Submitted:
08 June 2024
Posted:
11 June 2024
You are already at the latest version
Abstract
Plant-microbe interactions are essential for ecosystem health and agricultural productivity. Metabolomics has revolutionized our understanding of these interactions by providing powerful methods to decipher the complex chemical signals involved. This review highlights the various metabolomics techniques used to study plant-microbe interactions, such as mass spectrometry, nuclear magnetic resonance spectroscopy, and advanced imaging technologies. We also discuss the role of contemporary bioinformatics tools, extensive databases, and the increasing influence of artificial intelligence and machine learning in the analysis of metabolomics data. Integrating metabolomics with other omics fields, including transcriptomics, proteomics, and metagenomics, allows for a comprehensive understanding of the molecular mechanisms underlying plant-microbe interactions. The review emphasizes the significance of fluxomics in elucidating the dynamic nature of metabolic networks. While there are challenges in data analysis, integration, and interpretation, metabolomics has profoundly expanded our knowledge of the role metabolites play in shaping plant-microbe interactions. As metabolomics technologies advance and integrate further with other omics approaches, we anticipate groundbreaking discoveries that will provide deeper insights into these complex interactions, ultimately leading to innovative strategies for sustainable agriculture and biotechnology.
Keywords:
metabolomics
; plant-microbe interactions
; ecosystems
; agricultural productivity
; omics integration
1. Introduction
Plant-microbe interactions are fundamental to the functioning of terrestrial ecosystems, significantly influencing nutrient cycling, plant growth promotion, and disease resistance [1,2]. These interactions shape the dynamics and productivity of ecosystems, influencing the availability and acquisition of nutrients, as well as the resilience of plants against biotic and abiotic stresses [3] (Figure 1). Metabolomics has emerged as a powerful tool for uncovering the intricate interplay between plants and microbes [4,5]. This cutting-edge approach allows researchers to comprehensively analyze the vast array of metabolites produced by both organisms, shedding light on the complex chemical dialogues that govern their interactions [6,7]. By providing a comprehensive snapshot of the metabolic landscape, metabolomics enables the identification and characterization of key metabolites involved in these intricate relationships [8,9].
High-throughput metabolomics techniques, such as mass spectrometry (MS) and nuclear magnetic resonance (NMR) spectroscopy, allow for the rapid and comprehensive profiling of metabolites, enabling researchers to capture the dynamic changes in metabolic profiles that occur during plant-microbe interactions [10,11]. These advanced analytical techniques, coupled with bioinformatics tools for data processing and analysis, have revolutionized our understanding of plant-microbe interactions by facilitating the identification of novel metabolites and the elucidation of their biological functions [12,13].
Metabolomics has been instrumental in identifying signaling molecules, such as phytohormones, quorum-sensing compounds, and specialized metabolites, that mediate plant-microbe interactions [14]. These signaling molecules play crucial roles in the recognition, establishment, and regulation of both beneficial and pathogenic associations between plants and microbes [14,15]. By comparing the metabolic profiles of plants and microbes under different interaction scenarios, researchers can pinpoint the key metabolites involved in specific responses and unravel the underlying molecular mechanisms [16,17].
Furthermore, metabolomics has provided valuable insights into the metabolic basis of plant defense responses against pathogens [17]. By comparing the metabolic profiles of resistant and susceptible plant genotypes, researchers can identify metabolites associated with defense responses, antimicrobial compounds, and virulence factors [18,19]. This knowledge is crucial for developing strategies to enhance crop resistance and mitigate the impact of plant diseases, contributing to sustainable agricultural practices [18]. Metabolomics has also shed light on the metabolic exchanges that occur during the establishment and maintenance of beneficial plant-microbe associations, such as those involving plant growth-promoting rhizobacteria (PGPR) and arbuscular mycorrhizal fungi (AMF) [20,21]. These studies have revealed the complex network of metabolic interactions that underlie these mutualistic relationships, paving the way for the development of novel strategies to optimize plant growth and resilience [20,21].
In conclusion, metabolomics has emerged as a powerful tool for unraveling the complex chemical dialogues that govern plant-microbe interactions. By providing a comprehensive view of the metabolic landscape, metabolomics has shed light on the signaling molecules, defense compounds, and metabolic exchanges that shape these interactions (Figure 2). As metabolomics technologies continue to advance and integrate with other omics approaches, we can expect to gain even deeper insights into the fascinating world of plant-microbe interactions and their implications for sustainable agriculture and biotechnology [22].
2. Metabolomics Techniques in Plant-Microbe Interaction Studies
The application of high-throughput metabolomics in plant-microbe interactions has been facilitated by advances in analytical techniques, such as mass spectrometry (MS) and nuclear magnetic resonance (NMR) spectroscopy [23]. These techniques, coupled with bioinformatics tools for data processing and analysis, enable the rapid and sensitive detection and identification of a wide range of metabolites present in plant and microbial samples [10,22]. The sensitivity and high-throughput nature of these techniques allow for the rapid and comprehensive profiling of metabolites, enabling researchers to capture the dynamic changes in metabolic profiles that occur during plant-microbe interactions [11,12].
2.1. Mass Spectrometry-Based Approaches
Mass spectrometry (MS) has emerged as a powerful tool for metabolomics studies in plant-microbe interactions. MS-based approaches offer high sensitivity, selectivity, and the ability to analyze a wide range of metabolites simultaneously [24,25]. The most commonly used MS techniques in plant-microbe interaction studies include gas chromatography-mass spectrometry (GC-MS), liquid chromatography-mass spectrometry (LC-MS), and capillary electrophoresis-mass spectrometry (CE-MS) [10].
- a.
- Gas chromatography-mass spectrometry (GC-MS)
GC-MS is a widely used technique for the analysis of volatile and semi-volatile compounds in plant-microbe interactions [26]. This technique involves the separation of metabolites based on their volatility and polarity, followed by ionization and detection using MS [27,28]. GC-MS has been successfully applied to study the volatile organic compounds (VOCs) produced during plant-microbe interactions, such as those involved in plant defense responses and microbial communication [29]. For example, a study by Sharifi et al. (2018) used GC-MS to investigate the VOCs produced by the beneficial fungus Trichoderma harzianum and their effects on plant growth and defense responses [30].
- b.
- Liquid chromatography-mass spectrometry (LC-MS)
LC-MS is a versatile technique that allows for the separation and detection of a wide range of non-volatile metabolites, including primary and secondary metabolites [12]. This technique involves the separation of metabolites based on their interaction with a stationary phase and a mobile phase, followed by ionization and detection using MS [31,32]. LC-MS has been extensively used to study the metabolic changes occurring during plant-microbe interactions, such as those involved in plant defense responses and symbiotic associations [33,34]. For instance, a study by Stringlis et al. (2018) used LC-MS to investigate the role of coumarin exudation in shaping the root microbiome and promoting plant health [35].
- c.
- Capillary electrophoresis-mass spectrometry (CE-MS)
CE-MS is a powerful technique that combines the high separation efficiency of capillary electrophoresis with the sensitivity and selectivity of MS [36]. This technique involves the separation of metabolites based on their charge-to-size ratio in an electric field, followed by ionization and detection using MS [37]. CE-MS has been successfully applied to study the metabolic profiles of plants and microbes during their interactions, particularly in the context of plant defense responses and microbial virulence [38,39]. For example, a study by Bringel and Couée (2015) used CE-MS to investigate the metabolic responses of Arabidopsis thaliana to the bacterial pathogen Pseudomonas syringae [40].
2.2. Nuclear Magnetic Resonance (NMR) Spectroscopy
Nuclear magnetic resonance (NMR) spectroscopy is another powerful technique used in metabolomics studies of plant-microbe interactions [41]. NMR spectroscopy provides detailed structural information on metabolites and allows for the simultaneous detection of a wide range of compounds in a sample [41]. One of the main advantages of NMR spectroscopy is its non-destructive nature, which enables the analysis of living tissues and the monitoring of metabolic changes over time [42]. For example, a study by Rivero et al. (2015) used high-resolution magic angle spinning (HR-MAS) NMR spectroscopy to investigate the metabolic changes in tomato roots colonized by the arbuscular mycorrhizal fungus Rhizophagus irregularis [34]. The authors identified several metabolites, such as amino acids and organic acids, that were involved in the nutrient exchange between the plant and the fungus.
