
Submitted:
21 June 2024
Posted:
21 June 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Gc protein-derived Macrophage Activating Factor (GcMAF), a powerful immunostimulant endowed with anti-cancer and anti-angiogenetic activities, offers significant advantages when combined with radiation therapy. A stronger immune response improves the effectiveness of radiation therapy by allowing the body to better eliminate residual cancer cells after treatment. In addition, by boosting the immune system, GcMAF mitigates some of the immunosuppressive side effects of radiation therapy, leading to faster recovery. In order to fully exploit the potential of GcMAF in cancer therapy, knowledge of the molecular interactions with its receptor is essential. This study proposes the first extracellular domain (residues 1-34) of the CCR1 protein as the GcMAF receptor. The CCR1 gene, expressed in monocytes and 168 other cell types or tissues, encodes this transmembrane protein. Electrostatic and hydrophobic interactions, along with hydrogen bonds mediate the molecular interactions between the TPT420-GalNAcELAK (or TPK420ELAK) sequences of GcMAF (or Gc2 protein variant) and the TTEDYDTTT sequence of its receptor.
Keywords:
radiation therapy
; GcMAF
; receptor
; immune system
; cancer
; autism
; COVID-19
Introduction
A serum protein of human plasma called group-specific component (Gc protein), or Vitamin D-binding protein, exhibits multifunctionality. It exists in three common variants: Gc1F, Gc1S, and Gc2. Notably, Gc1 carries a specific sugar chain (O-linked trisaccharide) attached to its Threonine residue at position 420 (Thr420). This trisaccharide consists of N-acetylgalactosamine (GalNAc), galactose and sialic acid in this specific order. (Nabeshima et al. 2020) Selective removal of sialic acid and galactose from Gc1 can convert it into a potent molecule known as Gc protein-derived macrophage activating factor (GcMAF), leaving only GalNAc linked to Thr420. The Gc2 variant lacks this sugar chain entirely since it has Lysine at position 420 instead of Threonine (T420K mutation); nevertheless, the Gc2 variant is able to activate macrophages in a manner superimposable to that of GcMAF (Nabeshima et al. 2020).
GcMAF holds promise as a potential therapeutic candidate for a number of conditions ranging from immunotherapy and antiangiogenic cancer treatment (Pacini et al. 2011; Ruggiero et al. 2014; Saburi et al. 2017), to autism (Bradstreet et al. 2012), neurological side effects of cancer chemotherapy (Morucci et al. 2015; Branca et al. 2015), and COVID-19 (Spadera and Spadera 2020). More specifically, GcMAF offers significant advantages when combined with radiation therapy. A stronger immune response improves the effectiveness of radiation therapy by allowing the immune system to better eliminate residual cancer cells after treatment. Elimination of residual cancer cells results from direct inhibition of cancer cell proliferation and cancer cell-stimulated angiogenesis as demonstrated by Pacini et al. (2012) as well as by macrophage-induced cancer cell apoptosis (Thyer et al. 2013). In addition, by boosting the immune system, GcMAF mitigates some of the immunosuppressive side effects of radiation therapy, leading to faster recovery. Finally, in analogy with what observed with anti-cancer chemotherapy (Morucci et al. 2015; Branca et al. 2015), GcMAF has the potential to minimize the neurological side effects of radiation therapy (Jacob et al. 2022). Therefore, identification of its human receptor and characterization of the molecular interactions between GcMAF and the receptor becomes crucial for understanding its therapeutic potential and developing targeted treatment strategies.
Materials and Methods
Sequences and structures of proteins and domains were studied using the database UniProt and, more specifically, the function designated “align” (UniProt Consortium 2023).
Results
Human Gc protein is known to interact with 65 unique human interactors, 8 of which have the characteristics of, or are associated with receptors. These are: nuclear receptor subfamily 2, group F, member 2; chemokine (C-C motif) receptor 1; cubilin (intrinsic factor-cobalamin receptor); G protein-coupled receptor 18; growth factor receptor-bound protein 2; ligand dependent nuclear receptor interacting factor 1; low density lipoprotein receptor-related protein 2; purinergic receptor P2X, ligand-gated ion channel, 6 (Huttlin et al. 2021). Of these, only chemokine (C-C motif) receptor 1 (CCR1, also known as HM145, LD78 receptor, Macrophage inflammatory protein 1-alpha receptor (MIP-1alpha-R), RANTES-R) has structure, cellular localization, and function consistent with the known biological and clinical effects of GcMAF. Consistent with the multiplicity of effects exerted by GcMAF in different tissues, CCR1 is expressed in 168 cell types and tissues in addition to monocytes (see Gene expression database Bgee, CCR1 - ENSG00000163823), and is the target of at least 11 different ligands (Tian et al. 2004).
