
Submitted:
08 June 2024
Posted:
11 June 2024
You are already at the latest version
Abstract
Family Parvoviridae consists of small, non-enveloped viruses with linear, single-stranded DNA genomes of approximately 4-6 kilobases, subdivided into 3 subfamilies: Parvovirinae, Densovirinae and Hamaparvovirinae. Parvoviruses of aquatic animals infect crustaceans, mollusks, and finfish. In this review, these parvoviruses, which are highly host-specific and are associated with mass morbidity and mortality in both farmed and wild aquatic animals are described. They include Cherax quadricarinatus densovirus (CqDV) in freshwater crayfish; Sea star-associated densovirus (SSaDV) in sunflower sea star on the Northeastern Pacific Coast; Clinch densovirus 1 in freshwater mussels in the Clinch River, Virginia, and Tennessee, USA, in subfamily Densovirinae; Hepatopancreatic parvovirus (HPV) and Infectious hypodermal and hematopoietic necrosis virus (IHHNV) in farmed shrimp worldwide; Syngnathid ichthamaparvovirus 1 in gulf pipefish in the Gulf of Mexico and parts of South America; and Tilapia parvovirus (TiPV) in farmed tilapia in China and Thailand, in the subfamily Hamaparvovirinae. In addition, virus megataxonomy has enabled the inclusion of novel parvoviruses detected in both diseased and healthy animals using metagenomic sequencing for virus discovery, such as the novel zander parvovirus from zander in Hungary, novel salmon parvovirus from sockeye salmon smolts in British Columbia-Canada, and Spawner-isolated mortality virus (SMV) from shrimp broodstock in Australia.
Keywords:
Parvoviridae
; Finfish
; Crustaceans
; Mollusks
; Pathogens
; Viruses
; Parvovirinae
; Densovirinae
; Hamaparvovirinae
1. Introduction
Members of the family Parvoviridae are small, non-enveloped viruses with linear, single-stranded (ss) DNA genomes of approximately 4-6 kb. The family is subdivided into 3 subfamilies: Parvovirinae, Densovirinae and Hamaparvovirinae. Viruses in Parvovirinae and Densovirinae are distinguished primarily by their respective ability to infect vertebrate hosts versus invertebrate hosts (insects, shrimp, and echinoderms). In contrast, those in the subfamily Hamaparvovirinae infect both vertebrate and invertebrate hosts, hence the subfamily name from the ancient Greek word meaning “together” [1]. Because of their small genome, parvoviruses require actively dividing host cells and are host and tissue-specific. Some cause diseases which range from sub-clinical to lethal. A few require co-infection with helper viruses from other families [2]. This review describes the reported aquatic animal parvoviruses and puts them in the context of terrestrial animal parvoviruses, both veterinary [3] and human parvoviruses [4], so as to add to the current knowledge on the family Parvoviridae.
Novel zander parvovirus from zander or pikeperch (Sander lucioperca), a freshwater fish in Hungary, potentially represents a new genus and a new species in the subfamily Parvovirinae [5] and possibly the first member in this subfamily to infect an aquatic animal host. The novel zander parvovirus was detected by viral metagenomics and PCR methods in fecal samples collected from fish showing no clinical signs, and the possibility of a dietary origin of the virus could not be excluded [5].
Members of the subfamily Densovirinae can be highly host-specific and lethal [6]. Mass mortality in invertebrates is a well-characterized consequence of densovirus infection, with Densovirinae examples of Cherax quadricarinatus densovirus (CqDV) now renamed Decapod aquambidensovirus 1 [previously Putative gill parvovirus] (genus Aquambidensovirus) in freshwater crayfish (Cherax quadricarinatus) [7,8], Sea star-associated densovirus in sunflower sea star (Pycnopodia helianthoides) on the Northeastern Pacific Coast [9], and Clinch densovirus 1 in freshwater mussel (Acrimonies pectorosa) in the Clinch River, Virginia, and Tennessee, USA [10].
Hamaparvovirinae members include hepatopancreatic parvovirus (HPV), now renamed Decapod hepanhamaparvovirus 1 (DHPV-1) in the genus Hepanhamaparvovirus [formerly genus Hepandensovirus]) and infectious hypodermal and hematopoietic necrosis virus (IHHNV) now renamed Decapod penstylhamaparvovirus 1 in genus Penstylhamaparvovirus [formerly genus Penstyldensovirus] of prawns and shrimp, and in the genus Ichthamaparvovirus, Syngnathid ichthamaparvovirus 1 of gulf pipefish (Syngnathus scovelli) and Tilapia parvovirus (TiPV) of Nile tilapia (Oreochromis niloticus). Hepanhamaparvoviruses are widespread and highly pathogenic, causing hepatopancreatic disease, and can constitute an economic threat in cultured shrimp populations on rare occasions when larvae from wild-caught shrimp are introduced [11]. Penstylhamaparvoviruses were first identified in the 1980s in Hawaii as pathogens responsible for an economically significant and virulent disease in farmed shrimp (infectious hypodermal and hematopoietic necrosis) [12,13]; they are widespread but are no longer a major economic problem because tolerant shrimp populations have been developed [14]. Ichthamaparvoviruses are parvoviruses of fish, including Tilapia parvovirus (TiPV), a recently identified viral infection associated with mass morbidity and mortality in farmed adult Nile tilapia in China [15] and farmed juvenile red hybrid tilapia in Thailand [16,17]. The novel fish parvovirus (tilapia parvovirus HMU-HKU-1) was first identified using next-generation sequencing (NGS) on fecal samples from crocodiles fed with tilapia [18].
1.1. Parvovirus Classification and Virion Properties
Parvoviruses are one of the smallest animal viruses known. “Parvus” is the Latin word for “small.” The virus particles are non-enveloped, T=1 icosahedra, 22-28 nm in diameter (Figure 1). They are extremely resistant to environmental conditions (pH 3-9; and 60oC for 1 hour) and survive for long periods outside the host cell, resulting in persistence in the environment, carriage on fomites, and wide dissemination [20]. The most reliable disinfection is achieved with household bleach (6% sodium hypochlorite). Most vertebrate parvoviruses hemagglutinate red blood cells.
The genome is linear ssDNA of positive sense or negative sense (or ambisense in subfamily Densovirinae) of approximately 4-6 kilobases. The genome encodes a large non-structural protein (NS1) with a helicase superfamily 3 (SF3) domain and the capsid protein (VP). Parvoviruses are classified in the same species if their NS1 proteins share more than 85% amino acid sequence identity while diverging greater than 15% from members of other genera and are classified in the same genus if they cluster together on phylogenetic analysis of their complete NS1 protein sequences and their SF3 helicase domains [1]. Thus, using these NS1-based criteria, the family Parvoviridae is subdivided into three subfamilies, Parvovirinae with ten genera that infect vertebrate hosts, Densovirinae with eight genera that infect invertebrate hosts, and Hamaparvovirinae with five genera that infect both vertebrate and invertebrate hosts. The evolutionary relationships of the different genera are shown in the phylogenetic tree in Figure 1 of Pénzes et al. [1]. At the megataxonomic level, the family Parvoviridae has been classified in the realm Monodnaviria, kingdom Shotokuvirae, phylum Cossaviricota, class Quintoviricetes, and order Piccovirales, based on its relations to other DNA virus taxa [1].
1.1.1. Parvovirus Genome Organization
The coding region in the genome is flanked by palindromic double-stranded hairpin ends required for replication, but many other characteristics vary between members of different genera. Figure 2 shows the genome organizations of members of the representative genera of the three subfamilies, Parvovirinae, Densovirinae, and Hamaparvovirinae [18]. All parvoviruses encode the non-structural (NS) protein gene(s) and genes of accessory proteins in negative-sense orientation on the left-hand side of the genome. In contrast, the structural capsid VP gene on the right-hand side may be in negative-sense orientation (subfamilies Parvovirinae and Hamaparvovirinae) or positive-sense orientation (i.e., ambisense genome in subfamily Densovirinae). The genome ends contain short, terminal repeats that form varying secondary structures, which create self-priming palindromic hairpin telomeres that function as viral DNA replication origins [21]. These secondary structures can either be the same or different at the 5′- and 3′-termini, leading to homotelomeric or heterotelomeric genomes, respectively and are consistent across a genus. Among the subfamilies Parvovirinae and Hamaparvovirinae, homotelomeric viruses package equal numbers of plus or minus stranded genomes in viral particles. A packaging bias toward one viral genome strand is observed in parvoviruses with heterotelomeric genomes. For example, some parvoviruses encapsidate only the negative-sense DNA strand (e.g., Carnivore protoparvovirus 1 such as Canine parvovirus, and genus Amdoparvovirus) [22]; others encapsidate different portions of either positive-sense or negative-sense DNA strand (e.g., Primate erythroparvovirus 1 or Human parvovirus B19) and members of genus Dependoparvovirus package both strands in separate particles in a ratio of 1:1). Many parvoviruses package predominantly negative DNA strands. The proportion of negative to positive strands packaged depends on the host cell infected. No parvovirus is known to package predominantly positive DNA strands; positive-strand DNA occurs in variable proportions, 1-50% (so the genome of parvoviruses is often described as ssDNA of negative sense). The NS gene forms one or more nonstructural proteins (NS1-NS3) via alternative mRNA splicing [23]. Parvovirus NS1 is a large multifunctional protein with strand and site-specific endonuclease (nicking) activity, ATPase activity, and SF3 helicase domain with 3′ to 5′ processivity, rolling circle replication initiator protein motifs, and DNA binding domains; it is absolutely required for virus replication [23] and also contributes to pathology [24]. Parvovirus capsids can be composed of up to 4 VPs (VP1-4) generated from a single ORF of the VP gene by alternative splicing, and all share a common large C-terminal region [23].
1.1.2. Virus Replication
Parvoviruses multiply in the nucleus producing large intranuclear inclusion bodies. Parvoviruses enter the cell by endocytosis and replicate their genome in the nucleus by “rolling hairpin replication,” a unidirectional single-strand displacement mechanism, using the viral self-priming hairpin telomeres as well as cellular polymerases, ligases, and other replication factors [25]. In many parvoviruses, the extended N-terminus of the VP1 protein includes the phospholipase A2 (PLA2) enzymatic domain that is essential for cell entry due to the need to release from the endosomal or lysosomal pathway [21,26]. Because of their small genome, parvoviruses lack a viral DNA polymerase and require actively dividing host cells, although they cannot induce cells into the S phase (Figure 3). Members of the genus Dependoparvovirus are defective and require helper adenovirus or herpesvirus for replication.
1.2. General Pathogenesis of Parvoviruses
In general, the disease outcome of a parvovirus infection is controlled by various factors [23]. For example, in the autonomous parvoviruses, virus replication is dependent on cellular functions provided in the S phase of the cell division cycle, i.e., when cellular DNA synthesis is occurring, a feature that is correlated with the pathogenic potential of these viruses, so tissues with high cellular division rates are disproportionately affected [23]. The parvoviruses found in aquatic animals are listed in Table 1.
2. Genus Aquambidensovirus
The term “Ambidensovirus” was coined in 2014 as the name of a genus in the family Parvoviridae containing members with an ambisense genome organization [41]. Aquambidensovirus contains those viruses known to infect aquatic animal hosts, which currently include two assigned species, Decapod aquambidensovirus 1, and Asteroid aquambidensovirus 1. Members of the genus share about 70% NS1 amino acid sequence identity with each other and ~30% with other members of the subfamily Densovirinae [1]. A novel densovirus, Clinch densovirus 1, had a 63% amino acid sequence identity with Periplaneta fuliginosa densovirus (AF192260), which infects smoky brown cockroaches (Periplaneta fuliginosa) in China, the closest phylogenetic relative in the GenBank database [10], and did not cluster with Asteroid aquambidensovirus 1 [1], suggesting it is a different species, possibly the third species in genus Aquambidensovirus.
2.1. Decapod aquambidensovirus 1 (Cherax Quadricarinatus Densovirus (CqDV))
Decapod aquambidensovirus 1 is the type species of the genus Aquambidensovirus, with only one virus, Cherax quadricarinatus densovirus (CqDV) [previously Putative gill parvovirus [7]. The virus was first identified in a single moribund freshwater crayfish (Cherax destructor) collected in South Australia [42]. It was subsequently identified in a redclaw crayfish (Cherax quadricarinatus) farm with chronic mortalities in mixed infection with presumptive hepatopancreatic reovirus [7] and in another commercial redclaw crayfish farm with mass mortality in northern Queensland, Australia [8]. The main histopathological lesion in Cherax quadricarinatus in the transmission trial conducted by Edgerton et al. [7] was hypertrophic gill nuclei. The mass mortality reported by Bowater et al. [8] occurred in juvenile animals over four weeks (96% cumulative mortality); affected crayfish were weak, anorexic, and lethargic. Experimentally inoculated crayfish showed gross signs of malaise, anorexia, and disorientation before dying, and the gills, cuticular epithelium, and epithelial cells of the foregut, midgut, and hindgut were the most heavily infected tissues; virions consistent with the parvovirus morphology were seen by electron microscopy in the enlarged nuclei of both naturally and experimentally infected crayfish [8]. In the infection trial of redclaw crayfish infected with CqDV conducted by Bochow [43], the clinical signs appeared from 17 to 57 days post-injection. The most obvious lesion consisted of blisters filled with a viscous, gelatinous substance on the inner membrane between the carapace and gills extending across the branchial cavity covering the top underside of the cephalothorax (shown in Figure 14.1 of Bochow [43]). Microscopically, large basophilic intranuclear inclusion bodies, characteristic of parvovirus infection, were present in ectodermal tissue cells of the gills, cuticular epithelium, and gastric sieve and hemocytes in the hemal spaces (shown in Figure 14.2 of Bochow [43]).