2.3. Imaging Techniques (e.g., MALDI-MS Imaging, NMR Imaging)
Imaging techniques, such as matrix-assisted laser desorption/ionization mass spectrometry (MALDI-MS) imaging and NMR imaging, have been increasingly used in metabolomics studies of plant-microbe interactions [43]. These techniques allow for the spatial visualization of metabolites within tissues and provide valuable insights into the localization of specific compounds during plant-microbe interactions [44]. For example, a study by Veličković et al. (2020) used MALDI-MS imaging to investigate the spatial distribution of metabolites in the roots of the model legume Medicago truncatula during its symbiosis with the nitrogen-fixing bacterium Sinorhizobium meliloti [45]. The authors identified several metabolites, such as flavonoids and lipids, that were specifically localized in the nodules formed during the symbiotic interaction. Similarly, a study by Pétriacq et al. (2017) used NMR imaging to investigate the metabolic changes in Arabidopsis thaliana leaves infected with the bacterial pathogen Pseudomonas syringae [46]. The authors identified several metabolites, such as sugars and amino acids, that accumulated in specific regions of the infected leaves and played important roles in the plant’s defense response.
2.4. Bioinformatics Tools and Databases for Metabolomics Data Analysis
Bioinformatics tools and databases play a crucial role in the analysis and interpretation of metabolomics data generated from plant-microbe interaction studies [47]. These tools enable the processing, normalization, and statistical analysis of large metabolomics datasets, as well as the identification of metabolites through comparisons with reference databases [47,48]. Some of the widely used bioinformatics tools for metabolomics data analysis include XCMS [49], MZmine [50], and MetaboAnalyst [51]. These tools provide various functionalities, such as peak detection, alignment, and integration, as well as statistical analysis and data visualization [49]. Statistical tools, such as principal component analysis (PCA) and partial least squares-discriminant analysis (PLS-DA), are used to identify significant metabolic differences between sample groups and to visualize the data [23]. In addition, several metabolomics databases, such as METLIN [52], MassBank [53], and the Human Metabolome Database (HMDB) [54], serve as repositories for metabolite information and facilitate the identification of compounds based on their mass spectra and other chemical properties [52,53].
The integration of high-throughput analytical techniques and bioinformatics tools has greatly enhanced the efficiency and reliability of metabolomics studies in plant-microbe interactions. These advancements have enabled researchers to generate comprehensive metabolic profiles, identify novel metabolites, and unravel the complex metabolic networks involved in these interactions [55]. As technology continues to evolve, it is expected that metabolomics will play an increasingly important role in understanding the molecular mechanisms underlying plant-microbe interactions and in developing strategies for sustainable agriculture and biocontrol [56].
2.5. AI and Machine Learning in Metabolomics for Plant-Microbe Interactions
- a.
- Applications of AI and machine learning in metabolomics
Artificial intelligence (AI) and machine learning (ML) have emerged as powerful tools for analyzing and interpreting the vast amounts of complex data generated by metabolomics studies in plant-microbe interactions [57]. These computational approaches can help in various aspects of metabolomics research, such as data preprocessing, feature selection, metabolite identification, and network analysis [58]. For example, AI and ML algorithms can be used to automate the preprocessing of raw metabolomics data, including peak detection, alignment, and normalization, thus reducing the time and effort required for manual data processing [59]. Moreover, these techniques can aid in the identification of discriminative metabolites and biomarkers that are associated with specific plant-microbe interaction outcomes, such as disease resistance or symbiotic efficiency [60].
- b.
- Machine learning algorithms for metabolomics data analysis
Several machine learning algorithms have been applied to analyze metabolomics data from plant-microbe interaction studies [61]. These algorithms can be broadly classified into supervised and unsupervised learning methods [23]. Supervised learning methods, such as partial least squares discriminant analysis (PLS-DA) and support vector machines (SVM), use labeled data to train models that can predict the class membership of new samples based on their metabolic profiles [62]. These methods have been used to identify metabolic signatures associated with specific plant-microbe interaction outcomes, such as disease resistance or symbiotic efficiency [63]. Unsupervised learning methods, such as principal component analysis (PCA) and hierarchical clustering, explore the inherent structure of the metabolomics data without using class labels [64]. These methods have been used to identify patterns and relationships among metabolites and samples, and to generate hypotheses about the underlying biological processes [65].
- c.
- Deep learning for metabolomics
Deep learning, a subfield of machine learning that uses artificial neural networks with multiple layers, has shown great promise in analyzing metabolomics data from plant-microbe interaction studies [66]. Deep learning algorithms, such as convolutional neural networks (CNNs) and recurrent neural networks (RNNs), can automatically learn hierarchical representations of the metabolomics data, capturing complex patterns and relationships among metabolites [67]. These algorithms have been used for various tasks in metabolomics, such as metabolite identification, biomarker discovery, and network analysis [68]. For example, Uppal et al. (2017) developed xMSannotator, an R package that utilizes recurrent neural networks (RNNs) for network-based annotation of metabolites from high-resolution metabolomics data. By integrating metabolic transformation rules and mass spectral patterns within an RNN framework, the package enables annotating metabolites, including those not found in existing databases, outperforming traditional annotation methods [69]. Another study by Pomyen et al. (2021) used deep learning to integrate metabolomics and transcriptomics data from rice plants infected with the fungal pathogen Magnaporthe oryzae, revealing novel insights into the metabolic reprogramming during the infection process [70].
- d.
- Challenges and future directions
Despite the promising applications of AI and machine learning in metabolomics for plant-microbe interactions, several challenges remain to be addressed. One of the main challenges is the limited availability of large, well-curated metabolomics datasets for training and validating AI and ML models [71]. The development of standardized protocols for data collection, processing, and reporting, as well as the creation of public repositories for metabolomics data, will be crucial for advancing the application of AI and ML in this field [72]. Another challenge is the interpretability of the AI and ML models, particularly deep learning models, which can be difficult to understand and explain [73]. The development of explainable AI techniques, such as attention mechanisms and interpretation tools, will be important for generating biologically meaningful insights from these models [74].
In conclusion, AI and machine learning have the potential to revolutionize the analysis and interpretation of metabolomics data in plant-microbe interaction studies. These computational approaches can help in various aspects of metabolomics research, from data preprocessing to biomarker discovery and network analysis. However, addressing the challenges related to data availability, standardization, and model interpretability will be crucial for realizing the full potential of AI and ML in this field. As these technologies continue to advance and integrate with other omics approaches, we can anticipate new breakthroughs in our understanding of the complex metabolic interactions between plants and microbes, and their applications in agriculture and biotechnology.
3. Integration of Metabolomics with Other Omics Approaches
Metabolomics provides a comprehensive snapshot of the metabolic profiles of both plants and their associated microbes, enabling the identification of key metabolites involved in these interactions [16,25]. By integrating metabolomics data with other omics approaches, such as transcriptomics and proteomics, researchers can gain a more comprehensive understanding of the molecular mechanisms underlying these interactions [75]. This multi-omics approach facilitates the elucidation of regulatory networks, metabolic pathways, and signaling cascades involved in these complex interactions, paving the way for innovative strategies in sustainable agriculture, ecosystem management, and environmental conservation [76].
3.1. Transcriptomics and Metabolomics
The integration of transcriptomics and metabolomics has proven to be a powerful approach for understanding the complex interactions between plants and microbes [77]. Transcriptomics provides information on gene expression changes, while metabolomics reveals the resulting metabolic alterations [78]. By combining these two omics techniques, researchers can gain a more comprehensive understanding of the molecular mechanisms underlying plant-microbe interactions [7]. For example, a study by Camañes et al. (2015) integrated transcriptomics and metabolomics to investigate the response of tomato plants to the beneficial bacterium Pseudomonas putida [79]. The authors identified several metabolites, such as amino acids and organic acids, that were differentially accumulated in the plants, and correlated these changes with the expression of specific genes involved in plant defense and growth promotion.
3.2. Proteomics and Metabolomics
Proteomics, the study of the entire set of proteins expressed by an organism [80], can provide valuable insights into the functional aspects of plant-microbe interactions [81]. Integrating proteomics with metabolomics can help elucidate the relationship between protein abundance and metabolic changes [82]. For instance, Balmer et al. (2013) used proteomics and metabolomics to investigate the response of maize leaves to the fungal pathogen Colletotrichum graminicola [83]. The authors identified several proteins and metabolites, such as PR proteins, phytoalexins, and oxylipins, that were differentially accumulated in the infected leaves and could contribute to the plant’s defense response against the pathogen.
