
Submitted:
07 June 2024
Posted:
10 June 2024
You are already at the latest version
Abstract
The inclusion of exogenous phytase in broiler P and Ca deficient diets in response to heightened concerns over excessive P excretion into the environment over the years has remarkably been documented. However, responses among these studies have been inconsistent because of the several factors affecting P utilization. For this reason, a systematic review with meta-analysis of results from forty-one studies published from 2000 to February 2024 was evaluated to: 1) quantitatively summarize the size of phytase effect on growth performance, bone strength, and mineralization in broilers fed P and Ca deficient basal diets and 2) estimate and explore the heterogeneity of the effect sizes of outcomes using sub-group and meta-regression analyses. The quality of the included studies was assessed using the Cochrane Collaboration’s SYRCLE Risk of Bias checklists for animal studies. Additionally, applying the random-effects models, Hedges’ g effect size of supplementation with phytase was calculated using the R software to determine the standardized mean difference (SMD) at a 95% confidence interval. Sub-group analysis and meta-regression further explored heterogeneity (PSMD ≤ 0.05, I2 > 50%, n ≥ 10). The meta-analysis showed that supplemental phytase increases ADFI and BWG and reduces FCR at each time point of growth (< 0.0001). Additionally, phytase supplementation consistently increased tibia ash, P and Ca, and bone strength (< 0.0001) of broilers fed P and Ca deficient diets. Results of the subgroup and meta-regression analyses showed that age and strain of broiler, dietary P source, and duration of phytase exposure significantly influence the effect size of phytase on growth and bone parameters. In conclusion, phytase can attenuate the effect of reducing dietary available phosphorus and calcium and improve ADFI, BWG, and reduced FCR, especially when added to starter diets. It further enhances bone ash, bone mineralization, and bone-breaking strength of broilers, even though the effects of bone ash and strength can be maximized in the starter phase of growth. However, the effect sizes of phytase were related to the age and strain of the broiler, dietary P source, and duration of phytase exposure rather than the dosage.
Keywords:
Broilers
; Calcium
; Exogenous enzyme
; Growth performance
; Phosphorus
; Mineral utilization
; Welfare
1. Introduction
Calcium and phosphorus, constituting the first (99%) and second (80%) stored minerals in the skeleton as hydroxyapatite, are vital for numerous essential functions [1]. Their presence in balanced proportions within diets is equally crucial. These macro minerals, whether fed separately or in conjunction, are pivotal in facilitating bone formation and fulfilling various metabolic requirements [2]. Yet, phosphorus (P) remains one of monogastric nutrition’s most widely researched nutrients due to its importance in several biochemical reactions and its debilitating environmental effects if not properly managed. [3]. Poultry and pigs, in particular, among non-ruminants, are known efficient feed converters to muscle but are unable to sufficiently utilize phosphorus (P) from plant-based feed ingredients [4,5,6], with significant amounts of P excreted in animal waste [7]. The peculiarity is that most of these feed ingredients constituting a substantial portion of the broiler diet are from the cereals and oilseed families [8], with P in the form of phytin P [9]. This P, approximately 50 - 85%, is bound as the salt form of phytic acid, called phytate [10,11,12].
Phytate, a phosphorus storage form in plants, serves as both a nutrient and an antioxidant but also acts as an antinutrient. It’s known to be less readily available to non-ruminants, like broilers [5,6]. Within the small intestines, phytate forms robust chelates with cations such as Ca2+ or Cu2+, creating insoluble complexes that impede the effectiveness of endogenous phytase in the avian gut. This, in turn, decreases their capacity to release P [3,10,11,12] and promotes interference with protein digestion [13]. Due to this reason, inorganic phosphate became necessary to tackle the low P bioavailability issue and satisfy the P requirement of poultry [14]. Adding inorganic P, an expensive and finite mineral resource, to diets that already may contain enough P, though in an unavailable form, results in total P levels above the requirement [14]. Consequently, much of the excess P in the diet is excreted in animal waste [15]. As excreted P is a function of total P, heightened concerns in the past decade over phosphorus pollution of the environment presented policies about P regulation in animal feeding operations [14,16,17]. Following the increasing awareness and concerns, cut downs on “insurance” P levels in animal feeds prompted poultry nutritionists to supplement feed additives, such as phytase in the nutrition of non-ruminants [18,19,20,21].
Phytase efficacy to release phytate-bound phosphorus and improve the bioavailability of phosphorus in poultry feed is well-known [22,23]. Supplementation with commercial exogenous phytase, a common practice in poultry nutrition, is established to reduce the addition of inorganic phosphorus (P) and calcium (Ca) in poultry feed, improve growth performance, nutrient utilization, bone mineralization, and increase amino acid digestibility [24,25] at diverse physiological stages, releasing P and another mineral (e.g., Ca, Zn, Fe, Cu) [8], as well as reducing environmental P pollution [4]. However, wide variations in supplemented phytase effectiveness on P availability, even at the same phytase dose and type of diet, are reported to contribute to unreliable diet formulation. [26]. More recent studies have indicated that the release and utilization of P due to supplemented phytase is variable, depending on the characteristic of the birds (species, age, and physiological conditions), dietary factors such as phytate concentration, the source of phytate, mineral concentrations, and the source, dosing and feeding duration of phytase. [27,28,29,30,31,32].
The effects of supplemental phytase in P and Ca deficient diets on growth performance, bone strength, and mineralization in broiler chickens have been extensively studied [33,34,35,36,37,38,39,40]. However, these studies were conducted under different conditions, making generalization from such results difficult. Meta-analysis (MA), a statistical method, combines numerical summary results of independent studies/trials to pool an estimate of the effect size measures. [41]. Compared to that reported in individual studies, when the results of independent studies are conflicting, MA arrives at a better estimate of the population effect size. Its use in exogenous phytase (EP) studies in broilers has focused on non-phytate phosphorus, phosphorus retention, interactive effects of Ca, vitamin D3, and Eimeria infections [42,43,44,45,46]. As the genetic selection of broiler chickens, new generations of EP, and new studies continue to proliferate in this research domain, evaluating P utilization in current broiler chickens’ strains, dietary P source, and physiological stage is imperative. Therefore, the objectives of this study were to 1) summarize the size of the supplemented exogenous phytase effect on growth performance, bone strength, and mineralization in broilers fed P and Ca deficient basal diets and 2) estimate and explore the heterogeneity of the effect sizes of outcomes using subgroup and meta-regression analyses. Hence, we hypothesized that reducing P and Ca in a diet supplemented with phytase would not compromise broilers’ performance, bone strength, and mineralization at different growth phases.
2. Materials and Methods
2.1. Literature Search and Study Screening
To thoroughly frame and construct the right testable questions, the Population, Interventions, Comparators, and Outcomes (PICO) elements suggested by Schmid, et al. [47] motivated the foundation of the review topic, the protocol, and the strategy for searching the relevant literature. In identifying the PICO elements, Population (broiler chicken), Interventions (varying doses of exogenous phytase supplemented to basal stand-alone P and Ca deficient diet), Comparators (stand-alone P and Ca deficient basal diet), and Outcomes (response variables including growth performance, bone strength, and mineralization) were established. The meta-analysis, with registration number INPLASY20240096, followed the Preferred Reporting Items for Systematic Reviews and Meta-Analysis (PRISMA) updated guidelines [48] for identifying, selecting, choosing, and including information, as shown in Figure 1.
To address the research question, “exogenous phytases phytate unlocking efficacy for potential muscle growth and bone mineralization in broiler at different growth phases,” a comprehensive literature search for articles published between 2000 and February 2024 was conducted using Web of Science (accessed on 21 February 2024), Scopus (accessed on 22 February 2024), ScienceDirect (accessed on 21 February 2024), PubMed (accessed on 22 February 2024), Poultry Science (accessed on 21 February 2024), and Google Scholar (accessed on 20 February 2024) online databases. In all the databases, the keywords “phytase supplementation”, “phosphorus”, “broiler chicken”, “growth”, “bone mineralization”, and “blood characteristics” were used.
2.2. Inclusion and Exclusion Criteria
Search results from the six databases collectively (1305) were pooled in Zotero (Version 6.0.36), and then duplicate articles were removed. To ensure the appropriateness of the remaining articles in addressing the current research question, two reviewers independently and thoroughly applied a two-step screening process. First, screening was performed using the title and abstract to exclude review papers, conference papers, stimulated studies (in vitro), and studies that did not include broilers and did not measure the response variables of interest. Papers that passed the title and abstract screening were assessed for eligibility in the second step based on the inclusion and exclusion criteria of the meta-analysis. Inclusion criteria: (1) peer-reviewed journal article published in English, (2) studies involving stand-alone P and Ca deficient basal diet supplemented with phytase, (3) studies on broiler strains at either starter or grower-finisher phase, (4) studies with a randomized allotment of broilers, (5) studies with a quantified dose of phytase, (6) studies that reported the means of the control and experimental group with variability measures (standard deviation or standard error of mean) and sample size, and (7) studies that reported the parameters of interest. The exclusion criteria included (1) challenged studies, (2) studies with phytase fed as a replacement in the diet of starter and grower-finisher broilers, and (4) studies with phytase combined with other exogenous enzymes or additives. Forty-one (41) full-text articles were used for the meta-analysis, as illustrated in Figure 1.
2.3. Data Extraction
After screening, two main categories of data were extracted independently by two team members from studies identified as relevant (Table 1). The characteristics and design features of studies including author, year, country, breed/strain, phase of growth, basal diet source of phosphorus and calculated chemical composition of the stand-alone P and Ca deficient basal diet, category of phytase, origin of phytase, phytase expression host, dosage of phytase, and supplementation duration were extracted from each study. The outcomes of interest came from two categories. The growth performance category included average daily feed intake (ADFI), average daily gain (ADG), body weight gain (BWG), and feed conversion ratio (FCR). The second category, bone strength and mineralization included bone-breaking strength (BBS), tibia ash (TA), tibia calcium (TCa), and tibia phosphorous (TP). The mean and standard deviation of the mean of all outcomes corresponding to phytase and control groups were extracted from each study. Each treatment comparison with the control was considered a separate trial for studies that supplemented more than one phytase dosage. The data extracted from eligible studies were compiled and constructed into a database using structured spreadsheets created in Google Sheets (Google LLC, USA).
2.4. Appraisal of Study and Assessment of Risk of Bias
The assessment of the eligible studies’ quality, validity, and potential risk of bias was conducted independently by two team members using Cochrane Collaboration’s Systematic Review Center for Laboratory Animal Experimentation’s (SYRCLE) Risk of Bias (RoB) checklists of items for animal studies [49]. The assessment items included random sequence generation (selection bias), baseline characteristics (selection bias), allocation concealment (selection bias), random housing (performance bias), blinding of participants and personnel (performance bias), random outcome assessment (detection bias), incomplete outcome data (attrition bias), selective reporting (reporting bias), and other biases. Discussions with a third researcher settled the disagreements in the assessment.
2.5. Statistical Analysis
2.5.1. Meta-Analysis (MA)
Sixteen (16) separate meta-analyses were carried out to combine estimates of phytase supplementation on growth performance, bone strength, and mineralization in broilers across studies using the “meta” and “metafor” packages of R (version 4.3.3, “Angel Food Cake”, R Foundation for Statistical Computing Platform). The means of the experimental units (control and treatment) were registered as continuous data, and their effect was calculated with Hedges’ g as the standardized mean difference (SMD), also called the effect size (ES), using methods for the random-effects model. The difference between the means of the treated and the control groups was standardized using the standard deviation (SD) of the group’s mean with and without exogenous phytase. The random-effects model was used to estimate the effect size since it is more conservative than the fixed-effects model. [50,51]. At a confidence interval (CI) of 95%, calculated SMD with a p-value ≤ 0.05 was declared statistically significant.
2.5.2. Heterogeneity Assessment
The effect size heterogeneity was assessed and quantified using the chi-square test (Q) and the I2 statistics. With relatively low power, the Q test of a p-value of ≤ 0.10 was considered significant heterogeneity. [52]. The I2 statistic, which expresses the proportion of between-study variability, on the other hand, was assessed using values ranging from 0 to 100%. Using the Borenstein, et al. [53] benchmark range for I2 statistics, I2 < 25%, 25% ≤ I2 ≤ 50%, 50% ≤ I2 < 75%, and 75% ≤ I2 ≤ 100% were interpreted as representing low, moderate, high, and very high levels of heterogeneity, respectively. Where substantial heterogeneity (I2 > 50%) among analyzed outcomes was detected in the MA, meta-ANOVA (subgroup analysis) and meta-regression analyses were utilized to determine the sources of heterogeneity between studies.

Table 1.
A summary of the characteristics of the studies included in the meta-analysis.
| Studies | Country | Broiler Strain | Dietary P Source1 | Origin | Phytase Expression Host | Category of Phytase |
|---|---|---|---|---|---|---|
| Ajuwon et al. [54] | Germany | Ross 308 | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Beeson et al. [4] | UK | Ross 308 | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Bello et al. [55] | Canada | Ross 308 | Corn-Soybean | Bacteria |
Buttiauxella sp. Citrobacter braakii |
6-phytase 6-phytase |
| Borda-Molina et al. [56] | Germany | Ross 308 | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Bowen et al. [57] | USA | Ross 708 | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Broch et al. [33] | Brazil | Cobb 500 | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Broch et al. [34] | Brazil | Cobb 500 | Corn-Soybean | Fungi | Aspergillus oryzae | 3-phytase |
| Broch et al. [58] | Brazil | Cobb 500 | Corn-Soybean | Fungi | -- | 3-phytase |
| Campasino et al. [35] | USA | Hubbard × Cobb 500 | Corn-Soybean | Fungi | Aspergillus oryzae | 6-phytase |
| Cowieson et al. [36] | UK | Ross | Corn-Soybean | Fungi | Aspergillus oryzae | 6-phytase |
| Cowieson et al. [37] | Poland | Ross 308 | WCS | Bacteria | Escherichia coli | 6-phytase |
| Dersjant-Li et al. [59] | USA | Cobb 500 | Corn-Soybean | Bacteria | Buttiauxella sp. | 6-phytase |
| Dersjant-Li et al. [38] | New Zealand | Ross 308 | CSRRB | Bacteria | Trichoderma reesei | 6-phytase |
| Dessimoni et al. [39] | Brazil | Cobb 500 | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Ennis et al. [60] | USA | Ross × Ross 708 | Corn-Soybean | Bacteria Bacteria Fungi |
Escherichia coli Buttiauxella sp. Aspergillus niger |
6-phytase 6-phytase 3-phytase |
| Gehring et al. [61] | USA | Ross × Ross 708 | WCS | Fungi | Aspergillus niger | 6-phytase |
| Ghahri et al. [62] | Iran | Ross 308 | Corn-Soy-DDGS | Bacteria | Escherichia coli | 6-phytase |
| Gulizia et al. [40] | USA | Ross 708 × YPM | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Hernandez et al. [63] | USA | Cobb 500 | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Houshyar et al. [64] | Iran | Ross 308 | WCS | Bacteria | Serratia odorifera | 3-phytase |
| Javadi eta al. [65] | Spain | Ross | WCS | Bacteria | Trichoderma reesei | 6-phytase |
| Jlali et al. [66] | France | Ross 308 | Wheat-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Józefiak et al. [67] | Poland | Ross 308 | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Karami et al. [68] | Germany | Ross 308 | Corn-Soybean | Bacteria | Buttiauxella sp. | 6-phytase |
| Kiarie et al. [69] | Canada | Ross 308 | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Kriseldi et al. [70] | USA | Yield Plus × Ross 708 | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Kwon et al. [71] | Korea | Ross 308 | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Liu et al. [72] | Australia | Ross 308 | Corn-Soybean | Fungi | Aspergillus oryzae | 6-phytase |
| Moita et al. [73] | USA | Ross 308 | Corn-Soybean | Fungi | Aspergillus niger | 6-phytase |
| Powell et al. [74] | USA | Ross × Ross 508 | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Ptak et al. [75] | Poland | Ross 308 | WRES | Fungi Bacteria |
Aspergillus ficcum Escherichia coli |
3-phytase 6-phytase |
| Ptak et al. [76] | Poland | Ross 308 | WRES | Bacteria | Escherichia coli | 6-phytase |
| Shang et al. [77] | Canada | Ross × Ross 308 | WCS | Bacteria | -- | 6-phytase |
| Shi et al. [78] | USA | Cobb 500 | Corn-Soybean | Bacteria | Buttiauxella sp. | 6-phytase |
| Walk et al. [79] | UK | Ross 308 | Wheat-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Walk et al. [13] | USA | Cobb 500 | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Walk and Olukosi, [80] | UK | Ross 308 | Wheat-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Walk and Poernama, [81] | UK | Lohman Indian River Straight-run | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Walk et al. [82] | USA | Cobb 500 | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| Woyengo et al. [83] | Canada | Ross | Corn-Soybean | Bacteria | -- | 6-phytase |
| Zhang et al. [84] | China | Cobb 500 | Corn-Soybean | Bacteria | Escherichia coli | 6-phytase |
| China | Ross 308 | Corn-Soybean | Bacteria | Citrobacter braakii | 6-phytase |
1 WCS: wheat-corn-soybean; CSRRB: corn-soybean-rapeseed-rice bran; WRES: wheat-rapeseed expeller-soybean.
2.5.3. Publication Bias
Publication bias was analyzed to confirm the study results’ validity and assess the risk of bias in individual studies. The funnel plots were drawn to visualize the bias, and Egger’s linear test was performed to evaluate the publication bias accurately with numerical data [85]. Tests to assess publication bias can be achieved when the variable to be considered is at least ten studies and when significant bias is detected with p ≤ 0.05 because it may lead to false-positive claims [86]. Consequently, funnel plots and Egger’s test were only performed for variables that met the criteria above. In cases where statistical evidence of publication bias was found, Duval and Tweedie’s “trim-and-fill” method was used to estimate the number of possible missing observations [87].
