
Submitted:
05 June 2024
Posted:
07 June 2024
You are already at the latest version
Abstract
The role that aquatic aerosols might play in inter-ecosystem exchange in freshwater riparian environments has largely been understudied. In these environments, where freshwater streams are used both as drinking water and for treated waste disposal, water features like waterfalls, downed trees, and increased streamflow can serve as bioaerosol producers. Such water features could have an important role in the bacterial colonization of surrounding surfaces, including the riparian phyllosphere. In this study, we explore the influence of a freshwater stream’s bacterial community composition and micropollution on riparian maple leaves exposed to bioaerosols produced from the stream. Using culture-based and non-culture-based techniques, we compared phylloplane microbial communities in riparian zones, adjacent non-riparian forested zones, and the surface waters of the stream. In this system, riparian zone maple leaf surfaces had higher bacterial counts than non-riparian zone trees. Using metagenomic profiling of the 16S rRNA gene, we found that, while microbial communities on leaves in both riparian zone and forested sites were diverse, riparian zone bacterial communities were significantly more diverse. In addition, we found that riparian leaf bacterial communities shared more amplicon sequence variants (ASVs) with stream bacterial communities than forest leaves, indicating that the riparian zone phyllosphere is likely influenced by bioaerosols produced from water surfaces.
Keywords:
phyllosphere
; phylloplane
; aerosol
; riparian
; freshwater
; HymenobacterSphingomonas
; Oxalobacter
; Acer rubrum
1. Introduction
The phyllosphere, or the above-ground external surfaces of plants, is thought to be the largest microbial habitat on earth [1]. Phyllosphere microbial communities (PMCs) heavily influence plant fitness and ecosystem function, and themselves serve as a major source of bioaerosols globally [2]. Understanding the assembly and structure of the PMC is therefore vital for effective management of food safety and security, understanding local and ecosystem-level function, and predicting impacts of climate change both locally and globally[3]. Despite these realities, key questions remain unanswered regarding environmental impacts on the sources for and function of PMCs [4].
Bioaerosols are a dominant source for phyllosphere colonization [5], representing all major earth biomes, including marine, soil, plant, and animal sources [6]. Dispersal and delivery of bioaerosols to leaf surfaces are controlled by geographical proximity to bioaerosol sources [7,8], meteorological conditions like wind speed, wind direction, and precipitation [1], and leaf topography [9]. PMCs are a diverse mixture of fungi, bacteria, archaea, algae, and viral particles, with bacterial communities often dominating [10,11].
The microbial content of ambient aerosols varies based on seasonal and geospatial influences [12,13]. Once aerosolized microbes deposit on the leaves of plants and trees, they become either short-term or long-term epiphytic residents [13,14], according to host and established PMC responses to these new microbes [15,16]. Recent studies have demonstrated that, despite geospatial variability in microbial content of aerosols, trees appear to have species-specific core microbial communities [10]. The influence and source for microbes not included in that core microbiome, and their interactions with that core microbiome, are not well understood.
Land-based aerosol sources are better studied, but water surfaces are also a major source of aerosols [17], through the action of bubbles bursting at marine and freshwater surfaces and launching microscopic droplets into the air [18,19,20,21,22,23,24,25]. Bubbles are introduced in both marine and freshwater environments through wind-wave interactions [26,27,28] wave-shore interactions [29]; human recreation (e.g., boating, swimming); and water features creating whitewater, including art installations, fountains, downed trees, waterfalls, and gas evolution, among others. Bioaerosols created from water surfaces can be important mechanisms for transfer/transport of viable microbes and nutrients to land-based surfaces and ecosystems [17,26,27,28]. It follows, then, that water systems would be an important influence on the microbial colonization of adjacent phyllospheres, particularly in riparian zones.
While there has been some study of the contribution of the phyllosphere to soil microbes, and discussion of the eventual impact on water microbes as water travels from land to waterways, the contribution of aquatic microbiomes to land-based microbiomes, including the phyllosphere, is still poorly understood. Studies of the movement of soil microbes from riparian zones into waterways have been conducted, e.g. [30], but do not include the phyllosphere. Dodds et al. [31] studied the potential transfer of PMC’s to waterways through run-off, but did not consider the role that waterways themselves have in influencing PMC’s to begin with.
The connection between water and air quality through microbial aerosols has been demonstrated mostly in marine environments, including coastal oceans [27,32,33,34], estuarine coastlines [28,35], and polluted shipping canals [36,37]. As these studies continue, it’s clear that what is in the water (including chemical and biological pollution) is also transferred to the air, where it travels inland to deposit on terrestrial surfaces. Studies of this phenomenon and its implications for freshwater systems, however, have not been conducted, particularly as it relates to the environmental distribution of water-bourne sewage-associated microbes and micropollution (genetic material, antibiotic resistance gene cassettes). The connection between riparian PMCs and water quality, until our study, is largely unknown.
The influence of air pollution on PMC structure and function has been studied demonstrating a relationship between proximity to traffic and other forms of air pollution and the degradation of PMC function [38,39,40]. Anthropogenic impacts on PMCs in urban arenas have included a reduction in alphaproteobacteria in urban environments [41], but a higher diversity of microbes than non-urban PMC’s overall. Leaves and trees are known to be effective filters of air pollution, and are sometimes even used to clean air [42]. Interestingly, there are many studies outlining how riparian zones and the phyllosphere in particular protect waterways from pesticide applications [43], but no studies to date examining the role the riparian zone may play in protecting inland ecosystems from the pollution present in the water itself.
Here, we explore the potential for riparian phyllospheres to intercept microbial aerosols contributed by rural/suburban freshwater systems known to contain treated and untreated sewage inputs. We compared Acer rubrum (red maple) PMC’s in a freshwater stream riparian zone adjacent to aerosol-creating water features (a historical dam spillway and a waterfall) to A. rubrum PMC’s in the same forest system, but located well above the freshwater stream. Due to proximity to the waterway and the aerosols created by the distinct water features, we expected to find a clear difference in PMC diversity and structure between riparian and further inland forest A. rubrum PMCs. Furthermore, we hypothesized that riparian zone phyllospheres would include detectable influence from the adjacent water surface, including shared aquatic microbial species, sewage indicators, and micropollution.
2. Materials and Methods
2.1. Study Sites
As previously described in De Santana et al. [44], this study was conducted on the Saw Kill, a 23.0 km tributary of the Hudson River, located in the upper Hudson Valley, during the summer of 2015. The Saw Kill Watershed drains 57 km2, including forest, wetland, agricultural, and suburban developed land (Wikipedia Saw Kill 2022). There are 2 permitted sewage outfalls (NYSPDES permit #’s NY0271420, NY0031925) on the Saw Kill, and one permitted drinking water facility (130,000 gallons per day) between the two sewage outfalls.
Four phyllosphere sampling sites (two riparian sites (adjacent to de Santana et al. stream sampling sites), and two forest sites > 65 m upslope of the water sampling sites) were chosen to represent “riparian” and inland “forest” sites (Figure 1). Three A. rubrum at each site were marked prior to the sampling campaign to ensure consistent collection from the same trees throughout (Figure 1). The riparian trees were exposed to constant production of aquatic aerosol through a dam spillway (upper riparian) and a waterfall (lower riparian). Previous preliminary aerosol monitoring conducted during a clear, windless day demonstrated a significant difference in aerosol particle number and size distribution between forest and riparian tree sites (Supplemental Figure S1), with the riparian zone containing significantly higher aerosol particles in the range known to harbor bacterial aerosols [45]. All sampling was conducted on ten sampling days between 6/22/15 and 7/22/15.
