
Submitted:
06 June 2024
Posted:
07 June 2024
You are already at the latest version
Abstract
The aim of the study is to obtain a more complete picture of the blood plasma melatonin concentration in the donkey species. To this purpose sampling and statistical processing were carried out in such a way that allowed to establish the annual and daily rhythm. Based on human observations, according to the hypothesis of the authors, the blood plasma melatonin concentration of pregnant individuals rises during late gestational period, before parturition. To confirm this, the melatonin concentration of pregnant and non-pregnant mares was monitored and compared. In regard to the circannual rhythm, the significantly lowest midnight melatonin value (27.67 pg/ml) is typical for the summer solstice. Under consideration of circadian changes, a significantly strongest melatonin production (45.16 pg/ml) can be observed on the night of the winter solstice (p < 0.001). Considering the gestational age, the blood plasma melatonin concentration (around 38 pg/ml) does not change as pregnancy progresses (p = 0.136). The results obtained in the study population of the domestic ass usefully expand the little knowledge previously gathered about the development of blood plasma melatonin concentration of this species.
Keywords:
circannual and circadian rhythm
; gestation age
; domestic donkey
1. Introduction
Melatonin is the regulator of the body's daily and annual physiological cycles, its role is especially important in reproductive processes, both in animals and humans. At the same time, melatonin production, influenced by the duration of illumination, not only determines seasonal and daily rhythm, changes in the reproductive system, but also has antioxidant and immunomodulating properties, and is also involved in some metabolic processes. The rods of the retina of the eye are responsible for black and white and twilight, the cones are responsible for color vision. Melatonin is produced in the pineal gland (epiphysis).
The light stimulus enters from the melanopsin-producing ganglion cells of the retina the suprachiasmatic nucleus of the hypothalamus via the retinohypothalamic tract [1], and then reaches the pineal gland via the sympathetic noradrenergic pathway, where melatonin is synthesized. Light inhibits the production and secretion of the hormone, so the highest concentration of melatonin can be measured during darkness. Melatonin is produced not only in the pineal gland, but extrapineal melatonin has also been detected in various organs: the gastrointestinal tract, liver, kidney, adrenal gland, heart, thymus, gonads, placenta, uterus, platelets, white blood cells and can also be found in other cells of the immune system. The synthesis of melatonin in the mitochondria of eukaryotic cells represents a unique protection against oxidative damage and plays an important role in maintaining the physiological functioning of cells.
The melatonin produced in the pineal gland modulates the functioning of the pituitary gland, including the production of oxytocin, vasopressin, and prolactin. In addition, melatonin affects sexual development and reproductive processes and activates the hypothalamus-pituitary-ovary axis (HPO) system [2]. The presence of melatonin is sensed by its receptors on the surface of the cells of both central nervous and peripheral tissues. Of the two types of melatonin receptors (MT1 and MT2) in mammals, MT1 is significantly more important in melatonin-controlled reproductive processes [3].
The short day in horses and donkeys inhibits the reproductive cycle through the HPO axis. During the non-estrous period, the hypothalamus GnRH- [4] and the pituitary gland LH-secretion [5] is low. The production of melatonin is an important indicator of the maintenance of anestrus. Removal of the pineal gland [6] or surgical transection of its sympathetic innervation [7], by interfering with melatonin production, delays the onset of the breeding season [8].
The domestic donkey (Equus asinus), similar to the horse (Equus caballus), is a seasonally polyestrous animal and has an endogenous circannual reproductive rhythm, the main seasonal changes (winter-summer) have a significant influence on the development of the internal rhythm, for which melatonin is the "calendar information" organization. The main regulator is the increasing daylight in spring, which reactivates the HPO-axis as a seasonal change and results in reduced melatonin concentration [9,10]. At the same time, the natural, physiological breeding season of the donkey lasts from March to May in the Northern Hemisphere, and its formation is influenced by the lengthening of the days, the temperature and the energy content of the feed. Year-round active cycles can be observed at the equator. Both the foaling period (1-2 days) and the pregnancy period (37-38 days) are longer in the donkey mare. Ovulation is spontaneous, it occurs on the 2-4th day oestrus. The first oestrus can be observed 9-11 days after foaling, which can cause the foal's diarrhea [11]. The length of pregnancy is one year, sometimes a little more when jennies becoming pregnant at the early period [12]. It is recommended to take advantage of foaling for conception, especially if we want to keep a short interval between two foalings [13]. Although information on mule pregnancy is sporadic, in practice both donkey and horse mules are infertile due to different parental chromosome numbers. Nevertheless, the mule's melatonin production also shows seasonality; in autumn, higher melatonin concentrations appear in the blood than in spring [14].
The increase in the number of long days and light hours affects not only the triggering of ovulation and the start of the cycle, but also the shedding of the winter coat and the time of foaling. The fetus developing inside the mother's body is initially under the hormonal control of the mother animal and is part of its internal balance. This is how it is affected by the mother's body temperature and the maternal hormones that are able to reach the fetus through the placenta. It is known that some representatives of the prolactin and GH hormone families pass into the mammalian fetus and enter the circulation there. The gradually increasing daily light in spring causes a decrease in melatonin production and an increase in the circulating prolactin concentration in horses [15]. This mechanism has not yet been investigated in donkeys.
