.; et al. Finding needles in haystacks: linking scientific names, reference specimens and molecular data for Fungi. Database 2014, bau061. [Google Scholar] [CrossRef] [PubMed]
- McNeill, J.; Barrie, F.R.; Buck, W.R.; Demoulin, V.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Marhold, K.; Prado, J.; et al. International Code of Nomenclature for algae, fungi, and plants. Regnum Vegetabile 2012, 154. [Google Scholar]
- Oliveira, A.G.; Desjardin, D.E.; Perry, B.A.; Stevani, C.V. Evidence that a single bioluminescent system is shared by all known bioluminescent fungal lineages. Photochem. Photobiol. Sci. 2012, 11, 848–852. [Google Scholar] [CrossRef] [PubMed]
- Karunarathna, S.C.; Mortimer, P.E.; Tibpromma, S.; Dutta, A.K.; Paloi, S.; Hu, Y.; Baurah, G.; Axford, S.; Marciniak, C.; Luangharn, T.; et al. Roridomyces phyllostachydis (Agaricales, Mycenaceae), a new bioluminescent fungus from Northeast India. Phytotaxa 2020, 459, 155–167. [Google Scholar] [CrossRef]
- Cortés-Pérez, A.; Guzmán-Dávalos, L.; Ramírez-Cruz, V.; Villalobos-Arámbula, A.R.; Ruiz-Sanchez, E.; Ramírez-Guillén, F. New Species of Bioluminescent Mycena Sect. Calodontes (Agaricales, Mycenaceae) from Mexico. J. Fungi 2023, 9, e902. [Google Scholar] [CrossRef] [PubMed]
- Labella, A.M.; Arahal, D.R.; Castro, D.; Lemos, M.L.; Borrego, J.J. Revisiting the genus Photobacterium: taxonomy, ecology and pathogenesis. Int. Microbiol. 2017, 20, 1–10. [Google Scholar]
- Burtseva, O.; Kublanovskaya, A.; Baulina, O.; Fedorenko, T.; Lobakova, E.; Chekanov, K. The strains of bioluminescent bacteria isolated from the White Sea finfishes: genera Photobacterium, Aliivibrio, Vibrio, Shewanella, and first luminous Kosakonia. J. Photoch. Photobio. B. 2020, 208, 111895. [Google Scholar] [CrossRef]
- de Busserolles, F.; Marshall, N.J. Seeing in the deep-sea: visual adaptations in lanternfishes. Philos. Trans. R. Soc. Lond., B, Biol. Sci. 2017, 372, 20160070. [Google Scholar] [CrossRef]
- Al-Handawi, M.B.; Polavaram, S.; Kurlevskaya, A.; Commins, P.; Schramm, S.; Carrasco-López, C.; Lui, N.M.; Solntsev, K.M.; Laptenok, S.P.; Navizet, I.; et al. Spectrochemistry of firefly bioluminescence. Chem. Rev. 2022, 122, 13207–13234. [Google Scholar] [CrossRef] [PubMed]
- Owens, A.; Van den Broeck, M.; De Cock, R.; Lewis, S.M. Behavioral responses of bioluminescent fireflies to artificial light at night. Front. Ecol. Evol. 2022, 10, 946640. [Google Scholar] [CrossRef]
- Elkhateeb, W.A.; Daba, G.M. Bioluminescent Mushrooms: Boon for Environmental Health. Environ. Sci. Arch. 2022, 1, 88–97. [Google Scholar]
- Adams, C.A.; Donald, M.L.; Swearingen, C.; Escudero, E.; Sourell, S.; Landrein, S.; Seas, C.; Mueller, G.; Chaverri, P. Let there be nightlights: the ecological role of bioluminescence in a Costa Rican mushroom. bioRxiv. 2023. [CrossRef]
- Kaskova, Z.M.; Dörr, F.A.; Petushkov, V.N.; Purtov, K.V.; Tsarkova, A.S.; Rodionova, N.S.; Mineev, K.S.; Guglya, E.B.; Kotlobay, A.; Baleeva, N.S.; et al. Mechanism and color modulation of fungal bioluminescence. Sci. Adv. 2017, 3, e1602847. [Google Scholar] [CrossRef]
- Kotlobay, A.A.; Sarkisyan, K.S.; Mokrushina, Y.A.; Marcet-Houben, M.; Serebrovskaya, E.O.; Markina, N.M.; Gonzalez, S.L.; Gorokhovatsky, A.Y.; Vvedensky., A.; Purtov, K.V.; et al. Genetically encodable bioluminescent system from fungi. Proc. Natl. Acad. Sci. 2018, 115, 12728–12732. [Google Scholar] [CrossRef]
- Chang, C.C.; Chen, C.Y.; Li, W.W.; Ka, W. Mycena jingyinga, Mycena luguensis, and Mycena venus: three new species of bioluminescent fungi from Taiwan. Taiwania 2020, 65, 396–406. [Google Scholar]
- Mitiouchkina, T.; Mishin, A.S.; Somermeyer, L.G.; Markina, N.M.; Chepurnyh, T.V.; Guglya, E.B.; Karataeva, T.A.; Palkina, K.A.; Shakhova, E.S.; Fakhranurova, L.I.; et al. Plants with genetically encoded autoluminescence. Nat. Biotechnol 2020, 38, 944–946. [Google Scholar] [CrossRef]
- Weinstein, P.; Delean, S.; Wood, T.; Austin, A.D. Bioluminescence in the ghost fungus Omphalotus nidiformis does not attract potential spore dispersing insects. IMA Fungus 2016, 7, 229–234. [Google Scholar] [CrossRef]
- Bermudes, D.; Petersen, R.H.; Nealson, K.H. Low-level bioluminescence detected in Mycena haematopus basidiocarps. Mycologia 1992, 84, 799–802. [Google Scholar] [CrossRef]
