
Submitted:
07 June 2024
Posted:
10 June 2024
You are already at the latest version
Abstract
The members within the genus Thermoanerobacter have been much less investigated for protein and amino acid metabolism as compared with carbohydrates. The present study investigates the biotechnological potential of Thermoanaerobacter mathranii (DSM 11426) concerning its ability to produce high energy alcohols from amino and fatty acids. End-product formation was analyzed from glucose in the presence and absence of butyric acid as well as from selected proteins, and all amino acids. T. mathranii did not degrade any of the proteins tested to a large extent but de-grade several amino acids, namely serine and the branched-chain amino acids (leucine, isoleu-cine, valine) when cultivated in the presence of thiosulfate. The main end products from the branched-chain amino acids were a mixture of their corresponding branched-chain fatty acids and alcohols. 13C2-labeled leucine revealed that the strains degraded the amino acid in the pres-ence of thiosulfate, producing 3-methyl-1-butyrate, which was then used as an electron acceptor which led to the accumulation of 3-methyl-1-butanol. The strain is highly ethanologenic, pro-ducing more than 1.2 mol ethanol per mol of glucose degraded. The strain was able to reduce volatile fatty acids during glucose fermentation to their corresponding alcohol, further suggest-ing this strain may be of greater biotechnological value beyond bioethanol production.
Keywords:
Thermoanaerobacter
; protein
; amino acids
; alcohols
; fatty acid reduction
1. Introduction
The production of biofuels and other biomolecules from renewable biomass has been an area of intense investigation over the past 40 years. The main attention on biofuel production has been on the utilization of carbohydrates. The urge to use complex biomass for the production of second-generation ethanol has led to research on the use of thermophilic anaerobic bacteria, mainly because of their broad substrate range facilitating the degradation of a wide variety of sugars present in such biomass. The proteinaceous materials are often a substantial portion of biomass, but has not been extensively investigated as a source of biofuels and other bio-manufactured products.
Among thermophilic anaerobic bacteria, Thermoanaerobacter species have been intensively investigated for their ability to produce ethanol from complex biomass because of their broad substrate spectrum. All species within Thermoanaerobacter are obligate anaerobes; that ferment various carbohydrates to ethanol, acetate, lactate, hydrogen, and carbon dioxide [1,2]. These bacteria originate from various habitats like hot springs, hydrothermal vents, and oil fields [3,4,5,6,7,8,9,10]. Most of the species within the genus can catabolize the hexoses and pentoses present in lignocellulosic and macroalgae biomasses as well as many of the various disaccharides commonly encountered, notably cellobiose. Paired with the ability to utilize starch and often xylan, Thermoanaerobacter species are excellent candidates for the production of biofuels such as hydrogen and ethanol from complex biomass. As an example, many Thermoanaerobacter species are highly ethanologenic, with T. ethanolicus, T. pseudethanolicus, Thermoanaerobacter strain J1, and Thermoanaerobacter strain AK15 demonstrating ethanol yields above 1.5. mol ethanol from one mole of glucose [11,12,13,14,15]. Additionally, many of these strains also show high yields of ethanol from hydrolysates from various lignocellulosic biomasses [16,17,18,19,20]. Thus, our knowledge of carbohydrate metabolism leading to the production of second-generation ethanol has increased considerably in the past two decades. However, our understanding of the role of protein and amino acid metabolism among thermophilic anaerobes is not explored fully, despite its importance in various ecosystems as a part of nutrient cycling [21,22,23].
Most of our early understanding of protein degradation under anaerobic conditions is derived from mesophilic anaerobes, particularly proteolytic pathogens such as Clostridum botulinium [24,25,26,27,28], Clostridium perfringens [29], Clostridium tetanmorphum [30] among others [31,32,33,34,35,36,37]. By comparison, studies on thermophilic anaerobes have been limited to several genera like Caloramator and Thermoanerobacter species of which several strains are known to be proteolytic to some extent although the specific usage patterns of strains are often limited due to relatively few proteins and proteogenic amino acids being tested [38,39,40]. The importance of the need of electron acceptors for protein and amino acid degradation has been known for some time due to the unfavorable thermodynamics involved and is important knowledge to understand the role of thermophilic bacteria in hot environments [41,42,43]. As an example, it is known that the acetogen Thermoanaerobacter kivui uses alanine in the presence of thiosulfate [44] although use of other amino acids was not reported. Coprothermobacter (formerly Thermobacteroides) proteolyticus, as the name suggests, can utilize gelatin in the presence of a methanogen resulting in fermentation products such as branched-chain fatty acids (BCFAs) associated with the degradation of branched-chain amino acids (BCAAs) [45].
While several studies by our group have previously focused upon the BCAA catabolism of Thermoanaerobacter strains, namely Thermoanaerobacter strain AK85 [46], T. pseudethanolicus [47], and T. brockii [48], there have been one comprehensive investigation of all twenty proteogenic amino acids using Thermoanaerobacter strain AK90 [49], which is not deposited in a publicly accessible culture collection. These studies have shown that species within the genus Thermoanaerobacter can degrade the BCAA in the presence of a hydrogen scavenging organisms, like hydrogenotrophic methanogens as well as by using thiosulfate in the medium to scavenge the electrons produced in the oxidative deamination of these amino acids. Interestingly, co-cultivating Thermoanaerobacter strains with a hydrogenotrophic methanogen during growth on BCAA resulted in the production of their corresponding BCFA only. However, by adding thiosulfate to the medium these bacteria produce a mixture of both their corresponding BCFA and branched-chain alcohols (BCOHs). The production of the alcohol seems thus to be dependent on the electron scavenging system by these bacteria.
In the early 1990s, Thermoanaerobacter strain A3, which would go on to be described as Thermoanaerobacter mathranii, was isolated from an alkaline hot spring (70°C, pH 8.5) in Hveragerði (SW Iceland) through the enrichment of sediment material in anaerobic media containing 0.2% w/v xylan [50]. The strain produced ethanol as a dominant end product from xylose with other end products being carbon dioxide, hydrogen, and lactate. Strain A3 was formally described as a new species, Thermoanaerobacter mathranii, several years later during which time one of the authors, Indra Mathrani, of the original paper describing this strain had passed away due to food poisoning, resulting in the strain bearing his namesake in his honor [4]. The strain grows between 50 and 75°C with an optimum for growth at 70°C and grows best at near neutral pH. Like other Thermoanaerobacter species, T. mathranii is a mixed acid producer although ethanol is the dominant end product from glucose. Notably, T. mathranii subsp. Alimentarius is not as sensitive to ethanol as other thermophilic anaerobes, being able to tolerate 4% v/v without prior adaptation, making it a promising candidate for bioethanol production [51]. The strain has been intensively investigated for its ethanol production purpose, both the wild type (Ahring et al., 1999) as well as several genetically modified strains that have originated from the A3 type strain [16,17]. To our best knowledge, protein and amino acid metabolism of T. mathranii has not been investigated in any detail earlier. T. mathranii under NCBI Bioproject PRJNA33329, was sequenced and annotated by the DOE Joint Genome Institute (JGI) and uploaded to NCBI. JGI performed the genomic annotation with Prokaryotic Genomic Annotation Pipeline (PGAP) program. The genome assembly is further available on NCBI with reference genome accession number ASM9296v1.
The present investigation is directed towards gaining insight into the protein and amino acid metabolism of Thermoanaerobacter mathranii. Of particular interest is the strain’s potential to generate the corresponding BCOH like previously reported Thermoanaerobacter strains as well as better understanding a potential reductive route from the fatty acids produced by BCAA catabolism to their corresponding primary alcohol.
2. Materials and Methods
2.1. Culture Medium and Preparation
All reagents were obtained from Sigma Aldrich unless otherwise noted. Keratin was locally obtained from milled and used without further preparation. Thermoanaerobacter mathranii (DSM 11426) was acquired from DSMZ and was cultivated in Basal Mineral (BM) medium prepared as previously described [52]; the medium consisted of (per liter): NaH2PO4 2.34 g, Na2HPO4 3.33 g, NH4Cl 2.2 g, NaCl 3.0 g, CaCl2 8.8 g, MgCl2 x 6H2O 0.8 g, yeast extract 2.0 g, resazurin 1 mg, trace element solution 1 mL, vitamin solution (DSM141) 1 mL and NaHCO3 0.8 g. The trace element solution consisted of the following on a per liter basis: FeCl2 x 4 H2O 2.0g, EDTA 0.5 g, CuCl2 0.03 g, H3BO3, ZnCl2, MnCl2 x 4 H2O, (NH4)Mo7O24, AlCl3, CoCl2 x 6 H2O, NiCl2, and 0.05 g, Na2S x 9 H2O 0.3 g, and 1 mL of concentrated HCl. The medium was prepared by adding the buffer to distilled water containing resazurin and boiled for 10 min and cooling under nitrogen flushing. The mixture was then transferred to serum bottles using the Hungate technique [53,54] and autoclaved (121°C) for 60 min. All other components of the medium were added separately through filter (0.45 µm) sterilized solutions. All experiments were conducted at 65°C and at pH of 7.0 with a liquid-gas (L-G) ratio of 1:1 unless otherwise noted. In all cases, experiments were performed in triplicate.
