
Submitted:
07 June 2024
Posted:
11 June 2024
You are already at the latest version
Abstract
Background: The incidence of metabolic dysfunction-associated steatohepatitis (MASH)-related hepatocellular carcinoma (HCC) is increasing annually with a rise in the incidence of obesity and metabolic syndrome. Based on preliminary reports regarding the potential association of HCC and periodontitis, this study aimed to analyze the involvement of periodontal bacteria as well as the oral and intestinal bacterial flora in MASH-related HCC (MASH-HCC). Methods: Forty-one patients with MASH and 19 with MASH-HCC who completed survey questionnaires, underwent periodontal examinations, and subjected to sample collection (saliva, mouth-rinsed water, feces, and peripheral blood) participated in the study. Bayesian network analysis was used to analyze the causation between various factors, including MASH-HCC, examinations, and bacteria. Results: The occupancy rate of the genus Fusobacterium in the intestinal bacterial flora was significantly higher in in the MASH-HCC group than in the MASH group (p = 0.002), whereas that of Butyricicoccus (p = 0.022) and Roseburia (p < 0.05) was significantly lower. Bayesian network analysis revealed the absence of periodontal pathogenic bacteria and enteric bacteria affecting HCC. However, HCC directly affected the periodontal bacterial species Porphyromonas gingivalis, Tannerella forsythia, Fusobacterium nucleatum, and Prevotella intermedia in the saliva, as well as the genera Lactobacillus, Roseburia, Fusobacterium, Prevotella, Clostridium, Ruminococcus, Trabulsiella, and SMB53 in the intestine. Furthermore, P. gingivalis in the oral cavity directly affected the genera Lactobacillus and Streptococcus in the intestine. Conclusion: MASH-HCC directly affects periodontal pathogenic and intestinal bacteria, and P. gingivalis may affect the intestinal bacteria associated with gastrointestinal cancer.
Keywords:
microbiota
; oral bacteria
; intestinal bacteria
; hepatocellular cancer
; metabolic dysfunction-associated steatohepatitis
; Porphyromonas gingivalis
1. Introduction
Hepatocellular carcinoma (HCC) evolves from chronic liver disease and accounts for 85% of all primary liver cancers [1]. Viral hepatitis was the primary cause, whereas advances in treatment have gradually lowered its incidence [2]. Meanwhile, the incidence of HCC (MASH-HCC) related to metabolic dysfunction-associated steatohepatitis (MASH, formerly known as NASH) [3] has increased over the years owing to an increase in the incidence of obesity and metabolic syndrome [4]. However, the pathogenesis of MASH-HCC remains unclear.
To date, the mechanism by which periodontal bacteria affect the entire body is hypothesized to be as follows: bacteria enter the bloodstream directly from periodontal pocket ulceration and affect organs outside the oral cavity [5,6]. In recent studies, another mechanism has been identified: oral bacteria in saliva are transferred enterally to the intestine by swallowing and affect the intestinal bacterial flora and metabolism [7,8]. The liver is an organ of the digestive system that is anatomically and physiologically connected to the enterohepatic circulation via the portal vein. Hence, periodontal bacteria and lipopolysaccharides (LPS) derived from periodontal bacteria in the saliva may be implicated in the pathogenic mechanism of MASH-HCC by affecting the intestinal flora.
In recent years, epidemiological reports have shown that periodontal pathogen is a risk factor in onset of various cancers and cancer-associated mortality [9]. Fusobacterium nucleatum was specifically detected in organs of the digestive system that are anatomically close to the oral cavity [10,11,12], and the presence of Porphyromonas gingivalis was correlated with the malignancy of esophageal cancer [13]. Only a few reports have shown an association between HCC and periodontal pathogen, such as high circulating reactive oxygen species levels in patients with HCC and periodontitis [14]. As far as we are aware, our recent report showing an association between MASH-HCC and salivary P. gingivalis, F. nucleatum, and IgA is the only report to date highlighting a relationship between MASH-HCC and periodontopathic bacteria [15].
Based on these reports, we hypothesized that MASH-HCC is associated with periodontopathic bacteria in the oral cavity. This study aimed to analyze the clinical parameters and oral and intestinal bacterial flora in patients with MASH and MASH-HCC to determine the relationship between MASH-HCC and periodontal bacteria.
2. Materials and Methods
2.1. Participants
Participants in this study included patients with MASH and MASH-HCC aged 20 years or older who attended or were admitted to the Department of Gastroenterology at Yokohama City University (YCU) Hospital between November 2020 and April 2022. Those who were taking antimicrobials within one month prior to periodontal examination and those with edentulous jaws were excluded from the study. This study was approved by the research ethics committee of Kanagawa Dental University (KDU) and YCU and was conducted at YCU Hospital in compliance with the Declaration of Helsinki. All participants were informed of the purpose, outline, safety, and protection of personal information of this study, and their written consent to participate in the study based on their free will was obtained. Initially, sixty-nine participants were enrolled, and data from sixty participants (forty-one with MASH and nineteen with MASH-HCC) for whom all testing and sample collection data were available were used for the analysis.
2.2. Background Information
The participants' gender, age and smoking status were interviewed using a questionnaire form. The dentist filled out the response form based on the participants’ responses. Body mass index (BMI) was calculated from the medical records by obtaining the height and weight values closest to the date of periodontal examination.
2.3. Periodontal Examination
Periodontal examinations were performed by two dentists from the Department of Periodontology at KDU. The probing depth and bleeding on probing were measured at the six probing points per tooth. The plaque index was recorded at the four points per tooth, and tooth mobility was evaluated. Probing was performed at constant pressure using a plastic probe (Contact Probe, Nihon Dental Laboratory Co., Ltd., Tokyo, Japan) with a probing pressure of 0.2 N. The dentists calibrated their probing tools in advance. A periodontal jaw model (P15FE-500HPRO-S2A1-GSF, Nissin, Kyoto, Japan) was used for calibration.
2.4. Sample Collection
Saliva samples used for IgA concentration assay were collected using the SALIVETⓇ (SARSTEDT, Nümbrecht, Germany). A polypropylene-polyethylene polymer sponge was held under the tongue for 2 min, and the saliva-containing sponge was returned to the tube. The tubes were quickly ice-cooled, centrifuged (1,200 × g, 20 min, 4 °C), and stored at –80 °C until analysis. Mouth-rinsed water was used to analyze the oral microbiota. Participants rinsed with saline for 10 s and then collected into tubes. The rinsed water was stored at –80 °C until analysis. Fecal samples were collected from the participants using a Mykinso fecal collection kitⓇ (Cykinso, Inc., Tokyo, Japan) containing a guanidine thiocyanate solution. Fecal samples were collected by the participants themselves according to the manufacturer's manual.
2.5. Medical Examination
Peripheral blood samples were collected on the same day as periodontal examination. Endotoxin, high-sensitivity C-reactive protein (CRP), aspartate transaminase (AST), alanine aminotransferase (ALT), and total bilirubin (T-Bil) levels were analyzed.
2.6. Assay of IgA Concentration in Saliva
IgA concentration in saliva were determined by enzyme-linked immunosorbent assay (ELISA) using a Human IgA ELISA Kit (Bethyl Laboratories, Inc., Montgomery, TX, USA). The ELISA was performed according to the manufacturer’s instructions.
2.7. DNA Preparation and Microbiota Analysis
DNA extraction and bacterial flora analysis of the mouth-rinsed water and fecal samples were performed at the Medical Laboratory (Cykinso, Inc.) [16,17]. DNA was extracted using an automated DNA extraction machine (GENE PREP STAR PI-1200A; Kurabo Industries Ltd., Osaka, Japan) according to the manufacturer’s protocol. Detailed sequencing methods are described in previous report [18]. Data processing and assignment were performed using the QIIME2 pipeline (version 2020.8), and based on the work of Fujihara et al. [19].
2.8. Bayesian Network Analysis and Classification Trees
A Bayesian network is a directed acyclic graph composed of a set of variables {X1, X2,...,XN} and a set of directed edges between them [20]. The details of the analytical methods are described in our previous report [21]. Because the Bayesian network could not be analyzed with missing values, we excluded one participant from the MASH group who had missing T-Bil values, and data from fifty-nine participants (forty patients with MASH and nineteen patients with MASH-HCC) were used for the analysis. Based on the results of Bayesian network analysis, a classification tree analysis was performed using rpart.