In the study by Altinsaat et al. (2009), the melatonin concentrations of horse mares and stallions in June (15h light:9h dark) were lower (23.52 and 17.22 pg/ml), but higher in December (9h light:15h dark) (42.41 and 37.68 pg/ml) [16]. The night serum melatonin concentration in mares is between 10 and 20 pg/ml based on the experience of several authors [17,18,19], with the exception of a few outliers (around 50 pg/ml). In the study conducted by Guillaume et al. (2006), in donkey mares the nocturnal melatonin concentration in autumn was with an average of 90 pg/ml [20]. The same authors reported even higher levels (average 169 pg/ml) in mule mares, confirming the previous evaluations of others [14]. So, donkeys have a higher average plasma melatonin concentration than horses. At the same time, the melatonin concentration in seminal plasma of donkey and horse stallions obtained during the breeding season (March to June) were similar but at low values (2.82 and 2.48 pg/ml, respectively) [21]. Messias et al. (2022), in Brazil, measured the concentration of melatonin in donkey milk collected during morning milking as 4-5 pg/ml [22]. To the best of our knowledge, studies conducted so far on natural blood melatonin concentration in donkeys are limited to these literatures.
The aim of the study is to obtain a more complete picture of the blood plasma melatonin concentration in the donkey species. The authors' assumption is that this species also has cyclicity typical of horses. To this end, sampling and statistical processing were carried out in such a way that the annual and daily rhythm could be established. Based on human observations, according to the other hypothesis of the authors, the blood plasma melatonin concentration of pregnant individuals rises during late gestational period before parturition. To prove this, the melatonin concentration of pregnant and non-pregnant mares was monitored and compared.
2. Materials and Methods
2.1. Study Design
The donkey is regarded as an autochthonous animal species in Hungary. In 2003, its breeding association was found, and its stud book contains about 4000 individuals with limited ancestry information. The utilization of donkeys in Hungary previously was principally linked to the activity of herding and keeping of sheep [23]. The main reasons for using donkeys were their high social ability, for the protection of livestock, use as pack animals and their ability to thrive on poor quality of pasture. This domestic donkey population is heterogenous in appearance, and the breed specific characteristics are less typical than in other European donkey breeds [24]. There is a difference in the body capacity according to the color variants; the grey animals are of significant less bod capacity than the brown and black ones. The height at the withers is about 117 cm on average, however it varies in large range. In order to unify the Hungarian Fallow Donkey breed, a reconstruction program has begun 2021 [25]).
The study was conducted in the nucleus stud of Hungarian Fallow Donkey located in Bőszénfa (Game Management Landscape Centre, Kaposvár Campus, Hungarian University of Agriculture and Life Sciences). The stud farm is located at latitude 46.23 and longitude 17.85. The average temperature in December and June is 2 and 20 degrees Celsius, respectively. The time of sunrise and sunset at the winter solstice is 07:29 and 16:05, and at the summer solstice 04:56 and 20:45. All jennies were kept under free range condition almost year-round and spent the day and night on the pasture. During wintertime their feed was supplemented with hay. They always had access to water.
The day before the sampling, the animals were herded into their free stall barn and kept there until the sampling was completed. For the examination 15 mares were selected randomly at the beginning and the same ones remained for continued monitoring. Sampling took place three times at a predetermined time: June 14, 2022, November 12-13, 2022, and April 16-17, 2023. All three sampling occasions covered a 24-hour period, that is, a new individual was sampled every hour and a half.
The number of animals and sampling ensures the scientific value of the investigation. In research when blood sample is to take repeatedly, like this, scientists are urged to reduce the number of experimental animals and follow the 3R principle when conducting research. The project was approved under animal experiment license number PE/EA/01444-6/2022.
2.2. Sampling and Determination of Melatonin
In the stable, the light intensity was always below 10 Lux at night and 100 Lux during the day which was determined with a light meter. The night sampling was done with use of red light at low illuminance near the animal. For daytime sampling, the animals remained in the barn, so that melatonin production remained at a detectable level.
About 8 milliliters of blood were taken from the jugular vein into vacutainers containing EDTA when sampling. Then, samples were kept below 5 °C. Within a day, samples were centrifuged at 2000g for 10 minutes. After harvesting plasma into Eppendorf tubes, it was immediately frozen and stored at -18 °C. For the determination of melatonin concentrations (pg/ml), a Tecan RE29301 RIA kit (RE29301, IBL Internation GmbH, Hamburg, Germany) containing a iodine-125 radioisotope labelled anti-melatonin antibody (indirect radioimmunoassay) was used according to the manufacturer's instructions. Samples (200 µl) were measured in duplicates and their average calculated. Measurements were made by using a gamma counter (LKB 1470).
Melatonin test results were pooled and then, a Kolmogorov-Smirnov one-sample test for normality was performed. This test confirmed the normal distribution of melatonin data (d = 0.1257, p > 0.20).
2.3. Statistical Processing
As background data set for processing of the melatonin results, the following variables were recorded and calculated:
- Identification number; The equine national life number, which was established by use of a microchip reader,
- Pedigree; The origin of the mares was known to the extent of two ancestral rows,
- Sampling date; The day, when the blood was collected,
- Sampling time; The hour and minute of blood collection,
- Days around winter solstice; The number of days between December 21, 2022, and the date of sampling (in a range between -160 and 128),
- Minutes around midnight; The period (in minutes) between midnight and exact taking of blood sample (in a range between -715 and 710),
- Pregnancy status; It is determined based on spring parturition in 2023: pregnant (foaled mares, n=8) or non-pregnant (mares did not foal, n=7),
- Foaling date; The day, when the parturition occurred,
- Days to foaling; It is the inverse of gestational age, the time between the date of foaling and the date of sampling, expressed in days. This approach can be considered favorable due to the variable length of pregnancy. The non-pregnant mares were given a simulated foaling of May 15, representing the average date of foaling for this stud.
The statistical processing of the obtained melatonin concentration was realized in two steps. First, they were corrected by eliminating the factors affecting them. With the help of an individual animal model [26], the effect of the serial number of sampling, the pregnancy status (as fixed effects), and the distance from the winter solstice, midnight, and foaling (as covariates) were filtered out. This is how the corrected values of the melatonin concentration were created, which are graphically illustrated according to circannual rhythm, circadian rhythm and throughout gestation. During the correction, the effect on which the course of melatonin concentration is presented was removed from the model.