- Danks, M.A.; Simpson, N.; Elliott, T.F.; Paine, C.T.; Vernes, K. Modeling mycorrhizal fungi dispersal by the mycophagous swamp wallaby (Wallabia bicolor). Ecol. Evol. 2020, 10, 12920–12928. [Google Scholar] [CrossRef]
- Elliott, T.F.; Truong, C.; Jackson, S.M.; Zúñiga, C.L.; Trappe, J.M.; Vernes, K. Mammalian mycophagy: A global review of ecosystem interactions between mammals and fungi. J. Syst. Evol. 2022, 9, 99–159. [Google Scholar] [CrossRef] [PubMed]
- Bechara, E.J.H. Bioluminescence: A fungal nightlight with an internal timer. Curr Biol. 2015, 25, R283–R285. [Google Scholar] [CrossRef] [PubMed]
- Dauner, L.A.P.; Karunarathna, S.C.; Tibpromma, S.; Xu, J.; Mortimer, P.E. Bioluminescent fungus Roridomyces viridiluminus sp. nov. and the first Chinese record of the genus Roridomyces from Southwestern China. Phytotaxa 2021, 487, 233–250. [Google Scholar] [CrossRef]
- Ke, H.M.; Lu, M.R.; Chang, C.C.; Hsiao, C.; Ou, J.H.; Taneyama, Y.; Tsai, I.J. Evolution and Diversity of Bioluminescent Fungi. Cham: Springer International Publishing, Evolution of Fungi and Fungal-Like Organisms, 2023; 275-294.
- Lingle, W.L. Bioluminescence and ligninolysis during secondary metabolism in the fungus Panellus stypticus. J. Biolum. Chemilum. 1993, 8, e100. [Google Scholar]
- Niskanen, T.; Lücking, R.; Dahlberg, A.; Gaya, E.; Suz, L.M.; Mikryukov, V.; Liimatainen, K.; Druzhinina, I.; Westrip, J.R.; Mueller, G.M.; et al. Pushing the frontiers of biodiversity research: Unveiling the global diversity, distribution, and conservation of fungi. Annu. Rev. Environ. Resour. 2023, 48, 149–176. [Google Scholar] [CrossRef]
- Rashhmi, M.; Kushveer, J.S.; Sarma, V.V. A worldwide list of endophytic fungi with notes on ecology and diversity. Mycosphere 2019, 10, 798–1079. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.; Martin, F.M.; Selosse, M.A.; Sanders, I.R. Mycorrhizal ecology, and evolution: the past, the present, and the future. New Phytol. 2015, 205, 1406–1423. [Google Scholar] [CrossRef]
- Vydryakova, G.A.; Gusev, A.A.; Medvedeva, S.E. Effect of organic and inorganic toxic compounds on luminescence of luminous fungi. Appl. Biochem. Microbiol. 2011, 47, 293–297. [Google Scholar] [CrossRef]
- Mihail, J.D. Bioluminescence patterns among North American Armillaria species. Fungal Biol. 2015, 119, 528–537. [Google Scholar] [CrossRef]
- Desjardin, D.E.; Oliveira, A.G.; Stevani, C.V. Fungi bioluminescence revisited. Photochem. Photobiol. Sci. 2008, 7, 170–182. [Google Scholar] [CrossRef]
- Tan, Z.J.; Xie, D.P.; Wang, Z.; Li, W.G.; Liu, S. The study on bioluminescence condition of Amillaria mellea. Acta laser biology sinica. 2001, 10, 1007–7146. (In Chinese) [Google Scholar]
- Patil, S.R.; Yadav, S.V. Photographic record of Armillaria mellea a bioluminescent fungus from Lonavala in western Ghats, India. J. Threat. Taxa 2022, 14, 20692–20694. [Google Scholar] [CrossRef]
- BITB. Bioluminescence in the Bush: Glow in the Dark Mushrooms in Stewart Island. Available online: https://www.myconeer.com/p/bioluminescence-in-the-bush-glow (accessed on 15 April 2021).
- Ainsworth, M. Searching for luminous mushrooms of the marsh fungus Armillaria ectypa. Field Mycology 2004, 5(4), 142–144. [Google Scholar] [CrossRef]
- Silva-Filho, A.G.; Mombert, A.; Nascimento, C.C.; Nóbrega, B.B.; Soares, D.M.; Martins, A.G.; Domingos, A.H.; Santos, I.; Della-Torre, O.H.; Perry, B.A.; et al. Eoscyphella luciurceolata gen. and sp. nov. (Agaricomycetes) shed light on Cyphellopsidaceae with a new lineage of bioluminescent fungi. J. Fungi 2023, 9, 1004. [Google Scholar] [CrossRef]
- Nimalrathna, T.; Tibpromma, S.; Nakamura, A.; Galappaththi, M.C.A.; Xu, J.; Mortimer, P.E.; Karunarathna, S.C. The case of the missing mushroom: A novel bioluminescent species discovered within Favolaschia in Southwestern China. Phytotaxa 2022, 539, 244–256. [Google Scholar] [CrossRef]
- Liu, P.G. Luminescent fungi. Chinese Biodiversity 1995, 3, 109–112. (In Chinese) [Google Scholar]
- Terashima, Y.; Neda, H.; Hiroi, M. ; Luminescent intensity of cultured mycelia of eight basidiomycetous fungi from Japan. Mushroom Science and Biotechnology 2016, 24, 176–181. [Google Scholar]
- Terashima, Y.; Takahashi, H.; Taneyama, Y. The fungal flora in Southwestern Japan: Agarics and boletes. Tokai Daigaku, Kanagawa, Japan, 2016, pp. 349.
