Submitted:
05 June 2024
Posted:
10 June 2024
You are already at the latest version
Abstract
Cysteamine HCl has been established as a potent ulcerogenic agent of the gastrointestinal (GI) system. GI dysfunction and olfactory deficits are the most common clinical symptoms of many movement disorders including Parkinson's disease (PD). Cysteamine HCl has been shown to interfere with dopamine, a neurotransmitter crucial for motor, olfactory, and cognitive functions. However, the reports on the effect of cysteamine HCl treatment on the behavior and dopamine system appear to be inconsistent. Therefore, we revisited the impact of cysteamine HCl on motor function in experimental mice using a battery of behavioral tests such as pole test (PT), beam walking test (BWT), and rotarod test (RDT), while the olfactory ability and cognitive functions were examined through food buried test (FBT) and Y maze. Furthermore, we investigated the effect of cysteamine HCl on the density of dopaminergic tyrosine hydroxylase (TH)-positive cells in the substantia nigra (SN) and olfactory bulb (OB) of experimental mice using immunohistochemistry. Results revealed that cysteamine HCl administration in mice induced significant impairments in motor balance and coordination, as their movement-related performance was markedly reduced in behavioral tasks. Mice exposed to cysteamine HCl showed a pronounced reduction in odor discrimination ability and cognitive impairments. Strikingly, the number of TH-positive neurons was found to be reduced in the SN and OB of the cysteamine HCl-treated group which is a bonafide neuropathogenic hallmark of PD. This study highlights the potential neurotoxic effects of cysteamine HCl in experimental brains and suggests further investigation into its role in pathogenesis of Parkinsonism.
Keywords:
Cysteamine HCl
; motor coordination
; olfaction
; rotarod
; tyrosine hydroxylase
; Parkinsonism
1. Introduction
Cysteamine HCl is a potent ulcerogenic agent, as it triggers the hypersecretion of hydrochloric acid (HCl) in the stomach, leading to gastritis and peptic ulcers [1]. The physiological levels of intestinal alkaline phosphatase play a crucial role in maintaining homeostasis and supporting the immune defence mechanism of the gastrointestinal (GI) tract [2]. Studies have shown that cysteamine-HCl can counteract the activity of alkaline phosphatase in mucosal cells of the gut. When alkaline phosphatase activity is inhibited in the GI tract, it can lead to dysbiosis and prolonged intestinal inflammation, followed by systemic activation of immune cells [3]. The pro-inflammatory factors discharged from the activated immune cells in the circulation can subsequently produce a negative impact on the brain [4]. As the aberrant circadian rhythm often coincides with GI disorders, cysteamine HCl-induced pathophysiology of the GI tract can result in sleep disorders leading to mood, neurocognitive, and neurological disorders [5,6]. Meanwhile, ulceration in the GI tract caused by cysteamine-HCl can also deteriorate the gut-brain axis contributing to behavioral and neurophysiological abnormalities [7,8]. As cysteamine HCl can cross the blood-brain barrier (BBB), it has become more apparent that it can also cause anxiety and neurocognitive impairments [9]. Furthermore, few animal studies have shown that cysteamine HCl treatment can provoke neurotoxicological effects similar to 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP), an authenticated neurotoxin known to induce the clinical symptoms of Parkinson's disease (PD) [7,10]. Cysteamine HCl competitively antagonizes various ion channels and neurotransmitter systems due to its structural resemblance to ethylamine, a precursor of norepinephrine, dopamine, and serotonin that serve as crucial neuromodulators of stress response, motor, mood, and cognitive functions [10,11]. In particular, cysteamine HCl-mediated GI dysfunction in association with alteration of the dopamine system has been an intense research focus [7]. Meanwhile, the deleterious effects of cysteamine HCl on neuroplasticity responsible for movement disorders have also been recognized, but to a lesser extent [12]. Considering its dopamine depletion capacity, few studies indicated that cysteamine HCl treatment induces cognitive deficits, while its effects on locomotive behavior remain inconclusive [9,12,13]. Notably, reduced levels of dopamine have been linked to underlying causes of movement disorders seen in prevalent neurodegenerative diseases such as PD [14,15]. Notably, many movement disorders have also been characterized by olfactory deficits, suggesting a potential neuropathogenic overlap between movement disorders and OB deficits[15,16,17]. Hence, it can be proposed that cysteamine HCl treatment may be associated with movement disorders and olfactory deficiencies due to its negative impact on the brain's dopamine system. Despite accumulating evidence suggesting a potential link between cysteamine HCl treatment and neurological functions, reports on its effects on motor function and the dopaminergic system remain inconsistent. Moreover, the effect of cysteamine HCl on olfactory functions has not been elucidated. Therefore, we revisited the effects of cysteamine HCl administration on motor, cognitive, and olfactory functions in experimental mice using a battery of behavioral paradigm. We also extended our investigations to assess its possible impact on the density of tyrosine hydroxylase (TH)-positive cells, the main dopamine-producing cells in the substantia nigra (SN), and the glomerular layer of the olfactory bulb (OB) using immunohistochemistry.
2. Materials and Methods
2.1. Animals and Treatment
Four to five-month-old male BALB/c strain mice were procured from Biogen laboratory, Bangalore, India. The animals were maintained in identical conditions at a temperature of 22-24°C and a 12-hour light-dark cycle in the animal house facility at Bharathidasan University. The mice (N=12) were divided into two experimental groups: Control (N=6) received normal water, while the Cysteamine HCl group (N=6) received intraperitoneal injections of 60 milligram (mg)/ kilogram (kg) body weight (BW) of cysteamine HCl for three alternative days for a total of three doses. After 14 days, all mice were subjected to behavioral tests such as the pole test, beam walking test, rotarod test, food buried test, and Y-maze and then perfused. The brains were collected and further processed for immunohistochemical analysis. All animal experiments were conducted following approval from the Institutional Animal Ethical Committee (IAEC), Bharathidasan University, under the regulations of the Committee for the Purpose of Control and Supervision of Experiments on Animal (CPCSEA), India (Ref No. BDU/IAEC/P10/2019 dated 30.11.2019).
2.2. Pole Test
A vertical pole measuring 60 centimeters (cm) in height and a diameter of 1.9 cm was used to assess motor function and coordination in the experimental mice. The base of the pole was positioned in a new cage and filled with fresh husk bedding. Each animal was gently placed onto the top of the pole, facing upward. The time taken to reorient themselves downwards (T-time) and the total time taken to descend to the bottom of the pole (T-total) were measured. Experimental animals that underwent 3 successful trials within 3 minutes duration were calculated.
2.3. Beam Walking Test
A beam consisting of a 1-meter length and 1.9 cm width raised 50 cm above the ground level was used to study motor coordination and gait in experimental mice. An empty cage was placed at the endpoint. The ground under the beam was spread with a spongy material to avoid accidental injuries. Initially, the mice were trained to acclimatize with an elevated beam in the training session. Then each animal was gently released on the beam and the time taken to cross the beam and the number of times falling were noted down. A total of three successful trials from each animal were considered to calculate latency to complete the task.
2.4. Rotarod Test
Next, the motor coordination and balance in experimental mice were evaluated using a semiautomated rotarod apparatus (Inco, Ambala, India). Initially, each mouse was placed onto the rotating rod gently and trained to walk at a speed of 3 revolutions per minute (rpm). During the test phase, the speed was gradually raised from 3 rpm to 25 rpm every 10 seconds within the span of 2 minutes, and latency to fall from the rotating rod was recorded.
2.5. Buried Food Test
In order to measure the ability of olfaction, experimental mice were subjected to buried food test. Prior to the experiments, the food pellets were removed from their home cage, and the animals were deprived of food for 48 hours. Then animals were exposed to a tiny portion of chocolate-flavored food. Two hours later, the chocolate-flavored food was kept underneath at the mid-end of a rectangular box filled with clean bedding 3 inches in height. The test arena was digitally divided into two zones using SMART 3.0 video tracking module. While the blue-colored zone 1 designated the entire area, the food-buried area was marked as green-colored zone 2. The animals were released into the arena and given 5 minutes to explore for three consecutive trials. The time taken by the animals to collect the buried food was recorded as latency.
2.6. Y Maze
To assess the exploration-based spatial memory in mice, a Y maze apparatus was digitally, divided into four zones namely zone 1, the middle area marked in a triangular-shaped pink color, the familiar areas zone 2 and zone 3 which were marked as blue color, and brown color respectively. The unfamiliar area, zone 4 was marked as green color. In the training phase, the green color zone 4 was closed to prevent entry. Each animal was placed in zone 1 and allowed to explore all areas for 5 minutes excluding zone 4 and the animals received three trials. The next day, the barrier to enter zone 4 was removed and each mouse was released into zone 1. The exploratory tendency of animals in all the sessions were captured using the SMART 3.0 video tracking module and the time spent by each mouse in an unfamiliar zone 4 was calculated.