CCR1 is a typical G-protein coupled transmembrane receptor constituted by 4 extracellular domains, 7 transmembrane helical domains, and 4 cytoplasmic domains (Shao et al. 2021). Since GcMAF and Gc2 are large molecules characterized by a great number of hydrophilic, polar amino acids (Figure 1), thus unable to cross the plasma membrane, the only permissible interactions are those with the extracellular domains of the receptor.
Identification of the extracellular domain and amino acid sequence responsible for binding the active site of GcMAF was made possible by the observation that both ThrGalNAC and Lysine in position 420 of, respectively, Gc1 and Gc2, bind to the active site of the receptor (for details, please see supplementary Figure 3 of Nabeshima et al. (2020)). Therefore, the most likely characteristic of the receptor binding site has to be the presence of negatively charged amino acids able to establish electrostatic and other non-covalent interactions with Lysine and GalNAc.
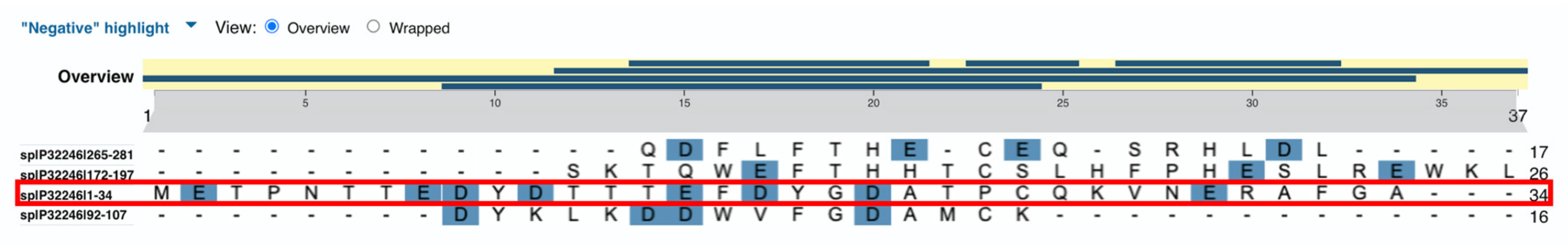
Figure 2 shows alignment of the sequences of the four extracellular domains of CCR1 where negatively charged amino acids are highlighted. The first domain (1-34; third line, outlined in red) has the highest number of negatively charged amino acids along with an unique sequence TTEDYDTTT where multiple Threonine residues flank Glutamic and Aspartic acid with a Tyrosine separating two Aspartic acid residues.
The sequence TTEDYDTTT of the first extracellular domain of CCR1 has all the features that make it the most likely candidate for binding both Lysine and GalNAc at position 420 of Gc2 and GcMAF, respectively.
Lysine can interact with Tyrosine, Glutamic, and Aspartic acid through ionic interactions and hydrogen bonding. Lysine side chain has a primary amine group that can become positively charged at physiological pH. In contrast, Glutamic, and Aspartic acid have side chains that can become negatively charged at physiological pH. Therefore, the positively charged Lysine side chain can attract the negatively charged side chains of glutamate and aspartate, forming strong ionic interactions. While ionic interactions are prominent, hydrogen bonding can also occur between Lysine and the other amino acids. The amine group of Lysine can act as a hydrogen bond donor. Tyrosine’s hydroxyl group on its side chain can act as a hydrogen bond acceptor. The carboxyl groups of Glutamic and Aspartic acid can also act as hydrogen bond acceptors. These hydrogen bonds, though weaker than ionic interactions, can contribute to stabilizing the ligand-receptor complex. While Tyrosine can’t participate in ionic interactions like Lysine, it can still form hydrogen bonds with Lysine’s amine group. The concomitant presence of ionic interactions with Glutamic and Aspartic acid can significantly increase the strength and specificity of the interaction between these amino acids.