- Liu, P.G.; Yang, Z.L. Studies of classification and geographic distribution of Laschia-complex from the Southern and Southeastern Yunnan, China. Acta Botanica Yunnanica 1994, 16, 47–52. (In Chinese) [Google Scholar]
- Chew, A.L.; Desjardin, D.E.; Tan, Y.S.; Musa, M.Y.; Sabaratnam, V. Bioluminescent fungi from peninsular Malaysia- A taxonomic and phylogenetic overview. Fungal Divers. 2014, 70, 149–187. [Google Scholar] [CrossRef]
- Yan, J.J.; Liu, X.R.; Xie, B.G.; Deng, Y.J. Isolation, identification, and characterization of Neonothopanus nambi (Basidiomycota, Fungi), a new record from China. Bulletin of Microbiology 2015, 42: 1703–1709.
- Desjardin, D.E.; Perry, B.A.; Lodge, D.J.; Stevani, C.V.; Nagasawa, E. Luminescent Mycena: New and noteworthy species. Mycologia 2010, 102, 459–477. [Google Scholar] [CrossRef] [PubMed]
- Chew, A.L.; Tan, Y.S.; Desjardin, D.E.; Musa, M.Y.; Sabaratnam, V. ; Four new bioluminescent taxa of Mycena sect. Calodontes from Peninsular Malaysia. Mycologia 2014, 106, 976–988. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.J.; Vargas-Isla, R.; Cabral, T.S.; Cardoso, J.S.; Andriolli, F.S.; Rodrigues, D.P.; Ikeda, T.; Clement, C.R.; Ishikawa, N.K. The amazonian luminescent Mycena cristinae sp. nov. from Brazil. Mycoscience 2021, 62, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Heinzelmann, R.; Baggenstos, H.; Rudolf, A. Is the bioluminescence in many Mycena species overlooked? – A case study from M. crocata, 2024. [Google Scholar] [CrossRef]
- Aravindakshan, D.M.; Kumar, T.K.A.; Manimohan, P. A new bioluminescent species of Mycena sect. Exornatae from Kerala State, India. Mycosphere 2012, 3, 556–561. [Google Scholar] [CrossRef]
- Desjardin, D.E.; Perry, B.A.; Stevani, C.V. New luminescent Mycenoid fungi (Basidiomycota, Agaricales) from Sao Paulo state, Brazil. Mycologia 2016, 108, 1165–1174. [Google Scholar] [PubMed]
- Cortés-Pérez, A.; Desjardin, D.E.; Perry, B.A.; Ramírez-Cruz, V.; Ramírez-Guillén, F.; Villalobos-Arámbula, A.R.; Rockefeller, A. New species, and records of bioluminescent Mycena from Mexico. Mycologia 2019, 111, 319–573. [Google Scholar] [CrossRef] [PubMed]
- Soares, C.C.; Cabral, T.S.; Vargas-isla, R.U.B.Y.; Cardoso, J.S.; Rodrigues, D.P.; Ishikawa, N.K.; Oliveira, J.J. Mycena lamprocephala, a new luminescent species from the Brazilian Amazon. Phytotaxa, 2024, 634(3), 187–203.
- Alves, M.H.; do Nascimento, C.C. Mycena margarita (Murrill) Murrill, 1916 (Basidiomycota: Agaricales: Mycenaceae): A bioluminescent agaric first recorded in Brazil. Check List 2014, 10, 239–243. [Google Scholar] [CrossRef]
- Desjardin, D.E.; Capelari, M.; Stevani, C. Bioluminescent Mycena species from São Paulo, Brazil. Mycologia 2007, 99, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Horak, E. Mycena rorida (Fr.) Quél. and related species from the Southern Hemisphere. Berichte. 1978, 88, 20–29. [Google Scholar] [CrossRef]
- Lee, S.M.L. The status of bioluminescent fungal species in Singapore. Nature in Singapore 2022, 1, e2022124. [Google Scholar]
- Tolgor, B. Notes on Basidiomycetes of Jilin province (VIII). Journal of Fungal Research 2007, 5, 72–74. [Google Scholar]
- Yang, Z.L.; Feng, B. The genus Omphalotus (Omphalotaceae) in China. Mycosystema 2013, 32, 545–556. [Google Scholar]
- Li, J.Z.; Hu, X.W. A new species of Lampteromyces from Hunan. Acta Sci Nat Univ Norm Hunan. 1993, 16, 2. [Google Scholar]
- OSSJ. Omphalotus subilludens – Southern Jack O' Lantern. Available online: https://www.texasmushrooms.org/en/omphalotus_subilludens.htm (accessed on 1 November 2013).
- Foerster, G.E.; Behrens, P.Q.; Airth, R.L. Bioluminescence and other characteristics of Collybia velutipes. Am. J. Bot. 1965, 52, 487–495. [Google Scholar] [CrossRef]
- Kushwaha, V.; Hajirnis, S. A review on bioluminescent fungi: A torch of curiosity. Int. J. Life Sci. 2016, A7, 107–110. https://oaji.net/articles/2017/736-1518519635.pdf.