2.9. Statistical Analysis
Statistical analyses were performed using SPSS Statistics (version 27.0; IBM, Tokyo, Japan) and R (version 3.5.1 (The R Project for Statistical Computing, Vienna, Austria, 2018). The Mann–Whitney’s U test was used for comparisons between two groups, except for gender and smoking status, which were verified by χ2 test. The Spearman's rank correlation coefficient was used for the correlation analysis. Statistical significance was set at p < 0.05.
3. Results
3.1. Participant's Information, Periodontal and Medical Examinations, and Salivary IgA Levels
Table 1 shows information on the participants, periodontal and medical status, and IgA concentration in saliva (the data is also reported in our previous study [15]). Compared with the MASH group, the MASH-HCC group was significantly older (p = 0.0004). Both groups showed similar periodontal examination results. The salivary IgA concentration was significantly lower in the MASH-HCC group than in the MASH group (p < 0.001). Endotoxin and T-Bil levels were significantly higher in the MASH-HCC group (p < 0.0001) than in the MASH group (p = 0.014).
3.2. Diversity and Composition of Bacterial Flora in the Saliva
Table 2 shows the diversity and abundance of the salivary microbiome. All bacterial phyla detected, 23 bacterial genera with an average occupancy greater than 0.5%, and 22 bacterial species with an average occupancy greater than 0.1% are presented. The Shannon index of the salivary microflora was significantly lower in the MASH-HCC group than in the MASH group (p = 0.03). Regardless of liver disease type, the predominant microorganisms at the phylum level in all participants were Bacillota, Bacteroidota, Pseudomonadota, Actinomycetota and Fusobacteriota. Similarly, the genera Streptococcus, Prevotella, Veillonella and Actinomyces predominated in both groups. Their occupancy rate was not significantly different between the two groups.
The proportions of P. gingivalis and F. nucleatum were higher in the MASH-HCC group than in the MASH group. However, only F. nucleatum showed a significant difference (p = 0.014). Conversely, the occupancy rate of Treponema denticola was significantly lower in the MASH-HCC group than in the MASH group (p = 0.02).
3.3. Diversity and Composition of Bacterial Flora in the Feces
Table 3 shows the diversity and abundance of bacterial flora in feces, including all bacterial phyla detected and 23 bacterial genera with top occupancy. The Shannon index was significantly lower in the MASH-HCC group than in the MASH group (p < 0.001). At the phylum level, both groups were dominated by Bacillota, Bacteroidota, Actinomycetota, and Pseudomonadota. However, occupancy rates were comparable between the two groups. Only Fusobacteriota was significantly more prevalent in the MASH-HCC group than in the MASH group (p = 0.002).
At the genus level, Bacteroides and Blautia dominated all participants, but their occupancy rates were comparable between the two groups. The occupancy rates of Butyricicoccus (p = 0.022) and Roseburia (p < 0.05) in the MASH-HCC group were significantly lower than those in the MASH group. Conversely, the occupancy rate of Fusobacterium in the MASH-HCC group was significantly higher than in the MASH group (p = 0.002).
3.4. Determination of Causal Effects Using Bayesian Network Analysis
Figure 1 shows the results of the Bayesian network analysis. We focused on the items that showed significant differences in the comparison between the two groups (Table 1, Table 2 and Table 3) and added the major oral periodontopathic bacterial species and the top 23 enterobacterial genera (Table 3), setting them as factors in the Bayesian network analysis.
The presence of HCC directly affected the following major periodontal bacterial species: P. gingivalis, Tannerella forsythia, F. nucleatum, and Prevotella intermedia in the saliva. It also affects salivary IgA concentrations. Furthermore, salivary IgA concentrations affect Prevotella intermedia in saliva.
HCC also directly affected the genera Lactobacillus, Roseburia, Fusobacterium, Prevotella, Clostridium, Ruminococcus, Trabulsiella, and SMB53 in the feces. In addition, P. gingivalis in the saliva directly affected Lactobacillus and Streptococcus in the feces and indirectly affected Blautia and Butyricicoccus. Moreover, salivary F. nucleatum affected Serratia in feces. Meanwhile, T-Bil level and age had a direct impact on HCC. The genus Oscillospira in the feces affected T-Bil.
3.5. Classification Tree to Assess Disease Type
The Bayesian network results showed that the two factors affecting HCC were T-Bil level and age. When the dependent variable was set as the presence or absence of HCC and the explanatory variables were set as T-Bil and age in a classification tree (Figure 2), the major factor affecting HCC was T-Bil, followed by age. HCC develops when T-Bil exceeds 1.35 mg/dl in the MASH. However, even if the T-Bil is less than 1.35 in MASH, HCC develops when the patient is over 77 years of age.
4. Discussion
This is the first study to analyze the relationship between oral periodontal pathogenic bacteria and intestinal bacteria in patients with MASH and MASH-HCC. In this study, the presence of HCC directly affected several periodontopathogenic bacteria in the saliva. In addition, a higher abundance of F. nucleatum in saliva was observed in the MASH-HCC group than in the MASH group. It has been reported that oral bacterial flora changes in patients with pancreatic cancer [22], and the presence of F. nucleatum in the oral cavity is elevated in patients with lung and colorectal cancers [23,24]. Cancer occurs when the systemic immune response decreases [25], and immune function clearly decreases in patients with cancer [26]. In addition, since the type and number of oral bacteria are related to systemic immune status [27], in this study, the decline in systemic immune function caused by MASH-HCC may have affected periodontal bacteria in the oral cavity, which is a remote organ.
The abundance of salivary F. nucleatum in the MASH-HCC group was higher than that of other periodontal pathogenic bacteria, except for P. gingivalis. F. nucleatum is an opportunistic bacterium present in the oral cavity of individuals without periodontal disease [28]. In addition, Leigh et al. [29] reported that opportunistic bacteria in the oral cavity increase owing to a decline in immune function. Hence, although there was no difference in periodontal conditions between the two groups in this study, the MASH-HCC group had decreased systemic immune function; therefore, the occupancy rate of F. nucleatum in the oral cavity may have been high.
Bayesian network analysis revealed that HCC directly affected several fecal bacteria; however, none of the fecal bacteria directly affected HCC. Among the intestinal bacteria directly affected by HCC, the genus Roseburia was found to have a lower abundance in the MASH-HCC group than in the MASH group, whereas the genus Fusobacterium had a higher abundance. Various studies have reported that gastrointestinal cancer is associated with the intestinal microbiome. The genus Roseburia was decreased in the intestinal microbiota of patients with colorectal and pancreatic cancer [30,31]. Although there is a known case of MASH that developed into MASH-HCC through liver cirrhosis [32], Roseburia occupancy was decreased in the intestinal microbiota of patients with liver cirrhosis [33]. The abundance of the genus Fusobacterium increases in the feces of patients with colorectal cancer [34,35]. The abundance of Fusobacterium increases, and that of Roseburia decreases when dysbiosis occurs in the intestinal microbiota [36]. The liver and intestinal microbiomes have a close bidirectional relationship [37], and dysbiosis occurs in patients with liver cancer [38]. Moreover, the Shannon Index, which shows the diversity of the intestinal microflora, decrease owing to dysbiosis [39]. The Shannon index was lower in the MASH-HCC group than in the MASH group in this study. Hence, it is likely that the MASH-HCC group in the present study had more advanced dysbiosis.
High-fat, high-glucose, and low-fiber Western diets are known to accelerate progression from MASH to MASH-HCC [40]. In addition, the Western diet increases the abundance of the genus Fusobacterium and decreases the abundance of the genus Roseburia in the intestine [41]. Thus, high-fat, high-glucose, and low-fiber diets that cause HCC may also affect the intestinal bacteria.
Our results showed that the genus Fusobacterium in feces did not affect HCC, and F. nucleatum in the saliva did not affect the genus Fusobacterium in feces. Guo et al. reported that F. nucleatum is increased in hepatocellular carcinoma tissues and that hepatocellular carcinoma is affected by F. nucleatum because methyltransferase-like protein 3 expression during F. nucleatum infection is involved in tumor progression [42]. Although not revealed in the present study, intestinal bacteria may affect HCC.