2.7. Immunohistochemical Assessment of TH Positive Dopaminergic Neurons in the SN and OB
The animals were anesthetized and perfused using 0.9% saline followed by 4% paraformaldehyde (PFA) (Himedia, India), and the brains were soaked in PFA for 24 hours. Further, the brains were stored in a 30% sucrose (SRL, India) solution at 4◦C for a week. Then the brains were cut into 30 micrometer (µm) sagittal sections using a dry ice-based sliding microtome (Weswox, India) and stored in a cryoprotectant solution. For the immunohistochemical analysis, 1 out of 12 brain sections were taken in a 12-well plate (Tarson, India) and washed thrice with 1× Tris-buffered saline (TBS) for 10 minutes, for antigen retrieval, the sections were treated with 10 millimolar (mM) sodium citrate buffer (Thermo Fisher Scientific. Waltham, MA, USA) at 65◦C for 90 minutes. After incubation, the sections were washed thrice with 1× TBS for 10 minutes. Then, the sections were blocked with 3% bovine serum albumin (BSA) (Himedia, India), for an hour before being treated with the primary antibody. The brain sections were transferred to a solution containing a rabbit α- TH antibody with the dilution of 1:250 (Cell Signaling Technology, Danvers, MA, USA) and incubated at 4◦C for 48 hours. After incubation with the primary antibody, the sections were washed thrice with 1× TBS for 10 minutes each. Further, brain sections were incubated with a goat α rabbit DyLightTM 594 secondary antibody with the dilution of 1:500 (Novus Biologicals, Colorado, USA), kept at 4◦C for 24 hours. The next day, the sections were washed twice with 1× TBS for 10 minutes. Further, the sections were placed on the microscopic slides and dried. The next day the specimens were sealed with ProLongTM Glass antifade mountant (Thermo Fisher Scientific, Waltham, MA, USA) and dried overnight. The slides were blind-coded, and the brain sections were analyzed and photographed using a fluorescence microscope (DM750, Leica Microsystems, Wetzler, Germany). The number of TH-positive cells was estimated in the SN and glomerular layer of OB in five brain sections from each animals using the ImageJ plugin with a cell counter. The average number of cells per section was calculated and compared among the control and Cysteamine HCl traded groups
2.8. Statistical Analysis
The values have been represented as mean ± standard deviation (SD). All the statistical analyses were made using the Graph pad prism. The statistical significance was assumed at p< 0.05 unless otherwise indicated.
3. Results
3.1. Cysteamine HCl Treated Mice Exhibited Delayed Motor Functions during the Pole Test
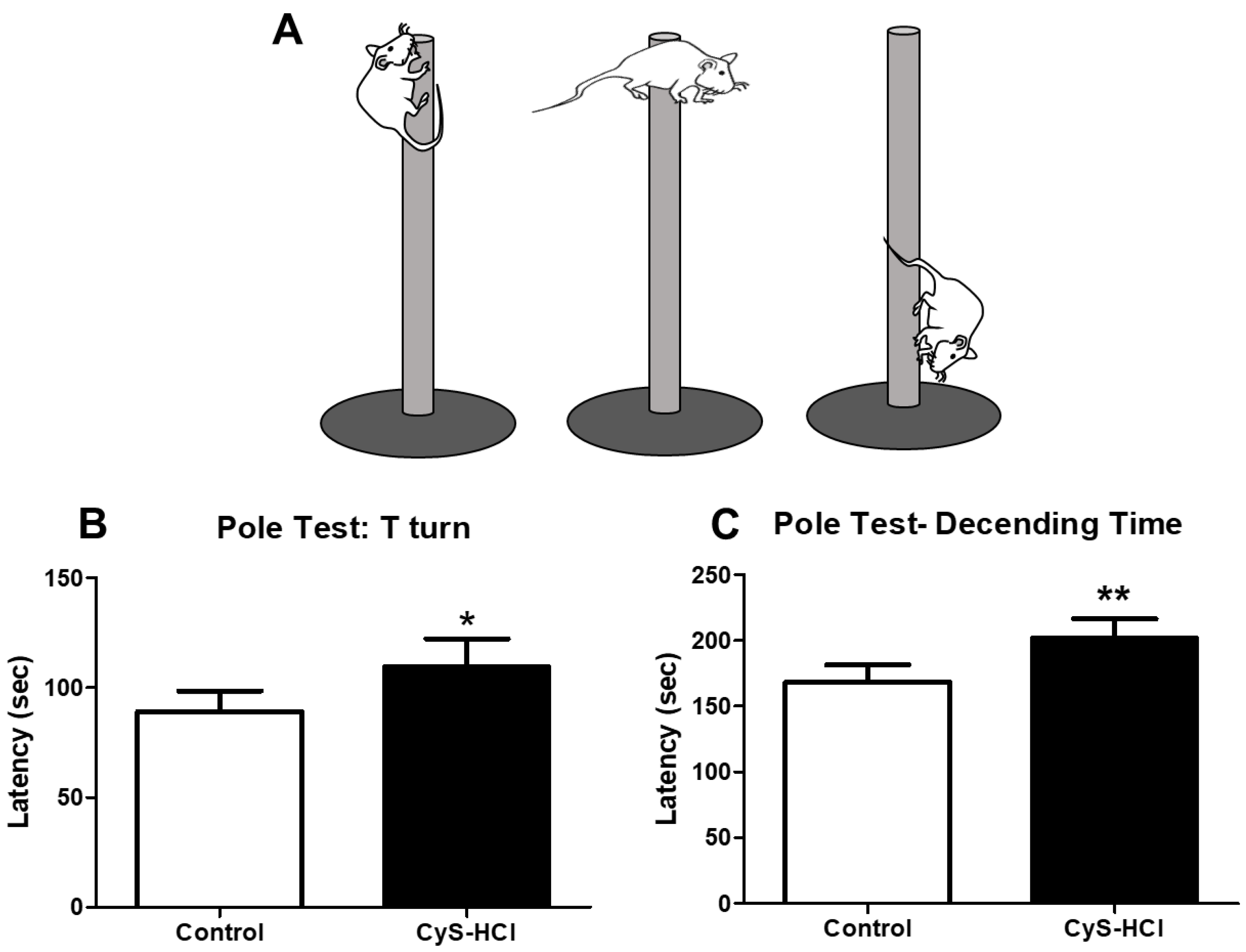
The pole test determines the movement disorder in the experimental animals in which the time taken by the animals to face the downward position is considered as T-turn. The cysteamine HCl-treated group displayed a significant delay in latency to orient downward when compared with the control group (Control 89.05 ± 9.50 Vs Cys-HCl 110±13). Subsequently, the latency to reach the base of the pole is significantly increased in the cysteamine HCl group than that of the control group (Control 168 ± 13 Vs Cys-HCl 201±15). Overall, the increased latency in both parameters for the cysteamine HCl-treated group suggests that cysteamine HCl impairs motor coordination and movement in the experimental animals. This could be due to its effects on the nervous system or specific brain regions responsible for motor control.
Figure 1.
Impaired the motor coordination upon Cysteamine HCl treatment in pole test. (A) Diagrammatic representation of motor activities of animals in the pole test (B) Bar graph depicts the latency of T-turn and (C) Latency of descending time by the experimental animals.
Figure 1.
Impaired the motor coordination upon Cysteamine HCl treatment in pole test. (A) Diagrammatic representation of motor activities of animals in the pole test (B) Bar graph depicts the latency of T-turn and (C) Latency of descending time by the experimental animals.