CqDV has one of the largest genomes in the family Parvoviridae (6334 nucleotides) (GenBank Accession # KP410261, [44]. Bochow [43] reported primers for molecular detection of CqDV by SYBR Green qPCR (CqDV 5 Fq 5′-CGCTGTGGAGAGTGCACTAGAGGC-3′; 2A Rq 5′-TCTGAATCAATCTCCTCACGATCGC-3′, amplicon size 281 bp). Analysis of the tissue tropism of CqDV in six organs using qPCR showed the branchial epithelium had the highest mean copy number mg-1 of tissue (1.22 x 106), followed by pleopod (1.19 x 106), antennal gland (5.53 x 105), gill (4.66 x 105), heart (1.32 x 105) and muscle (6.12 x 104) [43].
2.2. Asteroid aquambidensovirus 1 (Sea Star-Associated Densovirus (SSaDV))
The species Asteroid aquambidensovirus 1 comprises three viruses identified in sea stars and sea urchins (phylum Echinodermata) [9,45], all highly pathogenic [1]. Sea star-associated densovirus (SSaDV), the only one classified to date, was associated with an extensive outbreak of sea-star (asteroid) wasting disease (SSWD) (also known as “asteroid idiopathic wasting syndrome”) with mass mortality of captive asteroids in three species of common asteroid (Pycnopodia helianthoides, Pisaster ochraceus, and Evasterias troschelii) on the Northeastern Pacific Coast in 2013-2014 [9]. Environmental stressors may also play a role in the clinical outcome of infected sea stars [46,47] as a reexamination of the original metagenomic data found the virus to be commonly associated with apparently healthy or asymptomatic animals [48]. Clinical signs of SSWD include abnormal twisting of appendages, followed by the formation of white lesions and loss of turgor or a “deflated” appearance (deflation of arms and body), progressing to arm loss, necrosis, and rapid degradation leading to death [9,49].
Jackson et al. [48] reported primers for molecular detection of SSaDV by conventional PCR targeting VP1 (forward primer 5′-TGGCCACTCATCATGTCTCT-3′; reverse primer 5′-CTTGGGGTCCTTCATGAGC-3′). When this PCR assay was used to assess putative tissue tropism in three sea star species (Pisaster ochraceus, Evasterias troschelii, and Pisaster brevispinus), SSaDV was detected most frequently in the pyloric caeca (40/45 or 89%), followed by tube feet (17/47 or 36%), stomach (5/46 or 11%), body wall (5/47 or 11%), and gonads (4/42 or 10%) [48].
2.3. Clinch Densovirus 1
Clinch densovirus 1 is a novel densovirus linked to mass mortality in freshwater mussels, pheasantshell (Acrimonies pectorosa) in the Clinch River, Virginia and Tennessee, USA, since 2016 [10]. The virus was one of 17 novel viruses identified using hemolymph for metagenomic sequencing for virus discovery and the only one that was epidemiologically linked to morbidity. Clinch densovirus 1 is currently unassigned. Phylogenetic analysis showed it did not cluster with Asteroid aquambidensovirus 1 [1], suggesting it is a different species, possibly the third species in the genus Aquambidensovirus.
3. Genus Hepanhamaparvovirus
Hepanhamaparvovirus is one of three genera (together with genera Penstylhamaparvovirus and Ichthamaparvovirus) containing viruses known to infect aquatic hosts in the new subfamily Hamaparvovirinae (Figure 2). Members of this subfamily have an average of 30% amino acid sequence identity of their NS1 protein, and all species, like members of genus Aveparvovirus (subfamily Parvovirinae), lack the otherwise conserved phospholipase A2 domain in their VP1 proteins [1,23]. The genus Hepanhamaparvovirus has one species, Decapod hepanhamaparvovirus 1 (DHPV-1), which is also the type species of its former genus, Hepandensovirus.
3.1. Decapod hepanhamaparvovirus 1 (DHPV-1) (Hepatopancreatic Parvovirus (HPV))
The species Decapod hepanhamaparvovirus 1 (DHPV-1) was previously known as Hepatopancreatic parvovirus (HPV), formerly in the genus Hepandensovirus, and includes several genetically distinct strains from different shrimp and prawn species in different countries [50,51,52,53]. The genomes of the following ten strains have been completely sequenced: Penaeus monodon hepandensovirus 1 (PmoHDV1 (Thailand), GenBank Accession# DQ002873.1); Penaeus chinensis hepandensovirus (PchDV (China), Accession # NC_014357); Penaeus monodon hepandensovirus 2 (PmoHDV2 (Madagascar), Accession #s EU247528.1 and MT980830); Penaeus monodon hepandensovirus 3 (PmoHDV3 (Tanzania), Accession# EU588991.1); Penaeus merguiensis hepandensovirus (PmeDV (Australia), Accession# DQ458781.4); Penaeus monodon hepandensovirus 4 (PmoHDV4 (India), Accession# FJ410797.2); and Fenneropenaeus chinensis hepandensovirus (FchDV (South Korea), Accession# JN082231.1, (China) Accession# GU371276.1, and (Korea) Accession# AY008257). Another strain of DHPV-1 that has been refractory to PCR methods designed for DHPV-1 detection in P. monodon [54] occurs in cultivated giant river prawn Macrobrachium rosenbergii in Thailand [54,55] and Malaysia [56,57]. Most recently, Lee et al. [58] reported a novel genotype of DHPV-1 in P. vannamei (Pacific white shrimp) with approximately 70% sequence identity with all known DHPV-1s and with a unique ten amino acid deletion, and 3 and 1 amino acid insertions in the VP gene in a mixed infection with Enterocytozoon hepatopenaei in Taiwan. Thus, to date, four genotypes of DHPV-1 can be defined based on the VP gene sequence as shown in Figure 4 of Lee et al. [58]. Genotype I consists of strains from South Korea, China, Madagascar, and Tanzania; Genotype II strains from India, Indonesia, and Thailand; Genotype III strains from Australia and New Caledonia; and Genotype IV is the novel strain from Taiwan [58].
DHPV-1 is widespread and highly pathogenic, causing hepatopancreatic disease, and can constitute an economic threat in cultured shrimp populations on rare occasions when larvae from wild-caught shrimp are introduced. HPV was first described from farmed Penaeus merguiensis and P. indicus with a mixed infection with chlamydia in Singapore [59] and is considered to have later spread to wild shrimp in the Americas via importation of live infected Asian shrimp for aquaculture [60]. The host range of DHPV-1 includes at least 19 species of wild and cultured shrimp, prawns, and crabs worldwide [53]; it has been reported in many countries, including Australia, China, Korea, the Philippines, Indonesia, Malaysia, India, Kenya, Kuwait, Israel, and Taiwan, as well as from the Americas [53,58]. DPHV-1 was removed from the World Organization for Animal Health (WOAH) list of reportable pathogens as it was no longer associated with any significant negative economic repercussions in the aquaculture industry [61]. However, the virus remains in circulation in penaeid shrimp in China [61], Thailand [55], India [62], Taiwan [58], and several other countries, and can cause considerable losses in shrimp grow-out ponds without any obvious clinical manifestation [63]. Moreover, DPHV-1 infection is seldom observed alone in epizootics and has occurred in multiple infections with other more pathogenic viruses [32,53] which likely downplays its pathogenicity and economic significance. For example, there have been reports of co-infection of DHPV-1 and Enterocytozoon hepatopenaei (EHP) in P. vannamei [58,62], DHPV-1 and monodon baculovirus (MBV) in P. monodon [64,65], DHPV-1, MBV, and Yellow head virus (YHV) in P. monodon [66], and DHPV-1, MBV, and White spot syndrome virus (WSSV) in P. monodon [32].
DHPV-1 infects the epithelial cells of the hepatopancreas and midgut of shrimp, with infected individuals showing non-specific gross signs, including an atrophied hepatopancreas, anorexia, retarded growth, and reduced preening activities—resulting in epifouling in gills and appendages [67]; most DPHV-1-infected juvenile shrimp simply grow very slowly stopping at approximately 6 cm in length, weighing only about 5 g [30]. Mortalities during the larval stages have been reported in Australia in P. chinensis [31] and India in P. monodon [32]. Histopathology lesions in the hepatopancreas include basophilic inclusions within enlarged nuclei of tubule epithelial cells [30]. As is characteristic of autonomous parvoviruses, the actively dividing cells (E- and F-cells) at the distal ends of hepatopancreatic tubules show the most HPV inclusions [63,68].
Molecular diagnostic methods (conventional PCR, Real-time PCR, and in-situ hybridization) are instrumental in confirming the etiological role of DPHV-1 in hepatopancreas pathology. PCR methods are useful for screening cultured shrimp for HPV using harmless samples of small appendages or feces and for rapid and easy screening of large numbers of potential hosts and life stages as potential carriers [30]. To overcome the sequence diversity among different strains of DHPV-1 [50,51,52,53,69], Srisala et al. [55] developed a universal semi-nested PCR method to detect DHPV-1 in crustaceans by using primer sequences designed from the highly conserved region of the genome (nucleotide positions 1538 to 1887 in DHPV-1 NCBI accession no. DQ002873.1) (DHPV-U 1538 F 5′-CCTCTTGTTACATTTTACTC-3′; DHPV-U 1887 R 5′-GATGTCTTCTGTAGTCC-3′, amplicon size 350 bp, and DHPV-U 1622 F 5′-AAGTTTGCACAGTGGTTGT-3′, amplicon size 266 bp). These sequences were also used to make a DIG-labeled probe for in situ hybridization assays to localize DHPV-1 sequences in the histopathology lesions [55].
Currently, there is no anti-viral therapy for any viral diseases in shrimp. Therefore, biosecurity and genetically resistant lines remain the cornerstone in managing viral diseases [63]. Gene silencing using the RNA interference (RNAi) approach has been reported for both HPV [70] and IHHNV via injection. However, the oral delivery of RNAi molecules remains a challenge, and the utility of RNAi-based therapy has yet to be materialized in shrimp aquaculture [63].
4. Genus Penstylhamaparvovirus
The genus Penstylhamaparvovirus in the new subfamily Hamaparvovirinae with one species, Decapod penstylhamaparvovirus 1 [1], which is also the type species of its former genus, Penstyldensovirus, has four viruses (Penaeus stylirostris penstyldensovirus 1 & 2 and Penaeus monodon penstyldensovirus 1 & 2). Penaeus stylirostris penstyldensovirus 1 (PstDV1) is also known as the infectious hypodermal and hematopoietic necrosis virus (IHHNV) [2]. IHHNV was first identified as a pathogen responsible for an economically significant and virulent disease in farmed shrimp (infectious hypodermal and hematopoietic necrosis) in Hawaii, USA, in 1981 [12,13], which led to the collapse of penaeid shrimp farming in the Americas [71]. Phylogenetic analysis revealed the introduction of IHHNV to the American continent in the 1970s in imported P. monodon aquaculture stocks from Southeast Asia [72,73].
Based on 24 complete IHHNV genome sequences, the virus has been divided into five genotypes, infectious types I (in Australia), II (in the USA and Southeast Asia), and III (East Asia), and non-infectious types A (in Madagascar, Australia, Thailand, and India) and B (in Tanzania and Mozambique) [74]. The non-infectious form of IHHNV are endogenous viral element (EVE) sequences, lacking hairpins [75,76,77], that is inserted into the shrimp host genome following chronic or persistent infection [78] and could yield false-positive results for the diagnosis of shrimp infection with IHHNV. These sequences would likely have been eliminated unless they provide beneficial effects, such as EVE-derived immunity (EDI, e.g., mediated by TRIM5a and APOBEC). The IHHNV-EVE has been found integrated into chromosome 35 of the P. monodon genome, and IHHNV-EVE-related sequences are also present in P. vannamei [73,79].
IHHNV is listed by the World Organization for Animal Health (WOAH) [36] as an internationally notifiable disease due to its association with significant mortality in Pacific blue leg shrimp (Penaeus (Litopenaeus) stylirostris) and runt deformity syndrome (RDS) in Pacific white shrimp (Penaeus (Litopenaeus) vannamei). The virus is widespread [80] and has been reported in 32 countries to date in America, Asia, Oceania, and Africa [74,80,81,82]. In addition, it has been detected in about 30 species of shellfish, including in wild and farmed penaeid shrimps P. stylirostris [72], P. vannamei [83,84], P. monodon [85] and several others [74], in non-penaeid shrimp Macrobrachium rosenbergii [35,86], in crayfish Procambarus clarkii [87,88] and Cherax quadricarinatus [89], and crabs Hemigrapsus penicillatus, Neohelice granulate, Callinectes arcuatus, and Sesarma reticulatum [81,90,91,92] and in bivalve shellfish [93].
HHNV is still an important virus threatening shrimp aquaculture [74,82] even though tolerant shrimp populations have been developed [14,36]. IHHNV infection causes an acute disease with 80-100% mortality in postlarvae and juveniles of P. stylirostris [34] and postlarvae of M. rosenbergii [35]. The affected animals stop swimming, tumble, and then slowly sink to the bottom of the pond, and usually, they are ingested by healthy shrimps [94]. Very high mortality was reported in experimental IHHNV infection of crayfish P. clarkii (19/20 animals died) [87]. IHHNV infection causes chronic disease, runt-deformity syndrome (RDS), characterized by slow growth and deformities in the exoskeleton such as a bent (45o to 90o bend to left or right) or otherwise deformed rostrum, a deformed sixth abdominal segment, wrinkled antennal flagella, cuticular roughness, ‘bubble-heads’, and other cuticular deformities, without mortality, in juvenile of P. vannamei and P. monodon (Figure 4) [11,36,75,84,95,96], leading to 50% of the economic loss of shrimp industry [97]. IHHNV infection can cause 30%-90% growth retardation in juvenile P. vannamei [11,76]. Microscopic lesions include prominent intranuclear, Cowdry type A inclusion bodies characteristic of parvoviruses. The inclusion bodies observed in IHHNV occur in tissues of the ectodermal epithelium of fore- and hindgut, mesodermal origins like hematopoietic organs, antennal gland, and lymphoid organ [98]. Like other parvoviruses, IHHNV targets rapidly multiplying host cells, hence the severe infection of younger shrimp [76]. Adults of P. vannamei [99] and non-penaeid shrimp such as M. rosenbergii [74,86] serve as carriers of IHHNV without apparent clinical disease. Crayfish P. clarkii is also asymptomatic when naturally infected by IHHNV [88,100]. Red claw crayfish C. quadricarinatus could be a potential carrier of the virus [89], and crabs and bivalve shellfish can be asymptomatic carriers of IHHNV [74].
