
Submitted:
09 June 2024
Posted:
12 June 2024
You are already at the latest version
Abstract
Increased anthropogenic pressure on forest resources leads to deforestation and forest degradation, significantly limiting the regeneration capacity of native woody species and consequently the restoration of miombo woodlands in anthropized habitats within the rural area of Lubumbashi. This study assessed the diversity and natural regeneration capacity of miombo species through floristic inventories in three different habitats (unexploited forests, degraded forests, and post-cultivation fallows). The results reveal that for the adult’s stratum unexploited and degraded forests exhibit higher dendrometric (density, mean square diameter, basal area) and floristic parameter (taxa, genera, families) values compared to post-cultivation fallows. Furthermore, the regeneration of miombo woody species is higher in degraded forests (21 taxa; 105 juveniles/plot). However, regarding the sapling’s stratum (1 cm ≤ dbh < 10 cm), the three habitats display nearly similar situations. Additionally, the floristic composition and diversity of unexploited and degraded forests show a significantly higher similarity (76.50%) among them compared to these habitats and the post-cultivation fallows (56.00%). These findings indicate that miombo woodlands have the potential to regenerate and maintain floristic diversity. To sustain this natural regeneration capacity of miombo woody species and promote the restoration of forest mass and its floristic diversity, it is imperative to determine the rotation period after habitat exploitation and regulate anthropogenic activities and late bush fires, particularly in anthropized habitats at the village level.
Keywords:
Anthropogenic pressure
; deforestation
; forest degradation
; floristic diversity
; natural regeneration
; miombo woodlands.
1. Introduction
Forests constitute one of the most crucial terrestrial biomes on the planet, harboring 80% of terrestrial biodiversity [1,2] across approximately 4.06 billion hectares [3]. In Africa, forests cover 23% of the continent, totaling 675 million hectares [4], with nearly 10% of this area dominated by miombo woodlands [5,6]. Miombo woodlands are predominantly composed of woody species from the genera Brachystegia, Julbernardia, and Isoberlinia [7]. These woodlands span about 2.8 million km² in the Zambezi region [8], supporting the livelihoods of over 100 million rural and urban residents through the ecosystem services they provide [6,9]. Moreover, miombo woodlands boast significant biodiversity with high endemism rates, making them a conservation priority [10,11].
However, miombo woodlands are experiencing a reduction in area due to natural and particularly anthropogenic factors [12]. The combination of population growth and a deleterious socio-economic and political context, which forces local populations to heavily rely on forest resources for survival [13], leads to deforestation and degradation [14,15]. Furthermore, inadequate, and poorly enforced forestry legislation [16] results in unsustainable exploitation of forest resources, exacerbating deforestation and degradation [17].
The direct anthropogenic drivers of this change are primarily agriculture and charcoal production [15,18], both of which are itinerant [19]. Additional factors include the extraction of timber and craft wood, fuelwood, and late and repeated bushfires, all contributing to forest loss [20,21]. Consequently, the annual conversion rate of miombo woodlands ranges from 2% to 22% within the miombo ecoregion [21], with significantly higher rates in countries with intense anthropogenic pressure due to population poverty, such as the Democratic Republic of Congo (D.R. Congo). Despite its high forest potential, D.R. Congo has the highest annual deforestation rate in the Congo Basin, approximately -0.4% between 2001 and 2019 [22]. Furthermore, in the southeastern D.R. Congo, where miombo woodlands is the dominant vegetation unit [23], its coverage dropped from nearly 70% to 43% between 2000 and 2010 [24]. In this region, the miombo deforestation rate is even higher in rural areas adjacent to major cities, such as the rural area of Lubumbashi, which has a deforestation rate of 1.51% [15]. This situation contributes to environmental degradation and threatens the livelihoods of rural and urban populations dependent on miombo woodlands [2,9].
To address this deforestation and forest degradation, forest cover restoration is one of the recommended solutions [5,25-27]. Restoration involves adaptive processes that implement practices to restore ecological functionality and enhance human survival in deforested or degraded habitats [27]. This can be achieved through reforestation using fast-growing exotic woody species, which allows for the short-term reconstitution of vegetation cover and the availability of ecosystem services [19]. However, these exotic species pose a threat to native biodiversity and alter the original forest functions [28,29]. Therefore, using native woody species remains a viable alternative, ensuring the continuity of ecosystem service production by maintaining floristic composition, structure, and function [1]. This restoration typically involves nursery seedling production or facilitating natural regeneration in habitats. However, combined with logistical complexity management, nursery and the final establishment of seedlings in human-disturbed habitats can be costly, reducing its applicability [5]. In this perspective, promoting natural regeneration is a sustainable and optimal alternative to current forest loss [25]. Natural regeneration allows adult individuals in a plant community to replace themselves by establishing juveniles in the undergrowth (dbh < 10 cm) [30]. This regeneration, which ensures the persistence of woody species [31], is dependent on the disturbance gradient of habitats and the resilience of woody species to these disturbances [14].
Furthermore, several studies on the natural regeneration of forests in anthropized habitats have already been conducted in the miombo ecoregion [2,6,14,21,32-35]. However, these studies have predominantly focused on Southern Africa, while no research on natural regeneration has been initiated in the miombo woodlands of Central Africa, whose ecological and floristic characteristics increasingly differ from those of Southern African miombo [23]. Additionally, no study has been conducted to analyze the natural regeneration of miombo woodlands through forest inventory. This inventory technique remains reliable for assessing the capacity of woody species to regenerate and consequently restore forest cover [31,36]. Moreover, results on the natural regeneration capacity of woody species are valuable for forest management, sustainable biodiversity management [20], and implementing responses to human disturbances to ensure miombo woodlands resilience [14].
In this context, the present study was initiated to evaluate the natural regeneration capacity of miombo woody species in the rural area of Lubumbashi. It tests the hypothesis that (i) the density, average diameter, basal area, and floristic diversity differ among habitats due to anthropogenic disturbances. Higher values are expected in unexploited forests and lower values in post-cultivation fallows, with degraded forests in between. (ii) The regeneration capacity of miombo species is higher in degraded forests than in unexploited forests and post-cultivation fallows, due to the availability of resources (water, light, space) and lower intra/inter-specific competition and disturbances. (iii) The floristic diversity of habitats shows similarities. Higher similarities in floristic composition are expected between unexploited and degraded forests compared to post-cultivation fallows, due to the lower disturbances in these habitats compared to post-cultivation fallows.
2. Materials and Methods
2.1. Study Area
The present study was conducted in the rural area of Lubumbashi, located in southeastern D.R. Congo (Figure 1).
Situated at an altitude ranging from 1200 to 1300 meters, Lubumbashi and its rural surroundings have a Cw-type climate, characterized by a rainy season (November - March) and a dry season (May - September), separated by two transitional months (April and October) [37]. While the average annual temperatures in the latter half of the 20th century ranged between 17 and 26°C [7], recent observations indicate a warming trend [38]. Annual total precipitation varies between 1200 and 1300 mm [23]. Typically established on ferralitic soils [39], the miombo woodland is the dominant vegetation unit, although its cover is constantly declining primarily due to shifting agriculture, charcoal production, and increasing urbanization [12,15,18]. The population in the Lubumbashi region remains heavily dependent on natural resources, which are increasingly depleted by shifting agriculture and charcoal production (97.9% of the population), art wood carving (1.5%), artisanal timber exploitation (0.4%), and non-timber forest product collection (0.2%; [40]. Moreover, this population predominantly lives on less than USD 1.25 per day, indicating a high level of poverty and food insecurity [41].
Additionally, the village area of Lwisha, located approximately 80 km northwest of Lubumbashi, was selected as the study site. This village area was chosen due to its identification as a site with high anthropogenic activities, particularly agriculture, charcoal production, and mining [40,42]. Furthermore, the selection was guided by the availability of both unexploited and anthropized forest habitats, specifically those affected by charcoal production (degraded forests) and agriculture (post-cultivation fallows). Moreover, the village demonstrates weak implementation of the existing simple forest management plan, contributing to deforestation and forest degradation.
2.2. Methods
2.2.1. Sampling and Data Collection
To study the composition and floral diversity, three habitat types were chosen: unexploited forests (AUF), degraded forests (DFO), and post-cultivation fallows (FAL). These habitats are illustrated and described in the table below (Table 1).
For comparison purposes, the degraded forests and post-cultivation fallows were 4 to 5 years post-exploitation, corresponding to the optimal fallow period in the Lubumbashi region [45]. These anthropized habitats were selected based on a visual analysis of high-resolution Quick Bird images available for free on Google Earth [46].
In each habitat, four transects, each 500 meters long, were established along the four cardinal points (north, south, east, west) of the village. On each transect, four floristic inventory plots measuring 50 m x 20 m, spaced 100 meters apart, were set up [6]. Additionally, to assess the regeneration of miombo woody species and thus the restoration of this forest ecosystem in anthropized habitats, 80 subplots of 10 m x 5 m each were installed in each habitat [14]. This represents 20 subplots per transect and 5 subplots per inventory plot. The dimensions of the plots and subplots were determined based on previous studies ([6,14] indicating that 50 m x 20 m and 10 m x 5 m are adequate dimensions for floristic and forest regeneration studies in the miombo woodlands, respectively.
Furthermore, in each plot, all woody individuals with a diameter at breast height (dbh) ≥ 10 cm were inventoried. The diameter of these individuals was measured using a forestry tape [31]. Additionally, in these subplots, juvenile individuals (dbh < 10 cm) were inventoried, and their diameters measured. The inventory considered three groups: seedlings (dbh < 1 cm), saplings (1 cm ≤ dbh < 10 cm), and adults (dbh ≥ 10 cm) [47]. The first two strata consist of juvenile individuals (regeneration individuals) while the last stratum represents the adult population. It should be noted that seedlings were only counted. Moreover, juveniles from coppicing were not included in the inventory. The floristic inventories for this study were conducted from March 25 to June 29, 2023. During the inventories, the identification of unknown woody species was facilitated by comparing the collected herbarium specimens with existing floras (Flora of Zambia, Flora of Zimbabwe, and World Flora), specialized books, and various identification guides [7,48,49].
2.2.2. Data Analysis
The detailed analyses in this section focused on individuals inventoried in the three strata. However, the mean square diameter and basal area were not calculated for seedlings, as their diameters were not measured during the floristic inventory. Additionally, relative frequency and density, the natural regeneration index, indicator species identification, and alpha diversity were only applied to the regeneration strata.
To ensure homogeneity among plots of different ages within each habitat, the variability in terms of density and floral diversity was tested at a 5% significance level [50]. Thus, data collected on regeneration (dbh < 10 cm) in the subplots were extrapolated to the plot level, considering the ratio between the plot area (50 m × 20 m) and the cumulative area of the subplots within each plot (5 × (10 m × 5 m)). Furthermore, to characterize the three habitats, the density (N; Equation 1), quadratic diameter (DQ; Equation 2), and basal area (G/ha; Equation 3) of the inventoried adult individuals were calculated [6,31,51]. Density expresses the number of individuals inventoried per unit area (ha), while basal area represents the surface area occupied by woody individuals on this surface [31]. Additionally, the averages of woody plant species, genera, and families were calculated for these habitats [50].
where ni is the number of individuals of a species in a plot, and a a is the area of the plot expressed in hectares.
where n is the total number of measurements taken, and is each measurement squared.
with FE, the correction factor related to the plot area (m²), m, the number of woody individuals inventoried in the plot, and gi the basal area of each measured individual, calculated using the equation below (Equation 4) [31]:
with D, the diameter at breast height (DBH) of an individual, measured at 1.30 meters above the ground.
In addition, to assess the regeneration potential (dbh < 10cm) of the habitats, the frequency (f; Equation 5), relative frequency (RF; Equation 6), and relative density (RD; Equation 7) were calculated [6]. Frequency expresses the probability of a woody species being inventoried in each of the floristic inventory plots, while relative frequency is the proportion that a given species represents compared to all inventoried species. Relative density, on the other hand, expresses the proportion that individuals of a given species represent compared to the entire population of individuals in a forest stand [14].
with n being the total number of plots where the species has been inventoried and Np the total count of plots.
with f being the frequency of a woody species and F the sum of all frequencies.
where ni is the number of individuals of a species and N is the total count of all inventoried individuals.
Additionally, the Natural Regeneration Index (NRI), defined as the ratio between the number of juvenile individuals (dbh < 10 cm) and the number of adult individuals (dbh > 10 cm) of a species, was calculated [52]. When NRI < 1, the regeneration of the species in question is low, while when NRI ≥ 1, the regeneration is high [53]. Furthermore, to determine indicator species for the habitats in these first two strata, the Indicator Species Identification (IndVal) method [50] was used. It has the advantage of combining the frequency and abundance of a species to determine its indicative nature. Thus, a plant species is considered indicative of a group of plots when it is specific (absent or relatively less frequent in other plot groups) and faithful to that group (present in all or the majority of plots). This index is calculated for each species i in each group j using the following relationship (Equation 8); [54].
with Aij being the ratio between the number of individuals ij and the number of individuals j, while Bij is the ratio between the number of plots ij and the number of plots j. In this regard, Aij is the average abundance of species i within the surveys of group j compared to all groups (a measure of specificity), while Bij is the number of surveys where species i was inventoried within the surveys of group j (a measure of fidelity or constancy).