2.5.4. Meta-ANOVA and Meta-Regression
Meta-ANOVA tests were conducted to compare the effects of the broiler strains and basal diets’ dietary P source. Meta-regression analysis, on the other hand, was performed using effect sizes estimate (SMD) from each control and phytase supplementation treatment group comparison for each outcome (PSMD < 0.05, I2 > 50%, n ≥ 10) as the dependent variable, and phytase dosage (FTU/kg) and supplementation duration (days) as the independent (or explanatory) variable to examine the source of the meta-analysis’ detected heterogeneity.
2.5.5. Descriptive Statistics
Extracted data, including the calculated chemical composition of stand-alone P and Ca deficient basal diet, phytase dosage, and supplementation duration from the eligible studies, were analyzed using the descriptive statistics procedure of Minitab (Version 21.2, 2022).
3. Results
In this meta-analysis, the effects of exogenous phytase supplementation in dietary P and Ca deficient diet were investigated at three developmental stages (starter phase and grower and finisher phases combined) throughout the productive life span, starting with a variable P and Ca supply immediately after hatching. Broiler responses to phytase treatment were ascertained via performance, bone-breaking strength, and mineralization.
3.1. Appraisal of Study and Assessment of Risk of Bias
Figure 2 presents the risk of bias classification for the studies included in our meta-analysis based on the Cochrane Collaboration’s SYRCLE Risk of Bias tool for animal studies. Pertain to selection bias, all the included studies (41) were judged as having an unclear risk of bias for the sequence generation domain. However, their baseline characteristics were reported and weighed as a 100% low risk of bias. Allocation concealment of the domain of selection bias, on the other hand, constituted 2.44% for high risk and low risk of bias, respectively. However, most (95.12%) of the included studies showed an unclear risk. Regarding performance bias, all the included studies were appraised as having a low risk of bias (100%) for both random housing and blinding of the caregiver’s domain. In contrast, studies appraised and assessed showed almost equal proportion for low (48.78%) and unclear (43.90%) risk, with 7.32% reckoned as high risk in the case of random outcome assessment and blinding of outcome assessors (detection bias). Additionally, 17.07% of studies included in our meta-analysis were adjudged as unclear risk in describing the completeness of outcome data. Eligible studies of our meta-analysis were all considered free of selective outcome reporting and other problems and, therefore, appraised as 100% low risk of bias. In summary, of the 41 eligible studies in our meta-analysis, approximately 68.3% were judged as low risk of bias, unclear risk of bias (30%), and 1.7% constituted a high risk of bias.
3.2. Descriptive Statistics of Independent Variables
Descriptive statistics of the P and Ca deficient basal diet, phytase dosage, and supplementation duration in this meta-analysis are presented in Table 2. The average of the eligible studies formulated basal diets’ (negative diet) calculated chemical composition fed in (d 1 to 22) or (d 22 to 42) met or exceeded the NRC [88] nutrient specifications for young broilers, except for Ca and total P, which were reduced by 0.26 and 0.32 % of DM, respectively. In addition, the average phytase dosage supplemented was 1709 (FTU per Kg of diet), with a minimum of 120 and a maximum of 40500 in the starter phase, while the average (876) of the grower-finisher phase varied between 250 and 5000 FTU per Kg of diet. Regarding the duration of exposure to phytase, it ranged between 6 and 22 days and 10 and 33 days for the starter and grower-finisher phases, respectively.
3.3. Meta-Analysis
3.3.1. Growth Performance
The supplemental phytase effects on starter and grower-finisher meat-type chickens’ growth performance were summarized using random-effects meta-analysis models. The standardized mean difference estimates of ADFI, ADG, BWG, and FCR in the control and phytase supplementation treatment groups with corresponding heterogeneity estimates are presented in Table 3. In the starter phase (1-22 days), supplemental phytase increased ADFI (PSMD < 0.0001), ADG (PSMD = 0.0006), and BWG (PSMD < 0.0001). However, the estimate of FCR decreased with dietary supplementation of phytase. Similarly, in the grower-finisher phase (22-42 days), supplemental phytase significantly increased ADFI (PSMD = 0.0123) and BWG (PSMD = 0.002) but non-significantly decreased ADG (PSMD = 0.0699). Consistent with supplemental phytase’s influence on broiler chicken’s FCR as indicated in the starter group meat-type chicken, it significantly reduced (PSMD < 0.0001) in the grower-finisher phase. Supplementing phytase positively impacted growth responses in both phases, although the extent of phytase impact was much higher in the starter phase.
3.3.2. Bone Strength and Mineralization
The impact of supplemental phytase on broilers’ bone strength and mineralization responses was characterized using meta-analysis’s random-effects models and displayed in Table 4. Supplementation of the P and Ca deficient basal diet with exogenous phytase improved the BBS and tibia ash (PSMD < 0.0001), tibia Ca (PSMD = 0.0217), and tibia P (PSMD = 0.0024) of broilers. Similar observations of phytase improvement for BBS and tibia ash (PSMD < 0.0001), tibia Ca (PSMD = 0.0047), and tibia P (PSMD = 0.0036) were also recorded in the grower-finisher broiler. Notwithstanding supplemental phytase’s general improvement of bone strength and mineralization responses, its impact comparatively was reduced for BBS (SMD = 4.68 vs. 4.33) and tibia ash (SMD = 11.78 vs. 5.88) and heightened for tibia Ca (SMD = 4.35 vs 4.94) and tibia P (SMD = 5.26 vs 5.72) among broilers of the grower-finisher phase of growth.
3.4. Heterogeneity and Sensitivity Analysis (Publication Bias)
Table 3 and Table 4 indicate the X2 statistics (Q) test of heterogeneity and the corresponding proportion of total between-study variation of effect size estimates. In all the growth performance, bone strength, and mineralization responses measured in our meta-analysis, the estimates of the Q test revealed statistics with p-values ≤ 0.10, which were considered significant between-study variability with their corresponding proportions of heterogeneity (I2) ranging between 93.3 to 99.8%. With reference to the Borenstein, et al. [7] benchmark range for heterogeneity, the I2 statistics detected among analyzed outcomes in our MA were interpreted as representing very high levels of heterogeneity, as all the proportions fell within 75% ≤ I2 ≤ 100%. To address this concern, it was, therefore, imperative to explore what causes this heterogeneity.
On the other hand, visualized funnel plots (S17-20) and the p-values of all the measured outcomes in the MA using Egger’s regression asymmetry test revealed a significant bias (p ≤ 0.05). With sufficient evidence of publication bias detected among all the outcome variables in the MA, the results of the corrected publication bias by Duval and Tweedie’s “trim-and-fill” method (illustrated in Table 5), suggest that improvements seen in terms of effect size estimate of growth performance and bone strength and mineralization outcomes are actual effects of the supplemental phytase. Nonetheless, the magnitude of the effect sizes on measured responses in our MA, except BWG, were non-significantly reduced in our adjustment of publication bias (Table 5) compared to the observations in Table 3 and Table 4 in both phases of development.
3.5. Subgroup Analysis of Broiler Strain and Basal Diet’s Dietary P Source Association with Phytase Efficacy on Growth Performance, Bone Strength, and Mineralization
Subgroup analysis was performed to test the hypothesis that supplementing phytase in a P and Ca deficient diet is more effective among some strains of broilers as well as basal diets’ dietary P sources than others (i.e., studies in our meta-analysis do not stem from one overall population), assuming that they fall into different subgroups and that each subgroup has its actual overall effect [89]. Besides, in the case of a few studies within subgroups (say, five or fewer), estimates within subgroups are generally likely to be subject to greater sampling variation and, hence, are less precise [90,91]. Therefore, subgroups with just NC = 1 were not reported in our meta-analysis.
3.5.1. Broiler Strain and Growth Performance
The summary effects of the response of ADFI, ADG, BWG, and FCR, as illustrated in Table 6, revealed that the efficacy of phytase in a Ca and P deficient diet generally varied significantly (p < 0.0001) among broiler strain subgroups in both phases of production.
Among subgroups of starter broiler strains, supplementation with phytase consistently increased (p < 0.0001) the effects on ADFI, which was shown to be more pronounced and precise among some strains of broilers than others (Cobb 500 > Ross 308 > Ross > Ross × Ross 708 > Ross 708). Phytase’s positive effect on feed intake predictably translated to improved (p < 0.0001) response of broiler’s ADG, with much of it seen in Ross 308 compared to Cobb 500 strains. However, supplementation with phytase reduced (p < 0.0001) ADG in Ross strains. Interestingly, all subgroups of broiler strains recorded increased (p < 0.0001) BWG, with the observed effects pronounced among strains, including Cobb 500, Ross, Ross 708, Ross 308, Yield Plus × Ross 708, and Ross × Ross 708. Contrariwise to the effects of phytase intervention on starter broilers’ ADFI, ADG, and BWG, broilers’ FCR was significantly (p < 0.0001) decreased among Ross, Ross 708, Cobb 500, Ross 308, Hubbard × Cobb 500, Yield Plus × Ross 708, Ross × Ross 708, and Lohman Indian RSR, except in Ross 708 × YPM strain that recorded an increased. Similarly, among the growing-finishing broiler subgroups, ADFI (p = 0.0044), ADG (p < 0.0001), and BWG (p < 0.0001) indicators of growth were heightened in Cob 500 and Ross 308 strains, except the strain of Ross that recorded reduced effects. In contrast, a decreasing trend of FCR (p < 0.0001) was observed in Cobb 500, Ross 308, and Ross strains when fed P and Ca deficient basal diet supplemented with phytase.
It was observed that the impact of supplemented phytase in ameliorating the effects of the P and Ca deficiency on growth performance seemed to increase (ADFI, ADG, and BWG) and reduce (FCR) at each time point in both the starter and grower-finisher phases. However, the magnitude of phytase effects was pronounced and more precise in some strains of broilers than in others, generally during the starter phase than the grower-finisher phase.
3.5.2. Broiler Strain and Bone Strength and Mineralization
The subgroup analyses of broiler strains’ correlation with phytase effects on bone strength and mineralization in starter and grower-finisher phases are displayed in Table 7. Exogenous phytase supplementation to a low Ca and P-diet positively (p < 0.05) influenced bone mineralization indicators, including tibia ash, tibia Ca, and tibia phosphorus, but not bone-breaking strength in both phases of growth in our meta-analysis.
Bone-breaking strength (BBS) of broiler chicken supplemented with phytase at three weeks (p = 0.4776) and six weeks (p = 0.19) was not significantly associated with broiler strain, suggesting that effect sizes are consistent across subgroups. Yet, Ross 308, Cobb 500, and Ross strains’ tibia ash, calcium, and phosphorus were pronouncedly influenced by phytase supplementation in low Ca and P diets in both phases in our meta-analysis. Relatively, the magnitude of the effects of phytase varied with broiler strain and growth phase. Whereas BBS had reduced in both Cobb 500 and Ross 308 in the grower-finisher phase, the impact of supplemental phytase increased all bone mineralization indicators assessed in Cobb 500 strains. However, it yielded lower SMDs in Ross 308 in the grower-finisher phase.
3.5.3. Basal Diets’ Dietary P Source and Growth Performance
Statistics from subgroup analyses of basal diet’s dietary P as a moderator of phytase effects on responses of growth in broilers in both developmental stages in our meta-analysis are presented in Table 8. At three weeks, Table 8 shows that broilers’ ADFI (p = 0.0024), ADG (p < 0.0001), and FCR (p < 0.0001) varied significantly. Yet, there was no dietary P source influence on broilers’ BWG (p = 0.3167). Except for WRES (SMD = − 0.32), which decreased, supplemental phytase effects on broiler’s ADFI increased when fed corn-soybean (SMD = 15.33), wheat-soybean (SMD = 10.10), corn-soybean-DDGS (SMD = 8.99), CSRRB (SMD = 6.56), and wheat-corn-soybean (SMD = 1.86). Similarly, ADG increased when broilers were fed corn-soybean (SMD = 9.31) and wheat-soybean (SMD = 23.02) but reduced with wheat-corn-soybean (SMD = − 0.31) being fed. Body weight gain was not associated substantially with the basal diet’s dietary P source, signifying a consistent effect of phytase across the subgroups. In contrast, a general reducing effect of supplemental phytase on FCR is seen in all dietary P source subgroups, with substantial impact observed when corn-soybean (SMD = 10.79) and wheat-corn soybean (SMD = 4.35) were fed as basal diets.
Unlike the response pattern at three weeks, dietary P source as a moderator in our subgroup analysis significantly influenced ADFI (p < 0.0001), ADG (p = 0.0059), BWG (p = 0.0002), and FCR (p = 0.0005) at week 6 of growth. Supplemental phytase improved ADFI of broilers fed corn-soybean (SMD = 86.28), corn-soybean-DDGS (SMD = 39.35), and WRES (SMD = 0.62) but reduced intake when supplemented to wheat-corn-soybean (SMD = − 1.10). Comparable observations in the effects of phytase were revealed in the ADG and BWG of broilers fed wheat-corn-soybean but improved when supplemented to corn-soybean, corn-soybean-DDGS, and WRES. On the contrary, FCR decreased when phytase was augmented in all basal diets, with highly significant effects observed in corn-soybean and WRES. In general, feeding starter and grower-finisher broilers with a corn-soybean diet supplemented with phytase was much more beneficial and consistent in improving the growth performance of the broiler.
3.5.4. Basal Diets’ Dietary P Source and Bone Strength and Mineralization
The bone strength and mineralization of broilers supplemented with phytase are illustrated in Table 9. Basal diet’s dietary P source was a significant predictor for BBS (p = 0.0330), tibia ash (p < 0.0001), and tibia Ca (p = 0.5041) of broilers of 3 weeks, but not tibia P (p = 0.1534). Except in tibia Ca, the results indicated consistent effects necessitated by phytase in broilers fed corn-soybean, wheat-soybean, and wheat-corn-soybean diets. However, wheat-based diets realized a much greater effect size than corn-based diets in all responses. Like the starter phase, the effect size of BBS (p < 0.0001), tibia ash (p = 0.0007), tibia Ca (p < 0.0001), and tibia P (p < 0.0001) of broilers in week six was significantly predicted by the sources of P fed in our subgroup analysis. Although corn-soybean basal diet was associated with better effects of phytase in measured outcomes compared to wheat-corn-soybean, greater bone strength and mineralization are generally more associated with a wheat-soybean and WRES-based diet.
3.6. Effect of Phytase Dosage and Supplementation Duration on Broilers’ Growth Performance, Bone Strength, and Mineralization
A meta-regression analysis was carried out to investigate phytase dose-response and exposure duration to identify the summary trend emerging from multiple studies answering our research question. Analyses were performed to test whether there is any association, whether the response changes approximately at a constant rate throughout the observed outcome effects, and whether there is any substantial change in the outcome beyond the average treatment effects.
3.6.1. Meta-Regression Analysis of Phytase Dose-Response
The dose-response meta-analysis by regression models of broiler growth, bone strength, and mineralization per developmental phase are summarized in Table 10. During the starter phase (at three weeks), the covariate “phytase dosage” had no significant relationship (p > 0.05) with ADFI, ADG, BWG, or FCR. Likewise, the dosage of phytase supplemented to starter broilers had no significant relationship (p > 0.05) with BBS, tibia ash, and tibia calcium, except for tibia P, which was significantly changed (intercept = 9.5629; estimate = − 0.0032; p = 0.0467) with a unit increase in dosage, explaining 15.50% of the observed heterogeneity for tibia P. This observation is highlighted by the bubble plot (Figure 3), which showed a significant negative impact on tibia P due to increasing phytase dosage (500–3000 FTU per Kg of diet).
On the other hand, the results of the meta-regression models during the grower-finisher phase indicated a non-significant (p > 0.05) phytase dosage effect for growth outcomes (ADFI, ADG, BWG, and FCR) and bone strength and mineralization (BBS, tibia ash, tibia Ca, and tibia P).
3.6.2. Phytase Exposure Duration
Table 11 shows a summarized effect size investigating the association between phytase exposure duration with growth performance, bone strength, and mineralization arising from both developmental phases of broilers in the eligible studies of our MA. The results of the starter phase indicated that there was a non-significant (p > 0.05) association between phytase exposure duration and growth outcomes measured, except for FCR (intercept = − 29.0970; estimate = 1.1359; p = 0.0132), which improved significantly per unit increase in phytase duration. Equally, phytase exposure for a longer duration in broilers in the early phase of their development did not correlate significantly with bone-breaking strength and tibia ash but substantially lowered the effect size of tibia Ca (intercept = 19.4068; estimate = − 0.9962; p = 0.0074) and tibia P (intercept = 24.2718; estimate = − 1.2321; p <.0001). Phytase exposure duration as a predicting variable in our meta-regression models further explained 29.05% and 58% of the observed heterogeneity in tibia Ca and P accordingly.
