
Submitted:
09 June 2024
Posted:
13 June 2024
You are already at the latest version
Abstract
A novel means of applying radiotherapy in cancer treatment is the application of the radiation dose at a very high intensity for a very short time, FLASH radiotherapy (FLASH-RT). This technique involves exposure of tumors to >40 Gy/s, usually for less than one second. Studies conducted in cell and preclinical models suggest that FLASH-RT seems less damaging to normal tissues from adverse effects, relative to the same overall dose of radiation administered in conventional therapy (CONV-RT) which involves administration of lower levels of radiation repeated intermittently over a protracted period. In contrast, the susceptibility of tumor tissues to FLASH-RT is not diminished relative to CONV-RT. Within solid tumors both modes of dispensation of radiation produce an equivalent degree of cell damage. This differential between normal and malignant material has been found in isolated tissues, animal studies, and more recently in clinical trials. However, the classical radiation concept is that high energy linear transfer radiation (LET) is more damaging than the equivalent total dose of low LET. Thus, the susceptibility of cells should be greater to short-term exposure to high LET. This article discusses the potential reasons that may account for this discrepancy. While the relative protection given to untransformed tissues by FLASH-RT relative to tumor tissue is a major step forward in radiation therapy of cancer, the processes that lie behind this phenomenon are incompletely understood and are considered here.
Keywords:
Radiotherapy
; tumors
; FLASH radiation
; dosage
; frequency
1. Introduction
Radiotherapy has long been a major source of treatment for cancer, as a means of killing malignant cells. Over 50% of cancer patients currently receive such therapy [1]. Radiotherapy is generally used in conjunction with surgical procedures to minimize tumor burden, and chemotherapeutic agents also as a means of eradicating cancer cells. Many refinements in the design of this procedure have been made, always with the intent of minimizing harm to surrounding normal tissues while maintaining an intense destructive focus on to tumor tissue. A recent means of addressing this problem has been the development of FLASH-RT. The basic concept is the use of a very short pulse of high radiation, (around 40-100 Gy/sec) roughly 400-fold more intense than the lower level, longer exposures of radiation used in classical radiotherapy. The origins of this procedure have their roots in a 1959 paper reporting that a short burst of high levels radiation was less lethal to bacteria than the same dose administered over a longer time span [2]. The reduced toxicity caused by this ultra-high-dose short burst of radiation in comparison with a more extended lower dose rate, has been replicated in mammalian cell culture and then in intact animals [3,4]. In several of these reports, the relative sparing effect of FLASH-RT is described as being lost at dose rates below around 5-10Gy/sec [5,6].
A second unexpected discovery was that there was a divergence between the susceptibility of tumor cells to FLASH-RT relative to that of untransformed normal cells. The reduced lethality of FLASH-RT to normal cells was not replicated in tumor tissue. This is in sharp contrast to conventional radiotherapy where the vulnerability of cells, while being sensitive to the rate of mitosis, does not otherwise differ between normal and malignant cells.
Damage caused by conventional radiation has been thought of as being largely due to reactive oxygen species. A possible limitation has been that the hypoxic state of most tumor tissues may make them less susceptible to oxidative injury than normal cells [7]. In 2019, the first successful clinical application of FLASH-RT led to complete eradication of a multiresistant cutaneous lymphoma [8]. More recently, a range of studies on the effects of FLASH-RT have ensured that this strategy will have great impact on the future radiotherapeutic treatment of solid tumors. Several groups have reported the utility of this treatment in animal patients by veterinarians [3,9,10].
2. Accounting for the Differences between FLASH-RT and CONV-RT
There are two major unexplained issues concerning the selective advantage of FLASH-RT over CONV-RT that have not yet been fully explained. These are interrelated and overlap in some measure, but are conveniently discussed as separate problems.
2.1. Why Is FLASH-RT Less Lethal to Normal Cells Than the Same Dose of CONV-RT?
A given dose of radiation a higher dose-rate should be more toxic than the equivalent dosage applied for a longer period of time. This been an orthodox and well established view in radiation biology. and has led to the foundation of radiotherapy generally involving relatively low doses of radiation, applied at intervals. Exposure to extreme levels of gamma radiation over millisecond periods certainly seems especially lethal as judged by the effects of atomic bombs on Hiroshima and Nagasaki. In fact, FLASH has been described as a good model to simulate short duration high intensity radiation following detonating of a nuclear weapon [11].
2.1.1. Anoxia
However, there are now many reports of FLASH-RT being less damaging to normal cells than the equivalent dose of CONV-RT [12]. The mechanism underlying this is unclear but the dramatically sudden reduced oxygen tension resulting from FLASH-RT has been proposed to play a major part of the overall effect. It has long been known that radiation can lead to an anoxia-like state of target tissues [13], and that this becomes especially pronounced after high intensity radiation [14]. It has been posited that the rapid consumption of oxygen caused by FLASH-RT leads to transient hypoxia and this reduces the extent of production of harmful reactive oxygen species caused by the radiation relative to CONV-RT. Probably related to this, is the fact that FLASH does not induce neuroinflammation in the brain [15]. This is mediated by the lesser cell damage and lower induction of inflammatory cytokines provoked by FLASH [16]. Also, in the context of cerebral tissue, FLASH-RT has been reported as permitting maintenance of synaptic plasticity [17] and sparing of indices of behavioral and cognitive functioning [15]. Further evidence of the key role of sudden and intense anoxia in FLASH-RT is the finding that Induction of cerebral hyperoxia by means of carbogen inhalation prevented the neuroprotective effects of FLASH [15]. A clinical implication of this is that the higher levels of oxygen used during anesthesia can negate the protective consequences of FLASH-RT [18].
2.1.2. The Role of Reactive Oxygen Species
The sparing effect of FLASH has a biphasic distribution with respect to oxygen concentration. It is maximal at around at around 4% pO2 and falls off at oxygen concentrations above or below this [19]. A very low presence of oxygen in tissues can be insufficient to allow the formation of reactive oxygen species under either type of application of radiation [20].
