Submitted:
07 June 2024
Posted:
07 June 2024
You are already at the latest version
Abstract
Globally, Type 2 diabetes (T2D) and Cancer are the major cause of morbidity and mortality worldwide and is considered to be one of the most significant public health concerns of the 21st century. Over the next two decades, the global burden is expected to increase by approximately 60%. Several observational studies as well as clinical trials have demonstrated the health benefits of consuming whole grains and lower the risk of several chronic non communicable diseases including T2D and cancer. Cereals grains are the primary source of energy in the human diet. The most widely consumed pseudo cereals include (quinoa, amaranth and buckwheat) and cereals (wheat, rice and corn). From a nutritional perspective, both pseudo cereals and cereals are recognized for their complete protein, essential amino acids, dietary fibres and phenolic acids. The bran layer of the seed contains the majority of these components. Greater intake of whole grains than refined grains has been consistently linked to a lower risk of T2D and cancer. Due to their superior nutritional compositions, whole grains make them a preferred choice over refined grains. The modulatory effects of whole grains on T2D and cancer are also likely to be influenced by several mechanisms, some of these effects may be direct; while others involve altering the composition of gut microbiota, increasing the abundance of beneficial bacteria, and lowering harmful bacteria, increasing insulin sensitivity, lowering solubility of free bile acids, protein broken down into peptides and amino acids, producing short-chain fatty acids (SCFAs) and other beneficial metabolites that promote the proliferation in the colon which modulate the anti-diabetic and anti-cancer pathway. Thus the present review are two folded; first summarizes the recent knowledge about the nutritional composition and bioactive acids in pseudo cereals (quinoa, amaranth and buckwheat) and cereals (wheat, rice and corn); the second section summarizes and discusses the progress in recent human studies, such as observational (cross-sectional studies, case-control studies and cohort studies) and intervention studies in understanding their role in T2D and cancer including the potential mechanism. Overall according to the scientific data, whole grain consumption may reduce the incidence of T2D and cancer. Future studies should carry out randomized controlled trials to validate observational results and establish causality; in addition the current manuscript encourages to investigate the specific mechanisms by which whole grains exert their beneficial effects on health by examining the effects of different types of specific protein, dietary fibres and phenolic acids that might help to prevent or treat T2D and cancer.
Keywords:
pseudo cereals
; cereals
; protein
; dietary fibres
; phenolic acids
; T2D
; cancer
1. Introduction
Globally, Type 2 diabetes (T2D) and cancer are the leading causes of morbidity and mortality, and are considered as one of the most serious public health concerns of the 21st century. According to the International Diabetes Federation (IDF) 2021 report, estimated 537 million adults worldwide have diabetes, with 783 million expected to by 2045 [1]. Additionally, according to World Health Organization report (WHO) reports, every year 1.5 million deaths are directly attributed to diabetes worldwide in low and middle income countries [2]. The WHO 2023 report on cancer estimated that there were 10 million cancer related deaths and 20 million additional cases of the cancer worldwide. Globally, it is expected in 2040 that there will be over 30 million more cases of cancer worldwide, with the majority of cases in low and middle income countries [3]. Over the past several years, there has been a gradual increase in the number of cases and incidences of T2D and cancer. The increasing incidences of T2D and cancer worldwide represent a major risk to public health in under developed as well as developing countries. According to the American Institute for Cancer Research and U.S. Department of Health and Human Services declared the, ‘‘whole grains as an important part of a healthy diet’’ [4,5]. Several epidemiological studies have given convincing results for the significant role of the whole grain diet in lowering the incidence of T2D [6,7] and cancer [8–10]. In contrast, some researches on refined grains have found a negative relationship between high refined grain consumption and an increased risk of T2D [11–13] and cancer [14]. These findings support the health recommendations to replace refined grains with whole grains. Regarding to the American Association of Cereal Chemists International, whole grains are composed of intact ground, cracked or flaked kernels that contain a starchy endosperm, germ, and bran [15]. In contrast, refined grain products do not contain one or more integral kernel components.
Whole grains provide protein, dietary fiber, and phenolic acids, all of which are helpful to one's health [16–19]. Consuming whole grain protein can alter the composition of the gut microbiota, enhancing the helpful bacteria and suppressing harmful bacteria [20]. The stomach and small intestine digest whole grain protein, converting it into peptides and amino acids [20]. Amino acids are absorbed into the bloodstream and carried to the large intestine colon [21]. The gut microbiota in the colon ferments the amino acids, producing short chain fatty acids (SCFAs) and other metabolites that stimulate the proliferation of beneficial bacteria in the colon and also play an essential role in the reduction of T2D and cancer [18,22,23]. Furthermore, the bacterial gut population digests dietary fibers and produces SCFAs [24], which modulate gut hormone production, influence glucose and lipid metabolism which may reduce the risk of type 2 diabetes [24,25]. Furthermore, dietary fiber can remove nitrite from the stomach and reduce the amounts of nitroso compounds under high acid conditions, because nitrates enhance the risk of stomach cancer [26,27]. It has been demonstrated that phenolic acids and dietary fibres included in whole grains have a substantial impact on human health and protect against chronic non communicable diseases [28,29].
A comprehensive systematic review of 11 cohort studies published in 2022 by Ghanbari-Gohari et al, [7], discovered that whole grain consumption is related with a lower incidence of T2D in follow up of 463,282 individuals. The study found that intake of 50 g of whole grains was related with a 23% decreased incidence of T2D. The author indicated that increasing whole grains consumption might be more useful in lowering the risk of T2D. More recently, in 2024 a comprehensive review and meta-analysis of prospective trials in two prospective cohorts by Watling et al [8] explored the influence of whole grains on cancer and revealed that 16g/day of whole grains can reduce the risk of cancer. This study comprised 111,396 people from the prostate, lung, colorectal, and ovarian cancers and discovered that dietary fiber and whole grains were inversely related with liver cancer risk. The results of this study showed that consuming 10 g of dietary fiber and 16 g of whole grains per day has been associated with a decreased risk of cancer.
Consumption of whole grain products, which have much higher protein, dietary fibre, and phenolic contents than refined grains, and have a dietary fibre profile with a good balance of soluble and insoluble fibres, can surely make a significant impact in the reduction risk of diseases. In the meantime, nutritional approaches that emphasize grain intake are limited, particularly for whole grains [30–32]. While currently published recommendations are only available to the general population and are not influenced by the risk level of individual health outcomes [4,33], particular guidance for T2D and cancer prevention are urgently required.
Thus, the purpose of this review was to first discuss the nutritional compositions of pseudo-cereals (quinoa, amaranth, and buckwheat) and cereals (wheat, rice, and corn), as well as the essential and non-essential amino acids, dietary fibers (total, insoluble, and soluble), phenolic acids (Hydroxycinnamic acids and Hydroxybenzoic acids) and their antioxidant activates. Secondly, we have studied relationship between whole grain intake and the incidence of T2D and cancer from human studies, including observational and intervention studies, as well as the possible mechanism behind the intake of whole grains and T2D and cancer. Recent research indicates that whole grains including protein, dietary fiber, and phenolic acids may be more beneficial than individual isolated components. Future research should include more clinical trials, epidemiological and randomized controlled trials to validate observational results. Additionally, to investigate the specific mechanisms by which whole grains exert their beneficial effects on health by examining the effects of specific protein, dietary fibres and phenolic acids that might help to prevent or treat T2D and cancer.
2. Material and Methods
We have reviewed and summarized the nutritional characteristics of three targeted pseudo cereals (Quinoa, Amaranth and Buckwheat) and three cereals (Wheat, Rice and Corn) (Figure 1), including fat (%), ash (%), carbohydrate (%), crude protein (%), amino acid composition (g/100g), phenolic acids (µg/g) as well as their antioxidant activates. Following that, we have reviewer and discussed the recent findings concerning the link between whole grains consumption and chronic non communicable diseases like T2D and cancer. Then we have reviewed the current research progress on the relationship between whole grain consumption chronic non communicable diseases like T2D and Cancer. All of the materials for this review were searched in PubMed and Google Scholar, including human studies such as observational (cross-sectional, case-control, and cohort studies) and interventional research. In the current review, relevant data published in English language in peer-reviewed journals have been included for discussion. However, any information available in the form of conference abstracts, books, and unpublished studies was eliminated.
3. Nutritional Composition of Pseudo Cereals and Cereals
3.1. Fat in Pseudo Cereals and Cereals
In whole grains, groats contain the highest amount of fat. According to the Table 1 Amaranth contains the highest fat content (6.4 to 8.0%) [34–36] among all other grains followed by quinoa (4.9 to 7.5%) [37,38], buckwheat (1.4 to 7.4%) [39,40], corn (3.8-4.7%) [41,42], wheat (1.7-3.3%) [43–46], and rice (1.5-2.2%) [47,48] (Table 1). The highest fat level in pseudo cereal is due to a high concentration of unsaturated fatty acids, including linolenic acid. Among cereal grains corn contain the highest content of fat (3.8-4.7%) [41], which has primarily mono (30%) and poly-unsaturated (50%) lipids, with a little amount of saturated lipids (20%) [41,42]. Furthermore, fat is the second most important component of grain amaranth, with a larger concentration than other cereals. Fat is characterized by the high concentration of unsaturated fatty acids as well as saturated fatty acids. Linoleic acid is a major fatty acid in oil, making up more than half of the content of grain amaranth seeds [36]. The next highest is oleic acid (more than 20%), followed by palmitic acid (almost 20%) [49]. Amaranth contains two to three times the fat content of quinoa, buckwheat, and all other cereal grains, indicating how widely different grains are from one another.
3.2. Carbohydrates and Starch in Pseudo Cereals and Cereals
Carbohydrates are the primary nutritious components of pseudo-cereal grains, accounting for 60 to 80% of the seed dry weight (dw) [50–52]. Carbohydrates are essential in nutrition and affect various metabolic processes [25,53]. Quinoa, amaranth, and buckwheat, like other cereals, contain high levels of carbohydrates (50-75%), which are mostly composed of starch and non-starchy polysaccharides. According to the Table 1, we found that wheat contain the highest content of carbohydrate. Wheat (61.6-83.1%)[45,46,54–59], Buckwheat (63.1-82.1%) [50–52] [60–62], Rice (71.1-78.2%)[47,48,62] Quinoa (48.5-77.0%)[35,38,60,61], corn (65.0-74.3%) [33,41,42] and amaranth (63.1-70.0%) [34–36,50]. Starch is the primary carbohydrate, accounting for 58.1-64.2%, 65.0-75.0%, and 54.5-57.4% of total seed dry weight in quinoa, amaranth, and buckwheat, respectively [62]. Buckwheat starch contains more amylose (18.3-47% of total starch) than quinoa (11-12%) and amaranth (7.8-34.3%) [62]. Starch in the endosperm is a key component of rice grain, accounting for 50% to 90% of its dry weight [47,48]. The chemical composition of amaranth seeds is dominated by carbohydrates, with starch accounting for 50-60% of the overall mass of the seeds, or more than 90% of the carbohydrates present. Starch granules, which are found mostly in the endosperm, have a polygonal shape and a strong swelling capacity [63].
3.3. Protein in Pseudo Cereals and Cereals
The protein content and profile of all plants are determined by genotype and growing conditions. Compared to cereals, pseudo cereals offer superior nutritional value, primarily due to their higher protein levels, which vary from 9.1 to 16.7% quinoa [35,38,60], 13.1-21.5% for amaranth [34–36,50] and buckwheat 5.7–14.2% [50–52,60–62] and cereals wheat 8-19.0% [45,46,54–59], rice 6.8-7.3% [47,48,62] and corn 8.8-9.4% [41,42] Table 1.
The major protein fractions found in qunioa grains are 11S-type globulins and 2S albumins, which account for 27.9-60.2% and 13.2-42.3% of total seed proteins, respectively, followed by glutelins (18.1-31.6) and Prolamins comprise a lower protein proportion (0.5-19.3% of total seed protein) [63,64]. Amaranthine, an essential protein found in the globulins portion with a molecular weight of more than 300kDa and three subunits: pro amaranthine, one basic peptide, and one acidic [35]. Buckwheat seeds include primarily 8S and 13S globulin proteins. Furthermore, 2S globulins, glutelins, and prolamins are minor protein components of common and tartary buckwheat [22].
Quinoa is an excellent source of protein [65]. According to Abugoch's reports [66], the proteins found in quinoa grains include albumins (35%) and globulins (37%), with a smaller proportion of prolamins. Quinoa proteins are of equal grade to milk protein (casein) provide all essential amino acids [66,67]; thus, it is considered a complete diet [68]. Electrophoretic investigations revealed that quinoa proteins are composed of two major fractions: 11S-globulin and 2S-protein, 11S-globulin, also known as chenopodium [69], comprises nearly 37 percent of the total protein. This fraction consists of polypeptides with molecular weights of (22-23 and 32-39 kDa), as well as a relatively low concentration of amino acids (methionine and cysteine). The 2S-protein fraction has a molecular weight of 9 kDa and is rich in cysteine, arginine, and histidine, but low in methionine [69]. Proteins from amaranth seeds are rich in essential amino acids and have a high digestibility of around 90% [71–73]. Three primary proteins—albumin (40%), globulin (20%), and glutelin (25–30%)—as well as a little amount of prolamine (2-4%) are present in amaranth grains [74]. As a result of their inability to produce gluten, amaranth flour is recommended for those with celiac disease. Furthermore amaranth contain Lysine, which is a limit amino acid in cereals grains like wheat and others ranging from 363 to 421 mg/g N, the same quantity found in soy [71]. Additionally, compared to basic legumes (1.4% on average), amaranth proteins have a significant number of amino acids containing sulphur (2–5%), methionine, cystine, and cysteine [55,75,76].

Table 1.
Nutritional composition of pseudocerals (quinoa, amaranth and buckwheat); and cerals (wheat, rice and corn) grains in percentage (%).
Table 1.
Nutritional composition of pseudocerals (quinoa, amaranth and buckwheat); and cerals (wheat, rice and corn) grains in percentage (%).
| Pseudo cereals | ||||||||
| Whole grains | Moisture | Fat | Ash | Carbohydrates | Starch | Crude protein | References | |
| Quinoa | 8.2–13.1 | 4.9-7.5 | 4.1 | 48.5–77.0 | 58.1–64.2 | 9.1–16.7 | [35,37,38,60,62,76] | |
| Amaranth | 8.9–9.4 | 6.4 to 8.0 | 3.3 | 63.1–70.0 | 65.0–75.0 | 13.1–21.5 | [34–36,50,60,77] | |
| Buckwheat | 10-11 | 1.4- 7.4 | 1.33–3.09% | 62.1–82.1 | 54.5–57.4 | 5.7–14.2 | [35,39,40,51,52,60,78] | |
| Cereals | ||||||||
| Whole grains | Moisture | Fat | Ash | Carbohydrates | Starch | Crude protein | References | |
| Wheat | 12-13 | 1.72-3.3 | 1.7-1.9 | 62.6-83.1 | 60-75 | 8-19.0 | [43,45,46,54–59] | |
| Rice | 5.0-12.7 | 1.5-2.2 | 0.5-3.5 | 71.1-78.2 | 50- 90 | 6.8-7.3 | [47,48,62] | |
| Corn | 10.4 | 3.8-4.7 | 1.33-1.44 | 65.0-74.3 | 72.8 | 8.8-9.4 | [41,42] | |
It is known that buckwheat is an excellent source of lipids, antioxidants, organic acids, dietary fiber, phenolic acid, and high biological value proteins that do not form gluten and a balanced amino acid composition (high quantities of lysine and arginine, compared to cereals). Moreover, buckwheat seeds are mostly composed of two types of proteins: 8S and 13S globulins, also contain small amounts of 2S globulins, glutelins, and prolamins [79]. Buckwheat contains 5–14% protein, of which 35% is found in the endosperm, 55% in the embryo and the remaining portion is found in the shell [80,81]. On the other hand, cereal proteins contain 80–90% protein in the endosperm and 10-15% of the embryo [82].
Globally, rice as is the primary source of nutrition for the majority of people. According to Hirawan et al [47], 100 g of brown rice contains about 7.3 g of protein, 2.2 g of fat, and 71.1 g of carbohydrates. Furthermore, rice bran contains (10-15%) protein, making it more superior among other cereal grains due to its unique allergenic and anticancer attributes [83]. Similarly, whole corn possesses 8.8% protein, 3.8% fat, 1.33% ash, and 65.0% carbs and whole wheat contain 12.7% protein, 1.6% ash, 1.72% fat, and 61.6% carbs in100 g of whole wheat [43].
Table 1 represent the nutritional composition of pseudo cereal and cereals and the data indicates that buckwheat carbohydrate content ranges from 62.6 to 82.1% very close to that of wheat ranges 62.1 to 83.1%. According to the literature, wheat has the highest carbohydrate content while amaranth has the lowest, followed by buckwheat, rice, quinoa, and corn. In terms of protein content, Amaranth has the highest percentage (13.1-21.5%), while rice has the lowest (6.8-7.3%). The descending order of protein content of whole grains cereals and pseudo cereals is amaranth, wheat, quinoa, buckwheat corn, and rice, indicating that pseudo cereals have the highest protein content when compared to cereals. Furthermore, rice has the highest starch content among all other cereals grains, while buckwheat has the lowest; both wheat and amaranth have values of 60-75% and 65.0-75.0 % which are closely related to each other, respectively. Pseudo cereals are a popular trend in human diets because they are gluten-free (GF) and have high nutritional and nutraceutical value. Furthermore, consumption of these pseudo cereals plays an essential role in lowering the risk of numerous chronic non communicable diseases, such as T2D [6,7,24,25] and cancer [8–10,26,27]. In addition a recent, has highlighted the potential health benefits of pseudo cereals, presenting these crops as valuable resources for functional food creation [50,51]. However, more experiments are needed to confirm, carbohydrate, and protein contents of pseudo cereals and cereals.
3.4. Amino Acids in Pseudo Cereals and Cereals
Pseudo cereals have a balanced amino acid composition with greater amounts of lysine and arginine as compared to cereal grains, and have been recognized as a good source of high biological value proteins that do not produce gluten [84–87]. As previously discussed in the nutritional composition section of pseudo cereals and cereals, several studies have confirmed that quinoa contains high-quality protein and classified as gluten-free play a key role in celiac diseases [68]. Furthermore quinoa protein is a complete protein, containing all of the necessary amino acids, which are required for human growth and development and have a protein efficiency ratio comparable to milk casein [88]. Quinoa's main protein components are globulins and albumins [64,68]. Quinoa protein 2S-type albumin has a high cysteine, arginine, and histidine content [89]. Qunia contains the highest amount of Leucine 2.3-9.4 g/100 g, followed by Methionine 0.3-9.1 g/100 g protein, and the lowest concentration of Tryptophan 0.6-1.9 g/100 g protein in essential amino acids [61], while in non-essential amino acids, quinoa contains the highest concentration of Arginine, 6.9-13.6 g/100 g, and the lowest concentration of Cysteine, 0.1-2.7 g/100 g, respectively [61,64] Table 2. Amranth seeds proteins are rich in essential amino acids with a high digestibility of around 90% [70–72]. Amranth contain the highest conetnet of Lysine 4.8–8.00g/100 g followed by Leucine 4.2–6.9 g/100 g protein being lowest in Tryptophan 0.9–1.8 g/100 g in essential amino acid [50], while in non-essential amino acid Amaranth contain highest content of Glutamate, 14.4–17.7 g/100 g and lowest in Cysteine 2.1–3.6 g/100g respectively [36,55,63]. Furthermore, as discussed in the previous section, the major protein in buckwheat is a 1
3S globulin, which is regarded a rare vegetable protein with blood cholesterol-lowering properties. Buckwheat also includes lectins, which help to reduce the proliferation of both spontaneous and induced tumors [90]. Buckwheat has a balanced amino acid content, with a significant amount of lysine (4.2-8.6 g/100 g) and low levels of tryptophan (0.9-1.8 g/100 g) among essential amino acids amino acids [52,62], while contains the highest amount of Glutamate (23.2-24.4 g/100 g) and the lowest amount of Cysteine (0.8-3.5 g/100 g) among non-essential amino acids [52,61].