3.2. Cysteamine HCl Treated Mice Showed Impaired the Motor Coordination and Balance in Beam Walking Test
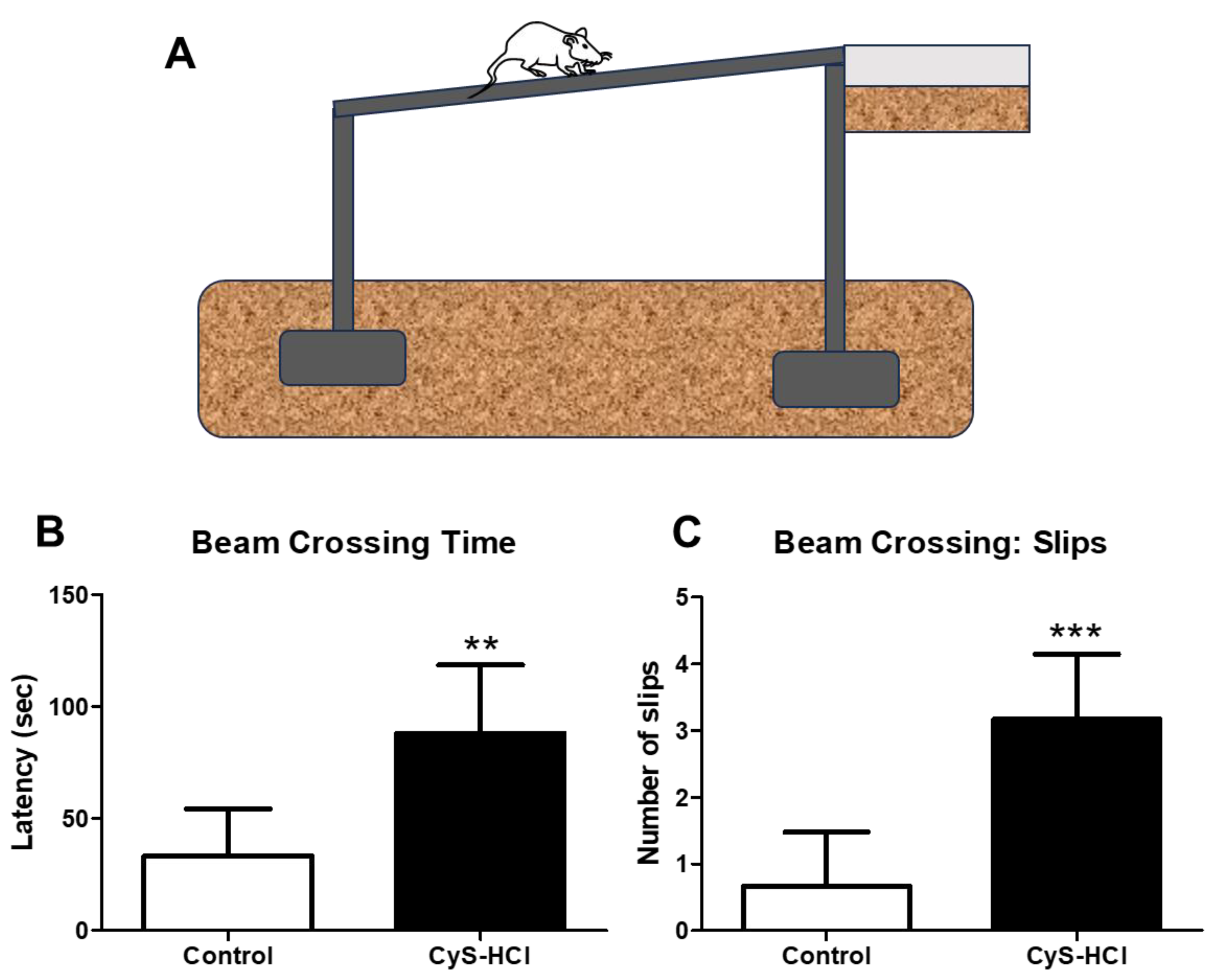
The beam walking test was employed to assess the motor coordination and balance of the experimental animals. The total time taken by the cysteamine HCl to traverse the raised beam was significantly increased when compared with the control group (Control 33 ± 21 Vs Cys-HCl 88±30). During the beam crossing, there was a notable increase in the number of foot slips observed in the cysteamine HCl-treated animals compared to the control group (Control 0.7 ± 0.8 Vs Cys-HCl 3 ±0.9). The outcome of the balance beam test indicates that cysteamine HCl impairs both motor coordination and balance in the experimental animals.
Figure 2.
Cysteamine HCl treatment impaired motor coordination and gait in the beam walking test. (A) Image depicts the beam walking test. (B) Bar graph indicates the latency to cross the beam and (C) Bar graph shows the number of foot slips during the beam crossing by the experimental animals.
Figure 2.
Cysteamine HCl treatment impaired motor coordination and gait in the beam walking test. (A) Image depicts the beam walking test. (B) Bar graph indicates the latency to cross the beam and (C) Bar graph shows the number of foot slips during the beam crossing by the experimental animals.
3.3. Cysteamine HCl Treatment Diminished the Locomotor Behavior in the Rotarod Test
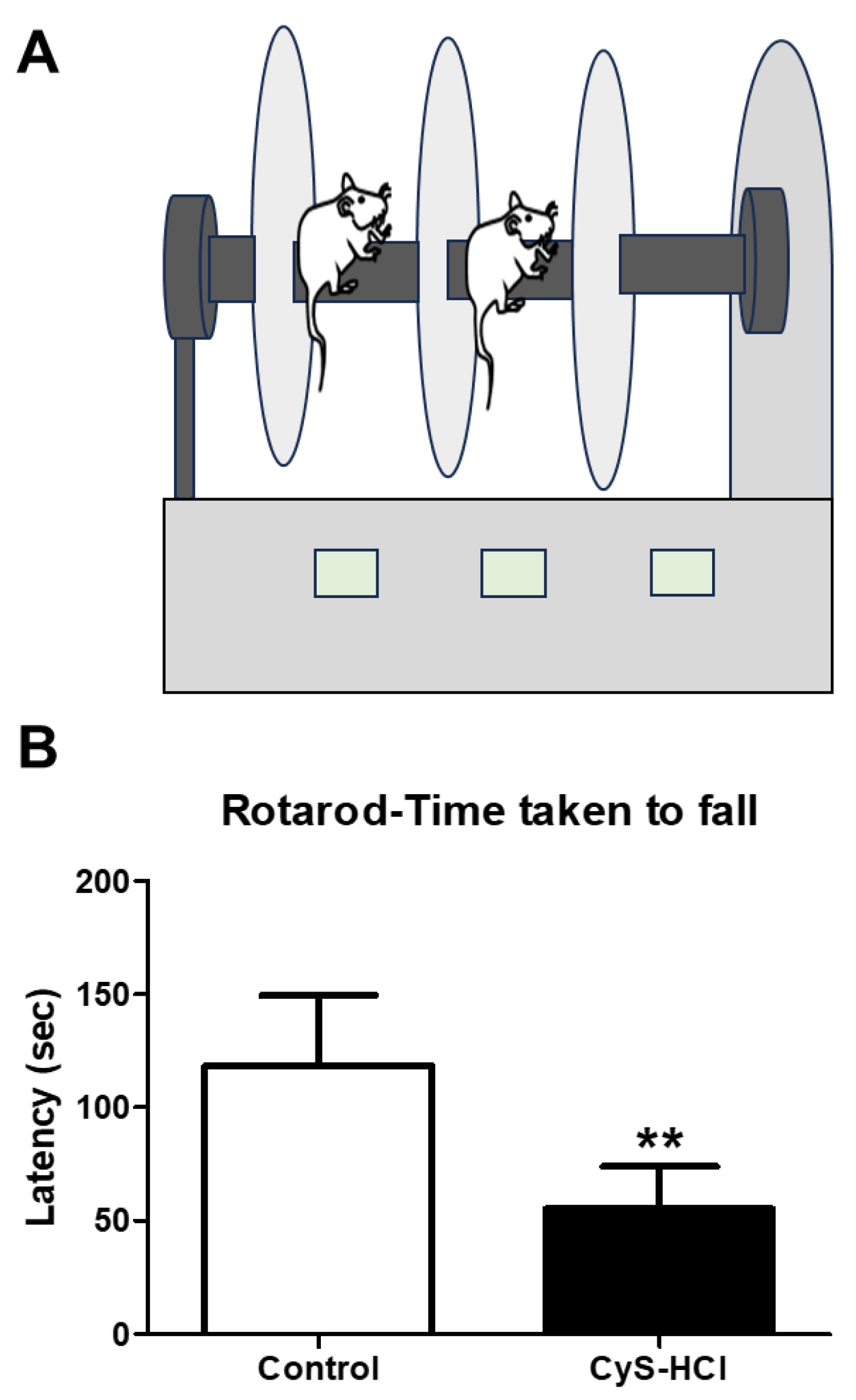
The rotarod test was used to assess the effect of cysteamine HCl on locomotor performance such as motor coordination and balance-based activities. While the control group showed better performance in the rotarod test, the latency on the rotating rod was considerably decreased in cysteamine HCl-treated animals (Control 118 ± 31 Vs Cys-HCl 55 ±18). The results from the rotarod test specify that cysteamine HCl-treated animals were not able to stay on the rod for a longer period, demonstrating poorer performance in terms of motor coordination, balance, and endurance. This observation line ups with the findings from the pole test and beam walking test, which also showed significant impairments in neurological functions responsible for movement and balance in the cysteamine HCl-treated group.
Figure 3.
Cysteamine HCl administration impaired the locomotory performance of mice in the rotarod test. (A) Illustration of the rotarod test and (B) gar graph represent the latency to fall during the rotarod test.
Figure 3.
Cysteamine HCl administration impaired the locomotory performance of mice in the rotarod test. (A) Illustration of the rotarod test and (B) gar graph represent the latency to fall during the rotarod test.