As illustrated by oxidative injury following ischemia-reperfusion, rapid re-oxygenation of anoxic cells can lead to formation of reactive oxygen species. Such an oxygen rebound event has been shown to occur after the sudden anoxia incurred after FLASH [21], and can be a source of the formation active oxidants existing momentarily. It may be that the different responses to FLASH and CONV radiation are related to the very transient high levels of free radicals caused by FLASH resulting in rapid detoxifying radical-radical interactions between peroxyl free radicals. This would lead to reduction of the overall presence of free radicals and could limit damage, including that incurred by migration of oxidant species to other sites [22]. This idea is supported by physicochemical modeling of reaction kinetics suggesting that peroxyl radical recombination is a major factor in enabling FLASH-RT protection [23]. As distances between charged particle trajectories are much closer with FLASH, detoxifying interactions between formed reactive oxidizing species such as peroxy radicals become more likely [24,25]. The fraction of damage due to secondary events is then likely to be reduced in FLASH-RT.
In contrast, CONV-RT may lead to a more prolonged presence of harmful short-lived reactive oxygen. The relatively diluted form of free radicals and their precursors generated in this manner, can prolong their existence and lead to diffusion of oxidizing species.
2.1.3. Damage to DNA
DNA can be harmed by radiation causing both double stranded or single stranded breaks in the DNA double helix. This can occur either by radiation energy directly impacting DNA, or by DNA being indirectly attacked by highly reactive and short lived species formed by reactive electrons in proximity to the DNA. In aqueous media, low energy radiation contributes more to indirect DNA damage than do high energy X-rays [26]. At very high doses and dose rates of radiation, where oxygen depletion in the cellular environment plays a major role, there may be a consequent decline in DNA damage due to a lesser ability to form reactive oxygen species [27]. This may be due to a saturation effect caused by the close proximity of the penetrant beams in FLASH [28].
In isolated plasmids, almost all DNA damage seems indirect since no potential scavenging molecules are present. In such defined isolated cell free media, FLASH reduced the extent of single stranded breaks (but not double-stranded breaks) in plasmid DNA in comparison to CONV [29,30]. The suspension of plasmids in a protein-free medium, containing only Tris buffer may account for the attenuation of the differences between conventional and FLASH radiation.
Non-lethal radiation damage to DNA, can lower overall cellular effectiveness and lead to emergence of cells of a senescence-associated secretory phenotype (SASP). These cells secrete inflammatory cytokines which can also further spread inflammatory events in nearby cells [31].
2.1.4. Inflammation
A major component of the lessening of overall radiation damage caused FLASH, may be attributable to reduced inflammation. FLASH-RT is far less toxic to circulating immune cells than is CONV-RT [6]. The evidence of FLASH-RT causing less inflammation seems to coincide with elevated alertness of T-cells which can inhibit tumor progression [32] and is likely due to a lower presence of reactive oxygen. In many instances the inhibition of non-productive and unfocused inflammation has been observed to bring about improved immune function [33].
Inflammation represents a response to altered oxidant conditions and to modifications of signaling pathways, and thus generally reflects a secondary consequence of metabolic change. In the case of FLASH, the shortened persistence of ROS production will also lead to reduction of the pathways of activation of the inflammatory response. This may account for the reduced inflammation and lower DNA damage found following FLASH [28].
There is evidence that the differential effect of FLASH involves its ability to maintain integrity of mitochondrial function such as mitochondrial membrane potential. In contrast standard radiation protocols lead to severe adverse morphological and functional changes in mitochondria [34,35]. Significantly, in this study, cell death following CONV-RT was largely by way of necrosis which leads to inflammation, while FLASH-RT led to apoptosis. Since necrosis leads to inflammatory events while apoptosis is a more organized means of dismantling cells, this difference could account for the lesser inflammation found with the FLASH procedure. The ability of FLASH-RT to induce regulated immune reactions rather than provoking uncontrolled inflammation has long term advantages. Thus, treatment of lung tissue of rodents with spatially focused microbeam irradiation at FLASH levels does not lead to radiation fibrosis even after one year [36].
FLASH-RT reduced the number of lung-derived fibroblasts that were converted into cells expressing indices of senescence relative to CONV. This is likely to underlie the prevention of radiation-generated fibrosis in lung by FLASH-RT [37]. Radiation-induced senescence can also be transferred to cells at remote sites perhaps by a bystander mechanism involving the circulation of exosomes [38]. Since the damage to DNA effected by FLASH treatment is less pronounced, this can reduce the formation of SASP-type cells [39]. These findings suggest that FLASH-RT is less potent in facilitating the premature aging events that have been ascribed to radiotherapy.
2.2. Why Are Tumor Cells More Sensitive Than Normal Cells to FLASH-RT?
Unlike normal tissues, tumor cells appear to be just as sensitive to FLASH-RT than to CONV-RT. This creates a beneficial disparity between the responses of the two cell types. Since higher dose rates of radiation are generally described producing greater damage to cells, FLASH-RT should cause a better tumor reaction than CONV-RT and this is indeed the case. The reasons underlying this divergence remain unresolved but several separate hypotheses are substantiated by credible evidence.
2.2.1. Tumor Metabolism is Unlike That of Normal Tissue
The excess sensitivity of tumor cells to FLASH-RT has some enigmatic aspects, as tumor tissues are often hypoxic and can live under conditions under which normal cells could not survive [4], yet the FLASH effect seems to involve transient but intense anoxia. Most solid tumors have a preferential utilization of anaerobic glycolysis as opposed to the emphasis on aerobic metabolism characteristic of normal cells. The less efficient energy metabolism of tumor cells is compensated for by enhanced glucose consumption [41,42]. This lack of dependence on high levels of oxygen accounts for the frequently encountered resistance to CONV-RT. In fact, tumor growth is attenuated by hyperoxia. However, tumor cells have a higher intrinsic pro-oxidant environment than normal cells and are more consistently subject to oxidative stress [43]. They are therefore more susceptible to free-radical induced death than are normal cells [44]. While hypoxia induces resistance of normal cells to FLASH-RT [45], tumor cells remain selectively vulnerable to FLASH-RT even under low oxygen tension. The solution to this paradox of selective susceptibility to tumor cells to FLASH-RT, may lie in altered signaling pathways leading to disruption of the cell cycle. It is likely that the metabolism of normal cells may bring about in a different response to FLASH-RT independently of oxygen concentration [27].
2.2.2. Differing Production and Disposition of Reactive Oxygen Species
Since hyperoxia can eliminate the differential response of normal and tumor cells to FLASH-RT in mice [15], oxygen tension is undoubtedly is involved in the varying reactions of normal and transformed cells to FLASH-RT. However, other factors are obviously involved and these remain largely unresolved despite several persuasive suggestions.
