
Submitted:
11 June 2024
Posted:
12 June 2024
You are already at the latest version
Abstract
Dairy products are an important source of protein and other nutrients in the Mediterranean diet. In these countries the most common source of milk for producing dairies are cow, goat, sheep and buffalo. Andalusia is traditionally the largest producer of goat milk in Spain. Kefir is a fermented product made from bacteria and yeasts which has health benefits beyond its nutritional properties. There is a lack of knowledge about the molecular mechanisms and metabolites that bring about these benefits. In this work, the combination of analytical techniques (GC-FID, UHPLC-MS-QToF, GC-QqQ-MS, and GC-ToF-MS) resulted in the detection of 105 metabolites in kefir produced with goat milk from two different thermal treatments (raw and pasteurized) fermented at four times (12, 24, 36, and 48 h, using 0 h as control). Of these, 27 metabolites differed between kefir produced with raw and pasteurized milk. These changes could be possibly caused by the effect of pasteurization on the microbial population in the starting milk. Some interesting molecules were identified, as shikimic acid, dehydroabietic acid, GABA and tyramine that could be related to antibacterial properties, enhance of immune system and arterial pressure. Moreover, a viability assay of the NIRS technique was performed to evaluate its use on monitoring the fermentation and classification of samples, which had resulted in a 90 % of correct classified samples according to their fermentation time. This study represents the most comprehensive metabolomic analysis of goat milk kefir so far, revealing the intricate changes in metabolites during fermentation and the impact of milk treatment.
Keywords:
fermented milk
; metabolome
; metabolites
; near infrared spectroscopy
; sample classification
1. Introduction
The consumption of goat milk and its derived products holds significant importance in the Mediterranean countries. Goat milk is very important in the renowned Mediterranean diet, valued for its nutritional richness and cultural significance throughout centuries [1]. Spain is one of the largest producers of goat livestock worldwide, and the use of goat milk and products like cheese is deeply rooted in the country’s culinary tradition. The high protein and mineral content of goat milk make it a valuable source of nutrients for the population, contributing to its importance in Spain’s agricultural and dietary landscape [2].
Kefir, a dairy product resulting from acid-alcoholic fermentation, has been increasingly recognized for its potential health benefits [3,4]. The consumption of this fermented milk has been associated with improved digestion, modulation of the immune system, and potential anti-inflammatory properties [5]. This has led to a growing interest in kefir as a functional food in Spain and worldwide. In particular, goat milk kefir is gaining attention due to the unique properties of goat milk and the potential synergistic effects of kefir fermentation on its nutritional value and health benefits [6]. While cow milk kefir has been extensively studied, goat milk kefir remains less explored despite goat milk’s nutritional advantages, such as easier digestibility, lower allergenicity and a favorable lipid profile when compared to cow milk [7,8].
Research in this omics field has made significant contributions to understanding the molecular features of milk kefir. Several studies have profiled the peptide composition of cow milk kefir, which has been the first and most characterized so far [9,10,11,12]. More recently, studies made by our research group have characterized goat and sheep milk kefir at different fermentation times using a peptidomics approach, shedding light on the dynamic changes in protein digestion during fermentation and identifying potential peptides with biological activity [13,14]. Another recent study has highlighted the intricate metabolic interactions and microbial community diversity of kefir grains, emphasizing the role of metabolomics in unraveling the complex metabolic processes underlying cow milk kefir fermentation [15]. Metabolomic analysis can play a crucial role in the biochemical characterization of kefir and other dairy products. By studying the complete set of small-molecule metabolites present in kefir, metabolomics may provide a comprehensive understanding of the metabolic processes occurring during fermentation and their impact on the properties of the final product. Also, metabolomic approaches enable the identification of bioactive compounds, flavor precursors, and metabolic pathways associated with the beneficial properties of foods, including kefir. This information can be highly valuable for quality control, product development, and understanding the potential health effects of kefir consumption [16].
This study focuses on the metabolomic analysis of goat milk kefir, aiming to characterize the differences between raw and pasteurized goat milk kefir throughout various fermentation times. For this, we have performed a comprehensive range of analytical techniques to characterize the metabolome in different conditions, as well as the near infrared spectroscopy (NIRS) technique to better characterize and also discriminate samples. Our metabolomics approach, which is to our knowledge the first performed in this product, offers a comprehensive way to understanding the biochemical changes during fermentation, providing insights into the nutritional and therapeutic potential of goat milk kefir.
2. Materials and Methods
2.1. Reagents
Methanol (hypergrade for LC-MS, LiChrosolv®), chloroform (for analysis, EMPARTA®), water (hypergrade for LC-MS, LiChrosolv®) and pyridine (for analysis, EMSURE®), were from Supelco (Merck KGaA, Darmstadt, Germany). Sigma-Aldrich Chemie (Sant Quentin Fallavier, France) supplied ribitol, methoxyamine hydrochloride (purity 98%), and pure standards for the target compounds. N-methyl-N-trimethylsilyl-trifluoroacetamide (MSTFA) was from Macherey-Nagel (Düren, Germany).