Primary bile acids increase in MASH-HCC, and Lactobacillus, which metabolizes them, has been reported to increase in the intestine [43]. Therefore, Bayesian network analysis revealed a direct effect of MASH-HCC on Lactobacillus spp in feces. Interestingly, not only HCC but also P. gingivalis in the saliva directly affected the genus Lactobacillus in feces. P. gingivalis is a typical periodontal pathogenic bacterium in the oral cavity that can affect intestinal bacteria and cause dysbiosis [44]. Park et al. [45] found that mice infected with P. gingivalis in the oral cavity showed increased levels of the intestinal phyla Actinobacteria and Deferribacteres. In addition, Nakajima et al. [46] reported that a single oral dose of P. gingivalis administered to mice increased Bacteroidetes and decreased Firmicutes in the intestine. Oral administration of P. gingivalis causes changes in the intestinal microbiota, impairs intestinal barrier function, and damages the liver [46]. Although the difference was not significant, the occupancy rate of salivary P. gingivalis was higher in the MASH-HCC group. Hence, in addition to HCC, the high abundance of P. gingivalis in saliva could have damaged the liver and altered the amount of primary bile acids, which could have affected the genus Lactobacillus.
We found that P. gingivalis in the saliva had a direct effect on the genus Streptococcus in the feces. In patients with atrophic gastritis in the gastric corpus who are at a high risk of gastric cancer, an increase in Streptococcus spp. in the stomach [47] and in the feces of patients with CRC [48] has been reported, and the genus Streptococcus is associated with digestive disorders. Thus, in the MASH-HCC group, an increase in P. gingivalis in the oral cavity can cause dysbiosis in the intestine, which may have affected Streptococcus spp.
The causal analysis indicated that salivary P. gingivalis caused a decrease in Blautia and Bacteroides via the genus Lactobacillus and Butyricicoccus via the genus Streptococcus. Studies have shown an association between these three intestinal bacteria and gastrointestinal cancer. Blautia spp. are decreased in the feces of liver cancer patients [49], Bacteroides spp. are decreased in the feces of mice that developed liver cancer due to a high-fat, high-cholesterol diet [50], and Butyricicoccus spp. are decreased in the feces of patients with esophageal cancer [51]. All of these bacterial genera are short-chain fatty acid (SCFA) producers [52,53,54]. McBrearty et al. [55] reported that since SCFAs have strong anti-inflammatory and antitumor effects, the administration of SCFA to mice delayed the development of hepatocellular carcinoma. These bacterial genera did not directly affect HCC but may have affected HCC via SCFA. Hence, reducing P. gingivalis in the oral cavity, which indirectly affects these three intestinal bacteria, may help prevent the development of MASH-HCC.
Our data demonstrated that salivary F. nucleatum affected the fecal Serratia spp., which is an opportunistic bacterium like F. nucleatum [27,56]. Lin et al. [57] reported increased levels of both oral F. nucleatum and gut opportunistic bacteria of a mouse model of ulcerative colitis. In the present study, patients with MASH-HCC would have had a generalized state of weakened immune system that made them susceptible to an increase in both the opportunistic bacteria F. nucleatum and the genus Serratia. Therefore, our results show that F. nucleatum in the oral cavity directly affects Serratia.
Interestingly, salivary IgA concentrations only affected P. intermedia in the saliva. Salivary IgA levels increase with the number of periodontal pathogenic bacteria and control them [58,59,60]. Despite this, the fact that salivary IgA concentration only affected P. intermedia in this study suggests that the effect of HCC on periodontal pathogenic bacteria in the oral cavity was greater than that of the salivary IgA concentration.
Of the two factors directly affecting HCC, one was blood T-Bil level, which was directly affected by fecal Oscillospira spp. T-Bil levels increase as liver function declines in patients with liver cancer [61]. Increased T-Bil levels have also been reported in rats with liver cancer [62]. Therefore, it is likely that the MASH-HCC group in this study showed a decline in liver function, resulting in high T-Bil levels. Furthermore, an increase in secondary bile acids produced by intestinal bacteria decreases liver function [63], and the level of secondary bile acids in feces is positively correlated with genus Oscillospira in feces [64]. The genus Oscillospira may have affected the increase in T-Bil levels by reducing liver function through secondary bile acids.
Age is another factor that directly affects HCC development. The median age of the patients in the MASH-HCC group was higher than that in the MASH group. Recently, Shimomura et al. [65] reported that patients with MASH-HCC were older and had lower antioxidant function than patients with MASH and that oxidative stress correlated with MASH activation markers, both of which were increased. However, young patients had lower levels of MASH activation markers because their antioxidant functions were preserved [65]. Hence, old age may be a major risk factor for MASH development.
The results of the classification tree analysis suggested that a T-Bil of 1.35 mg/ dL or higher in the MASH group was related to the occurrence of HCC. HCC occurrences at the age of 77 years or older, even when T-Bil is less than 1.35 mg/dL in MASH. Therefore, MASH patients with T-Bil of 1.35 mg/dL or higher, or older MASH patients with T-Bil less than 1.35 mg/dL, may require more medical assistance to prevent them from developing HCC. High T-Bil levels are the major factors affecting HCC. The genus Oscillospira, which elevates T-Bil levels, increases when dysbiosis occurs in the intestines [66]. Because P. gingivalis in the oral cavity causes intestinal dysbiosis [46], patients with MASH may require periodontal management to suppress the abundance of P. gingivalis in the oral cavity to prevent dysbiosis.
Limitations
The current study has several limitations. First, the number of participants was low. This is because it was difficult to recruit participants who met the inclusion criteria. Therefore, the number of participants in the two groups could not be matched. Second, differences were observed in the ages of participants in the MASH and MASH-HCC groups. Future studies will need to set the age of the participants higher in order to keep the age of both groups the same.
5. Conclusions
MASH-HCC directly affects periodontal pathogenic bacteria, salivary IgA, and intestinal bacteria. P. gingivalis may, directly and indirectly, affect the intestinal bacteria associated with gastrointestinal cancer.
Author Contributions
Conceptualization, M.M.; formal analysis, Y.Y. and Y.N.; investigation, T.M. (Takaaki Matsui) and R.T.; resources, T.K. (Takashi Kobayashi), M.Y., A.N. (Asako Nogami), T.K. (Takaomi Kessoku), M.T. (Muneaki Tamura) and Y.K.; data curation, T.M. (Toshiya Morozumi); writing—original draft preparation, T.M. (Takaaki Matsui), T.M. (Toshiya Morozumi), Y.Y. and T.K. (Takashi Kobayashi); writing—review and editing, M.Y. and N.A.; visualization, T.T. and K.A.; supervision, S.S., K.M., A.N. (Atsushi Nakajima) and M.K.; project administration, T.M. (Toshiya Morozumi); funding acquisition, M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by JSPS KAKENHI (grant number 19K10454).
Institutional Review Board Statement
This study was approved by the Ethics Committees of Kanagawa Dental University (No. 693) and Yokohama City University (B200100001).
Informed Consent Statement
Written informed consent was obtained from all participants involved in the study.
Data Availability Statement
All data obtained in this study is described in the revised manuscript.