In the second step, adjusted values were estimated for different specific times (transitions between seasons, parts of the days, and stages of lactation) on the corrected values by use of linear regression. So, adjustment was carried out for summer and winter solstices, and autumnal and vernal equinoxes. In regard to the day, adjusted values were computed for midday, afternoon 18:00, midnight, and morning 6:00. Regarding pregnancy, adjusted values were prepared for 300, 150, and 5 days before foaling. Adjusted values are also shown graphically, and additionally distance weighted least squares curve is fitted depending on the circannual rhythm, circadian rhythm and gestation. A statistical test (one-way ANOVA with Tukey honest significant difference test) was performed to compare them at midnight. As result, the mean and SEM (Standard Error of Mean) is displayed. For calculation of regression, statistical differences, and graphical presentation software Statistica (version 14., TIBCO Software Inc., Palo Alto, CA, USA) was applied [27].
3. Results
3.1. Circannual Rhythm
The corrected values for each sampling, the values adjusted for transitions between seasons, and the estimated annual distribution of the melatonin concentration is presented in Figure 1. In the mare study population, the highest melatonin concentration was found at winter solstice (about 32-33 pg/ml) and the lowest one at summer solstice (about 20 pg/ml). While during the autumn and spring equinoxes, the concentration of melatonin was roughly the same (27 and 25 pg/ml, respectively), time proportionally, between the values of the two solstices.
Melatonin concentration adjusted for the transitions between seasons and its statistical test are presented in Table 1. These values reflect the previously determined values, with the difference that they are higher, because these values are also adjusted for midnight. The significantly lowest value (27.67 pg/ml) is typical during the summer solstice, at midnight. During the autumn and spring equinoxes, similar concentrations (33.35 and 31.23 pg/ml, respectively), but significantly different from the values at the solstices, can be established. On the night of the winter solstice, the highest melatonin concentration (42.18 pg/ml) is formed in the blood plasma.
3.2. Circadian Rhythm
In Figure 2 the corrected values for each sampling, the values adjusted for times of the day, and the estimated circadian distribution of the melatonin concentration are presented. In the mares investigated, the highest melatonin concentration was observed at midnight (about 32 pg/ml) and the lowest ones at noon before and after midnight (about 17 and 20 pg/ml, respectively). At the same time, 6-6 hours before and after midnight the concentration of melatonin was similar (24 and 23 pg/ml, respectively).
Melatonin concentrations adjusted for the times of the day and winter solstice with statistical test are presented in Table 2. These values correspond to the previously presented values (see Figure 2). The values at noon (23.85 and 28.81 pg/ml, respectively) are significantly the lowest. The afternoon and morning values are in the medium range (33.17 and 32.63 pg/ml, respectively). In accordance with the previous (see Table 1), the strongest melatonin production (45.16 pg/ml) can be observed on the night of the winter solstice. Now, we emphasize that the values can be interpreted by themselves, per processing.
3.2. Gestational Age
The corrected values for each sampling, the values adjusted for specific days of gestation, and the estimated distribution of the melatonin concentration during the last 300 days before foaling is presented in Figure 3. The melatonin concentration of pregnant mares during this period of pregnancy can be considered constant in around 25 pg/ml. These values of the non-pregnant animals also stagnate at the same level during the control period.
There were no confirmed differences between the values of the melatonin concentration calculated at midnight during the study period, either according to gestational age or pregnancy status (Table 3). Their values are around 38 pg/ml and do not change as pregnancy progresses (p = 0.136). The apparent change can be regarded as a residual error despite correction for the season.
4. Discussion
We determined the change in blood plasma melatonin concentration during the whole year. Regarding the autumn period, Guillaume et al. (2006) also detected an increase in nocturnal melatonin concentration during two consecutive weeks [20]. According to Cozzi et al. (1991) the mule's melatonin production also shows cyclicity: in autumn, higher melatonin concentrations appear in the blood than in spring [14]. The same can be established in the present study, although the difference is not significant. However, the difference may have a physiological background since an intense decrease in melatonin concentration at the beginning of the year is a prerequisite for the start of the early reproductive period and estrus in donkeys.
The daily rhythm of blood plasma melatonin concentration was also confirmed in this study. However, it should be emphasized that the daytime brightness never exceeded 100 Lux during the examination. Otherwise, the production of melatonin would have decreased below the undetectable concentration (<0.5 pg/ml), as it was presented in the case of the horse [28]. A study done by Cozzi et al. (1991) in mules showed the circadian rhythm of melatonin production too. The rise of melatonin production started at 18 p.m. in fall and 22 p.m. in spring [14]. Interestingly the drop of melatonin production occurred around 7 a.m. to 8 a.m., at the same time, in both fall and spring. This study in mules showed that melatonin production follows a natural pattern of the daily light-dark cycle. In the horse species, several studies [17,29] also showed a significant 24-hour rhythm of the plasma melatonin level under natural conditions, i.e. a continuous light period of several hours is followed by a continuous dark period of several hours, and the sum of these constitutes the whole day. Taking samples every two hours from horses kept in darkness for 24 hours, Murphy et al. (2011) found no change in serum melatonin concentration over time [30]. Based on that data, they concluded that melatonin is not the only one that maintains circadian processes in the horse.
The last sample collection took place within two months before foaling. The correction was made for the more reliable time before parturition (5 days before parturition), not for the more uncertain parturition day. Knowledge of the melatonin concentration immediately before parturition and at the time of parturition requires further investigations. There is information about the nocturnal melatonin concentration in horse mares at parturition. Having the average foaling day at the beginning of March, this amounted to 36.35 pg/ml [31].