Wheat contain the highest amount of Leucine (4.1-6.3 g/100 g) and the lowest amount of Tryptophan (0.7-1.2 g/100 g) among essential amino acids [49,91,92], while it contains the highest amount of Serine (6.1-6.7 g/100 g) and the lowest amount of Proline (1.5-2.3 g/100 g) among non-essential amino acids [92–95]. Rice contain the highest amount of Phenylalanine 5.2-9.1 g/100 g, followed by Leucine 8.2-8.9 g/100 g protein and the lowest amount of Tryptophan 1.0-1.5 g/100 g in essential amino acids [49,91,96], while rice has the highest level of Glutamate 7.2-20.8 g/100 g and the lowest level of Cysteine 1.6-2.0 g/100 g in non-essential amino acids [93,96]. Furthermore corn contain the highest content of Phenylalanine 4.8-10.6 g/100 g, followed by Valine 3.6-5.1 g/100 g protein, and the lowest content of Methionine 0.6-0.7 g/100 g in essential amino acids [49,91,93,96], while corn has the highest content of Glutamate, 7.13-15.8 g/100 g, and the lowest content of Cysteine, 2.1-2.3 g/100 g, respectively [93,96,97].
Table 2.
Essential and non-essential amino acids in pseudo cereals and cereals.
| Essential amino acids | ||||||||||
| Pseudco Cereals | ||||||||||
| Whole grains | Th | Va | Ph | Is | Le | Me | Tr | Ly | Hi | References |
| Quinoa | 2.1–8.9 | 0.8–6.1 | 3.0–4.7 | 0.8–7.4 | 2.3-9.4 | 0.3–9.1 | 0.6–1.9 | 2.4–7.8 | 1.4–5.4 | [61] |
| Amaranth | 3.3–5.0 | 3.9–5.0 | 3.7–4.7 | 2.7–4.2 | 4.2–6.9 | 1.6–4.6 | 0.9–1.8 | 4.8–8.0 | 1.9-3.8 | [36,50] |
| Buckwheat | 3.9–4.0 | 2.3–6.1 | 1.3–7.2 | 1.1–4.1 | 2.2–7.6 | 0.5–2.5 | 0.7-1.8 | 4.2–8.6 | 1.8-4.9 | [52,63] |
| Cereals | ||||||||||
| wheat | 1.8-2.7 | 2.4-4.1 | 2.8-8.1 | 2.2-4.1 | 4.1-6.3 | 0.9-1.2 | 0.7-1.2 | 1.7-2.6 | 0.2- 1.3 | [49,91,92] |
| rice | 3.2-3.7 | 4.5-4.5 | 5.2-9.1 | 2.8-4.5 | 8.2-8.9 | 1.0-1.6 | 1.0-1.5 | 3.3-3.8 | 0.1-1.7 | [49,91,96] |
| corn | 1.1-4.0 | 3.6-5.1 | 4.8-10.6 | 2.3-4.6 | 1.3-3.8 | 0.6-0.7 | 0.6- 1.1 | 2.6-1.9 | 2.3- 2.6 | [49,91,93,96] |
| Non-essential amino acids | ||||||||||
| Pseudco Cereals | ||||||||||
| Whole grains | As | Glu | Se | Gly | Ar | Al | Ty | Cy | Pr | References |
| Quinoa | 8.0 | 13.2 | 3.4–5.7 | 2.2–6.1 | 6.9–13.6 | 3.2–5.7 | 2.5–3.7 | 0.1–2.7 | 2.3–5.5 | [61,64] |
| Amaranth | 7.3–10.7 | 14.4–17.7 | 4.9–9.3 | 6.7–15.2 | 8.7–15.6 | 3.5–6.2 | 3.3–3.7 | 2.1–3.6 | 2.82–4.6 | [33,55] |
| Buckwheat | 7.6–16.6 | 23.2–24.4 | 3.2–8.6 | 6.2–13.2 | 10.5-11.3 | 4.6–9.6 | 0.6–4.9 | 0.8–3.5 | 2.6–8.8 | [52,62] |
| Cereals | ||||||||||
| wheat | 4.2-6.6 | 2.8-3.5 | 6.1-6.9 | 4.6-6.31 | 4.7-7.2 | 3.8-5.4 | 1.8-3.8 | 1.4-3.0 | 1.5-2.3 | [92–95] |
| rice | 4.2-10.7 | 7.2-20.8 | 4.0-5.7 | 3.9-5.2 | 7.2-8.2 | 4.5-6.3 | 2.3-3.2 | 1.6-2.0 | 4.8-5.4 | [93,96] |
| corn | 4.7-6.0 | 7.13-15.8 | 5.0-6.4 | 2.5-4.0 | 4.3-10.3 | 5.1-7.9 | 3.0-4.8 | 2.1-2.3 | 1.1-2.8 | [93,96,98] |
Table 2: Amino acid compositions of pseudo cereals and cereals (g/100 g protein); T;Threonine, V;Valine, P;Phenylalanine, I;Isoleucine, L;Leucine, M;Methionine, T;Tryptophan, L;Lysine, A;Aspartate, G;Glutamate, S;Serine, H;Histidine, G;Glycine, A;Arginine, A;Alanine, T;Tyrosine, C;Cysteine, P;Proline.
As shown in Table 2, in the review of 9 essential amino acids (g/100 g protein) and 9 non-essential amino acids (g/100 g protein) in pseudo cereals quinoa, amaranth, and buckwheat; and cereals wheat, rice, and corn. In essential amino acids, except for quinoa, amaranth and buckwheat contains highest content of Lysine, while it is ranked fourth in quinoa. Furthermore lysine is most prevalent in pseudo cereal grains, and phenylalanine is most prevalent in cereal grains. Most interestingly, the lysine content of all pseudo cereal grains is closely related to each other, whereas cereal grains are lower in lysine. Methionine is second in quinoa, Leucine in amaranth, while leucine is ranked second in wheat and rice, and Valine is second in corn. In terms of non-essential amino acids, quinoa and wheat contain the most Arginine, while amaranth, buckwheat, rice, and corn contain the most Glutamate. Except for wheat, all other grains contain a small amount of Cysteine, which is ranked second in wheat, and the ranking becomes more variable in the rest of the essential amino acids found in pseudo cereals and cereals. Corn, quinoa, rice, buckwheat, wheat, and amaranth are in descending order of the 9 essential amino acids (g/100 g protein), whereas buckwheat, rice, amaranth, corn, quinoa, and wheat are in descending order of the 9 non-essential amino acids. Based on these comparisons, each grain has a unique amino acid composition. However, more research is needed on cereals, particularly pseudo cereals, which are a current trend in human diets because they are gluten-free (GF) grains with excellent nutritional and nutraceutical properties [68,84–86], to confirm the contents of essential and non-essential amino acids in protein of various whole grains.
- 1.
- Dietary fibers and phenolic acids in pseudo cereals and cereals, and their antioxidant properties.
Dietary fiber is defined by the European Food Safety Authority [99] as "non-digestible carbohydrates plus lignin, including non-starch polysaccharides". Functional fiber is made up of isolated, nondigestible carbohydrates that have physiological benefits in humans [100]. Dietary fibers are divided into two categories depending on their water solubility: soluble (e.g., β-glucan) and insoluble (e.g., arabinoxylan AX) [101]. AX molecules consist of a linear backbone of D-xylopyranosyl residues connected by β-(1-4) glycosidic interactions, α-L-arabinofuranosyl residues can be linked to D-xylopyranosyl residues at O-2 and O-3 locations (Figure 2) . AX has four structural elements: non-substituted, O-2 or O-3 monosubstituted, and disubstituted D-xylopyranosyl [102]. Ferulic acid may be esterified with arabinose residues at the O-5 position [103]. These ferulic acid structures can build bridges between AX chains, increasing the AX molecular weight while decreasing its water extractability. The presence of ester-linked hydroxycinnamic acids distinguishes graminaceous AXs. Trans-ferulic acid is the most abundant phenolic acid generated during alkaline hydrolysis of cereal grain cell wall material, present in considerably smaller amounts with cis-ferulic, sinapic, and trans- and cis-p-coumaric acids [104]. Ferulates and diferulates also interact with lignin, forming AX-lignin cross-links. However, on a molar basis, more than half of the ferulates in grains are monomeric and do not form dimers [105].
Amaranth, quinoa, and buckwheat are pseudo cereals with a long history of use as food sources having extremely interesting nutritional properties [106]. Pseudo cereals have grown in popularity as a component in gluten-free goods during the last decade. Although dietary fiber amount varies across quinoa species, quinoa contains around 7 to 26% TDF [35,61,66,107,108], whereas amaranth's total dietary fiber content varies between 9 and 21% (dry weight basis) [35,50,107,109,110]. Of the pseudo cereal grains, buckwheat has the lowest amount of dietary fiber [33,35,51,111] (Table 3). Comparing quinoa and amaranth contain higher amount of dietary fiber (~22-26%) than wheat (about 17%) [112,113], rice (about 10%) [114,115], corn (about 19%) [116,117]. In comparison to amaranth and quinoa, buckwheat groats have a lower total dietary fiber content (7–19.0%) [33,35,51,111], the water soluble fiber in buckwheat seeds is mostly classified as pectin, arabinogalactan, and xyloglucan [118]. Moreover, pectin was found in the outer and inner epidermis' cell walls as well as the endosperm of buckwheat seeds [118].
Phenolic acids are a type of secondary metabolite that belongs to a larger group of phenolic compounds found throughout the plant kingdom. They have been considered essential dietary ingredients contributing to flavor, color, and nutritional value. Phenolic compounds are distinguished by the presence of one or more aromatic rings linked by one or more hydroxyl groups. Benzoic acid and cinnamic acid derivatives are examples of phenolic acids. In general, "phenolic acids" refer to phenols with a single carboxylic acid activity. These naturally occurring phenolic acids have two distinct carbon structures: hydroxycinnamic and hydroxybenzoic. Although the fundamental structure stays constant, the number and location of hydroxyl groups on the aromatic ring dictate the diversity. Hydroxycinnamic acids are a group of aromatic carboxylic acid with a C6-C3 structure, whereas hydroxybenzoic acids are C6-C1 (Figure 2).
In (Table 3), we reviewed the phenolic acids in targeted cereal grains, indicating that cereal grains have a higher amount of hydroxycinnamic than pseudocereal grains. On the other hand, pseudo cereal grains have the highest amount of hydroxybenzoic acids compared to cereal grains. Quinoa grain contains the highest phenolic acid level with 1672-3083 µg/g dry weight [119], whereas buckwheat has the lowest amount contains 49.9 µg/g [120] followed by corn 601-1740 µg/g [121,122], wheat has 658-1171 µg/g [123], amaranth has 212-570 µg/g [50,51], and rice has 300-360 µg/g [121].
Hydroxycinnamic and hydroxybenzoic acids are phytochemicals found in certain foods.It should be highlighted that in humans, circulating hydroxybenzoic acids can be the result of bacterially mediated polyphenol metabolism in the lower intestine [124,125]].Whole grains cereals and pseudo-cereals have differing amounts of hydroxycinnamic and hydroxybenzoic acids. According to Table 2, the highest content of hydroxycinnamic acids is found in corn (5.7-1387.5 µg/g) [126,127], while the lowest content is found in amaranth (1.6-55.4 µg/g ) [128] followed by rice (1.0-301.7 µg/g) [121,129], wheat (3.4-195.0 µg/g) [129,130], quinoa (7.0 to 150.0 µg/g) [131,132], and buckwheat (1.7-122.8 µg/g) [120,133] respectively. Moreover, wheat contain the highest content of of hydroxybenzoic acids (7.5 to 230.6 µg/g) [129,130] and and corn the lowest (0.5-116.5 µg/g) [126,127] followed by quinoa (13.8-110.0 µg/g) [132], amaranth (1.8-173.5 µg/g) [128], wheat (1.2-118.0 µg/g) [120,133] and rice, which ranges from 2.8 to 115.6 µg/g [126,127].
The antioxidants activities of the targeted grains are summarized in Table 4, which shows that corn contain the highest antioxidant activities among all other pseudo cereals and cereal. Cereal bran is the rich source of antioxidants activities [135]. For example, DPPH corn contain the highest content 350.29 mmol Trolox/100g [71,136] followed by rice 180.41 mmol Trolox/100g [137–139] and wheat contain the lowest among all other grains (20.9 mmol Trolox/100g) [140–144].
Compared to other grains amaranth possess the highest content of ABTS and FRAP 179.8 µmolTrolox/100g, 147.4 µmolTrolox/100g [145–147]. The content become variable in rest of grains. Overall, maize contains the highest content of antioxidant activity [136,148–150], followed by amaranth [135,146,151], quinoa [147,151–153], wheat [140–144], Buckwheat [151,154–156], rice [137–139,144].
In summary, table 3 shows that quinoa has the greatest total dietary fibre content (7 to 26%) followed by amaranth (2 to 21%), while rice has the lowest dietary fibres level among cereals (2-10%). In pseudo cereals, quinoa has the highest amount of total dietary fibres and buckwheat has the lowest, whereas corn has the highest content of total dietary fibre and rice contain the lowest. The ranking become variable in the rest of grains. Furthermore, in (Table 3), we have summarized the phenolic acids in targeted grains, which indicate that the amount of hydroxycinnamic is more abundant in cereals grains than in pseudo cereals grains. On the other hand, pseudo cereal grains have a higher content of hydroxybenzoic acids than cereal grains. According to table 4, amaranth has the highest level of antioxidant activity in pseudo cereals, whereas corn has the highest content among cereal grains. Overall, maize contains the highest content of antioxidant activity followed by amaranth, quinoa, wheat, buckwheat and rice. The rank become variable i in the remaining grains. These comparisons indicate that each grain prefers a different synthesis pathway to the others, resulting in a distinct profile of bioactive compounds in grains. Whole grains contain various nutrients, including dietary fiber, phenolic acids, and antioxidants. Table 4 shows that maize has the highest antioxidant content, whereas rice has the lowest. The germ and bran contain the majority of the bioactive components, which are reducing during the grain refining process. However, further research is needed to verify their results. Whole-grain consumption has been associated to a lower risk of chronic noncommunicable illnesses such as type 2 diabetes [6,7,24,25] and cancer [8–10,26,27], since it improves glycemic management, blood lipids, body weight, and inflammation while lowering premature mortality [102,157]. However, more study and communication on these health advantages is needed to translate the science behind these beneficial effects into useful data. Because these health advantages are interconnected, frequently synergistic, and individual-specific, it is difficult to obtain solid evidence of the health impacts associated with any bioactive component found in grain. Nowadays, consumers today are attracted to value-added products and are health-conscious, so it will be beneficial for customers if supplements are added or replace the refined grains with whole grains of high nutritional value. Furthermore, public education is also necessary to encourage people to eat more whole grains at the recommended amounts.
- 2.
- Grains in modern time
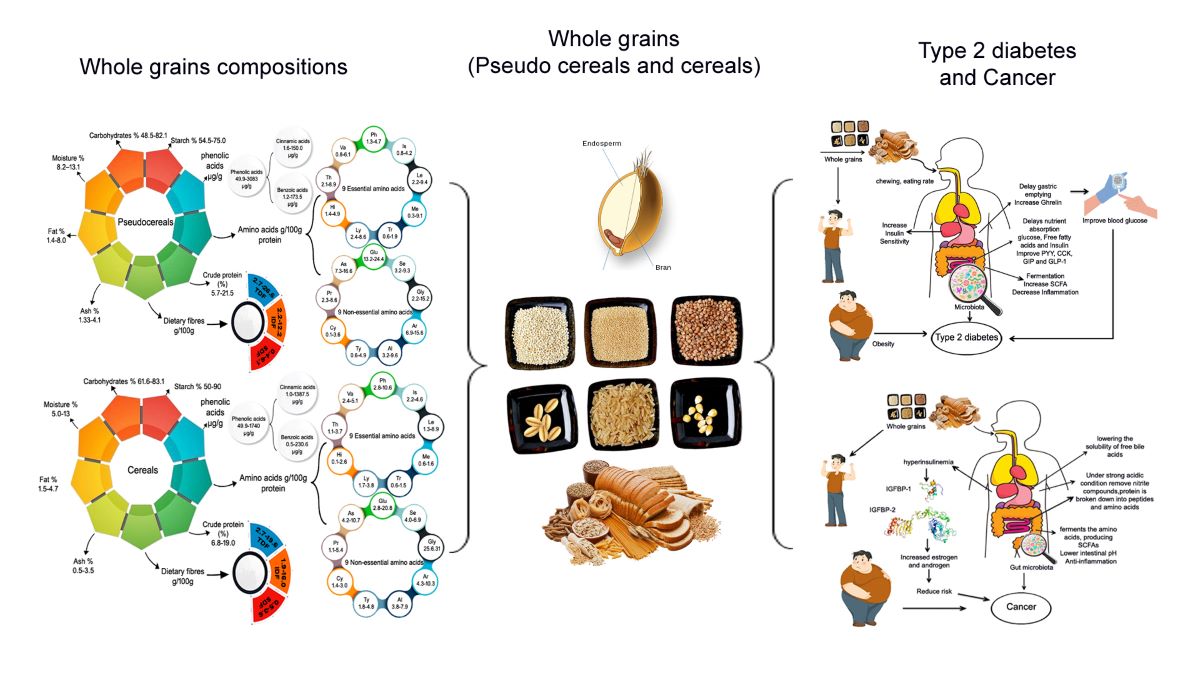
Whole grain Cereals and pseudo-cereals are currently popular in human diets due to their high nutritional and nutraceutical value. As previously discussed in details, whole grains have excellent nutritional properties have been linked to a reduced the risk of numerous chronic non communicable diseases, including T2D [6,7,24,25] and cancer [8–10,26,27]. In this context, it was critical to review the nutritional and functional profile of whole grains, which possess a high nutritional profile and are recognized as a super grain in a modern time. Whole grains pseudo cereals and cereals are known as "the grains of the a modern time" due to their high nutritional content [158]. Pseudo cerals Quinoa(Chenopodium quinoa Willd.), Amaranth (Amaranthus spp.) and Buckwheat (Fagopyrumesculentum Moench.) are promising crops for the future; they are gluten-free and have high nutritional and nutraceutical value [50,51,68]. Furthermore, recent study has highlighted the potential health advantages of pseudo cereals, confirming these crops as essential resources for functional food development [50,51]. Furthermore, whole grain cereals such as wheat (Triticum aestivum L., Triticum durum Desf.), rice (Oryza sativa L.), and corn (Zea mays L.) are a major source of energy worldwide. Cereal grains and its products have a long history of use by human. Cereals are staple foods that provide essential nutritional value in both low and middle income countries. Both cereals and pseudocereals are abundant in protein and carbohydrates and they contain a good balanced of essential amino acid composition that is characterized by an abundance of sulfur-rich amino acids, they are also a good source of dietary fiber and phenolic acids, with the highest amount of these compounds found in bran and germ (Figure 3). Consumers nowadays are health conscious and are attracted towards value-added products, so the addition/incorporation of supplements or completely replace the refined common cereals with whole grains of higher nutritional value will benefit the consumers. There is also a need to educate the public to increase their intake of whole grains to the recommended levels.