To compare the abundance and specific diversity of habitat regeneration, the Fisher alpha index (α; Equation 9) [2,55] was calculated. Additionally, the sampling effort of woody species in the regeneration was assessed by calculating the proportion between the inventoried species in the understory (Taxa_S) and the number of species according to the floristic richness estimator (Chao 1) [50].
with S being the total number of species; Ni the total abundance of species i, and ni the number of sites or plots where species i is present.
Furthermore, to identify statistical differences at the 5% significance level among the parameters characterizing the three habitats, the non-parametric Kruskal-Wallis test was applied to the results related to density, species richness, and types of disturbances. This test was chosen due to the non-normality of the data, confirmed by the Shapiro test [56]. In case of significant differences, the Dunn-Bonferroni post-hoc test was used for pairwise comparison of means of strata or habitats [57,58].
Finally, to compare the similarity of floristic lists among different habitats, non-metric multidimensional scaling analysis (NMDS) [59] was conducted and complemented by the Jaccard index (J; equation 9) [60].
where a is the total number of woody species inventoried in two habitats for comparison; b and c respectively represent the number of woody species inventoried in one of the two habitats but absent in the other habitat.
All these analyses were conducted using R software version 4.3.2. Pairwise comparisons of floristic diversity between strata and habitats were performed using Past 4.05 software, while the alpha diversity index was calculated using EstimateS version 9.1 software.
3. Results
3.1. Characterization of the Three Habitats Along an Anthropization Gradient
A total of 1,099 adult individuals were inventoried in the three habitats: 500 individuals in the unexploited forests, 456 individuals in the degraded forests, and 143 individuals in the post-cultivation fallows. These individuals belong to 60 species (unexploited forests: 36; degraded forests: 48; post-cultivation fallows: 32), 40 genera (unexploited forests: 27; degraded forests: 33; post-cultivation fallows: 25), and 25 families (unexploited forests: 17; degraded forests: 21; post-cultivation fallows: 16, Table 2).
Furthermore, in the unexploited forests, the families Fabaceae, Phyllanthaceae, and Dipterocarpaceae alone represent 57.77% of the inventoried species, accounting for 33.33%, 11.11%, and 8.33%, respectively. These families represent 89.00% of the inventoried individuals, with the Fabaceae family alone representing 76% of individuals. Among the individuals inventoried in the unexploited forests, 54.00% belong to the species Brachystegia wangermeeana (32.60%) and B. spiciformis (21.40%). Additionally, in the degraded forests, 62.50% of species belong to the families Fabaceae (33.33%), Phyllanthaceae (10.42%), Clusiaceae, Combretaceae, and Lamiaceae (6.25% each). The Fabaceae family is the most represented with 59.87% of inventoried individuals. In these degraded forests, 58.33% of individuals belong to the species B. wangermeeana (18.20%), B. spiciformis (12.28%), Albizia adianthifolia (10.53%), Diplorhynchus condylocarpon (6.58%), Uapaca kirkiana (6.14%), and Baphia bequaertii (4.61%). Finally, in the post-cultivation fallows, 65.63% of species belong to the families Fabaceae (34.38%), Phyllanthaceae (12.50%), Combretaceae, Loganiaceae, and Malvaceae (6.25% each). The Fabaceae family represents 67.83% of inventoried individuals in the post-cultivation fallows. 44.06% of inventoried individuals in the post-cultivation fallows belong to the species B. wangermeeana (21.68%), B. spiciformis, and A. adianthifolia (11.19% each). From these results, it is evident that the families Fabaceae and Phyllanthaceae are the most represented in the adult populations of these three habitats.
In general, the results regarding dendrometric and floristic parameters reveal significant differences between habitats. Specifically, the mean values of these dendrometric parameters are higher in the unexploited forests (Density: 312.50 individuals/ha; Mean diameter: 40.75 cm; Basal area: 16.78 m²/ha) and degraded forests (Density: 285.00 individuals/ha; Mean diameter: 32.57 cm; Basal area: 9.98 m²/ha) compared to the post-cultivation fallows (Density: 89.38 individuals/ha; Mean diameter: 28.84 cm; Basal area: 1.92 m²/ha). Moreover, floristic diversity is higher in the unexploited forests (10 species; 8 genera; 5 families) and degraded forests (12 species; 10 genera; 6 families) particularly (Table 3). These results suggest a significant influence of anthropization on dendrometric parameters of individuals and floristic diversity in these habitats.
3.2. Natural Regeneration of Miombo Woody Species in the Three Habitats
A total of 23,052 regeneration individuals, comprising 7,576 juveniles in unexploited forests, 9,044 juveniles in degraded forests, and 6,432 juveniles in post-cultivation fallows, were inventoried. The juveniles were distributed among 82 species (unexploited forests: 59; degraded forests: 69; post-cultivation fallows: 70), 59 genera (unexploited forests: 43; degraded forests: 48; post-cultivation fallows: 53), and 30 families (unexploited forests: 25; degraded forests: 25; post-cultivation fallows: 27).
In unexploited forests, seedlings (dbh<1 cm) were distributed among 54 species, 40 genera, and 25 families, while saplings (1 cm≤dbh<10 cm) were distributed among 43 species, 32 genera, and 19 families. Moreover, in degraded forests, seedlings belonged to 63 species, 43 genera, and 25 families, while saplings grouped into 44 species, 32 genera, and 19 families. In post-cultivation fallows, seedlings were grouped into 58 species, 44 genera, and 25 families, while saplings grouped into 53 species, 41 genera, and 21 families. Additionally, in unexploited forests, seedlings represented 87.49%, while saplings represented only 12.51%. In degraded forests and post-cultivation fallows, seedlings represented 73.64% and 72.70%, respectively. Furthermore, saplings represented only 14.30%, 35.80%, and 37.55% of seedlings in each habitat.
Moreover, in unexploited forests, 42.37% of inventoried species belonged to the Fabaceae and Phyllanthaceae families, with Fabaceae being the most represented at 30.51% of inventoried species. They represented 42.59% of seedlings and 46.51% of saplings. These two families accounted for 70.80% of inventoried juvenile individuals in unexploited forests, comprising 71.62% of seedlings and 65.40% of saplings. In degraded forests, the Fabaceae and Phyllanthaceae families represented 44.93% of inventoried woody species (dbh<1 cm: 42.86% and 1 cm≤dbh<10 cm: 47.73% of species) and 70.44% of inventoried juvenile individuals (dbh<1 cm: 70.19% and 1 cm≤dbh<10 cm: 71.14% of individuals). Furthermore, in post-cultivation fallows, the Fabaceae and Phyllanthaceae families represented 38.57% of inventoried species (dbh<1 cm: 37.93% and 1 cm≤dbh<10 cm: 43.40% of species) and 70.15% of inventoried juvenile individuals (dbh<1 cm: 69.25% and 1 cm≤dbh<10 cm: 72.52% of individuals).
Furthermore, for seedlings (dbh<1 cm), the relative frequency of woody species ranged from 0.35% to 5.56% (mean: 1.85 ± 1.55) in unexploited forests, while the relative density of these juvenile individuals varied from 0.06% to 12.61% (mean: 1.85 ± 2.91). The natural regeneration index ranged from 0.29 to 296.00 (mean: 25.48 ± 51.45). Regarding degraded forests, the relative frequency of woody species ranged from 0.31% to 4.93% (mean: 1.64 ± 1.33). The relative density of these species in degraded forests ranged from 0.06% to 20.48% (mean: 1.61 ± 3.27), and the natural regeneration index varied from 0.73 to 252.00 (mean: 28.12 ± 43.49). As for post-cultivation fallow lands, the relative frequency ranged from 0.39% to 6.29% (mean: 1.75 ± 1.65), while the relative density varied between 0.09% and 27.89% (mean: 1.75 ± 4.03). The natural regeneration index for these woody species ranged from 1.33 to 152.00 (mean: 33.67 ± 34.04).
In contrast, for saplings (1 cm≤dbh<10 cm), the relative frequency of species ranged from 0.69% to 10.42% (mean: 2.33 ± 2.00), while the relative density ranged from 0.42% to 17.30% (mean: 2.33 ± 3.40) in unexploited forests. The natural regeneration index for species in these unexploited forests varied from 0.87 to 24.00 (mean: 3.66 ± 4.33). In degraded forests, the relative frequency of woody species ranged from 0.52% to 6.71% (mean: 2.27 ± 1.65), while the density varied from 0.17% to 29.53% (mean: 2.27 ± 4.62). The regeneration index for species in these degraded forests ranged from 0.57 to 52.00 (mean: 8.97 ± 10.32). For post-cultivation fallows, the relative frequency ranged from 0.50% to 7.56% (mean: 1.88 ± 1.77), while the density oscillated between 0.23% and 23.46% (mean: 1.89 ± 3.56). The natural regeneration index ranged from 1.33 to 36.00 (mean: 11.57 ± 8.90).
Additionally, for the stratum of seedlings, the species Brachystegia spiciformis exhibited high frequencies across different habitats. This species showed similar relative frequencies to the species Baphia bequaertii and B. wangermeeana, respectively, in unexploited forests and degraded forests. Furthermore, the species Albizia adianthifolia, Baphia bequaertii, B. spiciformis, B. wangermeeana, and Isoberlinia angolensis represented 49.12% of seedlings inventoried in unexploited forests. In degraded forests, A. adianthifolia, B. spiciformis, and B. wangermeeana represented 42.82% of these juveniles, while these species represented 45.00% of juvenile individuals in post-cultivation fallows. In this stratum of seedlings, the species Psorospermum febrifugum, Ochna schweinfurthiana, and Rothmannia engleriana exhibited the highest natural regeneration index in unexploited forests, degraded forests, and post-cultivation fallows, respectively.
Moreover, for saplings, the species B. spiciformis exhibited a high relative frequency in unexploited forests and degraded forests, while the species B. wangermeeana showed a high relative frequency in post-cultivation fallows. Additionally, B. wangermeeana was the most represented species in all three habitats. The species B. spiciformis, B. wangermeeana, Diplorhynchus condylocarpon, and Pterocarpus angolensis represented 43.88% of inventoried juvenile individuals in unexploited forests. The species A. adianthifolia, B. bequaertii, B. spiciformis, B. wangermeeana, and D. condylocarpon represented 55.87% of juveniles inventoried in degraded forests. Furthermore, A. adianthifolia, Albizia antunesiana, B. bequaertii, B. spiciformis, B. wangermeeana, and Isoberlinia angolensis represented 56.72% of inventoried juveniles in post-cultivation fallows. Additionally, Olax obtusifolia, O. schweinfurthiana, and Albizia antunesiana exhibited the highest natural regeneration index in unexploited forests, degraded forests, and post-cultivation fallows, respectively.
Furthermore, in the seedlings’ stratum, miombo characteristic woody species (from the genera Brachystegia, Julbernardia, and Isoberlinia) represent 14.8% of total species in unexploited forests, 12.70% in degraded forests, and 10.35% in post-cultivation fallows. In the saplings’ stratum, these species account for 11.63%, 13.64%, and 15.09%, respectively. In terms of individuals, these species constitute 35.85% of seedlings in unexploited forests, 41.68% in degraded forests, and 43.63% in post-cultivation fallows. For saplings, they represent 36.71%, 44.97%, and 42.14%, respectively. The table below (Table 4) summarizes the top-five woody species for each calculated parameter in each habitat. The entire set of species studied is presented in the appendix (supplementary material 1).
However, other woody species, such as B. boehmii, were not inventoried in the adult stands of the different habitats but nevertheless have juveniles in the regeneration. These species represent 33.33% and 33.87% of the total inventoried species in the seedling’s stratum of unexploited forests and degraded forests, respectively, while they represent 52.63% in the same stratum of post-cultivation fallows. Additionally, these species represent 27.91%, 15.91%, and 39.62% of inventoried species in the sapling’s stratum of the three habitats.
Conversely, other species inventoried in the adult stands of the different habitats, such as Julbernardia globiflora, do not have seedlings (dbh<1 cm). These species, in turn, represent 10.82%, 14.58%, and 15.63% of adult stand species, respectively, in unexploited forests, degraded forests, and post-cultivation fallows. Furthermore, in the stratum of saplings, these species represent 16.22% and 22.92% in unexploited forests and degraded forests, respectively. Specifically, all species inventoried in the adult stratum of post-cultivation fallows have juveniles in the sapling’s stratum.
Moreover, in the stratum of seedlings, the species Garcinia huillensis, P. febrifugum, and Uapaca kirkiana are indicative of unexploited forests, while B. boehmii, Harungana madagascariensis, and Julbernardia paniculata are indicative of degraded forests. Phyllanthus muellerianus is the only indicative species of this stratum in post-cultivation fallows. In contrast, for saplings, the species O. obtusifolia is indicative of unexploited forests, while Diplorhynchus condylocarpon and Isoberlinia tomentosa are indicative of degraded forests. The species Pseudolachnostylis maprouneifolia is indicative of post-cultivation fallows.
The results related to the dendrometric and floristic parameters of regeneration strata are statistically similar between habitats, except for density per hectare, the number of individuals, species, and genera in seedlings stratum (Table 5), which show particularly low values in post-cultivation fallows. These results suggest that the regeneration potential is inversely proportional to the anthropogenic disturbances experienced by the habitats.
3.3. Comparison of Specific Richness of Woody Species Inventoried in Regeneration and Adult Stands
The comparison of floristic lists among different strata within each habitat reveals that the stratum of seedlings (dbh < 1 cm) notably differs from other strata, except in post-cultivation fallows. Additionally, the floristic list of saplings (1 cm ≤ dbh < 10 cm) is nearly included within that of the adult stratum in each habitat. This situation is particularly observed in unexploited and degraded forests. Conversely, in post-cultivation fallows, the floristic lists of regeneration strata, though largely distinct, are fully encompassed within that of adult individuals. Furthermore, the strata of seedlings and saplings in different habitats are increasingly similar to each other, respectively (Figure 2).