The circadian rhythm of melatonin secretion plays an important role in maintaining the fetus, having an anti-inflammatory effect. As shown in human, melatonin participates in the processes of implantation, the development of the placenta, and the maintenance of the neuroimmunoendocrine processes of the placenta, the purpose of which is the development of the vital functioning systems of the fetus. By regulating apoptosis, melatonin maintains the balance of cyto- and syncytiotrophoblast cells, thereby homeostasis of the placenta [32]. Melatonin produced in the placenta is secreted into the amniotic fluid, where it exerts antioxidant and anti-inflammatory effects [33]. In humans, blood melatonin levels increase during pregnancy, especially after 24 weeks, and reach their highest concentration before labor [34,35]. The circadian rhythm of gene expression was established directly in the uterus of pregnant and non-pregnant rodents, which highlighted the fact that melatonin triggers labor and parturition as a circadian signal. Labor begins in the late evening or early morning when melatonin secretion increases [36]. It has been shown to enhance oxytocin-induced uterine contractions by acting on protein kinase C via MT2 receptors [37]. Melatonin sensitizes myometrial myometrial cells to oxytocin, and its paracrine action facilitates simultaneous shortening of smooth muscle cells [38]. Melatonin exerts anxiolytic and analgesic effects via GHB (γ-hydroxybutyrate), MT1, MT2 and μ-opioid receptors [39]. When dim light is used at night, the peak concentration of melatonin is lower, and this prevents the formation of regular uterine contractions. It has also been observed in hospital conditions that the frequency of night births decreases with the use of artificial light at night [40].
5. Conclusions
The circannual and circadian determination of melatonin concentration was the first to be done in the donkey species and led to important results. It was found that the highest concentration of melatonin in the donkey mare under the test conditions (Northern hemisphere) is 45 pg/ml (at midnight on the winter solstice), which is roughly half of what was previously found [20]. The daily and annual rhythm is formed in the donkey species since values tend to be lower than this in the other seasons and times of the day. These findings show the great similarity of the donkey to the horse.
Since the donkey population was housed indoors during the experiment and the daylight remained below 100 Lux, melatonin values could also be measured during the day. However, as a result, the circannual and circadian runs appeared in a narrower range. In greater daylight, the values would probably have been scattered in a wider range from below.
It has been confirmed that the melatonin concentration does not change during the gestation period, unlike what human findings might have suggested. Determining the melatonin concentration around foaling still awaits as valuable research.
Author Contributions
András Gáspárdy (AG) and Sándor György Fekete (SF) conceived and supervised the investigation and had substantial inputs into the completion of manuscript. Levente Harmat (LH) and János Nagy (JN) participated in the collection of the biological samples and background information for processing and creation a first draft of the paper. Bence Somoskői (BS) and Alán Alpár (AA) performed the radioimmunoassay and contributed to the processing of literary sources. András Gáspárdy (AG) had substantial role in statistical data analysis. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the strategic research fund of the University of Veterinary Medicine Budapest (Grant No. SRF-001; Dr. Harmat Levente's Young Researcher Application “Determination of blood plasma melatonin concentration in donkeys depending on the circadian and circannual rhythm and the state of pregnancy 2023-2024, 1300000647”).
Institutional Review Board Statement
The project was approved under animal experiment license number PE/EA/01444-6/2022 in compliance with the Government Decree 40/2013 (II.14) on Animal experiments, Directive 2010/63/EU of the European Parliament and the Council on the Protection of animals used for scientific purposes, Act XXVIII of 1998 on the Protection and welfare of animals, and in compliance with Government Decree 66/2009 (IV.2) on the promulgation of the European Convention on the Protection of vertebrate animals used for experimental and other scientific purposes, signed in Strasbourg on March 18, 1986.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
The authors would like to thank they thank the Association of Hungarian Donkey Breeders (MSzE), especially the president Mihály Ernst, for making the background data of the donkey mares investigated available to us. The authors are grateful to Ágnes Keindl and Éva Dobozi for their laboratory assistance in handling and preparing the blood samples.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Schroeder, M.M.; Harrison, K.R.; Jaeckel, E.R.; Berger, H.N.; Zhao, X.; Flannery, M.P.; St Pierre, E.C.; Pateqi, N.; Jachimska, A.; Chervenak, A.P.; Wong, K.Y. The roles of rods, cones, and melanopsin in photoresponses of M4 intrinsically photosensitive retinal ganglion cells (ipRGCs) and optokinetic visual behavior. Front Cell Neurosci. 2018, 12:203. [CrossRef]
- Talpur, H.S.; Chandio, I.B.; Brohi, R.D.; Worku, T.; Rehman, Z.; Bhattarai, D.; Ullah, F.; JiaJia, L.; Yang, L. Research progress on the role of melatonin and its receptors in animal reproduction: A comprehensive review. Reprod Domest Anim. 2018, 53:831–849. [CrossRef]
- Gao, Y.; Zhao, S.; Zhang, Y.; Zhang, Q. Melatonin Receptors: A Key Mediator in Animal Reproduction. Vet Sci. 2022, 9:309. [CrossRef]
- Sharp, D.C.; Grubaugh, W.R. Use of push-pull perfusion techniques in studies of gonadotropin-releasing hormone secretion in mares. J Reprod Fertil Suppl. 1987, 35:289–296.
- Hart, P.J.; Squires, E.L.; Imel, K.J.; Nett, T.M. Seasonal variation in hypothalamic content of gonadotropin-releasing hormone (GnRH), pituitary receptors for GnRH, and pituitary content of luteinizing hormone and follicle-stimulating hormone in the mare. Biol Reprod. 1984, 30:1055–1062. [CrossRef]
- Grubaugh, W.; Sharp, D.C.; Berglund, L.A.; McDowell, K.J.; Kilmer, D.M.; Peck, L.S.; Seamans, K.W. Effects of pinealectomy in Pony mares. J Reprod Fertil Suppl. 1982, 32:293–295.