On the other hand, in grower-finisher broilers, the duration of phytase exposure was a non-significant (p > 0.05) contributing predictor of the association effects in all growth-measured outcomes of broilers-supplemented phytase. However, every unit change in the duration of supplementation increased the association’s effect size of tibia Ca (intercept = − 43.7178; estimate = 2.5215; p = 0.0034) and tibia P (intercept = − 56.0797; estimate = 3.2005; p = 0.0001) significantly, explaining 40.79% and 56.07% of the heterogeneity in our global study. Remarkable improvements in tibia mineralization are notable during the late phase of broiler development as compared to the starter phase, which reveals reducing effects as shown in the bubble plots (Figure 4). Notwithstanding these observations in tibia mineralization, supplementation duration did not substantially (p > 0.05) impact the effect size of bone-breaking strength and tibia ash.
4. Discussion
As documented by numerous studies, bone integrity and growth performance data have been commonly used to estimate and validate phytate P liberation by phytase, as these parameters provide additional useful mechanistic information [32]. More precisely, supplemental phytase effects on broiler chicken growth performance, bone strength, and mineralization have been completed in P and Ca inadequate diets with variable influence. To ascertain exogenous phytase global effects, meta-analyses focused on non-phytate phosphorus, phosphorus retention, and interactive effects of Ca, vitamin D3, and Eimeria infections have previously been explored [42,43,44,45,46]. Yet, as the genetic selection of broiler chickens, new generations of EP, and new studies continue to proliferate in this research domain, evaluating P utilization in current broiler chickens’ strains, dietary P source, and physiological stage is imperative.
4.1. Appraisal of Study and Assessment of Risk of Bias
A systematic review’s quality of evidence is as important as analyzing the data [89]. Indeed, according to Macleod, et al. [92], the quality and reliability of evidence are closely linked to the credibility of the data and the results of the individual studies included in the systematic review. A measure of a particular study’s quality by assessing its risk of bias is, thus, fundamental in the systematic review of a research question. Spanning between 2006 and 2024, the studies integrated into our review tail towards the period of surge in the call for the adoption and use of similar reporting standards in the description of the methods section of animal intervention data-driven studies. This notwithstanding, 30% of the review’s included studies were prone to an unclear risk of bias. The magnitude of unclear bias, significantly introduced by a lack of blinding, allocation concealment, sequence, and inadequate randomization in our appraisal, ascertains that similar reporting standards are not yet universal in animal intervention experimentation reporting. Our observation substantiates the arguments of Macleod et al. [92] and Kilkenny, et al. [93], who hold the view that most individual studies do not report blinding, allocation concealment, sequence generation, and randomization, which are not standard practices in animal experiment reporting. In agreement with our results, Nuamah et al. [89] and Ncho, et al. [94], both in a meta-analysis of animal intervention studies, found these risk domains as the main contributors of unclear bias. However, with an overall low risk of bias (68.3%) and just 1.7% high risk of bias, the evidence of our meta-analysis is credible and valid in measuring the overall effect size of phytase supplemented with inadequate P-and-Ca basal diets in broilers.
4.2. Impact of Exogenous Phytase on Growth Performance
As a general concept, the addition of phytase is well documented to hydrolyze phytate and reduce its anti-nutritional effects, thereby offering improved availability and digestibility of P, which have consistently been shown to support and enhance broiler performance and nutrient utilization when fed with P-deficient diets [11,23,95,96,97,98]. Our meta-analysis examined the growth performance of broilers from 1 to 42 days post-hatch. In the two 3-week periods of d 1 to 22 and d 22 to 42, performance indices, including ADFI and BWG, increased while FCR reduced during both periods, especially with a higher magnitude of phytase effect size in d 1 to 22 broilers fed corn-soybean dietary P source diet. This suggests that at all growth phases, phytase facilitates the bioavailability of enough phytate P from a P and Ca deficient diet to meet the requirement for growth.
Exogenous enzymes, including phytase influence on performance, generally correlate with increases in nutrient digestibility [83,95,99], as it is renowned for breaking phytate complexes to liberate P and other dietary essential nutrients, which in turn increase productive performance in birds with diets minimal in avP [100] or both Ca and avP [13]. The current global study thus confirms, as pointed out in the meta-analysis by Letourneau-Montminy et al. [45], Kermani et al. [44], and Shi et al. [78], whose analyses illustrated the positive effect of phytase on P utilization by broilers through improvement in growth performance, particularly BWG and FCR, with the most remarkable improvements realized in P-and Ca-deficient diets [101,102,103]. This claim, according to Tamim, et al. [104] and Gifford and Clydesdale [105], is accounted for by the less insoluble Ca-phytate complex formation in the gut owing to the lower level of Ca in the diet and the lower pH of the gut, which maintains a modest Ca-P balance in the diet. Poultry utilization of phytate P is inclined to the dietary Ca and P content [105,106,107] since Ca could quickly form a precipitate with phytate in the gut [108] and reduce the solubility of InsP6, making it less accessible to phosphatases [23]. For optimal broiler growth, feeding inadequate diets of P and Ca, the dietary calcium concentration, in essence, must be adjusted to the level that optimizes P utilization.
Regarding feed intake, broilers’ overall phytase effect size (Table 3) supports the claims that incremental phosphorus release is linked with feed intake increase, as P often acts as a regulator of feed intake [109,110]. Synergistically with other enzymes, supplemental phytase in poultry increases the transit rate through the gut, thus increasing feed intake [111]. Besides, the improved feed intake might also be due to the change in the viscosity of the diets [112], possibly attributed to the increased availability of P, other minerals, and nutrients, allowing a better-quality diet [113].
Supplemental phytase effect size in BWG and FCR in the present meta-analysis may be ascribed to improvement in P and Ca inadequate diet’s energy value, necessitated by the activity of the liberated phytate P. The improvement in BWG and FCR is possibly linked to the bird’s ability to utilize the energy of feed metabolize in the body owing to the diet’s increase in protein and amino acid digestibility, which correlates positively with growth performance indices [114]. It has been suggested by Humer, et al. [115] that the mechanism of energy improvements, particularly with phytase association, may be due to the dissolution of the phytate complexes, which promote protein absorption and increase carbohydrate and fat digestibility. Partly related to the improvement could be the exogenous phytase’s ability to offset endogenous losses to enhance metabolizable energy by lowering the energy required for maintenance, thus granting a significant amount of energy for growth. Confirming our observation further, the work of Abd El-Hack, et al. [116], ascribed superior broiler performance associated with phytase supplementation to increased energy released from the diet owing to a boost in nutrient digestibility via the release of nutrients bound to phytic acid and increased phosphorus utilization efficiency.
Contrary to the liberation excess P, the improvement in growth performance may be ascribed to the excess phytase dose (Table 2) supplemented in the starter phase. Thus, superdosing phytase in excess of commercial recommendation proposed further phytate hydrolysis per se results in improvements in BWG and FCR, particularly in young broilers [13]. Likewise, the extra-phosphoric effect of the phytase, arising from the liberation of myo-inositol, the final product of phytate dephosphorylation, might have partly contributed to growth improvement in both phases. Precisely, myo-inositol generated from dephosphorylation of the whole phytate increases broiler chickens’ growth rate and feed efficiency because of its crucial role in cell growth and metabolism, fat deposition and transport, protein deposition and transport, and gluconeogenesis. [117,118].
However, improvements in broiler performance by phytase supplementation of P and Ca inadequate diets are not solely mediated by the exogenous phytase but also depend on the feed composition, mineral content, source of the phytase, species and age of birds, and the endogenous microbiota that affects the pH range in the gut. [116,119]. In the present meta-analysis, although exogenous phytase’s impact to ameliorate the effects of the P and Ca deficiency on growth performance appeared to increase ADFI, ADG, and BWG and reduce FCR at each phase, the extent of its impact is strain (Table 7), age, and dietary P source-dependent (Table 8), with much higher impact observed in starter phase. The findings of the present study contradict those of Edwards Jr, et al. [120], who reported that the ability to utilize phytate P increases with the age of poultry, coupled with Peeler [121], who, in an earlier review, said that the biological availability of phytate phosphorus is intermediate for adult poultry but very low for young poultry. Converse to these assertions, a recent study by Abudabos [122] confirms our observation regarding phytate P utilization with age, affirming that the demand for P is more pronounced in young poultry for body weight gain. Moreover, the utilization of nutrients in chicks is more pronounced during the first two weeks because rapid development and growth of organs and tissues occur during this period [29,30]. In the case of broiler strain, significant prediction of growth performance in the present study inclines to the fact that the genetics of chicken influence the magnitude of indices of growth [123]. Cobb 500 and Ross 308 strains probably had a higher adaptive capacity to the deficient diet, and so increased their intestinal phytase and phosphatase activities, which generally improved performance and mineralization, touting them as efficient nutrient utilizers of low P and Ca diets supplemented with phytase.
Interestingly, the magnitude of growth responses throughout the production cycle of birds appears to also depend on the dietary P source, i.e., type of ingredient (Table 8) and supplementation duration (Table 11) other than the dosage of phytase (Table 10), as both explain a significant portion of the heterogeneity in our global study. Suggesting that phytase activity differs per the dietary P source, as each source shares distinct phytate susceptibility, probably because of their structure and storage of phytate. Consistent with Ravindran, et al. [124], the difference is predominantly due to the solubility of dietary P source at acidic pH, as phytase is more readily hydrolyzed by soluble phytate. Accordingly, the magnitude of the response to phytase is directly proportional to the concentration of phytate in ingredients, with poorly digestible feed ingredients showing more response to phytase than those with higher inherent digestibility. The discrepancy between the corn-SBM-based diet and wheat-SBM-based diet in our subgroup analysis may partly be associated with the propensity and storage site of phytate in the grains [125], which is more likely to complex wheat protein than that of maize [112]. Our observation was in line with Selle and Ravindran [23], that phytase hydrolyzes phytate in soybean meal and maize more readily than wheat.
Even though phytase in broiler diets has been supplemented until market weight, the findings from the present study suggest that the efficacy of phytase on growth response is more tailored to the duration of its supplementation and not the dosage. This supports the theory that longer feeding duration may result in broilers adapting to low-P diets to maintain P homeostasis adaptations for the digestive ability of the birds, resulting in higher digestibility of P [126,127,128].
4.3. Impact of Exogenous Phytase on Bone Strength and Mineralization
Bone ash, a highly sensitive metric used for dietary P availability evaluation [129], alongside bone mineralization, an efficient parameter for the prediction of the quality of the bone attributable to inorganic P release by phytase [32,130], are both recognized for their beneficial effects associated with broiler muscular development and performance. Besides, both bone parameters tend to be more precise and sensitive indicators to assess changes in Ca and P availability than live growth indices [108,131,132,133].
In this study, the low P and Ca diet supplemented with phytase increased bone strength and tibia ash, thus predictably increasing bone mineralization at each age. However, the magnitude of BBS and tibia ash decreased, whereas tibia Ca and P increased at d 42. Tibia ash is known to reduce the sensitivity of the bones to dietary changes as the bird ages [134]. The supplementation of phytase seems to be more influential in increasing tibia ash during the early phase of growth of broilers, confirming the evidence of Talaty, et al. [135] that bone mineral density peaked at four weeks of age. According to the authors, the tibia continued to grow, especially after the growth spurt at 3 to 4 weeks, but did not become denser in ash because of the increase in surface area. The observations of the present study are close to those reported in preceding studies [3,8,18,95], which implies that phytase can attenuate the effect of reducing available phosphorus and calcium while maintaining similar bone ash, bone mineralization, and bone-breaking strength of broilers. These observations may be due to the role of phytase and P, where phytase liberates phytate P in the low P and Ca diet. Almost 85% of P is retained in bone, yet Ca provision is required for P deposition through the formation of hydroxyapatite, a mineral phase of the bones [136]. With calcium accentuated as an essential nutritional factor in upholding bone strength, bone strength and mineralization will be reduced without a sufficient dietary calcium source due to blood hypocalcemia [137]. Hence, marginal P and Ca supply in broiler diets will adversely impact bone formation and mineralization, except in phytase supplementation [138]. Likewise, an additional release of bone-forming compounds, including zinc, amino acids, and other substances, according to Fernandes, et al. [139], also improves bone integrity.
Nonetheless, the outcomes of tibia P and Ca and tibia ash (to some extent) were not alone influenced by the phytase supplied but mainly by its duration of feeding (Table 11), dietary sources of P (Table 9), and strains of broilers (Table 7) in this study. This could be due to the increased negative impact of the inadequate diet on tibia ash over a longer period than the shorter feeding duration. Thus, there is an opportunity for continuous deposition of P and Ca on the bones during this developmental phase [95] from prolonged exposure to the ameliorative effects of phytase in the low P and Ca diet. Our findings support the assertion that skeletal integrity is influenced by rapid growth rate, genetics, the environment, management, nutrition, and age [137].
5. Conclusions
The systematic review using meta-analysis models indicates that supplementation of broiler P and Ca-deficient basal diets with phytase improved ADFI, BWG, and reduced FCR, especially when added to starter diets. Moreover, exogenous phytase supplementation supports bone strength and mineralization in broilers fed P and Ca-deficient diets at each time of growth. However, the effects of bone ash and strength can be maximized in the starter phase of development. However, the effect sizes were related to the age and strain of the broiler, dietary P source, and duration of phytase exposure rather than the dosage.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figures S1-S4: Starter growth performance; S5-S8: Grower-Finisher growth performance; S9-S12: Starter bone strength and mineralization; S13-S16: Grower-Finisher bone strength and mineralization; S17.:Trim and fill funnel plot assessment for testing and adjusting publication bias on starter broilers’ growth outcomes; S18: Trim and fill funnel plot assessment for testing and adjusting publication bias on grower-finisher broilers’ growth outcomes; S19: Trim and fill funnel plot assessment for testing and adjusting publication bias on starter broilers’ bone strength and mineralization outcomes; S20: Trim and fill funnel plot assessment for testing and adjusting publication bias on grower-finisher broilers’ bone strength and mineralization outcome.
Author Contributions
Conceptualization, E.N.; methodology, E.N., U.M.O., F.N.A.O. and N-J. C.; software, E.N.; validation, E.N., F.N.A.O. and N-J.C.; formal analysis, E.N. and U.M.O.; investigation, E.N., U.M.O., F.N.A.O. and N-J.C.; resources, E.N., U.M.O. and N-J. C; data curation, E.N. and U.M.O.; writing—original draft preparation, E.N.; writing—review and editing, E.N., U.M.O., E.J., Y.M., I.C., B.C., F.N.A.O., D.K., and N-J. C.; visualization, E.N., U.M.O., E.J., Y.M., I.C. and B.C. supervision, D.K. and N-J. C.; project administration, D.K. and N-J. C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by a grant from the “Cooperative Research Program for Agriculture Science & Technology Development (Project No. RS-2021-RD009994)”, Rural Development Administration, Republic of Korea.
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets used and analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
The author, Emmanuel Nuamah, is grateful for the financial support from the National Institute for International Education (NIIED) toward his studies in South Korea.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Suttle, N. Mineral nutrition of livestock; Cabi GB: 2022.