2.2. Substrate Utilization Spectrum
The ability of the strain to utilize amino acids were tested at 20 mM concentration in absence and presence of thiosulfate (20 mM). Protein degradation (casein, collagen, gelatine, keratin) was tested at 0.2% (w/v) concentration. Cultures were incubated for a period of 5 days at which time end products were analyzed. Experiments were done in 25 mL serum bottles with liquid – gas phase ratio of 1.0.
To investigate the electron flow during glucose fermentation, in the presence and absence of butyrate as an external electron acceptor, the strain was cultivated in BM medium on glucose (20 mM) only and on glucose (20 mM) with butyrate (20 mM). The experiment was performed in 57 mL serum bottle with a liquid phase ratio of 1.0.
2.3. NMR Experiments
2.4. Genome Search
To analyze the genome of T. mathranii subsp. mathranii A3, the reference genome assembly and PGAP annotation files were downloaded from NCBI’s genome database. The Refseq annotation file was searched by manual identification for relevant genes corresponding to the BCAA fermentation, carboxylic acid reduction, and ethanol fermentation pathways. The Expsasy Enzyme nomenclature database was applied in searching for alternative naming conventions of enzymes and determining the EC number. The protein sequences were cross referenced with the Uniprot database by BLAST, to confirm the enzymatic activity of the sequence in question. It is through this method that relevant enzymes were detected within the T. mathranii genome.
2.5. Analytical Methods
Hydrogen was analyzed by Perkin Elmer Auto System XL gas chromatograph according to [56]. Alcohols and volatile fatty acids were quantified by gas chromatography using a Perkin-Elmer Clarus 580 gas chromatograph as previously described [56]. Optical density was determined by measuring absorbance at 600 nm by a Perkin Elmer Lambda-25 UV-Vis spectrophotometer in a cuvette with a pathlength of 1 cm. Hydrogen sulfide was analyzed as described by Cline (1969) [57].
3. Results and Discussion
3.1. Enzymatic Activity
Unlike other genera commonly described in the literature, Thermoanaerobacter strains have not undergone some of the routine chemotaxonomic tests using commercially available screening kits as have many other mesophilic bacteria. In order to get an overview of the common carbohydrate and amino acid-related enzyme activities present among T. mathranii were evaluated; the use of the API ZYM strip proved to be facile although attempts to use the strips at lower temperatures did not produce good results supporting the thermophilic nature of the enzymes. T. mathranii was found to be positive for esterase, trypsin, acid phosphatase and napthol-AS-BI-phosophohydrolase (Supplementary Table S1).
Being highly saccharolytic, it is a little surprising that the strain was found negative on the enzymes typically involved in carbohydrate catabolism, such as α- and β-galactosidase, α- and β-glucosidase (Supplementary Table S1).
T. mathranii tests positive for the presence of an esterase that is active on four carbon substrates but seems to lack activity on C8 or C14 esters. While the explicit physiological function of esterases in Thermoanaerobacter is not known, it has been speculated that it may serve to help with deacylation of hemicellulose and other polymeric carbohydrates. Esterases have broad biotechnological potential and cold-active esterases have demonstrated broad catalytic promiscuity giving them a wide range of applications in organic synthesis. To the authors’ knowledge, few if any thermostable esterases have been exploited making this a potentially interesting avenue for future work. The presence of two phosphatases, acid phosphatase and napthol-AS-BI-phosophohydrolase, indicates that T. mathranii is capable of scavenging phosphate from its environment from both inorganic and organic sources. T. mathranii tested positive for trypsin, a common serine protease, which suggests that this strain may have some role in protein hydrolysis in its environment even if the strain itself does not seem to be highly proteolytic (see later). The trypsin produced by T. mathranii has not been previously described but warrants further investigation. The biotechnological potential of trypsin is well-established with this enzyme having found applications.
3.2. Degradation of Amino Acids
Much less attention has been paid to the capacity of thermophiles to degrade amino acids. Most of the information on amino acid catabolism among Clostridia has been gained from well-known proteolytic members of Clostridium sporogenes [36], Clostridium botulinum [31], and Clostridium sticklandii [32,33,34]. The degradation of amino acids is a complex process involving several oxidation and reduction steps and some amino acids can be degraded via multiple routes often with specific conditions being necessary. Generally, the most common pathways employ a to use a two-step mechanism involving a preliminary oxidative deamination of the amino acid, yielding a corresponding α-keto acid, which is then oxidatively decarboxylated to give one carbon shorter fatty acids (McInerney, 1988). However, under anaerobic conditions, this route is only possible for amino acids with high oxidation stages, such as serine and threonine [43]. The reduced amino acids, such as the BCAA (leucine, isoleucine, and valine) as well as alanine, are usually not degraded as single substrates under anaerobic conditions unless an electron scavenging element is added to the medium. In the 1990s, several investigations showed that these reduced amino acids were degraded when the amino acid degrading bacteria could dispose the electrons produced during oxidation of these amino acids to external electron accepter (either by co-cultivating the amino acid degrading bacterium with a hydrogenotrophic methanogen or through the use thiosulfate as a chemical electron acceptor). For instance, Fardeau et al. (1997) demonstrated that Thermoanaerobacter brockii degraded the BCAA only in the presence of thiosulfate, producing one carbon shorter BCFA [41]. Later investigations revealed that the BCAA were not only degraded to their corresponding fatty acid, but to a mixture of their corresponding BCFA and BCOH by Thermoanaerobacter brockii, and Caldanaerobacter subterraneaus subsp. yonseiensis, in the presence of thiosulfate [48].
In the present study, Thermoanaerobacter mathranii degraded serine and the three BCAAs (Figure 1), but only in the presence of thiosulfate. Apart from T. brockii some work has been done on other Thermoanaerobacter strains which reveals broad differences in the ratio of the alcoholic to carboxylic acid products. Table 1 shows selected data available in literature concerning leucine metabolism whereas Supplementary shows similar data for all three BCAAs BCAA. Compared to other Thermoanaerobacter strains, T. mathranii uses relatively few of the 20 proteogenic amino acids. For comparison, Thermoanaerobacter strain AK90 showed a greater variety of amino acids degraded, in addition to serine, six other amino acids were degraded in the presence of thiosulfate.
Similar to other Thermoanaerobacter strains, T. mathranii produced a mixture of BCFAs and BCOHs from the BCAAs and the concentration of the acid was always considerable higher than the alcohol in all cases (Figure 1).
Leucine was thus degraded to a mixture of 3-methyl-1-butyrate and 3-methyl-1-butanol, isoleucine to 2-methyl-1-butyrate and 2-methyl-1-butanol, and valine to 2-methyl-1-propionate and 2-methyl-1-propanol. The highest amount of BCFA and BCOH is accounted for valine degradation resulting in 1.73 and 10.90 mM of the 2-methyl-1-propanol and 2-methyl-1-propionate, respectively. T. mathranii generally has higher ROH to RCOOH ratios with ratios of 0.16 for valine, 0.33 for leucine, and 0.38 for isoleucine than other strains examined thus far while other members of Thermoanaerobacter (Table 1; Supplemental Table S2). Compared to other Thermoanaerobacter and Caldanaerobacter strains degrading BCAAs described in the literature to date, T. mathranii produces higher concentrations of BCOHs, such a 2-methyl-1-propanol from valine. For comparison, Thermoanaerobacter strains typically have ROH/RCOOH ratios between 0.05 to 0.22 for valine, 0.11 to 0.36 for leucine, and 0.09 to 0.31 for isoleucine when thiosulfate is used as the terminal electron acceptor (Table 1; Supplemental Table S2).
Later studies on other strains within the genera Thermoanaerobacter and Caldanaerobacter showed that this ability to produce a mixture of alcohols and acids from BCAA was common among both genera [49]. Investigations to understand in more detail the reaction pathway these bacteria use to produce both the acid and the alcohol have been done with Thermoanaerobacter strain AK85 showed that indeed the partial pressure of hydrogen was of great importance for the ratio of end products formed [46]. To demonstrate the conversion of leucine to a mixture of its corresponding fatty acid and alcohol T. mathranii was cultivated on 13C2-labeled leucine, without and with thiosulfate. When the strain was cultivated without thiosulfate, no labeled end products were formed (Figure 2A). Conversely, when thiosulfate was added, both C2 3-methyl-1-butyrate and C2 3-methyl-1-butanol were produced (Figure 2B).
To demonstrate whether the 3-methyl-1-butyrate can be converted to its corresponding alcohol the strain was finally cultivated on leucine with 13C1-labeled 3-methyl-1-butytate in the presence of thiosulfate. This resulted in the formation of 3-methyl-1-butanol showing that the fatty acid can indeed act as an electron acceptor instead of thiosulfate and produce the corresponding alcohol (Figure 2C). Earlier studies with Thermoanaerobacter strain AK85 and Thermoanaerobacter pseudethanolicus have also shown that by using 13C1 leucine as a substrate that BCAAs are first converted to the BCFAs which in turn is used as an electron donor and is converted to its corresponding alcohol [58].