2.2. Kefir Production
Kefir grains (Kefiralia, Gipuzkoa, Spain) were used to ferment goat milk, both raw and pasteurized at 74 ºC, provided by a local farm (Quesería “Los Peña”, Baena, Córdoba, Spain). Prior to the definite samples, the kefir grains were conditioned in the two types of milk (raw and pasteurized) for one week, adding the grains (5% w/v) in both types of milk, i.e., raw and pasteurized. After that, samples were prepared by adding the kefir grains (5% w/v) to either raw or pasteurized goat milk, at four different times (12, 24, 36, and 48 h), using the corresponding non-fermented milk (time 0) as control. Fermentations were performed in an incubator at 25 ºC under aerobic conditions and without shaking. Three biological replicates were made for each type of sample. The fermented milk, known as kefir, was separated from the grains using a sieve, and then the coagulated caseins and remaining microbial cells were removed by centrifugation at 4,500 × g for 10 min at 4 ºC. The samples were stored at -20 ºC in 50 mL conical centrifuge tubes until further metabolomics or NIRS analysis. Aliquots of non-fermented raw and pasteurized milk were also frozen to be subject to the same treatment as fermented samples.
2.3. Analysis of Fatty Acids by GC-FID
Cold extraction of fats from kefir samples was carried out according to a previously described method [17]. The resulting lipid fraction was subsequently subjected to a methylation reaction with 10% (w/v) KOH in methanol using vortex. After decanting, a small volume of the supernatant phase, containing the methyl esters, was taken in a vial. A gas chromatography equipment with a GC-FID Clarus 500 flame ionisation detector (PerkinElmer®) was used for fatty acid analysis. The capillary column used was a BPX70 (SGE Analytical ScienceTM), specific for methyl esters from fatty acids. Hydrogen was used as carrier gas at a pressure of 25 PSI. The injector and detector temperatures were set at 235 and 250 °C, respectively. For each analysis, 1 μL of sample was injected in split mode. The column oven temperature was maintained at 170 °C for the first 10 min, followed by a 5 min ramp to 235 °C, which was kept stable for an additional 3 min. Data acquisition and processing was performed using Total Chrome Navigator software (PerkinElmer®). The fatty acid profile was expressed as a % relative area of each peak recorded.
2.4. Analysis of Volatile Compounds by Headspace GC-QqQ-MS
For the analysis of volatile compounds with this technique, 1 g of kefir sample was weighed and 5 μL of a 100 ppm internal standard solution was added in a 10 mL vial. The analysis was carried out on a gas chromatographer coupled to a Scion TQ triple quadrupole GC-MS (QqQ) mass detector (Bruker). The GC was equipped with a DB-5MS capillary column (30 m length, 0.25 mm inner diameter, 0.25 μm phase thickness, Agilent). Incubation was performed at 80 °C for 12 min and 0.5 mL was injected from the headspace of the vial. The injector temperature was set at 225 °C and a 1-10 split. For the separation of the compounds, the following temperature program was set for the oven: 40 °C for the first 2 min, followed by a first temperature ramp of 10 °C/min until 175 °C was reached, and a second ramp of 35 °C/min until 275 °C was reached. The analyzer was operated in fullscan mode in the m/z range from 33 to 235. The source and transfer line temperature was set at 250 °C. The identification of each compound was carried out by comparison of its experimental mass spectra with those of the NIST library (https://chemdata.nist.gov).
2.5. Analysis of Non-Volatile Compounds by GC-ToF-MS
Kefir samples were processed as previously described [18]. A ribitol solution (0.2 mg/mL in ultrapure water) was used as internal quantitative standard. Briefly, approximately 80 mg of fresh samples were extracted with 480 µL of methanol: ribitol solution (4% v/v) and vortexed. Samples were shaken at 70 º C for 15 min, and centrifuged at 14000 rpm for 10 min (microcentrifuge 5415 R, Eppendorf AG, Hamburg Germany). Supernatants were transferred to new 2 ml eppendorf tubes and resuspended in 250 µL of chloroform and 500 µL of ultrapure water. After vortex and centrifugation at 4000 rpm for 15 min, 150 µL from the upper phase dried for at least 3 hours, without heating. The dried extracts were derivatized by using 40 µL of methoxyamine hydrochloride in pyridine (20 mg/ml, p/v), and shaken at 37 ºC for 2 h. After adding 70 µL MSTFA: Fatty acid methyl ester solution (20% v/v) (N-Methyl-N-trimethylsilyl-trifluoroacetamide, samples were shaken at 37 ºC for 30 min and transferred into GC vials. GC-TOF-MS analysis was performed using an Agilent 7890B GC system (Agilent Technologies, Palo Alto, CA, USA) with a Gerstel Multipurpose MPS autosampler (GERSTEL GmbH & Co.KG) and a Pegasus HT TOF-MS (Leco Corporation, St. Joseph, MI, USA). The analytical program employed for sample analysis was adopted from a previous study [18]. Three biological replicates for each sample were injected and the analyses were performed in random order to reduce the bias. The results for each metabolite were expressed using a relative index (RI), which was calculated according to the following equation:
2.6. Analysis of Non-Volatile Compounds by UHPLC-QToF-MS