However, the Jaccard similarity of floristic lists among different strata in the three habitats is depicted in the table below (Table 6). It is evident from this table that the lowest similarity is between the floristic list of seedlings in degraded forests and that of adults in post-cultivation fallows (42.00%), while the highest similarity is between the floristic lists of installed juveniles in unexploited forests and those in degraded forests (92%). Nonetheless, pairwise comparison of floristic lists of habitats (all strata combined) reveals that the floristic lists of unexploited and degraded forests exhibit a similarity of 76.50%, whereas that of post-cultivation fallows is 56.00% similar to unexploited and degraded forests, respectively. These results indicate that the floristic lists of habitats are influenced by natural factors (intra- and interspecific competition) and, particularly, by the extent of disturbances experienced by the habitats.
4. Discussion
4.1. Structure and Floristic Composition of Forest Strata and Stands Along the Anthropization Gradient
Dendrometric (density) and floristic parameters decrease according to the level of habitat disturbance (Table 2 and Table 3). This situation is attributed to both natural and particularly anthropogenic disturbances experienced by the human-modified habitats, primarily agriculture and charcoal production. Indeed, the conversion of forested lands into agroecosystems and charcoal production leads to deforestation and fragmentation of the miombo woodlands, particularly in the Lubumbashi region [15]. According to Refs. [18,61], anthropogenic activities, often carried out through rudimentary techniques, disrupt the ecological balance of ecosystems, subsequently affecting the structure and composition of the miombo woodlands. These results, found in the present study, corroborate those of other research conducted in the miombo ecoregion [2,14,21,35,50,62], showing that habitat floristic composition and diversity decline as anthropogenic disturbances increase.
Furthermore, the results of the present study were obtained through floristic inventories conducted on plots along a transect. This method has the advantage of reducing errors in alignment, angles, and width measurements that may arise when setting up continuous plots. Additionally, other methods, such as random sampling or systematic inventories, can be used and produce reliable results. However, these methods require more time and a larger team to conduct the floristic inventory, despite producing indisputable results [63]. Moreover, a square (rectangular) shape was used for inventory plots in the present study. This shape, for the same area, has the advantage of enumerating more individuals than other shapes and is most recommended in the open and dense forests of the tropical region [64,65]. However, the challenges of this shape lie in measuring distances, angles, and selecting their orientation when installing the plot. Additionally, this square shape often encounters numerous cases of uncertainty regarding whether border trees belong to the plot or not. Therefore, the circular shape would undoubtedly be more commonly used. This latter shape is the most objective in terms of measurements and results, as its installation does not require a preferred direction, thereby overcoming the challenges of the square shape as long as the circular plot is not too large [31]. Nevertheless, the inventory method, shape, and dimensions of the plots used in the present study have been employed by several authors in regeneration studies in the miombo ecoregion [6,14], justifying the choice of this methodology in the present study.
4.2. Regeneration of Miombo Woody Species Along the Anthropization Gradient
The regeneration potential (individual number and diversity) of habitats within the two regeneration strata remains higher in degraded forests than in unexploited forests and post-cultivation fallows (Table 4 and Table 5). These results demonstrate that the regeneration of woody species and subsequently the reconstitution of the miombo woodlands would be possible in anthropized habitats provided that human activities, especially agriculture, dendro-energy production, wood cutting, and bushfires, are prohibited. Additionally, these measures must be increasingly enforced in heavily disturbed habitats, such as post-cultivation fallows. The similarity in floristic richness between degraded and unexploited forests could be explained by the fact that the anthropogenic disturbances experienced by degraded forests, particularly related to the decrease in individual density through selective harvesting, make resources available in these habitats, such as space, water, and insolation [66]. This resource availability, coupled with low inter- and intraspecific competition, allows woody species, particularly pioneer and resilient species to anthropogenic disturbances (present in the herbaceous stratum and in the soil seed bank), to establish themselves [57]. In contrast to degraded forests, anthropogenic disturbances in agroecosystems are not only related to the loss of woody species density (tree cutting and stump removal) but also to the disruption of soil physicochemical and biological properties [19]. The combination of these disturbances negatively affects the regeneration potential and resilience capacity of woody species, potentially leading to savannization [67]. Furthermore, the regeneration potential in unexploited forests would depend on strong inter- and intraspecific competition for the aforementioned resources, primary factors in the establishment of plant species in habitats [68,69]. These results corroborate previous research [14,51,70,71] indicating that anthropized habitats exhibit high species richness in regeneration strata. These anthropized habitats are characterized by high environmental heterogeneity during early succession stages and high regeneration potential of miombo woody species [6]. However, these results do not support the findings of studies conducted, notably in Zimbabwe by [35], indicating that species richness is high in unexploited forests due to the absence of anthropogenic disturbances on a human scale. Nevertheless, the regeneration potential in unexploited forests depends on several factors, including inter/intra-specific competition for resources and minimal anthropogenic disturbances. Additionally, the vigor of adult trees (producing quality and sufficient seeds), the presence of animal species (facilitating seed dispersal), symbiotic interactions, good soil structure, and high nutrient availability also play crucial roles. These factors interact in complex ways, influencing the regeneration process in unexploited forests [68,72].
However, the impact of anthropogenic disturbances on species diversity in degraded forests is not systematically negative. Indeed, these disturbances could create new environmental conditions that sometimes favor increased plant diversity if these anthropogenic disturbances are of low intensity, limited duration, and characterized by minimal removal [73]. This situation has already been highlighted in previous studies conducted in the miombo ecoregion [34,74], demonstrating that floristic characteristics such as stand structure and species richness of anthropized habitats can reach values higher than those of unexploited mature forests after anthropogenic disturbances cease. Moreover, according to these same authors, these recorded disturbances have a long-term effect on the structure of populations and the specific richness of these habitats, keeping these parameters less similar compared to intact miombo woodlands after 20 to 35 years of regeneration [35].
Conversely, maintaining human pressure on natural resources even in heavily anthropized habitats compromises the regeneration of woody species and subsequently the reconstitution of the miombo woodlands, particularly in vulnerable anthropized habitats. Indeed, the distance from intact forests over an increasingly extensive radius [12] has led local communities to harvest woody species, furthermore through less sustainable practices [75], in anthropized areas near settlements for various needs [6]. In addition to this, late and repetitive bushfires [76] characterizing the miombo ecoregion [44,77-79] and particularly the Lubumbashi region [80], affect the natural regeneration of woody species in habitats. Moreover, the scarcity of species with high calorific value and the increase in charcoal production distance, in particular, induce the return of local communities to regenerating forest stands. This situation contributes to maintaining a high level of forest degradation [81] and decreases the potential for forest regeneration. Similarly, population growth and increased land pressure resulting from it have led local communities to shorten fallow periods to meet increased demand for basic necessities [45]. This further disrupts the process of woody species regeneration and ongoing miombo woodlands reconstitution in post-cultivation fallows [82]. These results are similar to those of studies conducted in Mozambique [6], showing that ongoing human activities in already anthropized habitats compromise the reconstitution of the miombo woodlands in these habitats.
4.3. Similarity between Floristic Lists Along the Anthropization Gradient
The floristic lists of unexploited forests and degraded forests exhibit higher similarities compared to post-cultivation fallows (Figure 2 and Table 6). This situation is attributed to the fact that during exploitation, agrosystems transitioning into post-cultivation fallows undergo anthropogenic disturbances that negatively impact dendrometric and floristic parameters, particularly. These findings corroborate studies conducted in the dense humid forest region [50,83], and specifically in the miombo ecoregion [6,35], demonstrating that anthropogenic activities in agrosystems negatively influence floristic diversity in post-cultivation fallows.
4.4. Implications for Sustainable Miombo Woodlands Restoration in Anthropized Landscapes
Anthropogenic disturbances affect the high potential for natural regeneration and resilience of different miombo woody species. To address this, Assisted Natural Regeneration (ANR) could be one solution. ANR involves deliberate protection of disturbed habitats against anthropogenic pressures and invasive plant species to accelerate natural forest succession processes leading to the reconstitution of a resilient and productive ecosystem [5,26]. It requires legislative reform and rules governing interactions between natural and social dynamics [84]. However, in the D.R. Congo, this reform would focus on access to natural resources, establishing reasonable rotation periods, regulating bushfires, and anthropogenic incursions into habitats at the end of their exploitation. ANR could be effective and less costly than reforestation and other revegetation strategies, provided there are seed sources in the restoration area [5]. This restoration technique has been successfully used in Ethiopia to restore forests over significant areas previously impacted by anthropogenic activities and has been proposed in Mozambique to restore miombo woodlands in anthropized habitats [5,74]. However, in regions with rapid population growth like the Lubumbashi region, implementing ANR can be challenging due to increased anthropogenic pressures on land and natural resources. To address this, zoning and defined collaborative restoration options involving local communities actively would be one solution to this situation [27,40].
Furthermore, reforestation and enrichment would be palliative solutions for anthropized habitats with low miombo resilience capacity after exploitation, such as post-cultivation fallows. Utilizing miombo woody species for reforestation and habitat enrichment would result in forest ecosystems with a structure, specific composition, and function almost similar to the previously exploited forest. In this regard, these restored habitats would continue to support the survival of both rural and urban populations by providing usual ecosystem services [85]. However, selecting fast-growing native species like Pterocarpus tinctorius Welw. and Combretum collinum Fresen. is necessary for short-term miombo woodlands reconstitution [86]. Nevertheless, similar to ANR habitats, reforested or enriched habitats should be protected from anthropogenic intrusions [6,35] and late, repetitive bushfires [80,87].
Furthermore, this study shows the current state of anthropized habitats regarding regeneration potential and subsequent forest reconstitution. However, it does not depict the successional dynamics of woody species in these habitats over the years following exploitation [21,35,51]. Additionally, the study does not show the distribution of these woody species based on their functional traits within different strata and habitats [50]. These missing ecological aspects would provide complementary information to the present results and remain important for the establishment of sustainable miombo woodlands management strategies.
5. Conclusions
The present study assessed the natural regeneration capacity of miombo woody species along a gradient of anthropization through floristic inventories in three different habitats, including one unexploited and two with varying levels of anthropization. The results confirm that density, mean diameter, basal area, taxa, genera, and families have high values in both unexploited and degraded forests. Indeed, significant differences were observed among the three habitats, with low values observed in fallows. Furthermore, these results confirm that the regeneration potential of miombo species and individuals’ number are high in degraded forests. Although low regeneration was observed in unexploited forests and post-cultivation fallows, except in the sapling’s stratum where regeneration in different habitats is almost equivalent. Additionally, our results indicate that there are similarities and dissimilarities in terms of floristic richness between habitats, as the floristic lists of unexploited forests and degraded forests show higher similarities than post-cultivation fallows. While our study did not characterize variations in dendrometric and floristic parameters according to the age of anthropized habitats, as well as the distribution of species in strata and habitats in terms of functional traits of woody species, our results show that the regeneration potential of miombo species depends on the intensity of anthropogenic disturbances experienced by habitats. To contribute to the regeneration of woody species and the reconstitution of miombo in anthropized habitats, appropriate legislation determining rotation periods and regulating repetitive bushfires and anthropogenic activities should be established. Additionally, inclusive reforestation and agroforestry activities using miombo woody species should be considered.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1
Funding
The research was funded by the project CHARLU (ARES-CCD, Belgium).
Data Availability Statement
The data related to the present study will be available upon request from the interested party.
Acknowledgments
The authors acknowledge the Academy of Research and Higher Education (ARES) and the Research Project for Development: "Strengthening the capacity for sustainable management of miombo woodlands by assessing the environmental impact of charcoal production and improving practices towards forest resources (PRD CHARLU)" for financial support for this study through the doctoral scholarship awarded to Dieu-donné N’tambwe Nghonda and Héritier Khoji Muteya.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Aerts, R. & Honnay, O. Forest restoration, biodiversity and ecosystem functioning. BMC Ecol. 2011, 11, 29. [CrossRef]
- Kalaba, F.K.; Quinn, C.H.; Dougill, A.J. & Vinya, R. Floristic composition, species diversity and carbon storage in charcoal and agriculture fallows and management implications in Miombo woodlands of Zambia. For. Ecol. Manag. 2013, 304, 99–109. [CrossRef]
- FAO. La Situation des forêts du monde 2022. Des solutions forestières pour une relance verte et des économies inclusives, résilientes et durables. FAO, Rome, Italie, 2022 ; 180 p. [CrossRef]
- Gonçalves, F.M.P.; Chisingui, A.V.; Luís, J.C.; Rafael, M.F.F.; Tchamba, J.J.; Cachissapa, M.J.; Caluvino, I.M.C.; Bambi, B.R.; Alexandre, J.L.M.; Chissingui, M.D.G.; Manuel, S.K.A.; Jacinto, H.D.; Finckh, M.; Meller, P.; Jürgens, N. & Revermann, R. First vegetation-plot database of woody species from Huíla province, SW Angola. VCS 2021, 2, 109–116. [CrossRef]
- Berrahmouni, N.; Regato, P. & Parfondry, M. Global guidelines for the restoration of degraded forests and landscapes in drylands: building resilience and benefiting livelihoods. FAO, Rome, Italie, 2015; 173 p.