3.3. Degradation of Proteins
Protein degradation by thermophilic anaerobic bacteria has received much less attention as compared to carbohydrates. Strictly anaerobic mesophiles capable of degrading protein have more attention such as Clostridium botulinum [28], Clostridium difficile [59],Clostridium perfringens [29]. Studies on thermophilic anaerobes have been limited to several genera like Caloramator and Thermoanaerobacter. The importance of electron acceptors for protein and amino acid degradation has been known for some time now and is important knowledge to understand the role of thermophilic bacteria in hot environments [42].
T. mathranii was tested for growth on four types of proteins, casein, collagen, gelatin, and keratin at concentrations of 0.2% w/v in the presence or absence of 20 mM of thiosulfate (Figure 3). The strain only produced acetate and ethanol slightly above control values for casein, collagen, gelatin as compared to yeast extract controls while cultivation on keratin resulted in end products similar to control values. However, addition of peptone and yeast extract enhanced both growth and end product formation by the strain (results not shown).
Interestingly, BCFAs and BCOHs were not above control values although as fermentation products were analyzed after five days, higher titers could have been higher if additional time for peptide hydrolysis is needed for complete degradation.
3.4. Conversion of Fatty Acids to Alcohols
Being able to reduce BCFAs to their corresponding alcohol during the degradation of the BCAAs, as shown above, is of interest and has led to investigation of the general ability of this phenomenon. As common among thermophilic anaerobes, Thermoanaerobacter and Caldanaerobacter dispose of their electrons produced during glucose (and other carbohydrates) oxidations to pyruvate to produce ethanol or lactate. Recent investigations have, however demonstrated that some Thermoanaerobacter strains can also reduce other electron acceptors like fatty acids which are converted to their corresponding alcohols [46,60,61,62]. This was tested for Thermoanaerobacter mathranii by cultivating the strain on glucose only and on glucose in the presence of butyrate. When the strain was cultivated on glucose only it led to a mixture of ethanol, acetate and lactate according to Equation (1):
1 Glucose → 1.23 Ethanol + 0.26 Acetate + 0.31 Lactate + 1.49 CO2 + 0.51 H2
When the strain was cultivated on glucose with the addition of 20 mM of butyrate the reaction stoichiometry changed according to Equation (2).
1 Glucose + 1 Butyrate → 0.85 Ethanol + 0.65 Acetate + 0.13 Lactate + 0.47 Butyrate + 1.50 CO2 + 0.07 H2 + 0.37 Butanol
Thus, as expected, the strain produces less ethanol and more acetate in the presence of butyrate as an electron acceptor and the fatty acid is partially converted to its corresponding alcohol, butanol. Instead of reducing pyruvate to only ethanol and lactate only, the reducing power is also used to reduce butyrate to its corresponding alcohol, butanol.
3.5. Genome and Pathway of Amino Acid Metabolism
T. mathranii’s genome was used to investigate the possible enzymes present that are responsible for the amino acid metabolism shown to be active. The strain only degraded serine and the BCAA in the present study. Serine was weakly degraded to a mixture of acetate and ethanol when used as a sole substrate but mainly to acetate when thiosulfate was added to the medium. The strain was shown positive for genes responsible for serine-O-acetyltransferase and serine hydrolyase. The BCAAs (leucine, isoleucine, valine) were only degraded when the strain was cultivated in the presence of thiosulfate. Genomic studies indeed show the presence of BCAA-specific ABC transporter permeases as well as BCAA transaminases. A proposed scheme for BCAA utilization and the production of BCFA and BCOH is shown in Figure 4. Supplementary Table S3 comprises the enzymes found present in the genome of T. mathranii.
Other genome studies show activity of several dehydrogenases, such as histidinol- homoserine-, and L-lactate dehydrogenases as well as several genes responsible for glutamate and histidine metabolism. These two amino acids were however not utilized by T. mathranii in the present study. Presence of enzymes involved in carboxylic acid reduction were investigated like acetaldehyde decarboxylase, alcohol dehydrogenase, acetate kinase, acetyl phosphotransferase, pyruvate ferredoxin oxidoreductase, pyruvate formate lyase and aldehyde reductase (Table 2, Supplementary Table S3) using Thermoanaerobacter psudethanolcus as a reference. Both strains were positive for most of the enzyme pathways investigated with the only major difference between the species is that T. pseudethanolicus is positive for aldehyde ferredoxin oxidoreductase but T. mathranii not, and T. mathranii is positive for BCAA ABC transporter permease but T. pseudethanolcus not (Table 2). Concerning the utilization of thiosulfate as an electron acceptor the strain is positive for thiosulfate transporter family protein and thiosulfide reductase family protein as expected being able to utilize thiosulfate as an electron scavenger with the reduced product being hydrogen sulfide as is a well known phenomenon of most Thermoanerobacter strains [5,6,8,63].
The strain was tested for the utilization of four proteins; casein, collagen, gelatine and keratin. The strain did only produce small amounts of end products from these proteins but was found to be positive for several protein and peptide degrading enzymes. For example the strain was positive for trypsin like peptidase domain containing protease, ATP-dependent Cip protease ATP binding subunit CipX and ATP-dependent protease ATPase subunit HsIU.
4. Conclusions
Here we confirm the activity of BCAA fermentation via 13C NMR and propose a mechanism for the synthesis of BCOHs in T. mathranii. The significance of this pathway has multiple implications across the genus. First, not all Thermoanaerobacter species within the genus exhibit BCAA transporters that are able to take advantage of this mechanism. T. pseudethanolicus, a highly studied organism, does not have BCAA transporters in their corresponding genomes. Thus, their ability to ferment branched chain alcohols are significantly reduced.
As shown, T. mathranii is able to serve as a platform for the bio-manufacturing of BCOHs for producing 3-methyl-1-butanol, 2-methyl-1-butanol, and 2-methyl-1-propanol from the corresponding BCAAs of leucine, isoleucine, and valine respectively. While two enzymes in the proposed pathway have yet to be identified for the decarboxylation step and the inferred thiosulfate reductase specifically; the presence of these enzymes are implied by the corresponding activity of the strain. These findings warrants further investigation in the annotated genome of T. mathranii to identify the exact enzymes responsible for the two aforementioned enzymatic reactions.
The data herein additionally showcases a potential for proteolytic activity within the strain. Not only does T. mathranii have a trypsin like peptidase (Table 2, Supplementary Table S2), the strain also expresses a specific BCAA transporter. By comparison, T. pseudethanolicus exhibits a similar genomic profile but, do not have an annotated BCAA transporter. In particular T. pseudethanolicus is able to perform a similar fermentation reaction using BCAA.
The presence of a BCAA transporter and trypsin like peptide can potentially indicate that T. mathranii’s role in the microbiome may be involved in the nitrogen cycling and alcohol formation. Further, T. mathranii was tested positive for trypsin activity. This hints that not only the strain is naturally proteolytic but may be involved in the nitrogen cycle. While the exact relationships between T. mathranii and other thermophilic species are speculated. it is undeniable that this strain has displayed some level of proteolytic activity and should facilitate more research into the proteolytic capacity of the strain.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Supplemental Table S1. API ZYM assay results for Thermoanaerobacter mathranii as compared to T. pseudethanolicus; Supplemental Table S2 – Comparison of end products formed from branched-chain amino acid fermentation by Caldanaerobacter and Thermoanaerobacter strains; Supplemental Table S3 - Thermoanaerobacter manthranii genes.
Author Contributions
Conceptualization, J.O, S.M.S, C.A.; methodology, J.O, S.M.S.; software, S.M.S, C.A.; validation, J.O, S.M.S.; formal analysis, J.O, S.M.S.; investigation, J.O, S.M.S.; resources, J.O.; data curation, S.M.S, C.A.; writing—original draft preparation, J.O, S.M.S.; writing—review and editing, J.O, S.M.S, C.A.; visualization, S.M.S.; supervision, J.O.; project administration, J.O. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data are available in the Supplementary Materials associated with this work.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Stackebrandt, E. The Family Thermoanaerobacteraceae. In The Prokaryotes; Springer Berlin Heidelberg: Berlin, Heidelberg, 2014; pp. 413–419. [Google Scholar]
- Wiegel, J.; Tanner, R.; Rainey, R. An Introduction to the Family Clostridiaceae. In Prokaryotes; 2006; pp. 654–678.