2.2. Surface water and phyllosphere sampling
Surface water samples were collected as per de Santana et al. [44] in sterile and acid-washed 2 L Nalgene bottles from mid-channel within 0.5 m of the stream surface at each site. All samples were placed on ice and transported to the laboratory for analysis within 2 hours of sampling. A list of all water samples along with their physical characterization (i.e., GPS coordinates, date, temperature, salinity, conductance, particulate) is available in de Santana et al. [44].
We used methods similar to Laforest-Lapointe et al. [24] and Tang et al. [46] to sample A. rubrum leaf PMC. Tree leaves collected during the sampling campaign (mid-summer) were fully mature, but not senescing, minimizing leaf age influence on PMC’s. For each sampling event, three leaves were randomly collected from each tree at 250 – 300 cm above ground-level. The leaves were gathered with sterilized forceps at the base of the stem and broken from the tree. Two leaves from each tree were immediately placed in a sterile 50 ml falcon tube to be later washed for use in culturable bacteria counts and bacterial DNA extraction, and one leaf was placed in a sterile, clear Nasco Whirl-Pak® bag for processing in the lab.
The leaf “wash,” representing both adaxial and abaxial leaf PMC, was obtained as follows: 15 ml of endotoxin-free Hyclone Hypure™ Cell Culture Grade water was pipetted into each of the falcon tubes containing two leaves from each tree. The tubes were then vortexed at maximum speed for 5 minutes, and then centrifuged at 1,300 rpm for 3 minutes. Supernatant from each was then transferred into a new sterile tube, with wash from all three trees per site pooled for downstream assays.
2.3. Bacterial analyses
2.3.1. Culture-based methods
Leaf surface area (LSA) was estimated by outlining sterile bagged leaves on grid paper using a light table to minimize disturbance of attached microbial communities. These leaves (one per tree) were then “printed” onto LB agar media with 0.004 g Amphotericin B (LB/AmB), in order to deter fungal growth [47]. Specifically, the adaxial side of each leaf was pressed gently onto the agar surface using a sterilized stirring rod, and then removed and discarded. Bacterial colonies were counted after 96 hours of growth in the dark and at room temperature (22-25 ºC). Concentrations of culturable bacteria on leaf surfaces and in water samples were also determined by spreading 100 μl of pooled leaf “wash” onto agar plates with the same media described above. These plates were incubated along with the leaf-print plates.
2.3.2. Culture-independent methods
For each water sample, we filtered 750 ml of surface water through a 0.22 μm Sterivex filter. We then filtered the remaining pooled leaf wash (~14.9 ml) from each site through a 0.22 μm Sterivex filter. A filter of only sterile, endotoxin-free water was also made to serve as a control for extraction methodology and sequencing methodology. We then extracted total DNA from these filters using the PowerWater DNA Isolation kit (MoBio Laboratories Carlsbad, CA, USA), now available as the DNeasy PowerWater DNA Isolation Kit (QIAGEN, Hilden, Germany). DNA concentrations in the extractions were measured using a Thermo Fisher Nanodrop 2000 (Thermo Scientific, Willmington, DE). All extractions of water and leaf filters contained more DNA than the control filter.
We quantified the presence of the 16s rRNA genes in water and leaf extractions using quantitative PCR (i.e., qPCR) following the procedure described in de Santana et al. [44]. In summary, we processed each sample in triplicate using the primers described in Gaze et al. [48] and the PowerUp SYBR Green Master Mix (Applied Biosystems, Foster City, CA, USA). Samples were then ran using the Bio-Rad CFX96 Real-Time PCR Detection System (Bio-Rad Laboratories, Hercules, CA, USA) with an internal standard curve constructed from a serial dilution of the Escherichia coli strain SK4903. Data files are available in de Santana et al. [44].
To characterize the overall bacteria community structure in each sample, we amplified the V4 region of our 16S rRNA gene amplicon sequencing library using primers 515F and 806R, as outlined in the Earth Microbiome Project [49]. Samples were sequenced on the Illumina Miseq platform using 250-bp paired ends by Wright Labs (Huntingdon, PA, USA) and are described in de Santana et al. [44]. Taxonomy was established using different curated databases and taxonomic assignment algorithms (Figure S1). Raw reads are/will be publicly available on the Sequence Reads Archives of the NCBI (accession number: TBD).
2.4. Bioinformatic and Statistical analyses
As described in de Santana et al. [44], raw Illumina sequences were filtered and trimmed with Trimmomatic, ver. 0.39,21 using the following parameters: ILLUMINACLIP:TruSeq3-PE.fa:2:30:10 LEADING:3 TRAILING:3 SLIDINGWINDOW:4:15 M INLEN:100. Subsequent steps were performed using QIIME2, ver 2021.222. DADA2 [50] was used to denoise paired reads (using the following parameters: --p-trunc-len-f 250, --p-trunc-len-r 233) with the median non-chimeric read count per sample estimated as 21,741 (Supplemental Table: Qiime2_Denoising_stats.tsv [x]). Alpha and beta-diversity analyses were performed on rarefied and non-rarefied amplicon sequence variant (ASV) lists using phyloseq [51]. Because they did not show differences in trends, we are providing the non-rarefied results in this paper, according to McMurdie et al. [52]. Finally, culture-based bacterial measurements and qPCR results were pooled by sample site type (riparian vs. forest) and differences were determined using a t-test. All statistical analyses were performed using R version 4.3.3 [53].
3. Results
3.1. Culture-based and culture-independent quantification of leaf PMCs
We first investigated whether the microbial load on leaves sampled in riparian sites differed from leaves sampled from forest sites. We compared PMC density by estimating the total CFU counts on the leaf surface through the leaf prints. While adaxial leaf surface area did not differ significantly between riparian and forest sites, we found that adaxial CFU density varied significantly between the two ecosystems (t = -4.3; DF = 21.11; P < 0.001; Figure 2A). The riparian phyllosphere had, on average, more than 3 times the abundance of bacterial CFU’s per cm2 as the forest phyllospheres. Interestingly, we found that CFU’s grown from leaf washes (which would include abaxial communities) did not differ significantly between forest and riparian leaves (t = 0.29; DF = 30.8; P = 0.77; Figure 2B). These results suggest there might be an additional CFU source for the adaxial side of leaves in the riparian zone.
We then investigated bacterial load in leaf wash using qPCR, a culture-independent technique, to account for the large proportion of bacteria that cannot grow on agar or in normal laboratory conditions. We found that the abundance of 16S rRNA gene was significantly higher in the riparian phyllosphere than in forest phyllospheres (t = -2.17; DF = 22.1; P = 0.04; Figure. 2C). Interestingly, we found the presence of IntI1 on both riparian and forest leaves (Figure 2D), but the difference was not significant (t = 0.27, DF = 25.76, p = 0.79). Crucially, combining culture-based and culture-independent metrics, our results strongly suggest that bacterial communities are more abundant in riparian ecosystems and even more so on the adaxial surface of leaves. These results again suggest that there exists a separate bacterial source for riparian leaf PMCs, and confirms the utility of employing both traditional culture-based methods and culture-independent techniques in studying leaf PMCs.