3.4. Cysteamine HCl-Treated Animals Showed Deteriorated Olfactory Behavior
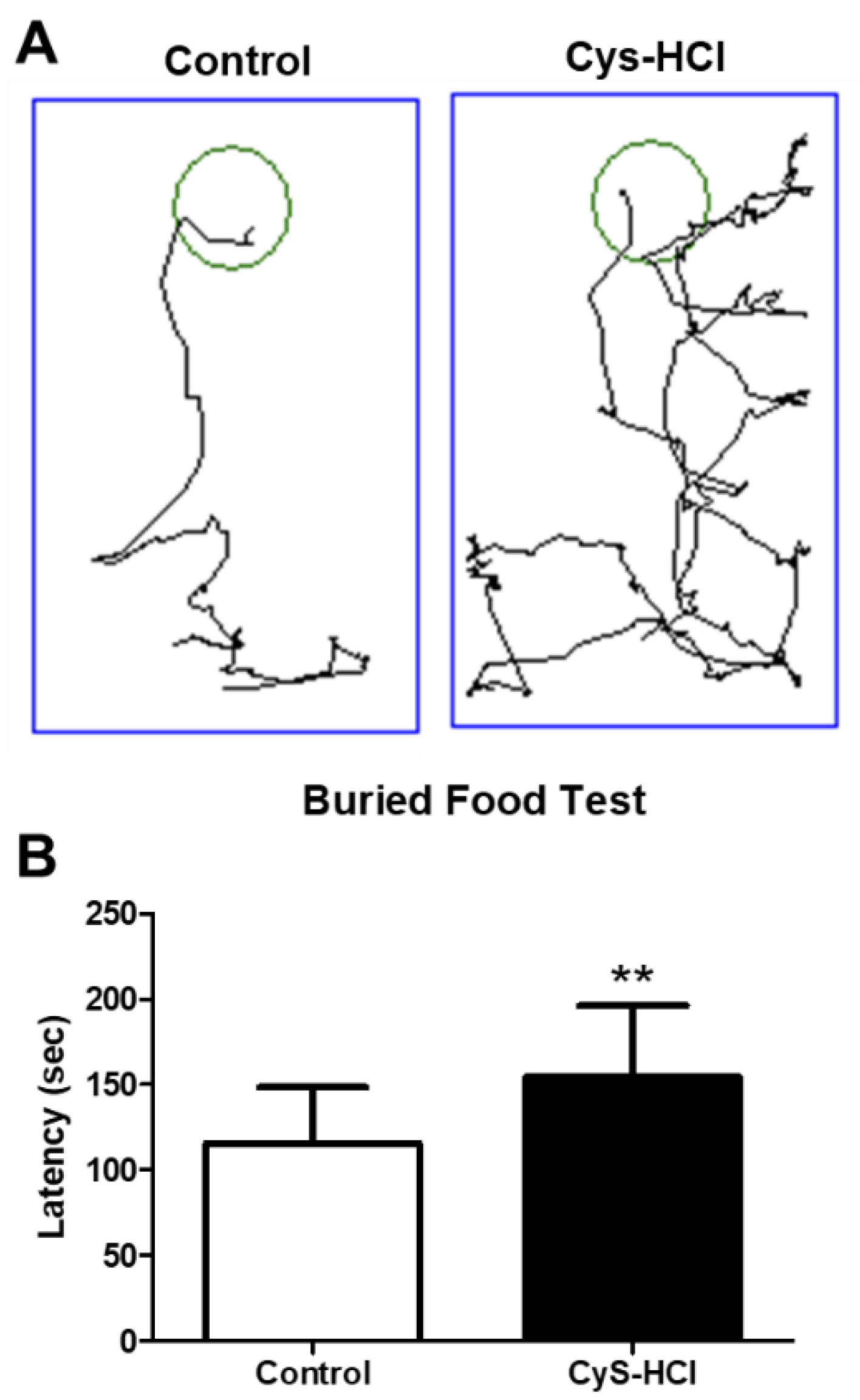
The buried food test was conducted to assess the effect of cysteamine HCl on olfactory function as it indicates the ability and drive of animals to sense the buried food. The cysteamine HCl-treated animals took significantly longer time to locate and retrieve the buried food compared to the control group (Control 116 ± 33 Vs Cys-HCl 154 ±42). This prolonged latency indicates that impediment of cysteamine HCl treatment with neural control of olfaction, as the treated animals were less efficient in using their sense of smell to find the hidden food.
Figure 4.
Cysteamine HCl treatment impeded the olfactory behavior in the buried food test. (A) Representative tracking image of buried food test of control and cysteamine HCl treated mice. The green color shows the hidden food zone and the blue color represents the overall test arena. (B) Bar graph shows the time taken to find the buried food by the experimental animals.
Figure 4.
Cysteamine HCl treatment impeded the olfactory behavior in the buried food test. (A) Representative tracking image of buried food test of control and cysteamine HCl treated mice. The green color shows the hidden food zone and the blue color represents the overall test arena. (B) Bar graph shows the time taken to find the buried food by the experimental animals.
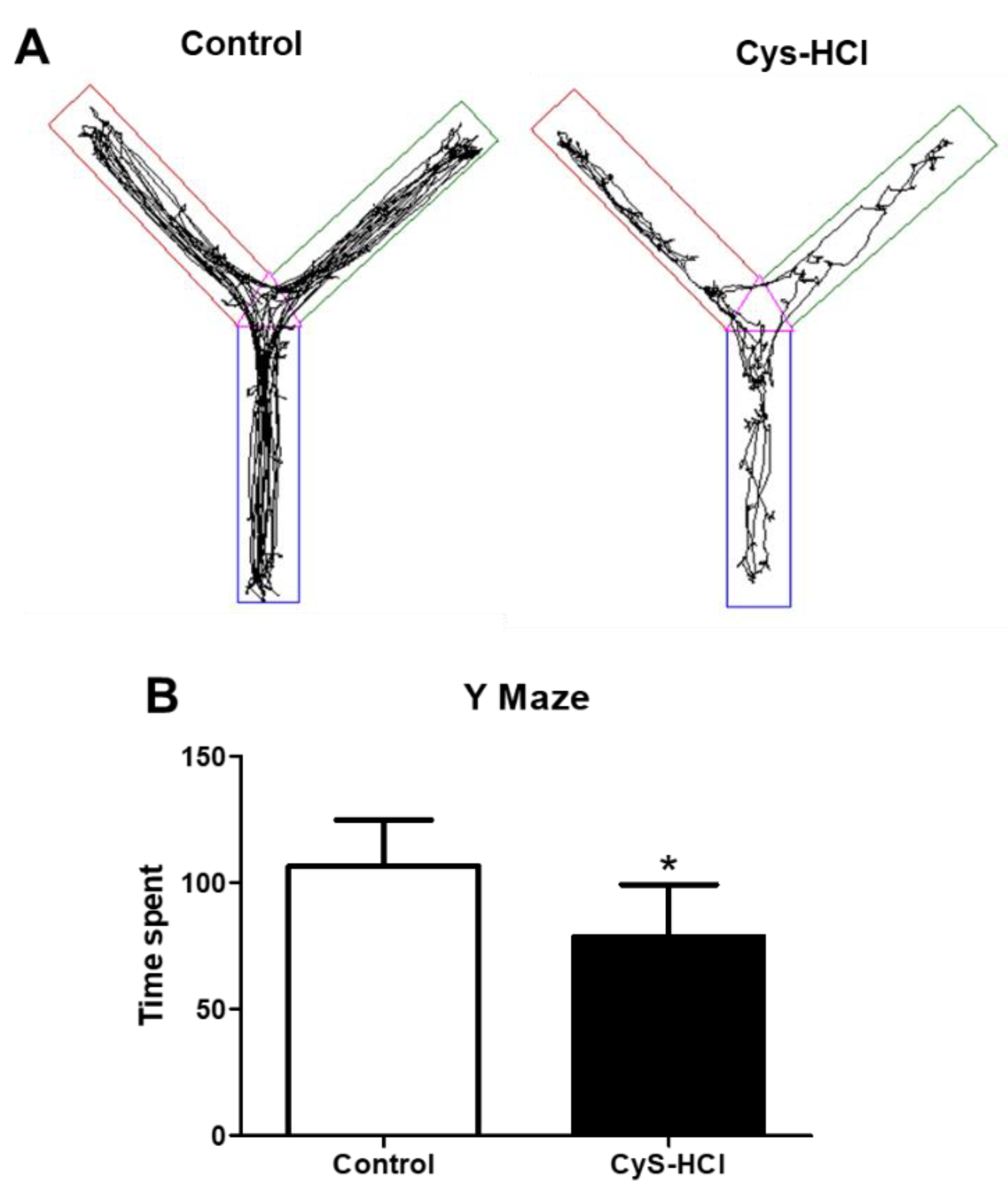
3.5. Impaired Spatial Memory in Cysteamine HCl Treatment during the Y Maze Test
The Y-maze test is used to evaluate the ability of animals to remember and explore new environments based on spatial reference, which is indicative of their cognitive function. In such a task, the control group spent considerable time exploring the unfamiliar zone-4, indicating normal curiosity and memory function. In contrast, the cysteamine HCl-treated animals spent significantly less time in the unfamiliar zone-4, suggesting impaired spatial memory or reduced exploratory behavior (Control 106 ± 18 Vs Cys-HCl 79 ±21). This suggests potential cognitive side effects of cysteamine HCl, highlighting the need for further research into its neurobiological effects and the mechanisms underlying these behavioral changes.
Figure 5.