- Sharp, D.C.; Vernon, M.W.; Zavy, M.T. Alteration of seasonal reproductive patterns in mares following superior cervical ganglionectomy. J Reprod Fertil Suppl. 1979, 27:87–93.
- Kilmer, D.M.; Sharp, D.C.; Berglund, L.A.; Grubaugh, W.; McDowell, K.J.; Peck, L.S. Melatonin rhythms in Pony mares and foals. J Reprod Fertil Suppl. 1982, 32:303–307.
- Nagy, P.; Guillaume, D.; Daels, P. Seasonality in mares. Anim Reprod Sci. 2000, 60-61:245–262. [CrossRef]
- Murphy, B.A. Circadian and circannual regulation in the horse: internal timing in an elite athlete. J Equine Vet Sci. 2019, 76:14–24. [CrossRef]
- Fielding, D. Reproductive characteristics of the jenny donkey - Equus asinus: A review. Trop Anim Health Prod. 1988, 20:161–166. [CrossRef]
- Galisteo, J.; Perez-Marin, C.C. Factors affecting gestation length and estrus cycle characteristics in Spanish donkey breeds reared in southern Spain. Theriogenology. 2010, 74:443–450. [CrossRef]
- Carluccio, A.; Gloria, A.; Robbe, D.; Veronesi, M.C.; De Amicis, I.; Cairoli, F.; Contri, A. Reproductive characteristics of foal heat in female donkeys. Animal. 2017, 11:461–465. [CrossRef]
- Cozzi, B.; Morei, G.; Ravault, J.P.; Chesneau, D.; Reiter, R.J. Circadian and seasonal rhythms of melatonin production in mules (Equus asinus x Equus caballus). J Pineal Res. 1991, 10:130–135. [CrossRef]
- O’Brien, C.; Darcy-Dunne, M.R.; Murphy, B.A. The effects of extended photoperiod and warmth on hair growth in ponies and horses at different times of year. PLoS ONE. 2020, 15:e0227115. [CrossRef]
- Altinsaat, Ç.; Üner, A.G.; Sulu, N.; Ergün, A.: Seasonal variations in serum concentrations of melatonin, testosterone, and progesterone in Arabian horse. Ankara Üniv Vet Fak Derg. 2009, 56:19–24. [CrossRef]
- Guerin, M.V.; Deed, J.R.; Kennaway, D.J.; Matthews, C.D. Plasma melatonin in the horse: Measurements in natural photoperiod and in acutely extended darkness throughout the year. J Pineal Res. 1995, 19:7–15. [CrossRef]
- Haritou, S.J.A.; Zylstra, R.; Ralli, C.; Turner, S.; Tortonese, D.J. Seasonal changes in circadian peripheral plasma concentrations of melatonin, serotonin, dopamine and cortisol in aged horses with Cushing's disease under natural photoperiod. J Neuroendocrinol. 2008, 20:988–996. [CrossRef]
- Rapacz, A.; Lewczuk, B.; Prusik, M.; Raś, A. Diurnal rhythm of plasma melatonin level in mares from spring equinox to summer solstice. Bull Vet Inst Pulawy. 2010, 54:693–699.
- Guillaume, D.; Zarazaga, L.; Malpaux, B.; Chemineau, P. Variability of plasma melatonin level in pony mares (Equus caballus), comparison with the hybrid: mules and with jennies (Equus asinus). Reprod Nutr Dev. 2006, 46:633–639. [CrossRef]
- González-Arto, M.; Vicente-Carrillo, A.; Martínez-Pastor, F.; Fernández-Alegre, E.; Roca, J.; Miró, J.; Rigau, T.; Rodríguez-Gil, J.E., Pérez-Pé, R.; Muiño-Blanco, T.; Cebrián-Pérez, J.A.; Casao, A. Melatonin receptors MT1 and MT2 are expressed in spermatozoa from several seasonal and nonseasonal breeder species. Theriogenology. 2016, 86(8):1958–1968. [CrossRef]
- Messias, T.B.O.N.; Sant’Ana, A.M.S.; Araújo, E.O.M.; Rangel, A.H.N.; Vasconcelos, A.S.E.; Salles, H.O.; Morgano, M.A.; Silva, V.S.N.; Pacheco, M.T.B.; Queiroga, R.C.R.E. Milk from Nordestina donkey breed in Brazil: Nutritional potential and physicochemical characteristics in lactation. Int Dairy J. 2022, 127:105291. [CrossRef]
- Ernst, J. The donkey. In: Living Heritage, Old Historical Hungarian Livestock; Bodó, I. ed.; Agroinform Publishing and Printing Ltd., Budapest, Hungary, 2004; pp. 30–33.
- Lénárt, Z.; Ernst, M.; Gáspárdy, A. Preliminary results on body conformation of Hungarian Fallow Donkey. Danub Anim Genet Resour. 2017, 2:53–58.
- Harmat, L.; Kuncicky, A.; Lénárt, Z.; Ernst, M.; Nagy, J.; Gáspárdy, A. Conformation traits of Hungarian Fallow Donkey mares according to their basic colour. Danub Anim Genet Resour. 2022, 7:17–21.
- Pedigree viewer (2015) version 6.5f. http://bkinghor@une.edi.au.
- TIBCO Software Inc. (2020) Data Science Workbench. Statistica version 14. http://tibco.com.