- National Research Council. Nutrient requirements of poultry; National Academies Press, 1994. [Google Scholar]
- Babatunde, O.O.; Adeola, O. A time-series effect of phytase supplementation on phosphorus utilization in growing and finishing pigs fed a low-phosphorus diet. Journal of Animal Science 2022, 100, skab350. [Google Scholar] [CrossRef] [PubMed]
- Beeson, L.; Walk, C.; Bedford, M.; Olukosi, O. Hydrolysis of phytate to its lower esters can influence the growth performance and nutrient utilization of broilers with regular or super doses of phytase. Poultry Science 2017, 96, 2243–2253. [Google Scholar] [CrossRef] [PubMed]
- Witzig, M.; Camarinha da Silva, A.; Green-Engert, R.; Hoelzle, K.; Zeller, E.; Seifert, J.; Hoelzle, L.E.; Rodehutscord, M. Spatial variation of the gut microbiota in broiler chickens as affected by dietary available phosphorus and assessed by T-RFLP analysis and 454 pyrosequencing. PLoS One 2015, 10, e0143442. [Google Scholar]
- Zeller, E.; Schollenberger, M.; Witzig, M.; Shastak, Y.; Kühn, I.; Hoelzle, L.E.; Rodehutscord, M. Interactions between supplemented mineral phosphorus and phytase on phytate hydrolysis and inositol phosphates in the small intestine of broilers. Poultry Science 2015, 94, 1018–1029. [Google Scholar] [CrossRef] [PubMed]
- Kiarie, E.; Walsh, M.; Nyachoti, C. Performance, digestive function, and mucosal responses to selected feed additives for pigs. Journal of Animal Science 2016, 94, 169–180. [Google Scholar] [CrossRef]
- Babatunde, O.; Cowieson, A.; Wilson, J.; Adeola, O. Influence of age and duration of feeding low-phosphorus diet on phytase efficacy in broiler chickens during the starter phase. Poultry Science 2019, 98, 2588–2597. [Google Scholar] [CrossRef] [PubMed]
- Nelson, T.S. The hydrolysis of phytate phosphorus by chicks and laying hens. Poultry Science 1976, 55, 2262–2264. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Angel, R.; Kim, S.-W.; Brady, K.; Yu, S.; Plumstead, P. Impacts of dietary calcium, phytate, and nonphytate phosphorus concentrations in the presence or absence of phytase on inositol hexakisphosphate (IP6) degradation in different segments of broilers digestive tract. Poultry Science 2016, 95, 581–589. [Google Scholar] [CrossRef]
- Orban, J.; Adeola, O.; Stroshine, R. Microbial phytase in finisher diets of White Pekin ducks: Effects on growth performance, plasma phosphorus concentration, and leg bone characteristics. Poultry Science 1999, 78, 366–377. [Google Scholar] [CrossRef]
- Selle, P.H.; Cowieson, A.J.; Ravindran, V. Consequences of calcium interactions with phytate and phytase for poultry and pigs. Livestock science 2009, 124, 126–141. [Google Scholar] [CrossRef]
- Walk, C.; Bedford, M.; Santos, T.; Paiva, D.; Bradley, J.; Wladecki, H.; Honaker, C.; McElroy, A. Extra-phosphoric effects of superdoses of a novel microbial phytase. Poultry Science 2013, 92, 719–725. [Google Scholar] [CrossRef]
- Tamim, N.M.; Angel, R. Phytate phosphorus hydrolysis as influenced by dietary calcium and micro-mineral source in broiler diets. Journal of Agricultural and Food Chemistry 2003, 51, 4687–4693. [Google Scholar] [CrossRef] [PubMed]
- Adeola, O. Nutrient management procedures to enhance environmental conditions: an introduction. Journal of Animal Science 1999, 77, 427–429. [Google Scholar] [CrossRef]
- Maguire, R.O.; Sims, J.T. Soil testing to predict phosphorus leaching. Journal of environmental quality 2002, 31, 1601–1609. [Google Scholar] [CrossRef]
- Sims, J.; Simard, R.; Joern, B. Phosphorus loss in agricultural drainage: Historical perspective and current research. Journal of Environmental Quality 1998, 27, 277–293. [Google Scholar] [CrossRef]
- Denbow, D.; Ravindran, V.; Kornegay, E.; Yi, Z.; Hulet, R. Improving phosphorus availability in soybean meal for broilers by supplemental phytase. Poultry Science 1995, 74, 1831–1842. [Google Scholar] [CrossRef] [PubMed]
- Harper, A.; Kornegay, E.; Schell, T. Phytase supplementation of low-phosphorus growing-finishing pig diets improves performance, phosphorus digestibility, and bone mineralization and reduces phosphorus excretion. Journal of Animal Science 1997, 75, 3174–3186. [Google Scholar] [CrossRef]
- Huff, W.; Moore Jr, P.; Waldroup, P.; Waldroup, A.; Balog, J.; Huff, G.; Rath, N.; Daniel, T.; Raboy, V. Effect of dietary phytase and high available phosphorus corn on broiler chicken performance. Poultry Science 1998, 77, 1899–1904. [Google Scholar] [CrossRef]
- Yi, Z.; Kornegay, E.; Ravindran, V.; Denbow, D. Improving phytate phosphorus availability in corn and soybean meal for broilers using microbial phytase and calculation of phosphorus equivalency values for phytase. Poultry Science 1996, 75, 240–249. [Google Scholar] [CrossRef]
- Cowieson, A.; Hruby, M.; Pierson, E.M. Evolving enzyme technology: impact on commercial poultry nutrition. Nutrition Research Reviews 2006, 19, 90–103. [Google Scholar] [CrossRef]
- Selle, P.H.; Ravindran, V. Microbial phytase in poultry nutrition. Animal feed Science and Technology 2007, 135, 1–41. [Google Scholar] [CrossRef]
- Borda-Molina, D.; Zuber, T.; Siegert, W.; Camarinha-Silva, A.; Feuerstein, D.; Rodehutscord, M. Effects of protease and phytase supplements on small intestinal microbiota and amino acid digestibility in broiler chickens. Poultry science 2019, 98, 2906–2918. [Google Scholar] [CrossRef] [PubMed]
- Zanu, H.; Keerqin, C.; Kheravii, S.; Morgan, N.; Wu, S.; Bedford, M.; Swick, R. Influence of meat and bone meal, phytase, and antibiotics on broiler chickens challenged with subclinical necrotic enteritis: 1. growth performance, intestinal pH, apparent ileal digestibility, cecal microbiota, and tibial mineralization. Poultry Science 2020, 99, 1540–1550. [Google Scholar] [CrossRef] [PubMed]
- Kebreab, E.; Hansen, A.V.; Strathe, A.B. Animal production for efficient phosphate utilization: from optimized feed to high efficiency livestock. Current Opinion in Biotechnology 2012, 23, 872–877. [Google Scholar] [CrossRef] [PubMed]
- Adeola, O.; Cowieson, A. Board-invited review: opportunities and challenges in using exogenous enzymes to improve nonruminant animal production. Journal of Animal Science 2011, 89, 3189–3218. [Google Scholar] [CrossRef] [PubMed]
- Akter, M.; Iji, P.; Graham, H. Increasing zinc levels in phytase-supplemented diets improves the performance and nutrient utilization of broiler chickens. South African Journal of Animal Science 2017, 47, 648–660. [Google Scholar] [CrossRef]
- Batal, A.; Parsons, C. Effects of age on nutrient digestibility in chicks fed different diets. Poultry Science 2002, 81, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Nitsan, Z.; Ben-Avraham, G.; Zoref, Z.; Nir, I. Growth and development of the digestive organs and some enzymes in broiler chicks after hatching. British Poultry Science 1991, 32, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Perryman, K.; O’Neill, H.M.; Bedford, M.; Dozier III, W. Methodology affects measures of phosphorus availability in growing broilers: Effects of calcium feeding strategy and dietary adaptation period length on true ileal phosphorus digestibility and predicted endogenous phosphorus losses. Poultry Science 2017, 96, 611–621. [Google Scholar] [CrossRef]
- Dersjant-Li, Y.; Awati, A.; Schulze, H.; Partridge, G. Phytase in non-ruminant animal nutrition: a critical review on phytase activities in the gastrointestinal tract and influencing factors. Journal of the Science of Food and Agriculture 2015, 95, 878–896. [Google Scholar] [CrossRef]
- Broch, J.; dos Santos, E.C.; Damasceno, J.L.; Nesello, P.d.O.; de Souza, C.; Eyng, C.; Pesti, G.M.; Nunes, R.V. Phytase and phytate interactions on broilers’ diet at 21 days of age. Journal of applied poultry research 2020, 29, 240–250. [Google Scholar] [CrossRef]
- Broch, J.; Savaris, V.; Wachholz, L.; Cirilo, E.; Tesser, G.; Pacheco, W.; Eyng, C.; Pesti, G.; Nunes, R. Influence of phytate and phytase on performance, bone, and blood parameters of broilers at 42 days old. South African Journal of Animal Science 2021, 51, 160–171. [Google Scholar] [CrossRef]
- Campasino, A.; York, T.; Wyatt, C.; Bedford, M.; Dozier III, W. Effect of increasing supplemental phytase concentration in diets fed to Hubbard× Cobb 500 male broilers from 1 to 42 days of age. Journal of Applied Poultry Research 2014, 23, 705–714. [Google Scholar] [CrossRef]
- Cowieson, A.; Acamovic, T.; Bedford, M. Supplementation of corn–soy-based diets with an Eschericia coli-derived phytase: effects on broiler chick performance and the digestibility of amino acids and metabolizability of minerals and energy. Poultry Science 2006, 85, 1389–1397. [Google Scholar] [CrossRef]
- Cowieson, A.; Ptak, A.; Maćkowiak, P.; Sassek, M.; Pruszyńska-Oszmałek, E.; Żyła, K.; Świątkiewicz, S.; Kaczmarek, S.; Józefiak, D. The effect of microbial phytase and myo-inositol on performance and blood biochemistry of broiler chickens fed wheat/corn-based diets. Poultry Science 2013, 92, 2124–2134. [Google Scholar] [CrossRef] [PubMed]
- Dersjant-Li, Y.; Abdollahi, M.R.; Bello, A.; Waller, K.; Marchal, L.; Ravindran, V. Effects of a novel consensus bacterial 6-phytase variant on the apparent ileal digestibility of amino acids, total tract phosphorus retention, and tibia ash in young broilers. Journal of Animal Science 2022, 100, skac037. [Google Scholar] [CrossRef] [PubMed]
- Dessimoni, G.V.; Sakomura, N.K.; Donato, D.C.Z.; Goldflus, F.; Ferreira, N.T.; Dalólio, F.S. Effect of supplementation with Escherichia coli phytase for broilers on performance, nutrient digestibility, minerals in the tibia and diet cost. Semina: Ciências Agrárias 2019, 40, 767–780. [Google Scholar] [CrossRef]
- Gulizia, J.; Rueda, M.; Ovi, F.; Bonilla, S.; Prasad, R.; Jackson, M.; Gutierrez, O.; Pacheco, W. Evaluate the effect of a commercial heat stable phytase on broiler performance, tibia ash, and mineral excretion from 1 to 49 days of age assessed using nutrient reduced diets. Journal of Applied Poultry Research 2022, 31, 100276. [Google Scholar] [CrossRef]
- Khan, S. Meta-Analysis; Springer Nature Singapore Pte Ltd.: Singapore, 2020; Volume 1, p. 15. [Google Scholar]
- Bougouin, A.; Appuhamy, J.; Kebreab, E.; Dijkstra, J.; Kwakkel, R.; France, J. Effects of phytase supplementation on phosphorus retention in broilers and layers: A meta-analysis. Poultry Science 2014, 93, 1981–1992. [Google Scholar] [CrossRef]
- Faridi, A.; Gitoee, A.; France, J. A meta-analysis of the effects of nonphytate phosphorus on broiler performance and tibia ash concentration. Poultry Science 2015, 94, 2753–2762. [Google Scholar] [CrossRef]
- Kermani, Z.A.; Taheri, H.; Faridi, A.; Shahir, M.; Baradaran, N. Interactive effects of calcium, vitamin D3, and exogenous phytase on phosphorus utilization in male broiler chickens from 1 to 21 days post-hatch: A meta-analysis approach. Animal Feed Science and Technology 2023, 295, 115525. [Google Scholar] [CrossRef]
- Letourneau-Montminy, M.-P.; Narcy, A.; Lescoat, P.; Bernier, J.; Magnin, M.; Pomar, C.; Nys, Y.; Sauvant, D.; Jondreville, C. Meta-analysis of phosphorus utilization by broilers receiving corn-soyabean meal diets: influence of dietary calcium and microbial phytase. Animal 2010, 4, 1844–1853. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Wang, J.; Kim, W.K. Interactive effects of calcium, phosphorus, and exogenous phytase on growth performance and bone ash in broilers under Eimeria or necrotic enteritis infections: a systemic review and meta-analysis. Journal of Applied Poultry Research 2024, 100422. [Google Scholar] [CrossRef]
- Schmid, C.H.; Stijnen, T.; White, I. Handbook of meta-analysis; CRC Press: 2020.
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. International journal of surgery 2021, 88, 105906. [Google Scholar] [CrossRef] [PubMed]
- Hooijmans, C.R.; Rovers, M.M.; De Vries, R.B.; Leenaars, M.; Ritskes-Hoitinga, M.; Langendam, M.W. SYRCLE’s risk of bias tool for animal studies. BMC medical research methodology 2014, 14, 1–9. [Google Scholar] [CrossRef]
- Fleiss, J.L. Review papers: The statistical basis of meta-analysis. Statistical methods in medical research 1993, 2, 121–145. [Google Scholar] [CrossRef] [PubMed]
- Riley, R.D.; Higgins, J.P.; Deeks, J.J. Interpretation of random effects meta-analyses. BMJ 2011, 342. [Google Scholar] [CrossRef] [PubMed]
- Egger, M.; Davey Smith, G. Principles and procedures. Systematic reviews in health care: meta-analysis in context. London: BMJ Books 2001.
- Borenstein, M.; Higgins, J.P.; Hedges, L.V.; Rothstein, H.R. Basics of meta-analysis: I2 is not an absolute measure of heterogeneity. Research synthesis methods 2017, 8, 5–18. [Google Scholar] [CrossRef]
- Ajuwon, K.M.; Sommerfeld, V.; Paul, V.; Däuber, M.; Schollenberger, M.; Kühn, I.; Adeola, O.; Rodehutscord, M. Phytase dosing affects phytate degradation and Muc2 transporter gene expression in broiler starters. Poultry Science 2020, 99, 981–991. [Google Scholar] [CrossRef]
- Bello, A.; Dersjant-Li, Y.; Korver, D. The efficacy of 2 phytases on inositol phosphate degradation in different segments of the gastrointestinal tract, calcium and phosphorus digestibility, and bone quality of broilers. Poultry Science 2019, 98, 5789–5800. [Google Scholar] [CrossRef]
- Borda-Molina, D.; Vital, M.; Sommerfeld, V.; Rodehutscord, M.; Camarinha-Silva, A. Insights into broilers’ gut microbiota fed with phosphorus, calcium, and phytase supplemented diets. Frontiers in Microbiology 2016, 7, 228625. [Google Scholar] [CrossRef] [PubMed]
- Bowen, K.; Jackson, M.; Ayres, V.; Boltz, T.; Lynch, E.; Moritz, J. Performance, carcass quality, tibia ash, and mineral digestibility responses of Ross 708 broilers to increasing dose of two commercially available mixer-added phytases. Journal of Applied Poultry Research 2022, 31, 100264. [Google Scholar] [CrossRef]
- Broch, J.; Nunes, R.V.; Eyng, C.; Pesti, G.M.; de Souza, C.; Sangalli, G.G.; Fascina, V.; Teixeira, L. High levels of dietary phytase improves broiler performance. Animal Feed Science and Technology 2018, 244, 56–65. [Google Scholar] [CrossRef]
- Dersjant-Li, Y.; Archer, G.; Stiewert, A.; Brown, A.; Sobotik, E.; Jasek, A.; Marchal, L.; Bello, A.; Sorg, R.; Christensen, T. Functionality of a next generation biosynthetic bacterial 6-phytase in enhancing phosphorus availability to broilers fed a corn-soybean meal-based diet. Animal Feed Science and Technology 2020, 264, 114481. [Google Scholar] [CrossRef]
- Ennis, C.; Gehring, C.; Bedford, M.; Wyatt, C.; Wamsley, K. Strategies to determine the efficacy of multiple phytase use at low activities using Ross x Ross 708 male broilers from 0 to 14 d. Journal of Applied Poultry Research 2020, 29, 977–994. [Google Scholar] [CrossRef]
- Gehring, C.; Bedford, M.; Dozier III, W. Effects of step-up and step-down phytase regimens on performance and processing yields of male broilers from 1 to 35 d of age. Journal of Applied Poultry Research 2014, 23, 252–259. [Google Scholar] [CrossRef]
- Ghahri, H.; Rostami, D.; Zandiyeh, M.A.; Abbasi, R.H. The effects of phytase on performance, serum mineral levels, enzyme activities and immune function of broilers fed nutritionally marginal diets. Middle East J Sci Res 2012, 11, 1481–1490. [Google Scholar]
- Hernandez, J.R.; Gulizia, J.P.; Adkins, J.B.; Rueda, M.S.; Haruna, S.I.; Pacheco, W.J.; Downs, K.M. Effect of Phytase Level and Form on Broiler Performance, Tibia Characteristics, and Residual Fecal Phytate Phosphorus in Broilers from 1 to 21 Days of Age. Animals 2022, 12, 1952. [Google Scholar] [CrossRef]
- Houshyar, M.; Saki, A.A.; Alikhani, M.Y.; Bedford, M.R.; Soleimani, M.; Kamarehei, F. Approaches to determine the efficiency of novel 3-phytase from Klebsiella pneumoniae and commercial phytase in broilers from 1 to 14 d of age. Poultry Science 2023, 102, 103014. [Google Scholar] [CrossRef]
- Javadi, M.; Cerisuelo, A.; Cambra-López, M.; Macías-Vidal, J.; Donadeu, A.; Dupuy, J.; Carpintero, L.; Ferrer, P.; Pascual, J.J. The Duration of the Trial Influences the Effects of Mineral Deficiency and the Effective Phytase Dose in Broilers’ Diets. Animals 2022, 12, 1418. [Google Scholar] [CrossRef]
- Jlali, M.; Kidd, M.T.; Cozannet, P.; Yavuz, B.; Preynat, A. Efficacy of a new biosynthetic 6-phytase in broilers on phosphorus balance and growth efficiency. Animal Feed Science and Technology 2024, 309, 115886. [Google Scholar] [CrossRef]
- Józefiak, D.; Ptak, A.; Kaczmarek, S.; Maćkowiak, P.; Sassek, M.; Slominski, B. Multi-carbohydrase and phytase supplementation improves growth performance and liver insulin receptor sensitivity in broiler chickens fed diets containing full-fat rapeseed. Poultry Science 2010, 89, 1939–1946. [Google Scholar] [CrossRef] [PubMed]