Impaired spatial memory in cysteamine HCl treated group during the Y maze test. (A) The image represents the video-tracking Y-maze of control and cysteamine HCl-treated mice. The pink color indicates zone 1, the green color represents zone 2 and the novel arm, the brown color indicates zone 3 and the blue color represents zone 4. (B) The Bar graph indicates the time spent by the experimental animals in the novel zone.
Figure 5.
Impaired spatial memory in cysteamine HCl treated group during the Y maze test. (A) The image represents the video-tracking Y-maze of control and cysteamine HCl-treated mice. The pink color indicates zone 1, the green color represents zone 2 and the novel arm, the brown color indicates zone 3 and the blue color represents zone 4. (B) The Bar graph indicates the time spent by the experimental animals in the novel zone.
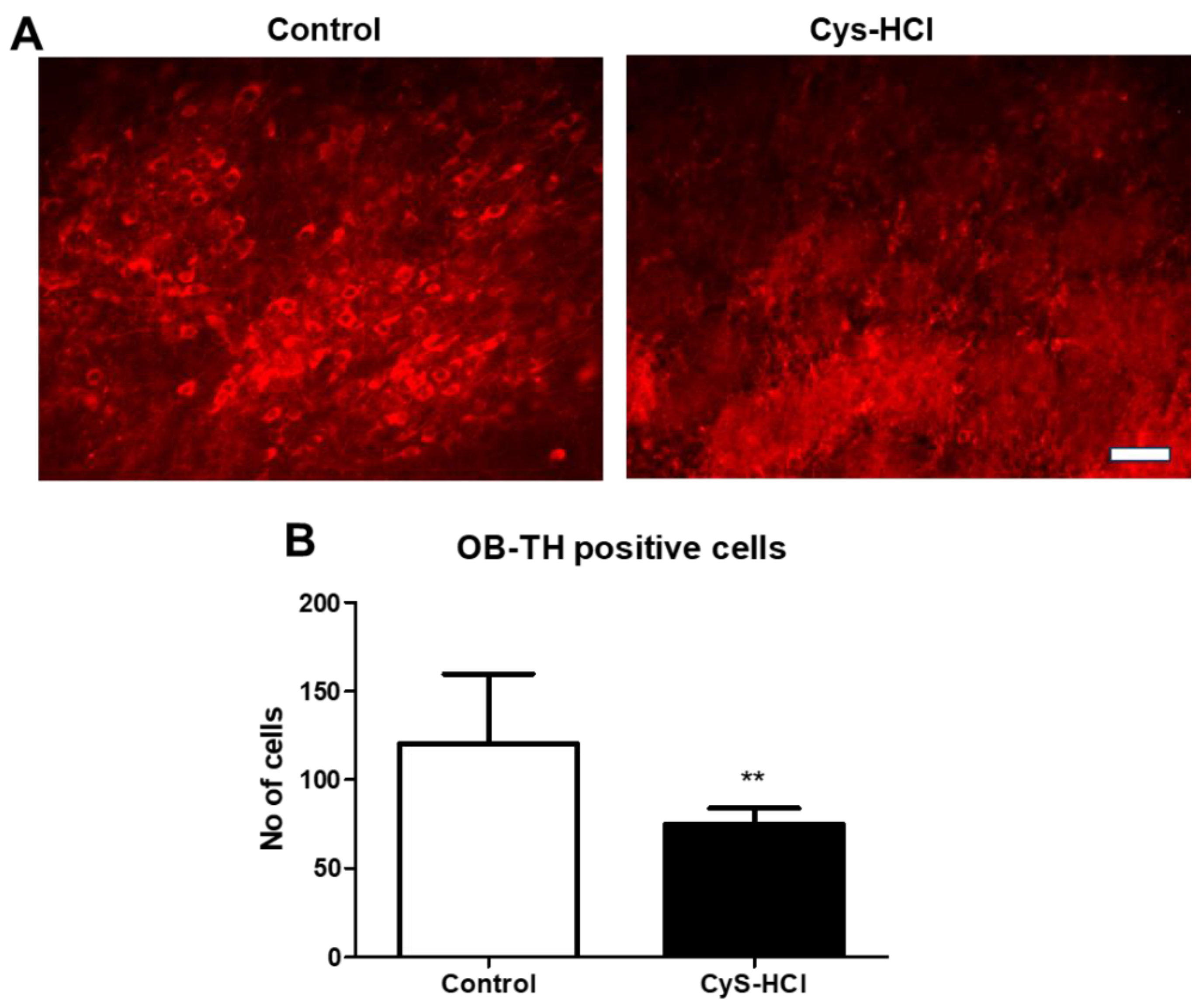
3.6. Cysteamine HCl Treatment Decreased the Number of TH-Positive Cells in the Substantia Nigra and Olfactory Bulb
TH is the rate-limiting enzyme involved in the synthesis of dopamine in the brain. TH-immunopositive cells are indicative of dopaminergic neurons in SN and OB [18]. Dopaminergic neurons play crucial roles in various brain functions, including motor control, motivation, reward processing, and regulation of mood, cognition, and olfaction [19]. Hence, the effect of cysteamine HCl on the number of TH-positive cells was evaluated in the SN and OB. Immunohistochemical assessment revealed that cysteamine HCl treatment significantly decreased the number of TH-positive cells in both the SN (Control 212 ± 33 Vs Cys-HCl 149 ±32) and OB (Control 121 ± 39 Vs Cys-HCl 75 ±9). This indicates that cysteamine HCl treatment could adversely affect the survival of dopaminergic neurons, with potential implications for disease conditions like PD.
Figure 6.
Cysteamine HCl treatment decreased the number of TH-positive cells in the substantia nigra (A) The represented immunofluorescence images of TH-positive cells in the Substantia nigra of control and cysteamine HCl treated mice. (B) Bar graph highlights the number of TH-positive cells in the substantia nigra of the experimental animals. Scale bar 25µm.
Figure 6.
Cysteamine HCl treatment decreased the number of TH-positive cells in the substantia nigra (A) The represented immunofluorescence images of TH-positive cells in the Substantia nigra of control and cysteamine HCl treated mice. (B) Bar graph highlights the number of TH-positive cells in the substantia nigra of the experimental animals. Scale bar 25µm.
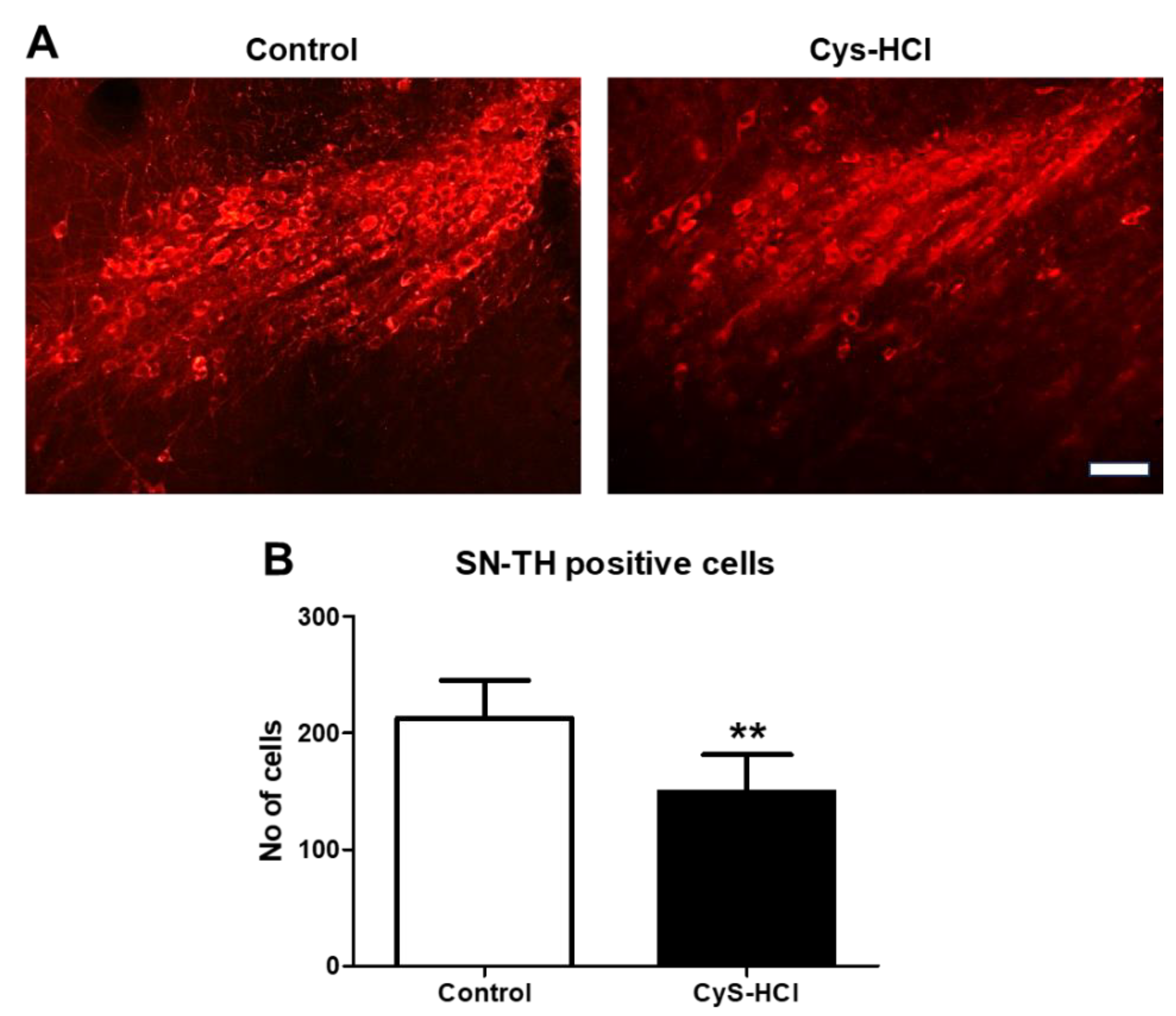
Figure 7.