5. Intake of of Whole Grains and Human Health
5.1.1. Relationship between Intake of Pseudo Cereals and Cereals; and T2D
T2D is a leading cause of morbidity and mortality globally, particularly in low-income countries [159].According to the International Diabetes Federation 2021 report, 537 million adults worldwide suffer from diabetes, with the predication of 783 million will be affected by 2045, indicating an increased risk in the coming decades [1]. Furthermore, according to a WHO report, worldwide about 422 million people have T2D and 1.5 million deaths are directly related to T2D each year. The number of cases and prevalence of T2D have steadily increased over the last few decades [2]. T2D is associated with a number of lifestyle factors including diet, physical activity, obesity, alcohol, smoking, and a poor diet, among them diet is the most important factor [160]. Whole grains pseudo cereals and cereals are essential parts of a healthy diet [161], contain the outer bran, germ, and inner endosperm [162], all of which are high in dietary fiber, antioxidants, and micronutrients [18], which have attracted researchers’ interest in investigating the influence of consuming whole grains on human health [163]. Intake of whole grains pseudo cereals and cereals were found beneficial for T2D [17,164–173] (Table 5). In contrast, some researches on refined grains have found a relationship between high refined grain consumption and an increased risk of T2D [174–177]. In a 4-week prospective and double-blind randomized trial on T2D found that consuming 25 g of quinoa flakes significantly reduced blood triglycerides, low density lipoprotein, and total cholesterol [164]. The authors have also found to have antioxidant abilities in rats on a high-fructose diet, decrease blood glucose, triglycerides, low density lipoprotein, and total cholesterol. Furthermore A recent study by Khan et al. [165] found that quinoa powder can significantly reduce blood glucose levels (10-40%) in an animal model over 5 weeks. The goal of this research was to examine the chemical composition, total phenolic and antioxidant activity of quinoa seed powder, and its effect on diabetic rats. The author discovered that consuming quinoa at various concentrations reduced blood and glucose levels from 236.7 mg/dl to 98.7 mg/dl at 40% quinoa and 120.3 mg/dl at 30% quinoa, with an average fasting level of 64.3 mg/dL. Furthermore, rats fed quinoa seed powder had significantly decreased thyroid hormone levels (T3 and T4). Kasozi et al. [166] conducted an animal study to determine the key changes in s100a1 protein levels, as well as antioxidant and histopathologic changes in blood, renal, and hepatic tissues of male diabetic Wistar rats. This study discovered that grain amaranth supplementation increased expression of the s100a1 calcium transport proteins, which leads to enhanced calcium homeostasis in tissues. The significant antioxidant activity of grain amaranth stimulates increased s100a1 protein levels, resulting in improved tissue protection against oxidative stress, which is frequent in T2D. Furthermore, [17] investigated the inhibitory activity of protein hydrolyzates derived from amaranth grain and their effect on postprandial hyperglycemia found that protein hydrolyzate can enhance glucose tolerance. Increased plasma insulin levels in both acute and chronic conditions. These studies give evidence for grain amaranth's mechanism of action in the treatment of T2D. In another animal study, [167] found that dietary SDF supplementation decreased fasting blood glucose levels, increased oral glucose tolerance, increased liver glycogen and insulin levels, and improved serum and liver lipid profiles [168]. Furthermore, Kyrø et al., [169] investigated the relationship between whole-grain intake and T2D, demonstrated that individual who consume 16 g of whole grains per day have an 11% and 7% decreased risk of type 2 diabetes, respectively. In this regard [170] found that consuming black grain wheat (>69 g/d) for 5 weeks an increase in TNF-α and IL-6 levels, indicating a potential risk reduction for T2D. A higher intake of whole grains and various commonly consumed whole grain foods, such as whole grain breakfast cereal, dark bread, brown rice, added bran, and wheat germ, was substantially related with a decreased risk of T2D. These findings provide support to the present recommendations for increasing whole grain consumption as part of a healthy diet to prevent T2D [171]. On the other hand, higher consumption of white rice is linked to an increased risk of incidence of T2D. In this regard, several studies have demontrated the association between white rice consumption and an increased risk of T2D [174–177]. Furthermore, Huang et al [172] found that, purple corn have anti-diabetic benefits by protecting pancreatic β-cells, increasing insulin production, and activating protein kinase in the liver [172]. The author suggests that it may be beneficial in the prevention of T2D and its complications. In another similar study, Consumption of maize and wheat peptide decreased serum levels and pancreatic gene expression of IL-6 and insulitis, while increasing pancreatic β-cell areas, pancreatic gene expression of IL-10, and serum levels of serine and histidine [173]. Meanwhile, the serum levels of serine and histidine were significantly increased in mice treated with this peptide mixture. These findings indicated that a combination of corn and wheat peptides could prevent T2D.
5.1.2. Potential Mechanism of Whole Grains and T2D
Potential factors behind the relationship between whole grain consumption and a lower prevalence of T2D include their nutritional value, protein, dietary fiber, and phenolic acid. Consuming whole grains may reduce the incidence of T2D through various mechanisms. The quantity of chewing required when eating whole grains varies according on particle size and structural integrity. Increased chewing may improve satiety by increasing stomach distention and stimulating gut hormone responses. Whole grain proteins slow carbohydrate digestion and absorption, lowering postprandial glucose levels and increasing insulin sensitivity [178]. Whole grain protein has anti-inflammatory properties, which help to reduce chronic inflammation that causes insulin resistance and T2D [179,180]. Whole grain protein feeds the healthy gut bacteria, which produces short-chain fatty acids that increase glucose metabolism and insulin sensitivity. Whole grain protein promotes satiety, which leads to weight management and a lower risk of T2D [181]. Many wholegrain components can help improve glucose metabolism, hence preventing T2D. Dietary fiber, in particular, has received a lot of attention since whole grains are high in fiber. Previous studies have shown that people who consume more whole grains gain less weight [182]. Dietary fibers (especially viscous fibers) can promote stomach distension, which stimulates satiety signals and increases hormones involved in body weight regulation, energy homeostasis, and glucose control. Wholegrain fiber has the ability to increase gastric distension and delay intestinal transit time, thereby stimulating satiety signals and increasing hormones involved in energy homeostasis and plasma glucose control (Ghrelin, peptide PYY; CCK: cholecystokinin; GIP: gastric inhibitory peptide; GLP-1: glucagon-like peptide) (Figure 4) [182]. Furthermore, fiber from whole grains inhibits nutrient absorption (glucose, free fatty acids) at the intestinal level, lowering insulin demand and stimulating fat oxidation, all of which contribute to fat storage reduction. Wholegrain changes gut microbiota composition and stimulate fiber fermentation, which results in SCFA synthesis. Consuming whole grains can also help prevent T2D by improving insulin sensitivity in the body [183]. Furthermore, consuming whole grains can help improve blood glucose levels by increasing satiety signals and associated hormones. Dietary fibers can control postprandial blood glucose levels by delaying stomach emptying, increasing transit time, and glucose absorption [184]. Aside from fiber, whole grains can reduce the incidence of T2D by lowering inflammatory markers such as C-reactive protein [180]. Higher levels of specific liver enzymes, such as aspartate aminotransferase, can make the body more susceptible to T2D, although these markers can be maintained within normal limits by taking whole grains [185]. Furthermore, the bacterial gut population digests insoluble fibers and produces SCFAs, which can mediate gut hormone production, impact glucose and lipid metabolism, and contribute in the risk of T2D [24,25]. Some bioactive compounds found in whole grains may play a beneficial role in T2D [186,187]. In particular, phenolic compounds and their antioxidant and anti-inflammatory properties may help to reduce the development and progression of T2D by inhibiting oxidative stress, inflammatory cytokine transcription, and chronic low-grade inflammation [188,189], thereby improving insulin sensitivity [190]. A polyphenol-rich diet has recently been found to increase glucose tolerance and insulin sensitivity in non-diabetics while also lowering the postprandial triglyceride response [191]. Finally, bioactive substances included in whole grains may help to enhance insulin sensitivity and slow the development and progression of type 2 diabetes by reducing oxidative stress, inflammatory cytokine transcription, and subclinical inflammations.
In summary: The global incidence of T2D has become a significant risk to human health in both under developing and developed countries. Diet is the most important component in minimizing the risk and controlling the development of T2D. Epidemiological studies suggest the consumption of whole grains in lowering the risk of T2D. Because the whole grains are rich in protein, dietary fibres and phenolic acids. We discovered a strong inverse link between whole grain consumption and T2D. Furthermore, quinoa seeds have significant benefits over other crops in terms of human nutrition and health maintenance. Quinoa seeds should be recommended for commercial use in everyday routines since they have the potential to provide additional protection against T2D. Moreover Grain amaranth supplementation boosted the calcium content of the food and improved calcium signaling in diabetic rats' blood, kidneys, and liver. This study provides evidence for the mechanism of action of grain amaranth in the treatment of T2D, as it is widely utilized in various underdeveloped countries. More research, however, is needed to determine the role of other components found in amaranth, as this would provide more comprehensive information of the synergistic and antagonistic roles performed by diverse pathway mechanisms. Furthermore, the authors emphasized the calcium concentration in grain amaranth; nevertheless, the effects of oxalates, exogenous calcium, and other chemical elements under feed supplementation would provide additional information that would guide prospective clinical studies. On the other hand, study on white rice consumption is related with a considerably increased risk of type 2 diabetes [174–177], and higher intake of brown rice consumption was significantly associated with a decreased risk of type 2 diabetes. The findings of several studies support the current recommendations to increase whole grain consumption as part of a healthy diet for the prevention of type 2 diabetes [17,164–173]. Furthermore Whole grains' modulatory effects on T2D are likely regulated by a variety of pathways, including the effects of protein, dietary fiber, and phenolic acids. This represents a significant research gap that must be filled by well-designed and randomized clinical studies. However, researchers have discovered largely contrasting results in this subject, emphasizing the need for additional investigation. More research on each of the impacts of bran, protein, dietary fiber, and phenolic acids on T2D is needed to fill the gap in the link between whole grains and T2D.
5.1.1. Relationship between Intake of Pseudo Cereals and Cereals; and Cancer
Cancer is the second leading cause of morbidity and mortality and is a major public health problem in the modern time,. According to the WHO Cancer report 2023, there were an estimated 20 million new cancer cases and 10 million deaths worldwide in the last few years [3]. WHO have predicated, over the next two decades, the cancer burden will increase by around 60%, putting additional straining on health-care systems, individuals, and communities. The global burden is expected to rise to around 30 million new cancer cases by 2040, with the greatest increases occurring in under developed and developed countries. Consumption of whole grain cereals has been found to be associated with a lower risk of multiple types of cancer in numerous studies, including colon, colectral, prostrate, and other types [16,18,192–202] (Table 6). In contrast, some research on refined grains has found a relationship between high refined grain consumption and a higher risk of cancer [203,204]. These findings complement health recommendations to replace refined grains with whole grains. In a randomized and double-blind trial control study conducted by [192] on quinoa has been found to reduce cholesterol, indicated that postmenopausal women who consumed 25 g of quinoa flakes daily, rather than maize flakes, had reduced total cholesterol and LDL but higher Glutathione levels [193]. Furthermore Vilcacundo et al [194] studied the anti-cancer activity of quinoa peptides, demonstrated that peptides released from quinoa seeds were shown to have the ability to inhibit tumor growth in a variety of colon cancer cell lines, including Caco-2, HT-29, and HTC-116, as well as the capacity to scavenge free radicals [194]. Additionally this study indicated that the anti-cancer effect were because of suppression of α-amylase, α-glucosidase, and dipeptidyl peptidase IV by inhibition of enzymes. Moreover, in a recent study by Guo et al.[205] indicated that quinoa peptides produced by in vitro digestion had ACE inhibitory activity, which has antihypertensive effects on rats. Peptides from raw and heat-treated amaranth seeds have also shown anticancer activity by apoptosis and cell growth suppression, using HT-29 colon cancer cells and breast cancer cell lines [206]. In another study House et al. [16] found that amaranth tricolor contains compounds that suppress tumor cell growth, also amaranth seed proteins contain peptides associated with the reduction risk of cancer. In this regard, [80] investigated the effect of amaranth on human breast cells, suggesting that amaranth may be an excellent source of bioactive peptides with antioxidant activity and intriguing anticancer properties. It was noted that buckwheat polysaccharides were found to reduce the amplification of human PC-3 prostate cancer cells rather than directly affecting their proliferation and increasing the secretion of anti-inflammatory biomarkers [194]. Additionally Hiroyuki Tomotake et al [195] investigated the physiologic features of high protein buckwheat flour by measuring its effects on blood cholesterol and body fat in rats and cholesterol gallstone formation in mice. This study discovered that buckwheat protein significant lowers the cholesterol level in liver, reducing the risk of colon cancer. In a human study of 1372 colorectal cancer cases by kyro et al., [196] discovered that high intake of whole grains phenolic acids were associated with a decreased incidence of colon cancer but not with total colorectal cancer, proximal colon cancer, or rectal cancer. . Furthermore, in epidemiological study of Buescher et al [197] demonstrated that, whole grain consumption is associated with decreased colon cancer risk. This study investigated the impact of whole and refined wheat diets on colon carcinogenesis and related risk factors in an animal model found that whole grains had a significant effect on precancerous lesions and aberrant crypt foci, while refined grains had no effect. To date, several in vivo investigations have shown that phytochemicals can suppress cancers of the gastrointestinal tract by modulating oxidative stress. For example, in a rat model of hepato-carcinogenesis, phytic acid was demonstrated to boost Glutathione S-t\zzransferases activity and reduce lipid peroxidation, and these benefits were coupled with a reduction in the emergence of hepato-carcinogenesis markers [198]. This shows that phytic acid may inhibit liver tumor development by reducing oxidative stress. In another study, phytic acid was shown to boost Glutathione S-transferases activity, resulting in a reduction in the frequency of colon tumors in a rat model [199]. Furthermore, supplementation with hydoxycinnamic acid, an active rice bran phenolic acid, protects colonic tissues from colon carcinogenesis produced by the pro-carcinogen in an animal model. This study found that hydoxycinnamic acid had significant chemopreventive efficacy against 1,2-dimethylhydrazine-induced colon cancer[200]. Reynoso-Camacho et al [201] investigate the preventive impact of corn tortillas against colon cancer development. The study found that eating tortillas, particularly those made from white and blue maize, significantly reduced the risk of adenocarcinoma by up to 77.5%. In an animal study of 36 male transgenic rats for 8 weeks by Long et al [201] found that purple corn reduce the incidence of adenocarcinoma in the lateral prostate and inhibit the progression of prostate cancer. The author discovered that purple corn inhibits the expression of Cyclin D1 and downregulates the activation of p38 MAPK and Erk1/2. Since purple corn color is a mixture, determining its active component should help in the understanding and usage of purple corn color for prostate cancer chemoprevention. The results suggested that cyanidin-3-glucoside and pelargonidin-3-glucoside are the active compounds in corn.
5.2.2. Potential Mechanism of Whole Grains and Cancer
Intake of whole grains has been shown to lower the risk of obesity and improve metabolic problems, as well as lower the risk of cancer [25,53]. Obesity, particularly central/visceral obesity, causes insulin resistance and prolonged compensatory hyperinsulinemia, which has been shown to induce mitogenic effects and lead to cancer risk by activating both the insulin receptor and the insulin-like growth factor binding protein 1 and 2 receptor (Figure 5). Which then increased estrogen and androgen levels have also been linked to the development of cancer? Second the stomach and small intestine digest whole grain protein into peptides and amino acids [20]. Amino acids enter the bloodstream and travel to the colon (large intestine) [21].The gut microbiota in the colon ferments amino acids, resulting in SCFAs and other metabolites [22]. Consuming whole grain protein may alter the composition of the gut microbiota, increasing the abundance of beneficial bacteria while decreasing the abundance of harmful bacteria, which inhibit cancer cell growth and proliferation; induce apoptosis, reduces inflammation and oxidative stress [20]. Third, whole grains contain a variety of phytochemicals, which have been linked to cancer prevention [207]. Fourth, whole grains provide a good source of dietary fibres, which dilute carcinogens and reduce their absorption in the intestinal epithelium by increasing feces production and shortening the intestine's transit time. Dietary fibre can be fermented in the colon to produce SCFAs such as butyrate. Butyrate is the preferred fuel for mucosal cells, and it can stimulate apoptosis and anti-tumor activity, slowing tumor growth. They also lower the pH of the intestine, which reduces the solubility of free bile acids and thus their carcinogenicity.
Furthermore, dietary fiber can remove nitrite from the stomach and reduce the levels of nitroso compounds under high acid conditions. Nitrates increase the risk of stomach cancer [26]. Fourth, whole grains have antioxidant and anti-inflammatory properties, which can improve blood sugar response and reduce insulin resistance, lowering the risk of cancer [208,209]. Whole grains are high in antioxidants and phenolic acids, which can help prevent oxidative damage [186]. Whole grain foods contain high levels of bioactive compounds with anti-carcinogenic properties [210]. The combined anti-carcinogenic effects of these bioactive compounds are likely to explain many of the findings from these studies.
In summary, several epidemiological studies have discovered an inverse relationship between whole grain consumption and various types of cancer; longitudinal studies are needed to investigate cancer development and progression. Whole grains are a complex food matrix containing a variety of bioactive compounds that function together to prevention of cancer. It is thus difficult to determine which constituent is responsible for protection; several studies suggestion the beneficial effect of whole grain dietary fibre together with phenolic acids may have beneficial effect in reduction of cancer. For example, some whole grains positive effects are primarily dependent on protein, dietary fibre, and phenolic acids, but the content varies by grain and is also found in other foods consumed by high whole users. Although refined grains have a low total dietary fiber content, refining removes proportionally more insoluble fiber than soluble fiber. Challenges appear in a diverse area, so it is critical to determine which of these components will provide the most protection. More research is needed to fully understand their effects, whether bran, germ, protein, dietary fibers, or specific phenolic acids linked to a lower risk of cancer. Furthermore, the link between the consumption of whole grains and other types of cancers, such as breast, pancreatic, oral, and pharyngeal cancer, is less studied, and the results are often conflicting. The exact mechanism and independent effect of whole-grain bran, protein, phenolic acids, and dietary fibres should be investigated.