3.2. Comparing bacterial communities in riparian and forest ecosystems
3.2.1. Riparian leaf PMCs show higher level of diversity
In addition to comparing total bacterial load between the riparian and forest ecosystems, we then compared the bacterial communities in terms of diversity and composition. From 39 leaf wash samples, including abaxial and adaxial surfaces, we found a total of 3975 amplicon sequence variants (ASVs), or bacterial types (Supplemental Table S1). Overall, we identified 25 different phyla, 147 orders, and 582 genera among the bacterial ASVs (Table S2). While a full list of taxonomic groups is available in Supplemental Table S2, the most common bacterial phyla were Proteobacteria (74.2 ± 2.9%) followed by Bacteroidota (14.4 ± 0.85%), Actinobacteriota (4.78 ± 0.81%), and Firmicutes (3.44 ± 1.2%). Beijerinckiaceae (14.4 ± 1.0%), was the most frequent bacterial family followed by Hymenobacteraceae (10.1 ± 1.1%), Comamonadaceae (9.78 ± 3.4%), Oxalobacteraceae (8.88 ± 2.0%), Enterobacteriaceae (7.51 ± 7.1%), and Sphingomonadaceae (6.08 ± 1.1%) (Supplemental Figure S2).
Using unique ASVs, or bacterial types, as the taxonomic unit, we then investigated whether bacterial communities associated with leaves sampled in riparian locations are different from bacterial communities sampled on leaves in forest ecosystems. On average, we found that riparian bacterial communities harbored a higher number of ASVs, i.e., 136.33 ± 54.86, than bacterial communities found on forest leaf surface, i.e., 105.22 ± 35.32 (F(1,34) = 5.39; P = 0.03). We found a similar difference when considering the Chao1 index, which predicts the number of bacterial types using sampling depth; the number of predicted ASV was once again higher in riparian microbial communities (F(1,34) = 4.69; P = 0.04; Figure 3A).
To account for possible differences in evenness between bacterial types, we also compared diversity using Shannon’s Diversity index. We again found a higher diversity in riparian leaf PMC (F(1,34) = 8.56; P = 0.006; Figure 3B). The latter indicates that the riparian ecosystems host a larger number of bacterial types, and that these bacterial types are more equally distributed.
3.2.2. Bacterial communities in riparian ecosystems are distinct from forest ecosystems.
We then investigated whether leaf PMC population structure differed based on ASV taxonomic identity and relative abundance. First, we conducted a non-metric multidimensional scaling analysis using the Bray-Curtis dissimilarity matrix, which roughly identifies the proportion of similarities between each community pair. As expected, we found that the PMCs differed significantly between the riparian and forest leaves (F(1,36) = 2.38; R2 = 0.06; P = 0.004). We also compared the microbial communities according to their phylogenetic overlap. This measure, which can be weighted to account for the abundance of each ASV in addition to their phylogenetic distance, is predicted to be a better reflection of the functional diversity present in the population; close phylogenetic matches are predicted to have more similar metabolism and ecological functions. Conducting a non-metric multidimensional scaling analysis on the weighted UniFrac scores, first assessing finer changes in phylogenetic distance and abundance, we found a statistical difference between the forest and riparian leaf PMCs (F(1,35) = 3.10; R2 = 0.08; P = 0.01). Then, we repeated the analysis with unweighted UniFrac, and found again that riparian and forest PMCs differed significantly (F (1,35) = 1.43; R2 = 0.04; P = 0.04), confirming that leaf PMCs in the riparian and forest areas were fundamentally different at the composition level and likely at the functional level as well.
Given the significant difference in overall population structure between the forest and riparian leaf PMCs, we were then interested in investigating which ASVs differed between riparian and forest communities. Overall, we found 186 ASVs that changed in relative abundance between the forest and riparian areas (DESeq2; adj-P ≤ 0.05), including 161 ASVs that were higher in abundance in the riparian area. While the most important observed differences were the over-abundance of Hymenobacter sp. and Sphingomonas sp. in the riparian area, we also found an increased abundance of wastewater indicators and potential human pathogens in the riparian area, including Clostridium sp., Enterococcus sp., Klebsiella sp., Mycoplasma sp., and Yersinia sp (Supplemental Figure S3)..
Interestingly, using methods as per Dueker et al. [35] to identify bacterial genera known to be primarily associated with sewage, we found that 80% of riparian bacterial communities (n = 20), and 78% of forest bacterial communities (n = 18) samples harbored sewage-related bacterial genera (Figure 4). While the mean relative abundance of these sewage associated bacteria was 1.7% in riparian PMC samples and 0.4% in the forest PMC samples, this difference was only marginally significant (t = -1.76; DF = 20.63; P = 0.09).
3.5. Water-phyllosphere connections
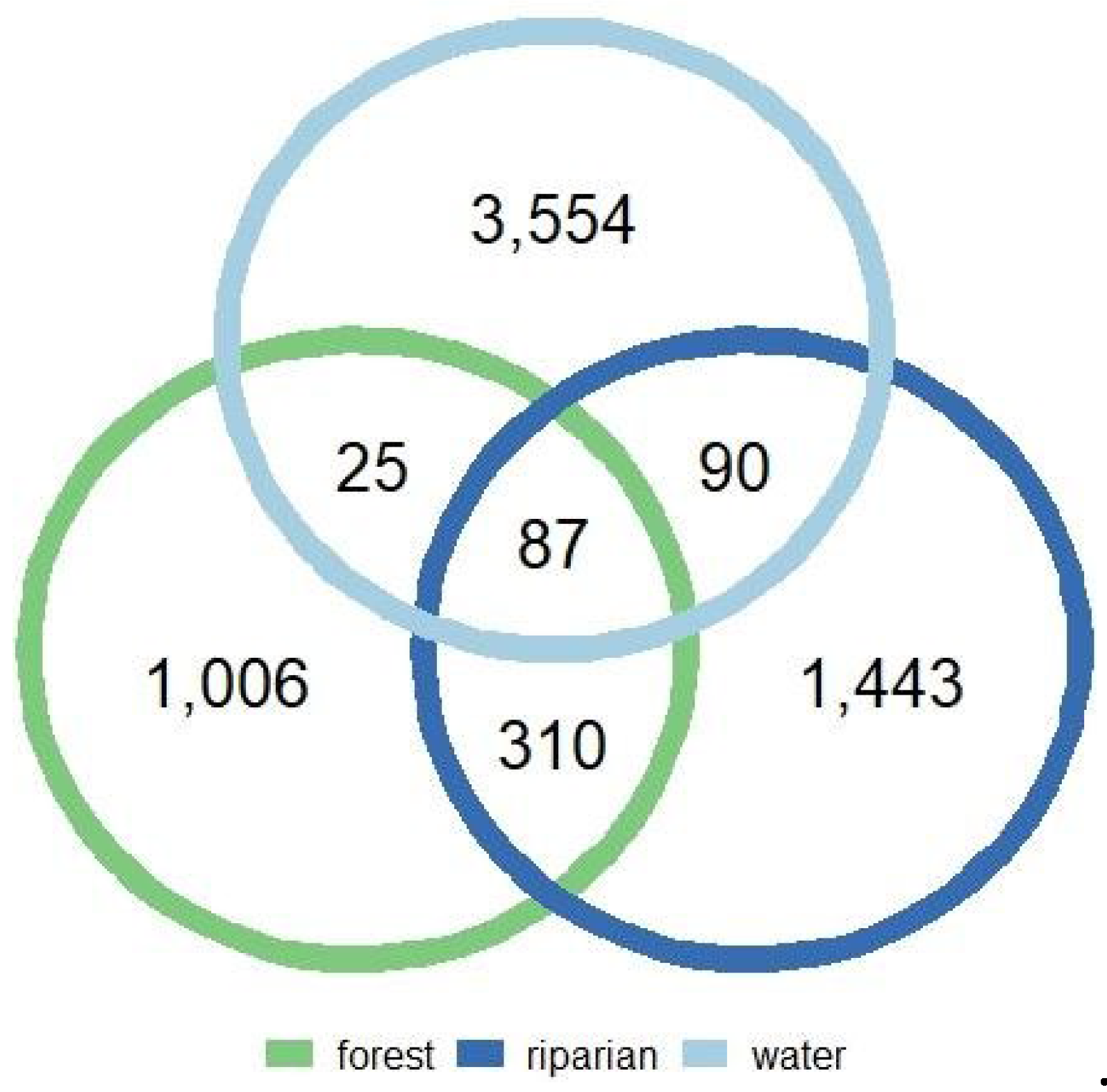