The short but powerful hypoxia encountered with FLASH is unlike the continuous hypoxia found with many solid tumors. High dose rates if irradiation elevate the rare of local energy deposition leading to severe depletion of intracellular pO2. The consequences of this may be more critical under the already relatively anoxic conditions characterizing tumor tissue. Thus, already hypoxic tumor tissues where hypoxia is previously established, may be unable to withstand further FLASH-induced hypoxia [46]. Normal cells have a lower oxidant load and a higher catalase content than tumor tissue and thus can detoxify hydrogen peroxide more rapidly and this ability leads to a lower appearance of intensely oxidant species which persist much longer in transformed cells [47]. The differing response of normal and malignant tissue to FLASH may involve a combination of reactions to initial depletion of oxygen and subsequent relative persistence of harmful reactive oxygen species in tumor tissue [47].
Another factor the distinguishes tumors from normal tissues is that the iron content in the form of low molecular weight complexes, is higher in tumor tissues. This redox labile iron catalyzes key Fenton transformations instigate to greater formation of reactive oxygen species in tumors [48].
There is undoubtedly a range of intrinsic differences between the metabolic events triggered by ROS in normal tissues and tumor tissues, and this initiated differing responses to antioxidants administered after excess oxidant activity. While tumor cells are often harmed by high doses of antioxidants, normal cells remain unaffected [49]. Taking advantage of this difference has been proposed as a means of enhancing the divergence of responses to FLASH-RT relative to CONV-RT [19,50].
2.2.3. Reduction of Bystander Effects by FLASH
Normal tissues which are near sites of tumor are frequently damaged by radiation procedures. These “bystander” effects are not confined to tissues adjacent to the tumor undergoing therapy, but can also be manifested at sites that are more distant. For example, radiation therapy for prostate cancer increases the risk of a range of tumors in other tissues [51]. This problem is particularly relevant in treatment of pediatric cancer where survivors can have a long life expectancy [52]. Excess micronuclei were induced when isolated cells are exposed to serum from survivors of the Chernobyl 20 years after their exposure to high levels of radiation. The overall viability of such cells was reduced relative to those expose to control serum from an unexposed population. [53]. This illustrates the persistence of changes incurred after exposure to intense radiation. The mechanisms underlying these indirect consequences are complex but likely to involve lasting genetic changes, release of inflammatory cytokines such as IL-6 and TNF-alpha from the target irradiated cells, and upregulation of oxidative metabolism. Some of the relatively low impact of FLASH on induction of inflammation appears to have a genomic basis relating to differential expression of pro- and anti-inflammatory genes [16]. Since bystander signals can remain active in tissues for extended periods of time, other genetic and epigenetic factors are also probably involved [54]. The very short time interval employed in FLASH-RT will reduce the fraction of cells in the circulation that are exposed to radiation. It has been estimated that 100 times more blood volume is subjected to irradiation by CONV-RT than by FLASH-RT and this large difference could account for a significant proportion of the reduced damage effected by FLASH-RT [55]. This difference may help to maintain immune surveillance in an effective state following FLASH rather than having it descend into non-targeted inflammation.
2.2.4. Immune Responses
The extent of T-cell incursion into tumors was increased by FLASH-RT, suggesting a more effective immune attack upon transformed cells [56]. Activation of myosin light chain proteins (MLPs) promotes the invasiveness of tumors [57,58] and is also involved in the activation of double-stranded DNA breaks [56]. CONV-RT but not FLASH-RT activates MLP. Pharmacological inhibition of MLP activation enables CONV-RT to mimic some of the advantages of FLASH-RT, including promotion of penetrance of active immune cells into tumor tissue and in inhibition of repair of double stranded DNA [56]. This reveals DNA repair and effective immune responses to comprise a significant component of the distinctive efficacy of FLASH-RT. The dissimilar response of the immune system to FLASH-RT in comparison to CONV-RT may also account for the selective susceptibility of tumor cells. The improved recruitment of lymphocytes found in FLASH-RT may markedly favor the destruction of tumor cells [59].
3. Outcome
Overall, FLASH-RT has a multitude of protective effects on biological tissues that differ from CONV-RT including limiting inflammatory cell infiltration, lowering generation of inflammatory factors, and decreasing the extent of radiation-induced fibrosis. Desirable protective effects of FLASH have now been reported in a comprehensive variety of organ systems [60]. Stem cell function is preserved and beneficial immune responses tolerated. In the brain, synaptic connectivity is and measures of cerebral function are maintained.
Both of the two questions posited above, remain incompletely answered, and challenge orthodox thinking in this area. Despite the fact that the mechanisms underlying the difference between FLASH-RT and conventional more prolonged radiotherapy, are not fully accounted for, the tactic of delivering radiation to cancer patients at a high dose rate for a short time should have major effect on improving the line of attack of cancer treatment.
Since there is no distinction between the susceptibility of normal and tumor tissues to conventional radiation, application of this CONV-RT relies on directing the radiation beam to a very focused target. The efficacy of treatment relies solely on precisely distinguishing between the location of healthy and tumor tissues. In the case of brain, stereotactic surgery involving bring many sources of radiation to a focal point, the “gamma knife” has been employed. However, since brain tumors while often not metastatic, are very comingled with brain tissue, the three month survival rate following this procedure remains under 10 months [61]. In contrast FLASH-RT does not rely solely on spatial separation of normal and abnormal tissue regions but takes advantage of their differential metabolic characteristics.