Conclusion
Whole grains pseudo cereals and cereals are known as "the grains of a modern time" due to their high nutritional and bioactive components. In term of protein content, amaranth contain the highest content of protein and lowest in rice, followed by wheat, quinoa, buckwheat corn, indicating that pseudo cereals contain the highest content of protein as compare to cereals. Corn, quinoa, rice, buckwheat, wheat and amaranth, are in descending order of the 9 Essential amino acids (g/100 g protein); while buckwheat, rice, amaranth, corn, quinoa and wheat are in descending order of the 9 non-Essential amino acids (g/100 g protein) . In pseudo cereals, quinoa possesses the highest content of total dietary fibers and phenolic acids, while buckwheat has the lowest. In cereal grains corn contain the highest content of total dietary fibres and rice contain the lowest, while wheat contain the highest content of phenolic acids and rice contain the lowest. The ranks become variable in the remaining cereals. Additionally, we discovered that cereal grains contain greater amounts of hydroxycinnamic acid than pseudo cereal grains. These compounds are considered to be primarily responsible for the beneficial effects on human health. Several epidemiological studies support the consumption whole grains over refined grains. Since, whole grains improve blood lipids, body weight, inflammation, and glycemic control while lowering premature mortality, we have found a strong inverse relationship between whole grain consumption and decreased risk of T2D and cancer. Additionally, the modulatory effects of whole grains on cancer and T2D might be contributed via a number of mechanisms, such as the actions of phenolic acids, dietary fibers, and protein. Future studies should carry out randomized controlled trials to validate observational results and establish causality; in addition the current manuscript encourages to investigate the specific mechanisms by which whole grains exert their beneficial effects on health by examining the effects of different types of specific protein, dietary fibres and phenolic acids that might help to prevent or treat T2D and cancer.
Funding
We acknowledge the financial support from National Natural Science Foundation of China (32172259), the Key Research and Development Project of Henan Province (231111111800), the Program for the Top Young Talents of Henan Associate for Science and Technology (2021), and the Innovative Funds Plan of Henan University of Technology (2021ZKCJ03).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the data are already provided in the main manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in study design, the decision to publish and the preparation of the manuscript.
References
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; et al. Global, Regional and Country-Level Diabetes Prevalence Estimates for 2021 and Projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [CrossRef]
- World Health Organization report (WHO) diabetes reports predication by 2025, (Available online); https://www.who.int/health-topics/diabetes (accessed on 14 May 2024).
- World Health Organization report (WHO) cancer report 2023, predication of cancer by 2040, (Available online); https://www.paho.org/en/campaigns/world-cancer-day-2023-close-care-gap#:~:text=Globally%2C%20there%20were%20an%20estimated,10%20million%20deaths%20from%20cancer (accessed on 14 May 2024).
- World Cancer Research Fund; American Institute for Cancer Research. Diet, Nutrition, Physical Activity and Cancer: A Global Perspective. Available online: http://dietandcancerreport.org (accessed on 14 May 2024).
- U.S. Department of Health and Human Services; U.S. Department of Agriculture. 2015–2020 Dietary Guidelines for Americans 8th Edition. Available online: http://health.gov/dietaryguidelines/2015/guidelines/ (accessed on 14 May 2024).
- Sastre, M.; Cimbalo, A.; Mañes, J.; Manyes, L. Gut Microbiota and Nutrition: Strategies for the Prevention and Treatment of Type 2 Diabetes. J. Med. Food 2024, 27, 97-109. [CrossRef]
- Ghanbari-Gohari, F.; Mousavi, S.M.; Esmaillzadeh, A. Consumption of whole grains and risk of type 2 diabetes: A comprehensive systematic review and dose–response meta-analysis of prospective cohort studies. Food Sci. Nutr. 2022, 10, 1950-1960. [CrossRef]
- Watling, C.Z.; Wojt, A.; Florio, A.A.; Butera, G.; Albanes, D.; Weinstein, S.J.; McGlynn, K.A. Fiber and whole grain intakes in relation to liver cancer risk: An analysis in two prospective cohorts and systematic review and meta-analysis of prospective studies. Hepatology 2024, 10-1097.
- Song, X.; Feng, X.; Chen, S.; Dai, Y.; Huang, H.; Li, X.; Liu, L. Potential impact of time trend of whole grain intake on burden of major cancers in China. Prev. Med. 2023, 175, 107674. [CrossRef]
- Gaesser, G.A. Whole grains, refined grains, and cancer risk: A systematic review of meta-analyses of observational studies. Nutrients 2020, 12, 3756. [CrossRef]
- Aune, D.; Norat, T.; Romundstad, P.; Vatten, L.J. Whole grain and refined grain consumption and the risk of type 2 diabetes: A systematic review and dose–response meta-analysis of cohort studies. Eur. J. Epidemiol. 2013, 28, 845-858. [CrossRef]
- Gaesser, G.A. Refined grain intake and risk of type 2 diabetes. Mayo Clin. Proc. 2022, 97, 1428-1436. [CrossRef]
- Mahzari, M.; Mamun, A. Does Consumption of Refined Carbohydrates Predict the Incidence of Type 2 Diabetes Mellitus? A Systematic Review and Meta-Analysis. Rom. J. Diabetes Nutr. Metab. Dis. 2020, 27, 168-179.
- Ernest, D.K.; Lemus, H.; Hsu, F.C.; Pierce, J.P.; Wu, T. The Independent and Joint Associations of Whole Grain and Refined Grain with Total Mortality among Breast Cancer Survivors: A Prospective Cohort Study. Nutrients 2022, 14, 3333. [CrossRef]
- AACCI. Definition of Whole Grain. 1999. (http://www.aaccnet.org/definitions/wholegrain.asp) (accessed on 19May 2024).
- House, N. C.; Puthenparampil, D.; Malayil, D,; & Narayanankutty, A. Variation in the polyphenol composition, antioxidant, and anticancer activity among different Amaranthus species. South Afr. J. of Bot., 2020. 135, 408-412. [CrossRef]
- Rao, V., & Poonia, A. (2023). Protein characteristics, amino acid profile, health benefits and methods of extraction and isolation of proteins from some pseudocereals—a review. Food Production, Processing and Nutrition, 5(1), 37. [CrossRef]
- Joye, I.J. Dietary fibre from whole grains and their benefits on metabolic health. Nutrients 2020, 12, 3045. [CrossRef]
- J.; Khan, M.Z.; Ma, Y.; Meng, Y.; Mushtaq, A.; Shen, Q.; Xue, Y. Overview of the Composition of Whole Grains’ Phenolic Acids and Dietary Fibre and Their Effect on Chronic Non-Communicable Diseases. Int. J. Environ. Res. Public Health 2022, 19, 3042.
- Zhao, J.; Zhang, X.; Liu, H.; Brown, M.A.; Qiao, S. Dietary Protein and Gut Microbiota Composition and Function. Curr. Protein Pept. Sci. 2019, 20, 145–154. [CrossRef]
- Bröer, S.; Fairweather, S.J. Amino Acid Transport across the Mammalian Intestine. Compr. Physiol. 2018, 9, 343–373.
- Sánchez-Alcoholado, L., Ramos-Molina, B., Otero, A., Laborda-Illanes, A., Ordóñez, R., Medina, J. A., ... & Queipo-Ortuño, M. I. (2020). The role of the gut microbiome in colorectal cancer development and therapy response. Cancers, 12(6), 1406. [CrossRef]
- Baky, M. H., Salah, M., Ezzelarab, N., Shao, P., Elshahed, M. S., & Farag, M. A. (2024). Insoluble dietary fibers: structure, metabolism, interactions with human microbiome, and role in gut homeostasis. Critical reviews in food science and nutrition, 64(7), 1954-1968. (link unavailable).
- Hernández, G.; et al. The Short-Chain Fatty Acid Acetate in Body Weight Control and Insulin Sensitivity. Nutrients 2019, 11, 1943.
- Bach Knudsen, K.E. Microbial Degradation of Whole-Grain Complex Carbohydrates and Impact on Short-Chain Fatty Acids and Health. Adv. Nutr. 2015, 6, 206–213. [CrossRef]
- Idehen, E.; Tang, Y.; Sang, S. Bioactive Phytochemicals in Barley. J. Food Drug Anal. 2017, 25, 148–161. [CrossRef]
- Aune, D.; et al. Dietary Fibre, Whole Grains, and Risk of Colorectal Cancer: Systematic Review and Dose–Response Meta-Analysis of Prospective Studies. BMJ 2011, 343.
- Călinoiu, L.F.; Vodnar, D.C. Whole Grains and Phenolic Acids: A Review on Bioactivity, Functionality, Health Benefits and Bioavailability. Nutrients 2018, 10, 1615. [CrossRef]
- de Melo Ribeiro, P.V.; Andrade, P.A.; Hermsdorff, H.H.M.; Dos Santos, C.A.; Cotta, R.M.M.; Estanislau, J.D.A.S.G.; de Oliveira Campos, A.A.; Rosa, C.D.O.B. Dietary Non-Nutrients in the Prevention of Non-Communicable Diseases: Potentially Related Mechanisms. Nutrition 2019, 66, 22–28. [CrossRef]
- U.S. Department of Health and Human Services and U.S. Department of Agriculture (2022) 2020–2025 Dietary Guidelines for Americans. https://www.dietaryguidelines.gov. (accessed on 19May 2024).
- Public Health England,2016; https://www.gov.uk/government/publications/the-eatwell-guide. (accessed on 19May 2024).
- Health Canada,Canada’s Food Guide, 2020;; https://food-guide.canada.ca/en/healthy-eating-recommendations/limit-highly-processed-foods/choosing-healthy-menu-options/. (accessed on 19May 2024).
- Mir, N.A.; Riar, C.S.; Singh, S. Nutritional Constituents of Pseudo Cereals and Their Potential Use in Food Systems: A Review. Trends Food Sci. Technol. 2018, 75, 170–180. [CrossRef]
- De Bock, P.; Daelemans, L.; Selis, L.; Raes, K.; Vermeir, P.; Eeckhout, M.; Van Bockstaele, F. Comparison of the Chemical and Technological Characteristics of Wholemeal Flours Obtained from Amaranth (Amaranthus sp.), Quinoa (Chenopodium quinoa) and Buckwheat (Fagopyrum sp.) Seeds. Foods 2021, 10, 651.
- Alonso-Miravalles, L.; O'Mahony, J.A. Composition, Protein Profile and Rheological Properties of Pseudocereal-Based Protein-Rich Ingredients. Foods 2018, 7, 73. [CrossRef]
- Venskutonis, P.R.; Kraujalis, P. Nutritional Components of Amaranth Seeds and Vegetables: A Review on Composition, Properties and Uses. Compr. Rev. Food Sci. 2013, 12, 381–412. [CrossRef]
- Ando, H.; Chen, Y.C.; Tang, H.J.; Shimizu, M.; Watanabe, K.; Mitsunaga, T. Food Components in Fractions of Quinoa Seed. Food Sci. Technol. Res. 2002, 8, 80–84. [CrossRef]
- Marmouzi, I.; El Madani, N.; Charrouf, Z.; Cherrah, Y.; Faouzi, M.E.A. Proximate Analysis, Fatty Acids and Mineral Composition of Processed Moroccan Chenopodium quinoa Willd. and Antioxidant Properties According to the Polarity. Phytothérapie 2015, 13, 110–117.
- Pirzadah, T.B.; Malik, B. Pseudocereals as Super Foods of 21st Century: Recent Technological Interventions. J. Agric. Food Res. 2020, (2), 100052. [CrossRef]
- Qin, P.; Wang, Q.; Shan, F.; Hou, Z.; Ren, G. Nutritional Composition and Flavonoids Content of Flour from Different Buckwheat Cultivars. Int. J. Food Sci. Technol. 2010, 45, 951–958. [CrossRef]
- Siyuan, S.; Tong, L.; Liu, R. Corn Phytochemicals and Their Health Benefits. Food Sci. Human Wellness 2018, 7, 185–195. [CrossRef]
- Ramashia, S.E.; Anyasi, T.A.; Gwata, E.T.; Meddows-Taylor, S.; Jideani, A.I.O. Processing, Nutritional Composition and Health Benefits of Finger Millet in Sub-Saharan Africa. Food Sci. Technol. 2019, 39, 253–266. [CrossRef]
- Oliveira, L.C.; Rosell, C.M.; Steel, C.J. Effect of the Addition of Whole-Grain Wheat Flour and of Extrusion Process Parameters on Dietary Fibre Content, Starch Transformation and Mechanical Properties of a Ready-to-Eat Breakfast Cereal. Int. J. Food Sci. Technol. 2015, 50, 1504–1514.
- Huda, M.N.; Lu, S.; Jahan, T.; Ding, M.; Jha, R.; Zhang, K.; Zhou, M. Treasure from Garden: Bioactive Compounds of Buckwheat. Food Chem. 2021, 335, 127653. [CrossRef]
- Kulathunga, J.; Reuhs, B. L.; Zwinger, S.; & Simsek, S. Comparative study on kernel quality and chemical composition of ancient and modern wheat species: Einkorn, emmer, spelt and hard red spring wheat. Foods, 2021. 10(4), 761. [CrossRef]
- Shewry, P.R.; Hawkesford, M.J.; Piironen, V.; Lampi, A.M.; Gebruers, K.; Boros, D.; Ward, J.L. Natural Variation in Grain Composition of Wheat and Related Cereals. J. Agric. Food Chem. 2013, 61, 8295–8303. [CrossRef]
- Hirawan, R.; Ser, W.Y.; Arntfield, S.D.; Beta, T. Antioxidant Properties of Commercial, Regular- and Whole-Wheat Spaghetti. Food Chem. 2010, 119, 258–264. [CrossRef]
- Kumar, A.; Metwal, M.; Kaur, S.; Gupta, A.K.; Puranik, S.; Singh, S.; Gupta, S.S.; Babu, B.K.; Sood, S.; Yadav, R. Nutraceutical Value of Finger Millet [Eleusine coracana (L.) Gaertn.] and Their Improvement Using Omics Approaches. Front. Plant Sci. 2016, \(7\), 1–14. [CrossRef]
- Rastogi, A.; Shukla, S. Amaranth: A New Millennium Crop of Nutraceutical Values. Crit. Rev. Food Sci. Nutr. 2013, 53, 109–125. [CrossRef]
- Joshi, D.C.; et al. From Zero to Hero: The Past, Present and Future of Grain Amaranth Breeding. Theor. Appl. Genet. 2018, 131, 1807–1823. [CrossRef]
- Joshi, D.C.; et al. Revisiting the Versatile Buckwheat: Reinvigorating Genetic Gains through Integrated Breeding and Genomics Approach. Planta 2019, 250, 783–801. [CrossRef]
- Manzoor, M., Hami, A., Pakhtoon, M. M., Batool, A., Zaffar, A., Sudan, J., ... & Zargar, S. M. (2023). Genetic variability of buckwheat (Fagopyrum spp.) genotypes for nutritional and nutraceutical traits. The Nucleus, 66(2), 1-9. [CrossRef]
- Seo, C.R.; Yi, B.; Oh, S.; et al. Aqueous Extracts of Hulled Barley Containing Coumaric Acid and Ferulic Acid Inhibit Adipogenesis in Vitro and Obesity in Vivo. J. Funct. Foods 2015, 12, 208–218. [CrossRef]
- Podeszwa, T.; Harasym, J.; Czerniecki, P.; Kopacz, M. Congress Wort Analysis from Commercial Buckwheat Malt Mixtures with RSM. Nauk. Inzynierskie i Technol. 2016, \(3\), 77–89.
- Aderibigbe, O.R.; Ezekiel, O.O.; Owolade, S.O.; Korese, J.K.; Sturm, B.; Hensel, O. Exploring the Potentials of Underutilized Grain Amaranth (Amaranthus spp.) along the Value Chain for Food and Nutrition Security: A Review. Crit. Rev. Food Sci. Nutr. 2020, 62, 656–669.
- Gebremariam, M.M.; Zarnkow, M.; Becker, T. Teff (Eragrostis tef) as a Raw Material for Malting, Brewing and Manufacturing of Gluten-Free Foods and Beverages: A Review. J. Food Sci. Technol. 2014, 51, 2881–2895. [CrossRef]
- de Sousa, T.; Ribeiro, M.; Sabença, C.; Igrejas, G. The 10,000-Year Success Story of Wheat! Foods 2021, 10, 2124.
- Kaplan Evlice, A.; Cetiner, B.; Pehlivan, A.; & Kara, R. Wheat Quality. In Advances in Wheat Breeding: Towards Climate Resilience and Nutrient Security. Springer Nature. 2024 (pp. 453-477).
- Zhygunov, D., Sots, S., Barkovska, Y., Liu, J., Wang, F., Liu, X., ... & Li, X. (2022). INFLUENCE OF GRAIN QUALITY INDICATORS ON THE FLOUR QUALITY INDICATORS AT THE LABORATORY MILLING. Grain Products and Mixed Fodder’s, 22(1).\. [CrossRef]
- Pereira, E.; et al. Chemical and Nutritional Characterization of Chenopodium quinoa Willd. (Quinoa) Grains: A Good Alternative to Nutritious Food. Food Chem. 2019, 280, 110–114. [CrossRef]
- Nowak, V.; et al. Assessment of the Nutritional Composition of Quinoa (Chenopodium quinoa Willd.). Food Chem. 2016, 193, 47–54. [CrossRef]
- Thanh-Tien, N.N.; et al. Nutritional Composition, Bioactive Compounds, and Diabetic Enzyme Inhibition Capacity of Three Varieties of Buckwheat in Japan. Cereal Chem. 2018, 95, 615–624. [CrossRef]
- Guo, H.; Hao, Y.; Yang, X.; Ren, G.; & Richel, A. Exploration on bioactive properties of quinoa protein hydrolysate and peptides: a review. Crit. Rev. in Food Sci. and Nutr. 2023. 63(16), 2896-2909. [CrossRef]
- Dakhili, S.; et al. Quinoa Protein: Composition, Structure and Functional Properties. Food Chem. 2019, 299, 125121.
- Fotschki, B.; Ju´skiewicz, J.; Jurgo ´nski, A.; Amarowicz, R.; Opyd, P.; Bez, J.; Muranyi, I.; Petersen, I.L.; Llopis, M.L. Protein-Rich Flours from Quinoa and Buckwheat Favourably Affect the Growth Parameters, Intestinal Microbial Activity and Plasma Lipid Profile of Rats. Nutrients 2020, 12, 2781.
- Abugoch, L.E.; Romero, N.; Tapia, C.A.; Silva, J.; Rivera, M. Study of Some Physicochemical and Functional Properties of Quinoa (Chenopodium quinoa Willd) Protein Isolates. J. Agric. Food Chem. 2008, 56, 4745–4750. [CrossRef]
- Vega-Galvez, A.; Miranda, M.; Vergara, J.; Uribe, E.; Puente, L.; Martinez, E.A. Nutrition Facts and Functional Potential of Quinoa (Chenopodium quinoa Willd.): An Ancient Andean Grain: A Review. J. Sci. Food Agric. 2010, 90, 2541–2547. [CrossRef]
- Okon, O. G. (2021). The nutritional applications of quinoa seeds. Biology and Biotechnology of Quinoa: Super Grain for Food Security, 35-49. [CrossRef]
- Brinegar, C.; Sine, B.; Nwokocha, L. High-Cysteine 2S Seed Storage Proteins from Quinoa (Chenopodium quinoa). J. Agric. Food Chem. 1996, 44, 1621–1623. [CrossRef]
- Singh, A., & Punia, D. (2020). Characterization and nutritive values of amaranth seeds. Current Journal of Applied Science and Technology, 39(3), 27-33. [CrossRef]
- Manassero, C.A.; Añón, M.C.; Speroni, F. Development of a High Protein Beverage Based on Amaranth. Plant Foods Hum. Nutr. 2020, 75, 599–607. [CrossRef]
- Arslan-Tontul, S.; Uslu, C.C.; Mutlu, C.; Erba¸s, M. Expected Glycemic Impact and Probiotic Stimulating Effects of Whole Grain Flours of Buckwheat, Quinoa, Amaranth and Chia. J. Food Sci. Technol. 2021, 59, 1460–1467.