3.3. Metagenomics and Metabolomics
Metagenomics, the study of the collective genomes of microorganisms in a particular environment, has been increasingly used to investigate the diversity and functional potential of plant-associated microbial communities [84]. Integrating metagenomics with metabolomics can provide a more comprehensive understanding of the metabolic capabilities of these communities and their interactions with host plants [85]. For example, a study by Gao et al. (2018) integrated metagenomics and metabolomics to investigate the microbial communities associated with the medicinal plant Salvia miltiorrhiza [86]. The authors identified several bacterial and fungal taxa that were enriched in the plant roots and correlated their presence with the production of specific bioactive compounds, such as tanshinones and salvianolic acids.
3.4. Fluxomics: A Complementary Approach to Study Plant-Microbe Interactions
In addition to metabolomics, fluxomics has emerged as a complementary approach to study plant-microbe interactions. Fluxomics is the quantitative analysis of metabolic fluxes, which are the rates of metabolite interconversion within a biological system [87,88]. This approach provides insights into the dynamic nature of metabolic networks and helps to identify the key metabolic pathways and regulatory mechanisms [89]. Fluxomics techniques, such as 13C metabolic flux analysis (13C-MFA) and flux balance analysis (FBA), allow researchers to trace the flow of metabolites through metabolic pathways and to quantify the flux distributions within a biological system [90,91]. These techniques involve the use of stable isotope labeling, such as 13C, to track the fate of metabolites as they are transformed and transported within the plant-microbe system [92,93].
By combining fluxomics with metabolomics, researchers can gain a more comprehensive understanding of the metabolic reprogramming that occurs during plant-microbe interactions [94]. For example, a study by Schwender et al. (2004) used 13C-MFA to investigate the metabolic fluxes in the interaction between the plant pathogen Agrobacterium tumefaciens and its host plant, Arabidopsis thaliana [95]. The authors found that the infection led to a redirection of carbon fluxes towards the synthesis of specific amino acids and organic acids, which were essential for the growth and virulence of the pathogen [95]. Fluxomics has also been applied to study the metabolic interactions between plants and beneficial microbes, such as arbuscular mycorrhizal fungi (AMF) and rhizobia [96]. For instance, Rich et al. (2017) employed 13C-MFA to study the metabolic interactions between the model legume Lotus japonicus and the nitrogen-fixing bacterium Mesorhizobium loti during root nodule symbiosis [97]. The authors discovered that the bacterium supplied the plant with fixed nitrogen in exchange for dicarboxylic acids and amino acids derived from the plant’s photosynthetic activity, and that the carbon allocation to the bacterium was modulated by the plant’s nitrogen status.
Fluxomics can also provide valuable insights into the metabolic basis of plant-microbe interactions under different environmental conditions, such as drought, salinity, and nutrient deficiency [98]. By quantifying the changes in metabolic fluxes under these conditions, researchers can identify the key metabolic pathways and regulatory mechanisms that contribute to plant adaptation and resilience [99,100]. By combining fluxomics with metabolomics, researchers can gain a more comprehensive understanding of the metabolic reprogramming that occurs during these interactions, and identify the key metabolic pathways and regulatory mechanisms that underlie plant-microbe communication and adaptation to environmental stresses. As fluxomics technologies continue to advance, we can expect to see more applications of this approach in the study of plant-microbe interactions and their implications for sustainable agriculture and biotechnology [101,102].
3.5. Examples of Multi-Omics Studies in Plant-Microbe Interactions
Several studies have demonstrated the power of integrating multiple omics approaches to unravel the complexity of plant-microbe interactions. For example, a study by Liu et al. (2017) integrated transcriptomics, proteomics, and metabolomics to investigate the response of maize plants to the beneficial fungus Trichoderma harzianum [103]. The authors identified several genes, proteins, and metabolites that were differentially expressed in the plants and contributed to their enhanced growth and stress tolerance. Similarly, Schläpfer et al. (2017) employed metagenomics, metabolomics, and transcriptomics to explore the interactions between the model plant Arabidopsis thaliana and its root-associated microbial communities [104]. The authors identified various bacterial and fungal taxa that were enriched in the plant rhizosphere and linked their presence to the expression of specific plant genes and the accumulation of defense-related metabolites, such as glucosinolates and camalexin. These multi-omics studies highlight the power of integrating different omics approaches to gain a more holistic understanding of the complex interactions between plants and microbes.
4. Challenges and Future Perspectives
Despite the significant advancements in metabolomics and its applications in plant-microbe research, several challenges remain. These include the complexity of data analysis and interpretation, the need for robust computational tools and databases, and the integration of multi-omics data across different spatial and temporal scales [23]. Overcoming these challenges will be crucial for unlocking the full potential of metabolomics in unraveling the intricate chemical dialogues that govern plant-microbe interactions.
4.1. Standardization of Metabolomics Protocols and Data Reporting
One of the major challenges in metabolomics studies of plant-microbe interactions is the lack of standardized protocols and data reporting guidelines [105]. This inconsistency makes it difficult to compare and integrate results from different studies, hindering the progress in this field [106]. Efforts have been made to establish standardized protocols for sample preparation, data acquisition, and data analysis in metabolomics [107]. For example, the Metabolomics Standards Initiative (MSI) has developed guidelines for reporting metabolomics experiments, including minimum information about a metabolomics experiment (MIAMET) [108]. Adherence to these guidelines will improve the reproducibility and comparability of metabolomics studies in plant-microbe interactions.
4.2. Advancements in Computational Tools and Databases
One major challenge in interpreting metabolomics data is the identification of metabolites and their biological roles in plant-microbe interactions [109]. The diversity of metabolites produced by plants and microbes, combined with the limited coverage of metabolite databases, makes it difficult to accurately identify and annotate the detected compounds [110]. Moreover, the functional characterization of identified metabolites requires a deep understanding of the biological context and the integration of metabolomics data with other omics approaches [111].
The rapid growth of metabolomics data generated from plant-microbe interaction studies has necessitated the development of advanced computational tools and databases for data processing, analysis, and interpretation [112]. Recent advancements in machine learning and artificial intelligence have enabled the development of more powerful tools for metabolomics data analysis [113]. For instance, deep learning algorithms have been used to improve the accuracy of metabolite identification and quantification [114]. Additionally, the integration of metabolomics databases with other omics databases, such as genomics and transcriptomics, has facilitated the construction of comprehensive metabolic networks and the identification of novel metabolic pathways [115].
4.3. Omics
The integration of metabolomics with transcriptomics and proteomics provides a more comprehensive view of the molecular mechanisms underlying plant-microbe interactions [116]. By combining these omics approaches, researchers can identify the genes, proteins, and metabolites involved in specific interaction processes and unravel their regulatory networks [117]. For instance, a study by Coppola et al. (2019) integrated metabolomics and transcriptomics to investigate the response of tomato plants to the beneficial fungus Trichoderma harzianum, revealing the coordinated regulation of defense and growth-related pathways [118].
However, the integration of multi-omics data poses its own challenges, such as data heterogeneity, dimensionality, and the need for advanced computational tools [119]. The development of bioinformatics pipelines and statistical methods for data integration, such as network analysis and machine learning algorithms, is essential for extracting biologically relevant information from the integrated datasets [115,120].
5. Conclusion
5.1. Summary of Key Points
Metabolomics has emerged as a powerful tool for investigating the complex interactions between plants and microbes. This review has highlighted the various metabolomics techniques, such as mass spectrometry and nuclear magnetic resonance spectroscopy, that have been employed to study plant-microbe interactions. We have discussed the application of metabolomics in elucidating the metabolic changes that occur during plant-pathogen interactions, identifying resistance-related metabolites, and assisting in the breeding of disease-resistant crops. Furthermore, we have explored the role of metabolomics in understanding the metabolic profiles of plant-symbiont associations, identifying signaling metabolites, and unraveling the establishment and functioning of symbioses. The integration of metabolomics with other omics approaches, such as transcriptomics, proteomics, and metagenomics, has provided a more comprehensive understanding of the molecular mechanisms underlying plant-microbe interactions.
5.2. Significance of Metabolomics in Understanding Plant-Microbe Interactions
Metabolomics has significantly advanced our understanding of the complex chemical dialogues that mediate plant-microbe interactions. By providing a comprehensive view of the metabolic changes that occur during these interactions, metabolomics has shed light on the diverse roles of primary and specialized metabolites in shaping the outcomes of these relationships. Metabolomics has enabled the identification of key metabolites involved in plant defense responses, symbiotic associations, and growth promotion, as well as the discovery of novel bioactive compounds with potential applications in agriculture and biotechnology. The integration of metabolomics with other omics approaches has further enhanced our ability to unravel the intricate networks of molecular interactions between plants and microbes, paving the way for the development of novel strategies for crop improvement and sustainable agriculture.