Finally, we assessed a possible connection between the phyllosphere and the water ecosystem located near the riparian trees sampled. To do so, we compared the bacterial community composition identified in the riparian and forest ecosystem to the bacterial community composition isolated from the Saw Kill stream (Figure 5). We found that only 87 ASVs were shared across all ecosystems which accounted for 6.0%, 8.6%, and 2.4% of all ASVs in riparian, forest, and stream microbiomes respectively (Supplemental Table S3). We also found that riparian and forest bacterial communities shared 310 ASVs (Supplemental Table S4) (Figure 4), which accounts for 21.5% and 30.8% of all ASVs found in riparian and forest communities respectively. The latter also represents the highest proportion of shared ASVs between two ecosystems. Interestingly, we found that riparian microbiomes shared 90 ASVs (Supplemental Table S5) with the stream microbiome while forest communities shared only 25 ASVs (Supplemental Table S6) with the stream microbiome. Using the non-parametric Fisher’s Exact test, we found that riparian bacterial communities share a significantly higher proportion of ASVs with the water surface bacteria communities than do forest bacterial communities (Fisher’s Exact test: adj-P < 0.01).
4. Discussion
Here, we investigated the possible link between bacterial communities found on the phyllospheres of trees to those found in surface waters of a freshwater stream system. Overall, we found that the leaves of A. rubrum near the stream (and a water feature creating aquatic aerosols) sustained higher numbers of culturable and total bacteria than A. rubrum in the surrounding forest. In addition, we found that the riparian leaf bacterial communities were compositionally different and more diverse than the communities isolated from forest leaves. Crucially, we also found that the riparian PMCs shared a significantly higher proportion of bacterial types with surface water bacteria communities than did the forest sites. These results, while limited to A. rubrum, suggest that there exists a strong link between riparian leaf PMCs and adjacent surface water bacterial content in freshwater ecosystems.
Studies of temperate forest leaf PMCs, and A. rubrum in particular, are sparse. To our knowledge, there has not previously been a study of riparian leaf PMC’s for A. rubrum. However, the PMC’s in our study had prominent representation of bacterial families previously noted as core PMC constituents in two temperate forests (including A. rubrum) in Québec, Canada [54]. These included Beijerinckiaceae, Enterobacteriacea, Oxalobacteraceae, and Hymenobactereacea [54,55]. Perhaps unsurprisingly, in both Québec PMC’s and our study’s PMCs, these bacterial families were dominated by bacterial genera commonly detected in aerosols, including Hymenobacter [56,57,58,59], Sphingomonas [59,60], and Oxalobacter [59].
A separate study observing leaf PMC’s across an urban gradient (including A. rubrum) used several sampling sites on the shore of the Lachine Canal in Montréal, Canada [41]. In this study, A rubrum PMC diversity increased with urban intensity, and leaf bacterial communities of A. rubrum were more impacted by urban intensity than other trees sampled (including hackberry (Celtis occidentalis), white ash (Fraxinus americana), sugar maple (Acer saccharum), and white spruce (Picea glauca) [41]. While the trees we sampled are located in a managed forest/stream area, the riparian zone trees were along a stream impacted by urban/suburban upstream activities [44].
Given the difference in overall population structure we found between riparian and forest A. rubrum, we then investigated what ASVs differ between riparian and forest communities. Our results suggest that the bacterial communities in each zone differ both at a broad phylogenetic level and at a finer scale, indicating an important difference in terms of community composition and likely at the functional level as well.
When looking at specific changes in ASVs, we found that riparian communities had a higher number of bacteria taxa and unique bacteria taxa not found in the forest ecosystem. We also found that the riparian communities were more evenly distributed, as measured as a higher Shannon Diversity Index. Interestingly, Laforest-Lapointe et al. [54] found that Québec forest A. rubrum communities had a mean Shannon Diversity Index of 3.7 +- 0.06, but in their subsequent study of urban exposure effects, near-shore A. rubrum leaf PMC Shannon Diversity Indices increased with urban exposure [41], ranging from 3.75 at low urban intensity to ~4.3 at medium urban intensity. Here, we found that riparian zone A. rubrum supported a significantly higher mean Shannon Diversity index, i.e.4.1, than the forest zone, i.e. 3.4, which is similar to that described by Laforest-Lapointe. Further study is needed to understand the drivers of this difference, but proximity to water aerosol sources and what they represent (e.g., urban influence through upstream sewage and runoff) may have significant impacts on A. rubrum PMCs.
We also found that A. rubrum PMCs in our study include water pollution-associated bacteria. While aerosolization of water pollution into coastal air is a known phenomenon [32,35,36,37], it is likely that a similar phenomenon is occurring in this freshwater stream system. Interestingly, we did find an increased abundance (albeit with marginal significance) of bacteria associated with wastewater and infectious disease in riparian leaf PMCs. The riparian leaf PMCs also had higher numbers of enterococcus (sewage indicating bacteria also detected in the stream using culture-based techniques [44]) than the forest leaf PMCs. While we did not determine the sources for PMC bacteria in this study, this finding indicates a detectable link between what is on riparian leaves and what is in adjacent waterways, which could occur either through bioaerosols originating from Sthe waterway impacting on the leaves, or the leaves providing these bacteria to the stream through runoff or re-aerosolization through wind and rain interactions. While more work is needed to explicitly determine the mechanisms of connection, our study does establish the possibility for many pathways to exist.