- Karami, M.; Karimi, A.; Sadeghi, A.; Zentek, J.; Goodarzi Boroojeni, F. Evaluation of interactive effects of phytase and benzoic acid supplementation on performance, nutrients digestibility, tibia mineralisation, gut morphology and serum traits in male broiler chickens. Italian Journal of Animal Science 2020, 19, 1428–1438. [Google Scholar] [CrossRef]
- Kiarie, E.; Woyengo, T.; Nyachoti, C. Efficacy of new 6-phytase from Buttiauxella spp. on growth performance and nutrient retention in broiler chickens fed corn soybean meal-based diets. Asian-Australasian Journal of Animal Sciences 2015, 28, 1479. [Google Scholar] [CrossRef] [PubMed]
- Kriseldi, R.; Walk, C.; Bedford, M.; Dozier III, W. Inositol and gradient phytase supplementation in broiler diets during a 6-week production period: 1. effects on growth performance and meat yield. Poultry Science 2021, 100, 964–972. [Google Scholar] [CrossRef] [PubMed]
- Kwon, C.; Lee, J.; Kim, D.; Kil, D.Y. Determination and validation of available phosphorus equivalency of Escherichia coli-derived phytase in broiler diets. Journal of Applied Poultry Research 2022, 31, 100298. [Google Scholar] [CrossRef]
- Liu, S.Y.; Cowieson, A.J.; Selle, P.H. The influence of meat-and-bone meal and exogenous phytase on growth performance, bone mineralisation and digestibility coefficients of protein (N), amino acids and starch in broiler chickens. Animal Nutrition 2016, 2, 86–92. [Google Scholar] [CrossRef]
- Moita, V.H.C.; Duarte, M.E.; Kim, S.W. Supplemental effects of phytase on modulation of mucosa-associated microbiota in the jejunum and the impacts on nutrient digestibility, intestinal morphology, and bone parameters in broiler chickens. Animals 2021, 11, 3351. [Google Scholar] [CrossRef] [PubMed]
- Powell, S.; Johnston, S.; Gaston, L.; Southern, L. The effect of dietary phosphorus level and phytase supplementation on growth performance, bone-breaking strength, and litter phosphorus concentration in broilers. Poultry Science 2008, 87, 949–957. [Google Scholar] [CrossRef]
- Ptak, A.; Józefiak, D.; Kierończyk, B.; Rawski, M.; Żyła, K.; Świątkiewicz, S. Effect of different phytases on the performance, nutrient retention and tibia composition in broiler chickens. Archives Animal Breeding 2013, 56, 1028–1038. [Google Scholar] [CrossRef]
- Ptak, A.; Bedford, M.R.; Świątkiewicz, S.; Żyła, K.; Jozefiak, D. Phytase modulates ileal microbiota and enhances growth performance of the broiler chickens. PloS One 2015, 10, e0119770. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Rogiewicz, A.; Patterson, R.; Slominski, B.; Kim, W. The effect of phytase and fructooligosaccharide supplementation on growth performance, bone quality, and phosphorus utilization in broiler chickens. Poultry Science 2015, 94, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Wang, J.; White, D.; Martinez, O.J.T.; Kim, W.K. Impacts of phytase and coccidial vaccine on growth performance, nutrient digestibility, bone development, and intestinal gene expression of broilers fed a nutrient reduced diet. Poultry Science 2023, 102, 103062. [Google Scholar] [CrossRef] [PubMed]
- Walk, C.; Bedford, M.; Olukosi, O. Effect of phytase on growth performance, phytate degradation and gene expression of myo-inositol transporters in the small intestine, liver and kidney of 21 day old broilers. Poultry science 2018, 97, 1155–1162. [Google Scholar] [CrossRef] [PubMed]
- Walk, C.; Olukosi, O. Influence of graded concentrations of phytase in high-phytate diets on growth performance, apparent ileal amino acid digestibility, and phytate concentration in broilers from hatch to 28 D post-hatch. Poultry Science 2019, 98, 3884–3893. [Google Scholar] [CrossRef] [PubMed]
- Walk, C.; Poernama, F. Evaluation of phytase, xylanase, and protease in reduced nutrient diets fed to broilers. Journal of Applied Poultry Research 2019, 28, 85–93. [Google Scholar] [CrossRef]
- Walk, C.; Santos, T.; Bedford, M. Influence of superdoses of a novel microbial phytase on growth performance, tibia ash, and gizzard phytate and inositol in young broilers. Poultry Science 2014, 93, 1172–1177. [Google Scholar] [CrossRef] [PubMed]
- Woyengo, T.; Slominski, B.; Jones, R. Growth performance and nutrient utilization of broiler chickens fed diets supplemented with phytase alone or in combination with citric acid and multicarbohydrase. Poultry Science 2010, 89, 2221–2229. [Google Scholar] [CrossRef]
- Zhang, Q.; Walk, C.L.; Sorbara, J.-O.B.; Cowieson, A.J.; Stamatopoulos, K. Comparative effects of two phytases on growth performance, bone mineralization, nutrient digestibility and phytate-P hydrolysis of broilers. Journal of Applied Poultry Research 2022, 3, 100247. [Google Scholar] [CrossRef]
- Egger, M.; Smith, G.D.; Schneider, M.; Minder, C. Bias in meta-analysis detected by a simple, graphical test. Bmj 1997, 315, 629–634. [Google Scholar] [CrossRef]
- Ioannidis, J.P.; Trikalinos, T.A. The appropriateness of asymmetry tests for publication bias in meta-analyses: a large survey. Cmaj 2007, 176, 1091–1096. [Google Scholar] [CrossRef] [PubMed]
- Duval, S.; Tweedie, R. A nonparametric “trim and fill” method of accounting for publication bias in meta-analysis. Journal of the american statistical association 2000, 95, 89–98. [Google Scholar]
- Council, N.R. Nutrient requirements of poultry: 1994, National Academies Press, 1994.
- Nuamah, E.; Poaty Ditengou, J.I.C.; Hirwa, F.; Cheon, I.; Chae, B.; Choi, N.-J. Dietary Supplementation of Tannins: Effect on Growth Performance, Serum Antioxidant Capacity, and Immunoglobins of Weaned Piglets—A Systematic Review with Meta-Analysis. Antioxidants 2024, 13, 236. [Google Scholar] [CrossRef] [PubMed]
- Borenstein, M.; Hedges, L.V.; Higgins, J.P.; Rothstein, H.R. Introduction to meta-analysis; John Wiley & Sons: 2011.
- Higgins, J.P.; Green, S. Cochrane handbook for systematic reviews of interventions. 2008.
- Macleod, M.R.; Fisher, M.; O’collins, V.; Sena, E.S.; Dirnagl, U.; Bath, P.M.; Buchan, A.; Van Der Worp, H.B.; Traystman, R.; Minematsu, K. Good laboratory practice: preventing introduction of bias at the bench. Stroke 2009, 40, e50–e52. [Google Scholar] [CrossRef] [PubMed]
- Kilkenny, C.; Parsons, N.; Kadyszewski, E.; Festing, M.F.; Cuthill, I.C.; Fry, D.; Hutton, J.; Altman, D.G. Survey of the quality of experimental design, statistical analysis and reporting of research using animals. PloS One 2009, 4, e7824. [Google Scholar] [CrossRef] [PubMed]
- Ncho, C.M.; Kim, S.-H.; Rang, S.A.; Lee, S.S. A Meta-analysis of probiotic interventions to mitigate ruminal methane emissions in cattle: implications for sustainable livestock farming. Animal 2024, 101180. [Google Scholar] [CrossRef] [PubMed]
- Dilger, R.; Onyango, E.; Sands, J.; Adeola, O. Evaluation of microbial phytase in broiler diets. Poultry Science 2004, 83, 962–970. [Google Scholar] [CrossRef] [PubMed]
- Onyango, E.; Bedford, M.; Adeola, O. Efficacy of an evolved Escherichia coli phytase in diets of broiler chicks. Poultry Science 2005, 84, 248–255. [Google Scholar] [CrossRef]
- Paiva, D.; Walk, C.; McElroy, A. Dietary calcium, phosphorus, and phytase effects on bird performance, intestinal morphology, mineral digestibility, and bone ash during a natural necrotic enteritis episode. Poultry Science 2014, 93, 2752–2762. [Google Scholar] [CrossRef]
- Proszkowiec-Weglarz, M.; Angel, R. Calcium and phosphorus metabolism in broilers: Effect of homeostatic mechanism on calcium and phosphorus digestibility. Journal of Applied Poultry Research 2013, 22, 609–627. [Google Scholar] [CrossRef]
- Olukosi, O.A.; Cowieson, A.J.; Adeola, O. Energy utilization and growth performance of broilers receiving diets supplemented with enzymes containing carbohydrase or phytase activity individually or in combination. British Journal of Nutrition 2008, 99, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Pirgozliev, V.; Bedford, M.; Acamovic, T.; Mares, P.; Allymehr, M. The effects of supplementary bacterial phytase on dietary energy and total tract amino acid digestibility when fed to young chickens. British Poultry Science 2011, 52, 245–254. [Google Scholar] [CrossRef]
- Akyurek, H.; Ozduven, M.L.; Okur, A.A.; Koc, F.; Samli, H.E. The effects of supplementing an organic acid blend and/or microbial phytase to a corn-soybean based diet fed to broiler chickens. African Journal of Agricultural Research 2011, 6, 642–649. [Google Scholar]
- Cowieson, A.; Wilcock, P.; Bedford, M. Super-dosing effects of phytase in poultry and other monogastrics. World’s Poultry Science Journal 2011, 67, 225–236. [Google Scholar] [CrossRef]
- Liu, N.; Ru, Y.; Cowieson, A.; Li, F.; Cheng, X.C. Effects of phytate and phytase on the performance and immune function of broilers fed nutritionally marginal diets. Poultry Science 2008, 87, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Tamim, N.; Angel, R.; Christman, M. Influence of dietary calcium and phytase on phytate phosphorus hydrolysis in broiler chickens. Poultry science 2004, 83, 1358–1367. [Google Scholar] [CrossRef]
- Gifford, S.R.; Clydesdale, F.M. Interactions among calcium, zinc and phytate with three protein sources. Journal of Food Science 1990, 55, 1720–1724. [Google Scholar] [CrossRef]
- Mohammed, A.; Gibney, M.; Taylor, T. The effects of dietary levels of inorganic phosphorus, calcium and cholecalciferol on the digestibility of phytate-P by the chick. British Journal of Nutrition 1991, 66, 251–259. [Google Scholar] [CrossRef]
- Scheideler, S.E.; Sell, J. Utilization of phytate phosphorus in laying hens as influenced by dietary phosphorus and calcium. 1987.
- Singh, P. Significance of phytic acid and supplemental phytase in chicken nutrition: a review. World’s Poultry Science Journal 2008, 64, 553–580. [Google Scholar] [CrossRef]
- Santos, F.; Hruby, M.; Pierson, E.; Remus, J.; Sakomura, N. Effect of phytase supplementation in diets on nutrient digestibility and performance in broiler chicks. Journal of Applied Poultry Research 2008, 17, 191–201. [Google Scholar] [CrossRef]
- Viveros, A.; Brenes, A.; Arija, I.; Centeno, C. Effects of microbial phytase supplementation on mineral utilization and serum enzyme activities in broiler chicks fed different levels of phosphorus. Poultry Science 2002, 81, 1172–1183. [Google Scholar] [CrossRef] [PubMed]
- Watson, B.; Matthews, J.; Southern, L.; Shelton, J. The effects of phytase on growth performance and intestinal transit time of broilers fed nutritionally adequate diets and diets deficient in calcium and phosphorus. Poultry science 2006, 85, 493–497. [Google Scholar] [CrossRef]
- Selle, P.; Ravindran, V.; Caldwell, A.; Bryden, W. Phytate and phytase: consequences for protein utilisation. Nutrition Research Reviews 2000, 13, 255–278. [Google Scholar] [CrossRef]
- Amerah, A.; Plumstead, P.; Barnard, L.; Kumar, A. Effect of calcium level and phytase addition on ileal phytate degradation and amino acid digestibility of broilers fed corn-based diets. Poultry Science 2014, 93, 906–915. [Google Scholar] [CrossRef] [PubMed]
- Sibbald, I.R.; Wolynetz, M.S. Comparison of three methods of excreta collection used in estimation of energy and nitrogen excretion. Poultry Science 1986, 65, 78–84. [Google Scholar] [CrossRef]
- Humer, E.; Schwarz, C.; Schedle, K. Phytate in pig and poultry nutrition. Journal of animal physiology and animal nutrition 2015, 99, 605–625. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hack, M.E.; Alagawany, M.; Arif, M.; Emam, M.; Saeed, M.; Arain, M.A.; Siyal, F.A.; Patra, A.; Elnesr, S.S.; Khan, R.U. The uses of microbial phytase as a feed additive in poultry nutrition–a review. Annals of Animal Science 2018, 18, 639–658. [Google Scholar] [CrossRef]
- Huber, K.; Zeller, E.; Rodehutscord, M. Modulation of small intestinal phosphate transporter by dietary supplements of mineral phosphorus and phytase in broilers. Poultry Science 2015, 94, 1009–1017. [Google Scholar] [CrossRef]
- Lee, S.; Bedford, M. Inositol-An effective growth promotor? World’s Poultry Science Journal 2016, 72, 743–760. [Google Scholar] [CrossRef]
- El Enshasy, H.; Dailin, D.J.; Abd Manas, N.H.; Azlee, N.İ.W.; Eyahmalay, J.; Yahaya, S.A.; Abd Malek, R.; Siwapiragam, V.; Sukmawati, D. Current and future applications of phytases in poultry industry: A critical review. Journal of Advances in VetBio Science and Techniques 2018, 3, 65–74. [Google Scholar] [CrossRef]
- Edwards, H.M., Jr.; Palo, P.; Sooncharernying, S.; Elliot, M. Factors influencing the bioavailability of phytate phosphorus to chickens. Nutrient availability: chemical and biological aspects 1989, 271–276. [Google Scholar]
- Peeler, H. Biological availability of nutrients in feeds: availability of major mineral ions. Journal of Animal Science 1972, 35, 695–712. [Google Scholar] [CrossRef] [PubMed]
- Abudabos, A.M. Intestinal phytase activity in chickens (Gallus Domesticus). Afr. J. Microbiol. Res 2012, 6. [Google Scholar]
- Sebola, N.; Mlambo, V.; Mokoboki, H.; Muchenje, V. Growth performance and carcass characteristics of three chicken strains in response to incremental levels of dietary Moringa oleifera leaf meal. Livestock Science 2015, 178, 202–208. [Google Scholar] [CrossRef]
- Ravindran, V.; Bryden, W.; Kornegay, E. Phytates: Occurrence, bioavailability and implications in poultry nutrition. 1995.
- Ravindran, V.; Selle, P.; Bryden, W. Effects of phytase supplementation, individually and in combination, with glycanase, on the nutritive value of wheat and barley. Poultry Science 1999, 78, 1588–1595. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Angel, R.; Kim, S.-W.; Jiménez-Moreno, E.; Proszkowiec-Weglarz, M.; Plumstead, P. Age and adaptation to Ca and P deficiencies: 2. Impacts on amino acid digestibility and phytase efficacy in broilers. Poultry Science 2015, 94, 2917–2931. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Angel, R.; Kim, S.-W.; Jiménez-Moreno, E.; Proszkowiec-Weglarz, M.; Plumstead, P. Impacts of age and calcium on phytase efficacy in broiler chickens. Animal Feed Science and Technology 2018, 238, 9–17. [Google Scholar] [CrossRef]
- Yan, F.; Angel, R.; Ashwell, C.; Mitchell, A.; Christman, M. Evaluation of the broiler’s ability to adapt to an early moderate deficiency of phosphorus and calcium. Poultry Science 2005, 84, 1232–1241. [Google Scholar] [CrossRef]
- Shastak, Y.; Rodehutscord, M. Determination and estimation of phosphorus availability in growing poultry and their historical development. World’s Poultry Science Journal 2013, 69, 569–586. [Google Scholar] [CrossRef]
- Pereira, R.; Menten, J.; Romano, G.; Silva, C.; Zavarize, K.; Barbosa, N. Efficiency of a bacterial phytase to release phytate phosphorus in broiler chicken diets. 2012.
- Gautier, A.; Walk, C.; Dilger, R. Effects of a high level of phytase on broiler performance, bone ash, phosphorus utilization, and phytate dephosphorylation to inositol. Poultry Science 2018, 97, 211–218. [Google Scholar] [CrossRef]
- Moss, A.F.; Chrystal, P.; Dersjant-Li, Y.; Liu, S.; Selle, P. The ranked importance of dietary factors influencing the performance of broiler chickens offered phytase-supplemented diets by the Plackett-Burman screening design. British Poultry Science 2019, 60, 439–448. [Google Scholar] [CrossRef] [PubMed]
- Sacakli, P.; Sehu, A.; Ergun, A.; Genc, B.; Selcuk, Z. The effect of phytase and organic acid on growth performance, carcass yield and tibia ash in quails fed diets with low levels of non-phytate phosphorus. Asian-Australasian Journal Of Animal Sciences 2006, 19, 198–202. [Google Scholar] [CrossRef]
- Karimi, A.; Bedford, M.; Sadeghi, G.; Ghobadi, Z. Influence of dietary non-phytate phosphorous levels and phytase supplementation on the performance and bone characteristics of broilers. Brazilian Journal of Poultry Science 2011, 13, 43–51. [Google Scholar] [CrossRef]
- Talaty, P.; Katanbaf, M.; Hester, P. Life cycle changes in bone mineralization and bone size traits of commercial broilers. Poultry Science 2009, 88, 1070–1077. [Google Scholar] [CrossRef] [PubMed]
- WPSA. Mineral requirements for poultry- mineral requirements and recommendations for growing birds. World’s Poultry Science Journal 1985, 41, 252–258. [Google Scholar] [CrossRef]
- Rath, N.; Huff, G.; Huff, W.; Balog, J. Factors regulating bone maturity and strength in poultry. Poultry Science 2000, 79, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Olukosi, O.; Kong, C.; Fru-Nji, F.; Ajuwon, K.; Adeola, O. Assessment of a bacterial 6-phytase in the diets of broiler chickens. Poultry Science 2013, 92, 2101–2108. [Google Scholar] [CrossRef]
- Fernandes, J.; Horn, D.; Ronconi, E.; Buzim, R.; Lima, F.; Pazdiora, D. Effects of phytase superdosing on digestibility and bone integrity of broilers. Journal of Applied Poultry Research 2019, 28, 390–398. [Google Scholar] [CrossRef]
Figure 1.