Acknowledgments
The authors thank the clinical staff of the Yokohama City University Hospital for their assistance.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nat. Rev. Dis. Primers 2021, 7, 6. [Google Scholar] [CrossRef]
- Zopf, S.; Kremer, A.E.; Neurath, M.F.; Siebler, J. Advances in hepatitis C therapy: What is the current state—What come’s next? World J. Hepatol. 2016, 8, 139–147. [Google Scholar] [CrossRef]
- Rinella, M.E.; Lazarus, J.V.; Ratziu, V.; Francque, S.M.; Sanyal, A.J.; Kanwal, F.; Romero, D.; Abdelmalek, M.F.; Anstee, Q.M.; Arab, J.P.; et al. A multisociety Delphi consensus statement on new fatty liver disease nomenclature. J. Hepatol. 2023, 79, 1542–1556. [Google Scholar] [CrossRef]
- Rumgay, H.; Arnold, M.; Ferlay, J.; Lesi, O.; Cabasag, C.J.; Vignat, J.; Laversanne, M.; McGlynn, K.A.; Soerjomataram, I. Global burden of primary liver cancer in 2020 and predictions to 2040. J. Hepatol. 2022, 77, 1598–1606. [Google Scholar] [CrossRef] [PubMed]
- Whittle, E.; Leonard, M.O.; Harrison, R.; Gant, T.W.; Tonge, D.P. Multi-method characterization of the human circulating microbiome. Front. Microbiol. 2018, 9, 3266. [Google Scholar] [CrossRef]
- Emery, D.C.; Cerajewska, T.L.; Seong, J.; Davies, M.; Paterson, A.; Allen-Birt, S.J.; West, N.X. Comparison of blood bacterial communities in periodontal health and periodontal disease. Front. Cell. Infect. Microbiol. 2020, 10, 577485. [Google Scholar] [CrossRef] [PubMed]
- Arimatsu, K.; Yamada, H.; Miyazawa, H.; Minagawa, T.; Nakajima, M.; Ryder, M.I.; Gotoh, K.; Motooka, D.; Nakamura, S.; Iida, T.; et al. Oral pathobiont induces systemic inflammation and metabolic changes associated with alteration of gut microbiota. Sci. Rep. 2014, 4, 4828. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, K.; Kamada, N. Exploring the oral-gut linkage: Interrelationship between oral and systemic diseases. Mucosal Immunol. <italic>in press</italic>. 2023. [Google Scholar] [CrossRef]
- Nwizu, N.; Wactawski-Wende, J.; Genco, R.J. Periodontal disease and cancer: Epidemiologic studies and possible mechanisms. Periodontol. 2000 2020, 83, 213–233. [Google Scholar] [CrossRef]
- Kostic, A.D.; Gevers, D.; Pedamallu, C.S.; Michaud, M.; Duke, F.; Earl, A.M.; Ojesina, A.I.; Jung, J.; Bass, A.J.; Tabernero, J.; et al. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res. 2012, 22, 292–298. [Google Scholar] [CrossRef]
- Boehm, E.T.; Thon, C.; Kupcinskas, J.; Steponaitiene, R.; Skieceviciene, J.; Canbay, A.; Malfertheiner, P.; Link, A. Fusobacterium nucleatum is associated with worse prognosis in Lauren’s diffuse type gastric cancer patients. Sci. Rep. 2020, 10, 16240. [Google Scholar] [CrossRef] [PubMed]
- Parhi, L.; Alon-Maimon, T.; Sol, A.; Nejman, D.; Shhadeh, A.; Fainsod-Levi, T.; Yajuk, O.; Isaacson, B.; Abed, J.; Maalouf, N.; et al. Breast cancer colonization by Fusobacterium nucleatum accelerates tumor growth and metastatic progression. Nat. Commun. 2020, 11, 3259. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Li, S.; Ma, Z.; Liang, S.; Shan, T.; Zhang, M.; Zhu, X.; Zhang, P.; Liu, G.; Zhou, F.; et al. Presence of Porphyromonas gingivalis in esophagus and its association with the clinicopathological characteristics and survival in patients with esophageal cancer. Infect. Agent Cancer 2016, 11, 3. [Google Scholar] [CrossRef] [PubMed]
- Tamaki, N.; Takaki, A.; Tomofuji, T.; Endo, Y.; Kasuyama, K.; Ekuni, D.; Yasunaka, T.; Yamamoto, K.; Morita, M. Stage of hepatocellular carcinoma is associated with periodontitis. J. Clin. Periodontol. 2011, 38, 1015–1020. [Google Scholar] [CrossRef]
- Takuma, R.; Morozumi, T.; Yamamoto, Y.; Kobayashi, T.; Matsui, T.; Yoneda, M.; Kessoku, T.; Nogami, A.; Tamura, M.; Kamata, Y.; et al. Association between non-alcoholic steatohepatitis-related hepatocellular carcinoma and periodontopathic bacteria: A cross-sectional pilot study. Appl. Sci. 2023, 13, 3893. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Watanabe, S.; Kameoka, S.; Shinozaki, NO.; Kubo, R.; Nishida, A.; Kuriyama, M.; Takeda, AK. A cross-sectional analysis from the Mykinso Cohort Study: establishing reference ranges for Japanese gut microbial indices. Biosci Microbiota Food Health. 2021, 40, 123–134. [Google Scholar] [CrossRef]
- Fujihara, H.; Matsunaga, M.; Ueda, E.; Kajiwara, T.; Takeda, AK.; Watanabe, S.; Baba, K.; Hagihara, K.; Myowa, M. Altered Gut Microbiota Composition Is Associated with Difficulty in Explicit Emotion Regulation in Young Children. Microorganisms. 2023, 11, 2245. [Google Scholar] [CrossRef] [PubMed]
- Maglogiannis, I.; Zafiropoulos, E.; Platis, A.; Lambrinoudakis, C. Risk analysis of a patient monitoring system using Bayesian Network modeling. J. Biomed. Inform. 2006, 39, 637–647. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Morozumi, T.; Hirata, T.; Takahashi, T.; Fuchida, S.; Toyoda, M.; Nakajima, S.; Minabe, M. Effect of periodontal disease on diabetic retinopathy in type 2 diabetic patients: A cross-sectional pilot study. J. Clin. Med. 2020, 9, 3234. [Google Scholar] [CrossRef] [PubMed]
- Nagata, N.; Nishijima, S.; Kojima, Y.; Hisada, Y.; Imbe, K.; Miyoshi-Akiyama, T.; Suda, W.; Kimura, M.; Aoki, R.; Sekine, K.; et al. Metagenomic identification of microbial signatures predicting pancreatic cancer from a multinational study. Gastroenterology 2022, 163, 222–238. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Lu, J.; Beck, J.D.; Moss, K.L.; Prizment, A.E.; Demmer, R.T.; Porosnicu Rodriguez, K.A.; Joshu, C.E.; Michaud, D.S.; Platz, E.A. Periodontal and other oral bacteria and risk of lung cancer in the atherosclerosis risk in communities (ARIC) study. Cancer Epidemiol. Biomarkers Prev. 2023, 32, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Tortora, S.C.; Bodiwala, V.M.; Quinn, A.; Martello, L.A.; Vignesh, S. Microbiome and colorectal carcinogenesis: Linked mechanisms and racial differences. World J. Gastrointest. Oncol. 2022, 14, 375–395. [Google Scholar] [CrossRef] [PubMed]
- Fulop, T.; Larbi, A.; Witkowski, J.M.; Kotb, R.; Hirokawa, K.; Pawelec, G. Immunosenescence and cancer. Crit. Rev. Oncog. 2013, 18, 489–513. [Google Scholar] [CrossRef]