- Wassink, E.C. Observations on the luminescence in fungi I, including a critical review of the species mentioned as luminescent in literature. Recueil des travaux botaniques néerlandais 1948, 4, 150–212. [Google Scholar]
- Desjardin, D.E.; Capelari, M.; Stevani, C.V. A new bioluminescent Agaric from São Paulo, Brazil. Fungal Divers. 2005, 18, 9–14. [Google Scholar]
- Stevani, C.V.; Oliveira, A.G.; Mendes, L.F.; Ventura, F.F.; Waldenmaier, H.E.; Carvalho, R.P.; Pereira, T.A. Current status of research on fungal bioluminescence: biochemistry and prospects for ecotoxicological application. Photochem. Photobiol. 2013, 89, 1318–1326. [Google Scholar] [CrossRef] [PubMed]
- Peterson, R.H.; Bermudes, D. Intercontinental compatibility in Panellus stypticus with a note on bioluminescence. Persoonia 1992, 14, 457–463. [Google Scholar]
- Arya, C.P.; Ratheesh, S.; Pradeep, C.K. New record of luminescent Mycena chlorophos (Mycenaceae) from Western Ghats of India. Studies in Fungi 2021, 6, 507–513. [Google Scholar] [CrossRef]
- Dutta, A.; Gupta, S.; Roy, J.K.; Ahmed, M.F. New distribution record of Roridomyces cf. phyllostachydis (Agaricales: Mycenaceae), a bioluminescent fungus from Namdapha National Park, Arunachal Pradesh. J. Threat. Taxa 2023, 3, 22920–22923. [Google Scholar] [CrossRef]
- Shih, Y.S.; Chen, C.Y.; Lin, W.W.; Kao, H.W. Mycena kentingensis, a new species of luminous mushroom in Taiwan, with reference to its culture method. Mycol. Prog. 2014, 13, 429–435. [Google Scholar] [CrossRef]
- Ke, H.M.; Lee, H.H.; Lin, C.I.; Liu, Y.C.; Lu, M.R.; Hsieh, J.A.; Chang, C.C.; Wu, P.H.; Lu, M.J.; Li, J.Y.; et al. Mycena genomes resolve the evolution of fungal bioluminescence. Proc. Natl. Acad. Sci. 2020, 117, 31267–31277. [Google Scholar] [CrossRef]
- Palkina, K.A.; Balakireva, A.V.; Belozerova, O.A.; Chepurnykh, T.V.; Markina, N.M.; Kovalchuk, S.I.; Tsarkova, A.S.; Mishin, A.S.; Yampolsky, I.V.; Sarkisyan, K.S. Domain truncation in hispidin synthase orthologs from non-bioluminescent fungi does not lead to hispidin biosynthesis. Int. J. Mol. Sci. 2023, 24, e1317. [Google Scholar] [CrossRef] [PubMed]
- Kahlke, T.; Umbers, K.D.L. Bioluminescence. Curr. Biol. 2016, 26, R313–R314. [Google Scholar] [CrossRef]
- Wilson, T.; Hastings, J.W. Bioluminescence: Living lights, lights for living. Harvard University Press 2013, 208. [Google Scholar] [CrossRef]
- Liu, X.; Wang, M.; Liu, Y. Chemistry in fungal bioluminescence: Theoretical studies on biosynthesis of luciferin from caffeic acid and regeneration of caffeic acid from oxidized luciferin. J. Fungi 2023, 9, 369. [Google Scholar] [CrossRef]
- Khakhar, A.; Starker, C.G.; Chamness, J.C.; Lee, N.; Stokke, S.; Wang, C.; Swanson, R.; Rizvi, F.; Imaizumi, T.; Voytas, D.F. Building customizable auto-luminescent luciferase-based reporters in plants. ELife 2020, 9, e52786. [Google Scholar] [CrossRef]
- Dubois, R. Note sur la fonction photogénique chez la Pholas dactylus, CR. Seances Soc. Biol. Paris. 1887, 39, e564. [Google Scholar]
- Airth, R.L.; McElroy, W.D. Light emission from extract of luminous fungi. J Bacteriol. 1959, 77, 249–250. [Google Scholar] [CrossRef]
- Airth, R.L.; Foerster, G.E. The isolation of catalytic components required for cell-free fungal bioluminescence. Arch. Biochem. Biophys. 1962, 97, 567–573. [Google Scholar] [CrossRef]
- Shimomura, O. Bioluminescence: Chemical principles and methods. World Scientific Publishing Co. Pte. Lt, Singapore, 2006; 500. [CrossRef]
- Oliveira, A.G.; Stevani, C.V. The enzymatic nature of fungal bioluminescence. Photochem. Photobiol. Sci. 2009, 8, 1416. [Google Scholar] [CrossRef] [PubMed]
- Purtov, K.V.; Petushkov, V.N.; Baranov, M.S.; Mineev, K.S.; Rodionova, N.S.; Kaskova, Z.M.; Tsarkova, A.S.; Petunin, A.I.; Bondar, V.S.; Rodicheva, E.K.; et al. The chemical basis of fungal bioluminescence. Angew. Chem. Int. Ed. Engl. 2015, 54, 8124–8128. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.Y.; Liu, Y.J. Chemistry in fungal bioluminescence: A theoretical study from Luciferin to light emission. J. Org. Chem. 2021, 86, 1874–1881. [Google Scholar] [CrossRef]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism-from biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Rokas, A.; Wisecaver, J.H.; Lind, A.L. The birth, evolution and death of metabolic gene clusters in fungi. Nat. Rev. Microbiol. 2018, 16, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Park, M.J.; Shim, D.; Ryoo, R. De novo genome assembly of the bioluminescent mushroom Omphalotus guepiniiformis reveals an Omphalotus-specific lineage of the luciferase gene block. Genomics 2022, 114, e110514. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.G.; Stevani, C.V.; Waldenmaier, H.E.; Viviani, V.; Emerson, J.M.; Loros, J.J.; Dunlap, J.C. Circadian control sheds light on fungal bioluminescence. Curr. Biol. 2015, 25, 964–968. [Google Scholar] [CrossRef] [PubMed]
- Sivinski, J.M. Arthropods attracted to luminous fungi. Psyche: A J. Entomol. 1981, 88, 383–390. [Google Scholar] [CrossRef]
- Mishara, M.; Srivastava, D. Bioluminescent fungi: Reviewing nature's riddle. J. Mycopathol. Res. 2021, 59, 199–206. [Google Scholar]
- Jabr, F. The secret history of bioluminescence. Hakai Magazine, 2016. Available online: https://hakaimagazine.com/features/secret-history-bioluminescence/ (accessed on 14th July 2023).