- Olawoye, B.; Kadiri, O.; & Oluwajuyitan, T. D. Grain Amaranth: Processing, Health Benefits and Applications. In Cereals, 2021. (pp. 221-234).
- Schmidt, D.; Verruma-Bernardi, M.R.; Forti, V.A.; Borges, M.T.M.R. Quinoa and Amaranth as Functional Foods: A Review. Food Rev. Int. 2021, 1–20. [CrossRef]
- Malik, M.; Sindhu, R.; Dhull, S. B.; Bou-Mitri, C.; Singh, Y.; Panwar, S.; & Khatkar, B. S. Nutritional composition, functionality, and processing technologies for Amaranth. J. of Food Proc. and Pres., 2023, 1-24. [CrossRef]
- Pandya, A.; Thiele, B.; Köppchen, S.; Zurita-Silva, A.; Usadel, B.; & Fiorani, F. Characterization of Bioactive Phenolic Compounds in Seeds of Chilean Quinoa (Chenopodium quinoa Willd.) Germplasm. Agronomy, 2023.13(8), 2170. [CrossRef]
- Orona-Tamayo, D.; Paredes-Lopez, O. Amaranth Part 2—Sustainable Crop for the 21st Century: Food Properties and Nutraceuticals for Improving Human Health. In SPS; 2024; pp. 413–441.
- Liu, T.; Hou, G.G. Trends in Whole Grain Processing Technology and Product Development. In Whole Grains, 2019; pp. 257–279.
- Zhou, Y.; et al. Structural and Antioxidant Analysis of Tartary Buckwheat (Fagopyrum tartaricum Gaertn.) 13S Globulin. J. Sci. Food Agric. 2019, 1220-1229. [CrossRef]
- Taniya, M.S.; Reshma, M.V.; Shanimol, P.S.; Krishnan, G.; Priya, S. Bioactive Peptides from Amaranth Seed Protein Hydrolysates Induced Apoptosis and Antimigratory Effects in Breast Cancer Cells. Food Bioscience 2020, 35, 100588. [CrossRef]
- Fan, X.; Guo, H.; Teng, C.; Zhang, B.; Blecker, C.; Ren, G. Anti-Colon Cancer Activity of Novel Peptides Isolated from In Vitro Digestion of Quinoa Protein in Caco-2 Cells. Foods 2022, 11, 194. [CrossRef]
- Prabhakar, B.N., Suneetha, W.J., Devi, S.S., & Shreeja, K. (2021). Effect of germination on nutritional composition of common buckwheat (Fagopyrum esculentum Moench). International Research Journal of Pure and Applied Chemistry, 22(1), 1-7. [CrossRef]
- Fabian, C.; Ju, Y.H. A Review on Rice Bran Protein: Its Properties and Extraction Methods. Crit. Rev. Food Sci. Nutr. 2011, 51, 816–827. [CrossRef]
- Deng, Y.; Lim, J.; Lee, G.H.; Nguyen, T.T.H.; Xiao, Y.; Piao, M.; Kim, D. Brewing Rutin-Enriched Lager Beer with Buckwheat Malt as Adjuncts. J. Microbiol. Biotechnol. 2019, 29, 877–886. [CrossRef]
- Cadenas, R.; Caballero, I.; Nimubona, D.; Blanco, C.A. Brewing with Starchy Adjuncts: Its Influence on the Sensory and Nutritional Properties of Beer. Foods 2021, 10, 1726. [CrossRef]
- Chrungoo, N.K.; Chettry, U. Buckwheat: A Critical Approach towards Assessment of Its Potential as a Super Crop. Indian J. Genet. Plant Breed 2021, 81, 1–23. [CrossRef]
- Dabija, A.; Ciocan, M. E.; Chetrariu, A.; & Mîrzan, D. Comparative evaluation of the physico-chemical characteristics of buckwhweat malt and barley malt. Int. Multidiscip. Sci.2021, 21, 51-58.
- Comino, I.; Moreno, M.D.; Real, A.; Rodriguez-Herrera, A.; Barro, F.; Sousa, C. The Gluten-Free Diet: Testing Alternative Cereals Tolerated by Celiac Patients. Nutrients 2013, 5, 4250–4268. [CrossRef]
- Filho, A.M.M.; Pirozi, M.R.; Borges, J.T.D.S.; Pinheiro Sant’Ana, H.M.; Chaves, J.B.P.; Coimbra, J.S.D.R. Quinoa: Nutritional, Functional, and Antinutritional Aspects. Crit. Rev. Food Sci. Nutr. 2017, 57, 1618–1630. [CrossRef]
- Chettry, U.; Chrungoo, N.K. Beyond the Cereal Box: Breeding Buckwheat as a Strategic Crop for Human Nutrition. Plant. Foods Hum. Nutr. 2021, 76, 399–409. [CrossRef]
- Promising Das, S. Amaranths: The Crop of Great Prospect. In Amaranthus: A Promising Crop of Future; Springer: Singapore, 2016; pp. 13–48.
- Shobana, S.; Krishnaswamy, K.; Sudha, V.; Malleshi, N.G.; Anjana, R.M.; Palaniappan, L.; Mohan, V. Finger Millet (Ragi, Eleusine coracana L.): A Review of Its Nutritional Properties, Processing, and Plausible Health Benefits. Adv. Food Nutr. Res. 2013, 69, 1–39.
- Hou, Y.; He, W.; Hu, S.; Wu, G. Composition of Polyamines and Amino Acids in Plant-Source Foods for Human Consumption. Amino Acids 2019, 51, 1153–1165. [CrossRef]
- Kabthymer, R.H.; Karim, M.N.; Hodge, A.M.; de Courten, B. High Cereal Fibre but Not Total Fibre Is Associated with a Lower Risk of Type 2 Diabetes: Evidence from the Melbourne Collaborative Cohort Study. Diabetes Obes. Metab. 2023, 25(7), 1911–1921.
- Jovanovski, E.; Nguyen, M.; Kurahashi, Y.; Komishon, A.; Li, D.; Thanh, H.H.V.; Vuksan, V. Are All Fibres Created Equal with Respect to Lipid Lowering? Comparing the Effect of Viscous Dietary Fibre to Non-Viscous Fibre from Cereal Sources: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Br. J. Nutr. 2023, 129(7), 1180–1192. [CrossRef]
- Pastor-Cavada, E.; Drago, S.R.; González, R.J.; Juan, R.; Pastor, J.E.; Alaiz, M.; Vioque, J. Effects of the Addition of Wild Legumes (Lathyrus annuus and Lathyrus clymenum) on the Physical and Nutritional Properties of Extruded Products Based on Whole Corn and Brown Rice. Food Chem. 2011, 128(4), 961–967. [CrossRef]
- Gong, K.; Chen, L.; Li, X.; Sun, L.; Liu, K. Effects of Germination Combined with Extrusion on the Nutritional Composition, Functional Properties and Polyphenol Profile and Related in Vitro Hypoglycemic Effect of Whole Grain Corn. J. Cereal Sci. 2018, 83, 1–8. [CrossRef]
- Schulze, M.B.; Schulz, M.; Heidemann, C.; Schienkiewitz, A.; Hoffmann, K.; Boeing, H. Fiber and Magnesium Intake and Incidence of Type 2 Diabetes: A Prospective Study and Meta-Analysis. Arch. Intern. Med. 2007, 167(9), 956–965.
- EFSA Panel on Dietetic Products, Nutrition, and Allergies (NDA). Scientific Opinion on Dietary Reference Values for Carbohydrates and Dietary Fibre. EFSA Journal,2010; 8(3), 1462.
- Meghwal, M.; Kadeppagari, R.K. Dietary Fibers and Their Role as Functional Food for Human Health: Food Fibers and Human Health. In Exam. Devel., Reg., and Con. of Funct. Foods, 2017; pp. 29–44.
- Zeng, Z.; Liu, C.; Luo, S.; Chen, J.; Gong, E. The Profile and Bioaccessibility of Phenolic Compounds in Cereals Influenced by Improved Extrusion Cooking Treatment. PLoS One 2016, 11(8), e0161086. [CrossRef]
- Dhingra, D.; Michael, M.; Rajput, H.; Patil, R.T. Dietary Fibre in Foods: A Review. J. Food Sci. Technol. 2012, 49, 255–266. [CrossRef]
- El-Seedi, H.R.; El-Said, A.M.A.; Khalifa, S.A.M.; Göransson, U.; Bohlin, L.; Borg-Karlson, A.K.; Verpoorte, R. Biosynthesis, Natural Sources, Dietary Intake, Pharmacokinetic Properties, and Biological Activities of Hydroxycinnamic Acids. J. Agric. Food Chem. 2012, 60(44), 10877–10895. [CrossRef]
- Vitaglione, P.; Napolitano, A.; Fogliano, V. Cereal Dietary Fibre: A Natural Functional Ingredient to Deliver Phenolic Compounds into the Gut. Trends Food Sci. Technol. 2008, 19, 451–463. [CrossRef]
- Bunzel, M.; Ralph, J.; Marita, J.M.; Hatfield, R.D.; Steinhart, H. Diferulates as Structural Components in Soluble and Insoluble Cereal Dietary Fibre. J. Sci. Food Agric. 2001, 81, 653–660. [CrossRef]
- Malleshi, N.G.; Agarwal, A.; Tiwari, A.; Sood, S. Nutritional Quality and Health Benefits. Millets and pseudocereal, 2021; pp. 159–168.
- Srichuwong, S.; Curti, D.; Austin, S.; King, R.; Lamothe, L.; Gloria-Hernandez, H. Physicochemical Properties and Starch Digestibility of Whole Grain Sorghums, Millet, Quinoa and Amaranth Flours, as Affected by Starch and Non-Starch Constituents. Food Chem. 2017, 233, 1–10. [CrossRef]
- Pulvento, C.; Riccardi, M.; Lavini, A.; Iafelice, G.; Marconi, E.; D’Andria, R. Yield and Quality Characteristics of Quinoa Grown in Open Field Under Different Saline and Non-Saline Irrigation Regimes. J. Agron. Crop Sci. 2012, 198, 254–263. [CrossRef]
- Alvarez-Jubete, L.; Arendt, E.K.; Gallagher, E. Nutritive Value and Chemical Composition of Pseudocereals as Gluten-Free Ingredients. Int. J. Food Sci. Nutr. 2009, 60, 240–257. [CrossRef]
- Robin, F.; Théoduloz, C.; Srichuwong, S. Properties of Extruded Whole Grain Cereals and Pseudocereals Flours. Int. J. Food Sci. Technol. 2015, 50, 2152–2159. [CrossRef]
- Steadman, K.J.; Burgoon, M.S.; Lewis, B.A.; Edwardson, S.E.; Obendorf, R.L. Buckwheat Seed Milling Fractions: Description, Macronutrient Composition and Dietary Fibre. J. Cereal Sci. 2001, 33, 271–278. [CrossRef]
- De Santis, M.A.; Kosik, O.; Passmore, D.; Flagella, Z.; Shewry, P.R.; Lovegrove, A. Comparison of the Dietary Fibre Composition of Old and Modern Durum Wheat (Triticum turgidum spp. durum) Genotypes. Food Chem. 2018, 244, 304–310. 1. [CrossRef]
- Rainakari, A.-I.; Rita, H.; Putkonen, T.; Pastell, H. New Dietary Fibre Content Results for Cereals in the Nordic Countries Using AOAC 2011.25 Method. J. Food Compos. Anal. 2016, 51, 1–8. [CrossRef]
- Amalraj, A.; Pius, A. Influence of Oxalate, Phytate, Tannin, Dietary Fiber, and Cooking on Calcium Bioavailability of Commonly Consumed Cereals and Millets in India. Cereal Chem. 2015, 92, 389–394. [CrossRef]
- Prasad, V.S.S.; Hymavathi, A.; Babu, V.R.; Longvah, T. Nutritional Composition in Relation to Glycemic Potential of Popular Indian Rice Varieties. Food Chem. 2018, 238, 29–34. [CrossRef]
- Prasanthi, P.S.; Naveena, N.; Vishnuvardhana Rao, M.; Bhaskarachary, K. Compositional Variability of Nutrients and Phytochemicals in Corn after Processing. J. Food Sci. Technol. 2017, 54, 1080–1090. [CrossRef]
- Gartaula, G.; Dhital, S.; Netzel, G.; Flanagan, B.M.; Yakubov, G.E.; Beahan, C.T.; Collins, H.M.; Burton, R.A.; Bacic, A.; Gidley, M.J. Quantitative Structural Organisation Model for Wheat Endosperm Cell Walls: Cellulose as an Important Constituent. Carbohydr. Polym. 2018, 196, 199–208. [CrossRef]
- Zhu, F. Dietary fiber polysaccharides of amaranth, buckwheat and quinoa grains: A review of chemical structure, biological functions and food uses. Carbohydrate Polymers,2020. 248, 116819. [CrossRef]
- Mudgil, P.; Omar, L.S.; Kamal, H.; Kilari, B.P.; Maqsood, S. Multi-Functional Bioactive Properties of Intact and Enzymatically Hydrolysed Quinoa and Amaranth Proteins. LWT 2019, 110, 207–213. [CrossRef]
- Zhang, G.; Xu, Z.; Gao, Y.; Huang, X.; Zou, Y.; Yang, T. Effects of Germination on the Nutritional Properties, Phenolic Profiles, and Antioxidant Activities of Buckwheat. J. Food Sci. 2015, 80, H1111–9. [CrossRef]
- Adom, K.K.; Liu, R.H. Antioxidant Activity of Grains. J. Agric. Food Chem. 2002, 50, 6182–6187. [CrossRef]
- Mattila, P.; Pihlava, J.-M.; Hellstrom, J. Contents of Phenolic Acids, Alkyl- and Alkenylresorcinols, and Avenanthramides in Commercial Grain Products. J. Agric. Food Chem. 2005, 53, 8290–8295. [CrossRef]
- Li, L.; Shewry, P.R.; Ward, J.L. Phenolic Acids in Wheat Varieties in the HEALTHGRAIN Diversity Screen. J. Agric. Food Chem. 2008, 56, 9732–9739. [CrossRef]
- Clifford, M.N.; van der Hooft, J.J.; Crozier, A. Human Studies on the Absorption, Distribution, Metabolism, and Excretion of Tea Polyphenols. Am. J. Clin. Nutr. 2013, 98 (6 Suppl): 1619S–1630S. [CrossRef]
- Sánchez-Patán, F.; Barroso, E.; van de Wiele, T.; et al. Comparative in Vitro Fermentations of Cranberry and Grape Seed Polyphenols with Colonic Microbiota. Food Chem. 2015, 183, 273–782. [CrossRef]
- Kandil, A.; Li, J.; Vasanthan, T.; Bressler, D.C. Phenolic Acids in Some Cereal Grains and Their Inhibitory Effect on Starch Liquefaction and Saccharification. J. Agric. Food Chem. 2012, 60, 8444–8449. [CrossRef]
- Žilic, S.; Serpen, A.; Akıllıoğlu, G.; Gökmeng, V.; Vancetovic, J. Phenolic Compounds, Carotenoids, Anthocyanins, and Antioxidant Capacity of Colored Maize (Zea mays L.) Kernels. J. Agr. Food Chem. 2012, 60, 1224–1231.
- Bang, J.H.; Jo, I.H.; Sebastin, R.; Jeong, W.T.; Oh, S.; Heo, T.Y.; et al. Comparative Analysis of Polyphenolic Compounds in Different Amaranthus Species: Influence of Genotypes and Harvesting Year. Antioxidants 2024, 13, 501. [CrossRef]
- Irakli, M.N.; Samanidou, V.F.; Biliaderis, C.G.; Papadoyannis, I.N. Development and Validation of an HPLC-Method for Determination of Free and Bound Phenolic Acids in Cereals after Solid-Phase Extraction. Food Chem. 2012, 134, 1624–1632. [CrossRef]
- Hithamani, G.; Srinivasan, K. Bioaccessibility of Polyphenols from Wheat (Triticum aestivum), Sorghum (Sorghum bicolor), Green Gram (Vigna radiata), and Chickpea (Cicer arietinum) as Influenced by Domestic Food Processing. J. Agric. Food Chem. 2014, 62, 11170–11179.
- Gómez-Caravaca, A.M.; Iafelice, G.; Verardo, V.; Marconi, E.; Caboni, M.F. Influence of Pearling Process on Phenolic and Saponin Content in Quinoa (Chenopodium quinoa Willd). Food Chem. 2014, 157, 174–178. [CrossRef]
- Repo-Carrasco-Valencia, R.; Hellström, J.K.; Pihlava, J.M.; Mattila, P.H. Flavonoids and Other Phenolic Compounds in Andean Indigenous Grains: Quinoa (Chenopodium quinoa), Kañiwa (Chenopodium pallidicaule), and Kiwicha (Amaranthus caudatus). Food Chem. 2010, 120, 128–133. [CrossRef]
- Sedej, I.; Sakač, M.; Mandić, A.; Mišan, A.; Tumbas, V.; Čanadanović-Brunet, J. Buckwheat (Fagopyrum esculentum Moench) Grain and Fractions: Antioxidant Compounds and Activities. J. Food Sci. 2012, 77, C954–C959. [CrossRef]
- Barbhai, M; D.;& Hymavathi, T. V. Nutrient, phytonutrient and antioxidant potential of selected underutilized nutri-cereal brans. J. Food Measur. and Charac. 2022, 16(3), 1952-1966. [CrossRef]
- Patel, S. Cereal Bran: The Next Super Food with Significant Antioxidant and Anticancer Potential. Mediterr. J. Nutr. Metab. 2012, 5, 91–104.
- Feregrino-Pérez, A.A.; Mercado-Luna, A.; Murillo-Cárdenas, C.A.; González-Santos, R.; Chávez-Servín, J.L.; Vargas-Madriz, A.F.; Luna-Sánchez, E. Polyphenolic Compounds and Antioxidant Capacity in Native Maize of the Sierra Gorda of Querétaro. Agronomy 2024, 14, 142. [CrossRef]
- Imam, M.U.; Musa, S.N.A.; Azmi, N.H.; Ismail, M. Effects of White Rice, Brown Rice and Germinated Brown Rice on Antioxidant Status of Type 2 Diabetic Rats. Int. J. Mol. Sci. 2012, 13, 12952–12969. [CrossRef]
- Gong, E.S.; Luo, S.; Li, T.; Liu, C.; Zhang, G.; Chen, J.; Zeng, Z.; Liu, R.H. Phytochemical Profiles and Antioxidant Activity of Processed Brown Rice Products. Food Chem. 2017, 232, 67–78. [CrossRef]
- Ranjkesh, N.; Daliri, M. S.; Mazloum, P.; Mousavi, A.; & Rameeh, V. Evaluation of physicochemical characteristics and antioxidant properties of elite rice (Oryza sativa L.). Cer. Res. Communi. 2021, 49, 485-491. [CrossRef]
- Çelik, E.E.; Gökmen, V. Effects of Fermentation and Heat Treatments on Bound-Ferulic Acid Content and Total Antioxidant Capacity of Bread Crust-Like Systems Made of Different Whole Grain Flours. J. Cereal Sci. 2020, 102978.