- Collins, M.D.; Lawson, P.A.; Willems, A.; Cordoba, J.J.; Fernandez-Garayzabal, J.; Garcia, P.; Cai, J.; Hippe, H.; Farrow, J.A. The Phylogeny of the Genus Clostridium: Proposal of Five New Genera and Eleven New Species Combinations. Int J Syst Bacteriol 1994, 44, 812–826. [Google Scholar] [CrossRef] [PubMed]
- Larsen, L.; Nielsen, P.; Ahring, B.K. Thermoanaerobacter mathranii Sp. Nov., an Ethanol-Producing, Extremely Thermophilic Anaerobic Bacterium from a Hot Spring in Iceland. Arch Microbiol 1997, 168, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-E.; Jain, M.K.; Lee, C.; Zeikus, J.G. Taxonomic Distinction of Saccharolytic Thermophilic Anaerobes: Description of Thermoanaerobacterium xylanolyticum Gen. Nov., Sp. Nov., and Thermoanaerobacterium saccharolyticum Gen. Nov., Sp. Nov.; Reclassification of Thermoanaerobium. Int J Syst Bacteriol 1993, 43, 41–51. [Google Scholar] [CrossRef]
- Wagner, I.D.; Wiegel, J. Diversity of Thermophilic Anaerobes. Ann N Y Acad Sci 2008, 1125, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Slobodkin, A.I.; Tourova, T.P.; Kuznetsov, B.B.; Kostrikina, N.A.; Chernyh, N.A.; Bonch-Osmolovskaya, E.A. Thermoanaerobacter siderophilus Sp. Nov., a Novel Dissimilatory Fe(III)-Reducing, Anaerobic, Thermophilic Bacterium. Int J Syst Bacteriol 1999, 49, 1471–1478. [Google Scholar] [CrossRef] [PubMed]
- Kozianowski, G.; Canganella, F.; Rainey, F.A.; Hippe, H.; Antranikian, G. Purification and Characterization of Thermostable Pectate-Lyases from a Newly Isolated Thermophilic Bacterium, Thermoanaerobacter italicus Sp. Nov. Extremophiles 1997, 1, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Kublanov, I. V; Tsiroulnikov, K.B.; Kaliberda, E.N.; Rumsh, L.D.; Haertle, T.; Bonch-Osmolovskaya, E.A. Keratinase of an Anaerobic Thermophilic Bacterium Thermoanaerobacter Sp. Strain 1004-09 Isolated from a Hot Spring in the Baikal Rift Zone. Microbiology (N Y) 2009, 78, 67–75. [Google Scholar] [CrossRef]
- Bonch-Osmolovskaya, E.A.; Miroshichenko, L.M.; Chernykh, N.A.; Kostrikina, N.A.; Pikuta, E. V; Rainey, F.A. Reduction of Elemental Sulfur by Moderately Thermophilic Organotrophic Bacteria and the Description of Thermoanaerobacter sulfurophilus Sp. Nov. Mikrobiologiya 1997, 66, 581–587. [Google Scholar]
- Jessen, J.E.J.; Orlygsson, J. Production of Ethanol from Sugars and Lignocellulosic Biomass by Thermoanaerobacter J1 Isolated from a Hot Spring in Iceland. J Biomed Biotechnol 2012, 1869–1882. [Google Scholar] [CrossRef]
- Onyenwoke, R.U.; Kevbrin, V. V.; Lysenko, A.M.; Wiegel, J. Thermoanaerobacter pseudethanolicus Sp. Nov., a Thermophilic Heterotrophic Anaerobe from Yellowstone National Park. Int J Syst Evol Microbiol 2007, 57, 2191–2193. [Google Scholar] [CrossRef]
- Wiegel, J.; Ljungdahl, L.G. Thermoanaerobacter ethanolicus Gen. Nov., Spec. Nov., a New, Extreme Thermophilic, Anaerobic Bacterium. Arch Microbiol 1981, 128, 343–348. [Google Scholar] [CrossRef]
- Orlygsson, J.; Scully, S.M. Biotechnological Prospects of Thermoanerobacter AK15: End-Product Formation from Carbohydrates, Amino Acids, and Lignocellulosic and Macroalgae Hydrolysates. Int J Mol Sci 2024, 25, 3490. [Google Scholar] [CrossRef] [PubMed]
- Clarkson, S.M.; Hamilton-Brehm, S.D.; Giannone, R.J.; Engle, N.L.; Tschaplinski, T.J.; Hettich, R.L.; Elkins, J.G. A Comparative Multidimensional LC-MS Proteomic Analysis Reveals Mechanisms for Furan Aldehyde Detoxification in Thermoanaerobacter pseudethanolicus 39E. Biotechnol Biofuels 2014, 7, 165. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, T.I.; Ahring, B.K. Evaluation of Continuous Ethanol Fermentation of Dilute-Acid Corn Stover Hydrolysate Using Thermophilic Anaerobic Bacterium Thermoanaerobacter BG1L1. Appl Microbiol Biotechnol 2007, 77, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Georgieva, T.I.; Mikkelsen, M.J.; Ahring, B.K. High Ethanol Tolerance of the Thermophilic Anaerobic Ethanol Producer Thermoanaerobacter BG1L1. Cent Eur J Biol 2007, 2, 364–377. [Google Scholar] [CrossRef]
- Tomás, A.F.; Karakashev, D.; Angelidaki, I. Thermoanaerobacter pentosaceus Sp. Nov., an Anaerobic, Extreme Thermophilic, High Ethanol-Yielding Bacterium Isolated from Household Waste. Int J Syst Evol Microbiol 2013, 63, 2396–2404. [Google Scholar] [CrossRef]
- Tomás, A.F.; Karagöz, P.; Karakashev, D.; Angelidaki, I. Extreme Thermophilic Ethanol Production from Rapeseed Straw: Using the Newly Isolated Thermoanaerobacter pentosaceus and Combining It with Saccharomyces Cerevisiae in a Two-Step Process. Biotechnol Bioeng 2013, 110, 1574–1582. [Google Scholar] [CrossRef]
- Sittijunda, S.; Tomas, A.F.; Reungsang, A.; O-Thong, S.; Angelidaki, I. Ethanol Production from Glucose and Xylose by Immobilized Thermoanaerobacter pentosaceus at 70°C in an up-Flow Anaerobic Sludge Blanket (UASB) Reactor. Bioresour Technol 2013, 143, 598–607. [Google Scholar] [CrossRef] [PubMed]
- McInerney, M.J. Anaerobic Hydrolysis and Fermentation of Fats and Proteins. In Biology of Anaerobic Microorganisms; Zehnder, A.J.B., Ed.; John Wiley & Sons, Ltd.: New York, 1988; pp. 373–415.
- Andreesen, J.R.; Bahl, H.; Gottschalk, G. Introduction to the Physiology and Biochemistry of the Genus Clostridium. In Clostridia; Minton, N.P., Clarke, D.J., Eds.; Plenum Press: New York, 1989; pp. 27–62.
- Reid, S.J.; Stutz, H.E. Nitrogen Assimilation in Clostridia. In Handbook of Clostridia; Durre, P., Ed.; CRC Press: Boca Raton, 2005; pp. 239–260.
- Mitruka, B.M.; Costilow, R.N. Arginine and Ornithine Catabolism by Clostridium botulinum. J Bacteriol 1967, 93, 295–301. [Google Scholar] [CrossRef]
- Tjaberg, T.B. Proteases of Clostridium botulinum II. The Relationship Between Growth Medium and the Production of Proteases by Clostridium botulinum Types A, B, C, D, E and F. Acta Vet Scand 1973, 14, 193–200. [Google Scholar] [CrossRef]
- Tjaberg, T.B. Proteases of Clostridium botulinum I. Classification of Proteases and Literature Survey. Acta Vet Scand 1973, 14, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, B.R.; Sugiyama, H. Isolation and Characterization of Protease from Clostridium botulinum Type B. Biochim Biophys Acta 1972, 268, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Lynt, R.K.; Kautter, D.A.; Solomon, H.M. Differences and Similarities Among Proteolytic and Nonproteolytic Strains of Clostridium botulinum Types A, B, E and F: A Review. J Food Prot 1982, 45, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Hapchuk, L.T.; Perason, A.M.; Price, J.F. Effect of Proteolytic Enzyme Production by Clostridium perfringens upon Porcine Muscle. Food Chem 1979, 4, 213–223. [Google Scholar] [CrossRef]
- Laanbroek, H.J.; Smit, A.J.; Nulend, G.K.; Veldkamp, H. Competition for L-Glutamate Between Specialised and Versatile Clostridium Species. Arch Microbiol 1979, 66, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Deklevat, M.L.; Dasgupta, B.R. Purification and Characterization of a Protease from Clostridium botulinum Type A That Nicks Single-Chain Type A Botulinum Neurotoxin into the Di-Chain Form. J Bacteriol 1990, 172, 2498–2503. [Google Scholar] [CrossRef] [PubMed]
- Elsden, S.R.; Hilton, M.G. Volatile Acid Production from Threonine, Valine, Leucine and Isoleucine by Clostridia. Arch Microbiol 1978, 117, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Elsden, S.R.; Hilton, M.G. Amino Acid Utalization Patterns in Clostridial Taxonomy. Arch Microbiol 1979, 123, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Elsden, S.R.; Hilton, M.G.; Waller, J.M. The End Products of the Metabolism of Aromatic Amino Acids by Clostridia. Arch Microbiol 1976, 107, 283–288. [Google Scholar] [CrossRef]
- Schwartz, A.C.; Schäfer, R. New Amino Acids, and Heterocyclic Compounds Participating in the Stickland Reaction of Clostridium sticklandii. Archives of Mcrobiology 1973, 276, 267–276. [Google Scholar] [CrossRef]
- Stickland, L.H. Studies in the Metabolism of the Strict Anaerobes (Genus Clostridium). The Oxidation of Alanine by Cl. sporogenes. IV. The Reduction of Glycine by Cl. sporogenes. Biochemical Journal 1935, 29, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Stickland, L.H. CCXXXII. Studies in the Metabolism of the Strict Anaerobes (Genus Clostridium). I. The Chemical Reactions by Which Cl. sporogenes Obtains Its Energy. Biochemical Journal 1934, 28, 1746–1759. [Google Scholar] [CrossRef] [PubMed]
- Tarlera, S.; Stams, A.J.M. Degradation of Proteins and Amino Acids by Caloramator proteoclasticus in Pure Culture and in Coculture with Methanobacterium Thermoformicicum Z245. Appl Microbiol Biotechnol 1999, 53, 133–138. [Google Scholar] [CrossRef]
- Chrisostomos, S.; Patel, B.K.C.; Dwivedi, P.P.; Denman, S.E. Caloramator indicus Sp. Nov., a New Thermophilic Anaerobic Bacterium Isolated from the Deep-Seated Nonvolcanically Heated Waters of an Indian Artesian Aquifer. Int J Syst Bacteriol 1996, 46, 497–501. [Google Scholar] [CrossRef]
- Riessen, S.; Antranikian, G. Isolation of Thermoanaerobacter keratinophilus Sp. Nov., a Novel Thermophilic, Anaerobic Bacterium with Keratinolytic Activity. Extremophiles 2001, 5, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Fardeau, M.L.; Patel, B.K.C.; Magot, M.; Ollivier, B. Utilization of Serine, Leucine, Isoleucine, and Valine by Thermoanaerobacter Brockii in the Presence of Thiosulfate or Methanobacterium Sp. as Electron Accepters. Anaerobe 1997, 3, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Faudon, C.; Fardeau, M.L.; Heim, J.; Patel, B.; Magot, M.; Ollivier, B.; Orstom, D.M.; Provence, U. De; Hugo, V.; Cedex, M.; et al. Peptide and Amino Acid Oxidation in the Presence of Thiosulfate by Members of the Genus Thermoanaerobacter. Curr Microbiol 1995, 31, 152–157. [Google Scholar] [CrossRef]
- Örlygsson, J. The Role of Interspecies Hydrogen Transfer on Thermophilic Protein and Amino Acid Metabolism. Ph.D., Swedish University of Agricultural Sciences, 1994.