- Jain, N. In Meghalaya, scientists discover glowing mushrooms, used by locals as natural torches. 2020. Available online: https://scroll.in/article/978927/in-meghalaya-scientists-discover-glowing-mushrooms-used-by-locals-as-natural-torches (accessed on 14th July 2023).
- Lepp, H. Aboriginal use of fungi. 2013, Available online:. Available online: https://www.anbg.gov.au/fungi/aboriginal.html (accessed on 14th July 2023).
- Araújo-Gomes, N.; Zambito, G.; Johnbosco, C.; Calejo, I.; Leijten, J.; Löwik, C.; Karperien, M.; Mezzanotte, L.; Teixeira, L.M. Bioluminescence imaging on-chip platforms for non-invasive high-content bioimaging. Biosens. Bioelectron. 2023, 237, e115510. [Google Scholar] [CrossRef]
- Davies, K.A.; Welch, S.R.; Jain, S.; Sorvillo, T.E.; Coleman-McCray, J.D.; Montgomery, J.M.; Spiropoulou, C.F.; Albariño, C.; Spengler, J.R. Fluorescent and bioluminescent reporter mouse-adapted Ebola viruses maintain pathogenicity and can be visualized in vivo. J. Infect. Dis. 2023, p.jiad136. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, S.M.; Poziomek, E.J.; Engelmann, W.H.; Rogers, K.R. A review of environmental applications of bioluminescence measurements. Chemosphere 1995, 30, 2155–2197. [Google Scholar] [CrossRef]
- Gianfreda, L.; Rao, M.A. Interactions between xenobiotics and microbial and enzymatic soil activity. Crit. Rev. Environ. Sci. Technol. 2008, 3, 269–310. [Google Scholar] [CrossRef]
- Ventura, F.F.; Mendes, L.F.; Oliveira, A.G.; Bazito, R.C.; Bechara, E.J.; Freire, R.S.; Stevani, C.V. Evaluation of phenolic compound toxicity using a bioluminescent assay with the fungus Gerronema viridilucens. Environ Toxicol Chem. 2020, 39(8), 1558–1565. [Google Scholar] [CrossRef] [PubMed]
- Ventura, F.F.; Soares, D.M.; Bayle, K.; Oliveira, A.G.; Bechara, E.J.; Freire, R.S.; Stevani, C.V. Toxicity of metal cations and phenolic compounds to the bioluminescent fungus Neonothopanus gardneri. Environmental Advances, 2021, 4, p100044. [Google Scholar] [CrossRef]
- Rani, A.Q.; Zhu, B.; Ueda, H.; Kitaguchi, T. Recent progress in homogeneous immunosensors based on fluorescence or bioluminescence using antibody engineering. Analyst. 2023. [CrossRef] [PubMed]
- Resendiz-Sharpe, A.; Vanhoffelen, E.; Velde, G.V. Bioluminescence imaging, a powerful tool to assess fungal burden in live mouse models of infection. Antifungal immunity. 2023, 197–210. [Google Scholar] [CrossRef]
- Yadav, A.K.; Chan, J. Activity-based bioluminescence probes for in vivo sensing applications. Curr. Opin. Chem. Biol. 2023, 74, e102310. [Google Scholar] [CrossRef]
Figure 1.
The Venn diagram of common bioluminescent part-related 122 fungal species set. (A) Bioluminescence presents in mycelium, fruiting body and to determine whether bioluminescence is present when another part is reported to be bioluminescent fungi. (B) Bioluminescence presents in mycelium, different parts of the fruiting body (Pileus and stipe), and unspecified parts of the fruiting body.
Figure 1.
The Venn diagram of common bioluminescent part-related 122 fungal species set. (A) Bioluminescence presents in mycelium, fruiting body and to determine whether bioluminescence is present when another part is reported to be bioluminescent fungi. (B) Bioluminescence presents in mycelium, different parts of the fruiting body (Pileus and stipe), and unspecified parts of the fruiting body.