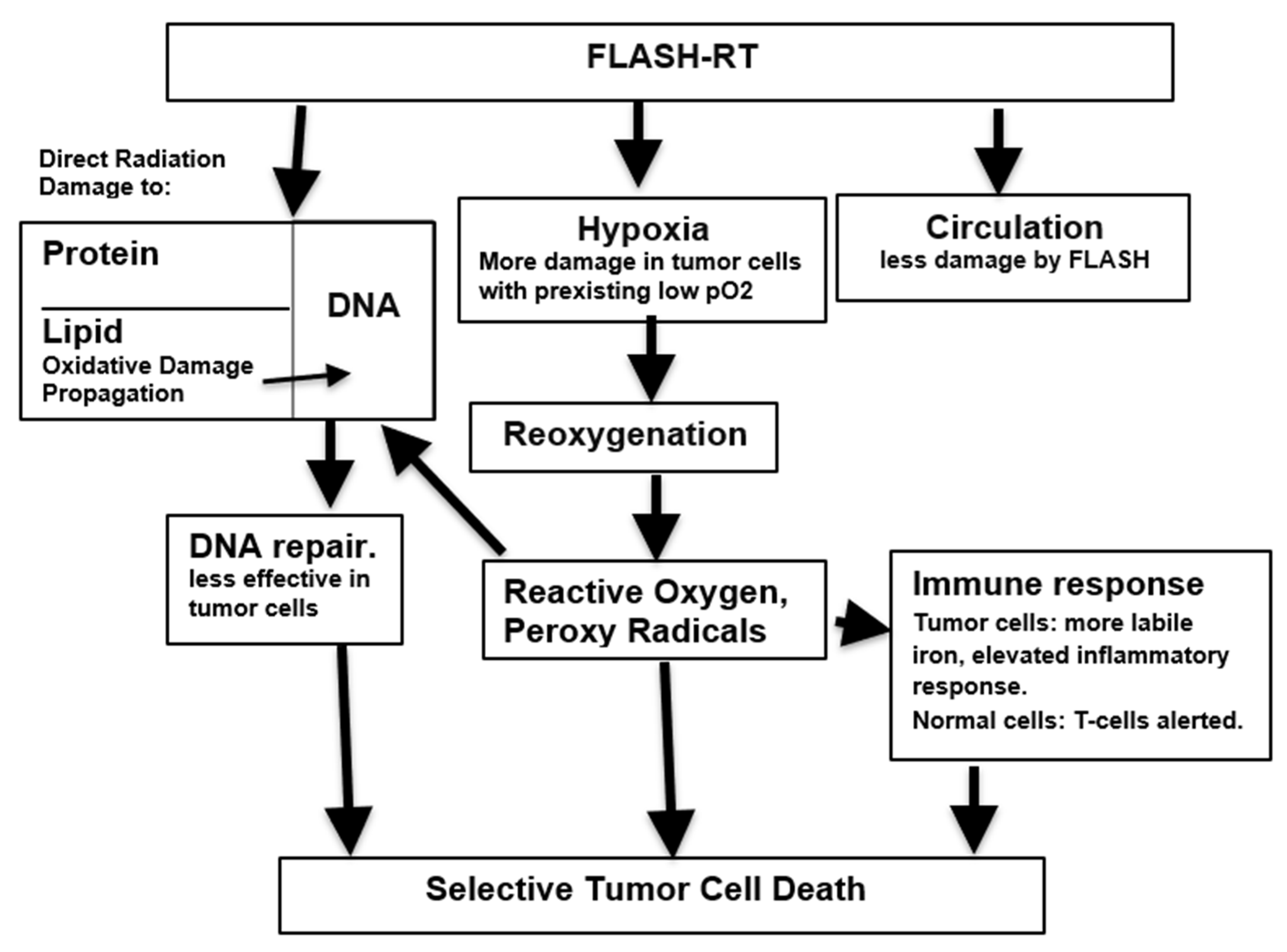
Any explanation of the mechanism of action leading to the advantageous properties of FLASH-RT must take into account the remarkable fact that they are have been described using several very different means of irradiation including electrons, protons and X-rays [62]. While many suggestions have been made to account for the FLASH phenomenon, these need not mutually exclusive but rather may reinforce each other. Figure 1 represents a possible integration of data that is inclusive, and places various hypotheses on a sequential lymphocytes from scale.
Research in this area is at a relatively early stage and several contradictory results have been reported. For example, FLASH has been found to have a sparing effect of some tumor cell lines under normoxic conditions [19]. There are especially conflicting data surrounding the effects of FLASH-RT on lymphocytes. In animal models of radiation-induced cardiac and splenic lymphopenia, high dose short duration exposure irradiation actually worsens the resulting injury, when compared with lower dose more prolonged exposure [63]. Similar absence of a sparing effect has been reported by others [64]. However, other reports describe a sparing effect of FLASH-RT lymphocytes from both experimental animals and from humans [65,66]. The normal oxygen pO2 within tissues is in the range of 3-7% considerably below that of ambient atmospheric pO2 levels of 18% [67]. Hyperoxia is known to block the advantageous effects of FLASH. Since many of the studies in isolated systems have been performed under normal concentrations of atmospheric oxygen, they are subject to the limitation that they are not replicating the more hypoxic conditions existing within intact tissues [68]. This could block the ability to observe the distinctive FLASH differential distinguishing it from CONV and may account for some of the conflicting data reported from isolated systems. Replication of exact physiological conditions prevailing in vivo is virtually impossible to achieve. Nevertheless, while it is more challenging to unravel mechanistic pathways in intact animals, perhaps this is the only means to fully establish exactly the sequence of events that lie behind the FLASH phenomenon. A report using an in vivo preparation, found that a single FLASH dose did not cause profound anoxia in intact muscle tissue. As a result, it was proposed that the difference in cellular damage found between FLASH-RT and CONV-RT, rather than being due to acute anoxia, is caused by the difference in the rate of tissue oxygen consumption between the two conditions. This can be higher in the case of CONV-RT [21].
Delayed undesirable effects such as osteonecrosis following treatment of canine oral cancer have been reported [10]. The treatment of human tumors requires an improved and more detailed understanding of the optimal parameters for many factors such as radiation spectrum, image guidance and underlying mechanisms. Unanswered considerations include the question of whether a high energy burst using a proton beam may be as effective as an electron beam [69]. The optimal variables for establishing these variables remain to be delineated before this treatment can be clinically applied on a broad scale. Protocols are currently being developed in more detail in order to allow safe testing and application of this procedure in humans [70].
It has been found that in an animal model of glioblastoma, while FLASH-RT produced less unwanted side effects than exposure to a standard radiation pattern, nevertheless, the overall survival rate was unaffected [71]. If such results were more widely confirmed, the value of FLASH-RT would be more limited than hoped. However, findings using experimental animals have generally found more lasting positive effects. It must be borne in mind that the FLASH approach is not necessarily devoid of potential harm to normal tissues if the wrong radiation protocols were chosen.