White spot syndrome virus (WSSV) is a differential diagnosis for IHHNV [96]. IHHNV has been reported to interfere with WSSV during mixed infection in penaeid shrimp, leading to higher survival rates compared to infection with WSSV alone [73,74,102,103]. Analysis of genetic diversity among IHHNV isolates in the Gulf of California found an unexpectedly high mutation rate that was comparable to that reported for RNA viruses, suggesting the potential for a new virulent strain to arise that might lead to epizootics similar to those observed in the early 1990s [104].
The preferred molecular method for diagnosing shrimp infection with IHHNV is conventional PCR using the primers IHHNV309F/R [105], which are recommended by the World Organization for Animal Health [36] to specifically detect infectious IHHNV forms and exclude the noninfectious related sequences. Dhar et al. [106] reported primers for molecular detection of IHHNV by SYBR Green qPCR (313F Forward 5′-AGGAGACAACCGACGACATCA-3′; 363R Reverse 5′-CGATTTCCATTGCTTCCATGA-3′). The SYBR Green qPCR was 2000-fold more sensitive than the conventional PCR [106]. Sequences of the whole IHHNV genome can be obtained using conventional PCR and the eight specific primers designed by Silva et al. [107]. In situ hybridization using IHHNV-specific DNA probes such as BA402 (available in kit form from DiagXotics Inc., 27 Cannon Rd., Wilton, CT 06897, USA) can be used on tissue sections preserved in Davidson’s, AFA or formalin [108].
IHHNV infection control and prevention are achieved through stocking good quality disease-free seeds, optimum rearing conditions, and good management practices on the farm [97].
5. Genus Ichthamaparvovirus
The genus Ichthamaparvovirus in the new subfamily Hamaparvovirinae (Figure 2) has at least two species, Syngnathid ichthamaparvovirus 1 (Syngnathus scovelli chapparvovirus) (the type species) and Tilapia parvovirus (TiPV). Syngnathus scovelli chapparvovirus has partially sequenced hairpins, suggesting that the genus is heterotelomeric [1].
5.1. Syngnathid ichthamaparvovirus 1
The near-complete genome sequence of Syngnathid ichthamaparvovirus 1 was identified in previously unreported DNA sequences by screening published whole genome sequencing (WGS) data of a gulf pipefish’s homogenized gill, muscle, and male brood pouch tissue (Syngnathus scovelli) [1,33]. Therefore, it is not known if this virus is associated with clinical disease. In a different syngnathid fish (family Syngnathidae), the tiger tail seahorse (Hippocampus comes), an endogenous viral element (EVE) with 70% NS1 amino acid sequence identity to Syngnathid ichthamaparvovirus 1 was found [33].
5.2. Tilapia Parvovirus (TiPV)
Tilapia parvovirus (TiPV) is an emerging pathogen in tilapia aquaculture, having been detected in China, Thailand and India. TiPV was first identified using next-generation sequencing (NGS) on fecal samples from crocodiles in China fed with tilapia [18]. Further investigation using a novel semi-nest PCR assay with primers targeting the viral NS1 gene detected the virus in tilapia intestine samples [18]. TiPV was subsequently associated with mass morbidity and 60-70% mortality in China’s farmed adult Nile tilapia (500-600 g) [15]. Clinical signs in affected fish included lethargy, anorexia, change in swim behavior, multifocal hemorrhage, and ocular lesions [15]. Microscopic lesions included splenic necrosis, encephalitis, nephritis, hepatitis, and gill branchitis [15] (Figure 5). Virions consistent with the parvovirus morphology were seen by electron microscopy in the cytoplasm and nucleus of cells of the heart, spleen, kidneys, brain, gills, and intestine [15]. The kidney and spleen were positive for ISH, PCR, and IFA [15]. The TiPV was isolated and propagated in tilapia brain cells (TiB) and induced a typical cytopathic effect (CPE) after three days post-infection (dpi) (Figure 6). This virus was used to infect adult tilapia experimentally, and clinical disease symptoms similar to those observed naturally were replicated [15]. In a more recent case of TiPV infection in adult Nile tilapia in Thailand, Dong et al. [109] reported Cowdry type A inclusion bodies in acinar cells of the pancreas as a diagnostic histopathological feature (i.e., pathognomonic of TiPV infection), which is also diagnostic of other parvoviral infections in shrimp and terrestrial species [109].
Recently, TiPV has been reported in mixed infections with Tilapia lake virus (TiLV), family Amnoonviridae, and polymicrobial infections in farmed juvenile red hybrid tilapia in Thailand [16,17], and in farmed Nile tilapia (Oreochromis niloticus) in India [110]. Dong et al. [109] reported TiPV in mixed infection with Streptococcus agalactiae in adult Nile tilapia in Thailand. In the disease outbreak investigated by Piewbang et al. [17], the mortality rate among farms reached 50-75%, with the most affected fish weighing 10-30 g, with lesser affected fish weighing 300-800 g. In the disease investigation reported by Yamkasem et al. [16], it was noted that differential diagnosis of TiPV and TiLV was difficult because there were no pathognomonic clinical signs, and no fish was found to be infected with TiPV alone. Moreover, the Thailand TiPV isolate (strain KU01-TH/2020’ (MW685502)) had a sequence identity of 98.74% to the virus first isolated in China (MT393593) [16]. In the report by Rajendran et al. [110], in one geographical region in India TiPV was detected along with TiLV and/or Aeromonas spp whereas in another region, fish were apparently healthy and only TiPV could be detected in the fish samples.
Liu et al. [15] reported primers for molecular detection of TiPV by PCR targeting the NS1 gene regions: conventional PCR (TiPV-F 5′-GAGATGGTGTGAAAATGAACGGG-3′; TiPV-R 5′-CTATCTCCTCGTTGCTCGGTGTATC-3′, amplicon size 534 bp) and SYBR Green qPCR (TiPV-Fq 5′-GCACCACAGCTGAGTACAAC-3′; TiPV-Rq 5′-AACTGCTCGGCTATCTCCTC-3′, amplicon size 134 bp). Analysis of TiPV distribution and viral loading in nine different tissues of naturally infected tilapia using qPCR showed the highest genome copies in kidney (3.5x107.32±0.21/mg) and spleen (4.2x107.12±0.35/mg) followed by intestine (4.3x106.37±0.36/mg), heart (5.1x106.25±0.27/mg), and brain (1.8x10 6.09±0.24/mg), with the lowest genome copies in gill (1.8x104.32±0.17/mg), liver (3.5x104.15±0.26/mg) and eye (2.8x103.78±0.32/mg) [15]. In addition, PCR revealed the prevalence of TiPV infection in six cities across three different provinces in China to range from 22.6% to 64.6% [15]. The spleen appears to be the best tissue for detecting TiPV by RT-PCR in apparently healthy tilapia [110].
The high pathogenicity of TiPV (highly fatal to adult tilapia [15]) dictates its host range be determined and adequate control measures developed against TiPV disease in tilapia aquaculture.
6. Unassigned
6.1. Novel Salmon Parvovirus from Sockeye Salmon
A full genome sequence of a novel salmon parvovirus—the first parvovirus to be identified in a fish species was obtained from sockeye salmon smolts from British Columbia-Canada. However, it is not known yet if it is associated with disease [37]. The novel salmon parvovirus was detected using NGS for virus discovery. It was found in a high load in sockeye salmon smolts migrating to the ocean, with load and prevalence generally declining from summer to fall. Prevalence varied greatly among stocks and years [37]. The full genome sequence of this novel salmon parvovirus is not publicly available, and its taxonomic location in the Parvoviridae family remains unassigned.
6.2. Novel Zander Parvovirus
Novel zander parvovirus (zander/M5/2015/HUN, OK236393) from zander or pikeperch (Sander lucioperca), a freshwater fish in Hungary, potentially represents a new genus and a new species in the subfamily Parvovirinae [5] (Figure 7). The novel zander parvovirus was detected by viral metagenomics and PCR methods in 3 out of 7 (42.8%) fecal samples collected from fish showing no clinical signs [5]. While the novel zander parvovirus may be the first member in the subfamily Parvovirinae to infect an aquatic animal host, its origin, whether from fish tissues or the fish diet, was not established [5]. Furthermore, one of the positive zander fecal samples also contained a potentially novel fish-origin picornavirus, family Picornaviridae [111], as a co-infection [5].
A sequence-specific screening primer pair (ZanderParvoscreen-F, 5′-GGCTAATCATCAAACAGGAAAGA-3′; ZanderParvo-screen-R, 5′-AGCTCC CACCACTTAATATCTT-3′) targeting the NS1 region of the viral genome identifies a 492 bp product [5].
6.3. Spawner-Isolated Mortality Virus (SMV)
Spawner-isolated mortality virus (SMV) belongs to the family Parvoviridae although its genome organization and relationship to other shrimp parvoviruses are unknown [113]. SMV causes mortalities in broodstock of Penaeus monodon with mid-crop mortality syndrome on grow-out farms in northern Australia [39,113,114] and redclaw crayfish Cherax quadricarinatus in northern Australia [115]. However, it is not known whether the virus is transferred from shrimp to crayfish or vice versa [115].
In laboratory challenge experiments, SMV was lethal to P. monodon, P. esculentus, P. japonicus, P. merguiensis, and Metapenaeus ensis, with mortality reaching 100% SMV [114]. In situ hybridization with an SMV-specific DNA probe detected the virus in endodermal tissues, including the distal ends of hepatopancreatic tubules, midgut and hindgut caecae, midgut, hindgut folds, and in the male reproductive tract, specifically in the terminal ampoule and the medial vas deferens as well as in the ovary and both the stromal matrix and spheroid cells of the lymphoid organ [114]. Owens et al. [39] investigated the association between the SMV status of P. monodon and P. merguiensis broodstock and the subsequent survival of their progeny. The prevalence in P. monodon spawners was 24% compared to 4% in P. merguiensis spawners, with 6.71% loss in ponds with postlarvae from SMV-positive spawners, supporting the hypothesis that SMV is vertically transmitted from spawners to postlarvae in hatcheries and causes reduced survival of progeny [39].
6.4. Crangon crangon Parvo-Like Virus (CcPaLV)
Crangon crangon parvo-like virus (CcPaLV) was discovered in a virome study of the European brown shrimp (Crangon crangon) using NGS [38]. That study described near-complete genomes of 16 novel viruses, most of which were distantly related to unclassified viruses or viruses belonging to the Picornavirales, Bunyavirales, Nudiviridae, Parvoviridae, Flaviviridae, Hepeviridae, Tombusviridae, Narnaviridae, Nodaviridae, and Sobemovirus. In addition, the study observed a difference in virome composition between muscle and hepatopancreatic tissue, suggesting a distinct tissue tropism of several of these viruses [38].
CcPaLV is highly divergent from known members of the subfamily Parvovirinae; it clusters between the subfamilies Parvovirinae and Densovirinae, although it is slightly more related to the subfamily Parvovirinae [38]. It remains to be seen whether this novel virus represents a novel subfamily in the family Parvoviridae or a very distinct genus in the subfamily Parvovirinae [38].
7. Concluding Remarks
From this review, it should be evident that the pathogenesis of aquatic animal (finfish, crustaceans, and mollusks) parvovirus diseases is comparable to that of terrestrial animal parvovirus diseases. These viruses cause diseases which range from sub-clinical to lethal such as mass morbidity and mortality in naïve animal populations, particularly in young (juvenile) animals. Virus replication takes place in the nucleus and requires host cell functions of late S phase or early G2 phase of the cell division cycle, a requirement for cycling cells that is the basis for many aspects of the pathogenesis of parvovirus infections. Because of their small genome, parvoviruses target tissues with a high cell turnover (i.e., autonomous parvoviruses) or require co-infection with helper viruses (i.e., defective parvoviruses). Moreover, the differences in the immune system of aquatic animals compared to terrestrial animals dictate that vaccination is not as effective in controlling aquatic animal parvoviruses as it is for terrestrial animal parvoviruses; in fact, it is not an option in crustaceans and mollusks.
Author Contributions
Conceptualization, F.K; investigation, data curation, writing—original draft preparation, F.K; writing—review and editing, F.K. M.K. and M.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author..
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Pénzes, J.J.; Söderlund-Venermo, M.; Canuti, M.; Eis-Hübinger, A.M.; Hughes, J.; Cotmore, S.F.; Harrach, B. 2020. Reorganizing the family Parvoviridae: a revised taxonomy independent of the canonical approach based on host association. Archives of Virology 165, 2133-2146. [CrossRef]
- Cotmore, S.F.; Agbandje-McKenna, M.; Canuti, M.; Chiorini, J.A.; Eis-Hubinger, A.-M.; Hughes, J.; Mietzsch, M.; Modha, S.; Ogliastro, M.; Penzes, J.J.; Pintel, D.J.; Qiu, J.; Soderlund-Venermo, M.; Tattersall, P.; Tijssen, P. ; ICTV Report Consortium, 2019. ICTV Virus Taxonomy Profile: Parvoviridae. Journal of General Virology 100, 367-368. [CrossRef]
- MacLachlan, N.J.; Dubovi, E.J. 2017. Chapter 12, Parvoviridae. In N. J. MacLachlan and E. J. Dubovi (ed.), Fenner’s Veterinary Virology, 5th ed. Academic Press, pp. 245-257. [CrossRef]
- Qiu, J.; Söderlund-Venermo, M.; Young, N.S. 2017. Human parvoviruses. Clinical Microbiology Reviews 30, 43-113. [CrossRef]
- Reuter, G.; Boros, Á.; Mátics, R.; Altan, E.; Delwart, E.; Pankovics, P. 2022. A novel parvovirus (family Parvoviridae) in a freshwater fish, zander (Sander lucioperca). Archives of Virology 167, 1163-1167. [CrossRef]
- Fédière, G. 2000. Epidemiology and pathology of Densovirinae, p. 1-11. In S. Faisst and J. Rommelaere (eds), Parvoviruses. From molecular biology to pathology and therapeutic uses. Karger, Basel, Switzerland. https://horizon.documentation.ird.fr/exl-doc/pleins_textes/pleins_textes_7/b_fdi_59-60/010025973.pdf.