- Ameja, L.G.; Ribeiro, N.S.; Sitoe, A. & Guillot, B. Regeneration and Restoration Status of Miombo Woodland Following Land Use Land Cover Changes at the Buffer Zone of Gile National Park’s Central Mozambique. Trees, For. People 2022, 9, 100290. [CrossRef]
- Malaisse, F. How to live and survive in Zambezian open forest (Miombo ecoregion). Les Presses Agronomiques de Gembloux, Gembloux, Belgique, 2010; 424 p.
- Chidumayo, E.N. Management implications of tree growth patterns in miombo woodlands of Zambia. For. Ecol. Manag. 2019, 436, 105–116. [CrossRef]
- Chirwa, P.W.; Larwanou, M.; Syampungani, S. & Babalola, F.D. Management and restoration practices in degraded landscapes of Eastern Africa and requirements for up-scaling. Int. For. Rev. 2015a, 17, 20–30. [CrossRef]
- Mittermeier, R.A.; Mittermeier, C.G.; Brooks, T.M.; Pilgrim, J.D.; Konstant, W.R.; Da Fonseca, G.A.B. & Kormos, C. Wilderness and biodiversity conservation. PNAS USA 2003, 100, 10309–10313. [CrossRef]
- Godlee, J.L.; Gonçalves, F.M.; Tchamba, J.J.; Chisingui, A.V.; Muledi, J.I.; Shutcha, M.N.; Ryan, C.M.; Brade, T.K. & Dexter, K.G. Diversity and Structure of an Arid Woodland in Southwest Angola, with Comparison to the Wider Miombo Ecoregion. Diversity 2020, 12, 140. [CrossRef]
- Useni, S.Y. ; Malaisse, F. ; Cabala, K.S. ; Munyemba, K.F. & Bogaert, J. Le rayon de déforestation autour de la ville de Lubumbashi (Haut-Katanga, RD Congo) : Synthèse. Tropicultura 2017, 35(3), 215–221. http://hdl.handle.net/2268/227664.
- Schneibel, A.; Stellmes, M.; Röder, A.; Finckh, M.; Revermann, R.; Frantz, D. & Hill, J. Evaluating the trade-off between food and timber resulting from the conversion of Miombo forests to agricultural land in Angola using multi-temporal Landsat data. Sci. Total Environ. 2016, 548–549, 390–401. [CrossRef]
- Gonçalves, F.M.P.; Revermann, R.; Cachissapa, M.J.; Gomes, A.L. & Aidar, M.P.M. Species diversity, population structure and regeneration of woody species in fallows and mature stands of tropical woodlands of southeast Angola. J. For. Res. 2018, 29, 1569–1579. [CrossRef]
- Khoji, M.H.; N’tambwe, N.D.; Mwamba, K.F.; Harold, S.; Munyemba, K.F.; Malaisse, F.; Bastin, J.-F.; Useni, S.Y. & Bogaert, J. Mapping and Quantification of Miombo Deforestation in the Lubumbashi Charcoal Production Basin (DR Congo): Spatial Extent and Changes between 1990 and 2022. Land 2023, 12, 1852. [CrossRef]
- Nansikombi, H.; Fischer, R.; Ferrer Velasco, R.; Lippe, M.; Kalaba, F.K.; Kabwe, G. & Günter, S. Can de facto governance influence deforestation drivers in the Zambian Miombo? For. Policy Econ. 2020, 120, 102309. [CrossRef]
- Kyale, K.J.; Wardell, D.A.; Mikwa, J.-F.; Kabuanga, J.M.; Monga Ngonga, A.M.; Oszwald, J. & Doumenge, C. Dynamique de la déforestation dans la Réserve de biosphère de Yangambi (République démocratique du Congo) : variabilité spatiale et temporelle au cours des 30 dernières années. Bois et For. Trop. 2019, 341, 15–28. [CrossRef]
- Cabala, K.S. ; Useni, S.Y. ; Amisi, M.Y.A. ; Munyemba, K.F. & Bogaert, J. Activités anthropiques et dynamique des écosystèmes forestiers dans les zones territoriales de l’Arc Cuprifère Katangais (RD Congo). Tropicultura 2022, 40(3/4), 2100 . [CrossRef]
- Reyniers, C. Agroforesterie et déforestation en République démocratique du Congo. Miracle ou mirage environnemental ? Mondes dév. 2019, 187, 113-132. [CrossRef]
- Abdourhamane, H. ; Morou, B. ; Rabiou, H. & Amhamane, A. Caractéristiques floristiques, diversité et structure de la végétation ligneuse dans le Centre-Sud du Niger : cas du complexe des forêts classées de Dan kada Dodo-Dan Gado. Int. j. biol. Chem. Sci. 2013, 7, 1048. [CrossRef]
- Syampungani, S.; Geldenhuys, C.J. & Chirwa, P.W. Regeneration dynamics of miombo woodland in response to different anthropogenic disturbances: forest characterisation for sustainable management. Agrofor. Syst. 2016, 90, 563–576. [CrossRef]
- Eba’a Atyi, R. ; Hiol Hiol, F. ; Lescuyer, G. ; Mayaux, P. ; Defourny, P. ; Bayol, N. ; Saracco, F. ; Pokem, D. ; Sufo Kankeu, R. & Nasi, R. Les forêts du bassin du Congo : état des forêts 2021. CIFOR, Bogor, Indonésie, 2022 ; 474 p. [CrossRef]
- Malaisse, F. ; Bogaert, J. ; Boisson, S. & Sikuzani, Y.U. La végétation naturelle d’Élisabethville (actuellement Lubumbashi) au début et au milieu du XXième siècle. Géo-Eco-Trop. 2021, 45(1), 41–51.
- Potapov, P.V.; Turubanova, S.A.; Hansen, M.C.; Adusei, B.; Broich, M.; Altstatt, A.; Mane, L. & Justice, C.O. Quantifying forest cover loss in Democratic Republic of the Congo, 2000–2010, with Landsat ETM+ data. Remote Sens. Environ. 2012, 122, 106–116. [CrossRef]
- Holl, K.D. & Aide, T.M. When and where to actively restore ecosystems? For. Ecol. Manag. 2011, 261, 1558–1563. [CrossRef]
- Rinaudo, T. ; Muller, A. & Morris, M. Manuel La Régénération Naturelle Assistée (RNA). Une ressource pour les gestionnaires de projets, les utilisateurs et tous ceux qui ont un intérêt à mieux comprendre et soutenir le mouvement pour la RNA. FMNR Hub, World Vision Australia, 2020 ; 241p.
- Awono, A. ; Assembe-Mvondo, S. ; Tsanga, R. ; Guizol, P. & Peroches, A. Restauration des paysages forestiers et régimes fonciers au Cameroun : Acquis et handicaps. Document Occasionnel 10. Bogor, Indonésie : Centre de recherche forestière internationale (CIFOR) ; et Nairobi, Kenya : Centre international de recherche en agroforesterie (ICRAF), 2023 ; 43 p. [CrossRef]
- Ramade, F. Eléments d’écologie. Ecologie appliquée : action de l’homme sur la biosphère. 7e édition, Dunod, France, 2012 ; 791p.
- Useni, S.Y.; Mpibwe, K.A.; Yona, M.J.; N’tambwe, N.D.; Malaisse, F. & Bogaert, J. Assessment of Street Tree Diversity, Structure and Protection in Planned and Unplanned Neighborhoods of Lubumbashi City (DR Congo). Sustainability 2022, 14, 3830. [CrossRef]
- Larson, J. E. & Funk, J. L. Regeneration: an overlooked aspect of trait-based plant community assembly models. J. Ecol. 2016, 104(5), 1284–1298.
- Rondeux, J. La mesure des arbres et des peuplements forestiers. 3ème édition. Les Presses Universitaires de Liège – Agronomie – Gembloux, Gembloux, Belgique, 2021 ; 738p. http://hdl.handle.net/2268/262622.
- Syampungani, S.; Tigabu, M.; Matakala, N.; Handavu, F. & Oden, P.C. Coppicing ability of dry miombo woodland species harvested for traditional charcoal production in Zambia: a win–win strategy for sustaining rural livelihoods and recovering a woodland ecosystem. J. For. Res. 2017, 28, 549–556. [CrossRef]
- Sangeda, A.Z. & Maleko, D.D. Regeneration Effectiveness Post Tree Harvesting in Natural Miombo Woodlands, Tanzania. J. Plant Sci. Agri. Res. 2018, 2(1), 10. http://www.imedpub.com/plant-sciences-and-agricultural-research/.
- Muvengwi, J.; Chisango, T.; Mpakairi, K.; Mbiba, M. & Witkowski, E.T.F. Structure, composition and regeneration of miombo woodlands within harvested and unharvested areas. For. Ecol. Manag. 2020, 458, 117792. [CrossRef]
- Montfort, F.; Nourtier, M.; Grinand, C.; Maneau, S.; Mercier, C.; Roelens, J.-B. & Blanc, L. Regeneration capacities of woody species biodiversity and soil properties in Miombo woodland after slash-and-burn agriculture in Mozambique. For. Ecol. Manag. 2021, 488, 119039. [CrossRef]
- Picard, N. & Gourlet-Fleury, S. Manuel de référence pour l’installation de dispositifs permanents en forêt de production dans le Bassin du Congo. COMIFAC, Yaoundé, Cameroun, 2008 ; 265 p. http://hal.cirad.fr/cirad-00339816.
- Mutondo, G.T. ; Kamutanda, D.K. & Numbi, A.M. Evaluation du bilan hydrique dans les milieux anthropisés de la forêt claire (région de Lubumbashi, Province du Haut-Katanga, R.D. Congo). Méthodologie adoptée pour l’estimation de l’évapotranspiration potentielle. Geo-Eco-Trop. 2018, 42, 1, 159–172.
- Kalombo, K. D. Évolution des éléments du climat en RDC : Stratégies d'adaptation des communautés de base, face aux événements climatiques de plus en plus fréquents. Éditions universitaires européennes, Sarrebruck, Germany, 2016 ; 220 p.
- Ngongo, M. L. ; Van Ranst, E. ; Baert, G. ; Kasongo, E. L. ; Verdoodt, A. ; Mujinya, B. B. & Mukalay, J. M. Guide des sols en République Démocratique du Congo, tome I : étude et gestion. Ed. Salama, Lubumbashi, République Démocratique du Congo, 2009 ; 260 p.
- N’tambwe, D.N.; Khoji, M.H.; Kasongo, K.B.; Kouagou, S.R.; Malaisse, F.; Useni, S.Y.; Masengo, K.W. & Bogaert, J. Towards an Inclusive Approach to Forest Management: Highlight of the Perception and Participation of Local Communities in the Management of miombo Woodlands around Lubumbashi (Haut-Katanga, D.R. Congo). Forests 2023a, 14, 687. [CrossRef]
- Cadre Intégré de Classification de la sécurité Alimentaire (IPC). Aperçu de l’Insécurité Alimentaire Aiguë de l’IPC. Kinshasa, République Démocratique du Congo, 2023.
- Khoji, M.H.; N’tambwe, N.D.; Malaisse, F.; Waselin, S.; Kouagou, R.S.; Cabala, K.S.; Munyemba, F.M.; Bastin, J.-F.; Bogaert, J. & Useni, S.Y. Quantification and Simulation of Landscape Anthropization around the Mining Agglomerations of Southeastern Katanga (DR Congo) between 1979 and 2090. Land 2022, 11, 850. [CrossRef]
- Ribeiro, N.S.; Katerere, Y.; Chirwa, P.W.; Grundy, I.M. Miombo Woodlands in a Changing Environment: Securing the Resilience and Sustainability of People and Woodlands. Springer International Publishing, Cham, Switzerland, 2020, 269 p. [CrossRef]
- Buramuge, V.A.; Ribeiro, N.S.; Olsson, L. & Bandeira, R.R. Exploring Spatial Distributions of Land Use and Land Cover Change in Fire-Affected Areas of Miombo Woodlands of the Beira Corridor, Central Mozambique. Fire 2023a, 6, 77. [CrossRef]
- Bolakonga, I.A.B. ; Nkulu, M.F.J. & Mushakulwa, W. Filières en République Démocratique du Congo : Maïs, riz, bananes plantains et pêche. Konrad Adenauer Stiftung, Kinshasa, République Démocratique du Congo, 2017 ; 321 p.
- Kalawu, S.M.; Ngoy, M.K.; Ombeni, I.; Mane, L. & Claude, P. Mapping the stratification of vegetation classes in the Miombo forests and assessing the accuracy of their classification in Katanga province in the Democratic Republic of the Congo. Int. j. sci. eng. Res. 2022, 13(2), 770–785.
- Ding, Y.; Zang, R.; Lu, X. & Huang, J. The impacts of selective logging and clear-cutting on woody plant diversity after 40 years of natural recovery in a tropical montane rain forest, south China. Sci. Total Environ. 2017, 579, 1683–1691. [CrossRef]
- Meerts, P.J. & Hasson, M. Arbres et Arbustes du Haut-Katanga. Editions Jardin Botanique de Meise: Brussels, Belgium, 2016 ; 386 p.
- Vollesen, K. & Merrett, L. A Photo Rich Field Guide to the (Wetter) Zambian Miombo Woodland: Based on Plants from the Mutinondo Wilderness Area, Northern Zambia. Ed. Oxford, Lusaka, Zambia, 2020; 1200 p.