Systematic literature search and selection process. The PRISMA diagram details the applied search and selection process of this study.
Figure 1.
Systematic literature search and selection process. The PRISMA diagram details the applied search and selection process of this study.
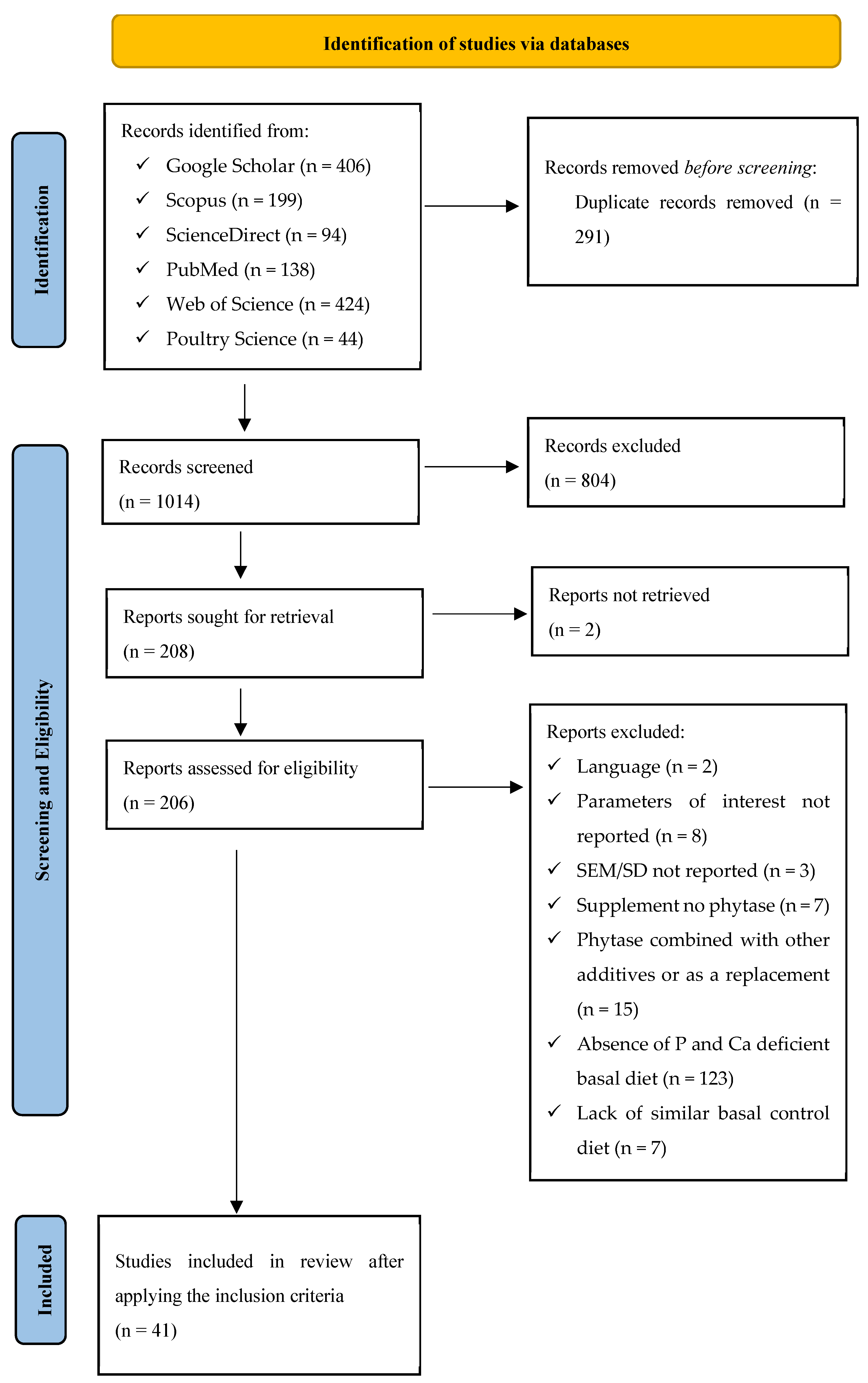
Figure 2.
A bar chart of the risk of bias classification of the included studies based on the Cochrane Collaboration’s SYRCLE Risk of Bias tool for animal studies.
Figure 2.
A bar chart of the risk of bias classification of the included studies based on the Cochrane Collaboration’s SYRCLE Risk of Bias tool for animal studies.
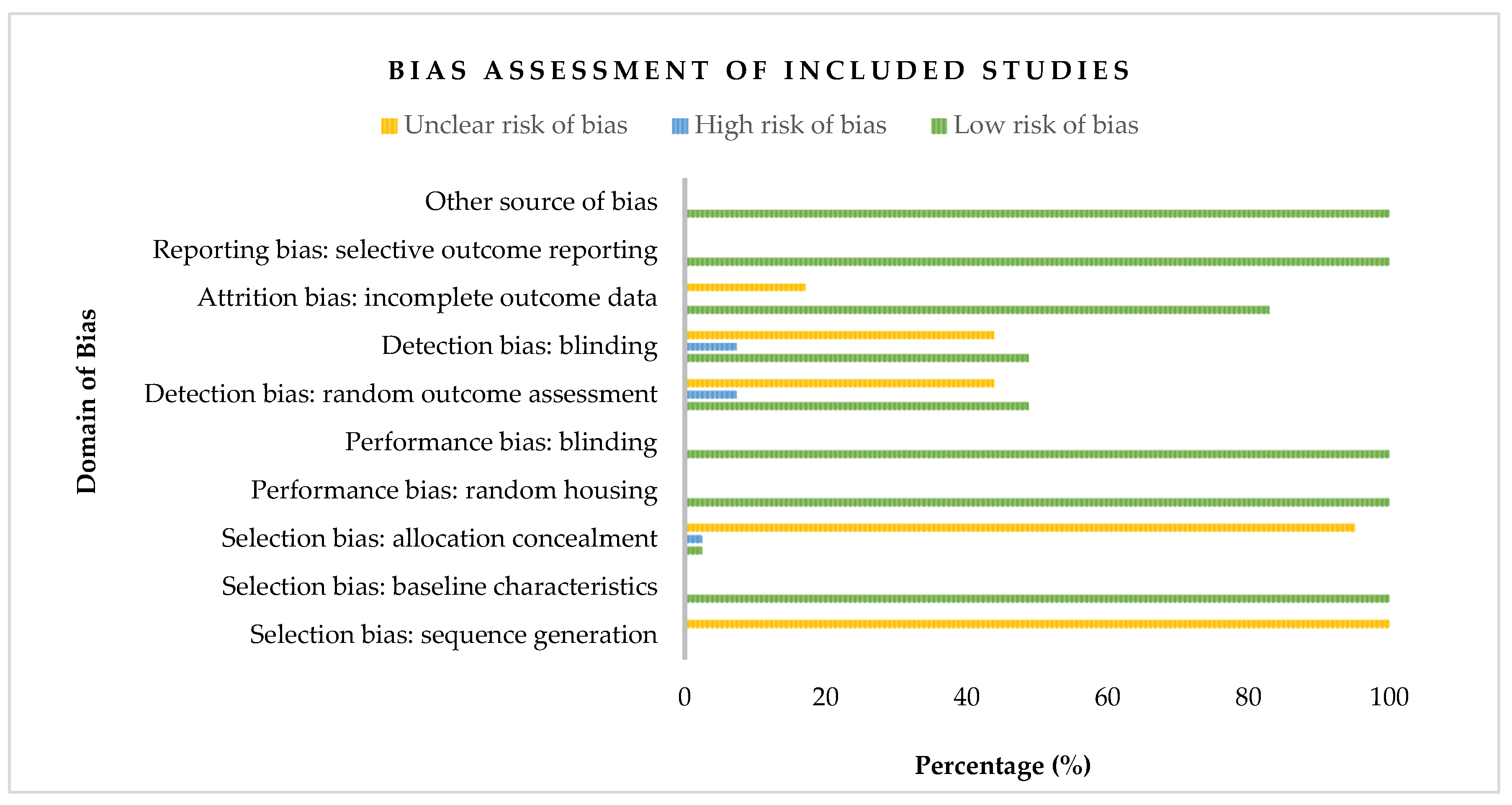
Figure 3.
A decreasing trend of tibia P’s standardized mean difference per unit increase of phytase dosage.
Figure 3.
A decreasing trend of tibia P’s standardized mean difference per unit increase of phytase dosage.
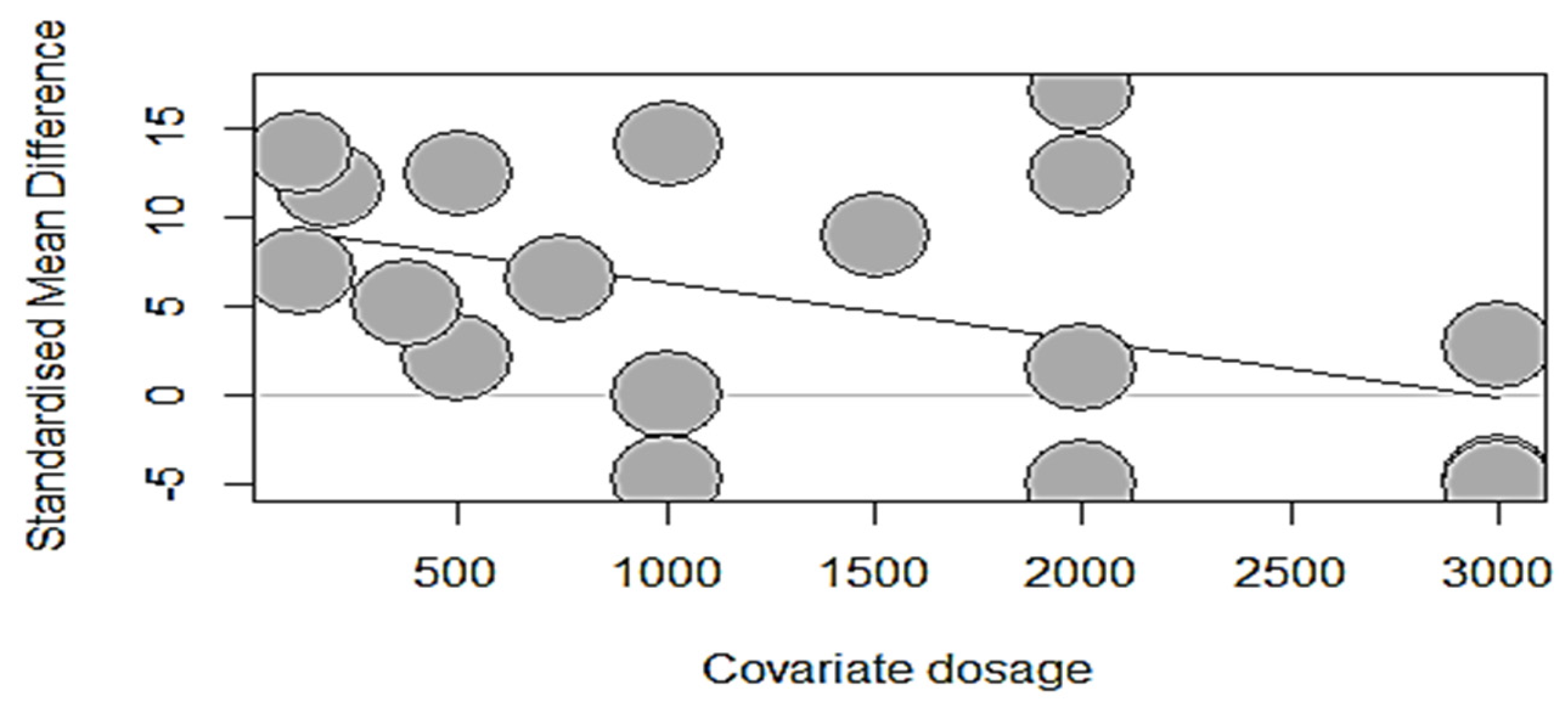
Figure 4.
Showing bubble plot of standardized mean difference of (a) starter FCR; (b) starter tibia Ca; starter tibia P; (d) grower-finisher tibia Ca; (e) grower-finisher tibia P associated with a unit change in duration of phytase exposure.
Figure 4.
Showing bubble plot of standardized mean difference of (a) starter FCR; (b) starter tibia Ca; starter tibia P; (d) grower-finisher tibia Ca; (e) grower-finisher tibia P associated with a unit change in duration of phytase exposure.
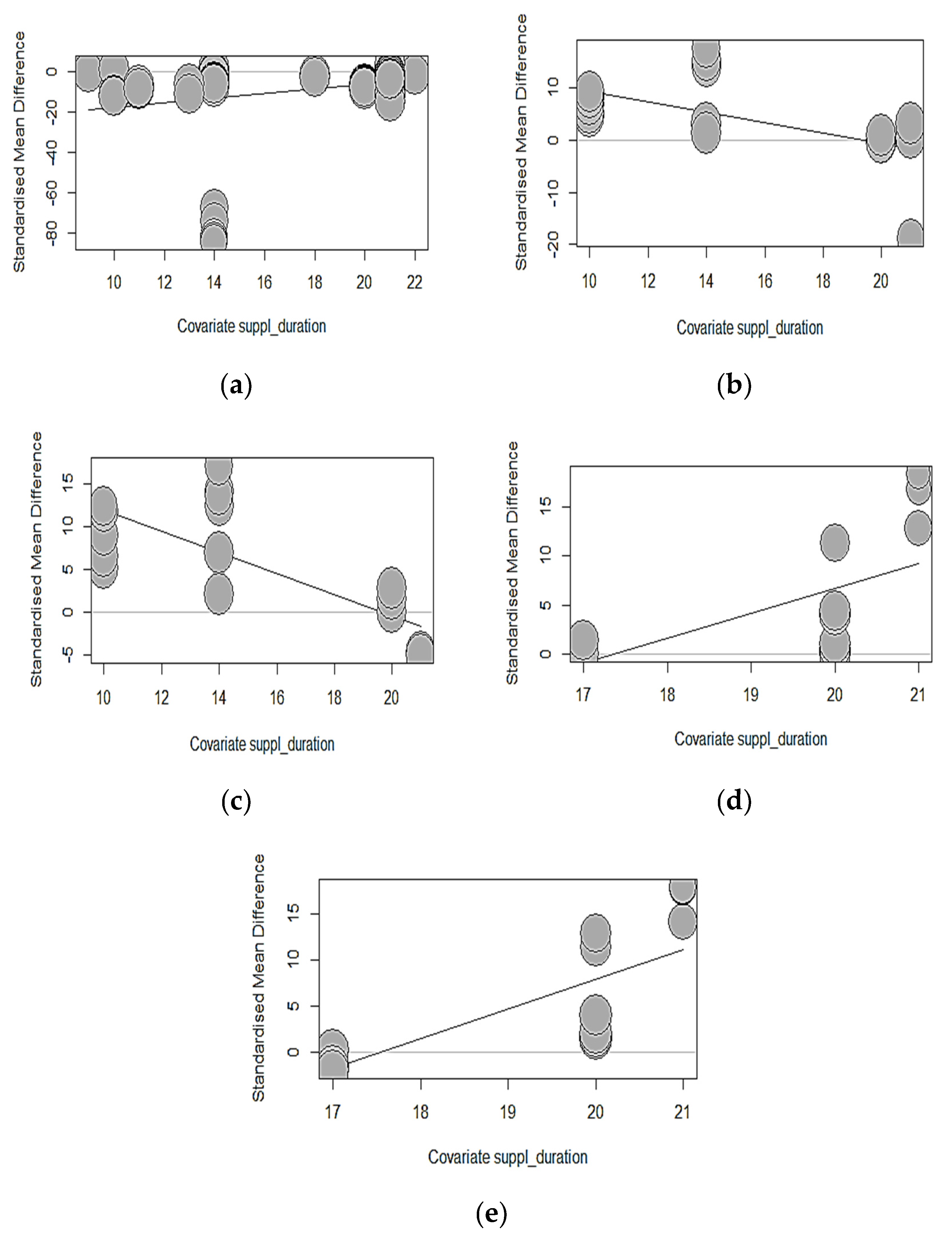
Table 2.
Descriptive statistics of the basal diet’s calculated composition, phytase dosage, and supplementation duration.
Table 2.