- Edgerton, B.F.; Webb, R.; Anderson, I.G.; Kulpa, E.C. 2000. Description of a presumptive hepatopancreatic reovirus, and a putative gill parvovirus, in the freshwater crayfish Cherax quadricarinatus. Diseases of Aquatic Organisms 41, 83-90. [CrossRef]
- Bowater, R.O.; Wingfield, M.; Fisk, A.; Condon, K.M.L.; Reid, A.; Prior, H.; Kulpa, E.C. 2002. A parvo-like virus in cultured redclaw crayfish Cherax quadricarinatus from Queensland, Australia. Diseases of Aquatic Organisms 50, 79-86. [CrossRef]
- Hewson, I.; Button, J.B.; Gudenkauf, B.M.; Miner, B.; Newton, A.L.; Gaydos, J.K.; Wynne, J.; Groves, C.L.; Hendler, G.; Murray, M.; Fradkin, S.; Breitbart, M.; Fahsbender, E.; Lafferty, K.D.; Kilpatrick, A.M.; Miner, C.M.; Raimondi, P.; Lahner, L.; Friedman, C.S.; Daniels, S.; Haulena, M.; Marliave, J.; Burge, C.A.; Eisenlord, M.E.; Harvell, C.D. 2014. Densovirus associated with sea-star wasting disease and mass mortality. Proceedings of the National Academy of Science, USA, 111, 17278-17283. [CrossRef]
- Richard, J.C.; Leis, E.; Dunn, C.D.; Agbalog, R.; Waller, D.; Knowles, S.; Putnam, J.; Goldberg, T.L. 2020. Mass mortality in freshwater mussels (Actinonaias Pectorosa) in the Clinch River, USA, linked to a novel densovirus. Sci. Rep. 10, 14498. [CrossRef]
- Kalagayan, H. et al. 1991. IHHN virus as an etiological factor in runt-deformity syndrome (RDS) of juvenile Penaeus vannamei cultured in Hawaii. Journal of the World Aquaculture Society 22, 235-243. [CrossRef]
- Lightner, D.V.; Redman, R.M.; Bell, T.A. 1983. Infectious hypodermal and hematopoietic necrosis, a newly recognized virus disease of penaeid shrimp. Journal of Invertebrate Pathology 42, 62-70. [CrossRef]
- Lightner, D.V.; Redman, R.M.; Bell, T.A.; Brock, J.A. 1983b. Detection of IHHN virus in Penaeus stylirostris and P. vannamei imported into Hawaii. Journal of World Mariculture Society 14, 212-225. [CrossRef]
- Tang, K.F.J.; Durand, S.; White, B.L.; Redman, R.M.; Pantoja, C.R.; Lightner, D.V. 2000. Postlarvae and juveniles of a selected line of Penaeus stylirostris are resistant to infectious hypodermal and hematopoietic necrosis virus infection. Aquaculture 190, 203-210. [CrossRef]
- Liu, W.; Zhang, Y.; Ma, J.; Jiang, N.; Fan, Y.; Zhou, Y.; Cain, K.; Yi, M.; Jia, K.; Wen, H.; Liu, W.; Guan, W.; Zeng, L. 2020. Determination of a novel parvovirus pathogen associated with massive mortality in adult tilapia. PLoS Pathogens 16, e1008765. [CrossRef]
- Yamkasem, J.; Tattiyapong, P.; Gorgoglione, B.; Surachetpong, W. 2021. Uncovering the first occurrence of Tilapia parvovirus in Thailand in tilapia during co-infection with Tilapia tilapinevirus. Transboundary and Emerging Diseases 00:1– 9. [CrossRef]
- Piewbang, C.; Tattiyapong, P.; Khemthong, M.; Lachroje, S.; Boonrungsiman, S.; Kasantikul, T.; Surachetpong, W.; Techangamsuwan, S. 2022. Dual infections of tilapia parvovirus (TiPV) and tilapia lake virus (TiLV) in multiple tilapia farms: Their impacts, genetic diversity, viral tropism, and pathological effects. Aquaculture 550, 737887. [CrossRef]
- Du, J.; Wang, W.; Chan, J.F.-W.; Wang, G.; Huang, Y.; Yi, Y.; Zhu, Z.; Peng, R.; Hu, X.; Wu, Y.; Zeng, J.; Zheng, J.; Cui, X.; Niu, L.; Zhao, W.; Lu, G.; Yuen, K.-Y.; Yin, F. 2019. Identification of a novel ichthyic parvovirus in marine species in Hainan Island, China. Frontiers in Microbiology 10:2815. [CrossRef]
- ViralZone <italic>Parvoviridae</italic>. Available at: https://viralzone.expasy.org/103?outline=all_by_species (accessed May 27, 2024).
- Gordon, J.C.; Angrick, E.J. 1986. Canine parvovirus: environmental effects on infectivity. Am J Vet Res. 47(7), 1464-1467.
- Cotmore, S.F.; Tattersall, P. 2014. Parvoviruses: Small does not mean simple. Annu Rev Virol. 1(1), 517-537. [CrossRef]
- Cotmore, S.F.; Tattersall, P. 2005. Genome packaging sense is controlled by the efficiency of the nick site in the right-end replication origin of parvoviruses minute virus of mice and LuIII. Journal of Virology 79, 2287-2300. [CrossRef]
- Jager, M.C.; Tomlinson, J.E.; Lopez-Astacio, R.A.; Parrish, C.P.; Van de Walle, G.R. 2021. Small but mighty: old and new parvoviruses of veterinary significance. Virology Journal 18, 210. [CrossRef]
- Chen, A.Y.; Qiu, J. 2010. Parvovirus infection-induced cell death and cell cycle arrest. Future Virology 5, 731-743. [CrossRef]
- Söderlund-Venermo, M. 2019. Emerging human parvoviruses: The rocky road to fame. The Annual Review of Virology 6, 71-91. [CrossRef]
- Kailasan, S.; Agbandje-McKenna, M.; Parrish, C.R. 2015. Parvovirus family conundrum: What makes a killer? Annu Rev Virol. 2, 425-450. [CrossRef]
- Deng, X.; Yan, Z.; Cheng, F.; Engelhardt, J.F.; Qiu, J. 2016. Replication of an autonomous human parvovirus in non-dividing human airway epithelium is facilitated through the DNA damage and repair pathways. PLOS Pathogens 12, e1005399. [CrossRef]
- Deng, X.; Xu, P.; Zou, W.; Shen, W.; Peng, J.; Liu, K.; et al. 2017. DNA damage signaling is required for replication of human bocavirus 1 DNA in dividing HEK293 cells. Journal of Virology 91, e01831-16. [CrossRef]
- Bucci, C.; Francoeur, M.; McGreal, J.; Smolowitz, R.; Zazueta-Novoa, V.; Wessel, G.M.; Gomez-Chiarri, M. 2017. Sea Star Wasting Disease in Asterias forbesi along the Atlantic Coast of North America. PLoS One. 12(12):e0188523. [CrossRef]
- Flegel, T.W. 2006. Detection of major penaeid shrimp viruses in Asia, a historical perspective with emphasis on Thailand. Aquaculture 258, 1-33. [CrossRef]
- Spann, K.M.; Aldard, R.D.; Hudson, D.A.; Dyecroft, S.B.; Jones, T.C.; Voigt, M.O.C. 1997. Hepatopancreatic parvo-like virus (HPV) of Penaeus japonicus cultured in Australia. Dis. Aquat. Org. 31, 239-241. [CrossRef]
- Manivannan, S.; Otta, S.K.; Karunasagar, I.; Karunasagar, I. 2002. Multiple viral infections in Penaeus monodon prawn post larvae in an Indian hatchery. Diseases of Aquatic Organisms 48, 233-236. [CrossRef]
- Pénzes, J.J.; de Souza, W.M.; Agbandje-McKenna, M.; Gifford, R.J. 2019. An ancient lineage of highly divergent parvoviruses infects both vertebrate and invertebrate hosts. Viruses 11, 525. [CrossRef]
- Vega-Heredia, S.; Mendoza-Cano, F.; Sanchez-Paz, A. ; 2012. The infectious hypodermal and haematopoietic necrosis virus: A brief review of what we do and do not know. Transboundary and Emerging Diseases 59, 95–105. [CrossRef]
- Hsieh, C.Y.; Chuang, P.C.; Chen, L.C.; Tu, C.; Chien, M.S.; Huang, K.C.; Kao, H.F.; Tung, M.C.; Tsai, S.S. ; 2006. Infectious hypodermal and haematopoietic necrosis virus (IHHNV) infections in giant freshwater prawn, Macrobrachium rosenbergii. Aquaculture 258, 73-79. [CrossRef]
- WOAH (World Organization for Animal Health). 2021. Infection with infectious hypodermal and haematopoietic necrosis virus. Chapter 2.2.4. In Manual of Diagnostic Tests for Aquatic Animals. pp 138-153. Available at: https://www.woah.org/fileadmin/Home/eng/Health_standards/aahm/current/2.2.04_IHHN.
- Miller, K.M.; Trudel, M.; Patterson, D.A.; Schulze, A.; Kaukinen, K.; Li, S.; Ginther, N.; Ming, T.; Tabata, A. ; 2013. Are smolts healthier in years of good ocean productivity? North Pacific Anadromous Fish Commission, Technical Report No. 9, 165-168. https://www.npafc.org/wp-content/uploads/TechReport9.
- Van Eynde, B.; Christiaens, O.; Delbare, D.; Shi, C.; Vanhulle, E.; Yinda, C.K.; Matthijnssens, J.; Smagghe, G. 2020. Exploration of the virome of the European brown shrimp (Crangon crangon). Journal of General Virology 101, 651-666. [CrossRef]
- Owens, L.; McElnea, C.; Snape, N.; Harris, L.; Smith, M. 2003. Prevalence and effect of spawner-isolated mortality virus on the hatchery phases of Penaeus monodon and P. merguiensis in Australia. Diseases of Aquatic Organisms 53, 101-106. [CrossRef]
- NCBI Taxonomy Browser. Available at: https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi.
- Cotmore, S.F.; Agbandje-McKenna, M.; Chiorini, J.A.; Mukha, D.V.; Pintel, D.J.; Qiu, J.; Soderlund-Venermo, M.; Tattersall, P.; Tijssen, P.; Gatherer, D. 2014. The family Parvoviridae. Archives of Virology 159, 1239-1247. [CrossRef]
- Edgerton, B.; Webb, R.; Wingfield, M. 1997. A systematic parvo-like virus in the freshwater crayfish Cherax destructor. Diseases of Aquatic Organisms 29:73-78. [CrossRef]
- Bochow, S. 2016. Characterisation of Cherax quadricarinatus densovirus: the first virus characterised from Australian freshwater crayfish. PhD thesis, James Cook University. Available at: https://researchonline.jcu.edu.au/48889/.