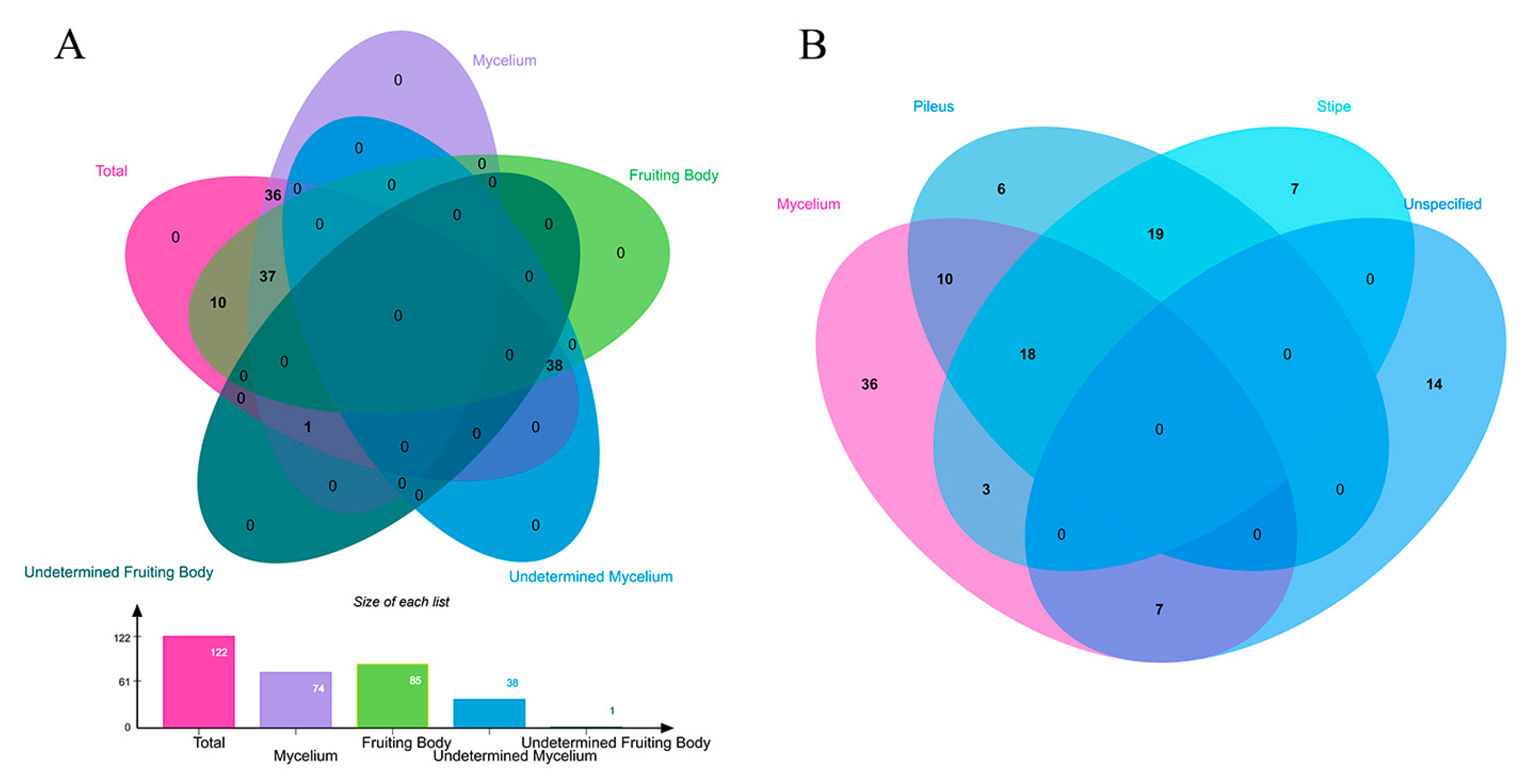
Figure 2.
The global distribution of bioluminescent fungi is based on the available literature; as the study was carried out, the richest fungi species were reported in Asia, followed by North and South America.
Figure 2.
The global distribution of bioluminescent fungi is based on the available literature; as the study was carried out, the richest fungi species were reported in Asia, followed by North and South America.

Figure 3.
Species percentage based on species counts reported from each lineage of bioluminescent fungi.
Figure 3.
Species percentage based on species counts reported from each lineage of bioluminescent fungi.

Figure 4.
Glowing piece of wood (a). Glowing mycelia on a wood in the Xishuangbanna Tropical Botanical Garden of China (b). Whole fruiting body and spores glowing in Favolaschia xtbgensis in Xishuangbanna Tropical Botanical Garden of China (c). Stipe glowing in Roridomyces phyllostachydis from Northeast India (d) (photo credit: Stephen Axford).
Figure 4.
Glowing piece of wood (a). Glowing mycelia on a wood in the Xishuangbanna Tropical Botanical Garden of China (b). Whole fruiting body and spores glowing in Favolaschia xtbgensis in Xishuangbanna Tropical Botanical Garden of China (c). Stipe glowing in Roridomyces phyllostachydis from Northeast India (d) (photo credit: Stephen Axford).


Table 1.
The list of bioluminescent fungi reported worldwide. Bioluminescence can be produced by entire fungi or, sometimes, only the mycelium, fruiting bodies, or spores.
Table 1.