In addition, Nabeshima et al. (2020) described the interaction of Thr418 of GcMAF and Gc2 with the receptor. The presence of multiple Threonine residues flanking the negatively charged amino acids in the sequence TTEDYDTTT of the first extracellular domain of CCR1 may be responsible for the interactions described by Nabeshima et al. (2020). Threonine residues facing each other on different proteins can interact with each other via hydrogen bonding and hydrophobic interactions. Each Threonine has a hydroxyl group on its side chain, which can act as both a hydrogen bond donor and acceptor. These hydroxyl groups can form hydrogen bonds with each other, creating a weak but stabilizing interaction between the two proteins. While not as strong as hydrogen bonding, weak hydrophobic interactions might occur. The side chain of Threonine has a methyl group; if the two methyl groups from the facing Threonine residues are positioned favorably, they experience attractive forces due to their hydrophobic nature.
The sequence TTEDYDTTT of the first extracellular domain of CCR1 also has all the features required to bind GalNAc. GalNAc can interact with Glutamic and Aspartic acid although the interaction wouldn’t involve electrical charge like with a positively charged amino acid such as Lysine. Hydrogen bonding is the most likely type of interaction between GalNAc and Glutamic and Aspartic acid. GalNAc. has hydroxyl groups on its sugar ring that can act as hydrogen bond donors whereas Glutamic and Aspartic acid both have carboxyl groups on their side chains that can act as hydrogen bond acceptors. In addition, GalNAc can interact with the Tyrosine that is flanked by Aspartic acid in the sequence TTEDYDTTT. Both Tyrosine and GalNAc have functional groups that can participate in hydrogen bonding. Tyrosine has a hydroxyl group on its side chain, which can act as a hydrogen bond donor. GalNAc has hydroxyl and carbonyl groups, which can act as hydrogen bond donors and acceptors, respectively. These interactions can occur between the side chain of Tyrosine and the hydroxyl or carbonyl groups of GalNAc, stabilizing a complex between the two molecules. The aromatic ring of Tyrosine can interact with the sugar ring of GalNAc through aromatic stacking interactions. These interactions can contribute to the overall stability of the complex. While both Tyrosine and GalNAc have hydrophilic groups, they also have some hydrophobic portions. Depending on the environment, weak hydrophobic interactions might occur between non-polar regions of the two molecules.
Discussion
Although the literature on GcMAF dates back to 1994 (Yamamoto et al. 1994), studies on its putative receptor are more recent. In 2013, the potential interaction between the vitamin D receptor (VDR) and GcMAF based on their structural features was described by Thyer et al. (2013). The authors hypothesized that the C-terminal 23 hydrophobic amino acids of VDR, located on the cytoplasmic side of the plasma membrane, interacted with the N-terminal 23 hydrophobic amino acids of GcMAF, positioned on the extracellular side. This interaction could potentially create a binding pocket for 1,25(OH)2D3 and oleic acid sandwiched between the two proteins. Such an interaction between GcMAF and VDR was proposed on the basis of previous observation that the response of human peripheral blood mononuclear cells to GcMAF was higher in cells from donors whose BsmI VDR polymorphism was the one that showed the highest response to 1,25(OH)2D3 and paricalcitol, a non-hypercalcemic vitamin D analogue (Pacini et al. 2012). Even though this hypothesis has not been disproven and might still be valid to explain some features of GcMAF responses, nevertheless, other hypotheses that have been put forward more recently merit consideration.
Nabeshima et al. (2020) propose that the GcMAF receptor is able to bind both the GcMAF characterized by GalNAc linked to Thr420 as well as the non-glycosylated Gc2, where Lysine at position 420 substitutes for GalNAc. In both cases, Thr418 participates in stabilizing the interaction. This hypothesis leads to the conclusion that there are at least two types of molecules with GcMAF activity, the bona fide GcMAF and Gc2. Therefore, the term GcMAF describes more of an activity rather than a single molecule. It is based on this hypothesis that the first extracellular domain of CCR1 was identified as the binding site for GcMAF and Gc2. However, if the term GcMAF might encompass an activity more than represent a single, well-defined molecule, then there are other molecules that have GcMAF activity, including sulfated polysaccharides such as chondroitin sulfate (Ruggiero et al. 2016), and fucoidan (Bakunina et al. 2018). Supporting this hypothesis, Pacini et al. (2011) observed that heparin inhibits the effects of GcMAF on human peripheral blood mononuclear cells. This inhibition might be due to competition for receptor binding, as Singh et al. (2015) demonstrated that heparin can bind to CCR1, and this binding is influenced by the sulfation pattern of heparin.