- Bochow, S.; Condon, K.; Elliman, J.; Owens, L. 2015. First complete genome of an Ambidensovirus; Cherax quadricarinatus densovirus, from freshwater crayfish Cherax quadricarinatus. Mar Genomics 3:305-312. [CrossRef]
- Gudenkauf, B.M.; Eaglesham, J.B.; Aragundi, W.M.; Hewson, I. 2014. Discovery of urchin-associated densoviruses (family Parvoviridae) in coastal waters of the Big Island, Hawaii. Journal of General Virology 95, 652-658. [CrossRef]
- Hewson, I.; Bistolas, K.S.I.; Quijano Cardé, E.M.; Button, J.B.; Foster, P.J.; Flanzenbaum, J.M.; Kocian, J.; Lewis, C.K. ; 2018. Investigating the complex association between viral ecology, environment, and northeast Pacific sea star wasting. Frontiers in Marine Science 5, 1-14. [CrossRef]
- Aquino, C.A.; Besemer, R.M.; DeRito, C.M.; Kocian, J.; Porter, I.R.; Raimondi, P.T.; Rede, J.E.; Schiebelhut, L.M.; Sparks, J.P.; Wares, J.P.; Hewson, I. 2021. Evidence that microorganisms at the animal-water interface drive sea star wasting disease. Frontiers in Microbiology 11, 610009. [CrossRef]
- Jackson, E.W.; Wilhelm, R.C.; Johnson, M.R.; Lutz, H.L.; Danforth, I.; Gaydos, J.K.; Hart, M.W.; Hewson, I. 2021. Diversity of sea star-associated densoviruses and transcribed endogenous viral elements of densovirus origin. Journal of Virology 95, e01594-20. [CrossRef]
- Eisenlord, M.E.; Groner, M.L.; Yoshioka, R.M.; Elliott, J.; Maynard, J.; Fradkin, S.; Turner, M.; Pyne, K.; Rivlin, N.; van Hooidonk, R.; Harvell, C.D. 2016. Ochre star mortality during the 2014 wasting disease epizootic: role of population size structure and temperature. Philosophical Transactions Royal Society London Series B Biological Science 371, 20150212. [CrossRef]
- La Fauce, K.A.; Elliman, J.; Owens, L. 2007. Molecular characterisation of hepatopancreatic parvovirus (PmergDNV) from Australian Penaeus merguiensis. Virology 5, 397-403. [CrossRef]
- Tang, K.F.J.; Pantoja, C.; Lightner, D.V. 2008. Nucleotide sequence of a Madagascar hepatopancreatic parvovirus (HPV) and comparison of genetic variation among geographic isolates. Diseases of Aquatic Organisms 80, 105-112. [CrossRef]
- Jeeva, S.; Kang, S.W.; Lee, Y.S.; Jang, I.K.; Seo, H.C.; Choi, T.J. 2012. Complete nucleotide sequence analysis of a Korean strain of hepatopancreatic parvovirus (HPV) from Fenneropenaeus chinensis. Virus Genes 44, 89-97. [CrossRef]
- Safeena, M.P.; Rai, P.; Karunasagar, I. 2012. Molecular biology and epidemiology of Hepatopancreatic parvovirus of Penaeid Shrimp. Indian Journal of Virology 23, 191-202. [CrossRef]
- Gangnonngiw, W.; Kiatpathomchai, W.; Sriurairatana, S.; Laisutisan, K.; Chuchird, N.; Limsuwan, C.; Flegel, T.W. 2009. A parvo-like virus in the hepatopancreas of freshwater prawns Macrobrachium rosenbergii cultivated in Thailand. Dis. Aquat. Org. 85, 167-173. [CrossRef]
- Srisala, J.; Thaiue, D.; Sanguanrut, P.; Aldama-Cano, D.J.; Flegel, T.W.; Sritunyalucksana, K. 2021. Potential universal PCR method to detect decapod hepanhamaparvovirus (DHPV) in crustaceans. Aquaculture 541, 736782. [CrossRef]
- Anderson, I.G.; Law, A.T.; Shariff, M.; Nash, G. 1990. A parvo-like virus in the giant freshwater prawn, Macrobrachium rosenbergii. Journal of Invertebrate Pathology 55, 447-449. [CrossRef]
- Lightner, D.V.; Redman, R.M.; Poulos, B.T.; Mari, J.L.; Bonami, J.R.; Shariff, M. 1994. Distinction of HPV-type virus in Penaeus chinensis and Macrobrachium rosenbergii using a DNA probe. Asian Fisheries Science 7, 267-272. [CrossRef]
- Lee, C.-F.; Chang, Y.-C.; Chiou, H.-Y.; Chang, H.-W. 2022. Concurrent infection of a novel genotype of hepatopancreatic parvovirus and Enterocytozoon hepatopenaei in Penaeus vannamei in Taiwan. Journal of Fish Diseases, 45, 1201–1210. [CrossRef]
- Chong, Y.C.; Loh, H. 1984. Hepatopancreas chlamydial and parvoviral infections of farmed marine prawns in Singapore. Singapore Veterinary Journal (Singapore) 9, 51-56.
- Lightner, D.V. ; 1996. A Handbook of Pathology and Diagnostic Procedures for Diseases of Penaeid Shrimp. World Aquaculture Society, Baton Rouge, LA. https://lib.ugent. 0021. [Google Scholar]
- Thitamadee, S.; Prachumwat, A.; Srisala, J.; Jaroenlak, P.; Salachan, P.V.; Sritunyalucksana, K.; Flegel, T.W.; Itsathitphaisarn, O. 2016. Review of current disease threats for cultivated penaeid shrimp in Asia. Aquaculture 452, 69-87. [CrossRef]
- Singaravel, V.; Gopalakrishnan, A.; Martin, G.G. ; 2021. Multiple infections of Enterocytozoon hepatopenaei and Hepatopancreatic parvovirus in pond-reared Penaeus vannamei in India. Aquaculture 545, 737232. [CrossRef]
- Dhar, A.K.; Cruz-Flores, R.; Caro, L.F.A.; Siewiora, H.M.; Jory, D. 2019. Diversity of single-stranded DNA containing viruses in shrimp. Virus Dis. 30, 43-57. [CrossRef]
- Flegel, T.; Thamavit, V.; Pasharawipas, T.; Alday-Sanz, V. 1999. Statistical correlation between severity of hepatopancreatic parvovirus infection and stunting of farmed black tiger shrimp (Penaeus monodon). Aquaculture 174, 197-206. [CrossRef]
- Chayaburakul, K.; Nash, G.; Pratanpipat, P.; Sriurairatana, S.; Withyachumnarnkul, B. 2004. Multiple pathogens found in growth-retarded black tiger shrimp Penaeus monodon cultivated in Thailand. Diseases of Aquatic Organisms 60, 89-96. [CrossRef]
- Chantanachookin, C.; Boonyaratanapalin, S.; Kasornchandra, J.; Direkbusarakom, S.; Ekpanithanpong, U.; Supamataya, K.; Siurairatana, S.; Flegel, T.W. 1993. Histology and ultrastructure reveal a new granulosis-like virus in Penaeus monodon affected by ‘‘yellowhead’’ disease. Diseases of Aquatic Organisms 17, 145-157. [CrossRef]
- Sukhumsirichart, W.; Attasart, P.; Boonsaeng, V.; Panyim, S. 2006. Complete nucleotide sequence and genomic organization of hepatopancreatic parvovirus (HPV) of Penaeus monodon. Virology 346, 266-277. [CrossRef]
- Cervellione, F.; McGurk, C.; Berger Eriksen, T.; Van Den Broeck, W. 2017. Use of computer-assisted image analysis for semi-quantitative histology of the hepatopancreas in whiteleg shrimp Penaeus vannamei (Boone). Journal of Fish Diseases 40, 1223-1234. [CrossRef]
- Phromjai, J.; Boonsaeng, V.; Withyachumnarnkul, B.; Flegel, T.W. 2002. Detection of hepatopancreatic parvovirus in Thai shrimp Penaeus monodon by in situ hybridization, dot blot hybridization and PCR amplification. Diseases of Aquatic Organisms 51, 227-232. [CrossRef]
- Attasart, P.; Kaewkhaw, R.; Chimwai, C.; Kongphom, U.; Panyim, S. 2011. Clearance of Penaeus monodon densovirus in naturally pre-infected shrimp by combined ns1 and vp dsRNAs. Virus Res. 159, 79-82. [CrossRef]
- Dhar, A.K.; Robles-Sikisaka, R.; Saksmerprome, V.; Lakshman, D.K. 2014. Biology, genome organization, and evolution of parvoviruses in marine shrimp. Advances in Virus Research 89, 85-139. [CrossRef]
- Lightner, D.V.; Redman, R.M. 1998. Shrimp diseases and current diagnostic methods. Aquaculture 164,201-220. [CrossRef]
- Tang, K.F.J.; Poulos, B.T.; Wang, J.; Redman, R.M.; Shih, H.-H.; Lightner, D.V. 2003. Geographic variations among infectious hypodermal and hematopoietic necrosis virus (IHHNV) isolates and characteristics of their infection. Diseases of Aquatic Organisms 53:91-99. [CrossRef]
- Yu, J.Y.; Yang, N.; Hou, Z.H.; Wang, J.J.; Li, T.; Chang, L.R.; Fang, Y.; Yan, D.C. 2021. Research progress on hosts and carriers, prevalence, virulence of infectious hypodermal and hematopoietic necrosis virus (IHHNV). Journal of Invertebrate Pathology 183, 107556. [CrossRef]
- Tang, K.F.J.; Lightner, D.V. 2006. Infectious hypodermal and hematopoietic necrosis virus (IHHNV)-related sequences in the genome of the black tiger prawn Penaeus monodon from Africa and Australia. Virus Res 118:185–191. [CrossRef]
- Rai, P.; Pradeep, B.; Safeena, M.P.; Karunasagar, I.; Karunasagar, I. 2009. Simultaneous presence of infectious hypodermal and hematopoietic necrosis virus (IHHNV) and type A virus-related sequence in Penaeus monodon from India. Aquaculture 295, 168-174. [CrossRef]
- Saksmerprome, V.; Jitrakorn, S.; Chayaburakul, K.; Laiphrom, S.; Boonsua, K.; Flegel, T.W. 2011. Additional random, single to multiple genome fragments of Penaeus stylirostris densovirus in the giant tiger shrimp genome have implications for viral disease diagnosis. Virus Research 160, 180-190. [CrossRef]
- Rusaini, La Fauce, K.A.; Elliman, J.; Bowater, R.O.; Owens, L. 2013. Endogenous Brevidensovirus-like elements in Cherax quadricarinatus: Friend or foe? Fish & Shellfish Immunology 396-399, 136-145. [CrossRef]
- Alcivar-Warren, A.; Bao, W.; Tang, K.F. 2022. Endogenous viral elements (EVE) of <italic>Decapod penstylhamaparvovirus 1</italic> (Infectious hypodermal and hematopoietic necrosis virus, IHHNV) – Implications for shrimp diagnosis. Aquaculture 2022, February 28 - March 4, 2022, San Diego, California. Abstract available at this link: https://www.was.org/Meeting/Program/PaperDetail/158479. (Accessed 7 June 2022).
- Salcedo-Mejía, L.A.; Durán-Ramirez, Y.; Velazco-Peña, R.Z.; Pinto, J.A.; Rebaza-Caballero, A. 2021. Near-complete genome sequences of 12 Peruvian strains of infectious hypodermal and hematopoietic necrosis virus infecting the shrimp Penaeus vannamei. Microbiol Resour Announc 10:e00169-21. [CrossRef]
- CEFAS (Centre for Environment Fisheries and Aquaculture Science). 2020. Infectious hypodermal and hematopoietic necrosis virus. Available at: https://www.cefas.co.uk/international-database-on-aquatic-animal-diseases/disease-data/?id=39 (accessed 29 June 2022).
- Bajaña, L.; Betancourt, I.; Bayot, B. 2022. Complete coding genome sequence of infectious hypodermal and hematopoietic necrosis virus isolated from Penaeus (Litopenaeus) vannamei shrimp in Ecuador. Microbiology Resource Announcements 11, 4. [CrossRef]
- Bell, T.A.; Lightner, D.V. 1984. IHHN virus: infectivity and pathogenicity studies in Penaeus stylirostris and Penaeus vannamei. Aquaculture 38, 185-194. [CrossRef]
- Chai, C.; Liu, Y.; Xia, X.; Wang, H.; Pan, Y.; Yan, S.; Wang, Y. 2016. Prevalence and genomic analysis of infectious hypodermal and hematopoietic necrosis virus (IHHNV) in Litopenaeus vannamei shrimp farmed in Shanghai, China. Arch Virol 161:3189–3201. [CrossRef]
- Lightner, D.V.; Redman, R.M.; Poulos, B.T.; Nunan, L.M.; Mari, J.L.; Hasson, K.W. 1997. Risk of spread of penaeid shrimp viruses in the Americas by the international movement of live and frozen shrimp. Revue Scientifique et Technique 16, 146-160. [CrossRef]
- Hazreen-Nita, M.K.; Kua, B.C.; Bhassu, S.; Othman, R.Y. ; 2012. Detection and genetic profiling of infectious hypodermal and haematopoietic necrosis virus (IHHNV) infections in wild berried freshwater prawn, Macrobrachium rosenbergii collected for hatchery production. Mol. Biol. Rep. 39, 3785-3790. [CrossRef]
- Chen, B.K.; Dong, Z.; Liu, D.P.; Yan, Y.B.; Pang, N.Y.; Nian, Y.Y.; Yan, D.C. ; 2017. Infectious hypodermal and haematopoietic necrosis virus (IHHNV) infection in freshwater crayfish Procambarus clarkii. Aquaculture 477, 76-79. [CrossRef]
- Chen, B.K.; Dong, Z.; Pang, N.Y.; Nian, Y.Y.; Yan, D.C. ; 2018. A novel real-time PCR approach for detection of infectious hypodermal and haematopoietic necrosis virus (IHHNV) in the freshwater crayfish Procambarus clarkii. J. Invertebr. Pathol. 157, 100-103. [CrossRef]
- Lee, C.; Choi, S.-K.; Jeon, H.J.; Lee, S.H.; Kim, Y.K.; Park, S.; Han, S.-Y.; Bae, S.; Kim, J.H.; Han, J.E. 2021. Detection of infectious hypodermal and hematopoietic necrosis virus (IHHNV, Decapod Penstylhamaparvovirus 1) in commodity Red Claw crayfish (Cherax quadricarinatus) imported into South Korea. Journal of Marine Science and Engineering 9, 856. [CrossRef]
- Yang, B.; Song, X.L.; Huang, J.; Shi, C.Y.; Liu, L. 2007. Evidence of existence of infectious hypodermal and hematopoietic necrosis virus in penaeid shrimp cultured in China. Vet. Microbiol. 120, 63-70. [CrossRef]
- Cavalli, L.S.; Batista, C.R.; Nornberg, B.F.S.; Mayer, F.Q.; Seixas, F.K.; Romano, L.A.; Marins, L.F.; Abre, P.C. 2013. Natural occurrence of White spot syndrome virus and Infectious hypodermal and hematopoietic necrosis virus in Neohelice granulata crab. J. Invertebr. Pathol. 114, 86-88. [CrossRef]
- Macías-Rodríguez, N.A.; Manon-Ríos, N.; Romero-Romero, J.L.; Camacho-Beltrán, E.; Magallanes-Tapia, M.A.; Leyva-López, N.E.; Hernández-López, J.; Magallón-Barajas, F.J.; Perez-Enriquez, R.; Sánchez-González, S.; Méndez-Lozano, J. 2014. Prevalence of viral pathogens WSSV and IHHNV in wild organisms at the Pacific Coast of Mexico. J. Invertebr. Pathol. 116, 8-12. [CrossRef]
- Wei, Y.W.; Fan, D.D.; Chen, J. 2017. The mussel Mytilus edulis L. as an important reservoir of infectious hypodermal and hematopoietic necrosis virus (IHHNV). Aquaculture Research 48, 758-759. [CrossRef]
- Rai, P.; Safeena, M.P.; Krabsetsve, K.; La-Fauce, K.; Owens, L.; Karunasagar, I. ; 2012. Genomics, molecular epidemiology and diagnostics of infectious hypodermal and hematopoietic necrosis virus. Indian Journal of Virology 23, 203-214. [CrossRef]
- Sellars, M.J.; Cowley, J.A.; Musson, D.; Rao, M.; Menzies, M.L.; Coman, G.J.; Murphy, B.S. 2019. Reduced growth performance of black tiger shrimp (Penaeus monodon) infected with infectious hypodermal and hematopoietic necrosis virus. Aquaculture 499:160-166. [CrossRef]
- DAWE (Australian Government Department of Agriculture, Water and the Environment). 2020. Infection with infectious hypodermal and haematopoietic necrosis virus (IHHNV). Available at: https://www.agriculture.gov.au/sites/default/files/documents/infection-infectious-hypodermal-haematopoietic-necrosis-virus.pdf.