Descriptive statistics of the basal diet’s calculated composition, phytase dosage, and supplementation duration.
| Starter Phase (1-22days) | Grower-Finisher Phase (22-42) | |||||||
|---|---|---|---|---|---|---|---|---|
| Variable | Mean | SEM | Minimum | Maximum | Mean | SEM | Minimum | Maximum |
| Crude Protein, % of DM | 21.91 | 0.22 | 19.40 | 26.33 | 20.48 | 0.47 | 17.80 | 23.00 |
| Metabolizable energy (MJ/kg) | 12.44 | 0.24 | 11.97 | 14.10 | 12.82 | 0.15 | 12.18 | 13.34 |
| Digestible Met + Cys, % of DM | 0.92 | 0.02 | 0.77 | 1.19 | 0.80 | 0.05 | 0.65 | 0.86 |
| Digestible Lysine, % of DM | 1.11 | 0.05 | 0.10 | 1.42 | 0.81 | 0.19 | 0.10 | 1.19 |
| Digestible Thr, % of DM | 0.78 | 0.01 | 0.63 | 0.89 | 0.71 | 0.03 | 0.6175 | 0.76 |
| Digestible Val, % of DM | 0.93 | 0.04 | 0.77 | 1.30 | 0.86 | 0.10 | 0.76 | 1.05 |
| Available Phosphorus, % of DM | 0.26 | 0.01 | 0.10 | 0.35 | 0.21 | 0.03 | 0.17 | 0.29 |
| Total P, % of DM | 0.53 | 0.01 | 0.36 | 0.65 | 0.47 | 0.04 | 0.41 | 0.60 |
| Calcium, % of DM | 0.74 | 0.02 | 0.40 | 0.95 | 0.64 | 0.04 | 0.47 | 0.74 |
| NPP, % of DM | 0.30 | 0.02 | 0.20 | 0.43 | 0.30 | 0.04 | 0.21 | 0.42 |
| Phytase dosage (FTU per Kg of diet) | 1709 | 397 | 120 | 40500 | 876 | 153 | 250 | 5000 |
| Supplementation duration, day | 16 | 0.68 | 6.00 | 22.00 | 20 | 1.48 | 10.00 | 33.00 |
Table 3.
Growth performance of starter and grower meat-type chickens supplemented with phytase.
| Random Effects Model | Heterogeneity | Egger’s Testb | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Parameter | n(NC) | SMD | 95% CI | p-Value | Chi-squared (Q) | I2 (%) | τ2 | p-Valuea | p-Value |
| Starter Phase (1-22 days) | |||||||||
| ADFI, g/day | 38(97) | 12.57 | 6.61, 18.54 | < 0.0001 | 25654.46 | 99.6 | 895.47 | 0.000 | < 0.0001 |
| ADG, g/day | 6(13) | 11.31 | 4.85, 17.76 | 0.0006 | 22789.26 | 99.8 | 140.72 | 0.000 | < 0.0001 |
| BWG, g | 38(96) | 10.76 | 8.13, 13.78 | < 0.0001 | 22789.26 | 99.6 | 171.11 | 0.000 | < 0.0001 |
| FCR, g/g | (90) | -8.99 | -12.59, -5.39 | < 0.0001 | 18235.57 | 99.5 | 300.91 | 0.000 | < 0.0001 |
| Grower-Finisher Phase (22-42 days) | |||||||||
| ADFI, g/day | (26) | 47.35 | 10.27, 84.42 | 0.0123 | 7353.11 | 99.7 | 9285.23 | 0.000 | < 0.0001 |
| ADG, g/day | 3(9) | 7.78 | -0.63, 16.19 | 0.0699 | 2676.45 | 99.7 | 165.52 | 0.000 | < 0.0001 |
| BWG, g | 11(22) | 7.67 | 3.62, 11.73 | 0.0002 | 5297.35 | 99.6 | 93.53 | 0.000 | 0.0118 |
| FCR, g/g | (24) | -4.30 | -6.38, -2.21 | < 0.0001 | 4387.11 | 99.5 | 26.65 | 0.000 | 0.0030 |
n: number of studies; NC: the number of comparisons between exogenous phytase treatment and deficient Ca and P control group; SMD: standardized mean difference; CI: confidence interval; Q: X2 statistics; I2: proportion of total variation of effect size estimate due to heterogeneity; τ2: heterogeneity variance of the actual effect. (Forest plots showing mean difference effects are presented in S1-8). a p-Value to X2 statistics (Q) test of heterogeneity. b Egger’s regression asymmetry test.
Table 4.
Bone strength and mineralization of starter and grower meat-type chickens supplemented with phytase.
Table 4.
Bone strength and mineralization of starter and grower meat-type chickens supplemented with phytase.
| Random Effects Model | Heterogeneity | Egger’s Testb | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Parameter | n(NC) | SMD | 95% CI | p-Value | Chi-squared (Q) | I2 (%) | τ2 | p-Valuea | p-Value |
| Starter Phase (1-22 days) | |||||||||
| BBS, Kgf/mm | 4(17) | 4.68 | 2.81, 6.54 | < 0.0001 | 225.00 | 92.9 | 14.41 | < 0.0001 | < 0.0001 |
| Tibia Ash, g/Kg DM | 16(51) | 11.78 | 8.65, 14.92 | < 0.0001 | 7144.72 | 99.3 | 125.08 | 0.000 | < 0.0001 |
| Tibia Ca, g/Kg DM | 4(17) | 4.35 | 0.64, 8.06 | 0.0217 | 550.00 | 97.1 | 59.77 | < 0.0001 | 0.0030 |
| Tibia P, g/Kg DM | 4(18) | 5.26 | 1.87, 8.65 | 0.0024 | 862.39 | 98.0 | 52.72 | < 0.0001 | 0.0089 |
| Grower-Finisher Phase (22-42 days) | |||||||||
| BBS, Kgf/mm | 3(11) | 4.33 | 2.59, 6.08 | < 0.0001 | 191.72 | 94.8 | 7.92 | < 0.0001 | 0.0013 |
| Tibia Ash, g/Kg DM | 6(21) | 5.88 | 3.13, 8.62 | < 0.0001 | 716.64 | 97.2 | 39.13 | < 0.0001 | 0.0005 |
| Tibia Ca, g/Kg DM | 4(13) | 4.94 | 1.51, 8.36 | 0.0047 | 179.30 | 93.3 | 37.63 | < 0.0001 | < 0.0001 |
| Tibia P, g/Kg DM | 4(13) | 5.72 | 1.86, 9.57 | 0.0036 | 229.97 | 94.8 | 47.75 | < 0.0001 | 0.0002 |
n: number of studies; NC: the number of comparisons between exogenous phytase treatment and deficient Ca and P control group; SMD: standardized mean difference; CI: confidence interval; Q: X2 statistics; I2: proportion of total variation of effect size estimate due to heterogeneity; τ2: heterogeneity variance of the actual effect; BBS: bone breaking strength. (Forest plots showing mean difference effects are presented in S9-16). a p-Value to X2 statistics (Q) test of heterogeneity. b Egger’s regression asymmetry test.
Table 5.
Corrected publication bias of phytase effect on broilers’ performance, bone strength, and mineralization.
Table 5.
Corrected publication bias of phytase effect on broilers’ performance, bone strength, and mineralization.
| Random Effects Model | |||||
|---|---|---|---|---|---|
| Parameter | NC | d.f. | SMDb | 95% CI | p-Valuec |
| Starter Phase (1-22 days) | |||||
| ADFI, g/d | 131 | 130 | 1.6779 | -5.04, 8.40 | 0.6244 |
| ADG, g/d | 18 | 17 | 1.8716 | -6.79, 10.53 | 0.6718 |
| BWG, g | 137 | 136 | 3.3592 | 0.31, 6.41 | 0.0307 |
| FCR, g/g | 124 | 123 | -2.0180 | -6.15, 2.11 | 0.3383 |
| BBS, Kgf/mm | 24 | 23 | 1.8186 | -0.59, 4.23 | 0.1387 |
| Tibia Ash, g/Kg DM | 74 | 73 | 2.0083 | -2.63, 6.65 | 0.3959 |
| Tibia Ca, g/Kg DM | 22 | 21 | 1.0343 | -2.93, 5.00 | 0.6093 |
| Tibia P, g/Kg DM | 24 | 23 | 0.9147 | -3.12, 4.95 | 0.6568 |
| Grower-Finisher Phase (22-42 days) | |||||
| ADFI, g/d | 35 | 34 | 1.1746 | -42.28, 44.62 | 0.9577 |
| ADG, g/d | 11 | 10 | 1.4517 | -9.31, 12.22 | 0.7916 |
| BWG, g | 29 | 28 | 2.0851 | -2.88, 7.05 | 0.4104 |
| FCR, g/g | 30 | 29 | -1.6657 | -4.26, 0.92 | -1.6657 |
| BBS, Kgf/mm | 16 | 15 | 2.2862 | 0.19, 4.38 | 0.0324 |
| Tibia Ash, g/Kg DM | 28 | 27 | 2.1719 | -1.74, 6.08 | 0.2761 |
| Tibia Ca, g/Kg DM | 16 | 15 | 1.7751 | -2.89, 6.44 | 0.4555 |
| Tibia P, g/Kg DM | 18 | 17 | 0.6856 | -4.28, 5.65 | 0.7868 |
NC: the number of comparisons between exogenous phytase treatment and deficient Ca and P control group; CI: confidence interval; d.f: degree of freedom. b standardized mean difference after adjustment for funnel plot asymmetry by the trim-and-fill method (random effects). c p-Value of effect estimate after adjustment for funnel plot asymmetry by the trim-and-fill method (random effects).
Table 6.
Meta-ANOVA of broiler strain association with measured growth outcomes.
| Random Effects Model | ||||||||
|---|---|---|---|---|---|---|---|---|
| Parameter | Covariate: Strain | NC | I2, % | SMD (95% CI) | τ2 | d.f | Q | p-Valuea |
| Starter Phase (1-22 days) | ||||||||
| ADFI, g/d | Ross 308 | 41 | 99.4 | 5.61 (3.67, 7.55) | 39.71 | 10 | 737.59 | < 0.0001 |
| Ross 708 | 6 | 98.8 | 1.35 (0.60, 2.09) | 0.86 | ||||
| Cobb 500 | 19 | 99.6 | 11.54 (5.09, 18.00) | 205.46 | ||||
| Hubbard × Cobb 500 | 4 | 83.4 | 151.73 (138.71, 164.75) | 146.01 | ||||
| Ross | 10 | 99.4 | 1.83 (-1.36, 5.02) | 26.32 | ||||
| Ross × Ross 708 | 5 | 99.6 | 1.70 (-1.51, 4.92) | 13.45 | ||||
| Ross 708 × YPM | 3 | 98.8 | 35.12 (23.05, 47.31) | 113.55 | ||||
| Yield Plus × Ross 708 | 5 | 99.6 | 2.56 (0.90, 4.23) | 3.60 | ||||
| Lohman Indian RSR | 2 | 96.1 | 11.54 (9.48, 13.60) | 2.12 | ||||
| ADG, g/d | Ross 308 | 6 | 99.9 | 15.52 (5.14, 25.91) | 168.09 | 3 | 264.88 | < 0.0001 |
| Cobb 500 | 3 | 89.6 | 18.65 (16.23, 21.06) | 4.10 | ||||
| Ross | 3 | 99.4 | -0.31 (-3.06, 2.43) | 5.85 | ||||
| BWG, g | Ross 308 | 43 | 99.4 | 7.67 (5.26, 9.72) | 46.42 | 10 | 1122.71 | < 0.0001 |
| Ross 708 | 6 | 96.0 | 7.69 (6.56, 8.81) | 1.90 | ||||
| Cobb 500 | 19 | 99.6 | 19.36 (8.12, 30.60) | 624.18 | ||||
| Hubbard × Cobb 500 | 4 | 83.4 | 20.70 (18.89, 22.52) | 2.86 | ||||
| Ross | 7 | 99.0 | 16.13 (11.23, 21.03) | 42.04 | ||||
| Ross × Ross 708 | 5 | 99.8 | 5.49 (-1.79, 12.77) | 68.80 | ||||
| Ross 708 × YPM | 3 | 97.4 | 2.81 (1.90, 3.73) | 0.63 | ||||
| Yield Plus × Ross 708 | 5 | 99.4 | 5.58 (3.42, 7.74) | 6.02 | ||||
| Lohman Indian RSR | 2 | 94.3 | 13.41 (11.31, 15.50) | 2.16 | ||||
| FCR, g/g | Ross 308 | 38 | 99.1 | -4.29 (-5.38, -3.21) | 11.37 | 10 | 1434.92 | < 0.0001 |
| Ross 708 | 6 | 82.8 | -7.68 (-8.22, -7.14) | 0.39 | ||||
| Cobb 500 | 15 | 99.4 | -4.84 (-6.92, -2.75) | 16.97 | ||||
| Hubbard × Cobb 500 | 4 | 82.2 | -7.18 (-7.83, -6.53) | 0.37 | ||||
| Ross | 10 | 99.6 | -48.69 (-72.95, -24.43) | 1511.60 | ||||
| Ross × Ross 708 | 5 | 99.6 | -2.66 (-5.84, 0.53) | 13.20 | ||||
| Ross 708 × YPM | 3 | 99.2 | 0.37 (-0.90, 1.65) | 1.26 | ||||
| Yield Plus × Ross 708 | 5 | 98.0 | -4.06 (-5.06, -3.06) | 1.27 | ||||
| Lohman Indian RSR | 2 | 99.3 | -3.19 (-5.15, -1.24) | 1.98 | ||||
| Grower-Finisher Phase (22-42 days) | ||||||||
| ADFI, g/d | Cobb 500 | 8 | 99.7 | 139.58 (43.64, 235.50) | 19111.51 | 3 | 13.11 | 0.0044 |
| Ross 308 | 12 | 99.0 | 10.46 (0.16, 20.76) | 328.99 | ||||
| Ross | 5 | 94.8 | -1.11 (-1.92, -0.29) | 0.80 | ||||
| ADG, g/d | Cobb 500 | 3 | 64.7 | 24.91 (23.24, 26.57) | 1.39 | 2 | 975.23 | < 0.0001 |
| Ross | 5 | 95.1 | -0.52 (-120, 0.16) | 0.54 | ||||
| BWG, g | Cobb 500 | 6 | 99.3 | 19.74 (13.07, 26.41) | 68.90 | 3 | 25.56 | < 0.0001 |
| Ross 308 | 11 | 99.4 | 4.16 (0.56, 7.76) | 36.26 | ||||
| Ross | 3 | 99.7 | 3.28 (-273.99, 842.31) | 242927.9 | ||||
| FCR, g/g | Cobb 500 | 6 | 99.7 | -8.39 (-11.19, -5.59) | 12.15 | 3 | 442.11 | < 0.0001 |
| Ross 308 | 12 | 98.7 | -4.72 (-7.63, -1.80) | 25.67 | ||||
| Ross | 5 | 17.4 | -0.13 (4.51, 5.46) | 0.0044 | ||||
NC: the number of comparisons between exogenous phytase treatment and deficient Ca and P control group; I2: proportion of total variation of the common effect size estimate due to heterogeneity; SMD: standardized mean difference; CI: confidence interval; Q: X2 statistics; d.f: degree of freedom; τ2: heterogeneity variance of the actual effect. a p-Value to test for subgroup differences (random effect).
Table 7.
Meta-ANOVA of broiler strain association with measured bone outcomes.
| Random Effects Model | ||||||||
|---|---|---|---|---|---|---|---|---|
| Parameter | Covariate: Strain | NC | I2, % | SMD (95% CI) | τ2 | d.f | Q | p-Valuea |
| Starter Phase (1-22 days) | ||||||||
| BBS, Kgf/mm | Cobb 500 | 10 | 93.5 | 4.12 (1.63, 6.60) | 15.31 | 1 | 0.50 | 0.4776 |
| Ross 308 | 7 | 91.1 | 5.52 (2.58, 8.46) | 14.56 | ||||
| Tibia Ash, g/Kg DM | Ross 708 | 3 | 89.2 | 4.11 (2.42, 5.79) | 1.92 | 5 | 20.68 | 0.0009 |
| Cobb 500 | 25 | 99.4 | 11.66 (7.84, 15.48) | 93.25 | ||||
| Ross × Ross 708 | 3 | 99.9 | 5.02 (-8.25, 18.28) | 137.28 | ||||
| Ross 708 × YPM | 3 | 0.0 | 4.88 (2.83, 6.92) | 0.00 | ||||
| Ross 308 | 16 | 95.5 | 19.00 (9.39, 28.62) | 366.62 | ||||
| Tibia Ca, g/Kg DM | Cobb 500 | 12 | 96.5 | 1.84 (-2.00, 5.68) | 45.13 | 1 | 5.38 | 0.0204 |
| Ross 308 | 5 | 97.3 | 10.50 (4.27, 16.72) | 48.66 | ||||
| Tibia P, g/Kg DM | Cobb 500 | 13 | 97.8 | 2.39 (-0.99, 5.77) | 37.91 | 1 | 17.67 | < 0.0001 |
| Ross 308 | 5 | 90.4 | 12.72 (9.29, 16.15) | 13.14 | ||||
| Grower-Finisher Phase (22-42 days) | ||||||||
| BBS, Kgf/mm | Ross 308 | 8 | 96.2 | 4.88 (2.49, 7.27) | 10.91 | 1 | 1.73 | 0.19 |
| Cobb 500 | 3 | 68.7 | 3.07 (1.83, 4.32) | |||||
| Tibia Ash, g/Kg DM | Ross 308 | 7 | 93.8 | 2.38 (-0.13, 4.89) | 10.87 | 3 | 68.22 | < 0.0001 |
| Ross 708 | 3 | 65.7 | 10.87 (8.60, 13.14) | 2.58 | ||||
| Cobb 500 | 6 | 91.3 | 13.00 (3.53, 22.46) | 133.92 | ||||
| Ross | 5 | 49.4 | 1.22 (0.51, 1.94) | 0.32 | ||||
| Tibia Ca, g/Kg DM | Ross 308 | 5 | 88.3 | 9.20 (1.27, 17.13) | 77.12 | 3 | 10.84 | 0.0044 |
| Cobb 500 | 3 | 27.5 | 6.28 (1.87, 10.70) | 14.05 | ||||
| Ross | 5 | 95.6 | 0.55 (0.02, 1.09) | 0.10 | ||||
| Tibia P, g/Kg DM | Cobb 500 | 3 | 93.6 | 9.20 (3.70, 14.69) | 21.37 | 2 | 13.41 | 0.0012 |
| Ross | 5 | 89.4 | 0.32 (-1.33, 1.97) | 3.17 | ||||
| Ross 308 | 5 | 95.5 | 9.61 (1.59, 17.62) | 78.58 | ||||
NC: the number of comparisons between exogenous phytase treatment and deficient Ca and P control group; I2: proportion of total variation of the common effect size estimate due to heterogeneity; SMD: standardized mean difference; CI: confidence interval; Q: X2 statistics; d.f: degree of freedom; τ2: heterogeneity variance of the actual effect. a p-Value to test for subgroup differences (random effects).