Another recent hypothesis concerning the GcMAF receptor comes from the article by Kirikovich et al. (2023) who demonstrated that, in a cell-free environment, GcMAF can interact with the protein encoded by the CLEC10A gene, a C-type lectin domain family 10 member A that is a single-pass type II membrane protein also known as Macrophage lectin 2. Although these results merit consideration since GalNAc is known to bind to this protein, it is worth noting that Macrophage lectin 2 is not listed among the interactors of human Gc protein and is difficult to envisage a role for this protein in angiogenesis, autism, or neuroprotection, conditions where GcMAF has proven effective. It is also possible that the interaction observed by Kirikovich et al. (2023) might be due to the “sticky” nature of Gc protein, a protein whose function is, among others, to bind actin as an actin scavenger (Otterbein et al. 2002). In addition, the mechanism proposed by Kirikovich et al. (2023) does not take into consideration the observation by Nabeshima et al. (2020) that Gc2 has GcMAF activity despite the absence of GalNAc. However, it is also possible that GcMAF binds different receptors either on the same cell or in different cell types. Given that GcMAF shows a number of biological effects that go well beyond activation of macrophages, this type of pleiotropy and redundancy cannot be ruled out.
Conclusion: Implications for Cancer Radiation Therapy
In a paradigm shift, radiation therapy is now understood to extend beyond local tumor control by inducing a systemic, or abscopal, effect on distant, untreated tumors. This newfound appreciation for radiation therapy’s ability to trigger an immune response has fueled the exploration of combining radiation therapy with immunotherapy (radiation-immunotherapy). However, several key questions remain, including the precise mechanisms of radiation therapy-immune system interaction, optimal radiation-immunotherapy treatment schedules, strategies to enhance the abscopal effect, and methods to overcome radiation-immunotherapy resistance. The concept of the “radscopal effect,” which utilizes low-dose radiation to reprogram the tumor microenvironment, presents a potential avenue to amplify the abscopal effect and overcome radiation-immunotherapy resistance. Collectively, these findings suggest that radiation therapy can be harnessed as a potent trigger of systemic antitumor immunity. When combined with immunotherapy, radiation therapy has the potential to become a cornerstone of curative and systemic treatment regimens for cancer patients (for rev. on radiation-immunotherapy, please see Zhang et al. 2022).
In the context of immunotherapy, GcMAF plays an unique role because of its multifaceted activities that target tumors from a variety of angles as reviewed in Ruggiero et al. (2014), and Saburi et al. (2017). The ongoing exploration of the GcMAF receptor holds significant promise for understanding its potential role in radiation-immunotherapy. The identification of the first extracellular domain of CCR1 as a binding site for both bona fide GcMAF and its unglycosylated variant, Gc2, suggests a broader concept of GcMAF activity beyond a single molecule. This opens doors to exploring the therapeutic potential of other molecules like sulfated polysaccharides that might share similar binding capabilities. Overall, the search for the GcMAF receptor sheds light on the intricate mechanisms of GcMAF activity. Future research focused on these recently identified interactions could pave the way for exploring the potential of GcMAF, as an immunomodulatory therapy to enhance the effectiveness of radiation therapy in cancer treatment.
Author Contributions
Marco Ruggiero, MD, PhD, National Coalition of Independent Scholars. 125 Putney Rd Battleboro, VT 05301, United States of America. email: marco.ruggiero@ncis.org
Funding
The author did not receive any funding for this study.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Ethics
This article is original and contains material that has not been submitted or published in any preprint server or scientific journal.
Acknowledgements
The author acknowledges the great work of Dr. Aldo Ruggiero, MD (1923-2006), pioneer of radiology in Prato, Italy, founder of the Studio Radiologico Ruggiero, source of boundless inspiration for this and many other scientific articles.