Finally, the fact that sewage bacteria and the micropollutant and antibiotic resistance gene IntI1 were present in a majority of water [44] and phyllosphere samples demonstrates that the A. rubrum phyllosphere could act as an extra-enteric reservoir for sewage-associated bacteria and antibiotic resistance genes, raising possible health concerns. On the other hand, our findings suggest that riparian zone vegetation could perhaps mitigate the long-distance transport of aerosolized water bacteria, which is a previously unappreciated role for riparian zones. Essentially, riparian zones may not only protect waterways from external pollutants (pesticides, herbicides, etc. [43]), but they may also be filtering and/or capturing aerosolized pollution from contaminated waterways.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Aerosol particle size distributions; Figure S2: Relative abundance of bacterial families in riparian and forest PMC’s: Figure S3: Aerosol particle size distributions Aerosol particle size distributions, and a link to detailed ASV taxonomy and grouping tables (S1-S6).
Author Contributions
Conceptualization, MED,BL, GPP; methodology, MED, BL, GPP; resources, MED, GPP; data curation, MED, BL, GPP; writing—original draft preparation, MED, BL, GPP; writing—review and editing, MED, GPP; funding acquisition, MED, BL. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Hudson River Foundation Polgar Fund.
Institutional Review Board Statement
Not applicable
Data Availability Statement
All data are accessible through the article, with further detail provided in Supplemental Materials. In addition, all stream water data are available through a Scientific Data paper by de Santana et al. [44]: https://www.nature.com/articles/s41597-022-01686-8#Sec12
Acknowledgments
The authors would like to thank the Bard Summer Research Institute for supporting this project as well as the Bard Biology Program, the Center for Environmental Sciences and Humanities. In addition, the authors thank Daniella Azulai and Haley Goss-Holmes for assistance in the field and Dylan Dahan for help with the qPCR analyses.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Morris CE, Kinkel LL, Lindow SE, Hecht-Poinar EI, Elliott VJ, editors. Fifty years of phyllosphere microbiology: significant contributions to research in related fields2002.
- Lighthart B, Shaffer BT. Increased airborne bacterial survival as a function of particle content and size. Aerosol Sci Technol. 1997;27(3):439-46. PubMed PMID: ISI:A1997XR30000011.
- Leveau JHJ, Beattie GA, Lindow SE, Mahaffee WF. Phyllosphere, Front and Center: Focus on a Formerly ‘Ecologically Neglected’ Microbial Milieu. Phytobiomes Journal. 2023;7(2):140-4. [CrossRef]
- Leveau J. A brief from the leaf: latest research to inform our understanding of the phyllosphere microbiome. Curr Opin Microbiol. 2019;49:41-9. [CrossRef]
- Vacher C, Hampe A, Porté AJ, Sauer U, Compant S, Morris CE. The Phyllosphere: Microbial Jungle at the Plant–Climate Interface. Annual Review of Ecology, Evolution and Systematics. 2016;47(Volume 47, 2016):1-24. [CrossRef]
- Bulgarelli D, Schlaeppi K Fau - Spaepen S, Spaepen S Fau - Ver Loren van Themaat E, Ver Loren van Themaat E Fau - Schulze-Lefert P, Schulze-Lefert P. Structure and functions of the bacterial microbiota of plants. (1545-2123 (Electronic)).
- Hanson CA, Fuhrman JA, Horner-Devine MC, Martiny JB. Beyond biogeographic patterns: processes shaping the microbial landscape. Nature reviews Microbiology. 2012;10(7):497-506. Epub 05/15. [CrossRef]
- PubMed PMID: 22580365. [CrossRef]
- Finkel OM, Burch Ay Fau - Elad T, Elad T Fau - Huse SM, Huse Sm Fau - Lindow SE, Lindow Se Fau - Post AF, Post Af Fau - Belkin S, et al. Distance-decay relationships partially determine diversity patterns of phyllosphere bacteria on Tamarix trees across the Sonoran Desert [corrected]. (1098-5336 (Electronic)).
- Yan K, Han W, Zhu Q, Li C, Dong Z, Wang Y. Leaf surface microtopography shaping the bacterial community in the phyllosphere: evidence from 11 tree species. Microbiological Research. 2022;254:126897. [CrossRef]
- Redford AJ, Bowers RM, Knight R, Linhart Y, Fierer N. The ecology of the phyllosphere: geographic and phylogenetic variability in the distribution of bacteria on tree leaves. Environ Microbiol. 2010;12(11):2885-93. PubMed PMID: WOS:000283737000002. [CrossRef]
- Lindow SE, Brandl MT. Microbiology of the phyllosphere. (0099-2240 (Print)).
- Lighthart B. The ecology of bacteria in the alfresco atmosphere. FEMS Microbiol Ecol. 1997;23(4):263-74. PubMed PMID: ISI:A1997XU71100001.
- Warren SD. Microorganisms of the Phyllosphere: Origin, Transport, and Ecological Functions. Frontiers in Forests and Global Change. 2022;5. [CrossRef]
- Koskella B. The phyllosphere. (1879-0445 (Electronic)).
- Rastogi G, Coaker GL, Leveau JHJ. New insights into the structure and function of phyllosphere microbiota through high-throughput molecular approaches. FEMS Microbiol Lett. 2013;348(1):1-10. [CrossRef]
- Schlechter RO, Miebach M, Remus-Emsermann MNP. Driving factors of epiphytic bacterial communities: A review. Journal of Advanced Research. 2019;19:57-65. [CrossRef]
- Burrows SM, Elbert W, Lawrence MG, Poschl U. Bacteria in the global atmosphere - Part 1: Review and synthesis of literature data for different ecosystems. Atmos Chem Phys. 2009;9(23):9263-80. PubMed PMID: ISI:000272689600013.
- Blanchard DC. The Ejection of Drops from the Sea and Their Enrichment with Bacteria and Other Materials - a Review. Estuaries. 1989;12(3):127-37. PubMed PMID: ISI:A1989AQ97400001.
- Blanchard DC. The production, distribution, and bacterial enrichment of the sea-salt aerosol. Air-sea exchange of gases and particles: Springer, Dordrecht; 1983. p. 407-54.
- Blanchard DC, Syzdek L. Bubbles and Water-to Air Transfer of Bacteria. Bull Amer Meteorol Soc. 1971;52(11):1136-41. PubMed PMID: ISI:A1971L248600015.
- Blanchard DC, Syzdek L. Mechanism for Water-to-Air Transfer and Concentration of Bacteria. Science. 1970;170(3958):626-&. PubMed PMID: ISI:A1970H672100024.
- Blanchard DC, Syzdek LD. Water-to-Air Transfer and Enrichment of Bacteria in Drops from Bursting Bubbles. Appl Environ Microbiol. 1982;43(5):1001-5. PubMed PMID: ISI:A1982NY69700005.
- Blanchard DC, Syzdek LD. Enrichment of Bacteria in Airborne Drops from Bubbles Bursting at the Surface of Bacterial-Laden Waters. Bull Amer Meteorol Soc. 1978;59(11):1516-. PubMed PMID: ISI:A1978GJ43200103.
- Blanchard DC, Syzdek LD. Concentration of Bacteria in Jet Drops from Bursting Bubbles. Journal of Geophysical Research. 1972;77(27):5087-&. PubMed PMID: ISI:A1972N516400004.
- Blanchard DC, Syzdek LD, Weber ME. Bubble Scavenging of Bacteria in Fresh-Water Quickly Produces Bacterial Enrichment in Airborne Jet Drops. Limnology and Oceanography. 1981;26(5):961-4. PubMed PMID: ISI:A1981MK65800016.