- Murphy, B.A.; Elliott, J.A.; Sessions, D.R.; Vick, M.M.; Kennedy, E.L.; Fitzgerald, B.P. Rapid phase adjustment of melatonin and core body temperature rhythms following a 6-h advance of the light/dark cycle in the horse. J Circadian Rhythms. 2007, 5:5. [CrossRef]
- Piccione, G.; Giannetto, C.; Bertolucci, C.; Refinetti, R. Daily rhythmicity of circulating melatonin is not endogenously generated in the horse. Biol Rhythm Res. 2013, 44:143–149. [CrossRef]
- Murphy, B.A.; Martin, A.-M.; Furney, P.; Elliott, J.A. Absence of a serum melatonin rhythm under acutely extended darkness in the horse. J Circadian Rhythms. 2011, 9:3. [CrossRef]
- Gáspárdy, A.; Gallagher, G.; Bartha, B.; Cseh, S.; Fekete, S.G.; Somoskői, B. Plasma melatonin concentration during the early post-partum period in Thoroughbred mares and their foals. Acta Vet Hung. 2023, 71:119–127. [CrossRef]
- Reiter, R.J.; Rosales-Corral, S.A.; Manchester, L.C.; Tan, D.-X. Peripheral reproductive organ health and melatonin: ready for prime time. Int J Mol Sci. 2013, 14:7231–7272. [CrossRef]
- Tarocco, A.; Caroccia, N.; Morciano, G.; Wieckowski, M.R.; Ancora, G.; Garani, G.; Pinton, P. Melatonin as a master regulator of cell death and inflammation: molecular mechanisms and clinical implications for newborn care. Cell Death Dis. 2019, 10:317. [CrossRef]
- Kivelä, A. Serum melatonin during human pregnancy. Acta Endocrinol (Copenh). 1991, 124:233-237. [PubMed]
- Ejaz, H.; Figaro, J.K.; Woolner, A.M.F.; Thottakam, B.M.V.; Galley, H.F. Maternal Serum Melatonin Increases During Pregnancy and Falls Immediately After Delivery Implicating the Placenta as a Major Source of Melatonin. Front Endocrinol (Lausanne). 2021, 11:623038. [CrossRef]
- Cagnacci, A.; Soldani, R.; Melis, G.B.; Volpe, A. Diurnal rhythms of labor and delivery in women: Modulation by parity and seasons. Am J Obstet Gynecol. 1998, 178:140–145. [CrossRef]
- Mårtensson, L.G.; Andersson, R.G.; Berg, G. Melatonin together with noradrenaline augments contractions of human myometrium. Eur J Pharmacol. 1996, 316:273–275. [CrossRef]
- Karpovitch, A.E.; Inna, E.; Moiseevich, K.I. In Melatonin: pregnancy and childbirth. MOJ Curr Res & Rev. 2018, 1:206‒210. [CrossRef]
- Wilhelmsen, M.; Amirian, I.; Reiter, R.J.; Rosenberg, J.; Gönegur, I. Analgesic effects of melatonin: a review of current evidence from experimental and clinical studies. J Pineal Res. 2011, 51:270–277. [CrossRef]
- Olcese, J.; Lozier, S.; Paradise, C. Melatonin and the circadian timing of human parturition. Reprod Sci. 2013, 20:168‒174. [CrossRef]
Figure 1.
Yearly distribution of plasma melatonin concentration in the investigated donkey population.
Figure 1.
Yearly distribution of plasma melatonin concentration in the investigated donkey population.
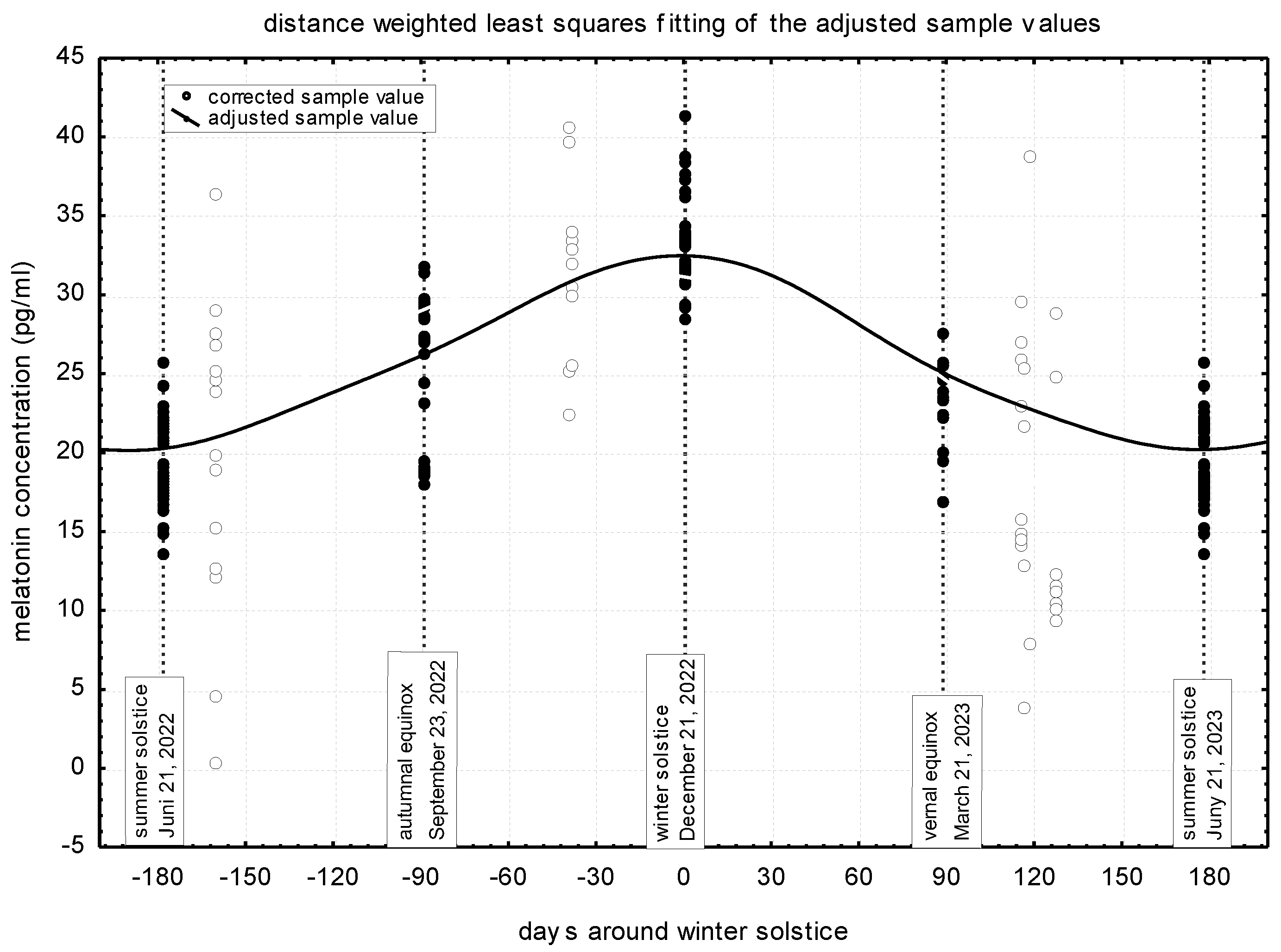
Figure 2.