A liquid-liquid pre-extraction was performed, in which 1 g of kefir sample was mixed with 10 mL of methanol. Also, 100 μL of a 10 ppm internal standard solution was added to compensate for systematic and random errors throughout the process. From the supernatant resulting from the extraction, a small fraction was taken, filtered to remove any remaining protein in suspension, and transferred to a vial for analysis. The analysis of polar metabolites was performed on a high-pressure liquid chromatograph equipment coupled to a G2XS quadrupole-time-of-flight (QToF) UHPLC-MS analyser (WatersTM). The analyser conditions, in positive electrospray ionisation and sensitivity mode, were as follows: capillary and cone voltages of 0.8 and 20 kV, respectively; mass acquisition range (m/z) 60-1,200 Da; cone gas and desolvation gas flow rates of 50 and 1000 L/h, respectively, and source and desolvation gas temperatures of 120 and 500 °C. The QToF was operated in MSE mode with a low collision energy of 6 eV, and a high energy ramp from 10 to 45 eV. For the acquisition, a 200 ppb solution of leucine (in acetonitrile/water 0.1% formic acid, 50:50), infused at 10 s intervals for internal mass correction, and a 0.5 mM solution of sodium formate (in 2-propanol/water, 90:10) was used as a reference and a 0.5 mM solution of sodium formate (in 2-propanol/water, 90:10) as a calibrant. An ACQUITY UPLC®BEH C18 column (2.1 × 50 mm, 1.7 μm) at 40 °C was used for metabolite separation. The mobile phase flow rate was 0.4 mL/min, using two mobile phases: A (0.1% formic acid) and B (methanol). An elution gradient of 18 min duration was used: 0-11 min 4% B: 11-13.8 min, 100%: 13.8-14 min, 100%: 14-18 min, 96%. In all cases, the volume of sample injected was 2 μL. Data acquisition and processing was performed using UNIFITM software. Tentative identification of polar metabolites was performed by m/z search, based on previously published work [19]. Results were expressed in relative areas based on the internal standard used.
2.7. NIRS Analysis
The kefir samples were scanned by reflectance on a Foss DS2500 NIRS spectrophotometer. The equipment monitors the VIS (visible) region between 400-1,100 nm as well as the NIR (near infrared) region between 1,100-2,500 nm. Therefore, the monitored region encompasses the entire region of the electromagnetic spectrum between 400 nm and 2,500 nm. 750 µL of each sample was deposited in the transport module with a circular gold-coated aluminium lid to promote the correct distribution of the kefir on the quartz glass and to avoid the presence of bubbles. The collected spectra were taken every 8.5 nm and the absorbance data were obtained as log(1/R), where R corresponds to the reflectance. Two spectra were obtained for each sample, giving a total of 60 spectra corresponding to 30 for each type of milk (raw and pasteurised) and 6 for each fermentation time (0, 12, 24, 36 and 48 h).
2.8. Data and Statistical Analysis
The software used for the analysis was RStudio version 2023.3.1.44619. The principal component analysis (PCA) and discriminant analysis of principal components (DAPC) were performed using relative area data or NIRS spectra using the following packages: stats, ggplot2, pls, FactoMiner, factoextra, MASS, tidymodels and caret. The NIRS spectra were previously normalized by SNV (Standard Normal Variate) and MSC (Multiple Scatter Correction), where MSC was chosen as the best normalization. The regions of the electromagnetic spectra used comprise the whole VIS + NIR region. The DAPC was run considering 15 PCs as the optimal number of PCs to retain. The confusion matrices of the classification models were obtained using the by Leave One Out Cross Validation (LOOCV). The statistical significance of differences was performed by one-way ANOVA with Tukey’s HSD post hoc test (p < 0.05) considering the following p-adjusted values: ns, p > 0.05; *, p < 0.05; **, p < 0.01; and ***, p < 0.001. Heatmaps were generated using the ComplexHeatmap and circlize packages. To represent metabolite data with relative area equal to 0, a value of 0.01 was added to the entire data matrix. The log2(Fold change) was then calculated and used to generate the heatmaps. The line plots were generated using ggplot2 and ggprism.
3. Results
3.1. Metabolomic Analysis of Goat Milk Kefir
With the aim of characterizing the qualitative and quantitative changes of metabolites during the fermentation time and the differences between raw and pasteurized milk in the production of kefir, we applied a range of analytical techniques to identify fatty acids (GC-FID), volatile (GC-QqQ-MS) and non-volatile compounds (UHPLC-ToF-MS, GC-ToF-MS). In total, we found 105 unique compounds in the ten different categories of samples—two thermal treatments, i.e., raw and pasteurized milk; and five fermentation times, i.e., 0, 12, 24, 36, and 48 h—, whose identities, and the technique with which they were identified are shown in Table 1. Out of the 105 metabolites, 21 were fatty acids identified by GC-FID; 26 volatile compounds were detected by GC-QqQ-MS; other 48 molecules were found by GC-ToF-MS; and 15 compounds were identified using UHPLC-ToF-MS. Five compounds were overlapping between these two latter techniques.
The three-dimensional principal component analysis (PCA) using all the metabolites detected with the above cited techniques explained the 67.02% of variance (40.41% for PC1, 16.91% for PC2 and 9.7% for PC3), clearly separating the two non-fermented milks (raw and pasteurized), but confounding some fermentation times, especially those at 36 and 48 h (Figure 1A). However, the discriminant analysis of principal components (DAPC) clearly separated the 10 different types of samples (Figure 1B), both according to the heat treatment of milk (raw vs pasteurized) and to the fermentation time (0, 12, 24, 36, and 48 h).