- Halvorsen, B.L.; Holte, K.; Myhrstad, M.C.; Barikmo, I.; Hvattum, E.; Remberg, S.F.; Wold, A.B.; Haffner, K.; Baugerød, H.; Andersen, L.F.; Moskaug, O. A Systematic Screening of Total Antioxidants in Dietary Plants. J. Nutr. 2002, 132, 461–471. [CrossRef]
- Serpen, A.; Gökmen, V.; Pellegrini, N.; Fogliano, V. Direct Measurement of the Total Antioxidant Capacity of Cereal Products. J. Cereal Sci. 2008, 48, 816–820. [CrossRef]
- Tufan, A.N.; Çelik, S.E.; Özyürek, M.; Güçlü, K.; Apak, R. Direct Measurement of Total Antioxidant Capacity of Cereals: QUENCHER-CUPRAC Method. Talanta 2013, 108, 136–142. [CrossRef]
- Adebo, O. A.; & Gabriela Medina-Meza, I. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules, 2020. 25(4), 927. [CrossRef]
- Kolniak-Ostek, J.; Kita, A.; Pęksa, A.; Wawrzyniak, A.; Hamułka, J.; Jeznach, M.; Danilčenko, H.; Jariene, E. Analysis of the Content of Bioactive Compounds in Selected Flours and Enriched Extruded Corn Products. J. Food Compos. Anal. 2017, 64, 147–155. [CrossRef]
- Peñarrieta, J.M.; Alvarado, J.A.; Åkesson, B.; Bergenståhl, B. Total Antioxidant Capacity and Content of Flavonoids and Other Phenolic Compounds in Canihua (Chenopodium pallidicaule): An Andean Pseudocereal. Mol. Nutr. Food Res. 2008, 52, 708–717. [CrossRef]
- Gorinstein, S.; Lojek, A.; Číž, M.; Pawelzik, E.; Delgado-Licon, E.; Medina, O.J.; Moreno, M.; Arnao Salas, I.; Goshev, I. Comparison of Composition and Antioxidant Capacity of Some Cereals and Pseudocereals. Int. J. Food Sci. Technol. 2008, 43, 629–637. [CrossRef]
- Loarca-Piña, G.; Neri, M.; Figueroa, J.D.D.; Castaño-Tostado, E.; Ramos-Gómez, M.; Reynoso, R.; Mendoza, S. Chemical Characterization, Antioxidant and Antimutagenic Evaluations of Pigmented Corn. J. Food Sci. Technol. 2019, 56, 3177–3184. [CrossRef]
- Yang, Z.; Zhai, W. Identification and Antioxidant Activity of Anthocyanins Extracted from the Seed and Cob of Purple Corn (Zea mays L.). Innov. Food Sci. Emerg. Technol. 2010, 11, 169–176. [CrossRef]
- Kostić, A.Ž.; Milinčić, D.D.; Špirović Trifunović, B.; Nedić, N.; Gašić, U.M.; Tešić, Ž.L.; et al. Monofloral Corn Poppy Bee-Collected Pollen—A Detailed Insight into Its Phytochemical Composition and Antioxidant Properties. Antioxidants 2023, 12, 1424. [CrossRef]
- Chlopicka, J.; Pasko, P.; Gorinstein, S.; Jedryas, A.; Zagrodzki, P. Total Phenolic and Total Flavonoid Content, Antioxidant Activity and Sensory Evaluation of Pseudocereal Breads. LWT-Food Sci. Technol. 2012, 46, 548–555. [CrossRef]
- Martínez-Villaluenga, C.; Peñas, E.; Hernández-Ledesma, B. Pseudocereal Grains: Nutritional Value, Health Benefits and Current Applications for the Development of Gluten-Free Foods. Food Chem. Toxicol. 2020, 137, 111178. [CrossRef]
- Li, L.; Lietz, G.; Seal, C.J. Phenolic, Apparent Antioxidant and Nutritional Composition of Quinoa (Chenopodium quinoa Willd.) Seeds. Int. J. Food Sci. Technol. 2021, 56, 3245–3254. [CrossRef]
- Gallardo, C.; Jimenez, L.; Garcia-Conesa, M.T. Hydroxycinnamic Acid Composition and in Vitro Antioxidant Activity of Selected Grain Fractions. Food Chem. 2006, 99, 455–463. [CrossRef]
- Đorđević, T.M.; Šiler-Marinković, S.S.; Dimitrijević-Branković, S.I. Effect of Fermentation on Antioxidant Properties of Some Cereals and Pseudo Cereals. Food Chem. 2010, 119, 957–963. [CrossRef]
- Inglett, G.E.; Chen, D.; Berhow, M.; Lee, S. Antioxidant Activity of Commercial Buckwheat Flours and Their Free and Bound Phenolic Compositions. Food Chem. 2011, 125, 923–929. [CrossRef]
- Pruett, A.; Aramouni, F. M.; Bean, S. R.; & Haub, M. D.Effect of Flour Particle Size on the Glycemic Index of Muffins Made from Whole Sorghum, Whole Corn, Brown Rice, Whole Wheat, or Refined Wheat Flours. Foods, 2023 12(23), 4188. [CrossRef]
- FAO. Food and Agriculture Organization Available online http://www.fao.org/alc/file/media/pubs/2011/cultivo_quinua_en. pdf (accessed on 19May 2024).
- Bommer, C.; Heesemann, E.; Sagalova, V.; Manne-Goehler, J.; Atun, R.; Bärnighausen, T.; Vollmer, S. The Global Economic Burden of Diabetes in Adults Aged 20–79 Years: A Cost-of-Illness Study. Lancet Diabetes Endocrinol. 2017, 5, 423–430. [CrossRef]
- Patel, B.; Unadkat, S.; Patel, H.; Rathod, M. Dietary Practices among Type 2 Diabetes Patients Visiting a Non-Communicable Disease (NCD) Clinic in a District of Western India: A Cross-Sectional Study. Cureus 2024, 16, 1. [CrossRef]
- Wang, Y.; Duan, Y.; Zhu, L.; Fang, Z.; He, L.; Ai, D.; Jin, Y. Whole Grain and Cereal Fiber Intake and the Risk of Type 2 Diabetes: A Meta-Analysis. Int. J. Mol. Epidemiol. Genet. 2019, 10, 38–46.
- Pasini, F.; Marzocchi, S.; Caboni, M. Whole Grain: An Open Issue. In Global Summit on Food Sci. and Technol.2024; pp. 1–2.
- Xiao, Y.; Ke, Y.; Wu, S.; Huang, S.; Li, S.; Lv, Z.; Yeoh, E.-K.; Lao, X.; Wong, S.; Kim, J.H.; Colditz, G.A.; Tamimi, R.M.; Su, X. Association between Whole Grain Intake and Breast Cancer Risk: A Systematic Review and Meta-Analysis of Observational Studies. Nutr. J. 2018, 17, 87. [CrossRef]
- Pasko, P.; Barton, H.; Zagrodzki, P.; Izewska, A.; Krosniak, M.; Gawlik, M.; Gorinstein, S. Effect of Diet Supplemented with Quinoa Seeds on Oxidative Status in Plasma and Selected Tissues of High Fructose-Fed Rats. Plant Foods Hum. Nutr. 2010, 65, 146–151. [CrossRef]
- Khan, R. A. R., Ali, M. U., Chand, S., Razzaq, M., Latif, M. U., Amjid, M., & Afzal, I. (2023). Anti-diabetic properties of Chenopodium quinoa on human health; an update and future prospects. One Health Triad, 2(168-175). [CrossRef]
- Kasozi, K.I.; Namubiru, S.; Safiriyu, A.A.; Ninsiima, H.I.; Nakimbugwe, D.; Namayanja, M.; Valladares, M.B. Grain amaranth is associated with improved hepatic and renal calcium metabolism in type 2 diabetes mellitus of male wistar rats. Evid.-Based Complement. Altern. Med. 2018, 2018, 4098942. [CrossRef]
- Qiu, J.; et al. Dietary Tartary Buckwheat Intake Attenuates Insulin Resistance and Improves Lipid Profiles in Patients with Type 2 Diabetes: A Randomized Controlled Trial. Nutr. Res. 2016, 36, 1392–1401. [CrossRef]
- Wu, W.; Li, Z.; Qin, F.; Qiu, J. Anti-Diabetic Effects of the Soluble Dietary Fiber from Tartary Buckwheat Bran in Diabetic Mice and Their Potential Mechanisms. Food Nutr. Res. 2021, 65. [CrossRef]
- Kyrø, C.; Tjønneland, A.; Overvad, K.; Olsen, A.; Landberg, R. Higher Whole-Grain Intake Is Associated with Lower Risk of Type 2 Diabetes among Middle-Aged Men and Women: The Danish Diet, Cancer, and Health Cohort. J. Nutr. 2018, 148, 1434–1444. [CrossRef]
- Liu, Y.; Qiu, J.; Yue, Y.; Li, K.; Ren, G. Dietary Black-Grained Wheat Intake Improves Glycemic Control and Inflammatory Profile in Patients with Type 2 Diabetes: A Randomized Controlled Trial. Ther. Clin. Risk Manag. 2018, 14, 247–256. [CrossRef]
- Hu, Y.; Ding, M.; Sampson, L.; Willett, W.C.; Manson, J.E.; Wang, M.; et al. Intake of Whole Grain Foods and Risk of Type 2 Diabetes: Results from Three Prospective Cohort Studies. BMJ 2020, 370. [CrossRef]
- Huang, B.; Wang, Z.; Park, J.H.; Ryu, O.H.; Choi, M.K.; Lee, J.Y.; et al. Anti-Diabetic Effect of Purple Corn Extract on C57BL/KsJ db/db Mice. Nutr. Res. Pract. 2015, 9, 22. [CrossRef]
- Sun, S.L.; Zhang, G.W.; Mu, H.Y.; Zhang, H.; Chen, Y.Q. The Mixture of Corn and Wheat Peptide Prevent Diabetes in NOD Mice. J. Diabetes Res. 2019. 163-170. [CrossRef]
- Hu, E.A.; Pan, A.; Malik, V.; Sun, Q. White Rice Consumption and Risk of Type 2 Diabetes: Meta-Analysis and Systematic Review. BMJ 2012, 344.
- Seah, J.Y.H.; Koh, W.P.; Yuan, J.M.; van Dam, R.M. Rice Intake and Risk of Type 2 Diabetes: The Singapore Chinese Health Study. Eur. J. Nutr. 2019, 58, 3349–3360. [CrossRef]
- Nanri, A.; Mizoue, T.; Noda, M.; Takahashi, Y.; Matsushita, Y.; Poudel-Tandukar, K.; et al. rice Intake and Type 2 Diabetes in Japanese Men and Women: The Japan Public Health Center–Based Prospective Study. Am. J. Clin. Nutr. 2011, 94, 884–891.
- Bhavadharini, B.; Mohan, V.; Dehghan, M.; Rangarajan, S.; Swaminathan, S.; Rosengren, A.; et al. White Rice Intake and Incident Diabetes: A Study of 132,373 Participants in 21 Countries. Diabetes Care 2020, 43, 2643–2650. [CrossRef]
- Schadow, A. M.; Revheim, I.; Spielau, U.; Dierkes, J.; Schwingshackl, L.; Frank, J& Rosendahl-Riise, H. The effect of regular consumption of reformulated breads on glycemic control: A systematic review and meta-analysis of randomized clinical trials. Advances in Nutrition, 2023 14(1), 30-43. [CrossRef]
- Milesi, G.; Rangan, A.; Grafenauer, S. Whole Grain Consumption and Inflammatory Markers: A Systematic Literature Review of Randomized Control Trials. Nutrients 2022, 14, 374. [CrossRef]
- Khan, J.; Gul, P.; Rashid, M.T.; Li, Q.; Liu, K. Composition of Whole Grain Dietary Fiber and Phenolics and Their Impact on Markers of Inflammation. Nutrients 2024, 16, 1047. [CrossRef]
- Kanata, M.C.; Yanni, A.E.; Koliaki, C.; Pateras, I.; Anastasiou, I.A.; Kokkinos, A.; Karathanos, V.T. Effects of Wheat Biscuits Enriched with Plant Proteins Incorporated into an Energy-Restricted Dietary Plan on Postprandial Metabolic Responses of Women with Overweight/Obesity. Nutrients 2024, 16, 1229. [CrossRef]
- Akhlaghi, M. The Role of Dietary Fibers in Regulating Appetite, an Overview of Mechanisms and Weight Consequences. Crit. Rev. Food Sci. Nutr. 2024, 64, 3139–3150. [CrossRef]
- Weickert, M.O.; Pfeiffer, A.F. Impact of Dietary Fiber Consumption on Insulin Resistance and the Prevention of Type 2 Diabetes. J. Nutr. 2018, 148, 7–12. [CrossRef]
- Lattimer, J.M.; Haub, M.D. Effects of Dietary Fiber and Its Components on Metabolic Health. Nutrients 2010, 2, 1266–1289.
- Choi, S.H.; Kim, B.T.; Shin, J.; Kim, K.N. Combined Effect of Serum Alanine Aminotransferase and Gamma-Glutamyltransferase on Incidence of Diabetes Mellitus: A Longitudinal Study. Medicine 2020, 99, 11.
- Fardet, A. New Hypotheses for the Health-Protective Mechanisms of Whole-Grain Cereals: What Is beyond Fibre? Nutr. Res. Rev. 2010, 23, 65–134.
- De Angelis, M.; Montemurno, E.; Vannini, L.; Cosola, C.; Cavallo, N.; Gozzi, G.; Maranzano, V.; Di Cagno, R.; Gobbetti, M.; Gesualdo, L. Effect of Whole-Grain Barley on the Human Fecal Microbiota and Metabolome. Appl. Environ. Microbiol. 2015, 81, 7945–7956. [CrossRef]
- Adom, K.K.; Sorrells, M.E.; Liu, R.H. Phytochemicals and Antioxidant Activity of Milled Fractions of Different Wheat Varieties. J. Agric. Food Chem. 2005, 53, 2297–2306. [CrossRef]
- Belobrajdic, D.P.; Bird, A.R. The Potential Role of Phytochemicals in Wholegrain Cereals for the Prevention of Type-2 Diabetes. Nutr. J. 2013, 12, 62. [CrossRef]
- Chen, S.; Zhao, X.; Ran, L.; Wan, J.; Wang, X.; Qin, J.; Shu, F.; Gao, Y.; Yuan, L.; Zhang, Q.; et al. Resveratrol Improves Insulin Resistance, Glucose and Lipid Metabolism in Patients with Non-Alcoholic Fatty Liver Disease: A Randomized Controlled Trial. Dig. Liver Dis. 2015, 47, 226–232. [CrossRef]
- Bozzetto, L.; Annuzzi, G.; Pacini, G.; Costabile, G.; Vetrani, V.; Vitale, M.; Griffo, G.; Giacco, A.; De Natale, C.; Cocozza, S.; et al. Polyphenol-Rich Diets Improve Glucose Metabolism in People at High Cardiometabolic Risk: A Controlled Randomized Intervention Trial. Diabetologia 2015, 58, 1551–1560. [CrossRef]
- De Carvalho, F.G.; Ovídio, P.P.; Padovan, G.J.; Jordão Junior, A.A.; Marchini, J.S.; Navarro, A.M. Metabolic Parameters of Postmenopausal Women after Quinoa or Corn Flakes Intake–A Prospective and Double-Blind Study. Int. J. Food Sci. Nutr. 2014, 65, 380–385.
- Vilcacundo, R.; Miralles, B.; Carrillo, W.; Hernández-Ledesma, B. In Vitro Chemopreventive Properties of Peptides Released from Quinoa (Chenopodium quinoa Willd.) Protein under Simulated Gastrointestinal Digestion. Food Res. Int. 2018, 105, 403–411.
- Lin, H.C.; Lin, J.Y. Immune Cell–Conditioned Media Suppress Prostate Cancer PC-3 Cell Growth Correlating with Decreased Proinflammatory/Anti-Inflammatory Cytokine Ratios in the Media Using 5 Selected Crude Polysaccharides. Integr. Cancer Ther. 2016, 15, NP13–NP25. [CrossRef]
- Tomotake, H.; Yamamoto, N.; Yanaka, N.; Ohinata, H.; Yamazaki, R.; Kayashita, J.; Kato, N. High Protein Buckwheat Flour Suppresses Hypercholesterolemia in Rats and Gallstone Formation in Mice by Hypercholesterolemic Diet and Body Fat in Rats because of Its Low Protein Digestibility. Nutrition 2006, 22, 166–173. [CrossRef]
- Kyrø, C.; Olsen, A.; Landberg, R.; Skeie, G.; Loft, S.; Åman, P.; Bueno-de-Mesquita, H.A. Plasma Alkylresorcinols, Biomarkers of Whole-Grain Wheat and Rye Intake, and Incidence of Colorectal Cancer. J. Natl. Cancer Inst. 2014, 106, djt352. [CrossRef]
- Buescher, M.I.; Gallaher, D.D. Wheat Color (Class), Not Refining, Influences Colon Cancer Risk in Rats. Nutr. Cancer 2014, 66, 849–856. [CrossRef]
- Dilworth, L., Stennett, D., & Omoruyi, F. (2023). Cellular and molecular activities of IP6 in disease prevention and therapy. Biomolecules, 13(6), 972; (link unavailable). [CrossRef]
- Khatiwada, J.; Verghese, M.; Davis, S.; Williams, L.L. Green Tea, Phytic Acid, and Inositol in Combination Reduced the Incidence of Azoxymethane-Induced Colon Tumors in Fisher 344 Male Rats. J. Med. Food 2011, 14, 1313–1320. [CrossRef]
- Sivagami, G.; Karthikkumar, V.; Balasubramanian, T.; Nalini, N. The Modulatory Influence of p-Methoxycinnamic Acid, an Active Rice Bran Phenolic Acid, against 1,2-Dimethylhydrazine-Induced Lipid Peroxidation, Antioxidant Status and Aberrant Crypt Foci in Rat Colon Carcinogenesis. Chem. Biol. Interact. 2012, 196, 11–22. [CrossRef]
- Reynoso Guerrero-Villanueva, G.; de Dios Figueroa, J.; Gallegos-Corona, M.A.; Mendoza, S.; Loarca-Pina, G.; Ramos-Gomez, M. Anticarcinogenic Effect of Corn Tortilla against 1,2-Dimethylhydrazine (DMH)-Induced Colon Carcinogenesis in Sprague–Dawley Rats. Plant Foods Hum. Nutr. 2015, 70, 146–152.
- Long, N.; Suzuki, S.; Sato, S.; Naiki-Ito, A.; Sakatani, K.; Shirai, T.; Takahashi, S. Purple Corn Color Inhibition of Prostate Carcinogenesis by Targeting Cell Growth Pathways. Cancer Sci. 2013, 104, 298–303. [CrossRef]
- Xu, Y.; Yang, J.; Du, L.; Li, K.; Zhou, Y. Association of Whole Grain, Refined Grain, and Cereal Consumption with Gastric Cancer Risk: A Meta-Analysis of Observational Studies. Foods; 2019, 7, 256-265.