Daily distribution of plasma melatonin concentration in the investigated donkey population.
Figure 2.
Daily distribution of plasma melatonin concentration in the investigated donkey population.
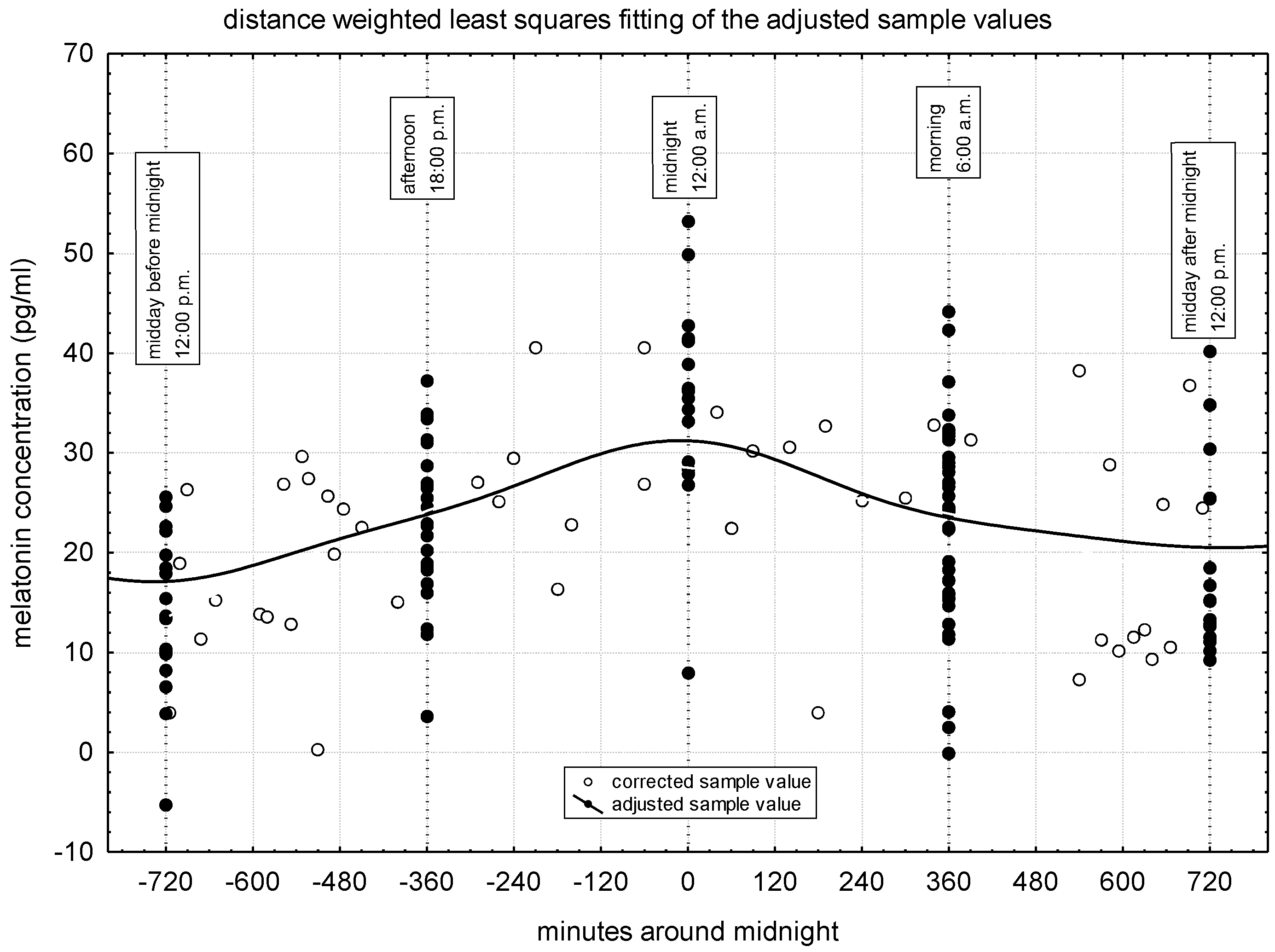
Figure 3.
Distribution of plasma melatonin concentration during gestation in pregnant mares and in non-pregnant controls.
Figure 3.
Distribution of plasma melatonin concentration during gestation in pregnant mares and in non-pregnant controls.