Figure 2 shows heatmaps and hierarchy cluster analyses of the found metabolites grouped according to the techniques with which they were identified (metabolite levels in all the samples are found in Supplementary Tables 1-5). Out of the 21 fatty acids detected, 20 decreased during fermentation (Figure 2A). In most cases, the decrease started in the first fermentation time measured, i.e., 12 h. Only one fatty acid, acid lignoceric, increased as the fermentation time progressed, being its increased more pronounced in raw milk kefir than in the pasteurized product, in which its augment was very slight. Regarding volatile compounds, some alcohols (ethanol and/or alkyl derivatives of other alcohols like propanol, butanol, hexanol or heptanol) appeared during the fermentation as the result of the yeast metabolism, as well as the major products of bacterial fermentation, i.e., acetic and lactic acid and/or their derivatives after reacting with ethanol (namely ethyl lactate and ethyl acetate) (Figure 2B). Other compounds including alkanes, ketones, aldehydes, organic acids and their ester derivatives were also found. For most cases, their increases were more pronounced in the raw milk kefir than in pasteurized milk kefir, although there were exceptions as the case of 2,3-butanedione. Figure 2C, 2D and 2E show the evolution of non-volatile molecules detected by LC-QToF-MS, and by GC-ToF-MS either using commercial standards (panel 2D) or by searching the mass spectra in libraries (panel 2E). Almost all amino acid detected clearly increased with fermentation, which is indicative of their released due to protein degradation by microbial proteases. Some sugars, like sucrose, glucose or glucose-6-phosphate, decreased, as well as lactose as expected. This was accompanied by the increase in lactic acid, as the result of lactic fermentation, and its derivative lactamide. Also, some molecules as GABA, shikimic acid, dehydroabietic acid and tyramine were also augmented as the fermentation progressed in both types of milk.
The evolution of some of the most relevant metabolites of lactic-alcoholic fermentation that occurred in our kefir samples is shown in Figure 3. Panels 3A-B show the changes of electron donors and final acceptors (i.e., sugars, lactic/acetic acid and/or derivatives), in which it is appreciated how sucrose, lactose and glucose decrease as lactic and acetic acid, as well as their ethyl esters, together with lactamide, increase. Panels 3C-D show the time-course appearance of alcohols due to alcoholic fermentation made by yeasts. It can be appreciated that the major alcohol found in kefir, i.e., ethanol, undergoes higher changes in pasteurized milk as the fermentation progresses, because of the lower level in the pasteurized non fermented milk compared to raw unfermented milk. Finally, panels 3E-F show the evolution of some key metabolites participating in neurotransmission or other processes. Of them, tyramine shows the most pronounced change, as it increased ca. 8-fold in both raw and pasteurized milk after 12 h fermentation.
3.2. NIRS Analysis of Goat Milk Kefir
We also used the NIR spectroscopy technique for characterizing the goat milk kefir and for the discrimination between the studied variables, i.e., thermal treatment of milk and fermentation time. The NIR spectra obtained corresponded to the typical ones of dairy products, with two predominant peaks at 1,450 and 1,930 nm, which correspond to water (Fig. S1). These peaks are so intense that prevent other characteristic molecules to highlight in the NIR spectrum of this type of products with a high-water content. However, the PCA loading plots show areas of the spectrum in which molecular bonds frequently found in milk-based products are present, like acetic acid, at 1,678 nm, or lactic acid, at 1,950 nm (Fig. S2).
Next, we performed a PCA of the NIRS data. The representation of the two first principal components, i.e., PC1 and PC2, explained the 88.65% of the variance (Figure 4A). In these analyses, non-fermented milks were clearly differentiated from the fermented products, although there was not a clear separation among fermentation times and thermal treatment of milk when considering the fermented milks at different times. As PCA is a non-supervised analysis, we carried out a discriminant analysis of principal components (DAPC) once established the different categories of samples, i.e., according to the type of thermally treated milk, and according to the fermentation time, both using the two and the three first principal components (Figure 4B). This analysis showed a clear distinction among fermentation times, but it did not discriminate so precisely between raw and pasteurized milk, as for some time points, groups corresponding to the same fermentation time were mixed.
3.3. Predictive Capacity of Metabolomics and NIRS to Discriminate Kefir Samples
Finally, the global data obtained (separately for metabolomics and NIRS) were used to predict the discrimination of kefir samples, both according to the thermal treatment of milk and to the fermentation time, using LOOCV model-based confusion matrices (Figure 5). This is a way to calculate the accuracy of classification of real samples according to predicted categories, serving as crossed validation of the techniques used for the generation of data. Regarding the whole metabolomics data, the model obtained a 93.3% accuracy (i.e., predicted correctly the classification) for both type of milk and fermentation time. When combining both factors, i.e., type of thermal treatment and fermentation time of milk, the model reached a 86.7% of sample classification (Figure 5A). With respect to NIRS data, the model classified the samples with a 90% accuracy according to the fermentation time, but the accuracy for classifying according to the thermal treatment of milk was lower (68.3%). Considering both variables, the sample classification lowered to 78.3% (Figure 5B).