The list of bioluminescent fungi reported worldwide. Bioluminescence can be produced by entire fungi or, sometimes, only the mycelium, fruiting bodies, or spores.
| Fungal Taxa | Distribution | Glowing Part | References | |||||
|---|---|---|---|---|---|---|---|---|
| Mycelium | Fruiting bodies | Cap | Stipe | Spores | ||||
| Armillaria Lineage | Armillaria borealis | Russia | + | / | / | / | / | [42] |
| Armillaria calvescens | USA | + | / | / | / | / | [43] | |
| Armillaria cepistipes | USA | + | / | / | / | / | [43] | |
| Armillaria fuscipes | Malaysia | + | / | / | / | / | [44] | |
| Armillaria gallica | Europe and USA | + | / | / | / | / | [44] | |
| Armillaria gemina | USA | + | / | / | / | / | [43] | |
| Armillaria mellea | China, Europe, India, and USA | + | / | / | / | / | [44,45,46] | |
| Armillaria nabsnona | USA | + | / | / | / | / | [43] | |
| Armillaria novae-zelandiae | New Zealand | + | / | / | / | / | [47] | |
| Armillaria ostoyae | Europe and USA | + | / | / | / | / | [44] | |
| Armillaria sinapina | USA | + | / | / | / | / | [43] | |
| Desarmillaria ectypa | Europe | + | + | + | / | / | [48] | |
| Desarmillaria tabescens | Europe and USA | + | + | / | / | / | [44] | |
| Eoscyphella Lineage | Eoscyphella luciurceolata | Brazil | ? | + | ? | ? | ? | [49] |
| Lucentipes Lineage | Mycena lucentipes | South America and ♣ | + | + | ? | + | / | [44] |
| Gerronema viridilucens | South America | + | + | + | + | / | [44] | |
| Mycenoid Lineage | Cruentomycena orientalis | Japan | + | + | + | + | / | [10] |
| Dictyopanus foliicola | Japan | + | + | / | / | / | [44] | |
| Favolaschia xtbgensis | China | + | + | + | + | + | [50] | |
| Favolaschia tonkinensis | China | ? | + | + | + | / | [51] | |
| Favolaschia peziziformis | Japan | ? | + | + | + | / | [52,53] | |
| Filoboletus manipularis | Africa, China, Sri Lanka and ♪ | ? | + | / | + | / | [54,55,56] | |
| Filoboletus hanedae | Japan | ? | + | / | + | / | [44] | |
| Filoboletus pallescens | ♪ | ? | + | ? | ? | ? | [44] | |
| Filoboletus yunnanensis | China | ? | + | ? | ? | / | [54,56,57] | |
| Gerronema glutinipes | Africa and China | ? | + | / | / | / | [54] | |
| Mycena abieticola | Brazil | ? | + | + | + | / | [57] | |
| Mycena aspratilis | Brazil and Puerto Rico | / | + | / | + | / | [57] | |
| Mycena asterina | South America | + | + | + | / | / | [44] | |
| Mycena cahaya | ♪ | + | + | + | + | / | [58] | |
| Mycena chlorophos | China, Japan, Pacific islands, Sri Lanka, and ♪ | + | + | + | + | / | [44,55] | |
| Mycena citricolor | South America and USA | + | / | / | / | / | [44] | |
| Mycena coralliformis | ♪ | + | / | / | / | / | [55] | |
| Mycena cristinae | Brazil | + | + | / | / | / | [59] | |
| Mycena crocata | Switzerland | + | / | / | / | / | [60] | |
| Mycena daisyogunensis | Japan | ? | + | ? | ? | / | [44] | |
| Mycena deeptha | India | + | / | / | / | / | [61] | |
| Mycena deformis | Brazil | + | / | / | / | / | [62] | |
| Mycena discobasis | Africa and South America | ? | + | + | + | / | [44] | |
| Mycena epipterygia | Europe, USA, and Japan | + | / | / | / | / | [44] | |
| Mycena fera | South America | ? | + | + | + | / | [44] | |
| Mycena flammifera | Japan | + | + | + | + | / | [52,53] | |
| Mycena fulgoris | Mexico | / | + | / | + | / | [63] | |
| Mycena galopus | Europe, USA, and Japan | + | / | / | / | / | [44] | |
| Mycena globulispora | Brazil, Mexico | ? | + | / | + | / | [62,63] | |
| Mycena gombakensis | ♪ | + | + | + | + | / | [55] | |
| Mycena guzmanii | Mexico | + | + | + | + | / | [63] | |
| Mycena haematopus | China, Europe, USA, Japan, and South America | + | + | + | / | / | [44] | |
| Mycena illuminans | Japan and ♪ | ? | + | + | / | / | [44,55] | |
| Mycena inclinata | Africa, China, Europe, and USA | + | / | / | / | / | [44] | |
| Mycena jingyinga | China | + | / | / | / | / | [29] | |
| Mycena kentingensis | China | + | + | + | / | / | [63] | |
| Mycena lacrimans | South America | ? | + | / | + | / | [44] | |
| Mycena lamprocephala | Brazil | + | + | + | + | ? | [64] | |
| Mycena lazulina | Japan | + | + | + | + | / | [52,53] | |
| Mycena luceata | Mexico | ? | + | + | / | ? | [19] | |
| Mycena luciferina | Mexico | ? | + | + | / | ? | [19] | |
| Mycena lucisnieblae | Mexico | + | / | / | / | ? | [19] | |
| Mycena luguensis | China | + | / | / | / | / | [29] | |
| Mycena lumina | Mexico | + | + | + | + | / | [63] | |
| Mycena luxaeterna | Brazil | + | + | / | + | / | [57] | |
| Mycena luxarboricola | Brazil | ? | + | + | + | / | [57] | |
| Mycena lux-coeli | Japan | ? | + | + | + | / | [44] | |
| Mycena luxfoliata | Japan | + | / | / | / | / | [52,53] | |