Advisory
No information in this paper is intended or implied to be a substitute for professional medical advice, diagnosis or treatment.
Conflicts of Interest
The author declares no conflict of interest in regard to the specific topics treated in this study.
References
- Bakunina I, Chadova O, Malyarenko O, Ermakova S. The Effect of Fucoidan from the Brown Alga Fucus evanescence on the Activity of α-N-Acetylgalactosaminidase of Human Colon Carcinoma Cells. Mar Drugs. 2018 May 10;16(5):155. [CrossRef]
- Bradstreet JJ, Vogelaar E, Thyer L. Initial observations of elevated alpha-N-acetylgalactosaminidase activity associated with autism and observed reductions from GC protein-macrophage activating factor injections. Autism Insights. 2012;4:31–8. [CrossRef]
- Branca JJ, Morucci G, Malentacchi F, Gelmini S, Ruggiero M, Pacini S. Effects of oxaliplatin and oleic acid Gc-protein-derived macrophage-activating factor on murine and human microglia. J Neurosci Res. 2015 Sep;93(9):1364-77. [CrossRef]
- Gene expression database Bgee, CCR1 - ENSG00000163823 https://www.bgee.org/gene/ENSG00000163823.
- Huttlin EL, Bruckner RJ, Navarrete-Perea J, Cannon JR, Baltier K, Gebreab F, Gygi MP, Thornock A, Zarraga G, Tam S, Szpyt J, Gassaway BM, Panov A, Parzen H, Fu S, Golbazi A, Maenpaa E, Stricker K, Guha Thakurta S, Zhang T, Rad R, Pan J, Nusinow DP, Paulo JA, Schweppe DK, Vaites LP, Harper JW, Gygi SP. Dual proteome-scale networks reveal cell-specific remodeling of the human interactome. Cell. 2021 May 27;184(11):3022-3040.e28. [CrossRef]
- Jacob J, Feuvret L, Simon JM, Ribeiro M, Nichelli L, Jenny C, Ricard D, Psimaras D, Hoang-Xuan K, Maingon P. Neurological side effects of radiation therapy. Neurol Sci. 2022 Apr;43(4):2363-2374. [CrossRef]
- Kirikovich SS, Levites EV, Proskurina AS, Ritter GS, Peltek SE, Vasilieva AR, Ruzanova VS, Dolgova EV, Oshihmina SG, Sysoev AV, Koleno DI, Danilenko ED, Taranov OS, Ostanin AA, Chernykh ER, Kolchanov NA, Bogachev SS. The Molecular Aspects of Functional Activity of Macrophage-Activating Factor GcMAF. Int J Mol Sci. 2023 Dec 12;24(24):17396. [CrossRef]
- Morucci G, Branca JJ, Gulisano M, Ruggiero M, Paternostro F, Pacini A, Di Cesare Mannelli L, Pacini S. Gc-protein-derived macrophage activating factor counteracts the neuronal damage induced by oxaliplatin. Anticancer Drugs. 2015 Feb;26(2):197-209. https://doi.org/10.1097/CAD.0000000000000177.