4. Discussion
In this work, we have performed the first metabolomic analysis of goat milk kefir, by combining different mass spectrometry-based techniques and NIRS for further differentiation of categories of the final product. Previous studies have focused on the characterization of kefir grains and the microbial composition of kefir [15,20,21] as well as in the identification of peptides derived from microbial digestion [9,10,11,12,13,14], but there is limited information on the metabolic changes that occur during milk fermentation by kefir grains. Although metabolomics has been widely applied to characterize other dairy products, like cheese or yogurt [19,22,23,24,25,26], to our knowledge most of the studies of metabolomic characterization of kefir published so far, either at a global or targeted level, have been carried out using cow milk [20,21,27,28]. Those involving goat milk kefir have been restricted to a limited group of compounds, mainly volatiles [29]. Therefore, our work provides the first insight into broader changes in the metabolite profiles of goat milk kefir, considering the fermentation time and different thermal treatments as factors affecting such variations.
Our metabolomic analysis identified 105 unique metabolites using different analytical techniques, revealing the complex biochemical composition of goat milk kefir. The significant changes in 86 metabolites with respect to fermentation time underscore the dynamic nature of kefir fermentation. Additionally, the differentiation between raw and pasteurized milk in the fermentation process, as evidenced by changes in 27 metabolites, suggests that milk treatment prior to fermentation can influence the metabolomic profile of kefir. The ability of NIRS to discriminate between types of milk and among fermentation times further demonstrates the potential of this technique in characterizing fermented foods.
Comparing our findings with existing literature reveals both consistencies and novelties in the metabolomic profile of goat milk kefir. Previous studies have similarly reported the dynamic changes in metabolites during kefir fermentation, reflecting the metabolic activities of the microbial community. However, our study extends the understanding by providing a detailed comparison between raw and pasteurized milk, highlighting how initial milk treatment can influence the fermentation outcome. This aspect has been less explored in previous research, offering a new perspective on the production of goat milk kefir. Microbes present in milk or those responsible for fermentation cause the appearance of volatile compounds conferring the characteristic flavor to dairy products [30,31]. Therefore, it is expected that killing of microorganisms by pasteurization will alter the metabolite profile.
By applying a comprehensive metabolomic analysis covering from non-polar to polar and volatile compounds, we found the major categories of metabolites described in cow milk kefir and in other dairy products: fatty acids; amino acids, sugars and other polar molecules; volatiles (alcohols, ketones, aldehydes, esters); organic acids; and other compounds, including polyols. Regarding fatty acids, these are compounds commonly found in fermented milks including yogurt and kefir of different species, as well as in cheese and butter [29,32]. Out of the 21 fatty acids identified in our work, 20 decreased during fermentation in both types of milk, as also described recently in cow milk kefir [15]. We also found that fermentation resulted in the release of single amino acids, as also described in cow milk kefir [15], which is clearly related to the protease activity of microbes present in the kefir grains. The gradual increase in the amino acid concentrations as the fermentation time progresses is in accordance with our previous findings of peptide release by a peptidomic approach [13]. Similar trends of sugars, alcohols, ketones, aldehydes and esters have been described in cow milk kefir [15,28], with some compounds increasing their concentrations during the fermentation time, and others decreasing. Interestingly, some compounds are exclusive of kefir fermentation due to the presence of yeast in the grains, as ethyl acetate or 3-methyl-1-butanol, as also described for cow milk kefir [28]. The main trend of organic acids found in our work was to decrease, as also described by Bourrie and cols. [28].
Other compounds identified in kefir have an already described bioactive profile. Thus, shikimic acid possesses antimicrobial activity against Staphylococcus aureus [33]. GABA is a neurotransmitter which has been also reported in cheese [25], with positive effects on insomnia, depression, the immune system, and blood pressure [34]. Dehydroabietic acid is a diterpenoid which increased up to 10-fold after 36 h of goat milk fermentation by kefir grains. This compound and its derivatives exhibit a wide range of bioactive properties, including antiviral, antitumor, wound regeneration, antimicrobial, or gastric protection [35]. Tyramine is a monoamine compound derived from the amino acid tyrosine. In foods, it is often produced by the decarboxylation of tyrosine during fermentation or decay. Fermented dairy products contain considerable amounts of this molecule. A large dietary intake of tyramine can increase blood pressure, so people taking monoamine oxidase inhibitors are recommended not to abuse from dairies to avoid possible hypertensive crises [36,37].
NIRS has been widely used for the characterization of foods, with multiple applications, including differentiation of products, maturation, or detection of fraudulent mixtures [38,39,40]. In dairies, its use is also habitual for the ease and cheapness of the technique compared to other massive analysis platforms [41,42,43,44]. In the present work, we have carried out a characterization of the kefir samples aiming at establishing differences between the two thermal treatments or fermentation times. The NIR spectra could distinguish with high accuracy the fermentation time variable, but the precision was lower when discriminating between raw and pasteurized milk. The NIRS data could be also integrated with metabolomics data, as the changes detected in the region 900-1,100 nm and in the large peak at 1,940 nm are associated to absorption changes mostly caused by N—H bonds. They may be related to the proteolytic activity of the microorganisms. This is supported by the metabolomics results obtained where an increase of free amino acids was observed as the fermentation time progresses, as already described previously at peptidomic level [13]. Also, changes in the 1,940 nm peak may be associated to alcohols. The changes observed in the final region of the spectrum between 2,000-2,400 nm would be caused by C—H bonds mostly attributed to lipids, in concordance with the decrease observed in almost all the fatty acid species detected over the fermentation time.