- Leigh, J.A.; Wolfe, R.S. Acetogenium kivui Gen. Nov., Sp. Nov., a Thermophilic Acetogenic Bacterium. Int J Syst Bacteriol 1983, 33, 866. [Google Scholar] [CrossRef]
- Ollivier, B.M.; Mah, R.A.; Ferguson, T.J.; Boone, D.R.; Garcia, J.L.; Robinson, R. Emendation of the Genus Thermobacteroides: Thermobacteroides proteolyticus Sp. Nov. a Proteolytic Acetogen from a Methanogenic Enrichment. International Journal of Systematic and Evolutionary Bacteriology 1985, 35, 425–428. [Google Scholar] [CrossRef]
- Scully, S.M.; Orlygsson, J. Branched-Chain Amino Acid Catabolism of Thermoanaerobacter Strain AK85 and the Influence of Culture Conditions on Branched-Chain Alcohol Formation. Amino Acids 2019, 51. [Google Scholar] [CrossRef]
- Scully, S.M.; Orlygsson, J. Branched-Chain Amino Acid Catabolism of Thermoanaerobacter pseudoethanolicus Reveals Potential Route to Branched-Chain Alcohol Formation. Extremophiles 2020, 24, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Scully, S.M.; Orlygsson, J. Branched-Chain Alcohol Formation from Branched-Chain Amino Acids by Thermoanaerobacter brockii and Thermoanaerobacter yonseiensis. Anaerobe 2014, 30, 82–84. [Google Scholar] [CrossRef]
- Scully, S.M.; Orlygsson, J. Amino Acid Metabolism of Thermoanaerobacter Strain AK90: The Role of Electron-Scavenging Systems in End Product Formation. J Amino Acids 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Sonne-Hansen, J.; Mathrani, M.; Ahring, B.K. Xylanolytic Anaerobic Thermophiles from Icelandic Hot-Springs. Appl Microbiol Biotechnol 1993, 38, 537–541. [Google Scholar] [CrossRef]
- Carlier, J.P.; Bonne, I.; Bedora-Faure, M. Isolation from Canned Foods of a Novel Thermoanaerobacter Species Phylogenetically Related to Thermoanaerobacter mathranii (Larsen 1997): Emendation of the Species Description and Proposal of Thermoanaerobacter mathranii Subsp. Alimentarus Su. Anaerobe 2006, 12, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Sveinsdottir, M.; Beck, S.R.; Orlygsson, J. Ethanol Production from Monosugars and Lignocellulosic Biomass by Thermophilic Bacteria Isolated from Icelandic Hot Springs. Icelandic Agricultural Science 2009, 22, 45–58. [Google Scholar]
- Hungate, R.E. A Roll Tube Method for Cultivation of Strict Anaerobes. In Methods in Microbiology; Norris, J.R., Ribbons, Eds.; Academic Press: New York, 1969; pp. 117–132. [Google Scholar]
- Miller, T.L.; Wolin, M.J. A Serum Bottle Modification of the Hungate Technique for Cultivating Obligate Anaerobes. Appl Microbiol 1974, 27, 985–987. [Google Scholar] [CrossRef]
- Scully, S.M. Amino Acid and Related Catabolism of Thermoanaerobacter Species. Ph.D., University of Iceland, 2019.
- Orlygsson, J.; Baldursson, S.R.B. Phylogenetic and Physiological Studies of Four Hydrogen-Producing Thermoanareobes from Icelandic Geothermal Areas. Icelandic Agricultural Science 2007, 93–105. [Google Scholar]
- Cline, J.D. Spectrophotometric Determination of Hydrogen Sulfide in Natural Waters. Limnol Oceanogr 1969, 14, 454–458. [Google Scholar] [CrossRef]
- Scully, S.M.; Orlygsson, J. Branched-Chain Amino Acid Catabolism of Thermoanaerobacter Strain AK85 and the Influence of Culture Conditions on Branched-Chain Alcohol Formation. Amino Acids 2019, Accepted. [Google Scholar] [CrossRef]
- Seddon, S. V; Hemingway, I.; Borriello, S.P. Hydrolytic Enzyme Production by Clostridium Difficile and Its Relationship to Toxin Production and Virulence in the Hamster Model. Journal of Medicinal Microbiology 1990, 31, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Hitschler, L.; Kuntz, M.; Langschied, F.; Basen, M. Thermoanaerobacter Species Differ in Their Potential to Reduce Organic Acids to Their Corresponding Alcohols. Appl Microbiol Biotechnol 2018, 102, 8465–8476. [Google Scholar] [CrossRef] [PubMed]
- Scully, S.M.; Brown, A.; Ross, A.B.; Orlygsson, J. Biotransformation of Organic Acids to Their Corresponding Alcohols by Thermoanaerobacter pseudoethanolicus. Anaerobe 2019, 57. [Google Scholar] [CrossRef] [PubMed]
- Scully, S.M.; Brown, A.E.; Mueller-Hilger, Y.; Ross, A.B.; Örlygsson, J. Influence of Culture Conditions on the Bioreduction of Organic Acids to Alcohols by Thermoanaerobacter pseudoethanolicus. Microorganisms 2021, 9, 1–24. [Google Scholar] [CrossRef]
- Lee, Y.-J.; Dashti, M.; Prange, A.; Rainey, F.A.; Rohde, M.; Whitman, W.B.; Wiegel, J. Thermoanaerobacter sulfurigignens Sp. Nov., an Anaerobic Thermophilic Bacterium That Reduces 1 M Thiosulfate to Elemental Sulfur and Tolerates 90 MM Sulfite. Int J Syst Evol Microbiol 2007, 57, 1429–1434. [Google Scholar] [CrossRef]
Figure 1.
End-product formation from BCAA fermentation by T. mathranii with and without thiosulfate (40 mM) after five-day cultivation (65°C, pH 7.0). Values represent the average of triplicate fermentations with standard deviation shown as error bars. Control contains yeast extract only.
Figure 1.
End-product formation from BCAA fermentation by T. mathranii with and without thiosulfate (40 mM) after five-day cultivation (65°C, pH 7.0). Values represent the average of triplicate fermentations with standard deviation shown as error bars. Control contains yeast extract only.