- Zébazé, D.; Gorel, A.; Gillet, J.-F.; Houngbégnon, F.; Barbier, N.; Ligot, G.; Lhoest, S.; Kamdem, G.; Libalah, M.; Droissart, V.; Sonké, B. & Doucet, J.-L. Natural regeneration in tropical forests along a disturbance gradient in South-East Cameroon. For. Ecol. Manag. 2023, 547, 121402. [CrossRef]
- Gonçalves, F.M.P.; Revermann, R.; Gomes, A.L.; Aidar, M.P.M.; Finckh, M. & Juergens, N. Tree Species Diversity and Composition of Miombo Woodlands in South-Central Angola: A Chronosequence of Forest Recovery after Shifting Cultivation. Int. J. For. Res. 2017, 2017, 1–13. [CrossRef]
- Hakizimana, P. ; Bangirinama, F. ; Havyarimana, F. ; Habonimana, B. & Bogaert, J. Analyse de l’effet de la structure spatiale des arbres sur la régénération naturelle de la forêt claire de Rumonge au Burundi. Bull. sci. Inst. Natl. Environ. Conserv. Nat. 2011, 9, 46–52.
- Melingui, J.B.N. ; Angoni, H. ; Claude, P.A. & Kono, L. Potentiel De Régénération Naturelle De Quelques Produits Forestiers Non Ligneux Prioritaires Dans Le Bassin De Production D’akom II (Sud Cameroun). World Wide j. multidiscip. Res. Dev. 2017, 4(2), 214–224. https://www.researchgate.net/publication/329245697.
- Dufrêne, M. & Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67(3), 345–366. https://www.jstor.org/stable/2963459.
- Colwell, R.K. & Elsensohn, J.E. EstimateS turns 20: statistical estimation of species richness and shared species from samples, with non-parametric extrapolation. Ecography 2014, 37, 609–613. [CrossRef]
- Razali, N.M. & Wah, Y.B. Power comparisons of Shapiro-Wilk, Kolmogorov-Smirnov, Lilliefors and Anderson-Darling tests. J. Stat. Model. and Anal. 2011, 2(1), 21-33.
- Gupta, B. & Mishra, T.K. Analysis of tree diversity and factors affecting natural regeneration in fragmented dry deciduous forests of lateritic West Bengal. Trop. Ecol. 2019, 60, 405–414. [CrossRef]
- Heinken, T.; Diekmann, M.; Liira, J.; Orczewska, A.; Schmidt, M.; Brunet, J.; Chytrý, M.; Chabrerie, O.; Decocq, G.; De Frenne, P.; Dřevojan, P.; Dzwonko, Z.; Ewald, J.; Feilberg, J.; Graae, B.J.; Grytnes, J.; Hermy, M.; Kriebitzsch, W.; Laiviņš, M.; Lenoir, J.; Lindmo, S.; Marage, D.; Marozas, V.; Niemeyer, T.; Paal, J.; Pyšek, P.; Roosaluste, E.; Sádlo, J.; Schaminée, J.H.J.; Tyler, T.; Verheyen, K.; Wulf, M. & Vanneste, T. The European Forest Plant Species List (EuForPlant): Concept and applications. J. Veg. Sci. 2022, 33, e13132. [CrossRef]
- Sawada, Y.; Imai, N.; Takeshige, R. & Kitayama, K. Relationships between tree-community composition and regeneration potential of Shorea trees in logged-over tropical rain forests. J. For. Res. 2022, 27, 222–229. [CrossRef]
- Albatineh, A.N. & Niewiadomska-Bugaj, M. Correcting Jaccard and other similarity indices for chance agreement in cluster analysis. Adv. Data Anal. Classi. 2011, 5, 179–200. [CrossRef]
- Useni, S.Y. ; Boisson, S. ; Cabala, K.S. ; Khonde, C.N. ; Malaisse, F. ; Halleux, J.-M. ; Bogaert, J. & Kankumbi, F.M. Dynamique de l’occupation du sol autour des sites miniers le long du gradient urbain-rural de la ville de Lubumbashi, RD Congo. Biotechnol. Agron. Soc. Environ. 2020, 24(1), 1-14. https://popups.uliege.be/1780-4507/index.php?id=18306.
- Kissanga, R.; Catarino, L.; Máguas, C.; Cabral, A.I.R. & Chozas, S. Assessing the Impact of Charcoal Production on Southern Angolan Miombo and Mopane Woodlands. Forests 2023, 15, 78. [CrossRef]
- Thiombiano, A.; Glele kakaï, R.; Bayen, P.; Boussim, J.I. & Mahamane, A. Méthodes et Dispositifs d’inventaires Forestiers en Afrique de l’Ouest : État des Lieux et Propositions Pour une Harmonisation. Ann. Sci. Agron. 2016, 20, 15–31.
- Houeto, G.; Glele Kakaï, R.; Salako, V.; Fandohan, B.; Assogbadjo, A.E.; Sinsin, B. & Palm, R. Effect of inventory plot patterns in the floristic analysis of tropical woodland and dense forest. Afr. J. Ecol. 2013, 52(3),257–264. [CrossRef]
- Salako, V. K.; Glele Kakaï, R. L.; Assogbadjo, A. E.; Fandohan, B.; Houinato, M. & Palm, R. Efficiency of inventory plot patterns in quantitative analysis of vegetation: a case study of tropical woodland and dense forest in Benin. South. For. 2013, 75(3), 137–143. [CrossRef]
- Chinder, G.B.; Hattas, D. & Massad, T.J. Growth and functional traits of Julbernardia globiflora (Benth) resprouts and seedlings in response to fire frequency and herbivory in miombo woodlands. S. Afr. J. Bot. 2020, 135, 476–483. [CrossRef]
- Useni, S.Y.; Khoji, M.H.; Langunu, S.; Gerardy, A. & Bogaert, J. Amplification of Anthropogenic Pressure Heavily Hampers Natural Ecosystems Regeneration within the Savanization Halo Around Lubumbashi City (Democratic Republic of Congo). Int. j. environ. Sci. nat. Resour. 2019, 17(2), 555958. [CrossRef]
- Finger, C.A.G.; Costa, E.A.; Hess, A.F.; Liesenberg, V. & Bispo, P.D.C. Simulating Sustainable Forest Management Practices Using Crown Attributes: Insights for Araucaria angustifolia Trees in Southern Brazil. Forests 2023, 14, 1285. [CrossRef]
- Pretzsch, H.; Del Río, M.; Arcangeli, C.; Bielak, K.; Dudzinska, M.; Ian Forrester, D.; Kohnle, U.; Ledermann, T.; Matthews, R.; Nagel, R.; Ningre, F.; Nord-Larsen, T.; Szeligowski, H. & Biber, P. Competition-based mortality and tree losses. An essential component of net primary productivity. For. Ecol. Manag. 2023, 544, 121204. [CrossRef]
- Chidumayo, E.N. Forest degradation and recovery in a miombo woodland landscape in Zambia: 22 years of observations on permanent sample plots. For. Ecol. Manag. 2013, 291, 154–161. [CrossRef]
- Lu, H.; Mohren, G.; Del Río, M.; Schelhaas, M.-J.; Bouwman, M. & Sterck, F. Species Mixing Effects on Forest Productivity: A Case Study at Stand-, Species- and Tree-Level in the Netherlands. Forests 2018, 9, 713. [CrossRef]
- Chirwa, P.W.; Larwanou, M.; Syampungani, S. & Babalola, F.D. Management and restoration practices in degraded landscapes of Southern Africa and requirements for up-scaling. Int. For. Rev. 2015b, 17, 31–42. [CrossRef]
- Puig, H. Diversité spécifique et déforestation : l’exemple des forêts tropicales humides du Mexique. Bois For. Trop. 2001, 268(2) : 41–55.
- Montfort, F. Dynamiques des paysages forestiers au Mozambique : étude de l’écologie du Miombo pour contribuer aux stratégies de restauration des terres dégradées. Thèse de doctorat, AgroParisTech, Paris, France, 2021 ; 189p.https://hal.science/tel-03524870.
- N’tambwe, N.D.; Biloso, M.A.; Malaisse, F.; Useni, S.Y.; Masengo, K.W. & Bogaert, J. Socio-Economic Value and Availability of Plant-Based Non-Timber Forest Products (NTFPs) within the Charcoal Production Basin of the City of Lubumbashi (DR Congo). Sustainability 2023b, 15, 14943. [CrossRef]
- Ryan, C.M. & Williams, M. How does fire intensity and frequency affect miombo woodland tree populations and biomass? Ecol. Appl. 2011, 21, 48-60. [CrossRef]
- Tarimo, B.; Dick, Ø.B.; Gobakken, T. & Totland, Ø. Spatial distribution of temporal dynamics in anthropogenic fires in miombo savanna woodlands of Tanzania. Carbon Balance Manag. 2015, 10, 18. [CrossRef]
- Van Wilgen, B.W.; De Klerk, H.M.; Stellmes, M. & Archibald, S. An analysis of the recent fire regimes in the Angolan catchment of the Okavango Delta, Central Africa. Fire Ecol. 2022, 18, 13. [CrossRef]
- Buramuge, V.A.; Ribeiro, N.S.; Olsson, L.; Bandeira, R.R. & Lisboa, S.N. Tree Species Composition and Diversity in Fire-Affected Areas of Miombo Woodlands, Central Mozambique. Fire 2023b, 6, 26. [CrossRef]
- Useni, S.Y.; Mpanda, M.M.; Khoji, M.H.; Cirezi, C.N.; Malaisse, F. & Bogaert, J. Vegetation Fires in the Lubumbashi Charcoal Production Basin (The Democratic Republic of the Congo): Drivers, Extent and Spatiotemporal Dynamics. Land 2023, 12, 2171. [CrossRef]
- Sola, P. ; Schure, J. ; Eba’a Atyi, R. ; Gumbo, D. & Okeyo, I. Politiques et pratiques en matière de bois-énergie dans certains pays d’Afrique subsaharienne – un examen critique. Bois For. Trop. 2019, 340, 27–41. [CrossRef]
- Mama, A. ; Bamba, I. ; Sinsin, B. ; Bogaert, J. & De Cannière, C. Déforestation, savanisation et développement agricole des paysages de savanes-forêts dans la zone soudano-guinéenne du Bénin. Bois For. Trop. 2014, 322(4), 65–76. [CrossRef]
- Meniko, T.H.J-P.P. ; Tshibamba, M. J. ; Sabongo, Y.P. ; Nshimba S.W.M.H. ; Dudu, A.B. ; Mate, M.J-P. & Bogaert, J. Caractérisation floristique de quatre habitats forestiers d’un gradient d’anthropisation à Masako. In Les forêts de la Tshopo : Écologie, histoire et composition ; Bogaert, J., Beeckman, H., De cannière, C., Defourny, P. et Ponette, Q., Eds ; Les Presses Universitaires de Liège, Belgique, 2020 ; 75–90.
- Guizol, P. ; Guizol, P. ; Diakhite, M. ; Seka, J. ; Mbonayem, L. ; Awono, A. ; Oyono, P.R. ; Ndikumagenge, C. ; Sonwa, D. ; Ndabirorere, S. ; Waitkuwait, E. ; Ngobieng, M.A. ; Tabi, P. & Essamba, L. La restauration des paysages forestiers (RPF) en Afrique centrale. In Les forêts du bassin du Congo : état des forêts 2021 ; Eba’a Atyi, R., Hiol Hiol, F., Lescuyer, G., Mayaux, P., Defourny, P., Bayol, N., Saracco, F., Pokem, D., Sufo Kankeu, R. & Nasi, R., Eds ; CIFOR, Bogor, Indonésie, 2022 ; 338–359.
- Talukdar, N.R., Choudhury, P., Barbhuiya, R.A. & Singh, B. Importance of Non-Timber Forest Products (NTFPs) in rural livelihood: A study in Patharia Hills Reserve Forest, northeast India. Trees For. and People 2021, 3, 100042. [CrossRef]
- Kaumbu, J.M.K.; Mpundu, M.M.M.; Kasongo, E.L.M.; Ngoy Shutcha, M.; Tekeu, H.; Kalambulwa A.N. & Khasa, D. Early Selection of Tree Species for Regeneration in Degraded Woodland of Southeastern Congo Basin. Forests 2021, 12, 117. [CrossRef]
- Giliba, R.A.; Mafuru, C.S.; Paul, M.; Kayombo, C.J.; Kashindye, A.M.; Chirenje, L.I. & Musamba, E.B. Human Activities Influencing Deforestation on Meru Catchment Forest Reserve, Tanzania. J. of Hum. Ecol. 2011, 33(1), 17–20. [CrossRef]
Figure 1.
City of Lubumbashi (grey polygon) and its rural area (white space surrounding Lubumbashi). The black dot indicates the village area of Lwisha, located approximately 80 km northwest of Lubumbashi. The geographical coordinates used for this mapping were obtained using a GPS device within the premises of the Lwisha village chief's office. The administrative boundaries on this map mark the borders between the DRC and Zambia, as well as between Kipushi Territory and other territories in Upper-Katanga Province.
Figure 1.
City of Lubumbashi (grey polygon) and its rural area (white space surrounding Lubumbashi). The black dot indicates the village area of Lwisha, located approximately 80 km northwest of Lubumbashi. The geographical coordinates used for this mapping were obtained using a GPS device within the premises of the Lwisha village chief's office. The administrative boundaries on this map mark the borders between the DRC and Zambia, as well as between Kipushi Territory and other territories in Upper-Katanga Province.