Limitations and Future Research
While our study provides valuable insights into the metabolomic profile of goat milk kefir, limitations exist, such as the need for validation of identified metabolites through targeted analysis. Future research should also explore the bioavailability and health effects of identified metabolites in clinical trials. Additionally, investigating the impact of fermentation variables, such as kefir grain composition and fermentation environment, could further elucidate the factors influencing the metabolomic profile of kefir.
5. Conclusions
This study represents the most comprehensive metabolomic analysis of goat milk kefir so far, revealing the intricate changes in metabolites during fermentation and the impact of milk treatment. Metabolomics achieves better classification results, but this requires simultaneous analysis of samples by several complex, high-cost and time-consuming analytical techniques. However, models obtained from NIRS data have a similar or lower classification capacity, although the analysis of samples by this technique is very fast, cheap and simple, which facilitates its implementation as a quality control tool. Our findings underscore the potential of goat milk kefir as a functional food, contributing to the understanding of its nutritional and therapeutic properties. By elucidating the metabolomic complexity of kefir, this research paves the way for future studies aimed at optimizing the health benefits of fermented foods. The identified metabolites, including significant changes in fatty acids and volatile and non-volatile compounds, have implications for the nutritional value and sensory properties of goat milk kefir. The differences observed between raw and pasteurized milk kefir suggest that selecting the appropriate milk treatment could tailor the health benefits and flavor profile of the final product. These findings contribute to the growing body of knowledge on fermented foods, supporting the development of functional foods that cater to specific health and dietary needs.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: NIRS spectra of goat milk kefir. A) Raw milk kefir; B) Pasteurized milk kefir; Figure S2: Principal component analysis (PCA) loading plots of the three first principal components of goat milk kefir; Table S1: Metabolite levels in kefir samples identified by GC-FID; Table S2: Metabolite levels in kefir samples identified by GC-QqQ-MS; Table S3: Metabolite levels in kefir samples identified by GC-MS-ToF using spectral libraries; Table S4: Metabolite levels in kefir samples identified by GC-MS-ToF using standards; Table S5: Metabolite levels in kefir samples identified by UHPLC-QToF-MS.
Author Contributions
Conceptualization, M.J.R.-O.; methodology, all the authors.; software, R.S.-R., J.G.-O., C.T. and S.O.; validation, all the authors; formal analysis, all the authors; investigation, R.S.-R., S.O. and M.J.R.-O.; resources, J.G.-O., S.O. and M.J.R.-O.; data curation, all the authors; writing—original draft preparation, M.J.R.-O.; writing—review and editing, all the authors; visualization, all the authors; supervision, S.O. and M.J.R.-O.; project administration, M.J.R.-O.; funding acquisition, not applicable. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
M.J.R.-O. is indebted to members of the AGR-164 group, headed by Prof. Jesús V. Jorrín-Novo, University of Córdoba, for lab support. GC-FID, GC-QqQ-MS, UHPLC-ToF-MS and NIRS analyses were carried out at the Central Facilities for Research Support (SCAI) of the University of Córdoba. GC-ToF-MS was performed at the Metabolomics Unit of the IHSM-CSIC-UMA.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Escareño, L.; Salinas-Gonzalez, H.; Wurzinger, M.; Iñiguez, L.; Sölkner, J.; Meza-Herrera, C. Dairy Goat Production Systems: Status Quo, Perspectives and Challenges. Trop. Anim. Health Prod. 2012, 45, 17–34. [Google Scholar] [CrossRef]
- Ministerio de Agricultura, Pesca y Alimentación Caracterización del sector ovino y caprino de leche en España; Madrid, Spain, 2022; https://www.mapa.gob.es/es/ganaderia/temas/produccion-y-mercados-ganaderos/caracterizacionovinoycaprinolechedatos2021_tcm30-562416.pdf.