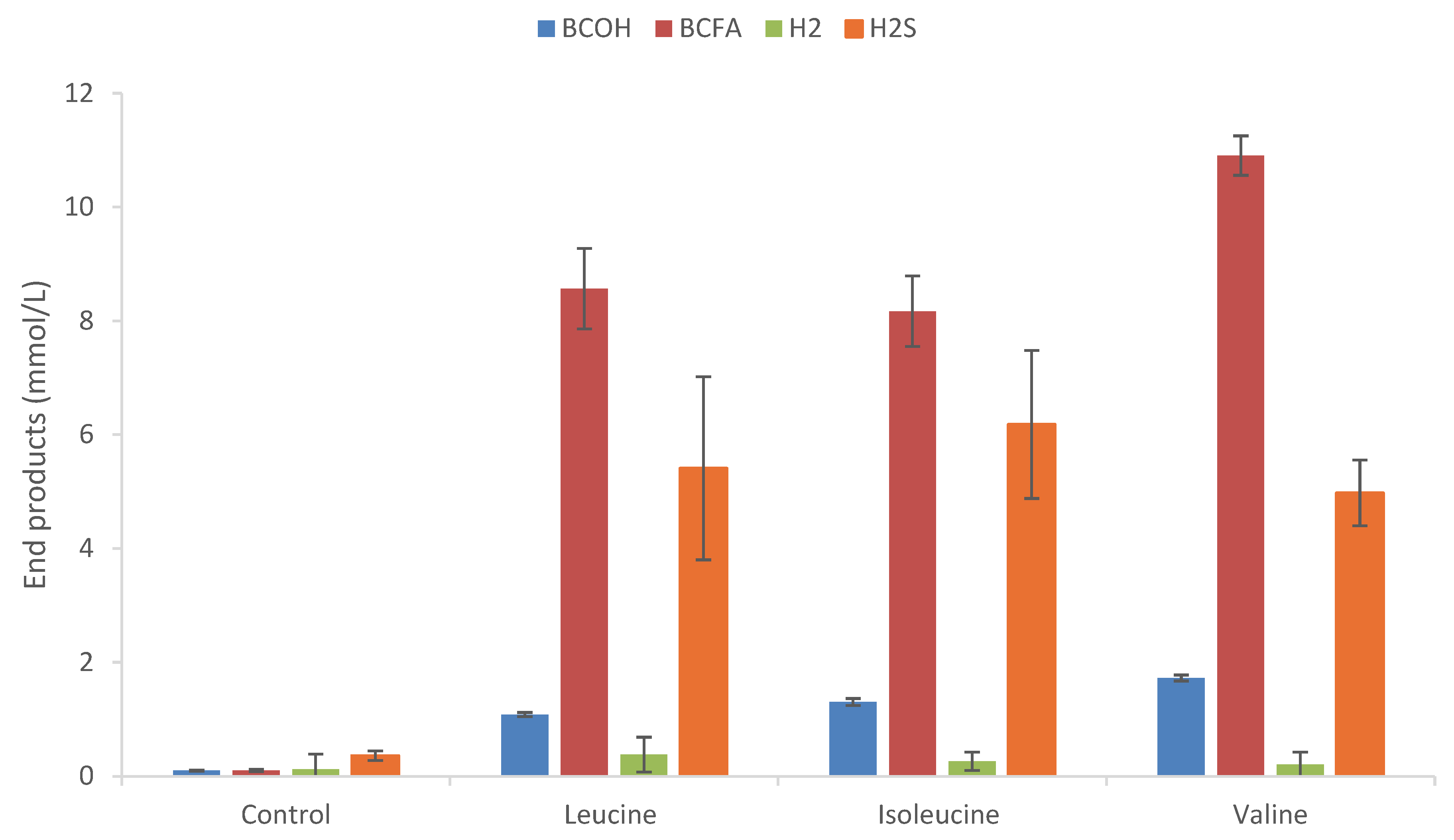
Figure 2.
13C NMR-spectrogram for the fermentation of 13C2 leucine by T. mathranii in the absence (a) and presence of thiosulfate (b) and 13C1 3-methyl-1-butyrate in the presence of unlabeled leucine and thiosulfate (c). Fermentations were performed at 65°C at pH 7.0 over 14 days. Blue, red, and magenta peaks correspond to the 13C-labeled carbon of the leucine, 3-methyl-butyrate, and 3-methyl-butanol, respectively.
Figure 2.
13C NMR-spectrogram for the fermentation of 13C2 leucine by T. mathranii in the absence (a) and presence of thiosulfate (b) and 13C1 3-methyl-1-butyrate in the presence of unlabeled leucine and thiosulfate (c). Fermentations were performed at 65°C at pH 7.0 over 14 days. Blue, red, and magenta peaks correspond to the 13C-labeled carbon of the leucine, 3-methyl-butyrate, and 3-methyl-butanol, respectively.
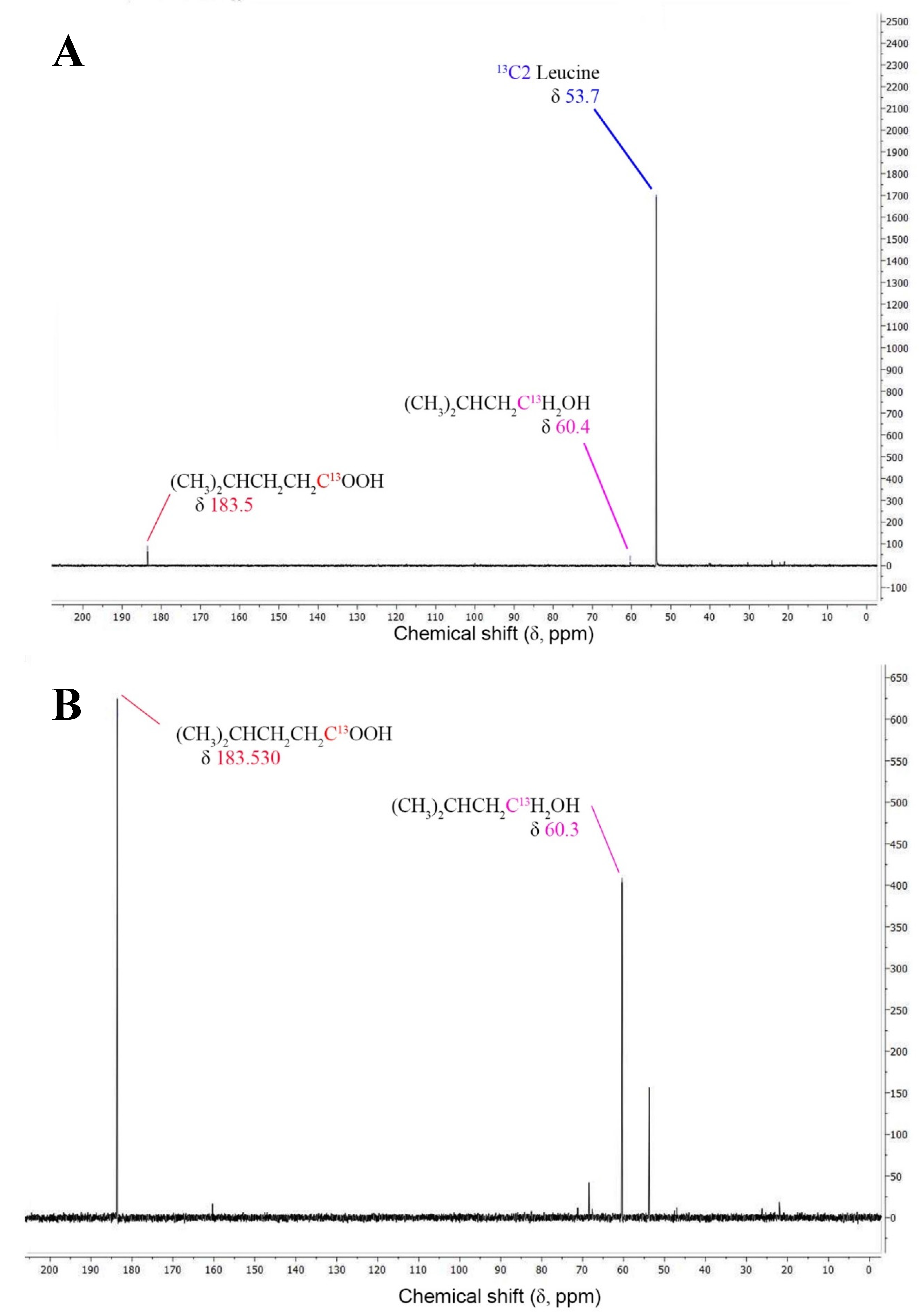
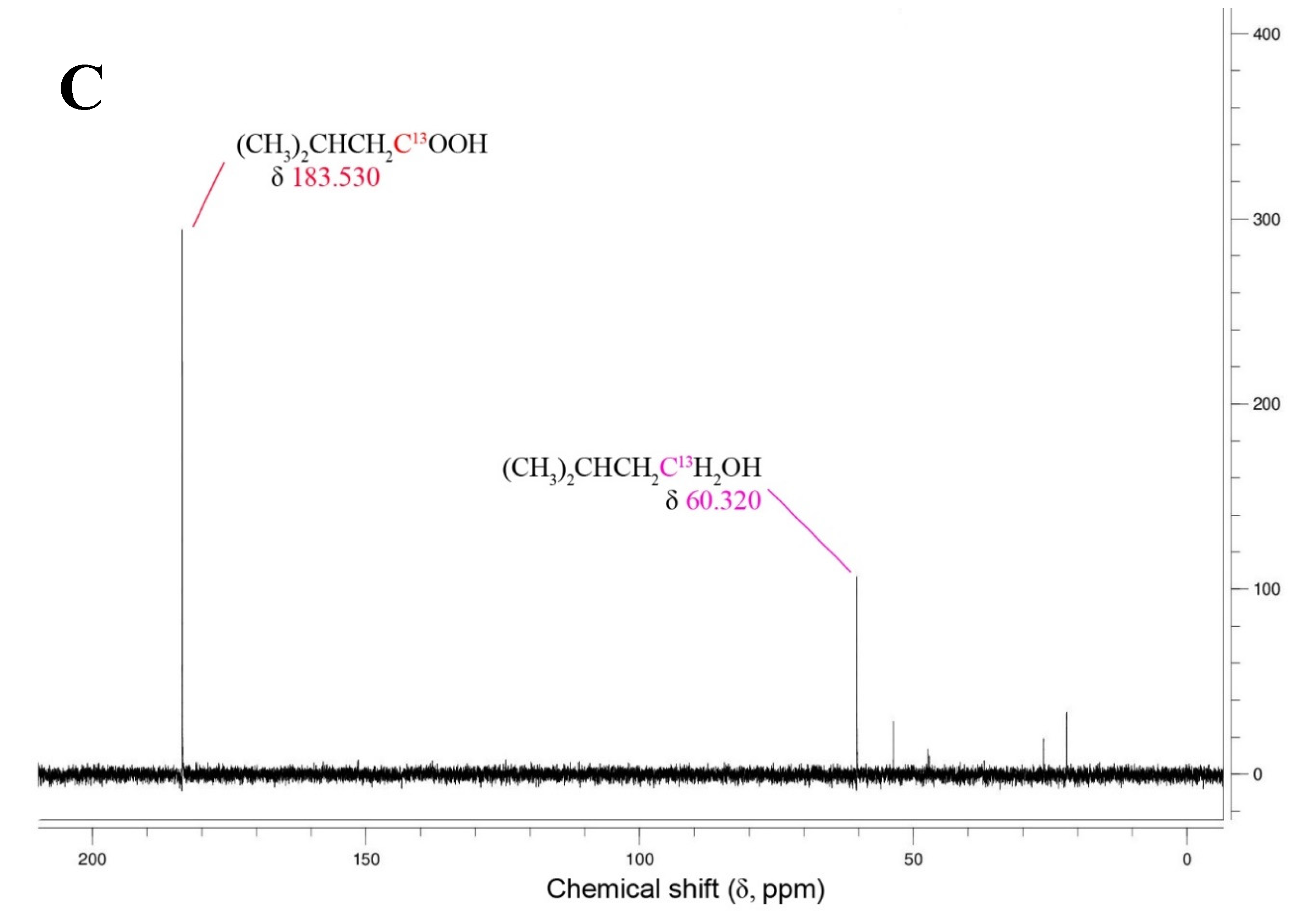
Figure 3.
Fermentation of casein, collagen, gelatine and keratin (0.2% w/v) by T. mathranii for five days cultivation (65°C, pH 7.0). Values represent the average of triplicate fermentations with standard deviation shown as error bars. Control contains yeast extract only.
Figure 3.
Fermentation of casein, collagen, gelatine and keratin (0.2% w/v) by T. mathranii for five days cultivation (65°C, pH 7.0). Values represent the average of triplicate fermentations with standard deviation shown as error bars. Control contains yeast extract only.
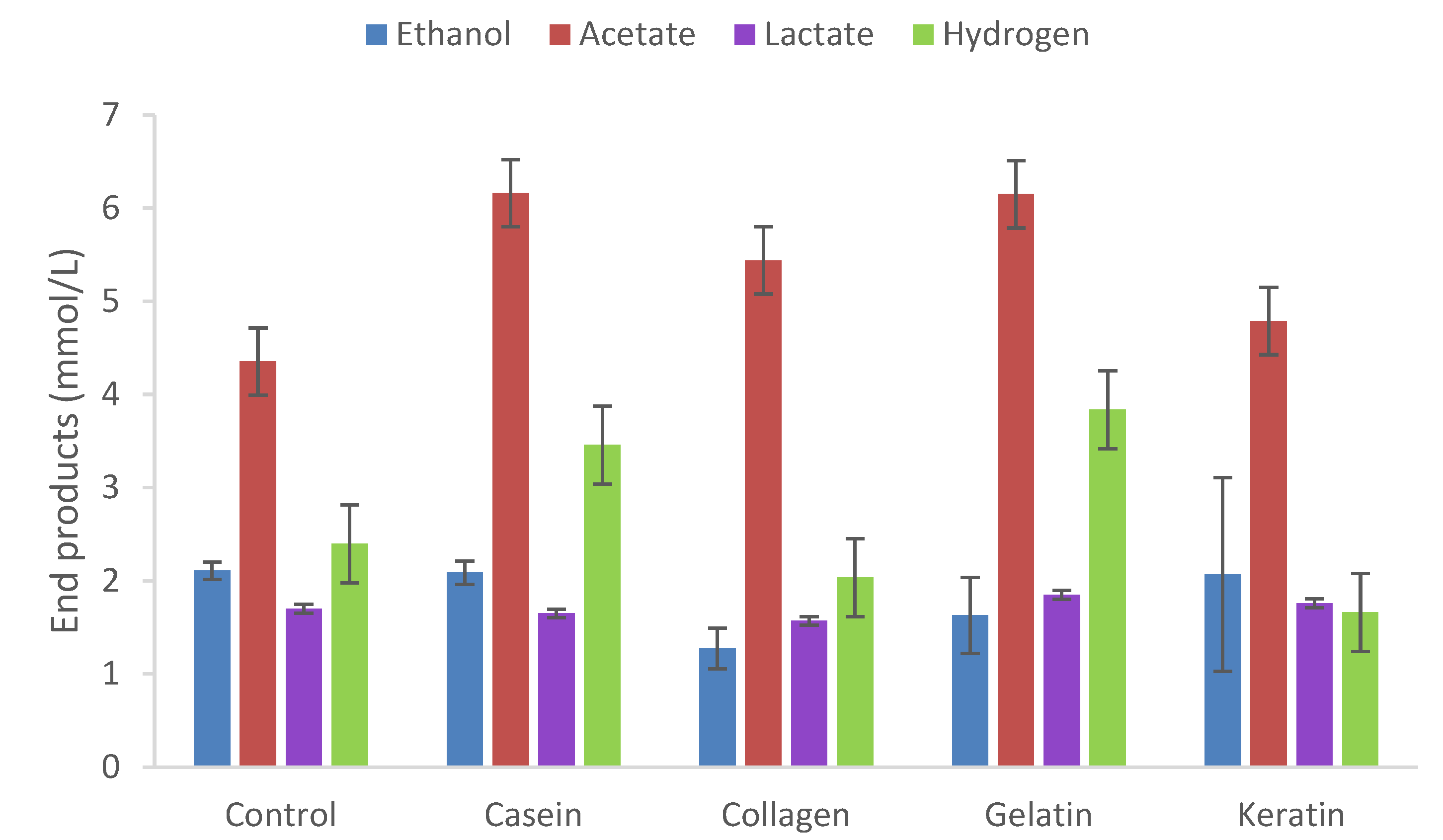
Figure 4.
Details of the proposed pathway for branched chain amino acid fermentation within T. mathranii in the presence of thiosulfate as a reducing agent. It should be noted, that this mechanism is being proposed for all BCAAs including: valine, leucine, and isoleucine. Within the figure, exogenous polypeptides from the environment are degraded by the trypsin like peptidase thus, releasing BCAAs. As displayed, BCAAs such as valine are imported into the cell with a BCAA ABC transporter. Valine subsequently undergoes transamination and decarboxylation, resulting in compounds C and B respectively. While T. mathranii exhibits a BCAA transaminase listed in