- Nabeshima Y, Abe C, Kawauchi T, Hiroi T, Uto Y, Nabeshima YI. Simple method for large-scale production of macrophage activating factor GcMAF. Sci Rep. 2020 Nov 5;10(1):19122. [CrossRef]
- Otterbein LR, Cosio C, Graceffa P, Dominguez R. Crystal structures of the vitamin D-binding protein and its complex with actin: structural basis of the actin-scavenger system. Proc Natl Acad Sci U S A. 2002 Jun 11;99(12):8003-8. [CrossRef]
- Pacini S, Morucci G, Punzi T, Gulisano M, Ruggiero M. Gc protein-derived macrophage-activating factor (GcMAF) stimulates cAMP formation in human mononuclear cells and inhibits angiogenesis in chick embryo chorionallantoic membrane assay. Cancer Immunol Immunother. 2011 Apr;60(4):479-85. [CrossRef]
- Pacini S, Punzi T, Morucci G, Gulisano M, Ruggiero M. Effects of vitamin D-binding protein-derived macrophage-activating factor on human breast cancer cells. Anticancer Res. 2012 Jan;32(1):45-52. [PubMed]
- Pacini S, Morucci G, Punzi T, Gulisano M, Ruggiero M, Amato M, Aterini S. Effect of paricalcitol and GcMAF on angiogenesis and human peripheral blood mononuclear cell proliferation and signaling. J Nephrol. 2012 Jul-Aug;25(4):577-81. [CrossRef]
- Ruggiero M, Ward E, Smith R, Branca JJ, Noakes D, Morucci G, Taubmann M, Thyer L, Pacini S. Oleic Acid, deglycosylated vitamin D-binding protein, nitric oxide: a molecular triad made lethal to cancer. Anticancer Res. 2014 Jul;34(7):3569-78. [PubMed]
- Ruggiero M, Reinwald H, Pacini S. Is chondroitin sulfate responsible for the biological effects attributed to the GC protein-derived Macrophage Activating Factor (GcMAF)? Med Hypotheses. 2016 Sep;94:126-31. [CrossRef]
- Saburi E, Saburi A, Ghanei M. Promising role for Gc-MAF in cancer immunotherapy: from bench to bedside. Caspian J Intern Med. 2017 Fall;8(4):228-238. [CrossRef]
- Shao Z, Shen Q, Yao B, Mao C, Chen LN, Zhang H, Shen DD, Zhang C, Li W, Du X, Li F, Ma H, Chen ZH, Xu HE, Ying S, Zhang Y, Shen H. Identification and mechanism of G protein-biased ligands for chemokine receptor CCR1. Nat Chem Biol. 2022 Mar;18(3):264-271. [CrossRef]
- Singh A, Kett WC, Severin IC, Agyekum I, Duan J, Amster IJ, Proudfoot AEI, Coombe DR, Woods RJ. The Interaction of Heparin Tetrasaccharides with Chemokine CCL5 Is Modulated by Sulfation Pattern and pH. J Biol Chem. 2015 Jun 19;290(25):15421-15436. [CrossRef]
- Spadera L, Spadera M. Potential role of GcMAF in suppressing the severity of COVID-19-induced immune responses: Lesson learned from HIV. Med Hypotheses. 2020 Nov;144:110293. [CrossRef]
- Thyer L, Ward E, Smith R, Fiore MG, Magherini S, Branca JJ, Morucci G, Gulisano M, Ruggiero M, Pacini S. A novel role for a major component of the vitamin D axis: vitamin D binding protein-derived macrophage activating factor induces human breast cancer cell apoptosis through stimulation of macrophages. Nutrients. 2013 Jul 8;5(7):2577-89. [CrossRef]
- Tian Y, New DC, Yung LY, Allen RA, Slocombe PM, Twomey BM, Lee MMK, Wong YH. Differential chemokine activation of CC chemokine receptor 1-regulated pathways: ligand selective activation of Galpha 14-coupled pathways. Eur J Immunol. 2004 Mar;34(3):785-795. [CrossRef]
- UniProt Consortium. UniProt: the Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2023 Jan 6;51(D1):D523-D531. [CrossRef]
- Yamamoto N, Lindsay DD, Naraparaju VR, Ireland RA, Popoff SN. A defect in the inflammation-primed macrophage-activation cascade in osteopetrotic rats. J Immunol. 1994 May 15;152(10):5100-7. [CrossRef] [PubMed]
- Zhang Z, Liu X, Chen D, Yu J. Radiotherapy combined with immunotherapy: the dawn of cancer treatment. Signal Transduct Target Ther. 2022 Jul 29;7(1):258. 10.1038/s41392-022-01102-y.
Figure 1.
Results of alignment of mouse Gc protein (VTDB_MOUSE. Upper line) and human Gc protein (VTDB_HUMAN. Lower line). Polar amino acids are highlighted.
Figure 1.
Results of alignment of mouse Gc protein (VTDB_MOUSE. Upper line) and human Gc protein (VTDB_HUMAN. Lower line). Polar amino acids are highlighted.

Figure 2.
Results of alignment of human CCR1 extracellular domains. The first domain (1-34) in the third line. Negatively charged amino acids are highlighted.
Figure 2.
Results of alignment of human CCR1 extracellular domains. The first domain (1-34) in the third line. Negatively charged amino acids are highlighted.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