- Sheng, C.C.; Han, F.Y. Immunoregulation effects of TIM-3 on tumors. Neoplasma 2019, 66, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Jungbauer, G.; Stähli, A.; Zhu, X.; Auber Alberi, L.; Sculean, A.; Eick, S. Periodontal microorganisms and Alzheimer disease – A causative relationship? Periodontol. 2000 2022, 89, 59–82. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Shi, T.; Li, Y.; Huang, L.; Yin, D. Fusobacterium nucleatum: The opportunistic pathogen of periodontal and peri-Implant diseases. Front. Microbiol. 2022, 13, 860149. [Google Scholar] [CrossRef] [PubMed]
- Leigh, J.E.; Shetty, K.; Fidel, P.L. Jr. Oral opportunistic infections in HIV-positive individuals: Review and role of mucosal immunity. AIDS Patient Care STDs 2004, 18, 443–456. [Google Scholar] [CrossRef]
- Borges-Canha, M.; Portela-Cidade, J.P.; Dinis-Ribeiro, M.; Leite-Moreira, A.F.; Pimentel-Nunes, P. Role of colonic microbiota in colorectal carcinogenesis: A systematic review. Rev. Esp. Enferm. Dig. 2015, 107, 659–671. [Google Scholar] [CrossRef]
- Zhou, W.; Zhang, D.; Li, Z.; Jiang, H.; Li, J.; Ren, R.; Gao, X.; Li, J.; Wang, X.; Wang, W.; et al. The fecal microbiota of patients with pancreatic ductal adenocarcinoma and autoimmune pancreatitis characterized by metagenomic sequencing. J. Transl. Med. 2021, 19, 215. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Kawaguchi, M.; Mikami, H.; Diao, P.; Zhang, X.; Zhang, Z.; Nakajima, T.; Iwadare, T.; Kimura, T.; Nakayama, J.; et al. Establishment of novel mouse model of dietary NASH rapidly progressing into liver cirrhosis and tumors. Cancers 2023, 15, 3744. [Google Scholar] [CrossRef] [PubMed]
- Nikkhah, A.; Ejtahed, H.S.; Ettehad Marvasti, F.; Taghavi, M.; Pakmehr, A.; Hajipour, F.; Larijani, B. The critical role of gut microbiota dysbiosis in skeletal muscle wasting: A systematic review. J. Appl. Microbiol. 2023, 134, lxac014. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.; Yang, X.; Zhang, R.; Li, J.; Xiao, X.; Hu, Y.; Chen, Y.; Yang, F.; Lu, N.; Wang, Z.; et al. Dysbiosis signature of fecal microbiota in colorectal cancer patients. Microb. Ecol. 2013, 66, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Tesolato, S.; Ortega-Hernández, A.; Gómez-Garre, D.; Claver, P.; De Juan, C.; De la Serna, S.; Paz, M.; Domínguez-Serrano, I.; Dziakova, J.; Rivera, D.; et al. Gut microbiota profiles in feces and paired tumor and non-tumor tissues from colorectal cancer patients. Relationship to the body mass index. PLOS ONE 2023, 18, e0292551. [Google Scholar] [CrossRef]
- Liu, P.; Yang, J.; Chen, Y.; Zhu, Y.; Tang, Y.; Xu, X.; He, H. Alterations of gut microbiota and metabolome in early chronic kidney disease patients complicated with hyperuricemia. Heliyon 2023, 9, e20328. [Google Scholar] [CrossRef]
- Rajapakse, J.; Khatiwada, S.; Akon, A.C.; Yu, K.L.; Shen, S.; Zekry, A. Unveiling the complex relationship between gut microbiota and liver cancer: Opportunities for novel therapeutic interventions. Gut Microbes 2023, 15, 2240031. [Google Scholar] [CrossRef]
- Huang, C.; Mei, S.; Zhang, X.; Tian, X. Inflammatory milieu related to dysbiotic gut microbiota promotes tumorigenesis of hepatocellular carcinoma. J. Clin. Gastroenterol. 2023, 57, 782–788. [Google Scholar] [CrossRef]
- Lee, S.J.; Kim, D.; Ann, H.W.; Han, M.; Lee, J.A.; Lee, Y.; Ahn, S.; Seo, H.W.; Kim, J.H.; Ahn, J.Y.; et al. Deciphering gut microbiota in patients with severe sepsis and septic shock. Shock 2024, 61, 28–33. [Google Scholar] [CrossRef]
- Hymel, E.; Vlock, E.; Fisher, K.W.; Farazi, P.A. Differential progression of unhealthy diet-induced hepatocellular carcinoma in obese and non-obese mice. PLOS ONE 2022, 17, e0272623. [Google Scholar] [CrossRef]
- Yang, L.; Li, Y.; Wang, S.; Bian, X.; Jiang, X.; Wu, J.; Wang, K.; Wang, Q.; Xia, J.; Jiang, S.; et al. Western diet aggravated carbon tetrachloride-induced chronic liver injury by disturbing gut microbiota and bile acid metabolism. Mol. Nutr. Food Res. 2021, 65, e2000811. [Google Scholar] [CrossRef]
- Guo, S.; Chen, F.; Li, L.; Dou, S.; Li, Q.; Huang, Y.; Li, Z.; Liu, W.; Zhang, G. Intracellular Fusobacterium nucleatum infection increases METTL3-mediated m6A methylation to promote the metastasis of esophageal squamous cell carcinoma. J. Adv. Res. in press. 2023. [Google Scholar] [CrossRef] [PubMed]
- Sydor, S.; Best, J.; Messerschmidt, I.; Manka, P.; Vilchez-Vargas, R.; Brodesser, S.; Lucas, C.; Wegehaupt, A.; Wenning, C.; Aßmuth, S.; et al. Altered microbiota diversity and bile acid signaling in cirrhotic and noncirrhotic NASH-HCC. Clin. Transl. Gastroenterol. 2020, 11, e00131. [Google Scholar] [CrossRef] [PubMed]
- Nasiri, K.; Amiri Moghaddam, M.; Etajuri, E.A.; Badkoobeh, A.; Tavakol, O.; Rafinejad, M.; Forutan Mirhosseini, A.; Fathi, A. Periodontitis and progression of gastrointestinal cancer: Current knowledge and future perspective. Clin. Transl. Oncol. 2023, 25, 2801–2811. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, I.; Han, S.J.; Kwon, S.; Min, E.J.; Cho, W.; Koh, H.; Koo, B.N.; Lee, J.S.; Kwon, J.S.; et al. Oral Porphyromonas gingivalis infection affects intestinal microbiota and promotes atherosclerosis. J. Clin. Periodontol. 2023, 50, 1553–1567. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, M.; Arimatsu, K.; Kato, T.; Matsuda, Y.; Minagawa, T.; Takahashi, N.; Ohno, H.; Yamazaki, K. Oral administration of P. gingivalis induces dysbiosis of gut microbiota and impaired barrier function leading to dissemination of enterobacteria to the liver. PLOS ONE 2015, 10, e0134234. [Google Scholar] [CrossRef] [PubMed]
- Conti, L.; Borro, M.; Milani, C.; Simmaco, M.; Esposito, G.; Canali, G.; Pilozzi, E.; Ventura, M.; Annibale, B.; Lahner, E. Gastric microbiota composition in patients with corpus atrophic gastritis. Dig. Liver Dis. 2021, 53, 1580–1587. [Google Scholar] [CrossRef]
- Qi, Z.; Zhibo, Z.; Jing, Z.; Zhanbo, Q.; Shugao, H.; Weili, J.; Jiang, L.; Shuwen, H. Prediction model of poorly differentiated colorectal cancer (CRC) based on gut bacteria. BMC Microbiol. 2022, 22, 312. [Google Scholar] [CrossRef]
- Chen, T.; Ding, R.; Chen, X.; Lu, Y.; Shi, J.; Lü, Y.; Tang, B.; Zhang, W.; Ye, C.; Yuan, M.; et al. Firmicutes and Blautia in gut microbiota lessened in chronic liver diseases and hepatocellular carcinoma patients: A pilot study. Bioengineered 2021, 12, 8233–8246. [Google Scholar] [CrossRef]