- Prado, M.R.; Blandón, L.M.; Vandenberghe, L.P.S.; Rodrigues, C.; Castro, G.R.; Thomaz-Soccol, V.; Soccol, C.R. Milk Kefir: Composition, Microbial Cultures, Biological Activities, and Related Products. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Bourrie, B.C.T.; Willing, B.P.; Cotter, P.D. The Microbiota and Health Promoting Characteristics of the Fermented Beverage Kefir. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Farag, M.A.; Jomaa, S.A.; El-Wahed, A.A.; El-Seedi, A.H.R. The Many Faces of Kefir Fermented Dairy Products: Quality Characteristics, Flavour Chemistry, Nutritional Value, Health Benefits, and Safety. Nutrients 2020, 12, 346. [Google Scholar] [CrossRef] [PubMed]
- Azizi, N.F.; Kumar, M.R.; Yeap, S.K.; Abdullah, J.O.; Khalid, M.; Omar, A.R.; Osman, M.A.; Mortadza, S.A.S.; Alitheen, N.B. Kefir and Its Biological Activities. Foods Basel Switz. 2021, 10, 1210. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.; Mora García, M.B. A 100-Year Review: Advances in Goat Milk Research. J. Dairy Sci. 2017, 100, 10026–10044. [Google Scholar] [CrossRef]
- Prosser, C.G. Compositional and Functional Characteristics of Goat Milk and Relevance as a Base for Infant Formula. J. Food Sci. 2021, 86, 257–265. [Google Scholar] [CrossRef]
- Ebner, J.; Aşçı Arslan, A.; Fedorova, M.; Hoffmann, R.; Küçükçetin, A.; Pischetsrieder, M. Peptide Profiling of Bovine Kefir Reveals 236 Unique Peptides Released from Caseins during Its Production by Starter Culture or Kefir Grains. J. Proteomics 2015, 117, 41–57. [Google Scholar] [CrossRef]
- Dallas, D.C.; Citerne, F.; Tian, T.; Silva, V.L.M.; Kalanetra, K.M.; Frese, S.A.; Robinson, R.C.; Mills, D.A.; Barile, D. Peptidomic Analysis Reveals Proteolytic Activity of Kefir Microorganisms on Bovine Milk Proteins. Food Chem. 2016, 197, 273–284. [Google Scholar] [CrossRef]
- Amorim, F.G.; Coitinho, L.B.; Dias, A.T.; Friques, A.G.F.; Monteiro, B.L.; Rezende, L.C.D.D.; Pereira, T.D.M.C.; Campagnaro, B.P.; De Pauw, E.; Vasquez, E.C.; et al. Identification of New Bioactive Peptides from Kefir Milk through Proteopeptidomics: Bioprospection of Antihypertensive Molecules. Food Chem. 2019, 282, 109–119. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, X.; Sun, Y.; Sun, X.; Guo, M. Differences in Protein Profiles of Kefir Grains from Different Origins When Subcultured in Goat Milk. J. Agric. Food Chem. 2022, 70, 7515–7524. [Google Scholar] [CrossRef]
- Izquierdo-González, J.J.; Amil-Ruiz, F.; Zazzu, S.; Sánchez-Lucas, R.; Fuentes-Almagro, C.A.; Rodríguez-Ortega, M.J. Proteomic Analysis of Goat Milk Kefir: Profiling the Fermentation-Time Dependent Protein Digestion and Identification of Potential Peptides with Biological Activity. Food Chem. 2019, 295, 456–465. [Google Scholar] [CrossRef]
- Dalabasmaz, S.; De La Torre, E.P.; Gensberger-Reigl, S.; Pischetsrieder, M.; Rodríguez-Ortega, M.J. Identification of Potential Bioactive Peptides in Sheep Milk Kefir through Peptidomic Analysis at Different Fermentation Times. Foods 2023, 12, 2974. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, X.; Yue, Y.; Du, G.; Chen, H.; Ning, M.; Yuan, Y.; Yue, T. Metagenomic Features of Tibetan Kefir Grains and Its Metabolomics Analysis during Fermentation. LWT 2023, 175, 114502. [Google Scholar] [CrossRef]
- Qin, C.; Liu, L.; Wang, Y.; Leng, T.; Zhu, M.; Gan, B.; Xie, J.; Yu, Q.; Chen, Y. Advancement of Omics Techniques for Chemical Profile Analysis and Authentication of Milk. Trends Food Sci. Technol. 2022, 127, 114–128. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.H.S. A SIMPLE METHOD FOR THE ISOLATION AND PURIFICATION OF TOTAL LIPIDES FROM ANIMAL TISSUES. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Osorio, S.; Do, P.T.; Fernie, A.R. Profiling Primary Metabolites of Tomato Fruit with Gas Chromatography/Mass Spectrometry. In Plant Metabolomics; Hardy, N.W., Hall, R.D., Eds.; Methods in Molecular Biology; Humana Press: Totowa, NJ, 2011; ISBN 978-1-61779-593-0. [Google Scholar]
- Settachaimongkon, S.; Van Valenberg, H.J.F.; Winata, V.; Wang, X.; Nout, M.J.R.; Van Hooijdonk, T.C.M.; Zwietering, M.H.; Smid, E.J. Effect of Sublethal Preculturing on the Survival of Probiotics and Metabolite Formation in Set-Yoghurt. Food Microbiol. 2015, 49, 104–115. [Google Scholar] [CrossRef]