| Mycena luxfoliicola | Mexico | + | + | + | + | / | [63] | |
| Mycena luxmanantlanensis | Mexico | + | + | + | / | ? | [19] | |
| Mycena luxperpetua | Puerto rico | + | + | + | + | / | [44] | |
| Mycena maculata | Africa, Europe, and USA | + | / | / | / | / | [44] | |
| Mycena margarita | Belize, Dominican Republic, Jamaica, Puerto rico, Brazil | / | + | + | + | / | [44,65] | |
| Mycena nebula | Mexico | ? | + | + | + | / | [63] | |
| Mycena nocticaelum | ♪ | + | + | + | / | / | [55] | |
| Mycena noctilucens | Pacific islands and ♪ | ? | + | + | + | / | [44,55] | |
| Mycena oculisnymphae | Brazil | / | + | + | + | / | [62] | |
| Mycena olivaceomarginata | Europe and USA | + | / | / | / | / | [44] | |
| Mycena perlae | Mexico | / | + | + | / | / | [63] | |
| Mycena polygramma | China, Europe, USA, Japan, and Africa | + | + | / | / | / | [44] | |
| Mycena pseudostylobates | Japan | + | ? | ? | ? | / | [44] | |
| Mycena pura | China, Europe, USA, Japan, and South America | + | / | / | / | / | [44] | |
| Mycena rosea | Europe | + | / | / | / | / | [44] | |
| Mycena roseoflava | New Zealand | + | + | / | + | / | [47] | |
| Mycena sanguinolenta | China, Europe, USA, and Japan | + | / | / | / | / | [44] | |
| Mycena seminau | ♪ | + | + | + | / | / | [58] | |
| Mycena silvaelucens | ♪ | ? | + | + | + | / | [44,58] | |
| Mycena sinar | ♪ | + | + | + | + | / | [58] | |
| Mycena singeri | South America and ♣ | ? | + | + | + | / | [44] | |
| Mycena sophiae | Mexico | + | / | / | / | ? | [19] | |
| Mycena sp. (PDD 80772) | New Zealand | ? | + | / | / | / | [44] | |
| Mycena sp. (SP #380150) | South America | + | + | / | / | / | [44,66] | |
| Mycena sp. (SP #380281) | South America | ? | + | / | / | / | [44,66] | |
| Mycena stellaris | Japan | + | + | + | + | / | [52,53] | |
| Mycena stylobates | Africa, China, Europe, USA, and Japan | + | / | / | / | / | [44] | |
| Mycena tintinnabulum | Europe | + | / | / | / | / | [44] | |
| Mycena venus | China | + | / | / | / | / | [29] | |
| Mycena zephirus | Europe | + | / | / | / | / | [44] | |
| Panellus luminescens | ♪ | + | + | + | + | + | [44,58] | |
| Panellus luxfilamentus | Sri Lanka and † | + | / | / | / | / | [58] | |
| Panellus pusillus | Africa, Australasia, China, Japan, USA, South America, and ♪ | ? | + | ? | ? | / | [44,51] | |
| Panellus stipticus | Africa, Australasia, China, Europe, Japan, USA, and South America | + | + | + | / | / | [44] | |
| Resinomycena fulgens | Japan | ? | + | + | + | / | [52,53] | |
| Resinomycena petarensis | Brazil | + | / | / | / | / | [62] | |
| Roridomyces irritans | Australasia | / | + | + | / | ? | [44] | |
| Roridomyces lamprosporus | Brazil, Ceylon, Malaysia, Papua New Guinea,Singapore, and Trinidad | / | + | / | / | + | [67] | |
| Roridomyces phyllostachydis | India | ? | + | / | + | / | [18,68] | |
| Roridomyces pruinosoviscidus | Australasia and ♪ | + | + | + | + | ? | [44,55] | |
| Roridomyces roridus | China, Europe, USA, South America, and Japan | + | / | / | / | / | [44,69] | |
| Roridomyces sublucens | Indonesia and † | / | + | + | + | / | [44] | |
| Roridomyces viridiluminus | China | + | + | + | + | / | [36] | |
| Omphalotus Lineage | Marasmiellus venosus | Japan | + | + | / | / | / | [52,53] |
| Marasmiellus lucidus | Japan | ? | + | + | + | / | [52,53] | |
| Neonothopanus gardneri | South America | ? | + | + | + | / | [44] | |
| Neonothopanus nambi | Australasia, China, South America, Thailand, ♪ and ♣ | ? | + | + | + | / | [44,55,56] | |
| Nothopanus noctilucens | Japan | ? | + | / | / | / | [44] | |
| Omphalotus guepiniiformis | China and Japan | + | + | / | / | / | [44,56,70] | |
| Omphalotus illudens | Europe and USA | + | + | + | / | / | [44] | |
| Omphalotus mangensis | China | ? | + | + | / | / | [44,56,71] | |
| Omphalotus nidiformis | Australasia | ? | + | + | + | / | [44] | |
| Omphalotus olearius | China and Europe | + | + | + | + | / | [44,56] | |
| Omphalotus olivascens | USA | / | + | / | / | / | [44] | |
| Omphalotus subilludens | USA | ? | + | / | / | ? | [72] | |
| Pleurotus decipiens | ♪ | ? | + | / | / | / | [44] | |
| Pleurotus nitidus | Japan | ? | + | / | / | / | [52,53] | |
| Ascomycota | Xylaria hypoxylon | Europe | ? | + | ? | ? | / | [44] |
Note: For Armillaria novae-zelandiae, Mycena roseoflava, and Omphalotus subilludens, there are no published literature as bioluminescent mushrooms, however, mushroom hunters have posted glowing mushroom photos of those species. ♪ = Refers to distribution within Malaysia, south Asia region; ♣ = Refers to distribution within Central America and the Caribbean region; + = glow; / = don’t glow;? = no reports.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