- Aller JY, Kuznetsova MR, Jahns CJ, Kemp PF. The sea surface microlayer as a source of viral and bacterial enrichment in marine aerosols. J Aerosol Sci. 2005;36(5-6):801-12. PubMed PMID: ISI:000229348600018.
- Dueker ME, Weathers KC, O'Mullan GD, Juhl AR, Uriarte M. Environmental Controls on Coastal Coarse Aerosols: Implications for Microbial Content and Deposition in the Near-Shore Environment. Environ Sci Technol. 2011;45(8):3386-92. PubMed PMID: ISI:000289341300029.
- Dueker ME, O’Mullan GD, Martinez J, Juhl AR, Weathers KC. Onshore wind speed modulates microbial aerosols along an urban waterfront. Atmosphere. 2017;8(11):215. [CrossRef]
- Baylor ER, Baylor MB, Blanchard DC, Syzdek LD, Appel C. Virus transfer from surf to wind. Science. 1977;198(4317):575-80. PubMed PMID: ISI:A1977DZ50800006.
- Lavy A, McGrath DG, Matheus Carnevali PB, Wan J, Dong W, Tokunaga TK, et al. Microbial communities across a hillslope-riparian transect shaped by proximity to the stream, groundwater table, and weathered bedrock. Ecology and Evolution. 2019;9(12):6869-900. [CrossRef]
- Dodds WK, Zeglin LH, Ramos RJ, Platt TG, Pandey A, Michaels T, et al. Connections and Feedback: Aquatic, Plant, and Soil Microbiomes in Heterogeneous and Changing Environments. BioScience. 2020;70(7):548-62. [CrossRef]
- Marks R, Kruczalak K, Jankowska K, Michalska M. Bacteria and fungi in air over the Gulf of Gdansk and Baltic sea. J Aerosol Sci. 2001;32(2):237-50. PubMed PMID: ISI:000166719100005.
- Dueker ME, O'Mullan GD, Weathers KC, Juhl AR, Uriarte M. Coupling of fog and marine microbial content in the near-shore coastal environment. Biogeosciences. 2012;9(2):803-13. [CrossRef]
- Evans SE, Dueker ME, (co-lead authors), Logan JR, Weathers KC. The biology of fog: results from coastal Maine and Namib Desert reveal common drivers of fog microbial composition. Sci Total Environ. 2019;647:1547-56. [CrossRef]
- Dueker ME, French S, O’Mullan GD. Comparison of Bacterial Diversity in Air and Water of a Major Urban Center. Frontiers in Microbiology. 2018;9.
- Dueker ME, O'Mullan GD. Aeration remediation of a polluted waterway increases near-surface coarse and culturable microbial aerosols. The Science of the total environment. 2014;478:184-9. Epub 2014/02/18. PubMed PMID: 24531127. [CrossRef]
- Dueker ME, O'Mullan GD, Juhl AR, Weathers KC, Uriarte M. Local environmental pollution strongly influences culturable bacterial aerosols at an urban aquatic Superfund site. Environ Sci Technol. 2012;46(20):10926-32. [CrossRef]
- Adomako MO, Yu F-H. Potential effects of micro- and nanoplastics on phyllosphere microorganisms and their evolutionary and ecological responses. Sci Total Environ. 2023;884:163760. [CrossRef]
- Ivashchenko KV, Korneykova MV, Sazonova OI, Vetrova AA, Ermakova AO, Konstantinov PI, et al. Phylloplane Biodiversity and Activity in the City at Different Distances from the Traffic Pollution Source. Plants [Internet]. 2022; 11(3).
- Mandal M, Das S, Roy A, Rakwal R, Jones OAH, Popek R, et al. Interactive relations between plants, the phyllosphere microbial community, and particulate matter pollution. Sci Total Environ. 2023;890:164352. [CrossRef]
- Laforest-Lapointe I, Messier C, Kembel Steven W. Tree Leaf Bacterial Community Structure and Diversity Differ along a Gradient of Urban Intensity. mSystems. 2017;2(6):10.1128/msystems.00087-17. [CrossRef]
- Guo L, Maghirang RG. Numerical Simulation of Airflow and Particle Collection by Vegetative Barriers. Engineering Applications of Computational Fluid Mechanics. 2012;6(1):110-22. [CrossRef]
- Wu S, Bashir MA, Raza Q-U-A, Rehim A, Geng Y, Cao L. Application of riparian buffer zone in agricultural non-point source pollution control—A review. Frontiers in Sustainable Food Systems. 2023;7. [CrossRef]
- de Santana CO, Spealman P, Azulai D, Reid M, Dueker ME, Perron GG. Bacteria communities and water quality parameters in riverine water and sediments near wastewater discharges. Scientific Data. 2022;9(1). [CrossRef]
- Chen XY, Ran PX, Ho KF, Lu WJ, Li B, Gu ZP, et al. Concentrations and Size Distributions of Airborne Microorganisms in Guangzhou during Summer. Aerosol Air Qual Res. 2012;12(6):1336-44. PubMed PMID: WOS:000313100300030. [CrossRef]
- Tang G, Fan Y, Li X, Tian R, Tang R, Xu L, et al. Effects of leaf properties on the counts of microbes on the leaf surfaces of wheat, rye and triticale. FEMS Microbiol Ecol. 2023;99(4):fiad024. [CrossRef]
- Odds FC. Interactions among amphotericin B, 5-fluorocytosine, ketoconazole, and miconazole against pathogenic fungi in vitro. (0066-4804 (Print)).
- Gaze WH, Zhang L, Abdouslam NA, Hawkey PM, Calvo-Bado L, Royle J, et al. Impacts of anthropogenic activity on the ecology of class 1 integrons and integron-associated genes in the environment. Isme Journal. 2011;5(8):1253-61. PubMed PMID: WOS:000295782200003. [CrossRef]
- Caporaso JG, Lauber CL, Walters WA, Berg-Lyons D, Huntley J, Fierer N, et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. The ISME Journal. 2012;6(8):1621-4. [CrossRef]
- Callahan BJ, McMurdie PJ, Rosen MJ, Han AW, Johnson AJA, Holmes SP. DADA2: High-resolution sample inference from Illumina amplicon data. Nature Methods. 2016;13(7):581-3. [CrossRef]
- McMurdie PJ, Holmes S. phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One. 2013;8(4):e61217. Epub 05/01. [CrossRef]
- 10.1371/journal.pone.0061217. Print 2013. PubMed PMID: 23630581.
- McMurdie PJ, Holmes S. Waste Not, Want Not: Why Rarefying Microbiome Data Is Inadmissible. PLoS Comput Biol. 2014;10(4). [CrossRef]
- 10.1371/journal.pcbi.1003531. PubMed PMID: 24699258.
- R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical.
- Computing, Vienna, Austria. 2018;URL https://www.R-project.org/.
- Laforest-Lapointe I, Messier C, Kembel SW. Tree phyllosphere bacterial communities: exploring the magnitude of intra- and inter-individual variation among host species. (2167-8359 (Print)).
- Laforest-Lapointe IA-O, Messier C, Kembel SW. Host species identity, site and time drive temperate tree phyllosphere bacterial community structure. (2049-2618 (Electronic)).