- Lightner, D.V. ; 2011. Virus diseases of farmed shrimp in the Western Hemisphere (the Americas): A review. Journal of Invertebrate Pathology 106, 110-130. [CrossRef]
- Jagadeesan, V.; Praveena, P.E.; Otta, S.K.; Jithendran, K.P. 2019. Classical runt deformity syndrome cases in farmed Penaeus vannamei along the east coast of India. Journal of Coastal Research, Special Issue No. 86, 107-111. [CrossRef]
- Zhang, C.; Yuan, J.F.; Shi, Z.L. ; 2007. Molecular epidemiological investigation of infectious hypodermal and hematopoietic necrosis virus and Taura syndrome virus in Penaeus vannamei cultured in China. Virologica Sinica 22, 380-388. [CrossRef]
- Nian, Y.-Y.; Chen, B.-K.; Wang, J.-J.; Zhong, W.-T.; Fang, Y.; Li, Z.; Zhang, Q.-S.; Yan, D.-C. 2020. Transcriptome analysis of Procambarus clarkii infected with infectious hypodermal and haematopoietic necrosis virus. Fish and Shellfish Immunology 98, 766-772. [CrossRef]
- Lee, D.; Yu, Y.-B.; Choi, J.-H.; Jo, A.-H.; Hong, S.-M.; Kang, J.-C.; Kim, J.-H. 2022. Viral shrimp diseases listed by the OIE: A review. Viruses 14, 585. [CrossRef]
- Bonnichon, V.; Lightner, D.V.; Bonami, J.R. 2006. Viral interference between infectious hypodermal and hematopoietic necrosis virus and white spot syndrome virus in Litopenaeus vannamei. Diseases of Aquatic Organisms 72, 179-184. [CrossRef]
- Molthathong, S.; Jitrakorn, S.; Joyjinda, Y.; Boonchird, C.; Witchayachamnarnkul, B.; Pongtippatee, P.; Flegel, T.; Saksmerprome, V. 2013. Persistence of Penaeus stylirostris densovirus delays mortality caused by white spot syndrome virus infection in black tiger shrimp (Penaeus monodon). BMC Veterinary Research 9, 33. [CrossRef]
- Robles-Sikisaka, R.; Bohonak, A.J.; McClenaghan, L.R., Jr.; Dhar, A.K. 2010. Genetic signature of rapid IHHNV (Infectious hypodermal and hematopoietic necrosis virus) expansion in wild penaeus shrimp populations. PLoS ONE 5, e11799. [CrossRef]
- Tang, K.F.; Navarro, S.A.; Lightner, D.V. 2007. PCR assay for discriminating between infectious hypodermal and hematopoietic necrosis virus (IHHNV) and virus-related sequences in the genome of Penaeus monodon. Diseases of Aquatic Organisms 74, 165-170. [CrossRef]
- Dhar, A.K.; Roux, M.M.; Klimpel, K.R. 2001. Detection and quantification of infectious hypodermal and hematopoietic necrosis virus and white spot virus in shrimp using real-time quantitative PCR and SYBR Green chemistry. Journal of Clinical Microbiology 39, 2835-2845. [CrossRef]
- Silva, D.C.D.; Nunes, A.R.D.; Teixeira, D.I.A.; Lima, J.P.M.S.; Lanza, D.C.F. 2014. Infectious hypodermal and hematopoietic necrosis virus from Brazil: sequencing, comparative analysis and PCR detection. Virus Research 189, 136-146. [CrossRef]
- Bower, S.M. 1996. Synopsis of infectious diseases and parasites of commercially exploited shellfish: Infectious hypodermal and haematopoietic necrosis virus (IHHNV) of penaeid shrimp. Available at: https://www.dfo-mpo.gc.ca/science/aah-saa/diseases-maladies/ihnvsp-eng.html (accessed 29 June 2022).
- Dong, H.T.; Sangpo, P.; Dien, L.T.; Mai, T.T.; Linh, N.V.; Del-Pozo, J.; Salin, K.R.; Senapin, S. 2022. Usefulness of the pancreas as a prime target for histopathological diagnosis of Tilapia parvovirus (TiPV) infection in Nile tilapia, Oreochromis niloticus. Journal of Fish Diseases, 45, 1323-1331. [CrossRef]
- Rajendran, K.V.; Sood, N.; Rao, B.M.; Valsalam, A.; Bedekar, M.K.; Jeena, K.; Pradhan, P.K.; Paria, A.; Swaminathan, T.R.; Verma, D.K.; Sood, N.K. 2023. Widespread occurrence of Tilapia parvovirus in farmed Nile tilapia Oreochromis niloticus from India. Journal of Fish Diseases, 00, 1–11. [CrossRef]
- Hargitai, R.; Pankovics, P.; Boros, Á.; Mátics, R.; Altan, E.; Delwart, E.; Reuter, G. 2021. Novel picornavirus (family Picornaviridae) from freshwater fishes (Perca fluviatilis, Sander lucioperca and Ameiurus melas) in Hungary. Archives of Virology 166, 2627-2632. [CrossRef]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. 2018. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol 4, vey016. [CrossRef]
- Fraser, C.A.; Owens, L. 1996. Spawner-isolated mortality virus from Australia Penaeus monodon. Diseases of Aquatic Organisms 27, 141-148. [CrossRef]
- Owens, L.; Haqshenas, G.; McElnea, C.; Coelen, R. 1998. Putative spawner-isolated mortality virus associated with mid-crop mortality syndrome in farmed Penaeus monodon from northern Australia. Diseases of Aquatic Organisms 34, 177-185. [CrossRef]
- Owens, L.; McElnea, C. 2000. Natural infection of the redclaw crayfish Cherax quadricarinatus with presumptive spawner-isolated mortality virus. Diseases of Aquatic Organisms 40, 219-223. [CrossRef]
Figure 1.
Parvoviridae. Schematic representation of a parvovirus particle showing the non-enveloped, round, T=1 icosahedral symmetry, 22-28 nm in diameter. The capsid consists of 60 copies of CP protein. (Reproduced from [19]. Source: SwissBioPics. The images are licensed under a Creative Commons Attribution 4.0 International (CC BY 4.0) License https://creativecommons.org/licenses/by/4.0/).
Figure 1.
Parvoviridae. Schematic representation of a parvovirus particle showing the non-enveloped, round, T=1 icosahedral symmetry, 22-28 nm in diameter. The capsid consists of 60 copies of CP protein. (Reproduced from [19]. Source: SwissBioPics. The images are licensed under a Creative Commons Attribution 4.0 International (CC BY 4.0) License https://creativecommons.org/licenses/by/4.0/).
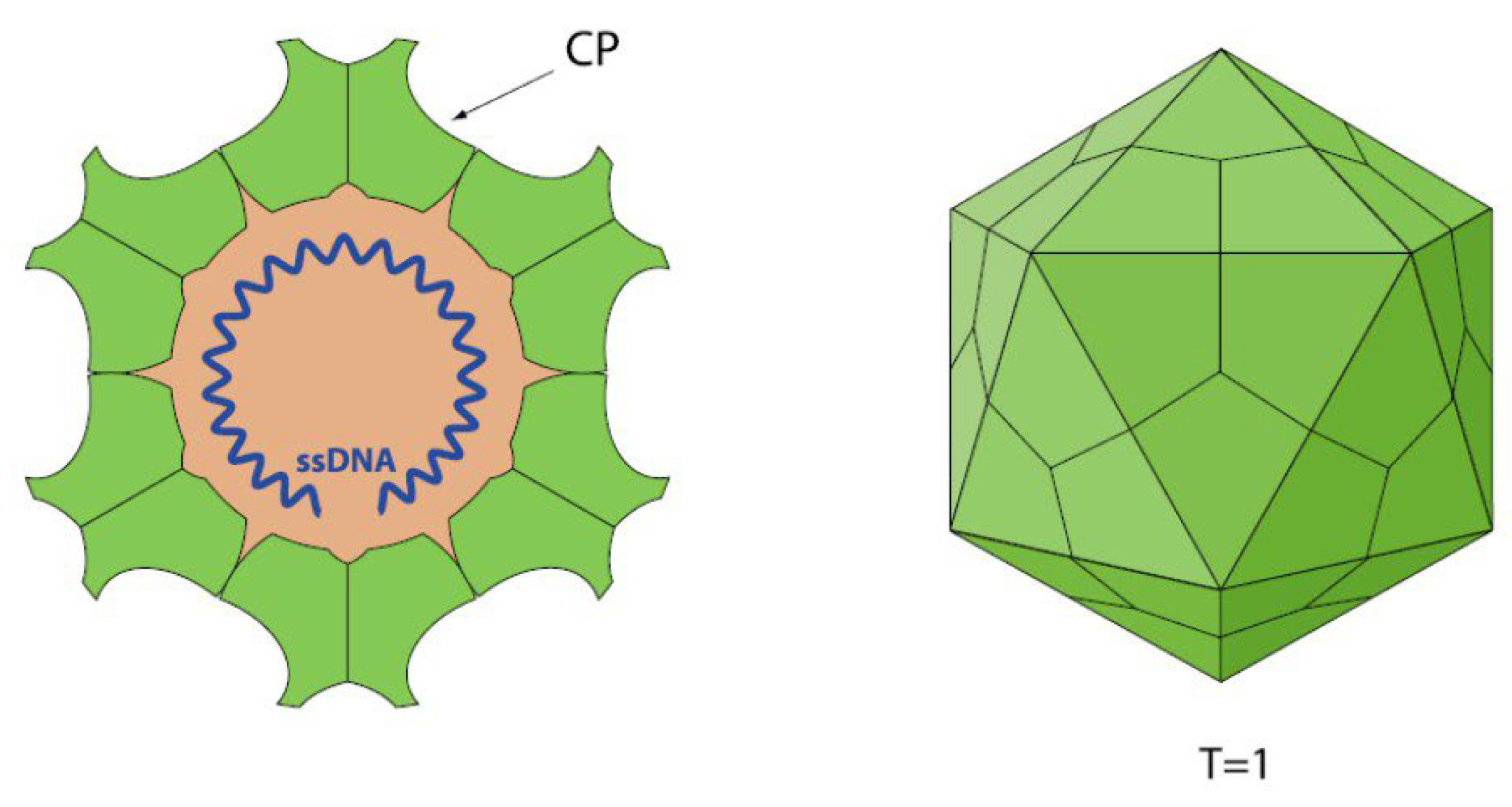
Figure 2.
Cladogram of subfamilies Hamaparvovirinae, Parvovirinae, and Densovirinae. Thegenome organizations of members of the representative genera of the three subfamilies are shown. The novel tilapia parvovirus HMU-HKU-1 discovered in this study was labeled with a red circle (●). The genera and subfamilies described in the newly proposed ICTV parvovirus taxonomic classification were highlighted in red. (Reproduced under Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/) from [18], Figure 2).
Figure 2.
Cladogram of subfamilies Hamaparvovirinae, Parvovirinae, and Densovirinae. Thegenome organizations of members of the representative genera of the three subfamilies are shown. The novel tilapia parvovirus HMU-HKU-1 discovered in this study was labeled with a red circle (●). The genera and subfamilies described in the newly proposed ICTV parvovirus taxonomic classification were highlighted in red. (Reproduced under Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/) from [18], Figure 2).
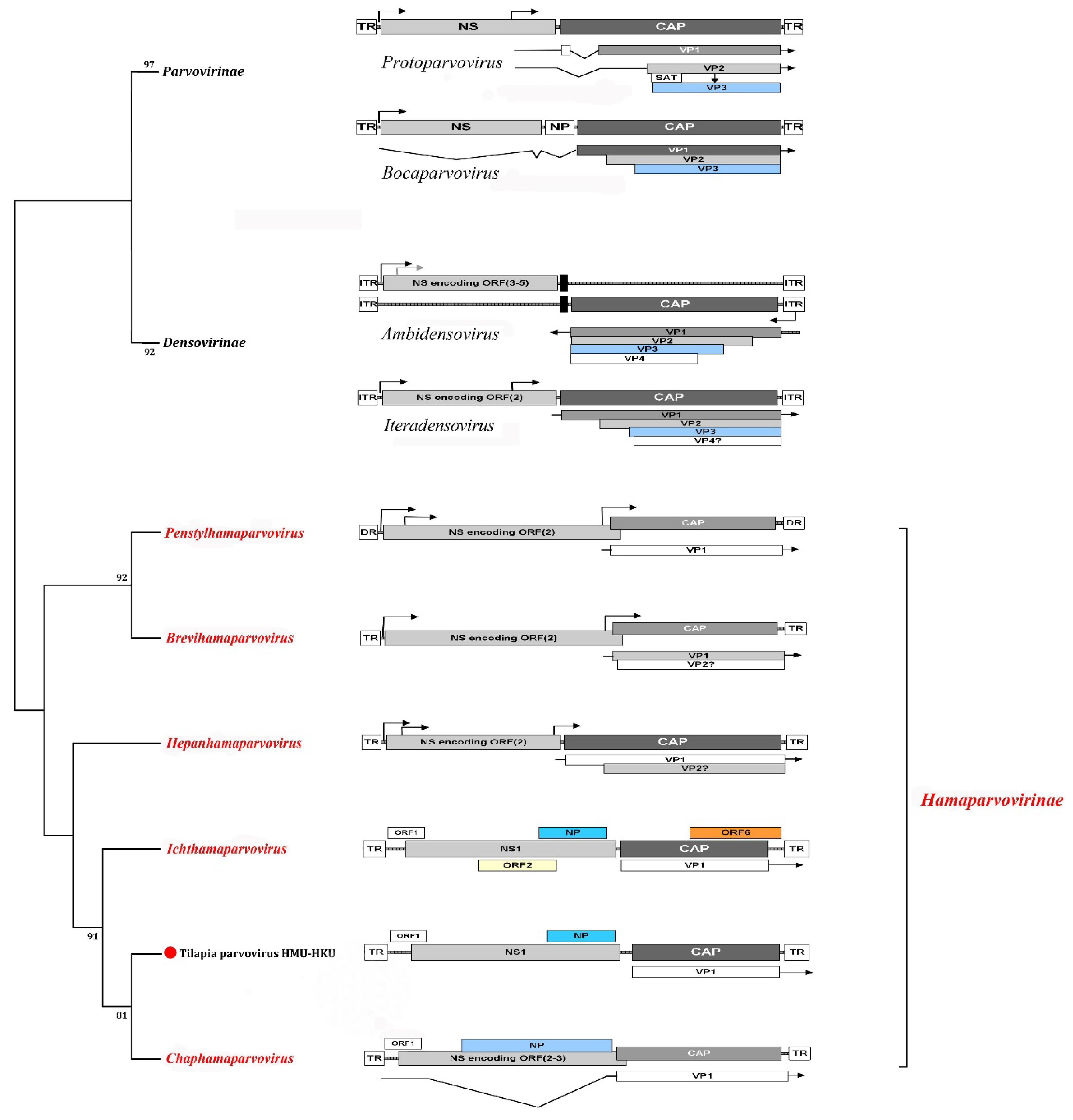
Figure 3.