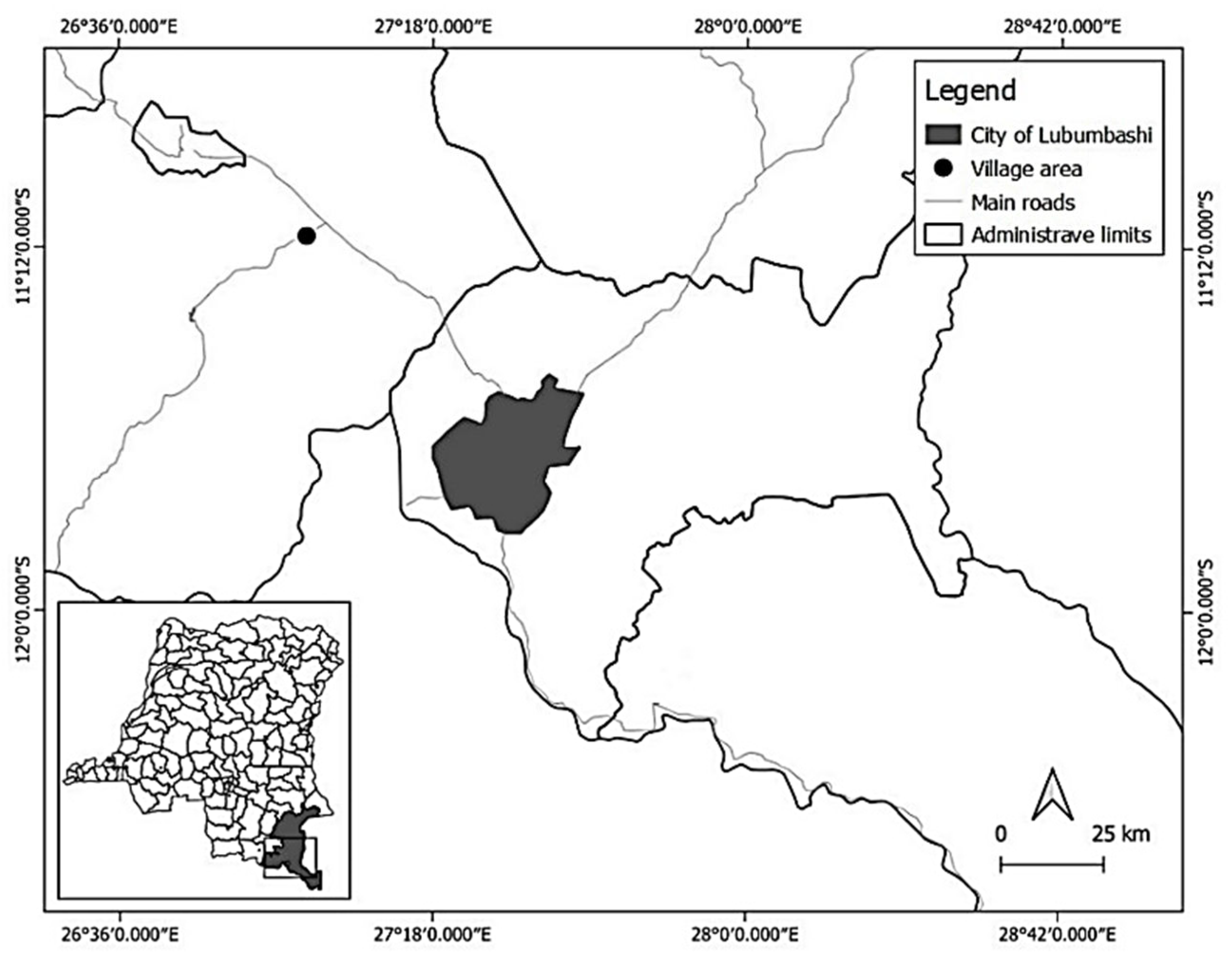
Figure 2.
Comparison of floristic richness among different habitats and diameter classes of inventoried individuals. A: Unexploited forests, B: Degraded forests, C: Post-cultivation fallows, D: comparison of Seedlings-Saplings strata. Acho : Acacia hockii, Acpo : Acacia polyacantha, Alad : Albizia adianthifolia, Alaf : Allophylus africanus, Alan : Albizia antunesiana, Alve : Albizia versicolor, Anbo : Anisophyllea boehmii, Anse : Annona senegalensis, Azga : Azanza garckeana, Babe : Baphia bequaertii, Boma : Bobgunnia madagascariensis, Bomo : Boscia mossambicensis, Brbo : Brachystegia boehmii, Brdu : Bridelia duvigneaudii, Brlo : Brachystegia longifolia, Brsp : Brachystegia spiciformis, Brta : Brachystegia taxifolia, Brwa : Brachystegia wangermeeana, Caab : Cassia abbreviata, Chba : Chrysophyllum bangweolense, Coco : Combretum collinum, Como : Combretum molle, Coze : Combretum zeyheri, Dico : Diplorhynchus condylocarpon, Dimw : Diospyros mweroensis, Ekbe : Ekebergia benguelensis, Eraf : Erythrophleum africanum, Faro : Faurea rochetiana, Fisp : Ficus sp, Fith : Ficus thonningii, Gahu : Garcinia huillensis, Gate : Gardenia ternifolia, Hafo : Haplocoelum foliolosum, Hama : Harungana madagascariensis, Hemo : Hexalobus monopetalus, Hyac : Hymenocardia acida, Isan : Isoberlinia angolensis, Isto : Isoberlinia tomentosa, Jugl : Julbernardia globiflora, Jupa : Julbernardia paniculata, Ladi : Lannea discolor, Laki : Landolphia kirkii, Mama : Marquesia macroura, Maob : Markhamia obtusifolia, Moaf : Monotes africanus, Moka : Monotes katangensis, Mucr : Multidentia crassa, Myae : Mystroxylon aethiopicum, Ocsc : Ochna schweinfurthiana, Olda : Oldfieldia dactylophylla, Olob : Olax obtusifolia, Pacu : Parinari curatellifolia, Pean: Pericopsis angolensis, Phka : Philenoptera katangensis, Phle : Phyllocosmus lemaireanus, Phmu : Phyllanthus muellerianus, Pith : Piliostigma thonningii, Pran : Protea angolensis, Psfe : Psorospermum febrifugum, Psma : Pseudolachnostylis maprouneifolia, Ptan : Pterocarpus angolensis, Ptti : Pterocarpus tinctorius, Roen : Rothmannia engleriana, Sarh : Salacia rhodesiaca, Sctr : Schrebera trichoclada, Star : Steganotaenia araliacea, Stco : Strychnos cocculoides, Stpu : Strychnos pungens, Stqu : Sterculia quinqueloba, Stsp : Strychnos sp, Stspi : Strychnos spinosa, Sygu : Syzygium guineense subsp macrocarpum, Temo : Terminalia mollis, Uabe : Uapaca benguelensis, Uaki : Uapaca kirkiana, Uani : Uapaca nitida, Uapi : Uapaca pilosa, Uaro : Uapaca robynsii, Vido : Vitex doniana, Vifi : Vitex fischeri, Vima : Vitex madiensis, Vimo : Vitex mombassae, Zaaf : Zanha africana. AUF: Unexploited forests, DFO: Degraded forests, FAL.
Figure 2.
Comparison of floristic richness among different habitats and diameter classes of inventoried individuals. A: Unexploited forests, B: Degraded forests, C: Post-cultivation fallows, D: comparison of Seedlings-Saplings strata. Acho : Acacia hockii, Acpo : Acacia polyacantha, Alad : Albizia adianthifolia, Alaf : Allophylus africanus, Alan : Albizia antunesiana, Alve : Albizia versicolor, Anbo : Anisophyllea boehmii, Anse : Annona senegalensis, Azga : Azanza garckeana, Babe : Baphia bequaertii, Boma : Bobgunnia madagascariensis, Bomo : Boscia mossambicensis, Brbo : Brachystegia boehmii, Brdu : Bridelia duvigneaudii, Brlo : Brachystegia longifolia, Brsp : Brachystegia spiciformis, Brta : Brachystegia taxifolia, Brwa : Brachystegia wangermeeana, Caab : Cassia abbreviata, Chba : Chrysophyllum bangweolense, Coco : Combretum collinum, Como : Combretum molle, Coze : Combretum zeyheri, Dico : Diplorhynchus condylocarpon, Dimw : Diospyros mweroensis, Ekbe : Ekebergia benguelensis, Eraf : Erythrophleum africanum, Faro : Faurea rochetiana, Fisp : Ficus sp, Fith : Ficus thonningii, Gahu : Garcinia huillensis, Gate : Gardenia ternifolia, Hafo : Haplocoelum foliolosum, Hama : Harungana madagascariensis, Hemo : Hexalobus monopetalus, Hyac : Hymenocardia acida, Isan : Isoberlinia angolensis, Isto : Isoberlinia tomentosa, Jugl : Julbernardia globiflora, Jupa : Julbernardia paniculata, Ladi : Lannea discolor, Laki : Landolphia kirkii, Mama : Marquesia macroura, Maob : Markhamia obtusifolia, Moaf : Monotes africanus, Moka : Monotes katangensis, Mucr : Multidentia crassa, Myae : Mystroxylon aethiopicum, Ocsc : Ochna schweinfurthiana, Olda : Oldfieldia dactylophylla, Olob : Olax obtusifolia, Pacu : Parinari curatellifolia, Pean: Pericopsis angolensis, Phka : Philenoptera katangensis, Phle : Phyllocosmus lemaireanus, Phmu : Phyllanthus muellerianus, Pith : Piliostigma thonningii, Pran : Protea angolensis, Psfe : Psorospermum febrifugum, Psma : Pseudolachnostylis maprouneifolia, Ptan : Pterocarpus angolensis, Ptti : Pterocarpus tinctorius, Roen : Rothmannia engleriana, Sarh : Salacia rhodesiaca, Sctr : Schrebera trichoclada, Star : Steganotaenia araliacea, Stco : Strychnos cocculoides, Stpu : Strychnos pungens, Stqu : Sterculia quinqueloba, Stsp : Strychnos sp, Stspi : Strychnos spinosa, Sygu : Syzygium guineense subsp macrocarpum, Temo : Terminalia mollis, Uabe : Uapaca benguelensis, Uaki : Uapaca kirkiana, Uani : Uapaca nitida, Uapi : Uapaca pilosa, Uaro : Uapaca robynsii, Vido : Vitex doniana, Vifi : Vitex fischeri, Vima : Vitex madiensis, Vimo : Vitex mombassae, Zaaf : Zanha africana. AUF: Unexploited forests, DFO: Degraded forests, FAL.