Cysteamine HCl treatment reduced the number of Tyrosine Hydroxylase (TH) in the glomerular layer of the olfactory bulb (A) Representative immunofluorescence images of TH-positive cells in the glomerular layer of the olfactory bulb of control and cysteamine HCl treated mice. (B) Bar graph shows the number of TH-positive cells in the olfactory bulb of the experimental animals. Scale bar 25µm.
Figure 7.
Cysteamine HCl treatment reduced the number of Tyrosine Hydroxylase (TH) in the glomerular layer of the olfactory bulb (A) Representative immunofluorescence images of TH-positive cells in the glomerular layer of the olfactory bulb of control and cysteamine HCl treated mice. (B) Bar graph shows the number of TH-positive cells in the olfactory bulb of the experimental animals. Scale bar 25µm.
4. Discussion
In an experimental quest to explore the pathogenic interplay between gastrointestinal (GI) disorders and neurological impairments, the current study revisited the deleterious effects of cysteamine HCl, an established ulcerogenic agent, on key neurobiological aspects in experimental mice using a battery of behavioral paradigms and immunohistochemical methods. The present study demonstrates that cysteamine-HCl administration has induced distinguishable movement disorders, olfactory dysfunction, and signs of cognitive impairment in experimental mice, accompanied by a marked reduction in dopamine-producing TH-positive cells in both the SN and OB. Strikingly, these findings suggest a compelling preclinical model of GI disorder induced by cysteamine-HCl, mirrors key symptomatic aspects of PD, including motor deficits, olfactory dysfunction, and dopaminergic neuronal loss. The use of various motor behavioral paradigms, such as the pole test and rotarod confirmed significant impairments in motor coordination and reduced locomotive activity levels. These deficits are likely due to the substantial loss of dopaminergic neurons in the SN, a hallmark of the neuropathology involved in PD.
Cysteamine HCl is an established chemical agent to model GI disorders in laboratory rodents [6,10]. While the exact mechanism for the ulcerogenic property of cysteamine HCl is not completely conferred, multiple experimental evidences suggest that it hyper-stimulates the secretion of HCl and gastrin in the gut leading to inflammation in the GI tract [1]. Moreover, subcutaneous administration of cysteamine in experimental rodents has been reported to decrease concentrations of somatostatin in the brain [20,21,22]. Somatostatin is a key hormone that interferes with the secretion of several other hormones important for growth, physiology, metabolism, and neurological functions [23]. The cysteamine HCl-induced reduced somatostatin-positive cells in the cortical and hippocampal regions of the brain of the Sprague-Dawley rats have been reported to be associated with considerable motor deficits and memory impairments [24]. Besides, Martin-Iverson, M T et al demonstrated local injections of cysteamine in the nucleus accumbens decreased the amphetamine-induced hyper-locomotor activities in association with reduced somatostatin levels [12]. While, the synergistic actions among somatostatin and dopamine on the hormonal regulation of the pituitary have been well established, classical radioimmunoassay-based experiments revealed a drastic reduction in somatostatin in the frontal cortex, hippocampus, and subjects with PD [25]. Moreover, pretreatment of somatostatin in experimental rat models mimicking PD has been reported to show resistance against lipopolysaccharide-induced neurodegeneration in SN by preventing the activation of microglia and accumulation of reactive oxygen species [26]. In a recent findings, we have demonstrated that administration of cysteamine HCl induced anxiety-like phenotype in the experimental animals in correlation with reduced neuronal density and increased activation of microglia cells in the hippocampus [8,27]. While in the intact brain, somatostatin plays a key role in olfaction and mood, affective disorders and olfactory dysfunctions have been established as non-motor clinical symptoms of many neurodegenerative disorders including PD [23,28,29]. Ample experimental evidence indicates that reduced levels of somatostatin can prompt olfactory impairments and anxiety-related disorders [23,28,30]. Therefore, cysteamine HCl-induced motor, reduced olfactory discrimination, and cognitive deficits noticed in the experimental animals could be in part linked to the reduced level of somatostatin in experimental animals.
Numerous research studies have shown that high prevalence of peptic ulcers in patients with anxiety and PD [31,32,33]. Considering that anxiety is the pathogenic overlap of PD, this linkage highlights the intricate interplay between the GI disorder and the pathogenesis of PD [33]. Cysteamine HCl administration has been reported to impair dopamine synthesis in the GI tract resulting in gastric ulceration in association with affective and neurodegenerative disorders [7]. Degeneration of dopamine-producing neurons in SN and OB is a prominent neuropathogenic hallmark of PD [34,35]. As a consequence, reduced levels of dopamine in the brain have been directly linked to motor deficits, olfactory dysfunction, and memory loss in PD [36]. Interestingly, S Szabo and CH Cho reported that cysteamine HCl exerts neurotoxicological effects similar to dopaminergic neurotoxin MPTP [10]. The conversion of MPTP into the toxic metabolite MPP+ by monoamine oxidase-B in astrocytes, followed by the accumulation of MPP+ in dopaminergic neurons due to its high affinity for dopaminergic transporter in SN in turn depletes ATP and increases oxidative stress, leading to neuronal degeneration [37]. Adequate experimental evidence supports the monoamine oxidase inhibitory effect of cysteamine HCl [38]. PD has been linked to the inhibition of mitochondrial monoamine oxidase, which might be highly relevant in the context of reduced levels of dopaminergic neurons in the brain of cysteamine HCl-treated mice [38,39]. Meantime, cysteamine administration-induced reduction of antioxidants in the experimental brains has become incorrectly evident [27]. The anticancer effect of cysteamine has been reported to be associated with mitochondrial degeneration linked to the accumulation of iron and this event activates peroxidase-positive autophagosomes, ultimately leading to cell death [40,41]. Considering these facts, cysteamine HCl can be proposed to induce iron-dependent, lipid peroxidation-driven ferroptosis, a mechanism that could be linked to the neurodegenerative processes observed in PD [42]. Moreover, Cysteamine-HCl is known to be a potent depigmenting molecule that reduces melanin synthesis through the inhibition of tyrosinase activity in the skin [43]. Strikingly, dopaminergic neurons generate a neuromelanin-like pigment, structurally similar to melanin, in the substantia nigra, which is associated with dopamine synthesis and is essential for neuroprotection [44]. Notably, the interaction of MPTP with neuromelanin is known to be a specific cause of the degeneration of dopaminergic neurons in the substantia nigra of the brain [45]. Considering the depigmentation nature of Cysteamine-HCl, it can also specifically target neuromelanin-producing dopaminergic neurons in the substantia nigra and olfactory bulb of experimental animals. Therefore, the reduced number of TH-positive dopaminergic neurons observed in the substantia nigra and olfactory bulb of cysteamine HCl-administered mice strongly supports the neuropathogenic changes related to PD.
In contrast, a few studies have shown that cysteamine HCl exhibits therapeutic benefits against transgenic and toxic experimental models of PD. For instance, Alberto Siddu et al. reported that cysteamine HCl treatment increased TH levels and improved motor performance in the transgenic Thy1-α-Syn model of PD, which is characterized by the overexpression of human α-synuclein [46]. However, this specific Thy1-α-Syn mice have been characterized by longer lifespan with no change the number of TH-positive neurons, no clear clinical signs of behavioral deficits, neurodegeneration, or motor neuron pathology [47,48,49]. According to Linjuan Sun et al., a low dose of cysteamine HCl ameliorated MPTP-induced neurodegeneration and oxidative stress, suggesting a neuroprotective role in a mouse model of PD [50]. Indeed, these findings are surprising, given that cysteamine HCl itself has been demonstrated to mimic the neurotoxic effects of MPTP. The reason for the contradictory reports on the effects of cysteamine HCl remains to be established [10]. Potential explanations for this discrepancy could include differences in experimental design, dosage, duration of administration, deferential production of metabolites and validation procedures. Therefore, further studies are needed to clarify these confounding factors and to better understand the underlying mechanisms by which cysteamine HCl exerts its beneficial or neurotoxic effects.