5.3. Outlook for Future Research Directions
Despite the significant progress made in applying metabolomics to study plant-microbe interactions, several challenges and opportunities remain for future research. The standardization of metabolomics protocols and data reporting guidelines is crucial for improving the reproducibility and comparability of studies in this field. The development of advanced computational tools and databases will facilitate the analysis and interpretation of the growing volume of metabolomics data generated from plant-microbe interaction studies. Future research should also focus on exploring the diversity of specialized metabolites produced by plants and their associated microbes, and elucidating their ecological roles in mediating these interactions. Finally, translating metabolomics findings into practical agricultural applications, such as marker-assisted breeding, metabolic engineering, and precision agriculture, will be essential for harnessing the power of plant-microbe interactions for sustainable crop production.
In conclusion, metabolomics has revolutionized our understanding of the complex chemical dialogues that underlie plant-microbe interactions. By providing a comprehensive view of the metabolic changes that occur during these interactions, metabolomics has opened up new avenues for discovering novel metabolites, unraveling the molecular mechanisms of symbioses and pathogenesis, and developing strategies for crop improvement. As metabolomics technologies continue to advance and integrate with other omics approaches, we can anticipate new breakthroughs in our understanding of the fascinating world of plant-microbe interactions and their applications in agriculture and biotechnology.
References
- Igarashi, D.; Bethke, G.; Xu, Y.; Tsuda, K.; Glazebrook, J.; Katagiri, F. Pattern-Triggered Immunity Suppresses Programmed Cell Death Triggered by Fumonisin B1. PLoS One 2013, 8, 1–2. [Google Scholar] [CrossRef]
- Bonfante, P.; Anca, I.-A. Plants, mycorrhizal fungi, and bacteria: a network of interactions. Annu. Rev. Microbiol. 2009, 63, 363–383. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; Van Der Putten, W.H. Going back to the roots: the microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, R.P.; Kopriva, S. Metabolic niches in the rhizosphere microbiome: new tools and approaches to analyse metabolic mechanisms of plant–microbe nutrient exchange. J. Exp. Bot. 2019, 70, 1087–1094. [Google Scholar] [CrossRef]
- Mhlongo, M.I.; Piater, L.A.; Madala, N.E.; Labuschagne, N.; Dubery, I.A. The chemistry of plant–microbe interactions in the rhizosphere and the potential for metabolomics to reveal signaling related to defense priming and induced systemic resistance. Front. Plant Sci. 2018, 9, 314703. [Google Scholar] [CrossRef]
- Kessler, A.; Kalske, A. Plant secondary metabolite diversity and species interactions. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 115–138. [Google Scholar] [CrossRef]
- van Dam, N.M.; Bouwmeester, H.J. Metabolomics in the rhizosphere: tapping into belowground chemical communication. Trends Plant Sci. 2016, 21, 256–265. [Google Scholar] [CrossRef]
- Boiteau, R.M.; Hoyt, D.W.; Nicora, C.D.; Kinmonth-Schultz, H.A.; Ward, J.K.; Bingol, K. Structure elucidation of unknown metabolites in metabolomics by combined NMR and MS/MS prediction. Metabolites 2018, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Mahieu, N.G.; Patti, G.J. Systems-level annotation of a metabolomics data set reduces 25 000 features to fewer than 1000 unique metabolites. Anal. Chem. 2017, 89, 10397–10406. [Google Scholar] [CrossRef]
- Allwood, J.W.; Ellis, D.I.; Goodacre, R. Metabolomic technologies and their application to the study of plants and plant-host interactions. Physiol. Plant. 2008, 132, 117–135. [Google Scholar] [CrossRef]
- Sumner, L.W.; Yang, D.S.; Bench, B.J.; Watson, B.S.; Li, C.; Jones, A.D. Spatially resolved plant metabolomics. Annu. Plant Rev. Vol. 43 Biol. Plant Metabolomics 2011, 43, 343–366. [Google Scholar]
- De Vos, R.C.; Moco, S.; Lommen, A.; Keurentjes, J.J.; Bino, R.J.; Hall, R.D. Untargeted large-scale plant metabolomics using liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2007, 2, 778–791. [Google Scholar] [CrossRef]
- Tenenboim, H.; Brotman, Y. Omic relief for the biotically stressed: metabolomics of plant biotic interactions. Trends Plant Sci. 2016, 21, 781–791. [Google Scholar] [CrossRef]
- Chen, L.; Schwier, M.; Krumbach, J.; Kopriva, S.; Jacoby, R.P. Metabolomics in plant-microbe interactions in the roots. Adv. Bot. Res. 2021, 98, 133–161. [Google Scholar]
- Lugtenberg, B.; Kamilova, F. Plant-growth-promoting rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [PubMed]
- Trethewey, R.N. Metabolite profiling as an aid to metabolic engineering in plants. Curr. Opin. Plant Biol. 2004, 7, 196–201. [Google Scholar] [CrossRef]
- Schenk, P.M.; Carvalhais, L.C.; Kazan, K. Unraveling plant–microbe interactions: can multi-species transcriptomics help? Trends Biotechnol. 2012, 30, 177–184. [Google Scholar] [CrossRef]
- Castro-Moretti, F.R.; Gentzel, I.N.; Mackey, D.; Alonso, A.P. Metabolomics as an emerging tool for the study of plant–pathogen interactions. Metabolites 2020, 10, 52. [Google Scholar] [CrossRef]
- Sade, D.; Shriki, O.; Cuadros-Inostroza, A.; Tohge, T.; Semel, Y.; Haviv, Y.; Willmitzer, L.; Fernie, A.R.; Czosnek, H.; Brotman, Y. Comparative metabolomics and transcriptomics of plant response to Tomato yellow leaf curl virus infection in resistant and susceptible tomato cultivars. Metabolomics 2015, 11, 81–97. [Google Scholar] [CrossRef]
- Vacheron, J.; Desbrosses, G.; Bouffaud, M.-L.; Touraine, B.; Moënne-Loccoz, Y.; Muller, D.; Legendre, L.; Wisniewski-Dyé, F.; Prigent-Combaret, C. Plant growth-promoting rhizobacteria and root system functioning. Front. Plant Sci. 2013, 4, 57135. [Google Scholar] [CrossRef]
- Pozo, M.J.; Van Der Ent, S.; Van Loon, L.C.; Pieterse, C.M.J. Transcription factor MYC2 is involved in priming for enhanced defense during rhizobacteria-induced systemic resistance in Arabidopsis thaliana. New Phytol. 2008, 180, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Wolfender, J.-L.; Rudaz, S.; Hae Choi, Y.; Kyong Kim, H. Plant metabolomics: from holistic data to relevant biomarkers. Curr. Med. Chem. 2013, 20, 1056–1090. [Google Scholar] [PubMed]
- Worley, B.; Powers, R. Multivariate analysis in metabolomics. Curr. metabolomics 2013, 1, 92–107. [Google Scholar] [PubMed]
- Villas-Bôas, S.G.; Mas, S.; Åkesson, M.; Smedsgaard, J.; Nielsen, J. Mass spectrometry in metabolome analysis. Mass Spectrom. Rev. 2005, 24, 613–646. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Fu, X. Reprogramming of Plant Central Metabolism in Response to Abiotic Stresses: A Metabolomics View. Int. J. Mol. Sci. 2022, 23, 5716. [Google Scholar] [CrossRef]
- Fiehn, O. Metabolomics by gas chromatography–mass spectrometry: Combined targeted and untargeted profiling. Curr. Protoc. Mol. Biol. 2016, 114, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Lisec, J.; Schauer, N.; Kopka, J.; Willmitzer, L.; Fernie, A.R. Gas chromatography mass spectrometry–based metabolite profiling in plants. Nat. Protoc. 2006, 1, 387–396. [Google Scholar] [CrossRef]
- Fu, X.; Xu, Y. Dynamic metabolic changes in arabidopsis seedlings under hypoxia stress and subsequent reoxygenation recovery. Stresses 2023, 3, 86–101. [Google Scholar] [CrossRef]
- Jud, W.; Winkler, J.B.; Niederbacher, B.; Niederbacher, S.; Schnitzler, J.-P. Volatilomics: a non-invasive technique for screening plant phenotypic traits. Plant Methods 2018, 14, 1–18. [Google Scholar] [CrossRef]
- Sharifi, R.; Lee, S.; Ryu, C. Microbe-induced plant volatiles. New Phytol. 2018, 220, 684–691. [Google Scholar] [CrossRef]