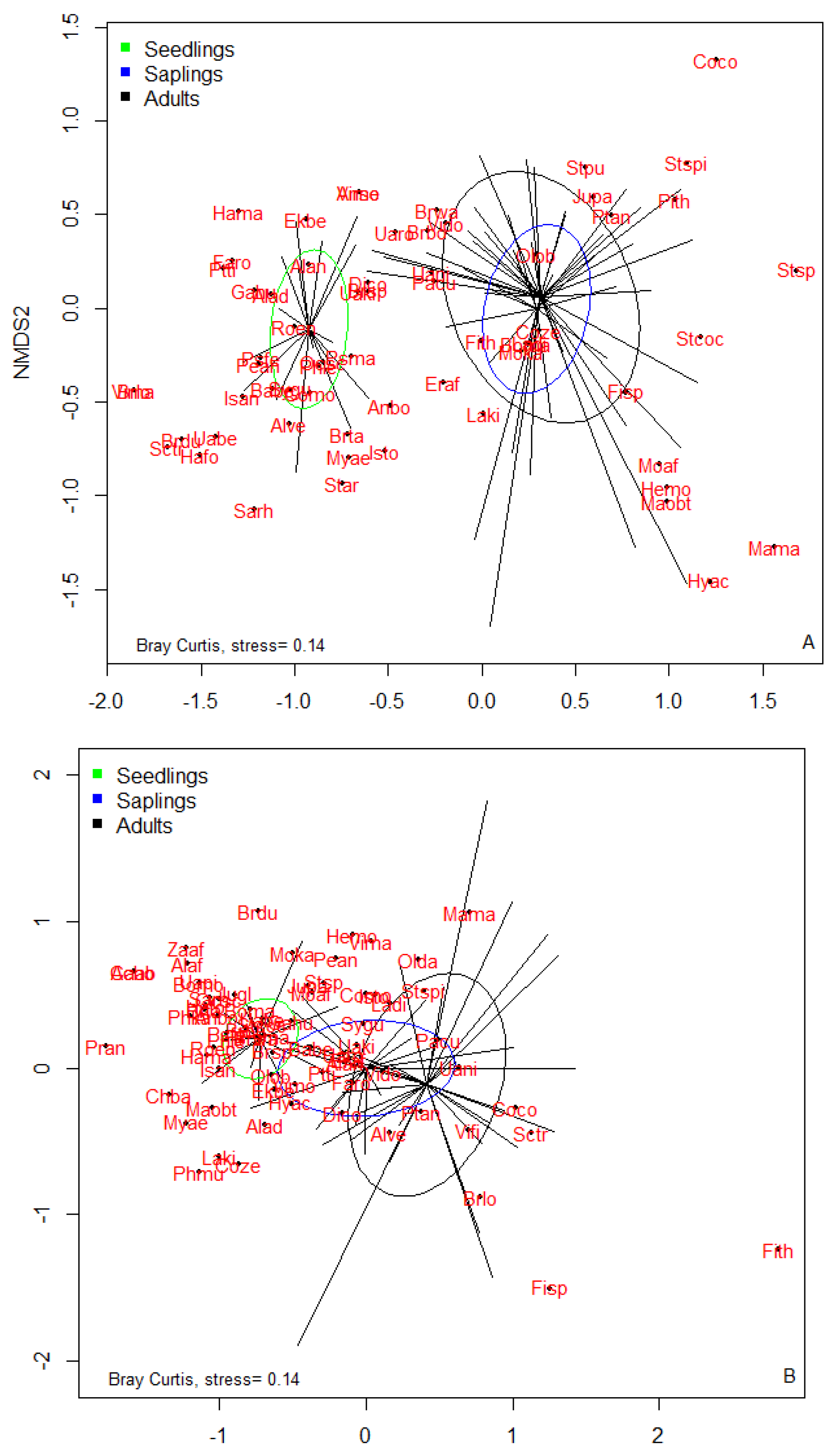
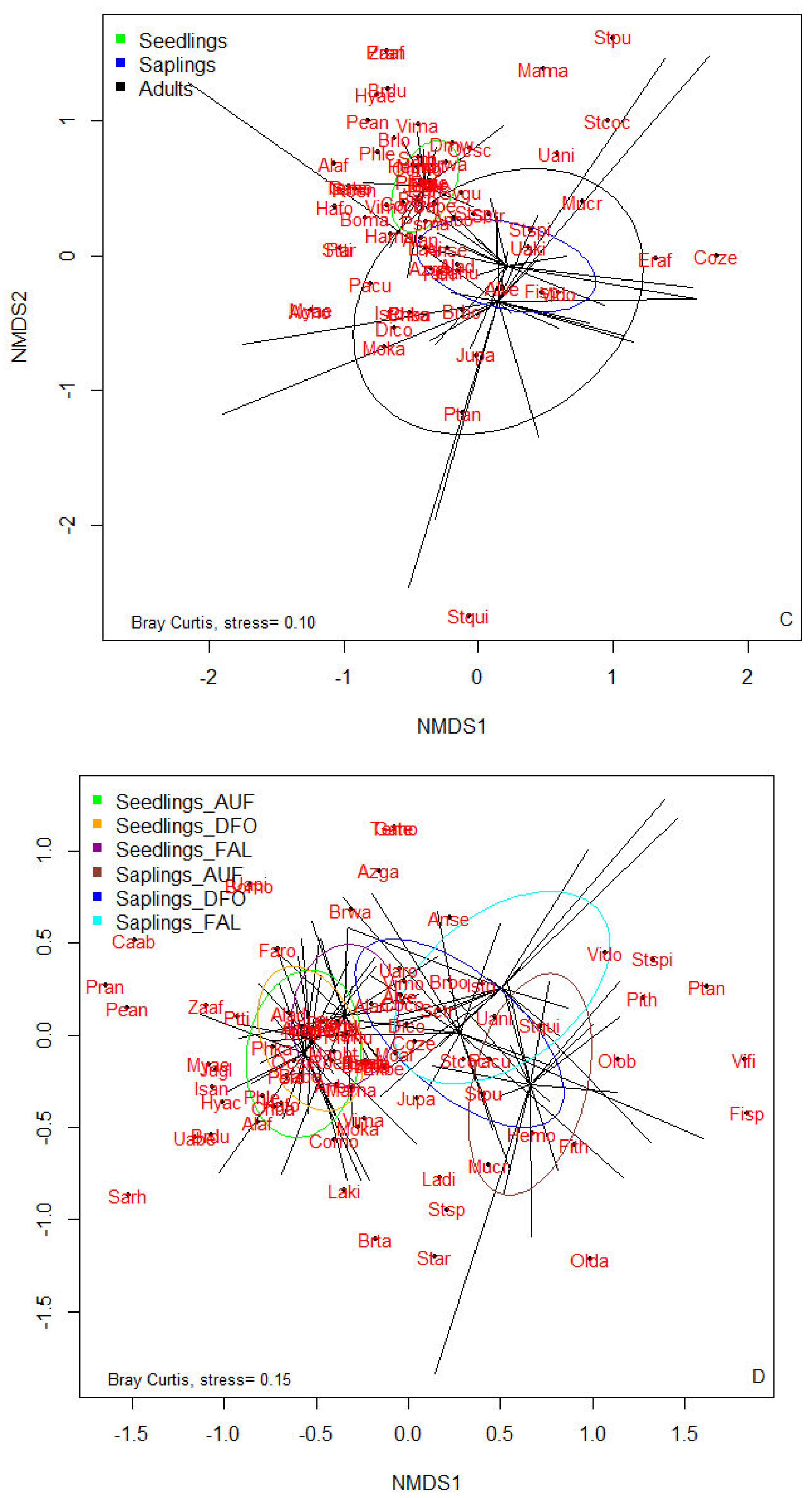

Table 1.
Presentation and description of the three surveyed habitats in Lwisha area.
| Habitat | Description |
|---|---|
Unexploited miombo forests
|
Are those not exploited for charcoal production or cultivated at a human scale [6]. These refer to the land characterized by vegetation dominated by a sparse herbaceous layer under a 10-20 m forest stratum. The canopy cover extends over at least 10-30% of the area, which spans between 0.05-1 hectares [15]. |
Degraded forests
|
Have been exploited for charcoal production [43] and correspond to forests where the capacity to provide ecosystem services has been significantly reduced due to decreased woody plant density and biodiversity. |
Post-cultivation follows
|
Fallows are habitats abandoned after subsistence farming. Referring to habitats severely damaged by excessive land use, degrading soil and vegetation, and delaying woody plant diversity recovery. Vegetation is primarily dominated by grasses [44]. |
Table 2.
Families, genera, and species of inventoried adult individuals in different habitats. AUF: unexploited forests, DFO: Degraded forests, FAL: Post-cultivation fallows. Letters indicate statistical differences between strata, -: the family was not represented during floristic inventories, n: sample size.
Table 2.
Families, genera, and species of inventoried adult individuals in different habitats. AUF: unexploited forests, DFO: Degraded forests, FAL: Post-cultivation fallows. Letters indicate statistical differences between strata, -: the family was not represented during floristic inventories, n: sample size.
| Families | AUF (%) | DFO (%) | FAL (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Genera n=27 | Species n=36 | Individuals n=500 | Genera n=33 | Species n=48 | Individuals n=456 | Genera n=25 | Species n=32 | Individuals n=143 | |
| Anacardiaceae | 3.70 | 2.78 | 0.80 | 3.03 | 2.08 | 2.63 | - | - | - |
| Anisophylleaceae | 3.70 | 2.78 | 1.00 | 3.03 | 2.08 | 0.88 | 4.00 | 3.13 | 1.40 |
| Annonaceae | - | - | - | - | - | - | 4.00 | 3.13 | 1.40 |
| Apocynaceae | 3.70 | 2.78 | 2.00 | 3.03 | 2.08 | 6.58 | 4.00 | 3.13 | 4.20 |
| Bignoniaceae | 3.70 | 2.78 | 0.60 | 3.03 | 2.08 | 0.22 | - | - | - |
| Celastraceae | 3.70 | 2.78 | 0.20 | 3.03 | 2.08 | 0.22 | - | - | - |
| Chrysobalanaceae | 3.70 | 2.78 | 1.20 | 3.03 | 2.08 | 3.51 | 4.00 | 3.13 | 2.80 |
| Clusiaceae | 3.70 | 2.78 | 0.20 | 9.09 | 6.25 | 1.10 | 4.00 | 3.13 | 0.70 |
| Combretaceae | 3.70 | 5.56 | 0.60 | 3.03 | 6.25 | 2.41 | 4.00 | 6.25 | 2.10 |
| Dipterocarpaceae | 7.41 | 8.33 | 7.20 | 6.06 | 4.17 | 5.04 | - | - | - |
| Fabaceae | 33.33 | 33.33 | 76.00 | 27.27 | 33.33 | 59.87 | 28.00 | 34.38 | 67.83 |
| Ixonanthaceae | 3.70 | 2.78 | 1.00 | 3.03 | 2.08 | 0.88 | 4.00 | 3.13 | 0.70 |
| Lamiaceae | 3.70 | 2.78 | 0.20 | 3.03 | 6.25 | 2.85 | 4.00 | 3.13 | 2.10 |
| Loganiaceae | 3.70 | 8.33 | 1.60 | 3.03 | 4.17 | 0.44 | 4.00 | 6.25 | 1.40 |
| Malvaceae | - | - | - | - | - | - | 8.00 | 6.25 | 3.50 |
| Meliaceae | 3.70 | 2.78 | 0.20 | 3.03 | 2.08 | 0.22 | - | - | - |
| Moraceae | - | - | - | 3.03 | 4.17 | 0.66 | - | - | - |
| Myrtaceae | - | - | - | 3.03 | 2.08 | 0.66 | 4.00 | 3.13 | 3.50 |
| Ochnaceae | 3.70 | 2.78 | 1.20 | 3.03 | 2.08 | 0.22 | 4.00 | 3.13 | 1.40 |
| Olacaceae | 3.70 | 2.78 | 0.20 | - | - | - | - | - | - |
| Oleaceae | - | - | - | 3.03 | 2.08 | 0.22 | 4.00 | 3.13 | 0.70 |
| Phyllanthaceae | 7.41 | 11.11 | 5.80 | 9.09 | 10.42 | 10.96 | 12.00 | 12.50 | 5.59 |
| Proteaceae | - | - | - | 3.03 | 2.08 | 0.44 | - | - | - |
| Rubiaceae | - | - | - | - | - | - | 4.00 | 3.13 | 0.70 |
| Total of frequencies | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 | 100.00 |
Table 3.
Dendrometric and floristic parameters of inventoried adult individuals in different habitats. Means ± Standard deviations. For a given parameter, habitats without common letters differ significantly at p < 0.05. AUF: Unexploited forests, DFO: Degraded forests, FAL: Post-cultivation fallows.
Table 3.
Dendrometric and floristic parameters of inventoried adult individuals in different habitats. Means ± Standard deviations. For a given parameter, habitats without common letters differ significantly at p < 0.05. AUF: Unexploited forests, DFO: Degraded forests, FAL: Post-cultivation fallows.
| Parameters | AUF | DFO | FAL |
|---|---|---|---|
| Dendrometric parameters | |||
| Density (individuals/ha) | 312.50±126.36a | 285.00±126.97a | 89.38±96.02b |
| Mean square diameter (cm) | 40.75±15.83a | 32.57±6.78b | 28.84±11.18b |
| Basal area (m2/ha) | 16.78±7.25a | 9.98±7.14a | 1.92±2.07b |
| Floristic parameters | |||
| Taxa_S/plot | 10.25±2.86a | 12.44±4.46a | 4.81±4.28b |
| Genera/plot | 8.44±2.16a | 9.88±3.36a | 3.94±3.17b |
| Families/plot | 4.67±1.96a | 5.88±1.86a | 2.69±2.06b |
Table 4.
Floristic list of the top five regenerative plant species, showing high relative frequency/density and natural regeneration index for each habitat (values in bold). The species list is presented in alphabetical order, and in case of tied values, the species concerned are counted as one. AUF: Unexploited forests, DFO: Degraded forests, FAL: Post-cultivation fallows. NRI: Natural regeneration index (ratio between juveniles and adults), dbh < 1 cm: Seedlings, 1 cm ≤ dbh < 10 cm: Saplings, - : Species not inventoried, n= sample size. RF (Relative Frequency) and RD (Relative Density) values are expressed in percentage. The entire set of species studied is presented in the appendix.
Table 4.