Summary of parvovirus replication requirements. (1) Most autonomous parvoviruses require mitotically active cells (S/G2 phase) to provide host replication factors to replicate their viral genome. (2) Recently, human bocavirus 1 (HBoV1) was demonstrated to replicate in non-dividing airway epithelial cells through the hijacking of DNA repair machinery [27,28]. (3) Dependoparvoviruses depend on co-infection with a helper virus to undergo productive replication in a host cell. (Reproduced under Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/ ) from [23], Figure 4).
Figure 3.
Summary of parvovirus replication requirements. (1) Most autonomous parvoviruses require mitotically active cells (S/G2 phase) to provide host replication factors to replicate their viral genome. (2) Recently, human bocavirus 1 (HBoV1) was demonstrated to replicate in non-dividing airway epithelial cells through the hijacking of DNA repair machinery [27,28]. (3) Dependoparvoviruses depend on co-infection with a helper virus to undergo productive replication in a host cell. (Reproduced under Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/ ) from [23], Figure 4).
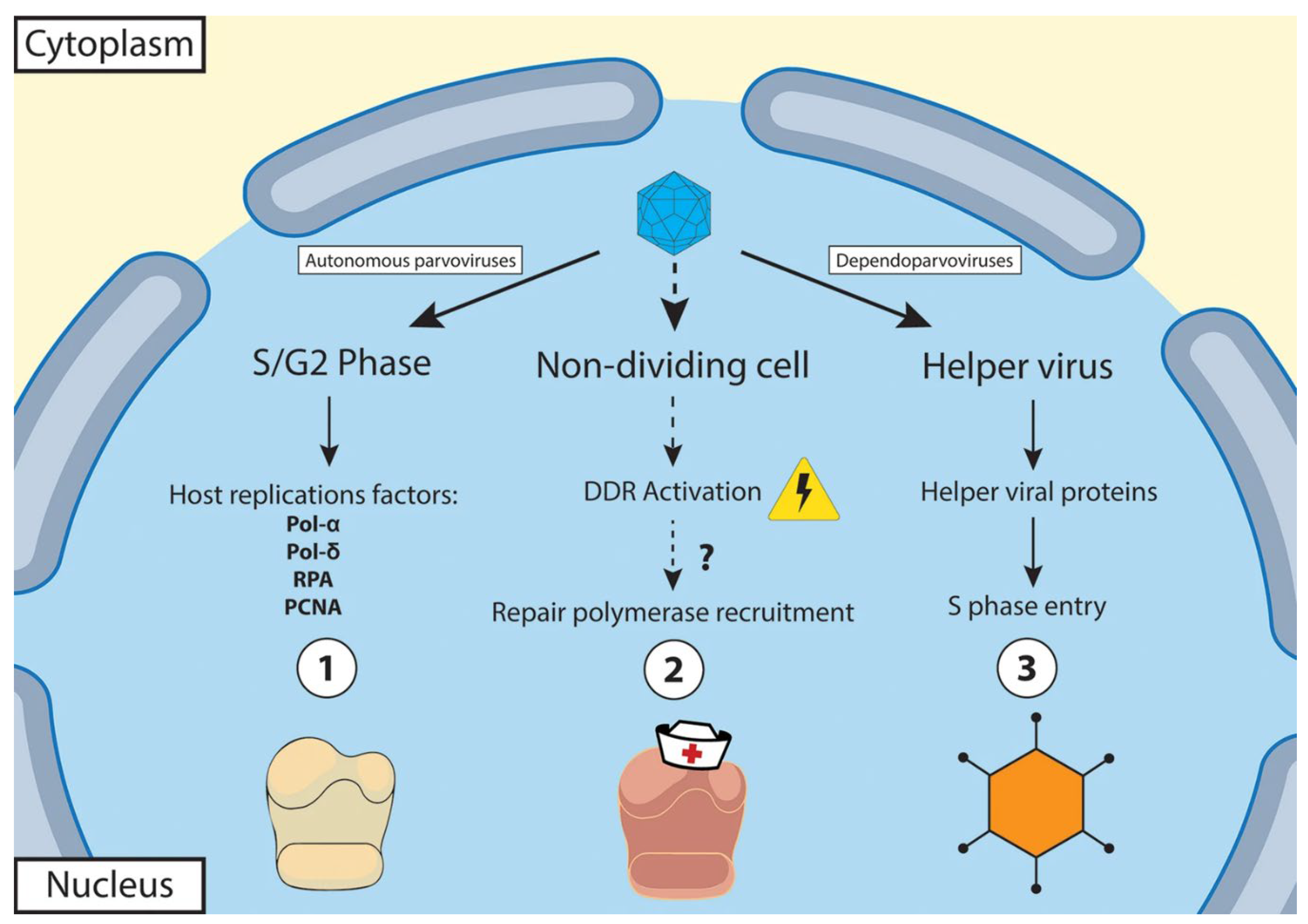
Figure 4.
Size variations observed in 50-day-old Penaeus monodon with infectious hypodermal and hematopoietic necrosis virus (IHHNV) (A,B) (Reprinted from Aquaculture, Vol. 289 (3-4), Rai, P.; Size variations observed in 50-day-old Penaeus monodon with infectious hypodermal and hematopoietic necrosis virus (IHHNV) (A,B). (Reproduced under Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/) from [101], Figure 10).
Figure 4.
Size variations observed in 50-day-old Penaeus monodon with infectious hypodermal and hematopoietic necrosis virus (IHHNV) (A,B) (Reprinted from Aquaculture, Vol. 289 (3-4), Rai, P.; Size variations observed in 50-day-old Penaeus monodon with infectious hypodermal and hematopoietic necrosis virus (IHHNV) (A,B). (Reproduced under Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/) from [101], Figure 10).

Figure 5.
Outbreaks, clinical symptoms, and pathological analysis. (A) Healthy tilapia; (B) Tilapia disease outbreak in cage-cultured results in massive mortality (August 2015; Jinmen, Hubei province, China); (C, D) Gross pathological signs of infected tilapia, including hemorrhages on the lower jaw, anterior abdominal, and the fin bases, accompanying with exophthalmos eyes and pronounced ocular lesions. (E) Healthy spleen; (F) Moderated diseased spleen infected at 3rd days after post TiPV-infection; (G, H) Severe diseased spleen infected at 5th days after post TiPV-infection. Lymphocytes (white arrow) and macrophages (white triangular arrowheads) in diseased spleen sinusoids, necrotic splenocytes (red arrow), virus inclusion body (black arrow), and melanomacrophage centers (asterisk) in affected spleen; (I) Healthy brain; (J) Moderated diseased brain infected at 3rd days after post TiPV infection, vacuolated neurons with marginated nuclear (red arrow); (K, L) Severe diseased brain infected at 5th days after post TiPV infection, vacuolated neurons with marginated nuclear (red arrow), lymphocytes (white arrow) in the blood vessel, edema of cerebral cortex (black arrow). HE staining. Bar = 20um (A, B, E, F), 50um (D), 5 um(C). (Reproduced under Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/) from [15], Figure 1).
Figure 5.
Outbreaks, clinical symptoms, and pathological analysis. (A) Healthy tilapia; (B) Tilapia disease outbreak in cage-cultured results in massive mortality (August 2015; Jinmen, Hubei province, China); (C, D) Gross pathological signs of infected tilapia, including hemorrhages on the lower jaw, anterior abdominal, and the fin bases, accompanying with exophthalmos eyes and pronounced ocular lesions. (E) Healthy spleen; (F) Moderated diseased spleen infected at 3rd days after post TiPV-infection; (G, H) Severe diseased spleen infected at 5th days after post TiPV-infection. Lymphocytes (white arrow) and macrophages (white triangular arrowheads) in diseased spleen sinusoids, necrotic splenocytes (red arrow), virus inclusion body (black arrow), and melanomacrophage centers (asterisk) in affected spleen; (I) Healthy brain; (J) Moderated diseased brain infected at 3rd days after post TiPV infection, vacuolated neurons with marginated nuclear (red arrow); (K, L) Severe diseased brain infected at 5th days after post TiPV infection, vacuolated neurons with marginated nuclear (red arrow), lymphocytes (white arrow) in the blood vessel, edema of cerebral cortex (black arrow). HE staining. Bar = 20um (A, B, E, F), 50um (D), 5 um(C). (Reproduced under Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/) from [15], Figure 1).
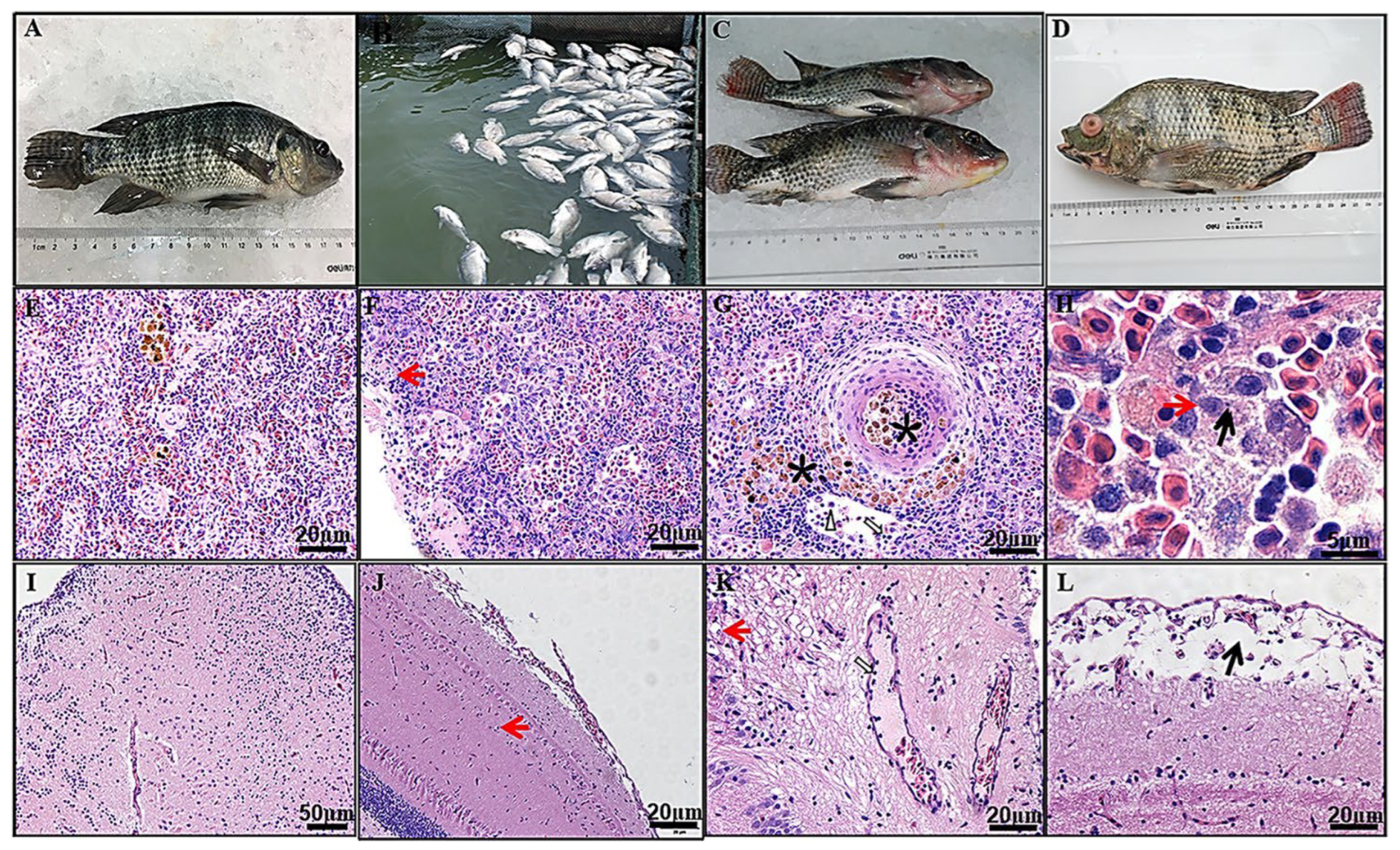
Figure 6.