- Mousavi, S.M.; Rigi, S.; Shayanfar, M.; Mohammad-Shirazi, M.; Sharifi, G.; Esmaillzadeh, A. Refined Grains Consumption Is Associated with a Greater Odds of Glioma. Nutrients; 2022, 25, 432-440. [CrossRef]
- Guo, H.; Hao, Y.; Richel, A.; Everaert, N.; Chen, Y.; Liu, M.; Yang, X.; Ren, G. Antihypertensive Effect of Quinoa Protein under Simulated Gastrointestinal Digestion and Peptide Characterization. J. Sci. Food Agric. 2020, 100, 5569–5576. [CrossRef]
- Fillería, S.G.; Tironi, V. Intracellular Antioxidant Activity and Intestinal Absorption of Amaranth Peptides Released Using Simulated Gastrointestinal Digestion with Caco-2 TC7 Cells. Food Biosci. 2021, 41, 101086. [CrossRef]
- Oh, H.; Kim, H.; Lee, D.H.; et al. Different Dietary Fibre Sources and Risks of Colorectal Cancer and Adenoma: A Dose–Response Meta-Analysis of Prospective Studies. Br. J. Nutr. 2019, 122, 605–615. [CrossRef]
- Makarem, N.; Nicholson, J.M.; Bandera, E.V.; et al. Consumption of Whole Grains and Cereal Fiber in Relation to Cancer Risk: A Systematic Review of Longitudinal Studies. Nutr. Rev. 2016, 74, 353–373. [CrossRef]
- Shikata, K.; Ninomiya, T.; Kiyohara, Y. Diabetes Mellitus and Cancer Risk: Review of the Epidemiological Evidence. Cancer Sci. 2013, 104, 9–14. [CrossRef]
- Pajari, A.M.; Freese, R.; Kariluoto, S.; Lampi, A.M.; Piironen, V. Bioactive Compounds in Whole Grains and Their Implications for Health. Whole Grains Health; 2021; pp. 301-336. [CrossRef]
Figure 1.
Images of whole grains included in the study (a) Pseudo-cereals, quinoa, amaranth and buckwheat: (b) Cereals, wheat, rice and corn.
Figure 1.
Images of whole grains included in the study (a) Pseudo-cereals, quinoa, amaranth and buckwheat: (b) Cereals, wheat, rice and corn.

Figure 2.
General structure of dietary fibres and phenolic acids (a) arabinoxylan structure from cereal grains. The (1→4)-linked β-Dxylopyranosyl backbone (in black) is substituted with side-chains, or branch elements, including O–3 and/or O–2-linked α-L-arabinofuranose units (in green) can be attached to the Xyl residues, which may be acylated on the O–5 position with ferulic acid (in red) to form feruloylated side-chains. β-(1-4) glycosidic linkage, (b) Cereal β-glucan is composed of cellotriosyl and cellotetraosyl units linked through β-1-3, β-1-4-linked d-glucose units (c) and (d) Structures of naturally occurring phenolic acids. (c) Hydroxybenzoic; (d), hydroxycinnamic structures. R1, R2, and R3 represents H, OH, and OCH3.
Figure 2.
General structure of dietary fibres and phenolic acids (a) arabinoxylan structure from cereal grains. The (1→4)-linked β-Dxylopyranosyl backbone (in black) is substituted with side-chains, or branch elements, including O–3 and/or O–2-linked α-L-arabinofuranose units (in green) can be attached to the Xyl residues, which may be acylated on the O–5 position with ferulic acid (in red) to form feruloylated side-chains. β-(1-4) glycosidic linkage, (b) Cereal β-glucan is composed of cellotriosyl and cellotetraosyl units linked through β-1-3, β-1-4-linked d-glucose units (c) and (d) Structures of naturally occurring phenolic acids. (c) Hydroxybenzoic; (d), hydroxycinnamic structures. R1, R2, and R3 represents H, OH, and OCH3.
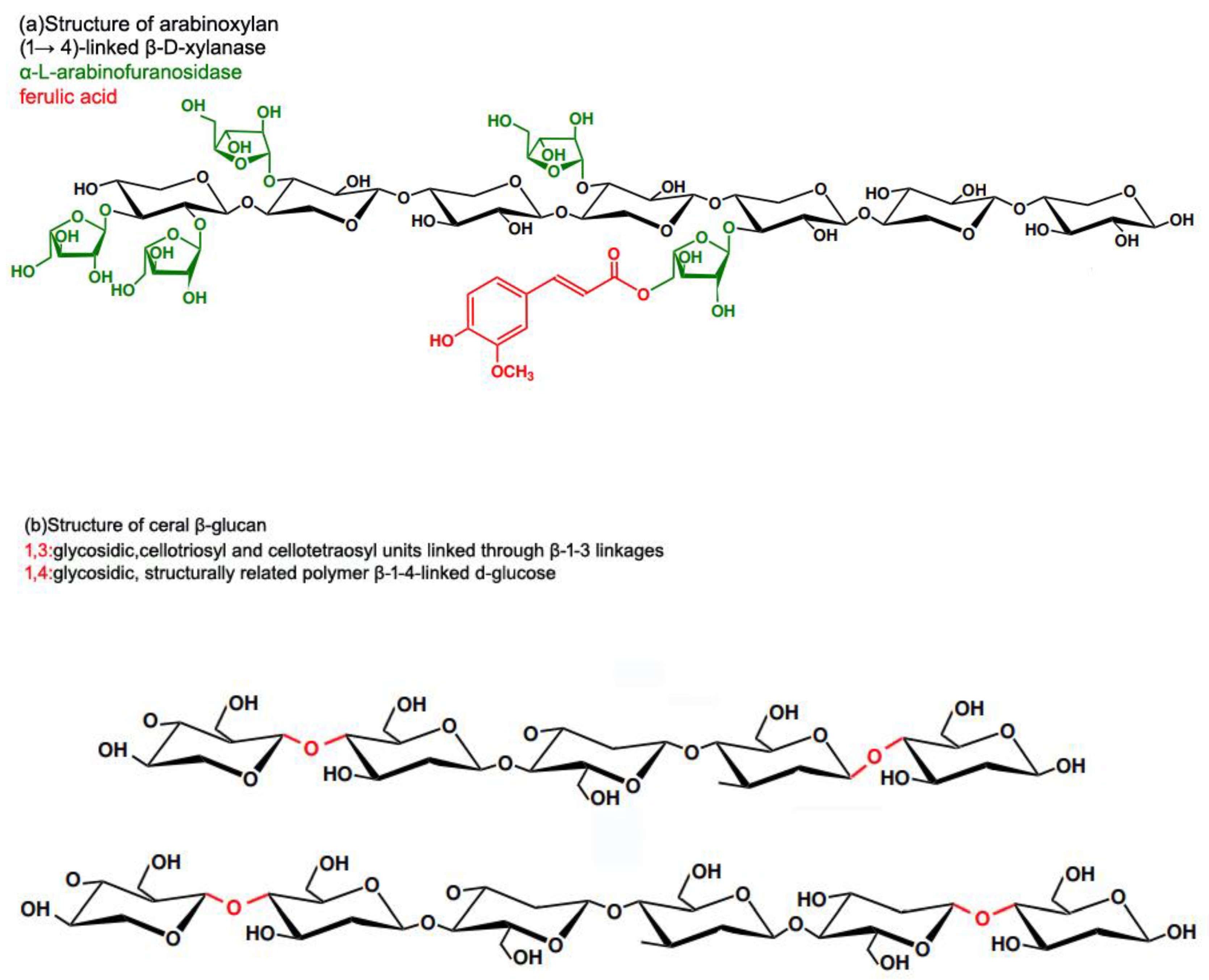
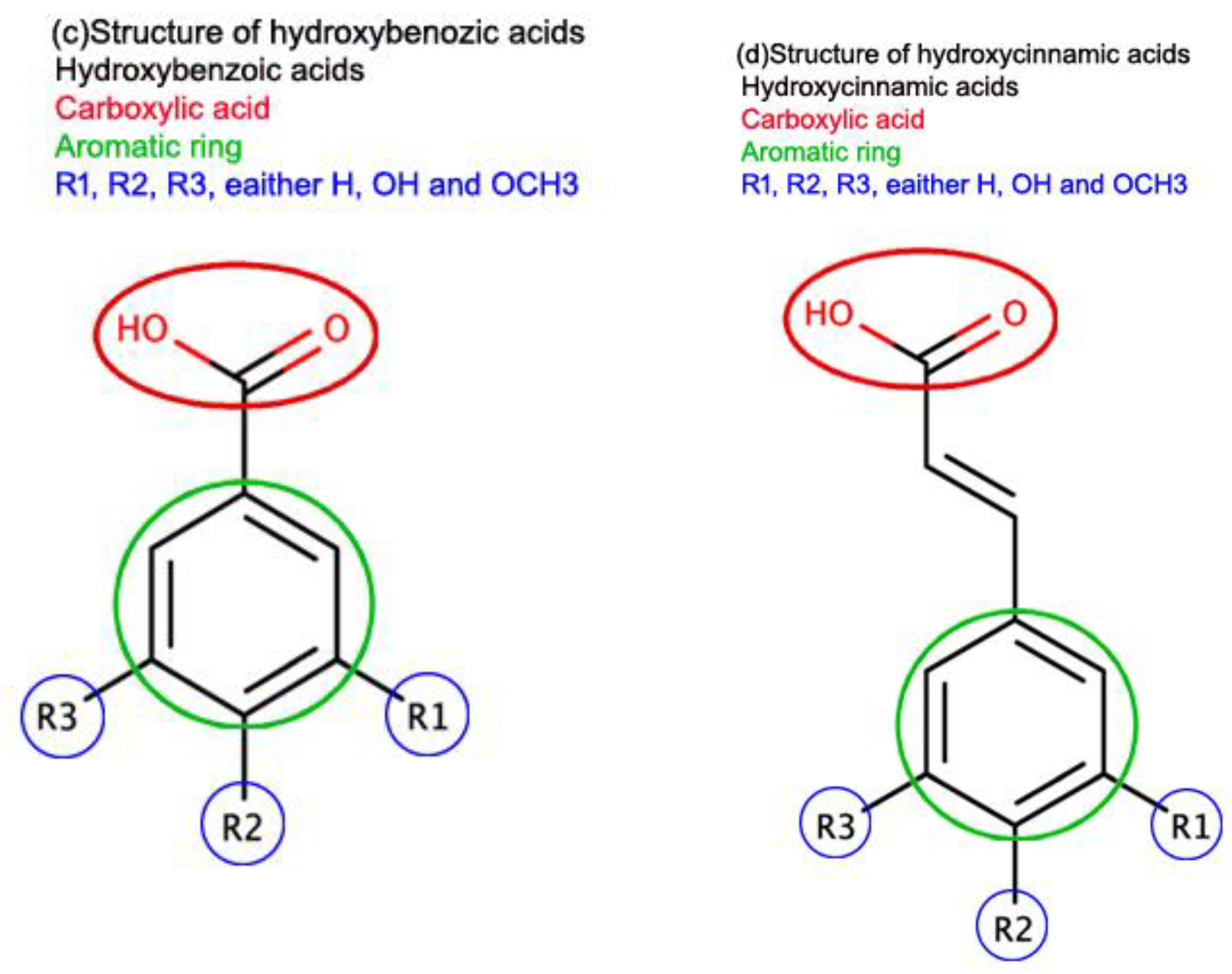
Figure 3.
Summary of Table 1, Table 2, Table 3 in graphical form TDF; Total dietary fibres, SDF; Soluble dietary fibrs, IDF; Insoluble dietary fibres, T;Threonine, V;Valine, P;Phenylalanine, I;Isoleucine, L;Leucine, M;Methionine, T;Tryptophan, L;Lysine, A;Aspartate, G;Glutamate, S;Serine, H;Histidine, G;Glycine, A;Arginine, A;Alanine, T;Tyrosine, C;Cysteine, P;Proline.
Figure 3.
Summary of Table 1, Table 2, Table 3 in graphical form TDF; Total dietary fibres, SDF; Soluble dietary fibrs, IDF; Insoluble dietary fibres, T;Threonine, V;Valine, P;Phenylalanine, I;Isoleucine, L;Leucine, M;Methionine, T;Tryptophan, L;Lysine, A;Aspartate, G;Glutamate, S;Serine, H;Histidine, G;Glycine, A;Arginine, A;Alanine, T;Tyrosine, C;Cysteine, P;Proline.
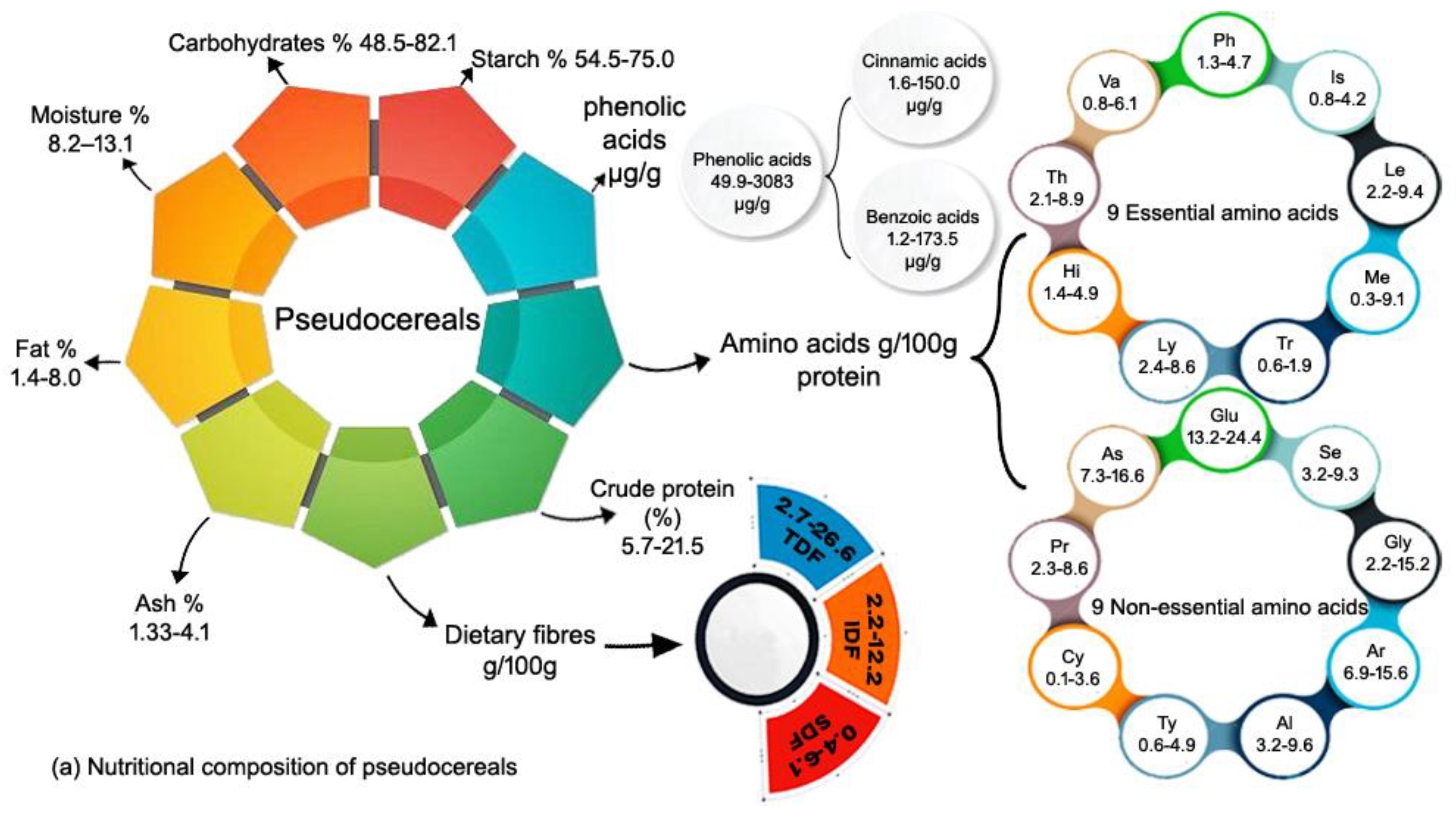
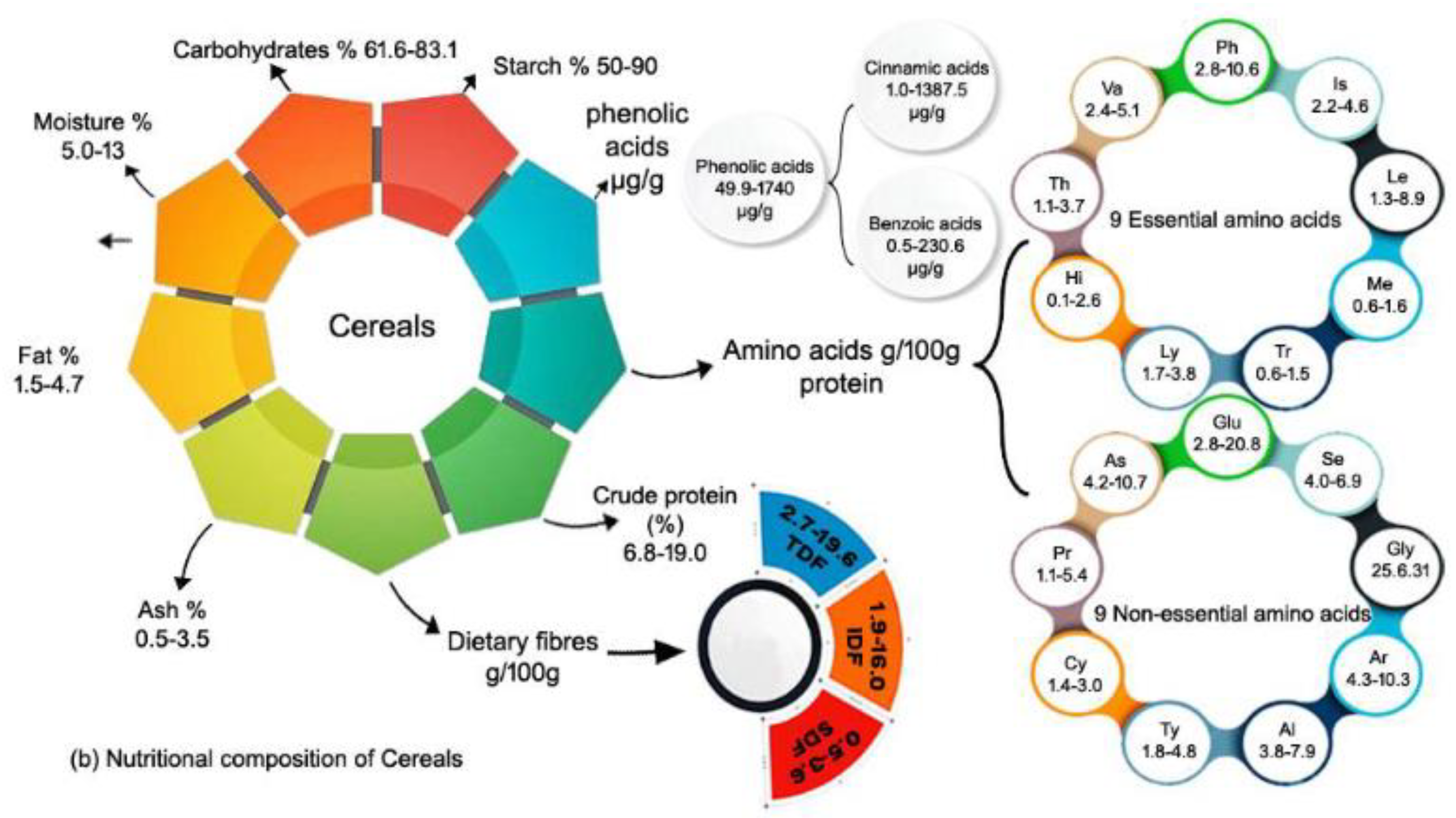
Figure 4.