- Zhang, X.; Coker, O.O.; Chu, E.S.; Fu, K.; Lau, H.C.H.; Wang, Y.X.; Chan, A.W.H.; Wei, H.; Yang, X.; Sung, J.J.Y.; et al. Dietary cholesterol drives fatty liver-associated liver cancer by modulating gut microbiota and metabolites. Gut 2021, 70, 761–774. [Google Scholar] [CrossRef]
- Cheung, M.K.; Yue, G.G.L.; Lauw, S.; Li, C.S.Y.; Yung, M.Y.; Ng, S.C.; Yip, H.C.; Kwan, H.S.; Chiu, P.W.Y.; Lau, C.B.S. Alterations in gut microbiota of esophageal squamous cell carcinoma patients. J. Gastroenterol. Hepatol. 2022, 37, 1919–1927. [Google Scholar] [CrossRef]
- Rodríguez-García, A.; Arroyo, A.; García-Vicente, R.; Morales, M.L.; Gómez-Gordo, R.; Justo, P.; Cuéllar, C.; Sánchez-Pina, J.; López, N.; Alonso, R.; et al. Short-chain fatty acid production by gut microbiota predicts treatment response in multiple myeloma. Clin. Cancer Res. in press. 2023. [Google Scholar] [CrossRef]
- Minnebo, Y.; Delbaere, K.; Goethals, V.; Raes, J.; Van de Wiele, T.; De Paepe, K. Gut microbiota response to in vitro transit time variation is mediated by microbial growth rates, nutrient use efficiency and adaptation to in vivo transit time. Microbiome 2023, 11, 240. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, B.; Zhao, X.; Zhang, P.; Guo, J.; Zhuang, Y.; Wang, S. Coffee leaf tea extracts improve hyperuricemia nephropathy and its associated negative effect in gut microbiota and amino acid metabolism in rats. J. Agric. Food Chem. 2023, 71, 17775–17787. [Google Scholar] [CrossRef]
- McBrearty, N.; Arzumanyan, A.; Bichenkov, E.; Merali, S.; Merali, C.; Feitelson, M. Short chain fatty acids delay the development of hepatocellular carcinoma in HBx transgenic mice. Neoplasia 2021, 23, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Fazio, G.; Galioto, F.; Ferlito, A.; Coronella, M.; Palmucci, S.; Basile, A. Cavitated pulmonary nodules in a female patient with breast cancer: Keep in mind Serratia marcescens’ infections. Respir. Med. Case Rep. 2021, 33, 101441. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Zhang, X.; Zhu, X.; Jiao, J.; Wu, Y.; Li, Y.; Zhao, L. Fusobacterium nucleatum aggravates ulcerative colitis through promoting gut microbiota dysbiosis and dysmetabolism. J. Periodontol. 2023, 94, 405–418. [Google Scholar] [CrossRef]
- Hägewald, S.; Bernimoulin, J.P.; Köttgen, E.; Kage, A. Salivary IgA subclasses and bacteria-reactive IgA in patients with aggressive periodontitis. J. Periodont. Res. 2002, 37, 333–339. [Google Scholar] [CrossRef]
- Yu, F.; Xu, Q.A.; Chen, W. A targeted fimA DNA vaccine prevents alveolar bone loss in mice after intra-nasal administration. J. Clin. Periodontol. 2011, 38, 334–340. [Google Scholar] [CrossRef]
- Chang, E.; Kobayashi, R.; Fujihashi, K.; Komiya, M.; Kurita-Ochiai, T. Impaired salivary SIgA antibodies elicit oral dysbiosis and subsequent induction of alveolar bone loss. Inflamm. Res. 2021, 70, 151–158. [Google Scholar] [CrossRef]
- Zhang, T.; Cheng, S.; Li, J.; Shang, Y.; Zheng, M. Evaluation of the effect of ultrasound interventional injection of cisplatin in the treatment of liver cancer. Am. J. Transl. Res. 2021, 13, 5603–5609. [Google Scholar] [PubMed]
- Li, T.; Yang, G.; Hao, Q.; Zhang, X.; Zhang, X. Daphnetin ameliorates the expansion of chemically induced hepatocellular carcinoma via reduction of inflammation and oxidative stress. J. Oleo Sci. 2022, 71, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Wang, H.; Bukhari, I.; Zhao, Y.; Huang, H.; Yu, Y.; Sun, X.; Mi, Y.; Mei, L.; Zheng, P. Effects of cholesterol-lowering probiotics on non-alcoholic fatty liver disease in FXR gene knockout mice. Front. Nutr. 2023, 10, 1121203. [Google Scholar] [CrossRef]
- Chen, W.; Wei, Y.; Xiong, A.; Li, Y.; Guan, H.; Wang, Q.; Miao, Q.; Bian, Z.; Xiao, X.; Lian, M.; et al. Comprehensive analysis of serum and fecal bile acid profiles and interaction with gut microbiota in primary biliary cholangitis. Clin. Rev. Allergy Immunol. 2020, 58, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Shimomura, Y.; Takaki, A.; Wada, N.; Yasunaka, T.; Ikeda, F.; Maruyama, T.; Tamaki, N.; Uchida, D.; Onishi, H.; Kuwaki, K.; et al. The serum oxidative/anti-oxidative stress balance becomes dysregulated in patients with non-alcoholic steatohepatitis associated with hepatocellular carcinoma. Intern. Med. 2017, 56, 243–251. [Google Scholar] [CrossRef]
- Defaye, M.; Nourrisson, C.; Baudu, E.; Lashermes, A.; Meynier, M.; Meleine, M.; Wawrzyniak, I.; Bonnin, V.; Barbier, J.; Chassaing, B.; et al. Fecal dysbiosis associated with colonic hypersensitivity and behavioral alterations in chronically Blastocystis-infected rats. Sci. Rep. 2020, 10, 9146. [Google Scholar] [CrossRef]
Figure 1.
Bayesian network with the graphical representation of causal relationships between factors. The analyzed factors consisted of a total of 32 items (4 items that showed significant differences in Table 1, 5 oral periodontal bacterial species, and 23 genera of top occupying intestinal bacteria). g_: enterobacterial genera, HCC: hepatocellular carcinoma. The source of the arrow is the cause and the destination is the effect. The numbers listed on the side of the arrows are Spearman's rank correlation coefficients (n = 59). Statistical superiority was defined as p < 0.05, in which case the numbers were marked with an *.
Figure 1.
Bayesian network with the graphical representation of causal relationships between factors. The analyzed factors consisted of a total of 32 items (4 items that showed significant differences in Table 1, 5 oral periodontal bacterial species, and 23 genera of top occupying intestinal bacteria). g_: enterobacterial genera, HCC: hepatocellular carcinoma. The source of the arrow is the cause and the destination is the effect. The numbers listed on the side of the arrows are Spearman's rank correlation coefficients (n = 59). Statistical superiority was defined as p < 0.05, in which case the numbers were marked with an *.
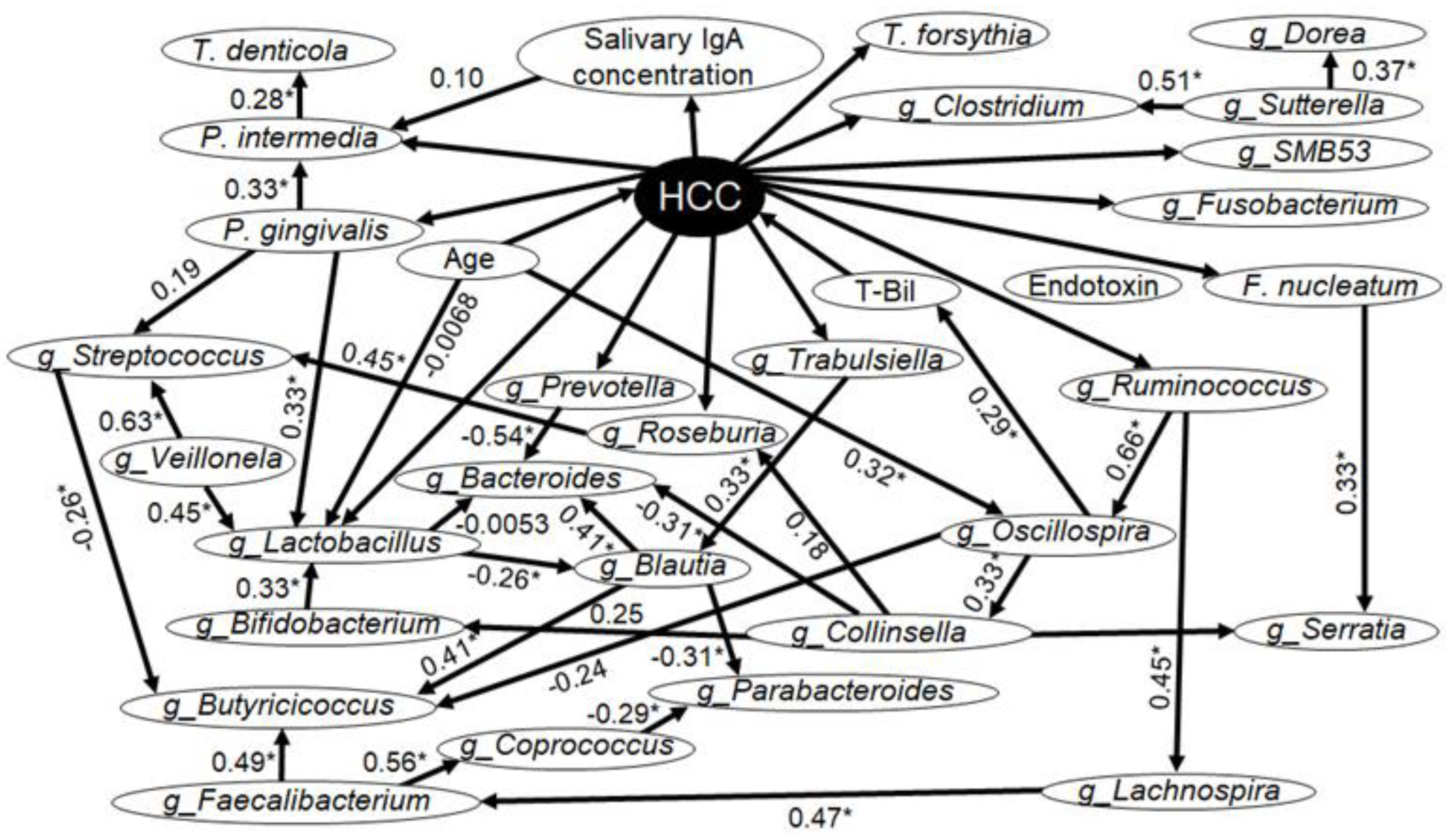
Figure 2.
Validated classification tree with hepatocellular carcinoma (HCC) as the dependent variable and total bilirubin (T-Bil) and age as the explanatory variables.
Figure 2.
Validated classification tree with hepatocellular carcinoma (HCC) as the dependent variable and total bilirubin (T-Bil) and age as the explanatory variables.