- Allwood, J.W.; Goodacre, R. An introduction to liquid chromatography–mass spectrometry instrumentation applied in plant metabolomic analyses. Phytochem. Anal. An Int. J. Plant Chem. Biochem. Tech. 2010, 21, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Freund, D.M.; Hegeman, A.D.; Cohen, J.D. Metabolic signatures of Arabidopsis thaliana abiotic stress responses elucidate patterns in stress priming, acclimation, and recovery. Stress Biol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Mhlongo, M.I.; Piater, L.A.; Steenkamp, P.A.; Labuschagne, N.; Dubery, I.A. Metabolic profiling of PGPR-treated tomato plants reveal priming-related adaptations of secondary metabolites and aromatic amino acids. Metabolites 2020, 10, 210. [Google Scholar] [CrossRef] [PubMed]
- Rivero, J.; Gamir, J.; Aroca, R.; Pozo, M.J.; Flors, V. Metabolic transition in mycorrhizal tomato roots. Front. Microbiol. 2015, 6, 598. [Google Scholar] [CrossRef] [PubMed]
- Stringlis, I.A.; Yu, K.; Feussner, K.; de Jonge, R.; Van Bentum, S.; Van Verk, M.C.; Berendsen, R.L.; Bakker, P.A.H.M.; Feussner, I.; Pieterse, C.M.J. MYB72-dependent coumarin exudation shapes root microbiome assembly to promote plant health. Proc. Natl. Acad. Sci. 2018, 115, E5213–E5222. [Google Scholar] [CrossRef] [PubMed]
- Ramautar, R.; Somsen, G.W.; de Jong, G.J. CE-MS in metabolomics. Electrophoresis 2009, 30, 276–291. [Google Scholar] [CrossRef]
- Monton, M.R.N.; Soga, T. Metabolome analysis by capillary electrophoresis–mass spectrometry. J. Chromatogr. A 2007, 1168, 237–246. [Google Scholar] [CrossRef]
- Guijas, C.; Montenegro-Burke, J.R.; Domingo-Almenara, X.; Palermo, A.; Warth, B.; Hermann, G.; Koellensperger, G.; Huan, T.; Uritboonthai, W.; Aisporna, A.E. METLIN: a technology platform for identifying knowns and unknowns. Anal. Chem. 2018, 90, 3156–3164. [Google Scholar] [CrossRef] [PubMed]
- Gamir, J.; Pastor, V.; Kaever, A.; Cerezo, M.; Flors, V. Targeting novel chemical and constitutive primed metabolites against P lectosphaerella cucumerina. Plant J. 2014, 78, 227–240. [Google Scholar] [CrossRef]
- Bringel, F.; Couée, I. Pivotal roles of phyllosphere microorganisms at the interface between plant functioning and atmospheric trace gas dynamics. Front. Microbiol. 2015, 6, 144430. [Google Scholar] [CrossRef]
- Kim, H.K.; Choi, Y.H.; Verpoorte, R. NMR-based plant metabolomics: where do we stand, where do we go? Trends Biotechnol. 2011, 29, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, P.; Kruger, N.J.; Ratcliffe, R.G. Metabolite fingerprinting and profiling in plants using NMR. J. Exp. Bot. 2005, 56, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Boughton, B.A.; Thinagaran, D. Mass spectrometry imaging (MSI) for plant metabolomics. Plant Metabolomics Methods Protoc. 2018, 241–252. [Google Scholar]
- Sturtevant, D.; Lee, Y.-J.; Chapman, K.D. Matrix assisted laser desorption/ionization-mass spectrometry imaging (MALDI-MSI) for direct visualization of plant metabolites in situ. Curr. Opin. Biotechnol. 2016, 37, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Veličković, D.; Agtuca, B.J.; Stopka, S.A.; Vertes, A.; Koppenaal, D.W.; Paša-Tolić, L.; Stacey, G.; Anderton, C.R. Observed metabolic asymmetry within soybean root nodules reflects unexpected complexity in rhizobacteria-legume metabolite exchange. ISME J. 2018, 12, 2335–2338. [Google Scholar] [CrossRef] [PubMed]
- Pétriacq, P.; Williams, A.; Cotton, A.; McFarlane, A.E.; Rolfe, S.A.; Ton, J. Metabolite profiling of non-sterile rhizosphere soil. Plant J. 2017, 92, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, A.; Kusano, M. Recent progress in the development of metabolome databases for plant systems biology. Front. Plant Sci. 2013, 4, 42957. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y. Metabolomics study on Arabidopsis thaliana abiotic stress responses for priming, recovery, and stress combinations. <bold>2018</bold>.
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinformatics 2010, 11, 1–11. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]
- Smith, C.A.; O’Maille, G.; Want, E.J.; Qin, C.; Trauger, S.A.; Brandon, T.R.; Custodio, D.E.; Abagyan, R.; Siuzdak, G. METLIN: A metabolite mass spectral database. Therapeutic Drug Monitoring 2005, 27, 747–751. [Google Scholar] [CrossRef]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K.; et al. MassBank: A public repository for sharing mass spectral data for life sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Feunang, Y.D.; Marcu, A.; Guo, A.C.; Liang, K.; Vázquez-Fresno, R.; Sajed, T.; Johnson, D.; Li, C.; Karu, N. HMDB 4.0: the human metabolome database for 2018. Nucleic Acids Res. 2018, 46, D608–D617. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.D. Plant metabolomics in a nutshell: potential and future challenges. Annu. Plant Rev. Vol. 43 Biol. Plant Metabolomics 2011, 43, 1–24. [Google Scholar]
- Tugizimana, F.; Piater, L.; Dubery, I. Plant metabolomics: A new frontier in phytochemical analysis. S. Afr. J. Sci. 2013, 109, 1–11. [Google Scholar] [CrossRef]
- Murmu, S.; Sinha, D.; Chaurasia, H.; Sharma, S.; Das, R.; Jha, G.K.; Archak, S. A review of artificial intelligence-assisted omics techniques in plant defense: current trends and future directions. Front. Plant Sci. 2024, 15, 1292054. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Berdine, G. Artificial intelligence in biomedical research. Southwest Respir. Crit. Care Chronicles 2023, 11, 62–65. [Google Scholar] [CrossRef]
- Misra, B.B.; Mohapatra, S. Tools and resources for metabolomics research community: A 2017–2018 update. Electrophoresis 2019, 40, 227–246. [Google Scholar] [CrossRef]
- Salinas, J.L.; Kissinger, J.C.; Jones, D.P.; Galinski, M.R. Metabolomics in the fight against malaria. Mem. Inst. Oswaldo Cruz 2014, 109, 589–597. [Google Scholar] [CrossRef]
- Gromski, P.S.; Muhamadali, H.; Ellis, D.I.; Xu, Y.; Correa, E.; Turner, M.L.; Goodacre, R. A tutorial review: Metabolomics and partial least squares-discriminant analysis–a marriage of convenience or a shotgun wedding. Anal. Chim. Acta 2015, 879, 10–23. [Google Scholar] [CrossRef]
- Mishra, P.; Singh, U.; Pandey, C.M.; Mishra, P.; Pandey, G. Application of student’s t-test, analysis of variance, and covariance. Ann. Card. Anaesth. 2019, 22, 407–411. [Google Scholar] [CrossRef]
- Abdel-Farid, I.B.; Jahangir, M.; Van Den Hondel, C.; Kim, H.K.; Choi, Y.H.; Verpoorte, R. Fungal infection-induced metabolites in Brassica rapa. Plant Sci. 2009, 176, 608–615. [Google Scholar] [CrossRef]
- Jolliffe, I.T.; Cadima, J. Principal component analysis: a review and recent developments. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2016, 374, 20150202. [Google Scholar] [CrossRef]
- Want, E.J.; Wilson, I.D.; Gika, H.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Holmes, E.; Nicholson, J.K. Global metabolic profiling procedures for urine using UPLC–MS. Nat. Protoc. 2010, 5, 1005–1018. [Google Scholar] [CrossRef]
- Lecun, Y.; Bengio, Y.; Hinton, G. Deep learning. Nature 2015, 521, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Ching, T.; Himmelstein, D.S.; Beaulieu-Jones, B.K.; Kalinin, A.A.; Do, B.T.; Way, G.P.; Ferrero, E.; Agapow, P.-M.; Zietz, M.; Hoffman, M.M. Opportunities and obstacles for deep learning in biology and medicine. J. R. Soc. interface 2018, 15, 20170387. [Google Scholar] [CrossRef]
- Liebal, U.W.; Phan, A.N.T.; Sudhakar, M.; Raman, K.; Blank, L.M. Machine learning applications for mass spectrometry-based metabolomics. Metabolites 2020, 10, 243. [Google Scholar] [CrossRef]
- Uppal, K.; Walker, D.I.; Jones, D.P. xMSannotator: an R package for network-based annotation of high-resolution metabolomics data. Anal. Chem. 2017, 89, 1063–1067. [Google Scholar] [CrossRef] [PubMed]
- Pomyen, Y.; Wanichthanarak, K.; Poungsombat, P.; Fahrmann, J.; Grapov, D.; Khoomrung, S. Deep metabolome: Applications of deep learning in metabolomics. Comput. Struct. Biotechnol. J. 2020, 18, 2818–2825. [Google Scholar] [CrossRef]
- Zhang, A.; Sun, H.; Yan, G.; Wang, P.; Wang, X. Metabolomics for biomarker discovery: moving to the clinic. Biomed Res. Int. 2015, 2015. [Google Scholar] [CrossRef]
- Spicer, R.A.; Salek, R.; Steinbeck, C. Compliance with minimum information guidelines in public metabolomics repositories. Sci. data 2017, 4, 1–8. [Google Scholar] [CrossRef]
- Samek, W.; Montavon, G.; Vedaldi, A.; Hansen, L.K.; Müller, K.-R. <italic>Explainable AI: interpreting, explaining and visualizing deep learning</italic>; Springer Nature, 2019; Vol. 11700; ISBN 3030289540.