Floristic list of the top five regenerative plant species, showing high relative frequency/density and natural regeneration index for each habitat (values in bold). The species list is presented in alphabetical order, and in case of tied values, the species concerned are counted as one. AUF: Unexploited forests, DFO: Degraded forests, FAL: Post-cultivation fallows. NRI: Natural regeneration index (ratio between juveniles and adults), dbh < 1 cm: Seedlings, 1 cm ≤ dbh < 10 cm: Saplings, - : Species not inventoried, n= sample size. RF (Relative Frequency) and RD (Relative Density) values are expressed in percentage. The entire set of species studied is presented in the appendix.
| Species | Family | dbh<1cm | 1 cm≤dbh<10 cm | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AUF (n=6628) | DFO (n=6660) | FAL (n=4676) | AUF (n=948) | DFO (n=2384) | FAL (n=1756) | ||||||||||||||
| RF | RD | NRI | RF | RD | NRI | RF | RD | NRI | RF | RD | NRI | RF | RD | NRI | RF | RD | NRI | ||
| Albiziaadianthifolia (Schumach.) W. Wight | Fabaceae | 4.17 | 6.76 | 29.87 | 4.62 | 9.01 | 12.50 | 5.50 | 5.39 | 15.75 | 4.86 | 3.38 | 2.13 | 5.17 | 6.71 | 3.33 | 5.54 | 5.47 | 6.00 |
| Albizia antunesiana Harms | Fabaceae | 4.17 | 4.16 | 23.00 | 3.08 | 2.52 | 12.92 | 5.11 | 3.25 | 50.67 | 2.78 | 1.69 | 1.33 | 4.13 | 1.85 | 3.38 | 4.54 | 6.15 | 36.00 |
| Anisophyllea boehmii Engl. | Anisophylleaceae | 3.13 | 1.39 | 18.40 | 2.77 | 2.88 | 48.00 | 1.97 | 1.71 | 40.00 | 3.47 | 2.95 | 5.60 | 2.07 | 0.67 | 4.00 | 7.06 | 3.19 | 28.00 |
| Baphia bequaertii De Wild. | Fabaceae | 5.56 | 7.91 | 23.82 | 4.31 | 5.29 | 16.76 | 5.11 | 6.50 | 50.67 | 3.47 | 3.38 | 1.45 | 5.68 | 5.70 | 6.48 | 3.53 | 7.06 | 20.67 |
| Brachystegia spiciformis Benth. | Fabaceae | 5.56 | 12.61 | 7.81 | 4.93 | 13.33 | 15.86 | 6.29 | 10.61 | 31.00 | 10.42 | 15.19 | 1.35 | 6.71 | 5.54 | 2.36 | 6.05 | 7.97 | 8.75 |
| Brachystegia wangermeeana De Wild. | Fabaceae | 4.51 | 10.80 | 4.39 | 4.93 | 20.48 | 16.43 | 5.90 | 27.89 | 42.06 | 6.94 | 17.30 | 1.01 | 6.20 | 29.53 | 8.48 | 7.56 | 23.46 | 13.29 |
| Combretum molle R. Br ex G. Don | Combretaceae | 2.43 | 1.21 | 40.00 | 0.92 | 0.42 | 9.33 | 3.54 | 3.08 | 72.00 | 0.69 | 0.84 | 4.00 | 1.03 | 0.50 | 4.00 | 1.51 | 1.37 | 12.00 |
| Combretum zeyheri Sond. | Combretaceae | - | - | - | 0.62 | 0.18 | 6.00 | - | - | 0.00 | 0.69 | 0.42 | - | 1.55 | 2.85 | 34.00 | 1.51 | 0.91 | 16.00 |
| Diplorhynchus condylocarpon (Müll. Arg.) Pichon | Apocynaceae | 2.78 | 2.96 | 19.60 | 4.31 | 2.46 | 5.47 | 1.18 | 0.86 | 6.67 | 3.47 | 5.91 | 5.60 | 5.68 | 8.39 | 6.67 | 2.02 | 1.82 | 5.33 |
| Ekebergia benguelensis Welw. ex C.DC. | Meliaceae | 2.08 | 0.54 | 36.00 | 1.54 | 0.36 | 24.00 | 2.36 | 1.45 | - | - | - | 0.00 | 1.55 | 0.50 | 12.00 | 2.02 | 1.14 | - |
| Garcinia huillensis Oliv. | Clusiaceae | 3.13 | 1.15 | - | 2.16 | 0.48 | 16.00 | 1.57 | 0.34 | 16.00 | - | - | - | - | - | 0.00 | 0.50 | 0.23 | 4.00 |
| Harungana madagascariensis Lam. ex Poir. | Clusiaceae | 1.04 | 0.36 | - | 2.77 | 2.04 | 136.00 | 1.57 | 1.28 | - | - | - | - | - | - | 0.00 | 1.01 | 0.46 | - |
| Hymenocardia acida Tul. | Phyllanthaceae | - | - | - | 1.23 | 1.26 | 42.00 | 1.18 | 5.99 | - | 0.69 | 0.42 | 1.55 | 1.34 | 16.00 | 1.01 | 0.46 | - | |
| Isoberlinia angolensis (Benth.) Hoyle & Brenan | Fabaceae | 3.82 | 11.04 | 45.75 | 4.62 | 5.23 | 38.67 | 3.54 | 4.53 | 42.40 | 4.86 | 2.95 | 1.75 | 2.07 | 4.53 | 12.00 | 3.53 | 6.61 | 23.20 |
| Isoberlinia tomentosa (Harms) Craib & Stapf | Fabaceae | 0.35 | 0.18 | - | - | - | 0.00 | - | - | 0.00 | - | - | - | 4.13 | 4.53 | 21.60 | 2.02 | 2.28 | 8.00 |
| Julbernardia paniculata (Benth.) Troupin | Fabaceae | 0.69 | 0.12 | 1.60 | 1.85 | 1.44 | 24.00 | - | - | 0.00 | 0.69 | 0.84 | 1.60 | 1.55 | 0.67 | 4.00 | 1.01 | 1.14 | 20.00 |
| Markhamia obtusifolia (Boulanger) Sprague | Bignoniaceae | 0.35 | 0.12 | 2.67 | 1.23 | 0.66 | 44.00 | 1.97 | 1.20 | - | 1.39 | 0.84 | 2.67 | 1.03 | 1.01 | 24.00 | - | - | - |
| Mystroxylon aethiopicum (Thunb.) Lœs. | Celastraceae | 0.69 | 0.24 | 16.00 | 0.92 | 0.90 | 60.00 | 0.39 | 0.09 | - | - | - | 0.00 | 1.03 | 0.50 | 12.00 | - | - | - |
| Ochna schweinfurthiana F. Hoffm. | Ochnaceae | 4.17 | 3.02 | 33.33 | 3.69 | 3.78 | 252.00 | 3.93 | 2.65 | 62.00 | 4.17 | 2.53 | 4.00 | 3.10 | 2.18 | 52.00 | 1.51 | 0.91 | 8.00 |
| Olax obtusifolia De Wild. | Olacaceae | 0.69 | 0.24 | 16.00 | 0.92 | 0.18 | - | - | - | - | 3.47 | 2.53 | 24.00 | - | - | - | - | - | - |
| Parinari curatellifolia Planch. ex Benth. | Chrysobalanaceae | 1.74 | 0.48 | 5.33 | 1.85 | 0.84 | 3.50 | 1.57 | 0.86 | 10.00 | 2.08 | 1.27 | 2.00 | 4.13 | 3.69 | 5.50 | 3.53 | 2.96 | 13.00 |
| Phyllocosmus lemaireanus (De Wild. & T. Durand) T. Durand & H. Durand | Ixonanthaceae | 2.78 | 2.41 | 32.00 | 3.08 | 2.58 | 43.00 | 3.93 | 1.97 | 92.00 | 2.78 | 2.53 | 4.80 | 2.58 | 1.01 | 6.00 | 0.50 | 0.23 | 4.00 |
| Pseudolachnostylis maprouneifolia Pax | Phyllanthaceae | 2.78 | 2.35 | 78.00 | 3.39 | 1.80 | 17.14 | 3.14 | 1.80 | 42.00 | 2.08 | 2.53 | 12.00 | 2.07 | 1.01 | 3.43 | 5.04 | 3.19 | 28.00 |
| Psorospermum febrifugum Spach | Clusiaceae | 4.51 | 4.47 | 296.00 | 2.77 | 2.04 | 68.00 | 4.32 | 1.88 | - | - | - | 0.00 | 1.03 | 0.34 | 4.00 | 3.02 | 1.37 | - |
| Pterocarpus angolensis DC. | Fabaceae | 0.35 | 0.06 | 0.29 | - | - | 0.00 | 0.39 | 0.17 | 1.60 | 5.56 | 5.49 | 3.71 | 3.10 | 1.17 | 3.50 | 3.02 | 1.59 | 5.60 |
| Rothmannia engleriana (K. Schum.) Keay | Rubiaceae | 3.82 | 2.53 | - | 2.16 | 1.50 | 4.72 | 3.25 | 152.00 | 1.39 | 0.84 | - | - | - | - | 1.01 | 0.46 | 8.00 | |
| Schrebera trichoclada Welw. | Oleaceae | 0.69 | 0.12 | - | - | - | 0.00 | 1.57 | 0.60 | 28.00 | - | - | - | - | - | 0.00 | 1.51 | 1.59 | 28.00 |
| Strychnos cocculoides Boulanger | Loganiaceae | 0.35 | 0.06 | 1.33 | 1.54 | 0.42 | 28.00 | 1.97 | 1.37 | 64.00 | 2.08 | 1.27 | 4.00 | 1.55 | 0.67 | 16.00 | 1.51 | 0.68 | 12.00 |
| Strychnos spinosa Lam. | Loganiaceae | - | - | 0.00 | - | - | - | 0.39 | 0.09 | - | 0.69 | 0.42 | 4.00 | 1.03 | 0.50 | - | 1.51 | 0.68 | - |
| Syzygium guineense (Willd.) DC. subsp. macrocarpum | Myrtaceae | 3.13 | 1.03 | - | 1.23 | 0.48 | 10.67 | 2.75 | 1.97 | 18.40 | 1.39 | 0.84 | - | 1.55 | 1.01 | 8.00 | 3.53 | 2.05 | 7.20 |
| Uapaca kirkiana Müll. Arg. | Phyllanthaceae | 4.51 | 3.14 | 13.00 | 2.46 | 2.70 | 6.43 | 0.79 | 0.34 | 4.00 | 4.17 | 3.80 | 2.25 | 3.62 | 2.35 | 2.00 | 2.02 | 1.37 | 6.00 |
| Vitex doniana Sweet | Lamiaceae | 0.35 | 0.06 | 4.00 | 0.62 | 0.18 | 2.40 | 0.79 | 0.34 | 5.33 | 0.69 | 0.42 | 4.00 | 1.03 | 0.34 | 1.60 | 2.02 | 1.14 | 6.67 |
| Vitex mombassae Vatke | Lamiaceae | 0.35 | 0.12 | - | 0.62 | 0.12 | 8.00 | 1.97 | 0.77 | - | - | - | - | 1.55 | 0.67 | 16.00 | - | - | - |
Table 6.
Jaccard similarity between floristic lists of different strata in the three habitats. AUF: unexploited forests, DFO: degraded forests, FAL: post-cultivation fallows, <1: Seedlings; ≥1: Saplings; ≥10: Adults. Relative values are presented in decimal form.
Table 6.
Jaccard similarity between floristic lists of different strata in the three habitats. AUF: unexploited forests, DFO: degraded forests, FAL: post-cultivation fallows, <1: Seedlings; ≥1: Saplings; ≥10: Adults. Relative values are presented in decimal form.
| AUF<1 | DFO<1 | FAL<1 | AUF≥1 | DFO≥1 | FAL≥1 | AUF≥10 | DFO≥10 | |
|---|---|---|---|---|---|---|---|---|
| DFO<1 | 0.65 | |||||||
| FAL<1 | 0.68 | 0.65 | ||||||
| AUF≥1 | 0.65 | 0.55 | 0.50 | |||||
| DFO≥1 | 0.71 | 0.59 | 0.55 | 0.92 | ||||
| FAL≥1 | 0.67 | 0.64 | 0.74 | 0.63 | 0.68 | |||
| AUF≥10 | 0.65 | 0.48 | 0.57 | 0.71 | 0.79 | 0.63 | ||
| DFO≥10 | 0.67 | 0.57 | 0.59 | 0.73 | 0.80 | 0.74 | 0.86 | |
| FAL≥10 | 0.56 | 0.42 | 0.50 | 0.71 | 0.67 | 0.63 | 0.60 | 0.63 |
Table 5.
Comparison of dendrometric parameters and species richness between the two classes of juvenile individuals inventoried in the three habitats. Means ± Standard Deviations. For a given parameter, habitats without common letters differ significantly at p < 0.05. AUF: Unexploited forests, DFO: Degraded forests, FAL: Post-cultivation fallows, dbh<1 cm: Seedlings, 1 cm ≤ dbh < 10 cm: Saplings, - : values were not calculated due to lack of relevant data.
Table 5.
Comparison of dendrometric parameters and species richness between the two classes of juvenile individuals inventoried in the three habitats. Means ± Standard Deviations. For a given parameter, habitats without common letters differ significantly at p < 0.05. AUF: Unexploited forests, DFO: Degraded forests, FAL: Post-cultivation fallows, dbh<1 cm: Seedlings, 1 cm ≤ dbh < 10 cm: Saplings, - : values were not calculated due to lack of relevant data.
| dbh<1 cm | 1 cm≤dbh<10 cm | |||||
|---|---|---|---|---|---|---|
| AUF | DFO | FAL | AUF | DFO | FAL | |
| Dendrometric parameters | ||||||
| Density (individuals/ha) | 4142.50±2176.33ab | 4185.00±1544.84a | 2935.00±1567.39b | 592.50±341.36a | 1490.00±1133.21a | 1110.00±954.82a |
| Mean square diameter (cm) | - | - | - | 7.52±0.66a | 7.16±0.48a | 7.06±0.49a |
| Basal area (m2/ha) | - | - | - | 2.49±1.26a | 5.76±4.36a | 4.25±3.71a |
| Floristic parameters | ||||||
| Individuals | 103.56±54.41ab | 104.63±38.62a | 73.38±39.18b | 14.81±8.53a | 37.25±28.33a | 27.75±23.87a |
| Taxa_S | 18.00±3.97ab | 20.50±3.41a | 16.06±3.99b | 16.88±6.91a | 19.13±10.83a | 19.19±14.62a |
| Genera | 15.25±4.09ab | 17.75±3.49a | 13.44±3.76b | 7.81±3.62a | 10.63±5.15a | 10.13±6.39a |
| Families | 9.56±2.78a | 10.88±2.83a | 9.31±2.89a | 5.19±2.83a | 6.50±3.18a | 6.63±3.91a |
| Chao-1 | 21.14±5.85a | 25.57±4.78a | 21.30±8.29a | 18.87±11.80a | 20.80±11.22a | 23.16±17.04a |
| Taxa_S/Chao-1 | 0.85 | 0.80 | 0.75 | 0.90 | 0.92 | 0.83 |
| Fisher_alpha | 6.78±1.66a | 8.57±3.13a | 6.99±2.11a | 10.74±6.17a | 9.96±11.91a | 8.71±5.79a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