Table 2 and Supplementary Table S3; the enzyme involved decarboxylation step is illusive. Compound B may be reduced by one of the several potential alcohol dehydrogenases (listed in Supp. Table S2) allowing the alcohol compound passively diffuse through the membrane. Alternatively, the aldehyde itself can passively diffuse through the membrane. For the reducing potential, thiosulfate is imported via a specific thiosulfate transporter. Again, no obvious thiosulfate reductase was found in the annotation. However, Ars sulfite reductase is present for cycling SO3 and S2- concentrations. Therefore, the presence of the thiosulfate reductase is implied. 2-methyl-1-propanol is A; 2-methyl-propanal is B; 2-methyl-1-propionate is C; Dashed lines represent passive diffusion.
Figure 4.
Details of the proposed pathway for branched chain amino acid fermentation within T. mathranii in the presence of thiosulfate as a reducing agent. It should be noted, that this mechanism is being proposed for all BCAAs including: valine, leucine, and isoleucine. Within the figure, exogenous polypeptides from the environment are degraded by the trypsin like peptidase thus, releasing BCAAs. As displayed, BCAAs such as valine are imported into the cell with a BCAA ABC transporter. Valine subsequently undergoes transamination and decarboxylation, resulting in compounds C and B respectively. While T. mathranii exhibits a BCAA transaminase listed in Table 2 and Supplementary Table S3; the enzyme involved decarboxylation step is illusive. Compound B may be reduced by one of the several potential alcohol dehydrogenases (listed in Supp. Table S2) allowing the alcohol compound passively diffuse through the membrane. Alternatively, the aldehyde itself can passively diffuse through the membrane. For the reducing potential, thiosulfate is imported via a specific thiosulfate transporter. Again, no obvious thiosulfate reductase was found in the annotation. However, Ars sulfite reductase is present for cycling SO3 and S2- concentrations. Therefore, the presence of the thiosulfate reductase is implied. 2-methyl-1-propanol is A; 2-methyl-propanal is B; 2-methyl-1-propionate is C; Dashed lines represent passive diffusion.