- Roldán DM, Kyrpides N, Woyke T, Shapiro N, Whitman WB, Králová S, et al. Hymenobacter artigasi sp. nov., isolated from air sampling in maritime Antarctica. Int J Syst Evol Microbiol. 2020;70(9):4935-41. [CrossRef]
- Kim Y, Subramanian P, Choi H, Weon HY, Kim S, Kwon SW, et al. Five novel Hymenobacter species isolated from air: Hymenobacter cellulosilyticus sp. nov., Hymenobacter cellulosivorans sp. nov., Hymenobacter aerilatus sp. nov., Hymenobacter sublimis sp. nov. and Hymenobacter volaticus sp. nov. LID. (1466-5034 (Electronic)). [CrossRef]
- Roldán DM, Kyrpides N, Woyke T, Shapiro N, Whitman WB, Králová S, et al. Hymenobacter caeli sp. nov., an airborne bacterium isolated from King George Island, Antarctica. LID (1466-5034 (Electronic)). [CrossRef]
- Erkorkmaz BA, Gat D, Rudich Y. Aerial transport of bacteria by dust plumes in the Eastern Mediterranean revealed by complementary rRNA/rRNA-gene sequencing. Communications Earth & Environment. 2023;4(1):24. [CrossRef]
- Xue H, Piao C-g, Wang X-z, Lin C-l, Guo M-w, Li Y. Sphingomonas aeria sp. nov., isolated from air. Int J Syst Evol Microbiol. 2018;68(9):2866-71. [CrossRef]
Figure 1.
Study sites near the Saw Kill (solid line, flow is right to left). Locations of A. rubrum used for leaf sampling denoted by white circles. Aerosol-creating waterfalls upstream of both riparian sites noted, along with the location of the outflow for the Bard College wastewater treatment plant.
Figure 1.
Study sites near the Saw Kill (solid line, flow is right to left). Locations of A. rubrum used for leaf sampling denoted by white circles. Aerosol-creating waterfalls upstream of both riparian sites noted, along with the location of the outflow for the Bard College wastewater treatment plant.

Figure 2.
Culture-based phyllosphere bacteria counts: (a) geometric mean and standard error of cfu’s per cm2 grown from leaf prints and normalized by leaf surface area (riparian n = 60, forest n = 60), (b) cfu’s per ml leaf wash (riparian n = 20, forest n = 20), and culture-independent (qPCR) abundances of (c) 16S copies (riparian n = 20, forest n = 20), and (d) IntI1 copies per ml leaf wash (riparian n = 20, forest n = 20). Statistically significant differences denoted by an asterisk (*).
Figure 2.
Culture-based phyllosphere bacteria counts: (a) geometric mean and standard error of cfu’s per cm2 grown from leaf prints and normalized by leaf surface area (riparian n = 60, forest n = 60), (b) cfu’s per ml leaf wash (riparian n = 20, forest n = 20), and culture-independent (qPCR) abundances of (c) 16S copies (riparian n = 20, forest n = 20), and (d) IntI1 copies per ml leaf wash (riparian n = 20, forest n = 20). Statistically significant differences denoted by an asterisk (*).
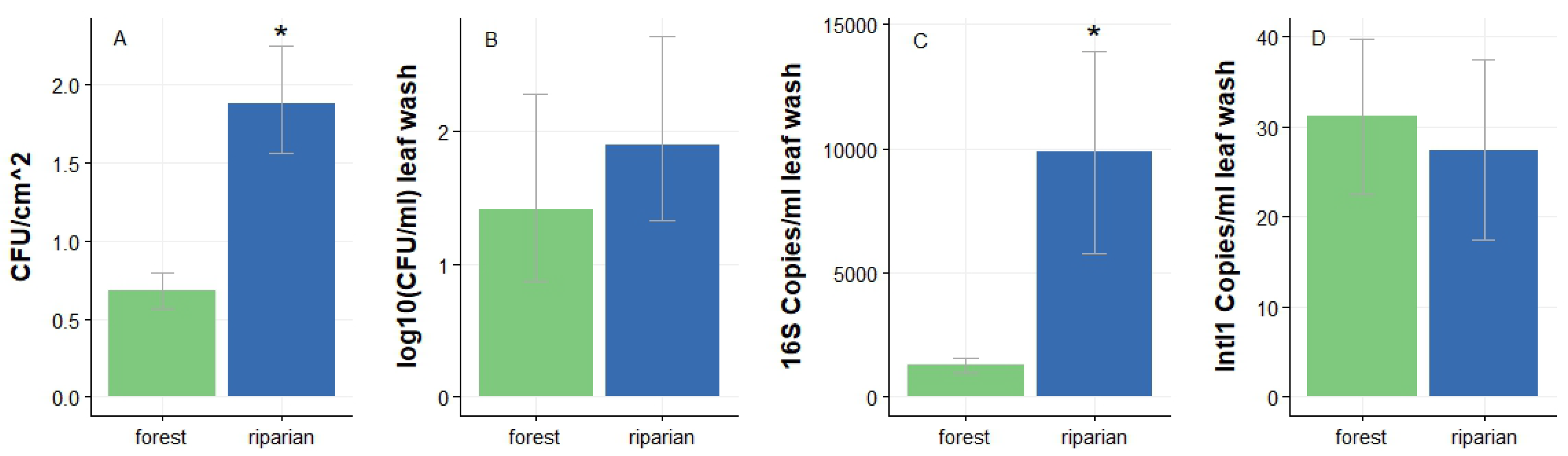
Figure 3.
Alpha diversity comparisons between forest leaf PMC samples (n = 18) and riparian leaf PMC samples (n = 20): (A) predicted ASV’s (Chao1 index) and (B) Shannon’s Diversity index calculated from non-rarefied samples. Boxes and lines denote data range and mean, and black points represent outliers. Green and dark blue points denote by-sample index value. Statistically significant difference denoted with an asterisk (*).
Figure 3.
Alpha diversity comparisons between forest leaf PMC samples (n = 18) and riparian leaf PMC samples (n = 20): (A) predicted ASV’s (Chao1 index) and (B) Shannon’s Diversity index calculated from non-rarefied samples. Boxes and lines denote data range and mean, and black points represent outliers. Green and dark blue points denote by-sample index value. Statistically significant difference denoted with an asterisk (*).