- Yang, F.; Du, M.; Hu, X. Evaluating explanation without ground truth in interpretable machine learning. arXiv Prepr. arXiv1907.06831 2019. [Google Scholar]
- Jastrebski, S.F.; Lamont, S.J.; Schmidt, C.J. Chicken hepatic response to chronic heat stress using integrated transcriptome and metabolome analysis. PLoS One 2017, 12, e0181900. [Google Scholar] [CrossRef] [PubMed]
- Weckwerth, W. Metabolomics in systems biology. Annu. Rev. Plant Biol. 2003, 54, 669–689. [Google Scholar] [CrossRef] [PubMed]
- Tang, J. Microbial metabolomics. Curr. Genomics 2011, 12, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Guijas, C.; Montenegro-Burke, J.R.; Warth, B.; Spilker, M.E.; Siuzdak, G. Metabolomics activity screening for identifying metabolites that modulate phenotype. Nat. Biotechnol. 2018, 36, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Camañes, G.; Scalschi, L.; Vicedo, B.; González-Bosch, C.; García-Agustín, P. An untargeted global metabolomic analysis reveals the biochemical changes underlying basal resistance and priming in Solanum lycopersicum, and identifies 1-methyltryptophan as a metabolite involved in plant responses to Botrytis cinerea and Pseudomonas sy. Plant J. 2015, 84, 125–139. [Google Scholar] [CrossRef]
- Fan, K.-T.; Xu, Y.; Hegeman, A.D. Elevated Temperature Effects on Protein Turnover Dynamics in Arabidopsis thaliana Seedlings Revealed by 15N-Stable Isotope Labeling and ProteinTurnover Algorithm. Int. J. Mol. Sci. 2024, 25, 5882. [Google Scholar] [CrossRef]
- Mathesius, U.; Mulders, S.; Gao, M.; Teplitski, M.; Caetano-Anollés, G.; Rolfe, B.G.; Bauer, W.D. Extensive and specific responses of a eukaryote to bacterial quorum-sensing signals. Proc. Natl. Acad. Sci. 2003, 100, 1444–1449. [Google Scholar] [CrossRef] [PubMed]
- Feussner, I.; Polle, A. What the transcriptome does not tell—proteomics and metabolomics are closer to the plants’ patho-phenotype. Curr. Opin. Plant Biol. 2015, 26, 26–31. [Google Scholar] [CrossRef]
- Balmer, D.; de Papajewski, D.V.; Planchamp, C.; Glauser, G.; Mauch-Mani, B. Induced resistance in maize is based on organ-specific defence responses. Plant J. 2013, 74, 213–225. [Google Scholar] [CrossRef]
- Berg, G.; Grube, M.; Schloter, M.; Smalla, K. Unraveling the plant microbiome: looking back and future perspectives. Front. Microbiol. 2014, 5, 71409. [Google Scholar] [CrossRef] [PubMed]
- Sasse, J.; Martinoia, E.; Northen, T. Feed your friends: do plant exudates shape the root microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Gao, W.; Sun, H.-X.; Xiao, H.; Cui, G.; Hillwig, M.L.; Jackson, A.; Wang, X.; Shen, Y.; Zhao, N.; Zhang, L. Combining metabolomics and transcriptomics to characterize tanshinone biosynthesis in Salvia miltiorrhiza. BMC Genomics 2014, 15, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ratcliffe, R.G.; Shachar-Hill, Y. Measuring multiple fluxes through plant metabolic networks. Plant J. 2006. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Fu, X.; Sharkey, T.D.; Shachar-Hill, Y.; Walker, B.J. The metabolic origins of non-photorespiratory CO2 release during photosynthesis: A metabolic flux analysis. Plant Physiol. 2021, 186, 297–314. [Google Scholar] [CrossRef] [PubMed]
- Schwender, J.; Goffman, F.; Ohlrogge, J.B.; Shachar-Hill, Y. Rubisco without the Calvin cycle improves the carbon efficiency of developing green seeds. Nature 2004, 432, 779–782. [Google Scholar] [CrossRef] [PubMed]
- Salon, C.; Raymond, P.; Pradet, A. Quantification of carbon fluxes through the tricarboxylic acid cycle in early germinating lettuce embryos. J. Biol. Chem. 1988, 263, 12278–12287. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Wieloch, T.; Kaste, J.A.M.; Shachar-Hill, Y.; Sharkey, T.D. Reimport of carbon from cytosolic and vacuolar sugar pools into the Calvin–Benson cycle explains photosynthesis labeling anomalies. Proc. Natl. Acad. Sci. 2022, 119, e2121531119. [Google Scholar] [CrossRef] [PubMed]
- Dieuaide-Noubhani, M.; Raffard, G.; Canioni, P.; Pradet, A.; Raymond, P. Quantification of Compartmented Metabolic Fluxes in Maize Root Tips Using Isotope Distribution from 13C-or 14C-Labeled Glucose∗. J. Biol. Chem. 1995, 270, 13147–13159. [Google Scholar] [CrossRef]
- Xu, Y.; Koroma, A.A.; Weise, S.E.; Fu, X.; Sharkey, T.D.; Shachar-Hill, Y. Daylength variation affects growth, photosynthesis, leaf metabolism, partitioning, and metabolic fluxes. Plant Physiol. 2024, 194, 475–490. [Google Scholar] [CrossRef]
- Kruger, N.J.; Masakapalli, S.K.; Ratcliffe, R.G. Strategies for investigating the plant metabolic network with steady-state metabolic flux analysis: lessons from an Arabidopsis cell culture and other systems. J. Exp. Bot. 2012, 63, 2309–2323. [Google Scholar] [CrossRef] [PubMed]
- Schwender, J.; Ohlrogge, J.B.; Shachar-Hill, Y. A flux model of glycolysis and the oxidative pentosephosphate pathway in developing Brassica napus embryos. J. Biol. Chem. 2003. [Google Scholar] [CrossRef] [PubMed]
- Bago, B.; Pfeffer, P.E.; Abubaker, J.; Jun, J.; Allen, J.W.; Brouillette, J.; Douds, D.D.; Lammers, P.J.; Shachar-Hill, Y. Carbon export from arbuscular mycorrhizal roots involves the translocation of carbohydrate as well as lipid. Plant Physiol. 2003, 131, 1496–1507. [Google Scholar] [CrossRef] [PubMed]
- Rich, M.K.; Schorderet, M.; Bapaume, L.; Falquet, L.; Morel, P.; Vandenbussche, M.; Reinhardt, D. The petunia GRAS transcription factor ATA/RAM1 regulates symbiotic gene expression and fungal morphogenesis in arbuscular mycorrhiza. Plant Physiol. 2015, 168, 788–797. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Cheung, C.Y.M.; Hilbers, P.A.J.; Van Riel, N.A.W. Flux balance analysis of plant metabolism: the effect of biomass composition and model structure on model predictions. Front. Plant Sci. 2016, 7, 537. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Schmiege, S.C.; Sharkey, T.D. The oxidative pentose phosphate pathway in photosynthesis: a tale of two shunts. New Phytol. 2024. [Google Scholar] [CrossRef] [PubMed]
- Wilson, N.J.; Smith-Moore, C.M.; Xu, Y.; Edwards, B.; Hovary, C. La; Barampuram, S.; Li, K.; Aslett, D.; Ji, M.; Lin, X. Introduction of a condensed, reverse tricarboxylic acid cycle for additional CO2 fixation in plants. bioRxiv 2022, 2003–2022. [Google Scholar]