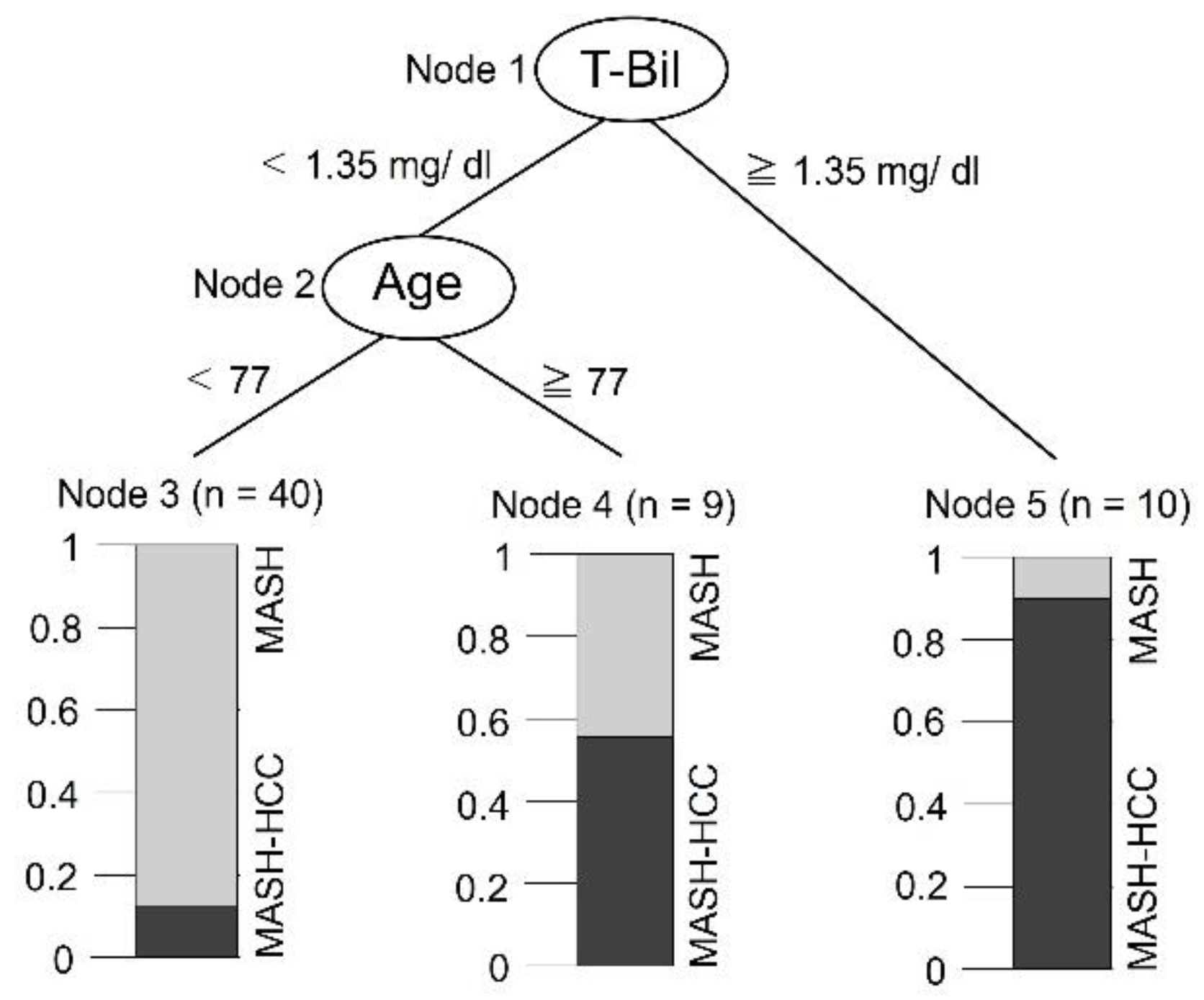

Table 1.
Demographic factors, periodontal and medical conditions, and salivary IgA levels.
| Parameter | MASH Group (N = 41) |
MASH-HCC Group (N = 19) |
p-Value |
|---|---|---|---|
| Gender (men/women) | 25/16 | 13/6 | 0.578 |
| Smoking status (+/−) | 9/32 | 4/15 | 0.973 |
| Age (years) | 59 (55–70) | 79 (64–82) | 0.0004* |
| BMI | 26.2 (22.3–31.5) | 27.7 (25.9–30.8) | 0.503 |
| Number of remaining teeth | 26 (21–27) | 25 (21–27) | 0.598 |
| PD (mm) | 2.8 (2.6–3.2) | 2.9 (2.6–3.2) | 0.653 |
| BOP (%) | 14.9 (10.7–24.1) | 15.4 (7.3–31.4) | 0.799 |
| Tooth mobility | 0 (0–0.1) | 0 (0–0.1) | 0.703 |
| PlI | 0.9 (0.8–1.4) | 0.9 (0.7–1.4) | 0.619 |
| Salivary IgA (ug/mL) | 231.7 (146.5–482.8) | 102.7 (85.8–168.9) | <0.001* |
| Endotoxin (EU) | 0.13 (0.08–0.17) | 0.22 (0.15–0.28) | <0.0001* |
| CRP (mg/dL) | 0.14 (0.09–0.48) | 0.13 (0.07–0.34) | 0.487 |
| AST (U/L) | 51 (27–62) | 38 (29–58) | 0.546 |
| ALT (U/L) | 53 (26–71) | 29 (23–44) | 0.094 |
| T-Bil (mg/dL) | 0.8 (0.6–1) | 1.3 (0.7–1.7) | 0.014* |
BMI, body mass index; PD, Probing depth; BOP, Bleeding on probing; PlI, Plaque index; CRP, C-reactive protein; AST, aspartate aminotransferase; ALT, alanine aminotransferase; T-Bil, total bilirubin. BOP (+) = 1; BOP (−) = 0. The figures for gender and smoking status indicate the number of individuals, and statistical analysis was performed by χ2 test. The presented values for the other items are medians (first quartile–third quartile), and the Mann-Whitney’s U test was used for statistical analysis (*p < 0.05).
Table 2.
Bacterial phyla, genera and species of the salivary microbiota.
| Parameter | MASH Group (N = 41) |
MASH-HCC Group (N = 19) |
p-Value |
|---|---|---|---|
| Diversity of bacterial flora | |||
| Shannon index | 6.72 (6.45–7.13) | 6.51 (6.05–6.80) | 0.03* |
| Phylum | |||
| Actinomycetota (%) | 11.7 (9.56–15.19) | 13.15 (10.38–14.25) | 0.661 |
| Bacillota (%) | 39.5 (36.94–43.06) | 42.7 (35.56–45.30) | 0.604 |
| Bacteroidota (%) | 24.94 (21.61–28.02) | 23.45 (20.54–30.08) | 0.450 |
| Campylobacterota (%) | 0.976 (0.618–1.50) | 1.13(0.806–1.57) | 0.418 |
| Cyanobacteria (%) | 0 (0–0) | 0 (0–0) | 0.928 |
| Desulfobacterota (%) | 0 (0–0.0346) | 0.0135 (0–0.0208) | 0.945 |
| Fusobacteriota (%) | 8.12 (3.94–9.81) | 8.23 (5.42–9.61) | 0.335 |
| Patescibacteria (%) | 1.83 (0.463–2.54) | 1.32 (0.337–2.01) | 0.167 |
| Pseudomonadota (%) | 13.93 (6.79–16.71) | 13.17 (3.01–17.39) | 0.727 |
| Spirochaetota (%) | 0.269 (0.025–0.638) | 0.061 (0.029–0.386) | 0.185 |
| Synergistetes (%) | 0.040 (0–0.103) | 0.020 (0.013–0.025) | 0.228 |
| Genus | |||
| Actinomyces (%) | 6.27 (3.62–8.05) | 5.39 (3.84–6.72) | 0.418 |
| Alloprevotella (%) | 1.54 (0.788–3.26) | 1.16 (0.691–1.91) | 0.348 |
| Atopobium (%) | 0.337 (0.165–0.774) | 0.495 (0.147–0.768) | 0.962 |
| Campylobacter (%) | 1.02 (0.650–1.50) | 1.13 (0.806–1.57) | 0.525 |
| Capnocytophaga (%) | 0.897 (0.282–1.85) | 0.976 (0.524–1.70) | 0.391 |
| Corynebacterium (%) | 0.306 (0.191–0.804) | 0.484 (0.146–0.931) | 0.340 |
| Fusobacterium (%) | 3.25 (1.84–5.07) | 4.48 (2.24–5.58) | 0.266 |
| Gemella (%) | 2.15 (1.14–3.07) | 2.04 (1.17–3.29) | 0.949 |
| Granulicatella (%) | 2.73 (1.86–3.73) | 2.39 (1.60–3.16) | 0.221 |
| Haemophilus (%) | 3.36 (1.70–5.34) | 3.96 (1.10–5.77) | 0.836 |
| Lactobacillus (%) | 0.00620 (0–0.239) | 0.122 (0–1.02) | 0.247 |
| Lautropia (%) | 0.141 (0–0.633) | 0.138 (0–0.831) | 0.572 |
| Leptotrichia (%) | 2.83 (1.36–4.58) | 2.23 (1.53–3.84) | 0.567 |
| Megasphaera (%) | 0.413 (0.151–0.873) | 0.536 (0.106–1.03) | 0.861 |
| Neisseria (%) | 5.20 (1.14–8.72) | 1.25 (1.03–9.20) | 0.505 |
| Peptostreptococcus (%) | 0.256 (0.0694–0.643) | 0.183 (0–0.664) | 0.262 |
| Porphyromonas (%) | 3.71 (1.72–7.14) | 3.98 (0.900–6.49) | 0.589 |
| Prevotella (%) | 16.1 (11.8–19.3) | 15.9 (11.6–20.6) | 0.799 |
| Rothia (%) | 3.09 (1.53–5.53) | 4.42 (2.38–5.42) | 0.409 |
| Streptococcus (%) | 20.3 (16.1–24.6) | 22.2 (16.0–27.0) | 0.340 |
| TM7x (%) | 0.729 (0.221–1.509) | 0.463 (0.0127–1.04) | 0.249 |
| Treponema (%) | 0.292 (0.0331–0.561) | 0.0611 (0.0302–0.225) | 0.0768 |
| Veillonella (%) | 7.98 (6.07–10.7) | 9.68 (6.30–12.4) | 0.204 |
| Species | |||
| Actinomyces israelii (%) | 0 (0–0) | 0 (0–0.0256) | 0.101 |
| Bifidobacterium dentium (%) | 0 (0–0.040) | 0 (0–0.0748) | 0.620 |