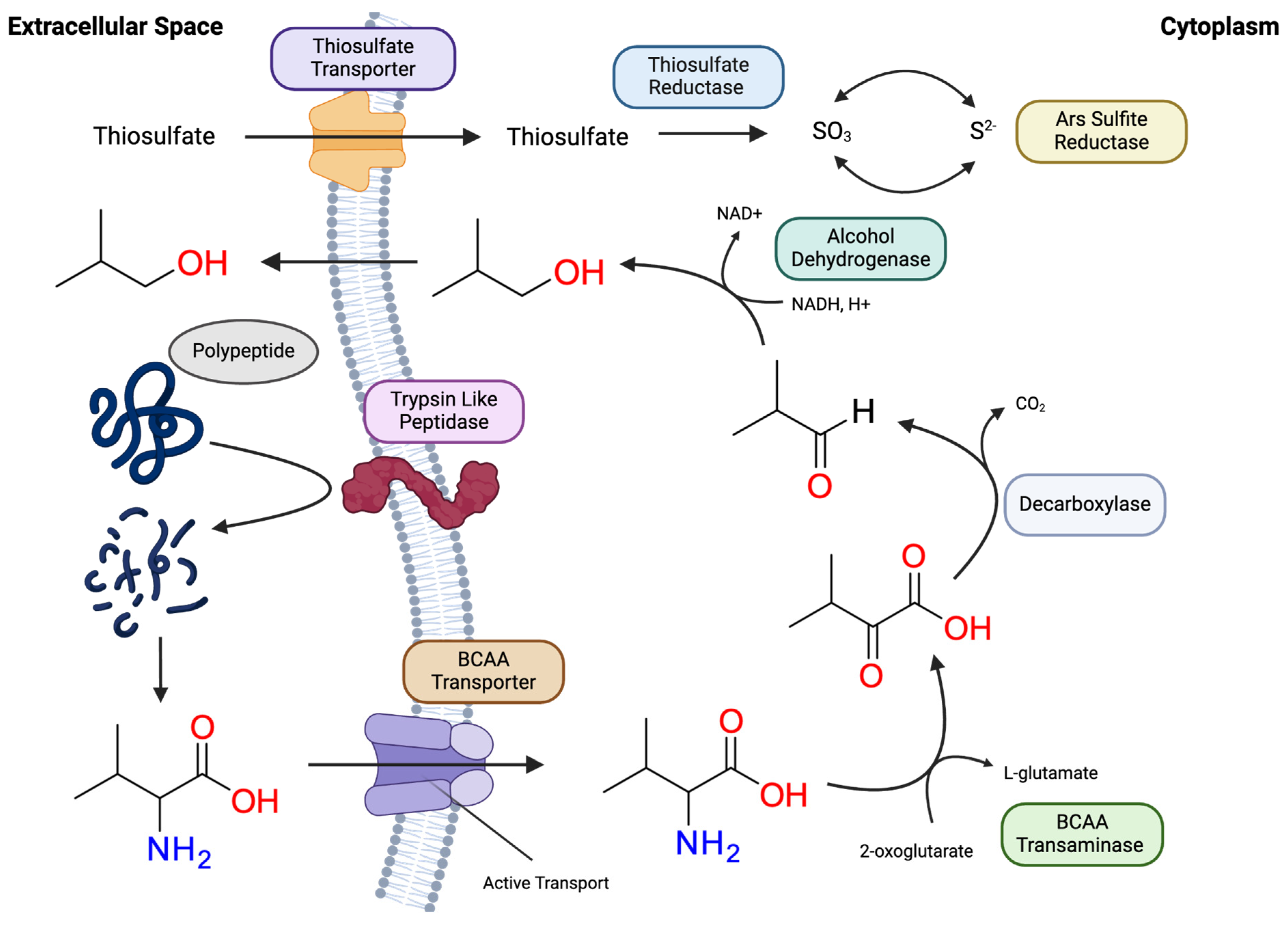

Table 1.
Comparison of end products formed from branched-chain amino acid fermentation by selected Thermoanaerobacter strains.
Table 1.
Comparison of end products formed from branched-chain amino acid fermentation by selected Thermoanaerobacter strains.
| Strain | Substrate | RCOOH (mM) | ROH (mM) | ROH/RCOOH ratio | Reference |
| T. mathranii (DSM 11426) | Leucine + S2O3 | 8.6 | 2.8 | 0.33 | This study |
| T. pseudethanolicus (DSM 2355) | Leucine + S2O3 | 10.3 | 1.2 | 0.12 | (Scully & Orlygsson, 2020) |
| Thermoanaerobacter brockii (DSM 1457) | Leucine + S2O3 | 13.6 | 1.5 | 0.11 | (Scully & Orlygsson, 2014) |
| T. ethanolicus (DSM 2246) | Leucine + S2O3 | 5.1 | 0.6 | 0.12 | (Scully et al., 2015) |
| Thermoanaerobacter strain AK68 | Leucine + S2O3 | 13.0 | 2.5 | 0.15 | (Scully et al., 2015) |
| Thermoanaerobacter strain AK85 | Leucine + S2O3 | 10.6 | 1.4 | 0.13 | (Scully & Orlygsson, 2019) |
| Thermoanaerobacter strain AK90 | Leucine + S2O3 | 9.7 | 3.5 | 0.36 | (Scully & Orlygsson, 2015) |
| Thermoanaerobacter strain AK152 | Leucine + S2O3 | 8.8 | 2.3 | 0.26 | (Scully et al., 2015) |
Table 2.
Comparison of selected genes present in T. mathranii and T. pseudethanolicus. Of important note is the lack of Aldehyde ferredoxin oxidoreductase (AOR) within T. mathranii. Indicating that the pyruvate fermentation mechanism must operate distinctly. Further, T. pseudethanolicus does not exhibit a BCAA transporter permease, while T. mathranii does. This is indicative that T. mathranii is able to utilize exogenous branched chain amino acids more readily than T. pseudethanolicus. Excluding the differences in AOR and BCAA transporters, both strains exhibit a similar genetic profile. Y and N indicates where the gene was found in the annotation for yes and no respectively. NA indicates that the gene was not found in any of the annotations. * indicates the gene has multiple subunits. [ ] are the total number of copies found within each respective strain.
Table 2.
Comparison of selected genes present in T. mathranii and T. pseudethanolicus. Of important note is the lack of Aldehyde ferredoxin oxidoreductase (AOR) within T. mathranii. Indicating that the pyruvate fermentation mechanism must operate distinctly. Further, T. pseudethanolicus does not exhibit a BCAA transporter permease, while T. mathranii does. This is indicative that T. mathranii is able to utilize exogenous branched chain amino acids more readily than T. pseudethanolicus. Excluding the differences in AOR and BCAA transporters, both strains exhibit a similar genetic profile. Y and N indicates where the gene was found in the annotation for yes and no respectively. NA indicates that the gene was not found in any of the annotations. * indicates the gene has multiple subunits. [ ] are the total number of copies found within each respective strain.
| Gene | Pathway(s) | T. mathranii | T. pseudethanolicus |
| Bifunctional aldehyde alcohol dehydrogenase | BCAA fermenation RCOOH reduction Ethanol fermentation |
Y | Y |
| Aldehyde ferredoxin oxidoreductase | RCOOH reduction Ethanol fermentation |
N | Y |
| Alcohol dehydrogenase | BCAA fermenation RCOOH reduction Ethanol fermentation |
Y [4] | Y [4] |
| BCAA transaminase | BCAA fermentation | Y | Y |
| BCAA ABC transporter permease | BCAA fermentation | Y | N |
| Oxaloacetate decarboxylase subunit alpha | BCAA fermentation | Y | Y |
| Butyrate kinase | RCOOH reduction | Y [2] | Y [2] |
| Butyryl phosphotransferase | RCOOH reduction | Y | Y |
| Pyruvate:ferredoxin oxidoreductase | Pyruvate fermenation | Y [2] | Y [2] |
| Acetate kinase | Pyruvate fermenation | Y | Y |
| Acetyl phosphotransferase | Pyruvate fermenation | Y | Y |
| YeeE/YedE thiosulfate transporter family protein | Thiosulfate utilization | Y [2] | Y [3] |
| Thiosulfate reductase | Thiosulfate utilization | NA | NA |
| Ars sulfite reductase* | Thiosulfate utilization | Y | Y |
| Trypsin-like peptidase | Protease | Y [2] | Y [2] |
* Indicates the gene has multiple subunits, [ ] are the total number of copies found within each respective.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