Potential mechanism of whole grains intake and reduce risk of T2D; peptide PYY; CCK: cholecystokinin; GIP: gastric inhibitory peptide; GLP-1: glucagon like peptide: SCFA, short chain fatty acids.
Figure 4.
Potential mechanism of whole grains intake and reduce risk of T2D; peptide PYY; CCK: cholecystokinin; GIP: gastric inhibitory peptide; GLP-1: glucagon like peptide: SCFA, short chain fatty acids.
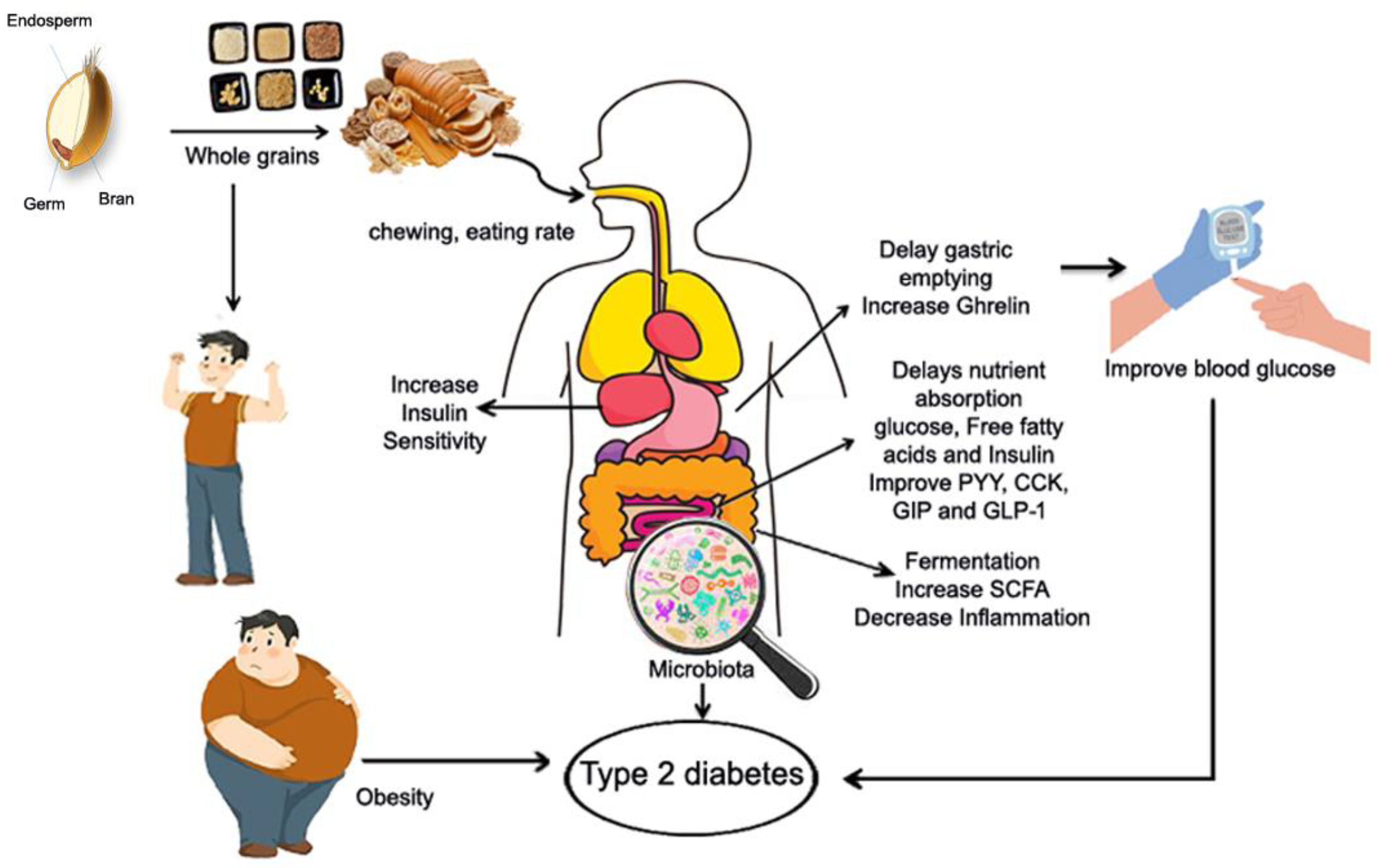
Figure 5.
Potential mechanism of whole grains intake and reduce risk of cancer insulin-like growth factor-binding protein 1 (IGFBP-1) insulin-like growth factor-binding protein 2 (IGFBP-2).
Figure 5.
Potential mechanism of whole grains intake and reduce risk of cancer insulin-like growth factor-binding protein 1 (IGFBP-1) insulin-like growth factor-binding protein 2 (IGFBP-2).
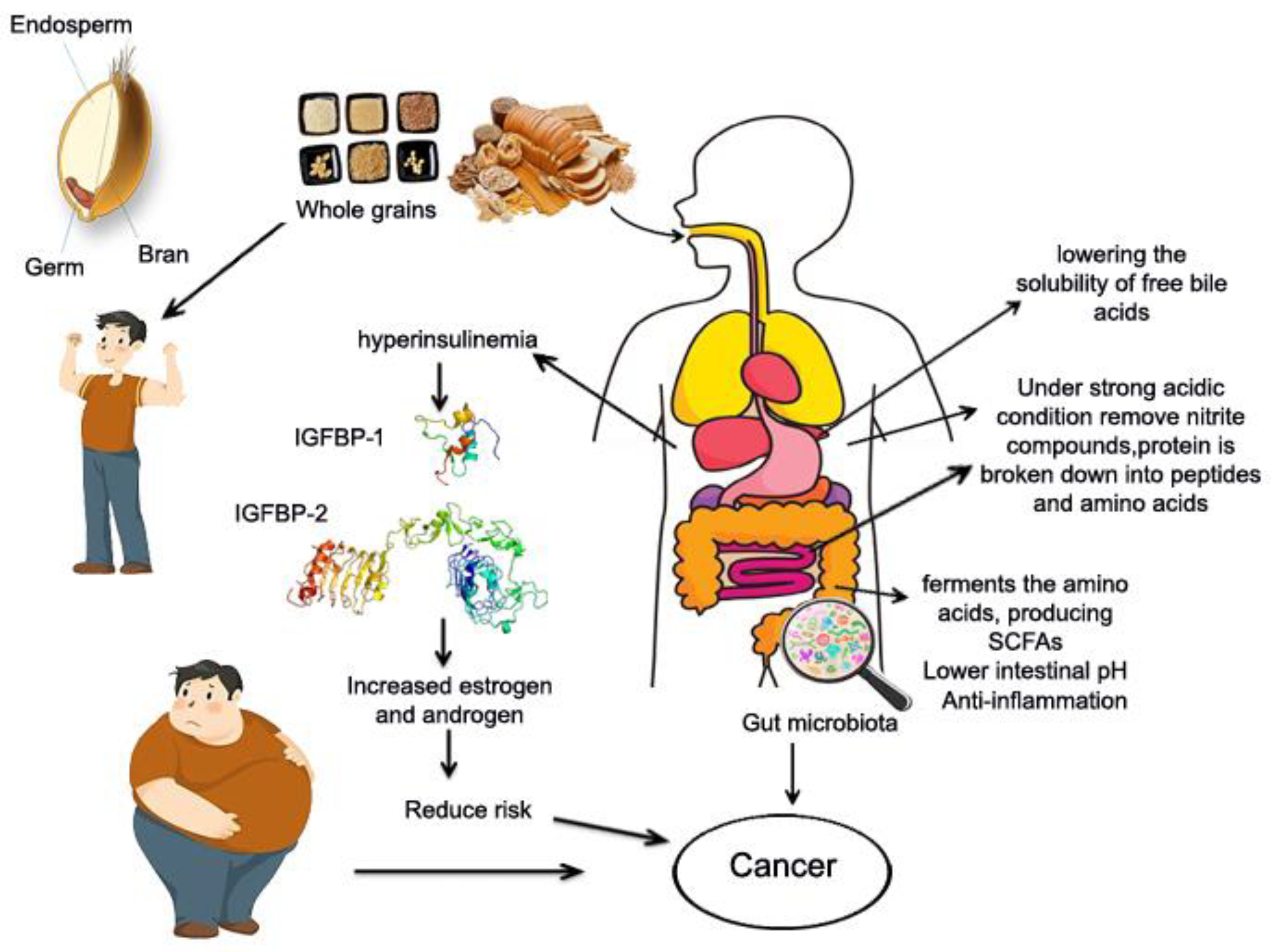
Table 3.
Dietary fibres, TDF; Total dietary fibres, IDF; Insoluble dietary fibres, SDF; Soluble dietary fibres, PA;phenolic acids, HC;hydroxycinnamic and HB;hydroxybenzoic acids contents of pseudocereals and cereals.
Table 3.
Dietary fibres, TDF; Total dietary fibres, IDF; Insoluble dietary fibres, SDF; Soluble dietary fibres, PA;phenolic acids, HC;hydroxycinnamic and HB;hydroxybenzoic acids contents of pseudocereals and cereals.
| Pseudo cereals | ||||||
| Whole grains | TDF | IDF | SDF | PA | HC | HB |
| Quinoa | 7.0–26.5 g/100g [35,61,108] |
9.9–12.2 g/100g [66,107] |
0.4–2.9 g/100g [66,108] | 1672–3083 µg/g[50] | 7.0 to 150.0 µg/g [131,132] | 13.8-110.0 µg/g [131] |
| Amaranth | 2.7–20.6 g/100g [35,50,109] | 7.7-9.1 g/100g [107,110] | 2.7-3.7 g/100g [107,110] | 212–570 µg/g [50,51] | 1.6-55.4 µg/g [127] | 1.8-173.5 µg/g [128] |
| Buckwheat | 10.3-19.0 g/100g [33,35,51] |
2.2-5.8 g/100g [33,111] | 4.8-6.1 g/100g [33,111] | 49.9 µg/g [120] | 1.7-122.8 µg/g [120,133] | 1.2-118.0 µg/g [120,133] |
| Cereals | ||||||
| Wheat | 9.2–17.0 g/100g [35,61,112] | 7.2–14.7 g/100g [112,113] | 1.4–2.9 g/100g [112,113] | 658–1171 µg /g [121] | 3.4-195.0 µg/g [129,130] | 7.5 to 230.6 µg/g [129,130] |
| Rice | 2.7–9.9 g/100g [114,115] | 1.9–5.4 g/100g [114,115] |
0.5–2.5 g/100g [114,115] | 300–360 µg/g [124] | 1.0-301.7 µg/g [122,129] | 2.8-115.6 µg/g [129,175] |
| corn | 13.1–19.6 g/100g [35,117] | 3.1–16.0 g/100g [35,116,117] | 1.5–3.6 g/100g [35,117] | 601–1740 µg/g [121,122] | 5.7-1387.5 µg/g [126,127] | 0.5–116.5 µg/g [126,127] |
Table 4.
Antioxidant activities of pseudo cereals and cereals.
| Pseudo cereals | |||||||
| Cereals type | DPPH | FRAP | ABTS | TEAC | CUPRAC | References | |
| Quinoa | 60.22 (mmol Trolox/100g) | 58.7 [mg Trolox/100 g dw | 8.61( µmolTrolox/100g) | 59.56 (µmolTrolox/100g) | 4.968(µmolTrolox /g) | [147,151–153] | |
| Amaranth | 134.8(mmol Trolox/100g) | 147.4(µmolTrolox/100g) | 179.8(µmolTrolox/100g) | _ | 5.11 (µmol Trolox /g | [145,147,151] | |
| Buckwheat | 80.80(mmol Trolox/100g) | 49.43(µmolTrolox/100g) | 58(mmolTrolox/100g) | 5.93 (µmol/100g) | 4.14 (µmol Trolox /g) | [151,154–156] | |
| Cereals | |||||||
| wheat | 20.9 (mmol Trolox/100g) | 70. µmol TE/g dw | 5.4(mmolTroex/100g) | 17.8 mmol TEAC/g | 13.44 (mmol Trolox /g) | [140–144] | |
| Rice | 180.41(mmol Trolox/100g) | 46.5(µmolTrolox/100g) | 1.78mgTEAC/g extract | 21.3 (mg Trolox equiv./100g | 3.21 (µmol Trolox /g) | [137–139,144] | |
| Corn | 350.29(mmolTrolox/100g) | 30.56(μmolTrolox/100g) | 92.69(μmol Trolox/100g) | 59.6 µg/ml | 22.78(mg/g) | [136,148–150] | |
Table 5.
Summary of whole grains intake and reduction risk of T2D, ↓; decrease, ↑; increase.
| Pseudo cereals | |||||||
| Whole grains | Action mechanism | Model | doses | Key findings | References | ||
| Quinoa | Anti-diabatic | 24 rats; 5 weeks | quinoa seeds in high-fructose diet | ↓ Blood glucose, ↓blood triglycerides, ↓low density lipoprotein, ↓ total cholesterol | [164] | ||
| Anti-diabatic | 55 Male albino rats; 6 weeks | quinoa seeds powder | ↓glucose levels, ↓thyroid hormones | [165] | |||
| Amranth | Anti-diabatic | 30 rats; 5 weeks | Amaranth grains | ↑Calcium in the diet and improved calcium signaling in blood, kidney, and liver of diabetic rats. ↑expression of the s100a1 calcium transport proteins | [166] | ||
| Anti-diabatic | 25 CDI mice; 4 weeks | Amranth protein hydrolyzate | ↑plasma insulin | [17] | |||
| Buckwheat | Anti-diabatic | 165 Diabetic Individuals;4 weeks |
Buckwheat |
↓ level of serum glucose, ↓fast insulin, ↓total cholesterol, ↓LDL cholesterol |
[167] | ||
| Anti-diabatic | 50 C57BL/6 mice; weeks | Buckwheat soluble dietary fibres | ↓ levels of fasting blood glucose, ↑ oral glucose tolerance, ↑ levels of liver glycogen and insulin, ↑ lipid profiles in both the serum and liver. | [168] | |||
| Cereals | |||||||
| Wheat | Anti-diabatic | 57,053 Men and women 50–65 y a | Whole grain Wheat | 16 g of whole wheat per day lower risk of type 2 diabetes and high whole-grain intake may have more benefits | [169] | ||
| Anti-diabatic | 120 patients of T2D | Black wheat grains intake (>69 g/d) for 5 weeks | ↑ glycemia and the inflammatory profile in T2D patients, ↓ glycated albumin, and prevented the increase in TNF-α and IL-6 levels | [170] | |||
| Rice | Anti-diabetic | 4618796 Men and women | Brown rice | higher consumption of total whole grains and the most commonly consumed whole grain foods was significantly associated with a lower risk of type 2 diabetes | [171] | ||
| diabatic | 45,411 male and female aged 45–74, 25,666 men 33,622 women age 45–75 y ,132,373 individuals age 35–70 y, 13284 cases, 2352384 participants | White rice | intake of white rice is associated with an increased risk of type 2 diabetes | [174–177] | |||
| corn | Anti-diabatic | 4 groups mice; 8 week | Purple corn | ↑ insulin secretion ↑ AMPK activation in the liver ,↑ phosphorylation of activated protein kinase, ↓phosphoenolpyruvate carboxykinase, ↓ glucose 6-phosphatase | [172] | ||
| Anti-diabatic | 6 mice; 1-6 week | corn | ↓ serum level ↓ pancreatic gene expression of IL-6 and insulitis, ↑ pancreatic β-cell areas, pancreatic gene, ↑ expression of IL-10 serum levels of serine and histidine | [173] | |||
Table 6.
Summary of whole grains intake and reduction risk of cancer, ↓; decrease, ↑; increase.
| Pseudo cereals | |||||
| Whole grains | Action mechanism | Model | doses | Key findings | References |
| Quinoa | Cancer | 35 females; 2 years | quinoa flakes daily | ↓interleukin-6, which is a marker of inflammation, ↓Tumor, ↓total cholesterol ↓serum triglyceride |
[192] |
| Colon cancer | in vitro gastrointestinal digestion model | Quinoa protein | Large peptides responsible for the colon cancer cell viability inhibitory activitySmaller peptides< 5 kDa with antiproliferative activity i and was the main responsible for the radical scavenging activity while peptides > 5 kDa showed greater anti-cancer effects. | [193] | |
| Amranth | Cancer | Animal model | Amranth protein and polyphenols | Inhibitory effect on tumour cell proliferation inhibition of histone acetylation | [16] |
| Breast cancer | Human breast cells | Amranth | The results indicated that the digested sample was capable of inhibiting cell growth and found that amaranth may be a good source of bioactive peptides with good antioxidant activity and promising anticancer activity. | [80] | |
| buckwheat | Prostate cancer | 10 weeks old mice | crude polysaccharides buckwheat | negative correlation between PC-3 cell viabilities and (interleukin [IL]-6 + tumor necrosis factor [TNF]-α)/IL-10 level ratios in the corresponding MCM, implying that macrophages suppress PC-3 cell growth through decreasing secretion ratios of proinflammatory/anti-inflammatory cytokines in a tumor microenvironment. | [194] |
| Colon cancer | Mice weight 70g and 28 g | High protein buckwheat flour | strong activities against cancer; In the design of studyL rats exposed to an artificially induced tumor were found to be protected against colon cancer by buckwheat protein | [195] |
|
| Cereals | |||||
| wheat | Colectral cancer | Human study 1372 colorectal cancer cases | Wheat phenolic acids | High concentrations of phenolic acids were associated with a lower incidence of distal colon cancer but not with overall colorectal cancer, proximal colon cancer, and rectal cancer. | [196] |
| Colon cancer | mice | Whole wheat vs refined wheat | Red wheat significantly fewer colonic precancerous lesions than soft white-fed, while refined grains have no reduction risk, oxygen radical and fecal bile acid concentration were highers than refined grains | [197] | |
| rice | Cancer | Rat model of liver cancer | Rice bran | ↑glutathione-S-transferase activity, ↓lipid peroxidation↓ level of placental glutathione-S-transferase -positive foci, a marker of hepato-carcinogenesis, ↓ number of colon tumours | [198,199] |
| Colectral cancer | Rat model of colorectal cancer | Rice bran phytochemicals | Reverse the effect of chemically-induced colorectal cancer in rats, by reducing level of lipid peroxidation and protein oxidation in liver, ↑ activity of superoxide dismutase, catalase and glutathione peroxidase ↑ glutathione, vitamin E and vitamin C levels ↓ number of aberrant crypt foci and colon tumours |
[200] | |
| corn | Cancer | 4–5 wk, male rats | corns | inhibition of β-glucuronidase activity, and induction of detoxifying enzymes in liver and colon, as well as a decrease in the expression of the two most important proliferative proteins (K-ras and β-catenin) involved in colon carcinogenesis | [201] |
| prostate cancer | 36 rats; 8 weeks | Purple corn | ↓ incidence of adenocarcinoma in the lateral prostate and slowed down the progression of prostate cancer. ↓ expression cyclin-dependent kinases, downregulation of the activation growth factor and cytokines |
[202] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