| Capnocytophaga gingivalis (%) | 0.345 (0.130–0.691) | 0.427 (0.154–1.25) | 0.193 |
| Dialister pneumosintes (%) | 0 (0–0.0821) | 0 (0–0.0587) | 0.917 |
| Fusobacterium nucleatum (%) | 0.189 (0.0200–0.443) | 0.362 (0.170–0.928) | 0.014* |
| Lactobacillus crispatus (%) | 0 (0–0) | 0 (0–0.00302) | 0.544 |
| Lactobacillus fermentum (%) | 0 (0–0.287) | 0 (0–0.150) | 0.661 |
| Lactobacillus gasseri (%) | 0 (0–0.0257) | 0 (0–0.122) | 0.165 |
| Lactobacillus salivarius (%) | 0 (0–0.00405) | 0 (0–0.0874) | 0.363 |
| Metamycoplasma hyosynoviae (%) | 0 (0–0.0576) | 0 (0–0.0599) | 1.000 |
| Porphyromonas endodontalis (%) | 0.457(0.138–0.982) | 0.195(0–0.377) | 0.0923 |
| Porphyromonas gingivalis (%) | 0.138 (0–1.26) | 0.400 (0–0.725) | 0.520 |
| Prevotella denticola (%) | 0.459 (0.0750–0.948) | 0.429 (0.0683–0.924) | 0.905 |
| Prevotella enoeca (%) | 0 (0–0.00780) | 0 (0–0) | 0.149 |
| Prevotella intermedia (%) | 0 (0–0.334) | 0 (0–0.158) | 0.532 |
| Prevotella nigrescens (%) | 0 (0–0.0243) | 0 (0–0) | 0.175 |
| Streptococcus anginosus (%) | 0.124 (0.0379–0.247) | 0.165 (0.0126–0.373) | 0.937 |
| Streptococcus mutans (%) | 0.0110 (0–0.0608) | 0.0685 (0–0.118) | 0.159 |
| Streptococcus pneumoniae (%) | 0.557 (0.277–0.998) | 0.376(0–1.43) | 0.260 |
| Streptococcus sobrinus (%) | 0 (0–0.0183) | 0(0–0.00838) | 0.919 |
| Tannerella forsythia (%) | 0.146 (0.0612–0.287) | 0.0827 (0.0200–0.328) | 0.661 |
| Treponema denticola (%) | 0.0638 (0–0.195) | 0 (0–0.0325) | 0.0223* |
Values are presented as medians (first quartile–third quartile), and the Mann-Whitney’s U test was used for statistical analysis (*p < 0.05).
Table 3.
Bacterial phyla and genera of the fecal microbiota.
| Parameter | MASH Group (N = 41) |
MASH-HCC Group (N = 19) |
p-Value |
|---|---|---|---|
| Diversity of bacterial flora | |||
| Shannon index | 5.79 (5.54–6.17) | 5.43 (4.92–5.56) | <0.001* |
| Phylum | |||
| Actinomycetota (%) | 5.92 (2.54–9.96) | 4.62 (1.91–9.57) | 0.515 |
| Bacillota (%) | 46.8 (42.1–51.1) | 45.9 (42.1–51.7) | 0.793 |
| Bacteroidota (%) | 37.3(31.5-42.6) | 35.7(31.2-41.2) | 0.634 |
| Campylobacterota (%) | 0(0-0) | 0(0-0) | 0.307 |
| Desulfobacterota (%) | 0.210(0.0138-0.687) | 0.203(0-0.753) | 0.719 |
| Fusobacteriota (%) | 0.0172 (0–0.452) | 1.11 (0.263–2.32) | 0.002* |
| Patescibacteria (%) | 0(0-0) | 0(0-0) | 0.580 |
| Pseudomonadota (%) | 4.98 (3.99–10.6) | 6.11 (3.46–12.3) | 0.861 |
| Spirochaetota (%) | 0 (0–0) | 0 (0–0) | 0.197 |
| Synergistetes (%) | 0 (0–0) | 0 (0–0) | 1.000 |
| Verrucomicrobia (%) | 0 (0–0) | 0 (0–0) | 0.776 |
| Genus | |||
| Bacteroides (%) | 29.8 (19.0–34.5) | 25.6 (19.4–29.4) | 0.360 |
| Bifidobacterium (%) | 3.84 (0.39–9.48) | 1.87 (1.02–7.72) | 0.836 |
| Blautia (%) | 7.16 (4.66–10.8) | 8.11 (1.31–10.4) | 0.424 |
| Butyricicoccus (%) | 0.540 (0.195–0.825) | 0.170 (0.029–0.370) | 0.022* |
| Clostridium (%) | 0.310 (0.165–0.820) | 0.270 (0.130–0.770) | 0.594 |
| Collinsella (%) | 1.83 (0.0100–2.98) | 1.20 (0.0100–3.00) | 0.930 |
| Coprococcus (%) | 1.51 (0.375–4.17) | 0.830 (0.140–2.00) | 0.133 |
| Dorea (%) | 2.70 (1.71–4.62) | 2.68 (0.460–4.05) | 0.259 |
| Faecalibacterium (%) | 1.68 (0–6.16) | 0.119 (0–3.45) | 0.282 |
| Fusobacterium (%) | 0.0100 (0–0.550) | 1.11 (0.260–2.00) | 0.002* |
| Lachnospira (%) | 0.479 (0.0150–1.65) | 0.070 (0.0100–1.00) | 0.292 |
| Lactobacillus (%) | 0.0200 (0–0.505) | 0.220 (0–8.54) | 0.104 |
| Oscillospira (%) | 0.610 (0.205–1.43) | 1.19 (0.290–1.75) | 0.500 |
| Parabacteroides (%) | 2.51 (1.26–5.93) | 3.42 (1.73–5.83) | 0.490 |
| Prevotella (%) | 0.0100 (0–0.0300) | 0.0200 (0.0100–0.850) | 0.074 |
| Roseburia (%) | 0.970 (0.380–1.76) | 0.360 (0.150–1.29) | <0.05* |
| Ruminococcus (%) | 1.38 (0.105–4.00) | 0.239 (0.110–1.80) | 0.294 |
| Serratia (%) | 0.0200 (0–0.560) | 0.0100 (0–4.36) | 0.562 |
| SMB53 (%) | 0.110 (0.0200–0.265) | 0.239 (0–0.650) | 0.707 |
| Streptococcus (%) | 0.790 (0.190–3.45) | 1.56 (0.210–4.42) | 0.634 |
| Sutterella (%) | 3.20 (0.985–4.12) | 2.13 (0.580–3.87) | 0.259 |
| Trabulsiella (%) | 0.270 (0.040–0.920) | 0.440 (0.090–2.39) | 0.499 |
| Veillonela (%) | 0.0500 (0–1.23) | 0.050 (0.0100–0.650) | 0.797 |
Values are presented as medians (first quartile–third quartile), and the Mann-Whitney’s U test was used for statistical analysis (* p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