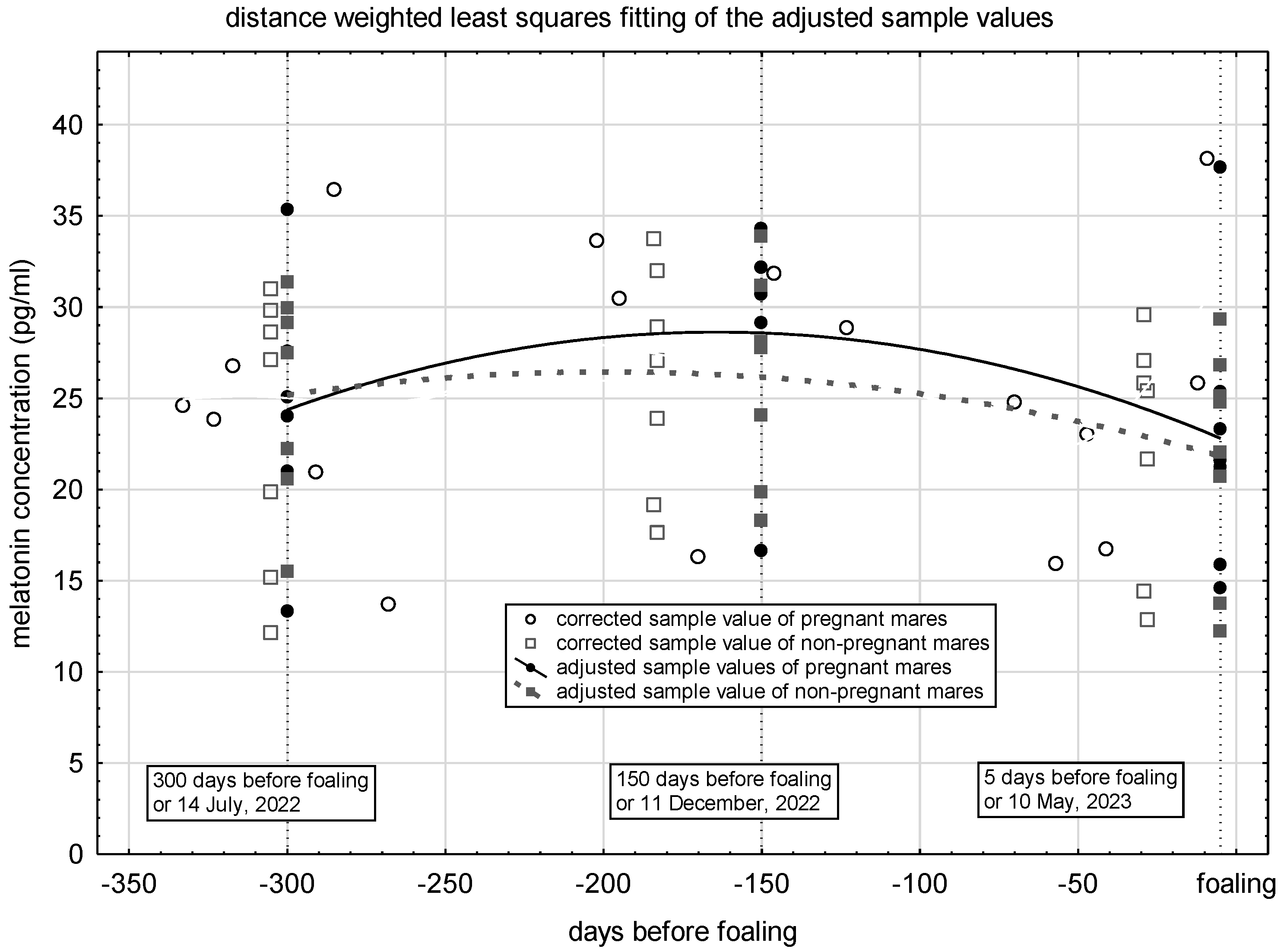

Table 1.
Melatonin concentration adjusted for the transitions between seasons and midnight.
|
Event p < 0.001 |
Melatonin concentration mean, pg/ml |
SEM* |
| Summer solstice, Juni 22, 2022, 12 a.m. | 27.67a | 0.459 |
| Autumnal equinox, September 24, 2022, 12 a.m. | 33.35b | 1.071 |
| Winter solstice, December 22, 2022, 12 a.m. | 42.18c | 0.523 |
| Vernal equinox, March 21, 2023, 12 a.m. | 31.23b | 0.683 |
* Standard error of mean. a-c different superscript letters differ significantly (Tukey honest significant difference test, P<0.05).
Table 2.
Daily melatonin concentration adjusted for the times of the day and winter solstice.
|
Dayparts p < 0.001 |
Melatonin concentration mean, pg/ml |
SEM* |
| Midday before midnight, 12:00 p.m., December 21, 2022 | 23.85a | 1.981 |
| Afternoon, 18:00 p.m., December 21, 2022 | 33.17b | 1.607 |
| Midnight, 12:00 a.m., December 22, 2022 | 45.16c | 2.460 |
| Morning, 06:00 a.m., December 22, 2022 | 32.63b | 1.652 |
| Midday after midnight, 12:00 p.m., December 22, 2022 | 28.81ab | 2.371 |
* Standard error of mean. a-c different superscript letters differ significantly (Tukey honest significant difference test, P<0.05).
Table 3.
The melatonin concentration adjusted for midnight according to specific days of gestation.
|
Days to foaling p = 0.136 |
Melatonin concentration mean, pg/ml |
SEM* |
| 300 days before foaling, p = 0.830 | 38.80 | 1.74 |
| Pregnant | 38.37 | 2.971 |
| Non-pregnant | 39.17 | 2.218 |
| 150 days before foaling, p = 0.523 | 41.17 | 1.757 |
| Pregnant | 42.58 | 3.107 |
| Non-pregnant | 40.16 | 2.169 |
| 5 days before foaling, p = 0.771 | 36.09 | 1.754 |
| Pregnant | 36.67 | 2.922 |
| Non-pregnant | 35.58 | 2.248 |
* Standard error of mean.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