Further understanding of the events that lead to the distinctive properties of FLASH-RT will allow refinement of this strategy and enhancement of its efficiency. Many parameters remain to be studied in more detail. A variant of FLASH therapy whereby treatment is divided into two consecutive exposures may have particular value in protection of the developing juvenile brain which is particularly sensitive to radiation-induced damage [72]. The relative insensitivity of normal tissue to FLASH suggests that higher overall doses than those currently used, might be employed, thus effecting great damage to tumor tissue. Surprisingly, the shielding nature of the FLASH effect has been observed after exposure of biological tissues to a wide range of particles including electrons, x-rays, and protons [16]. It is important that reports on this subject clearly define the exact physical nature of the beam used including dose rate and beam width. The importance of precise description of parameters used is paramount in this emerging and fluid field. In addition to pinpointing the most suitable conditions for application of the procedure, the relative suitability of various types of radiation remain to be unambiguously defined. However, even in the absence of a more comprehensive expansion of knowledge concerning FLASH, this novel technique is already proving to be of growing usefulness in clinical radiotherapy.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Begg, A.C.; Stewart, F.A.; Vens, C. Strategies to improve radiotherapy with targeted drugs. Nat Rev Cancer. 2011, 11, 239–253. [Google Scholar] [CrossRef]
- Dewey, D.L.; Boag, J.W. Modification of the oxygen effect when bacteria are given large pulses of radiation. Nature. 1959, 183, 1450–1451. [Google Scholar] [CrossRef]
- Vozenin M,C,; De Fornel, P. ; Petersson, K.; Favaudon, V.; Jaccard, M.; Germond, J.F.; Petit, B.; Burki, M.; Ferrand, G.; Patin, D.; Bouchaab,H.; Ozsahin, M.; Bochud, F.; Bailat, C.; Devauchelle, P.; Bourhis, J. The Advantage of FLASH Radiotherapy Confirmed in Mini-pig and Cat-cancer Patients. Clin Cancer Res. 2019, 25, 35–42.
- Lin, B.; Gao, F.; Yang, Y.; Wu, D.; Zhang, Y.; Feng, G.; Dai, T.; Du, X. FLASH Radiotherapy: History and Future. Front Oncol. 2021, 11, 644400. [Google Scholar] [CrossRef]
- Jin, J.Y.; Gu, A.; Wang, W.; Oleinick, N.L.; Machtay, M.; Spring Kong, F.M. Ultra-high dose rate effect on circulating immune cells: A potential mechanism for FLASH effect? Radiother Oncol. 2020, 149, 55–62. [Google Scholar] [CrossRef]
- Favaudon, V.; Labarbe, R.; Limoli, C.L. Model studies of the role of oxygen in the FLASH effect. Med Phys. 2022, 49, 2068–2081. [Google Scholar] [CrossRef]
- Hubenak, J.R.; Zhang, Q.; Branch, C.D.; Kronowitz, S.J. Mechanisms of injury to normal tissue after radiotherapy: a review. Plast Reconstr Surg. 2014, 133, 49e–56e. [Google Scholar] [CrossRef]
- Bourhis, J.; Sozzi, W.J.; Jorge, P.G.; Gaide, O.; Bailat, C.; Duclos, F.; Patin, D.; Ozsahin, M.; Bochud, F.; Germond, J.F.; Moeckli, R.; Vozenin, M.C. Treatment of a first patient with FLASH-radiotherapy. Radiother Oncol, 2019; 139, 18–22. [Google Scholar]
- Konradsson, E.; Szecsenyi, R.E.; Adrian, G.; Coskun, M. ; Børresen B.; Arendt, M.L.; Erhart, K.; Bäck,S.Å.; Petersson, K.; Ceberg, C. Evaluation of intensity-modulated electron FLASH radiotherapy in a clinical setting using veterinary cases. Med. Phys. 2023, 50, 6569–6579. [Google Scholar] [CrossRef]
- Børresen, B.; Arendt, M.L.; Konradsson, E.; Bastholm Jensen, K.; Bäck, S.Å.; Munck, A.f.; Rosenschöld, P.; Ceberg, C.; Petersson, K. Evaluation of single-fraction high dose FLASH radiotherapy in a cohort of canine oral cancer patients. Front Oncol. 2023, 13, 256760. [Google Scholar] [CrossRef]
- Swarts, S.G.; Flood, A.B.; Swartz, H.M. Implications of “flash” radiotherapy for biodosimetry. Radiat Prot Dosimetry. 2023, 199, 1450–1459. [Google Scholar] [CrossRef]
- Bourhis, J.; Montay-Gruel, P.; Gonçalves Jorge, P.; Bailat, C.; Petit, B.; Ollivier, J.; Jeanneret-Sozzi, W.; Ozsahin, M.; Bochud, F.; Moeckli, R.; Germond, J.F.; Vozenin, M.C. Clinical translation of FLASH radiotherapy: Why and how? Radiother Oncol. 2019, 139, 11–17. [Google Scholar] [CrossRef]
- Hendry, J.H.; Moore, J.V.; Hodgson, B.W.; Keene, J.P. The constant low oxygen concentration in all the target cells for mouse tail radionecrosis. Radiat Res. 1982, 92, 172–181. [Google Scholar] [CrossRef]
- Hall, E.J.; Brenner, D.J. The dose-rate effect revisited: radiobiological considerations of importance in radiotherapy. Int J Radiat Oncol Biol Phys. 1991, 21, 1403–1414. [Google Scholar] [CrossRef]
- Montay-Gruel, P.; Acharya, M.M.; Petersson, K.; Alikhani, L.; Yakkala, C.; Allen, B.D.; Ollivier, J.; Petit, B.; Jorge, P.G.; Syage, A.R.; Nguyen, T.A.; Baddour, A.A.D.; Lu, C.; Singh, P.; Moeckli, R.; Bochud, F.; Germond, J.F.; Froidevaux, P.; Bailat, C.; Bourhis, J.; Vozenin, M.C.; Limoli, C.L. Long-term neurocognitive benefits of FLASH radiotherapy driven by reduced reactive oxygen species. Proc Natl Acad Sci U S A. 2019, 116, 10943–10951. [Google Scholar] [CrossRef]