Morphology of the Tilapia brain cells (TiB) and cytopathic effect (CPE) induction on TiB induced by TiPV and transmission electron micrographs of the TiPV-infected TiB cells. (A) The TiB cells at passage 1, 10 days; (B) The TiB cells at passage 2, 3 days; (C) TiB cells infected with TiPV at passage 3 at 5 days post infection; (D) TiB cells infected with TiPV at passage 6 at 3 days post infection (Bar = 100 μm). (E) Virus particles existed in the cytoplasm and nuclei (white arrow), Nu: nucleus. (Bar, 1 μm); (F) High magnification of the region in the white rectangular box indicated in Panel A, virus particles aggregated in the cytoplasm (Bar, 200 nm); (G) The virus releasing at the plasma membrane of the TiB cell. (Bar, 200 nm); (H) Purified TiPV particles negatively stained with 2% phosphotungstic acid (Bar, 200nm). (Reproduced under Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/) from [15], Figure 2).
Figure 6.
Morphology of the Tilapia brain cells (TiB) and cytopathic effect (CPE) induction on TiB induced by TiPV and transmission electron micrographs of the TiPV-infected TiB cells. (A) The TiB cells at passage 1, 10 days; (B) The TiB cells at passage 2, 3 days; (C) TiB cells infected with TiPV at passage 3 at 5 days post infection; (D) TiB cells infected with TiPV at passage 6 at 3 days post infection (Bar = 100 μm). (E) Virus particles existed in the cytoplasm and nuclei (white arrow), Nu: nucleus. (Bar, 1 μm); (F) High magnification of the region in the white rectangular box indicated in Panel A, virus particles aggregated in the cytoplasm (Bar, 200 nm); (G) The virus releasing at the plasma membrane of the TiB cell. (Bar, 200 nm); (H) Purified TiPV particles negatively stained with 2% phosphotungstic acid (Bar, 200nm). (Reproduced under Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/) from [15], Figure 2).
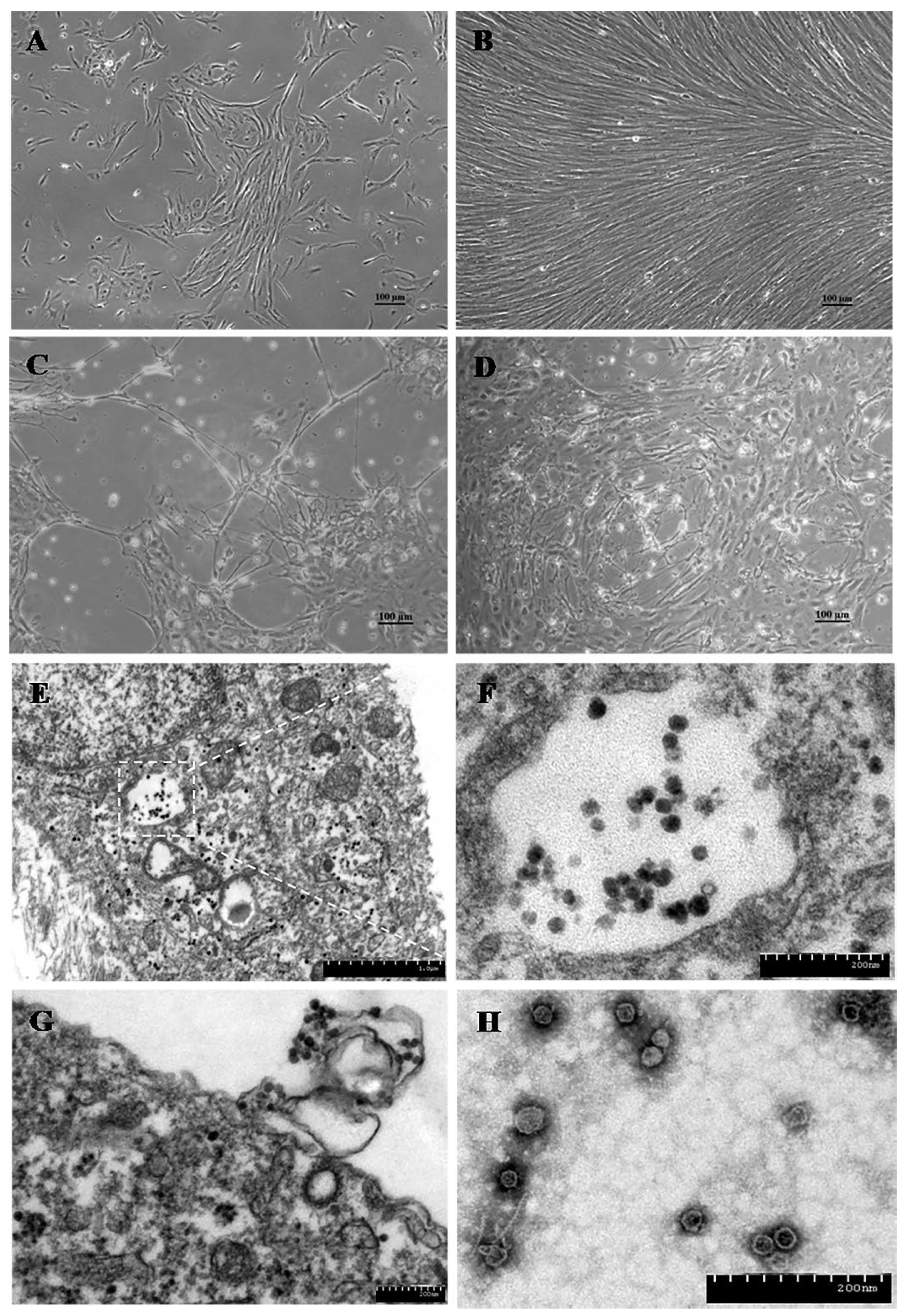
Figure 7.
Phylogenetic analysis of zander/M5/2015/HUN (OK236393, bold letters) and representatives of 10 genera in the subfamily Parvovirinae based on the ~460-aa-long tripartite helicase domain of NS1. The dendrogram was constructed based on an amino acid sequence alignment of tripartite helicase domains by Bayesian Evolutionary Analysis Utility version v1.10.4 (BEAST) [112] using the LG+I+G+F substitution model, a lognormal relaxed clock, and Youle process, throughout 10,000,000 generations. The tree is drawn to scale with branch lengths measured in the average number of substitutions per time unit. PV = parvovirus. (Reproduced under Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/) from [5], Figure 2).
Figure 7.
Phylogenetic analysis of zander/M5/2015/HUN (OK236393, bold letters) and representatives of 10 genera in the subfamily Parvovirinae based on the ~460-aa-long tripartite helicase domain of NS1. The dendrogram was constructed based on an amino acid sequence alignment of tripartite helicase domains by Bayesian Evolutionary Analysis Utility version v1.10.4 (BEAST) [112] using the LG+I+G+F substitution model, a lognormal relaxed clock, and Youle process, throughout 10,000,000 generations. The tree is drawn to scale with branch lengths measured in the average number of substitutions per time unit. PV = parvovirus. (Reproduced under Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/) from [5], Figure 2).
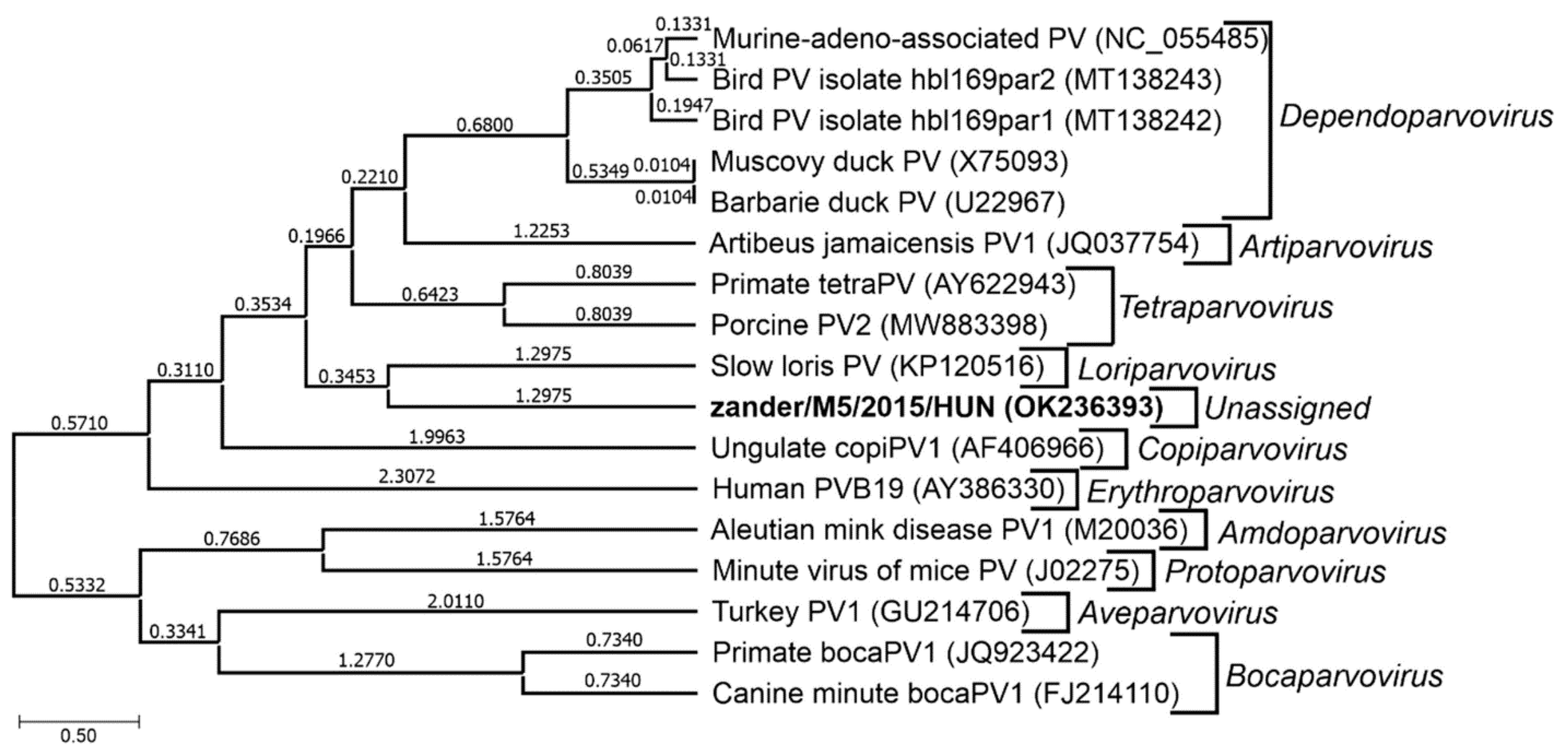

Table 1.
Summary of parvoviruses found in aquatic animals.
| Subfamily and Genus | Species1 | Virus common name (abbreviation) | Sampled host | Geographical area | Clinical disease | Reference |
| Densovirinae, Aquambidensovirus | Decapod aquambidensovirus 1 | Cherax quadricarinatus densovirus [previously known as Cherax quadricarinatus parvo-like virus (CqPlV)]; Cherax destructor systemic parvo-like virus (CdSPV) | redclaw crayfish Cherax quadricarinatus and Cherax destructor | Australia | Chronic mortality or mass mortality | [7,8] |
| Asteroid aquambidensovirus 1 | Sea star-associated densovirus (SSaDV) | sea stars and sea urchins | West coast of USA and Atlantic coast of North America | Originally associated with sea star wasting syndrome (SSWS) | [9,29] | |
| Clinch densovirus 1 | Freshwater mussels | Clinch River, Virginia, and Tennessee, USA | Episodic mass mortality | [10] | ||
| Hamaparvovirinae, Hepanhamaparvovirus | Decapod hepanhamaparvovirus 1 | Fenneropenaeus chinensis hepatopancreatic densovirus [previously known as Hepatopancreatic parvovirus (HPV)] | Prawns & Shrimp | Widespread | Reduced growth in juvenile shrimp; Mortalities during the larval stages. | [30-32] |
| Hamaparvovirinae, Ichtahamaparvovirus | Syngnathid ichthamaparvovirus 1 | Syngnathus scovelli chapparvovirus |
gulf pipefish | Gulf of Mexico and parts of South America | Not known | [33] |
| Tilapia parvovirus (TiPV) | Nile tilapia | China and Thailand | Mass morbidity and 60-70% mortality in adult Nile tilapia (500-600 g); mass mortality of 50-75% in juvenile red hybrid tilapia (10-30 g to 300-800 g). | [15-17] | ||
| unclassified Ichthamaparvovirus | ||||||
| Hamaparvovirinae, Penstylhamaparvovirus | Decapod penstylhamaparvovirus 1 | Penaeus stylirostris penstyldensovirus (PstDV 1&2); Penaeus monodon penstyldensovirus (PmoPDV 1&2) [previously known as Infectious hypodermal & hematopoietic necrosis virus (IHHNV)] | Prawns & Shrimp | Widespread | IHHN; 80-100% mortality in postlarvae and juveniles of Penaeus (Litopenaeus) stylirostris and postlarvae of Macrobrachium rosenbergii; runt-deformity syndrome (RDS) in juvenile of P. vannamei and P. monodon |
[34-36] |
| Unassigned | Novel salmon parvovirus | Sockeye salmon | BC-Canada | Not known | [37] | |
| Unassigned in subfamily Parvovirinae | Novel zander parvovirus (zander/M5/2015/HUN, OK236393) | Zander or pikeperch (Sander lucioperca) | Hungary | Not known | [5] | |
| Unassigned | Crangon crangon parvo-like virus 1 (CcPaLV 1) | Molluscs | Europe | Not known | [38] | |
| Unassigned | Spawner-isolated mortality virus (SMV) | Freshwater crayfish | Northern Australia | Mortalities in broodstock of Penaeus monodon with mid-crop mortality syndrome on grow-out farms | [39] |
1Species as listed in the NCBI Taxonomy Browser [40].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