Table 8.
Meta-ANOVA of basal diet’s dietary P source association with measured growth outcomes.
| Random Effects Model | ||||||||
|---|---|---|---|---|---|---|---|---|
| Parameter | Covariate: Dietary P Sourcea | NC | I2, % | SMD (95% CI) | τ2 | d.f | Q | p-Valueb |
| Starter Phase (1-22 days) | ||||||||
| ADFI, g/d | Corn-Soybean | 69 | 99.6 | 15.33 (7.03, 23.62) | 1233.91 | 5 | 18.52 | 0.0024 |
| Wheat-Corn-Soybean | 9 | 99.6 | 1.86 (-0.24, 0.03) | 2249.25 | ||||
| CSRRB | 2 | 99.2 | 6.56 (3.27, 9.86) | 5.44 | ||||
| Corn-Soybean-DDGS | 3 | 98.9 | 8.99 (2.54, 15.45) | 31.98 | ||||
| Wheat-Soybean | 11 | 99.8 | 10.10 (4.92, 15.28) | 76.63 | ||||
| WRES | 3 | 95.9 | -0.32 (-4.88, 4.24) | 15.68 | ||||
| ADG, g/d | Corn-Soybean | 6 | 99.8 | 9.31 (1.03, 17.59) | 106.92 | 2 | 36.83 | < 0.0001 |
| Wheat-Corn-Soybean | 3 | 99.4 | -0.31 (-3.06, 2.43) | 5.86 | ||||
| Wheat-Soybean | 4 | 99.4 | 23.02 (15.74, 30.31) | 54.77 | ||||
| BWG, g | Corn-Soybean | 72 | 99.6 | 11.35 (7.99, 14.70) | 209.70 | 5 | 5.89 | 0.3167 |
| Wheat-Corn-Soybean | 6 | 99.5 | 6.97 (2.06, 11.88) | 37.48 | ||||
| CSRRB | 2 | 90.5 | 8.61 (6.00, 11.23) | 3.23 | ||||
| Corn-Soybean-DDGS | 3 | 70.9 | 8.17 (6.84,9.51) | 0.98 | ||||
| Wheat-Soybean | 9 | 99.8 | 11.79 (4.20, 19.37) | 134.58 | ||||
| WRES | 4 | 92.8 | 6.40 (3.38,9.41) | 8.25 | ||||
| FCR, g/g | Corn-Soybean | 63 | 99.5 | -10.79 (-16.00, 5.58) | 442.82 | 6 | 488.97 | < 0.0001 |
| Wheat-Corn-Soybean | 9 | 99.6 | -4.35 (-7.80, -0.91) | 27.58 | ||||
| CSRRB | 2 | 0.0 | -12.04 (-13.12, -10.95) | 0 | ||||
| Corn-Soybean-DDGS | 3 | 78.9 | -3.31 (-4.08, -2.54) | 0.36 | ||||
| Wheat-Soybean | 7 | 99.5 | -4.85 (-7.07, -2.64) | 8.91 | ||||
| WRES | 4 | 73.8 | -8.21 (-10.89, -5.54) | 5.41 | ||||
| Corn-Soybean | 2 | 0.0 | -0.50 (-0.75, -0.24) | 0 | ||||
| Grower-Finisher Phase (22-42 days) | ||||||||
| ADFI, g/d | Corn-Soybean | 13 | 99.8 | 86.28 (17.50, 155.05) | 15976.48 | 3 | 30.93 | < 0.0001 |
| Wheat-Corn-Soybean | 6 | 93.5 | -1.10 (-1.76, -0.44) | 0.62 | ||||
| Corn-Soybean-DDGS | 3 | 96.9 | 39.35 (23.13, 55.57) | 196.29 | ||||
| WRES | 4 | 95.1 | 0.62 (-2.75, 3.99) | 11.36 | ||||
| ADG, g/d | Corn-Soyabean | 4 | 99.9 | 18.18 (4.89, 31.47) | 183.31 | 1 | 7.59 | 0.0059 |
| Wheat-Corn-Soybean | 5 | 95.1 | -0.52 (-120, 0.16) | 0.54 | ||||
| BWG, g | Corn-Soybean | 11 | 99.8 | 11.61 (5.00, 18.22) | 124.72 | 3 | 19.30 | 0.0002 |
| Wheat-Corn-Soybean | 4 | 98.9 | -2.38 (-6.20, 1.43) | 15.02 | ||||
| Corn-Soybean-DDGS | 3 | 98.5 | 7.91 (2.41, 13.42) | 23.19 | ||||
| WRES | 4 | 90.1 | 6.43 (2.59, 10.26) | 13.61 | ||||
| FCR, g/g | Corn-Soybean | 11 | 99.7 | -4.73 (-7.89, -1.58) | 28.44 | 3 | 17.94 | 0.0005 |
| Wheat-Corn-Soybean | 6 | 98.7 | -2.66 (-7.42, 2.09) | 35.21 | ||||
| Corn-Soybean | 3 | 81.9 | -0.91 (-1.49, -0.34) | 0.21 | ||||
| WRES | 4 | 93.2 | -8.30 (-12.36, -4.25) | 14.57 | ||||
NC: the number of comparisons between exogenous phytase treatment and deficient Ca and P control group; I2: proportion of total variation of the common effect size estimate due to heterogeneity; SMD: standardized mean difference; CI: confidence interval; Q: X2 statistics; d.f: degree of freedom; τ2: heterogeneity variance of the actual effect. a WRES: wheat-rapeseed expeller-soybean; CSRRB: corn-soybean-rapeseed rice bran. b p-Value to test for subgroup differences (random effects).
Table 9.
Meta-ANOVA of basal diet’s sources of phosphorus association with measured bone outcomes.
| Random Effects Model | ||||||||
|---|---|---|---|---|---|---|---|---|
| Parameter | Covariate: Dietary P Sourcea | NC | I2, % | SMD (95% CI) | τ2 | d.f | Q | p-Valueb |
| Starter Phase (1-22 days) | ||||||||
| BBS, Kgf/mm | Corn-Soybean | 13 | 91.9 | 3.65 (1.78, 5.51) | 11.22 | 1 | 4.55 | 0.0330 |
| Wheat-Soybean | 4 | 90.5 | 8.16 (4.45, 11.88) | 12.27 | ||||
| Tibia Ash, g/Kg DM | Corn-Soybean | 45 | 99.4 | 4.11 (2.42, 5.79) | 1.92 | 2 | 38.83 | < 0.0001 |
| Wheat-Soybean | 4 | 89.9 | 36.74 (30.47, 43.02) | 29.80 | ||||
| Tibia Ca, g/Kg DM | Corn-Soybean | 15 | 97.4 | 4.52 (0.27, 8.76) | 69.12 | 1 | 0.45 | 0.5041 |
| Wheat-Corn-Soybean | 2 | 0.0 | 3.05 (2.39, 3.73) | 0 | ||||
| Tibia P, g/Kg DM | Corn-Soybean | 16 | 98.0 | 4.65 (0.99, 8.31) | 54.70 | 1 | 2.04 | 0.1534 |
| Wheat-Corn-Soybean | 2 | 92.4 | 10.14 (3.55, 16.73) | 20.91 | ||||
| Grower-Finisher Phase (22-42 days) | ||||||||
| BBS, Kgf/mm | Corn-Soybean | 9 | 92.7 | 3.59 (2.27, 4.91) | 3.63 | 1 | 16.33 | < 0.0001 |
| WRES | 2 | 0.0 | 9.03 (6.75, 11.32) | |||||
| Tibia Ash, g/Kg DM | Corn-Soybean | 16 | 97.8 | 7.53 (3.96, 11.09) | 50.60 | 1 | 11.54 | 0.0007 |
| Wheat-Corn-Soybean | 5 | 49.4 | 1.22 (0.51, 1.94) | 0.32 | ||||
| Tibia Ca, g/Kg DM | Corn-Soybean | 5 | 95.2 | 3.73 (-0.08, 7.55) | 18.188 | 2 | 68.87 | < 0.0001 |
| Wheat-Corn-Soybean | 5 | 27.5 | 0.55 (0.02, 1.09) | 0.10 | ||||
| WRES | 3 | 13.7 | 15.44 (11.91, 18.97) | 2.02 | ||||
| Tibia P, g/Kg DM | Corn-Soybean | 5 | 96.0 | 5.55 (0.36, 10.74) | 33.66 | 2 | 72.40 | < 0.0001 |
| Wheat-Corn-Soybean | 5 | 89.4 | 0.32 (-1.33, 1.97) | 3.17 | ||||
| WRES | 3 | 0.0 | 16.12 (12.86, 19.38) | 0 | ||||
NC: the number of comparisons between exogenous phytase treatment and deficient Ca and P control group; I2: proportion of total variation of the common effect size estimate due to heterogeneity; SMD: standardized mean difference; CI: confidence interval; Q: X2 statistics; d.f: degree of freedom; τ2: heterogeneity variance of the actual effect. a WRES: wheat-rapeseed expeller-soybean; CSRRB: corn-soybean-rapeseed rice bran. b p-Value to test for subgroup differences (random effects).
Table 10.
Meta-regression of phytase dosage effects on growth, bone strength, and mineralization outcomes.
Table 10.
Meta-regression of phytase dosage effects on growth, bone strength, and mineralization outcomes.
| Dependent Parameter | Intercept | SE (p-Valuea) | Dosage | QM | d.f. | p-Valueb | R2 (%) |
|---|---|---|---|---|---|---|---|
| Starter Phase (1-22 days) | |||||||
| ADFI, g/d | 13.1238 | 3.27 (<.0001) | -0.0003 | 0.2153 | 1 | 0.6426 | 0.00 |
| ADG, g/d | 9.6103 | 5.82 (0.0985) | 0.0016 | 0.1300 | 1 | 0.7185 | 0.00 |
| BWG, g | 10.7360 | 1.44 (<.0001) | 0.0000 | 0.0015 | 1 | 0.9692 | 0.00 |
| FCR, g/g | -7.9007 | 1.95 (<.0001) | -0.0006 | 2.4100 | 1 | 0.1206 | 1.32 |
| BBS, Kgf/mm | 6.2370 | 1.86 (0.0008) | -0.0010 | 0.9577 | 1 | 0.3278 | 0.00 |
| Tibia Ash, g/Kg DM | 10.2937 | 2.86 (0.0003) | 0.0015 | 0.3922 | 1 | 0.5311 | 0.00 |
| Tibia Ca, g/Kg DM | 7.9141 | 3.07 (0.0100) | -0.0029 | 2.0975 | 1 | 0.1475 | 5.78 |
| Tibia P, g/Kg DM | 9.5629 | 2.69 (0.0004) | -0.0032 | 3.9557 | 1 | 0.0467 | 15.50 |
| Grower-Finisher Phase (22-42 days) | |||||||
| ADFI, g/d | 38.0642 | 25.93 (0.1421) | 0.0037 | 0.0332 | 1 | 0.8555 | 0.00 |
| ADG, g/d | 9.8424 | 10.87 (0.3652) | -0.0033 | 0.0436 | 1 | 0.8345 | 0.00 |
| BWG, g | 62.2857 | 54.22 (0.2507) | -0.0167 | 0.1632 | 1 | 0.6862 | 0.00 |
| FCR, g/g | -4.5661 | 1.51 (0.0025) | 0.0003 | 0.0647 | 1 | 0.7992 | 0.00 |
| BBS, Kgf/mm | 6.8364 | 2.03 (0.0008) | -0.0031 | 1.6752 | 1 | 0.1956 | 3.38 |
| Tibia Ash, g/Kg DM | 0.6803 | 3.24 (0.8339) | 0.0063 | 3.1101 | 1 | 0.0778 | 3.15 |
| Tibia Ca, g/Kg DM | 5.0380 | 4.31 (0.2425) | -0.0001 | 0.0002 | 1 | 0.9886 | 0.00 |
| Tibia P, g/Kg DM | 5.3695 | 4.81 (0.2646) | 0.0005 | 0.0078 | 1 | 0.9297 | 0.00 |
QM: coefficient of moderators; QM is considered significant at p-Value (≤ 0.05); d.f: degree of freedom; R2: the amount of heterogeneity accounted for by the covariate (moderator); SE: standard error of intercept. a p-value to the regression model intercept (random effects). b p-Value to the regression model moderator (dosage).
Table 11.
Meta-regression of duration of phytase exposure on growth, bone strength, and mineralization outcomes.
Table 11.
Meta-regression of duration of phytase exposure on growth, bone strength, and mineralization outcomes.
| Dependent Parameter | Intercept | SE (p-Valuea) | Duration | QM | d.f. | p-Valueb | R2 (%) |
|---|---|---|---|---|---|---|---|
| Starter Phase (1-22 days) | |||||||
| ADFI, g/d | -15.3207 | 15.33 0.3177) | 1.5427 | 3.4411 | 1 | 0.0636 | 2.50 |
| ADG, g/d | -16.1756 | 35.67 (0.6502) | 1.3331 | 0.5988 | 1 | 0.4390 | 0.00 |
| BWG, g | 1.3602 | 6.32 (0.8296) | 0.5302 | 2.3123 | 1 | 0.1284 | 1.31 |
| FCR | -29.0970 | 8.31 (0.0005) | 1.1359 | 6.14 | 1 | 0.0132 | 5.28 |
| BBS, Kgf/mm | -57.8350 | 50.41 (0.2512) | 3.0031 | 1.5383 | 1 | 0.2149 | 1.95 |
| Tibia Ash, g/Kg DM | 6.2928 | 8.19 (0.4423) | 0.2953 | 0.3922 | 1 | 0.4965 | 0.00 |
| Tibia Ca, g/Kg DM | 19.4068 | 5.85 (0.0009) | -0.9962 | 312.37 | 1 | 0.0074 | 29.05 |
| Tibia P, g/Kg DM | 24.2718 | 4.17 (<.0001) | -1.2321 | 22.8293 | 1 | <.0001 | 58.00 |
| Grower-Finisher Phase (22-42 days) | |||||||
| ADFI, g/d | -91.3932 | 144.35 (0.5266) | 6.9971 | 0.8589 | 1 | 0.3540 | 0.00 |
| ADG, g/d | -60.0350 | 66.49 (0.3665) | 3.5078 | 1.0447 | 1 | 0.3067 | 0.49 |
| BWG, g | 511.5840 | 267.52 (0.0558) | -24.73 | 3.0712 | 1 | 0.0797 | 9.11 |
| FCR | -5.7871 | 8.25 (0.4830) | 0.0787 | 0.0331 | 1 | 0.8557 | 0.00 |
| BBS, Kgf/mm | 7.7524 | 3.53 (0.0282) | -0.1255 | 0.9370 | 1 | 0.3331 | 0.00 |
| Tibia Ash, g/Kg DM | 14.4573 | 5.32 (0.0065) | -0.3716 | 2.8099 | 1 | 0.0937 | 10.34 |
| Tibia Ca, g/Kg DM | -43.7178 | 16.60 (0.0084) | 2.5215 | 8.5614 | 1 | 0.0034 | 40.79 |
| Tibia P, g/Kg DM | -56.0797 | 16.19 (0.0005) | 3.2005 | 14.4598 | 1 | 0.0001 | 56.07 |
QM: coefficient of moderators; QM is considered significant at p-Value (≤ 0.05); d.f: degree of freedom; R2: the amount of heterogeneity accounted for by the covariate (moderator); SE: standard error of intercept. a p-value to the regression model intercept (random effects). b p-Value to the regression model moderator (dosage).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