- Friedl, A.A.; Prise, K.M.; Butterworth, K.T.; Montay-Gruel, P.; Favaudon, V. Radiobiology of the FLASH effect. Med Phys. 2022, 49, 1993–2013. [Google Scholar] [CrossRef]
- Limoli, C.L.; Kramár, E.A.; Almeida, A.; Petit, B.; Grilj, V.; Baulch, J.E.; Ballesteros-Zebadua, P.; Loo, B.W. Jr.; Wood, M.A.; Vozenin, M.C. The sparing effect of FLASH-RT on synaptic plasticity is maintained in mice with standard fractionation. Radiother Oncol. 2023, 186, 109767. [Google Scholar] [CrossRef]
- Tavakkoli, A.D.; Clark, M.A.; Kheirollah, A.; Sloop, A.M.; Soderholm, H.E.; Daniel, N.J.; Petusseau, A.F.; Huang, Y.H.; Thomas, C.R. Jr.; Jarvis, L.A.; Zhang, R.; Pogue, B.W.; Gladstone, D.J.; Hoopes, P.J. Anesthetic Oxygen Use and Sex Are Critical Factors in the FLASH Sparing Effect. Adv Radiat Oncol. 2024, 9, 101492. [Google Scholar] [CrossRef]
- Adrian, G.; Konradsson, E.; Lempart, M.; Bäck, S.; Ceberg, C.; Petersson, K. The FLASH effect depends on oxygen concentration. Br J Radiol. 2020, 93, 20190702. [Google Scholar] [CrossRef]
- Moon, E.J.; Petersson, K.; Olcina, M.M. The importance of hypoxia in radiotherapy for the immune response, metastatic potential and FLASH-RT. Int J Radiat Biol. 2022, 98, 439–451. [Google Scholar] [CrossRef]
- El Khatib, M.; Van Slyke, A.L.; Velalopoulou, A.; Kim, M.M.; Shoniyozov, K.; Allu, S.R.; Diffenderfer, E.E.; Busch, T.M.; Wiersma, R.D.; Koch, C.J.; Vinogradov, S.A. Ultrafast Tracking of Oxygen Dynamics During Proton FLASH. Int J Radiat Oncol Biol Phys. 2022, 113, 624–634. [Google Scholar] [CrossRef]
- Labarbe, R.; Hotoiu, L.; Barbier, J.; Favaudon, V. A physicochemical model of reaction kinetics supports peroxyl radical recombination as the main determinant of the FLASH effect. Radiother Oncol. 2020, 153, 303–310. [Google Scholar] [CrossRef]
- Favaudon V, Labarbe R, Limoli CL. Model studies of the role of oxygen in the FLASH effect. Med Phys. 2022 Mar;49(3):2068-2081. Epub 2021 Aug 18. PMCID: PMC8854455. [CrossRef] [PubMed]
- Ramos-Méndez, J.; Domínguez-Kondo, N.; Schuemann, J.; McNamara, A.; Moreno-Barbosa, E.; Faddegon, B. LET-Dependent Intertrack Yields in Proton Irradiation at Ultra-High Dose Rates Relevant for FLASH Therapy. Radiat Res. 2020, 194, 351–362. [Google Scholar] [CrossRef]
- Perstin, A.; Poirier, Y.; Sawant, A.; Tambasco, M. Quantifying the DNA-damaging Effects of FLASH Irradiation With Plasmid DNA. Int J Radiat Oncol Biol Phys. 2022, 113, 437–447. [Google Scholar] [CrossRef]
- Alizadeh, E.; Sanz, A.G.; García, G.; Sanche, L. Radiation Damage to DNA: The Indirect Effect of Low Energy Electrons. J Phys Chem Lett. 2013, 4, 820–825. [Google Scholar] [CrossRef]
- Garty, G.; Obaid, R.; Deoli, N.; Royba, E.; Tan, Y.; Harken, A.D.; Brenner, D.J. Ultra-high dose rate FLASH irradiator at the radiological research accelerator facility. Sci Rep. 2022, 12, 22149. [Google Scholar] [CrossRef]
- Schüler, E.; Acharya, M.; Montay-Gruel, P.; Loo, B.W. Jr.; Vozenin, M.C.; Maxim, P.G. Ultra-high dose rate electron beams and the FLASH effect: From preclinical evidence to a new radiotherapy paradigm. Med Phys. 2022, 49, 2082–2095. [Google Scholar] [CrossRef]
- Small, K.L.; Henthorn, N.T.; Angal-Kalinin, D. Chadwick, A. L.; Santina, E.; Aitkenhead, A.; Kirkby, K.J.; Smith, R.J.; Surman, M.; Jones, J.; Farabolini, W.; Corsini, R.; Gamba, D.; Gilardi. A.; Merchant, M.J.; Jones, R.M. Evaluating very high energy electron RBE from nanodosimetric pBR322 plasmid DNA damage. Sci Rep. 2021, 11, 3341. [Google Scholar]
- Ohsawa, D.; Hiroyama, Y.; Kobayashi, A.; Kusumoto, T.; Kitamura, H.; Hojo, S.; Kodaira, S.; Konishi, T. DNA strand break induction of aqueous plasmid DNA exposed to 30 MeV protons at ultra-high dose rate. J Radiat Res. 2022, 63, 255–260. [Google Scholar] [CrossRef]
- Kumar, K.; Kumar, S.; Datta, K.; Fornace, A.J. Jr.; Suman, S. High-LET-Radiation-Induced Persistent DNA Damage Response Signaling and Gastrointestinal Cancer Development. Curr Oncol. 2023, 30, 5497–5514. [Google Scholar] [CrossRef]
- Iturri, L.; Bertho, A.; Lamirault, C.; Juchaux, M.; Gilbert, C.; Espenon, J.; Sebrie, C.; Jourdain, L.; Pouzoulet, F.; Verrelle, P.; De Marzi, L.; Prezado, Y. Proton FLASH Radiation Therapy and Immune Infiltration: Evaluation in an Orthotopic Glioma Rat Model. Int J Radiat Oncol Biol Phys. 2023, 116, 655–665. [Google Scholar] [CrossRef]
- Bondy, S.C. Mitochondrial Dysfunction as the Major Basis of Brain Aging. Biomolecules. 2024, 14, 402. [Google Scholar] [CrossRef]
- Zhou, X.; Li, N.; Wang, Y.; Wang, Y.; Zhang, X.; Zhang, H. Effects of X-irradiation on mitochondrial DNA damage and its supercoiling formation change. Mitochondrion. 2011, 11, 886–892. [Google Scholar] [CrossRef]