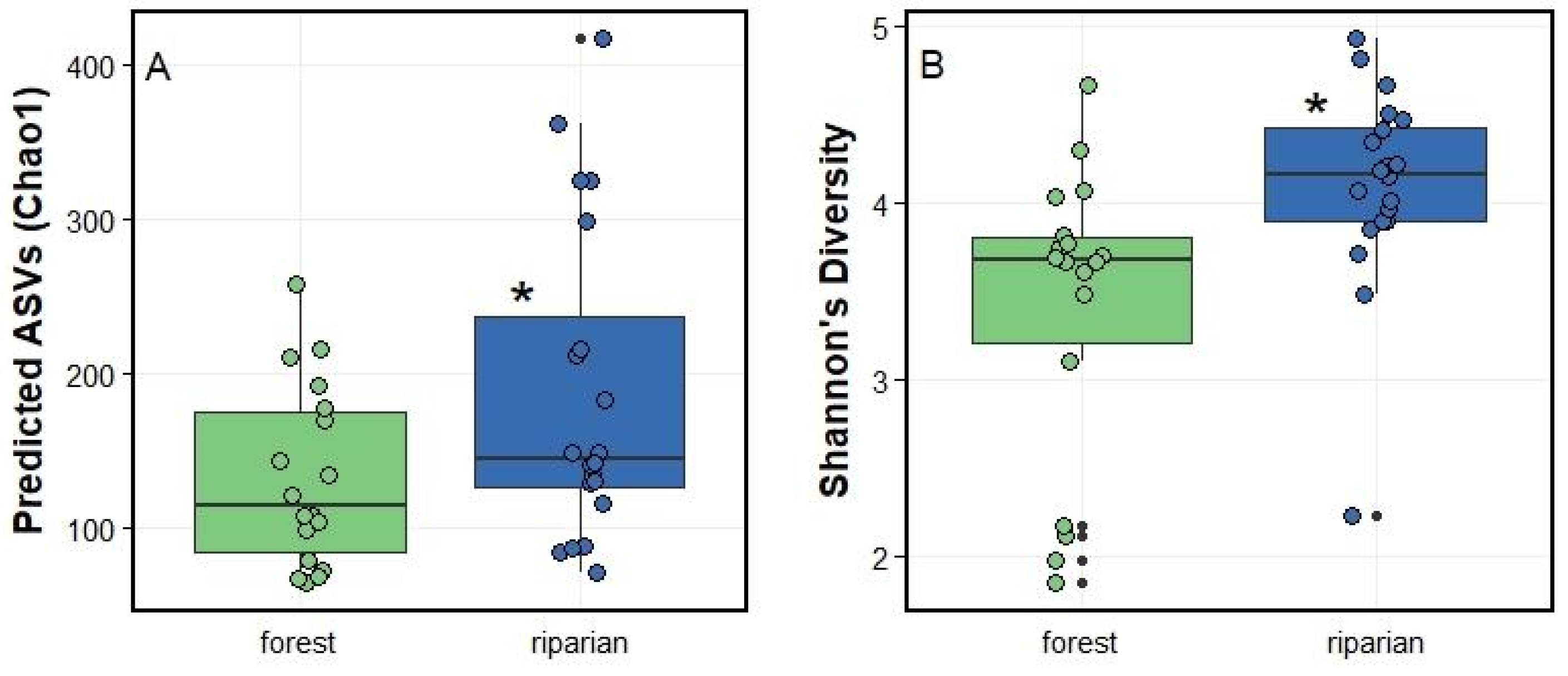
Figure 4.
Phylogenetic tree demonstrating the by-sample abundances of ASV’s identified as sewage-related (see Supplemental Table X) found on riparian (dark blue), and forest (green) phyllospheres. Each point represents a sample, point size relates to # of ASVs, ranging from 1 - 125.
Figure 4.
Phylogenetic tree demonstrating the by-sample abundances of ASV’s identified as sewage-related (see Supplemental Table X) found on riparian (dark blue), and forest (green) phyllospheres. Each point represents a sample, point size relates to # of ASVs, ranging from 1 - 125.
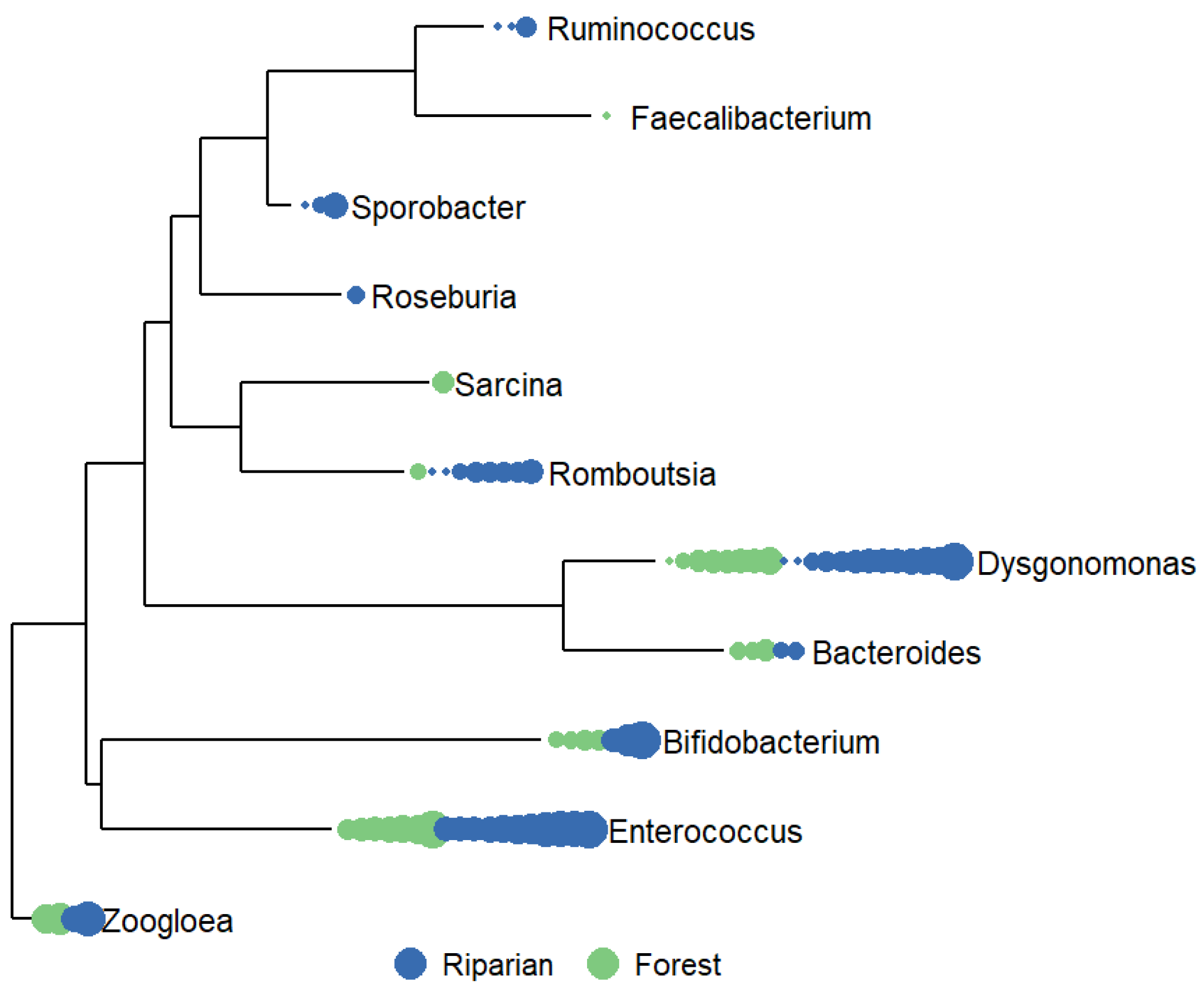
Figure 5.
Venn diagram demonstrating the number of shared ASVs between forest (green), riparian (dark blue), and water (light blue) microbial communities.
Figure 5.
Venn diagram demonstrating the number of shared ASVs between forest (green), riparian (dark blue), and water (light blue) microbial communities.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