Based on the impartment in motor, olfactory and cognitive behavioral outcomes in correlation with reduced dopamine neurons addressed by immunohistochemistry, the current study advocates the neurotoxicological effects of cysteamine HCl that recapitulates key clinical symptoms and pathological features of PD. The possible adverse effects associated with Cysteamine HCl tenement practiced for various disorder may not be excluded. This study warrants careful consideration and monitoring in clinical settings to ensure safety and optimize therapeutic outcomes related to Cysteamine HCl ad the results reinforce the importance of considering GI defects and-neurological defects related to PD.
5. Conclusion
Cysteamine HCl is a well-established chemical agent for inducing gastric ulcers in experimental animal models. Besides, its depigmentation properties in the skin are also well established, as it interferes with melanin production through antagonizing the activity of TH pathway. Hence, it can also impact neuromelanin in dopamine-producing neurons in the brain. Results obtained from the behavioral studies show that cysteamine HCl treatment impairs motor, olfactory and cognitive functions. In addition, cysteamine HCl administration has been found to the significantly reduced TH-positive cells in the SN and OB in experimental animals. This study provides robust evidence that cysteamine HCl-induced GI disorder in mice results in significant neurobiological impairments, particularly mirroring the dopaminergic deficits and motor dysfunction seen in PD. Eventually, Cysteamine HCl treatment can be considered to model experimental model of PD. At present, the possible adverse effects of cysteamine HCl in humans need to be thoroughly evaluated and monitored to ensure patient safety and optimize therapeutic outcomes.
Authors Contributions
Conceptualization, M. K; methodology, DB. S., A.P; validation, M.K., DB.S, JF.VA; and formal analysis, M.K., DB. S; investigation, M.K; resources; data curation, DB. S., A.P., JF. VA., M.K; writing—original draft preparation, DB. S., A.P; writing review and editing, DB. S., A.P., JF. VA., M.K; visualization, M.K., DB.S; supervision, M.K.; project administration, M.K; funding acquisition, M.K. All authors have read and agreed to publish the manuscript.
Funding Information
M.K has been supported by University Grants Commission-Faculty Recharge Programme (UGC-FRP), New Delhi, India. M.K. acknowledges RUSA 2.0 Biological Sciences, Bharathidasan University for their financial support. M.K gratefully acknowledge UGC-SAP and DST-FIST for the infrastructure of the Department of Animal Science, Bharathidasan University. Divya Bharathi Selvaraj is the recipient of RUSA 2.0 project fellowship (Ref. No. BDU/RUSA 2.0/TRP/BS/Date 22/04/2021). Jemi Feiona Vergil Andrews was supported as a project assistant from the project grant SERB EEQ/2016/000639.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of, Institutional Animal Ethics Committee (IAEC) and approved by the Institutional Ethics Committee of Bharathidasan University, Tiruchirappalli Reg. No. 418/GO/Re/S/01/CPCSEA as protocol no. BDU/IAEC/P10/2019 dated on 30.11.2019.
Data Availability Statement
All data needed to evaluate the conclusions are present in the paper.
Acknowledgement
We express our sincere gratitude to Professor. M. Anusiyadevi Jayachandran, Department of Biochemistry for the help in providing rotarod instrument and insightful suggestions. We thank Professor. K. Emmanuvel Rajan and Professor. R. Thirumurugan, Department of Animal Science for the support and extending the departmental Instrumentation facilities. The authors would like to thank Professor. B. Kadalmani for the valuable suggestion.
Informed Consent Statement
“Not applicable”
Conflicts of Interest
"The authors declare no conflict of interest."
References
- Selye, H.; Szabo, S. Experimental Model for Production of Perforating Duodenal Ulcers by Cysteamine in the Rat. Nature 1973, 244, 458–459. [Google Scholar] [CrossRef] [PubMed]
- Santos, G.M.; Ismael, S.; Morais, J.; Araújo, J.R.; Faria, A.; Calhau, C.; Marques, C. Intestinal Alkaline Phosphatase: A Review of This Enzyme Role in the Intestinal Barrier Function. Microorganisms 2022, 10, 746. [Google Scholar] [CrossRef] [PubMed]
- Bilski, J.; Mazur-Bialy, A.; Wojcik, D.; Zahradnik-Bilska, J.; Brzozowski, B.; Magierowski, M.; Mach, T.; Magierowska, K.; Brzozowski, T. The Role of Intestinal Alkaline Phosphatase in Inflammatory Disorders of Gastrointestinal Tract. Mediators Inflamm. 2017, 2017, 9074601. [Google Scholar] [CrossRef] [PubMed]
- Sochocka, M.; Diniz, B.S.; Leszek, J. Inflammatory Response in the CNS: Friend or Foe? Mol. Neurobiol. 2017, 54, 8071–8089. [Google Scholar] [CrossRef] [PubMed]
- Codoñer-Franch, P.; Gombert, M. Circadian Rhythms in the Pathogenesis of Gastrointestinal Diseases. World J. Gastroenterol. 2018, 24, 4297–4303. [Google Scholar] [CrossRef] [PubMed]
- Szabo, S.; Pihan, G. Development and Significance of Cysteamine and Propionitrile Models of Duodenal Ulcer. Chronobiol. Int. 1987, 4, 31–42. [Google Scholar] [CrossRef]
- Sikiric, P.; Boban Blagaic, A.; Krezic, I.; Zizek, H.; Kalogjera, L.; Smoday, I.M.; Vukovic, V.; Oroz, K.; Chiddenton, H.M.; Buric, S.; et al. From Selye’s and Szabo’s Cysteamine-Duodenal Ulcer in Rats to Dopamine in the Stomach: Therapy Significance and Possibilities. Pharmaceuticals 2023, 16, 1699. [Google Scholar] [CrossRef] [PubMed]
- Sri Rethinavel, H.; Selvaraj, D.B.; Balakrishnan, S.J.; Vergil Andrews, J.F.; Joseph, J.H.M.; Kandasamy, M. Omeprazole Treatment Manifests Anxiolytic Effects in a Cysteamine Hydrochloride Induced Mouse Model of Gastrointestinal Disorder. Heliyon 2022, 8, e09787. [Google Scholar] [CrossRef]
- Justino, L.; Welner, S.A.; Tannenbaum, G.S.; Schipper, H.M. Long-Term Effects of Cysteamine on Cognitive and Locomotor Behavior in Rats: Relationship to Hippocampal Glial Pathology and Somatostatin Levels. Brain Res. 1997, 761, 127–134. [Google Scholar] [CrossRef]
- Szabo, S.; Cho, C.H. From Cysteamine to MPTP: Structure-Activity Studies with Duodenal Ulcerogens. Toxicol. Pathol. 1988, 16, 205–212. [Google Scholar] [CrossRef]
- Terry, L.C.; Craig, R. Cysteamine Effects on Monoamines, Dopamine-Beta-Hydroxylase and the Hypothalamic-Pituitary Axis. Neuroendocrinology 1985, 41, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Martin-Iverson, M.T.; Radke, J.M.; Vincent, S.R. The Effects of Cysteamine on Dopamine-Mediated Behaviors: Evidence for Dopamine-Somatostatin Interactions in the Striatum. Pharmacol. Biochem. Behav. 1986, 24, 1707–1714. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.D.; Snyder, S.H. Therapeutic Applications of Cysteamine and Cystamine in Neurodegenerative and Neuropsychiatric Diseases. Front. Neurol. 2019, 10, 1315. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.D.; Yi, L.X.; Wang, D.Q.; Lim, T.M.; Tan, E.K. Role of Dopamine in the Pathophysiology of Parkinson’s Disease. Transl. Neurodegener. 2023, 12, 44. [Google Scholar] [CrossRef] [PubMed]
- Rethinavel, H.S.; Ravichandran, S.; Radhakrishnan, R.K.; Kandasamy, M. COVID-19 and Parkinson’s Disease: Defects in Neurogenesis as the Potential Cause of Olfactory System Impairments and Anosmia. J. Chem. Neuroanat. 2021, 115, 101965. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, S.; Horie, Y. Association Between Olfactory Impairment and Disease Severity and Duration in Parkinson’s Disease. Mov. Disord. Clin. Pract. 2020, 7, 820–826. [Google Scholar] [CrossRef]