- Guo, Z.; Buonanno, M.; Harken, A.; Zhou, G.; Hei, T.K. Mitochondrial Damage Response and Fate of Normal Cells Exposed to FLASH Irradiation with Protons. Radiat Res. 2022, 197, 569–582. [Google Scholar] [CrossRef]
- Trappetti, V.; Fazzari, J.; Fernandez-Palomo, C.; Smyth, L.; Potez, M.; Shintani, N.; de Breuyn Dietler, B.; Martin, O.A.; Djonov, V. Targeted Accumulation of Macrophages Induced by Microbeam Irradiation in a Tissue-Dependent Manner. Biomedicines. 2022, 10, 735. [Google Scholar] [CrossRef]
- Buonanno, M.; Grilj, V.; Brenner, D.J. Biological effects in normal cells exposed to FLASH dose rate protons. Radiother Oncol. 2019, 139, 51–55. [Google Scholar] [CrossRef]
- Elbakrawy, E.; Kaur Bains, S.; Bright, S.; Al-Abedi, R.; Mayah, A.; Goodwin, E.; Kadhim, M. Radiation-Induced Senescence Bystander Effect: The Role of Exosomes. Biology (Basel). 2020, 9, 191. [Google Scholar] [CrossRef]
- Fouillade, C.; Curras-Alonso, S.; Giuranno, L.; Quelennec, E.; Heinrich, S.; Bonnet-Boissinot, S.; Beddok, A.; Leboucher, S.; Karakurt, H.U.; Bohec, M.; Baulande, S.; Vooijs, M.; Verrelle, P.; Dutreix, M.; Londoño-Vallejo, A.; Favaudon, V. FLASH Irradiation Spares Lung Progenitor Cells and Limits the Incidence of Radio-induced Senescence. Clin Cancer Res. 2020, 26, 1497–1506. [Google Scholar] [CrossRef]
- Giaccia, A.J. Hypoxic Stress Proteins: Survival of the Fittest. Semin Radiat Oncol. 1996, 6, 46–58. [Google Scholar] [CrossRef]
- Adekola, K.; Rosen, S.T.; Shanmugam, M. Glucose transporters in cancer metabolism. Curr Opin Oncol. 2012, 24, 650–654. [Google Scholar] [CrossRef]
- Ganapathy-Kanniappan, S.; Geschwind, J.F. Tumor glycolysis as a target for cancer therapy: progress and prospects. Mol Cancer. 2013, 12, 152. [Google Scholar] [CrossRef]
- Aboelella, N.S.; Brandle, C.; Kim, T.; Ding, Z.C.; Zhou, G. Oxidative Stress in the Tumor Microenvironment and Its Relevance to Cancer Immunotherapy. Cancers (Basel). 2021, 13, 986. [Google Scholar] [CrossRef]
- Alanazi, A.; Jay-Gerin, J.P.; Blázquez-Castro, A. Is singlet oxygen involved in FLASH-RT? J Appl Clin Med Phys. 2023, 24, e13974. [Google Scholar] [CrossRef]
- Leavitt, R.J.; Almeida, A.; Grilj, V.; Montay-Gruel, P.; Godfroid, C.; Petit, B.; Bailat, C.; Limoli, C.L.; Vozenin, M.C. Acute Hypoxia Does Not Alter Tumor Sensitivity to FLASH Radiation Therapy. Int J Radiat Oncol Biol Phys. 2024, S0360-3016, 00320–00321. [Google Scholar] [CrossRef]
- Abolfath, R.; Grosshans, D.; Mohan, R. Oxygen depletion in FLASH ultra-high-dose-rate radiotherapy: A molecular dynamics simulation. Med Phys. 2020, 47, 6551–6561. [Google Scholar] [CrossRef]
- Ma, J.; Gao, H.; Shen, X.; Bai, X.; Tang, M. A FLASH model of radiolytic oxygen depletion and reactive oxygen species for differential tumor and normal-tissue response. medRxiv. 2023, 2023.10.20.23297337. [Google Scholar]
- Spitz, D.R.; Buettner, G.R.; Petronek, M.S.; St-Aubin, J.J.; Flynn, R.T.; Waldron, T.J.; Limoli, C.L. An integrated physico-chemical approach for explaining the differential impact of FLASH versus conventional dose rate irradiation on cancer and normal tissue responses. Radiother Oncol. 2019, 139, 23–27. [Google Scholar] [CrossRef]
- Prasad, K.N.; Sinha, P.K.; Ramanujam, M.; Sakamoto, A. Sodium ascorbate potentiates the growth inhibitory effect of certain agents on neuroblastoma cells in culture. Proc Natl Acad Sci U S A. 1979, 76, 829–832. [Google Scholar] [CrossRef]
- Prasad, K.N.; Bondy, S.C. FLASH Radiation vs. Conventional Radiation: Basis of Varying Effects and Potential Improvement of Their Differential Effects on Normal and Malignant Cells. J. Adv. Med. Sci. 2024. (in press) [Google Scholar]
- Wallis, C.J.; Mahar, A.L.; Choo, R.; Herschorn, S.; Kodama, R.T.; Shah, P.S.; Danjoux, C.; Narod, S.A.; Nam, R.K. Second malignancies after radiotherapy for prostate cancer: systematic review and meta-analysis. BMJ. 2016, 352, i851. [Google Scholar] [CrossRef]
- Casey, D.L.; Vogelius, I.R.; Brodin, N.P.; Roberts, K.B.; Avanzo, M.; Moni, J.; Owens, C.; Ronckers, C.M.; Constine, L.S.; Bentzen, S.M.; Olch, A. Risk of Subsequent Neoplasms in Childhood Cancer Survivors After Radiation Therapy: A PENTEC Comprehensive Review. Int J Radiat Oncol Biol Phys. 2024, 119, 640–654. [Google Scholar] [CrossRef]
- Marozik, P.; Mothersill, C.; Seymour, C.B.; Mosse, I.; Melnov, S. Bystander effects induced by serum from survivors of the Chernobyl accident. Exp Hematol. 2007, 35, 55–63. [Google Scholar] [CrossRef]
- Tang, H.; Cai, L.; He, X.; Niu, Z.; Huang, H.; Hu, W.; Bian, H.; Huang, H. Radiation-induced bystander effect and its clinical implications. Front Oncol. 2023, 13, 1124412. [Google Scholar] [CrossRef]
- Rudigkeit, S.; Schmid, T.E.; Dombrowsky, A.C.; Stolz, J.; Bartzsch, S.; Chen, C.B.; Matejka, N.; Sammer, M.; Bergmaier, A.; Dollinger, G.; Reindl, J. Proton-FLASH: effects of ultra-high dose rate irradiation on an in-vivo mouse ear model. Sci Rep. 2024, 14, 1418. [Google Scholar] [CrossRef]