- Libourel, I.G.L.; Shachar-Hill, Y. Metabolic Flux Analysis in Plants: From Intelligent Design to Rational Engineering. Annu. Rev. Plant Biol. 2008. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Pang, N.; He, L.; Xu, Y.; Fu, X.; Tang, Y.; Shachar-Hill, Y.; Chen, S. Re-Programing Glucose Catabolism in the Microalga Chlorella sorokiniana under Light Condition. Biomolecules 2022, 12, 939. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, N.; Qiu, M.; Feng, H.; Vivanco, J.M.; Shen, Q.; Zhang, R. Enhanced rhizosphere colonization of beneficial Bacillus amyloliquefaciens SQR9 by pathogen infection. FEMS Microbiol. Lett. 2014, 353, 49–56. [Google Scholar] [CrossRef]
- Schläpfer, P.; Zhang, P.; Wang, C.; Kim, T.; Banf, M.; Chae, L.; Dreher, K.; Chavali, A.K.; Nilo-Poyanco, R.; Bernard, T. Genome-wide prediction of metabolic enzymes, pathways, and gene clusters in plants. Plant Physiol. 2017, 173, 2041–2059. [Google Scholar] [CrossRef] [PubMed]
- Wolfender, J.-L.; Marti, G.; Thomas, A.; Bertrand, S. Current approaches and challenges for the metabolite profiling of complex natural extracts. J. Chromatogr. A 2015, 1382, 136–164. [Google Scholar] [CrossRef] [PubMed]
- Misra, B.B.; van der Hooft, J.J.J. Updates in metabolomics tools and resources: 2014–2015. Electrophoresis 2016, 37, 86–110. [Google Scholar] [CrossRef] [PubMed]
- Fiehn, O.; Robertson, D.; Griffin, J.; van der Werf, M.; Nikolau, B.; Morrison, N.; Sumner, L.W.; Goodacre, R.; Hardy, N.W.; Taylor, C. The metabolomics standards initiative (MSI). Metabolomics 2007, 3, 175–178. [Google Scholar] [CrossRef]
- Goodacre, R. Water, water, every where, but rarely any drop to drink. Metabolomics 2014, 10, 5–7. [Google Scholar] [CrossRef]
- Bowen, B.P.; Northen, T.R. Dealing with the unknown: metabolomics and metabolite atlases. J. Am. Soc. Mass Spectrom. 2010, 21, 1471–1476. [Google Scholar] [CrossRef] [PubMed]
- Kind, T.; Wohlgemuth, G.; Lee, D.Y.; Lu, Y.; Palazoglu, M.; Shahbaz, S.; Fiehn, O. FiehnLib: mass spectral and retention index libraries for metabolomics based on quadrupole and time-of-flight gas chromatography/mass spectrometry. Anal. Chem. 2009, 81, 10038–10048. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Mochamad Afendi, F.; Altaf-Ul-Amin, M.; Takahashi, H.; Nakamura, K.; Kanaya, S. Metabolomics of medicinal plants: the importance of multivariate analysis of analytical chemistry data. Curr. Comput. Aided. Drug Des. 2010, 6, 179–196. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Gatto, L.; Wang, R.; Sachsenberg, T.; Uszkoreit, J.; Leprevost, F. da V.; Fufezan, C.; Ternent, T.; Eglen, S.J.; Katz, D.S. Ten simple rules for taking advantage of Git and GitHub. PLoS Comput. Biol. 2016, 12, e1004947. [Google Scholar] [CrossRef]
- Mendez, K.M.; Broadhurst, D.I.; Reinke, S.N. The application of artificial neural networks in metabolomics: a historical perspective. Metabolomics 2019, 15, 142. [Google Scholar] [CrossRef]
- Rinaudo, P.; Boudah, S.; Junot, C.; Thévenot, E.A. biosigner: A New Method for the Discovery of Significant Molecular Signatures from Omics Data. Front. Mol. Biosci. 2016, 3, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Toubiana, D.; Puzis, R.; Wen, L.; Sikron, N.; Kurmanbayeva, A.; Soltabayeva, A.; del Mar Rubio Wilhelmi, M.; Sade, N.; Fait, A.; Sagi, M. Combined network analysis and machine learning allows the prediction of metabolic pathways from tomato metabolomics data. Commun. Biol. 2019, 2, 214. [Google Scholar] [CrossRef] [PubMed]
- Sauer, U.; Heinemann, M.; Zamboni, N. Getting closer to the whole picture. Science (80-. ). 2007, 316, 550–551. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, A.; Kusano, M.; Redestig, H.; Arita, M.; Saito, K. Integrated omics approaches in plant systems biology. Curr. Opin. Chem. Biol. 2009, 13, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Coppola, M.; Diretto, G.; Digilio, M.C.; Woo, S.L.; Giuliano, G.; Molisso, D.; Pennacchio, F.; Lorito, M.; Rao, R. Transcriptome and metabolome reprogramming in tomato plants by Trichoderma harzianum strain T22 primes and enhances defense responses against aphids. Front. Physiol. 2019, 10, 745. [Google Scholar] [CrossRef] [PubMed]
- Savoi, S.; Wong, D.C.J.; Degu, A.; Herrera, J.C.; Bucchetti, B.; Peterlunger, E.; Fait, A.; Mattivi, F.; Castellarin, S.D. Multi-omics and integrated network analyses reveal new insights into the systems relationships between metabolites, structural genes, and transcriptional regulators in developing grape berries (Vitis vinifera L.) exposed to water deficit. Front. Plant Sci. 2017, 8, 1124. [Google Scholar] [CrossRef]
- Wanichthanarak, K.; Fahrmann, J.F.; Grapov, D. Genomic, proteomic, and metabolomic data integration strategies. Biomark. Insights 2015, 10, BMI-S29511. [Google Scholar] [CrossRef]
Figure 1.
Key Plant-Microbe Interactions and Their Implications. The schematic diagram illustrates various plant-microbe interactions and their effects on plant health and growth. Beneficial interactions include induced systemic resistance and systemic acquired resistance mediated by epiphytic (phylloplane) microbes, plant growth promotion by rhizobacteria and fungi, and nitrogen fixation by symbiotic bacteria. Conversely, detrimental interactions involve leaf pathogens, root pathogens, and the uptake of C, N, and P from soil organic matter by endocytosis of microbes via root hairs. Mycorrhizal fungi engage in mutualistic associations, while carbon root exudates attract heterotrophic microbes. Secreting enzymes by microbes compete for nutrients, and increased or decreased nutrient availability modulates the endocytosis of microbes.
Figure 1.
Key Plant-Microbe Interactions and Their Implications. The schematic diagram illustrates various plant-microbe interactions and their effects on plant health and growth. Beneficial interactions include induced systemic resistance and systemic acquired resistance mediated by epiphytic (phylloplane) microbes, plant growth promotion by rhizobacteria and fungi, and nitrogen fixation by symbiotic bacteria. Conversely, detrimental interactions involve leaf pathogens, root pathogens, and the uptake of C, N, and P from soil organic matter by endocytosis of microbes via root hairs. Mycorrhizal fungi engage in mutualistic associations, while carbon root exudates attract heterotrophic microbes. Secreting enzymes by microbes compete for nutrients, and increased or decreased nutrient availability modulates the endocytosis of microbes.