- Bhatia-Dey, N.; Heinbockel, T. The Olfactory System as Marker of Neurodegeneration in Aging, Neurological and Neuropsychiatric Disorders. Int. J. Environ. Res. Public. Health 2021, 18, 6976. [Google Scholar] [CrossRef]
- Best, J.A.; Nijhout, H.F.; Reed, M.C. Homeostatic Mechanisms in Dopamine Synthesis and Release: A Mathematical Model. Theor. Biol. Med. Model. 2009, 6, 21. [Google Scholar] [CrossRef]
- Speranza, L.; di Porzio, U.; Viggiano, D.; de Donato, A.; Volpicelli, F. Dopamine: The Neuromodulator of Long-Term Synaptic Plasticity, Reward and Movement Control. Cells 2021, 10, 735. [Google Scholar] [CrossRef]
- Brown, M.R.; Fisher, L.A.; Sawchenko, P.E.; Swanson, L.W.; Vale, W.W. Biological Effects of Cysteamine: Relationship to Somatostatin Depletion. Regul. Pept. 1983, 5, 163–179. [Google Scholar] [CrossRef]
- Widmann, R.; Sperk, G. Cysteamine-Induced Decrease of Somatostatin in Rat Brain Synaptosomes in Vitro. Endocrinology 1987, 121, 1383–1389. [Google Scholar] [CrossRef] [PubMed]
- Haroutunian, V.; Mantin, R.; Campbell, G.A.; Tsuboyama, G.K.; Davis, K.L. Cysteamine-Induced Depletion of Central Somatostatin-like Immunoactivity: Effects on Behavior, Learning, Memory and Brain Neurochemistry. Brain Res. 1987, 403, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Kumar, U.; Singh, S. Role of Somatostatin in the Regulation of Central and Peripheral Factors of Satiety and Obesity. Int. J. Mol. Sci. 2020, 21, 2568. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, L.W.; Dokla, C.P. Morris Water Task Impairment and Hypoactivity Following Cysteamine-Induced Reductions of Somatostatin-like Immunoreactivity. Brain Res. 1989, 505, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Epelbaum, J.; Ruberg, M.; Moyse, E.; Javoy-Agid, F.; Dubois, B.; Agid, Y. Somatostatin and Dementia in Parkinson’s Disease. Brain Res. 1983, 278, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Bai, L.; Zhang, X.; Li, X.; Liu, N.; Lou, F.; Ma, H.; Luo, X.; Ren, Y. Somatostatin Prevents Lipopolysaccharide-Induced Neurodegeneration in the Rat Substantia Nigra by Inhibiting the Activation of Microglia. Mol. Med. Rep. 2015, 12, 1002. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, D.B.; Vergil Andrews, J.F.; Anusuyadevi, M.; Kandasamy, M. Ranitidine Alleviates Anxiety-like Behaviors and Improves the Density of Pyramidal Neurons upon Deactivation of Microglia in the CA3 Region of the Hippocampus in a Cysteamine HCl-Induced Mouse Model of Gastrointestinal Disorder. Brain Sci. 2023, 13, 266. [Google Scholar] [CrossRef] [PubMed]
- Saiz-Sanchez, D.; Ubeda-Bañon, I.; Flores-Cuadrado, A.; Gonzalez-Rodriguez, M.; Villar-Conde, S.; Astillero-Lopez, V.; Martinez-Marcos, A. Somatostatin, Olfaction, and Neurodegeneration. Front. Neurosci. 2020, 14, 96. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.M.; Koh, S.-B. Many Faces of Parkinson’s Disease: Non-Motor Symptoms of Parkinson’s Disease. J. Mov. Disord. 2015, 8, 92–97. [Google Scholar] [CrossRef]
- Lin, L.-C.; Sibille, E. Reduced Brain Somatostatin in Mood Disorders: A Common Pathophysiological Substrate and Drug Target? Front. Pharmacol. 2013, 4, 110. [Google Scholar] [CrossRef]
- Goodwin, R.D.; Keyes, K.M.; Stein, M.B.; Talley, N.J. Peptic Ulcer and Mental Disorders among Adults in the Community: The Role of Nicotine and Alcohol Use Disorders. Psychosom. Med. 2009, 71, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Çamcı, G.; Oğuz, S. Association between Parkinson’s Disease and Helicobacter Pylori. J. Clin. Neurol. Seoul Korea 2016, 12, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Higinbotham, A.S.; Kilbane, C.W. The Gastrointestinal Tract and Parkinson’s Disease. Front. Cell. Infect. Microbiol. 2024, 13, 1158986. [Google Scholar] [CrossRef] [PubMed]
- Vernier, P.; Moret, F.; Callier, S.; Snapyan, M.; Wersinger, C.; Sidhu, A. The Degeneration of Dopamine Neurons in Parkinson’s Disease: Insights from Embryology and Evolution of the Mesostriatocortical System. Ann. N. Y. Acad. Sci. 2004, 1035, 231–249. [Google Scholar] [CrossRef] [PubMed]
- Maiti, P.; Manna, J.; Dunbar, G.L. Current Understanding of the Molecular Mechanisms in Parkinson’s Disease: Targets for Potential Treatments. Transl. Neurodegener. 2017, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, S.; Arachchige, A.S.P.M. Depletion of Dopamine in Parkinson’s Disease and Relevant Therapeutic Options: A Review of the Literature. AIMS Neurosci. 2023, 10, 200–231. [Google Scholar] [CrossRef] [PubMed]
- Meredith, G.E.; Rademacher, D.J. MPTP Mouse Models of Parkinson’s Disease: An Update. J. Park. Dis. 2011, 1, 19–33. [Google Scholar] [CrossRef] [PubMed]
- Demarco, C.; Coletta, M.; Bombardieri, G. INHIBITION OF PLASMA MONOAMINE OXIDASE BY CYSTEAMINE. Nature 1965, 205, 176. [Google Scholar] [CrossRef] [PubMed]
- Cohen, G.; Farooqui, R.; Kesler, N. Parkinson Disease: A New Link between Monoamine Oxidase and Mitochondrial Electron Flow. Proc. Natl. Acad. Sci. U. S. A. 1997, 94, 4890–4894. [Google Scholar] [CrossRef]
- Fujisawa, T.; Rubin, B.; Suzuki, A.; Patel, P.S.; Gahl, W.A.; Joshi, B.H.; Puri, R.K. Cysteamine Suppresses Invasion, Metastasis and Prolongs Survival by Inhibiting Matrix Metalloproteinases in a Mouse Model of Human Pancreatic Cancer. PLoS ONE 2012, 7, e34437. [Google Scholar] [CrossRef]
- Lee, C.-M. A Review on the Antimutagenic and Anticancer Effects of Cysteamine. Adv. Pharmacol. Pharm. Sci. 2023, 2023, 2419444. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Cui, Y.; Chang, Y.-Z.; Yu, P. Ferroptosis-Related Factors in the Substantia Nigra Are Associated with Parkinson’s Disease. Sci. Rep. 2023, 13, 15365. [Google Scholar] [CrossRef] [PubMed]
- Atallah, C.; Charcosset, C.; Greige-Gerges, H. Challenges for Cysteamine Stabilization, Quantification, and Biological Effects Improvement. J. Pharm. Anal. 2020, 10, 499–516. [Google Scholar] [CrossRef]
- Zecca, L.; Tampellini, D.; Gerlach, M.; Riederer, P.; Fariello, R.G.; Sulzer, D. Substantia Nigra Neuromelanin: Structure, Synthesis, and Molecular Behaviour. Mol. Pathol. 2001, 54, 414–418. [Google Scholar] [PubMed]
- D’Amato, R.J.; Alexander, G.M.; Schwartzman, R.J.; Kitt, C.A.; Price, D.L.; Snyder, S.H. Evidence for Neuromelanin Involvement in MPTP-Induced Neurotoxicity. Nature 1987, 327, 324–326. [Google Scholar] [CrossRef] [PubMed]
- Siddu, A.; David, L.S.; Lauinger, N.; Chen, X.; Saint-Pierre, M.; Alpaugh, M.; Durcan, T.; Cicchetti, F. Beneficial Effects of Cysteamine in Thy1-α-Syn Mice and Induced Pluripotent Stem Cells with a SNCA Gene Triplication. Neurobiol. Dis. 2020, 145, 105042. [Google Scholar] [CrossRef] [PubMed]
- Chesselet, M.-F.; Richter, F.; Zhu, C.; Magen, I.; Watson, M.B.; Subramaniam, S.R. A Progressive Mouse Model of Parkinson’s Disease: The Thy1-aSyn (“Line 61”) Mice. Neurotherapeutics 2012, 9, 297–314. [Google Scholar] [CrossRef]
- Richter, F.; Stanojlovic, M.; Käufer, C.; Gericke, B.; Feja, M. A Mouse Model to Test Novel Therapeutics for Parkinson’s Disease: An Update on the Thy1-aSyn (“Line 61”) Mice. Neurotherapeutics 2023, 20, 97. [Google Scholar] [CrossRef]
- Rathke-Hartlieb, S.; Kahle, P.J.; Neumann, M.; Ozmen, L.; Haid, S.; Okochi, M.; Haass, C.; Schulz, J.B. Sensitivity to MPTP Is Not Increased in Parkinson’s Disease-Associated Mutant Alpha-Synuclein Transgenic Mice. J. Neurochem. 2001, 77, 1181–1184. [Google Scholar] [CrossRef]
- Sun, L.; Xu, S.; Zhou, M.; Wang, C.; Wu, Y.; Chan, P. Effects of Cysteamine on MPTP-Induced Dopaminergic Neurodegeneration in Mice. Brain Res. 2010, 1335, 74–82. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