- Kim, Y.E.; Gwak, S.H.; Hong, B.J.; Oh, J.M.; Choi, H.S.; Kim, M.S.; Oh, D.; Lartey, F.M.; Rafat, M.; Schüler, E.; Kim, H.S.; von Eyben, R.; Weissman, I.L.; Koch, C.J.; Maxim, P.G.; Loo, B.W. Jr.; Ahn, G.O. Effects of Ultra-high dose rate FLASH Irradiation on the Tumor Microenvironment in Lewis Lung Carcinoma: Role of Myosin Light Chain. Int J Radiat Oncol Biol Phys. 2021, 109, 1440–1453. [Google Scholar] [CrossRef]
- Foroumadi, R.; Rashedi, S.; Asgarian, S.; Mardani, M.; Keykhaei, M.; Farrokhpour, H.; Javanshir, S.; Sarallah, R.; Rezaei, N. Circular RNA MYLK as a prognostic biomarker in patients with cancers: A systematic review and meta-analysis. Cancer Rep (Hoboken). 2022, 5, e1653. [Google Scholar] [CrossRef]
- Zhou, R.; Chen, J.; Xu, Y.; Ye, Y.; Zhong, G.; Chen, T.; Qiu, L. PRPF19 facilitates colorectal cancer liver metastasis through activation of the Src-YAP1 pathway via K63-linked ubiquitination of MYL9. Cell Death Dis. 2023, 14, 258. [Google Scholar] [CrossRef]
- Bogaerts, E.; Macaeva, E.; Isebaert, S.; Haustermans, K. Potential Molecular Mechanisms behind the Ultra-High Dose Rate “FLASH” Effect. Int J Mol Sci. 2022, 23, 12109. [Google Scholar] [CrossRef]
- Tang, R.; Yin, J.; Liu, Y.; Xue, J. FLASH radiotherapy: A new milestone in the field of cancer radiotherapy. Cancer Lett. 2024, 587, 216651. [Google Scholar] [CrossRef]
- Park, H.; Chung, H.T.; Kim, J.W.; Dho, Y.S.; Lee, E.J. A 3-month survival model after Gamma Knife surgery in patients with brain metastasis from lung cancer with Karnofsky performance status ≤ 70. Sci Rep. 2023, 13, 13159. [Google Scholar] [CrossRef]
- Gao, Y.; Liu, R.; Chang, C.W.; Charyyev, S.; Zhou, J.; Bradley, J.D.; Liu, T.; Yang, X. A potential revolution in cancer treatment: A topical review of FLASH radiotherapy. J Appl Clin Med Phys. 2022, 23, e13790. [Google Scholar] [CrossRef]
- Venkatesulu, B.P.; Sharma, A.; Pollard-Larkin, J.M.; Sadagopan, R.; Symons, J.; Neri, S.; Singh, P.K.; Tailor, R.; Lin, S.H.; Krishnan, S. Ultra high dose rate (35 Gy/sec) radiation does not spare the normal tissue in cardiac and splenic models of lymphopenia and gastrointestinal syndrome. Sci Rep. 2019, 9, 17180. [Google Scholar] [CrossRef]
- Zhang, Q.; Gerweck, L.E.; Cascio, E.; Gu, L.; Yang, Q.; Dong, X.; Huang, P.; Bertolet, A.; Nesteruk, K.P.; Sung, W.; McNamara, A.L.; Schuemann, J. Absence of Tissue-Sparing Effects in Partial Proton FLASH Irradiation in Murine Intestine. Cancers (Basel). 2023, 15, 2269. [Google Scholar] [CrossRef]
- Cucinotta, F.A.; Smirnova, O.A. Effects of Flash Radiotherapy on Blood Lymphocytes in Humans and Small Laboratory Animals. Radiat Res. 2023, 199, 240–251. [Google Scholar] [CrossRef]
- Galts, A.; Hammi, A. FLASH radiotherapy sparing effect on the circulating lymphocytes in pencil beam scanning proton therapy: impact of hypofractionation and dose rate. Phys Med Biol. 2024, 69. [Google Scholar] [CrossRef]
- Wilson, J.D.; Hammond, E.M.; Higgins, G.S.; Petersson, K. Ultra-High Dose Rate (FLASH) Radiotherapy: Silver Bullet or Fool’s Gold? Front Oncol. 2020, 9, 1563. [Google Scholar] [CrossRef]
- Melia, E.; Parsons, J.L. DNA damage and repair dependencies of ionising radiation modalities. Biosci Rep. 2023, 43, BSR20222586. [Google Scholar] [CrossRef]
- Böhlen, T.T.; Germond, J.F.; Desorgher, L.; Veres, I.; Bratel, A.; Landström, E.; Engwall, E.; Herrera, F.G.; Ozsahin, E.M.; Bourhis, J.; Bochud, F.; Moeckli, R. Very high-energy electron therapy as light-particle alternative to transmission proton FLASH therapy - An evaluation of dosimetric performances. Radiother Oncol. 2024, 194, 110177. [Google Scholar] [CrossRef]
- Moeckli, R.; Gonçalves Jorge, P.; Grilj, V.; Oesterle, R.; Cherbuin, N.; Bourhis, J.; Vozenin, M.C.; Germond, J.F.; Bochud, F.; Bailat, C. Commissioning of an ultra-high dose rate pulsed electron beam medical LINAC for FLASH RT preclinical animal experiments and future clinical human protocols. Med Phys. 2021, 48, 3134–3142. [Google Scholar] [CrossRef]
- Liljedahl, E.; Konradsson, E.; Linderfalk, K.; Gustafsson, E.; Petersson, K.; Ceberg, C.; Redebrandt, H.N. Comparable survival in rats with intracranial glioblastoma irradiated with single-fraction conventional radiotherapy or FLASH radiotherapy. Front Oncol. 2024, 13, 1309174. [Google Scholar] [CrossRef]
- Allen, B.D.; Alaghband, Y.; Kramár, E.A.; Ru, N.; Petit, B.; Grilj, V.; Petronek, M.S.; Pulliam, C.F.; Kim, R.Y.; Doan, N.L.; Baulch, J.E.; Wood, M.A.; Bailat, C.; Spitz, D.R.; Vozenin, M.C.; Limoli, C.L. Elucidating the neurological mechanism of the FLASH effect in juvenile mice exposed to hypofractionated radiotherapy. Neuro Oncol. 2023, 25, 927–939. [Google Scholar] [CrossRef]
Figure 1.
Putative mechanisms underlying susceptibility of tumor cells to FLASH radiation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