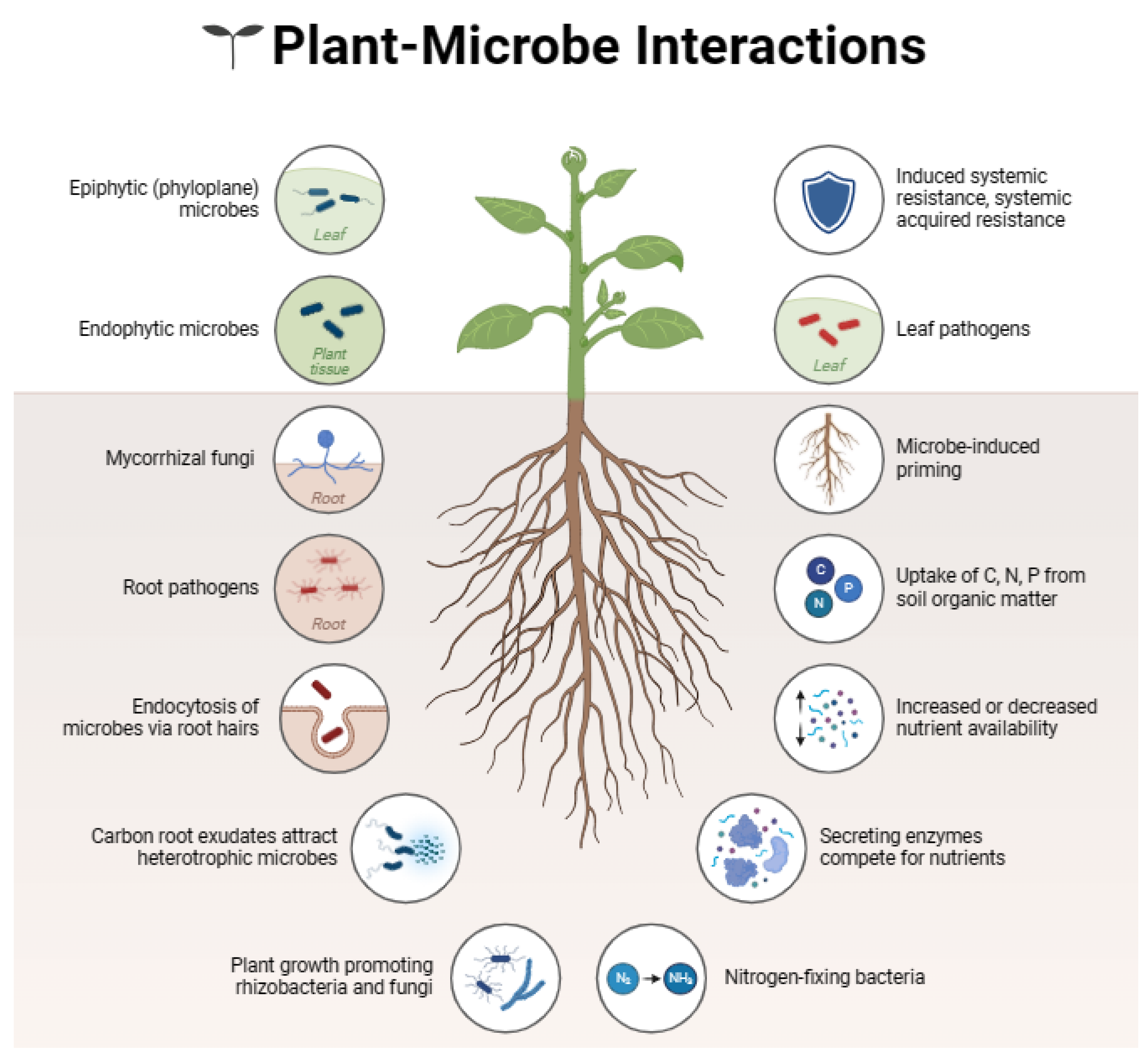
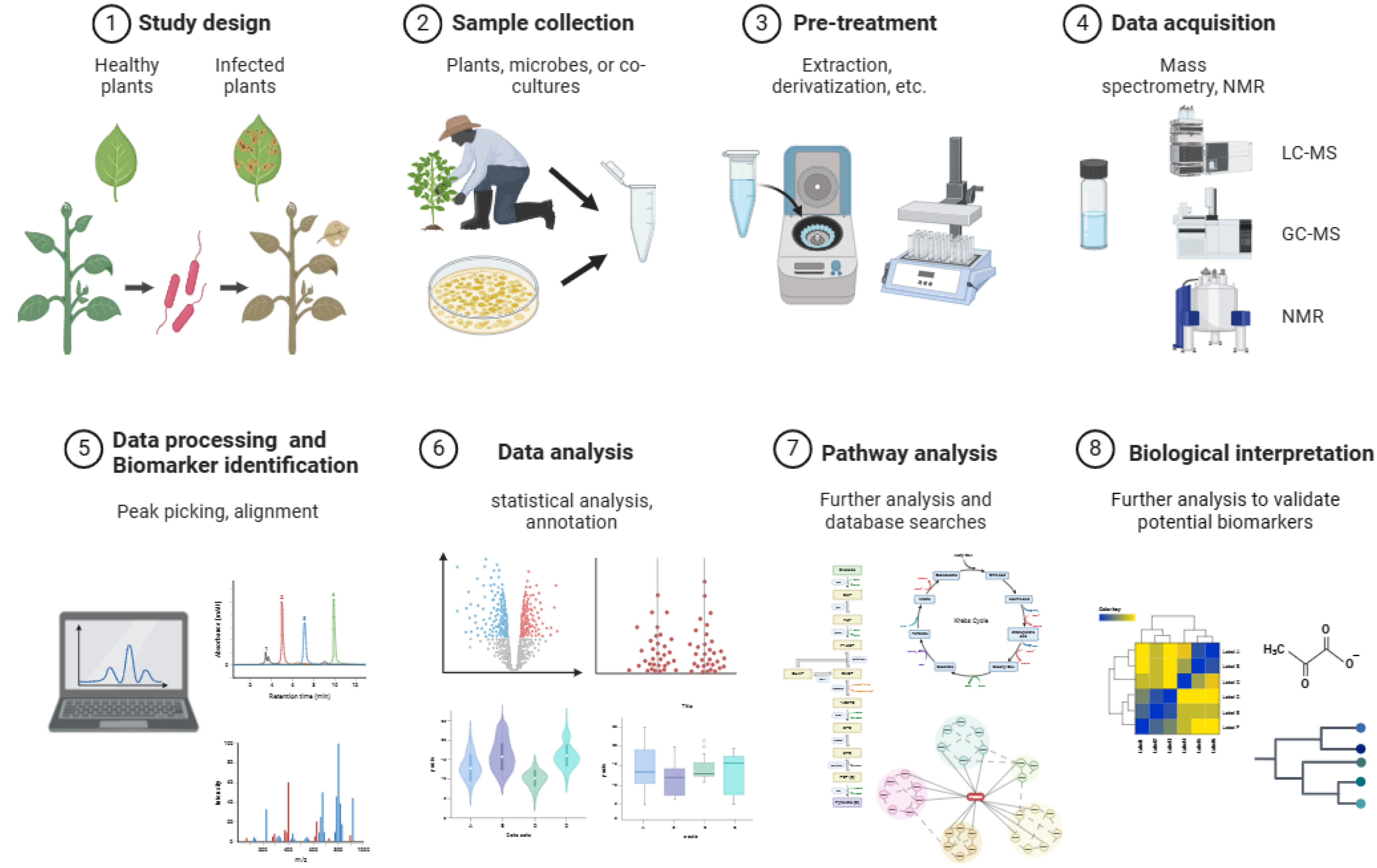
Figure 2.
Workflow of untargeted metabolomics for studying plant-pathogen interactions. The workflow begins with (1) careful study design, considering healthy and infected plants. (2) Sample collection involves obtaining plant, microbe, or co-culture samples. (3) Pre-treatment methods, such as extraction and derivatization, prepare the samples for (4) data acquisition using mass spectrometry and NMR spectroscopy. (5) Data processing and peak picking alignment convert raw data into a format suitable for (6) statistical analysis, including univariate and multivariate methods. (7) Pathway analysis and database searches identify metabolites and elucidate altered biological pathways. Finally, (8) biological interpretation integrates metabolomics data with other omics approaches to validate potential biomarkers and gain a systems-level understanding of the plant’s response to biotic stress.
Figure 2.
Workflow of untargeted metabolomics for studying plant-pathogen interactions. The workflow begins with (1) careful study design, considering healthy and infected plants. (2) Sample collection involves obtaining plant, microbe, or co-culture samples. (3) Pre-treatment methods, such as extraction and derivatization, prepare the samples for (4) data acquisition using mass spectrometry and NMR spectroscopy. (5) Data processing and peak picking alignment convert raw data into a format suitable for (6) statistical analysis, including univariate and multivariate methods. (7) Pathway analysis and database searches identify metabolites and elucidate altered biological pathways. Finally, (8) biological interpretation integrates metabolomics data with other omics approaches to validate potential biomarkers and gain a systems-level understanding of the plant’s response to biotic stress.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