- Batista, L.L.; Malta, S.M.; Guerra Silva, H.C.; Borges, L.D.F.; Rocha, L.O.; Da Silva, J.R.; Rodrigues, T.S.; Venturini, G.; Padilha, K.; Da Costa Pereira, A.; et al. Kefir Metabolites in a Fly Model for Alzheimer’s Disease. Sci. Rep. 2021, 11, 11262. [Google Scholar] [CrossRef]
- Walsh, A.M.; Crispie, F.; Kilcawley, K.; O’Sullivan, O.; O’Sullivan, M.G.; Claesson, M.J.; Cotter, P.D. Microbial Succession and Flavor Production in the Fermented Dairy Beverage Kefir. mSystems 2016, 1, e00052-16. [Google Scholar] [CrossRef] [PubMed]
- Pisano, M.B.; Scano, P.; Murgia, A.; Cosentino, S.; Caboni, P. Metabolomics and Microbiological Profile of Italian Mozzarella Cheese Produced with Buffalo and Cow Milk. Food Chem. 2016, 192, 618–624. [Google Scholar] [CrossRef] [PubMed]
- Caboni, P.; Maxia, D.; Scano, P.; Addis, M.; Dedola, A.; Pes, M.; Murgia, A.; Casula, M.; Profumo, A.; Pirisi, A. A Gas Chromatography-Mass Spectrometry Untargeted Metabolomics Approach to Discriminate Fiore Sardo Cheese Produced from Raw or Thermized Ovine Milk. J. Dairy Sci. 2019, 102, 5005–5018. [Google Scholar] [CrossRef] [PubMed]
- Murgia, A.; Scano, P.; Cacciabue, R.; Dessì, D.; Caboni, P. GC-MS Metabolomics Comparison of Yoghurts from Sheep’s and Goats’ Milk. Int. Dairy J. 2019, 96, 44–49. [Google Scholar] [CrossRef]
- Scano, P.; Cagliani, L.R.; Consonni, R. 1H NMR Characterisation of the Lipid Fraction and the Metabolite Profiles of Fossa (Pit) Cheese. Int. Dairy J. 2019, 90, 39–44. [Google Scholar] [CrossRef]
- Settachaimongkon, S.; Nout, M.J.R.; Antunes Fernandes, E.C.; Hettinga, K.A.; Vervoort, J.M.; Van Hooijdonk, T.C.M.; Zwietering, M.H.; Smid, E.J.; Van Valenberg, H.J.F. Influence of Different Proteolytic Strains of Streptococcus Thermophilus in Co-Culture with Lactobacillus Delbrueckii Subsp. Bulgaricus on the Metabolite Profile of Set-Yoghurt. Int. J. Food Microbiol. 2014, 177, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Beshkova, D.M.; Simova, E.D.; Frengova, G.I.; Simov, Z.I.; Dimitrov, Zh.P. Production of Volatile Aroma Compounds by Kefir Starter Cultures. Int. Dairy J. 2003, 13, 529–535. [Google Scholar] [CrossRef]
- Bourrie, B.C.T.; Diether, N.; Dias, R.P.; Nam, S.L.; De La Mata, A.P.; Forgie, A.J.; Gaur, G.; Harynuk, J.J.; Gänzle, M.; Cotter, P.D.; et al. Use of Reconstituted Kefir Consortia to Determine the Impact of Microbial Composition on Kefir Metabolite Profiles. Food Res. Int. 2023, 173, 113467. [Google Scholar] [CrossRef] [PubMed]
- Cais-Sokolińska, D.; Wójtowski, J.; Pikul, J.; Danków, R.; Majcher, M.; Teichert, J.; Bagnicka, E. Formation of Volatile Compounds in Kefir Made of Goat and Sheep Milk with High Polyunsaturated Fatty Acid Content. J. Dairy Sci. 2015, 98, 6692–6705. [Google Scholar] [CrossRef]
- Guzel-Seydim, Z.; Seydim, A.C.; Greene, A.K. Organic Acids and Volatile Flavor Components Evolved During Refrigerated Storage of Kefir. J. Dairy Sci. 2000, 83, 275–277. [Google Scholar] [CrossRef]
- Zhao, M.; Ma, H.; Hou, Y.; Li, J.; Zou, T.; Zhang, D.; Wen, R.; Li, H.; Song, H. Characterization of Key Odor-Active Off-Flavor Compounds in Aged Pasteurized Yogurt by Sensory-Directed Flavor Analysis. J. Agric. Food Chem. 2022, 70, 14439–14447. [Google Scholar] [CrossRef]
- Silva, T.; Pires, A.; Gomes, D.; Viegas, J.; Pereira-Dias, S.; Pintado, M.E.; Henriques, M.; Pereira, C.D. Sheep’s Butter and Correspondent Buttermilk Produced with Sweet Cream and Cream Fermented by Aromatic Starter, Kefir and Probiotic Culture. Foods 2023, 12, 331. [Google Scholar] [CrossRef]
- Bai, J.; Wu, Y.; Liu, X.; Zhong, K.; Huang, Y.; Gao, H. Antibacterial Activity of Shikimic Acid from Pine Needles of Cedrus Deodara against Staphylococcus Aureus through Damage to Cell Membrane. Int. J. Mol. Sci. 2015, 16, 27145–27155. [Google Scholar] [CrossRef]
- Cui, Y.; Miao, K.; Niyaphorn, S.; Qu, X. Production of Gamma-Aminobutyric Acid from Lactic Acid Bacteria: A Systematic Review. Int. J. Mol. Sci. 2020, 21, 995. [Google Scholar] [CrossRef]
- Hao, M.; Xu, J.; Wen, H.; Du, J.; Zhang, S.; Lv, M.; Xu, H. Recent Advances on Biological Activities and Structural Modifications of Dehydroabietic Acid. Toxins 2022, 14, 632. [Google Scholar] [CrossRef]
- Gillman, P.K. A Reassessment of the Safety Profile of Monoamine Oxidase Inhibitors: Elucidating Tired Old Tyramine Myths. J. Neural Transm. Vienna Austria 1996 2018, 125, 1707–1717. [Google Scholar] [CrossRef]
- Salter, M.; Kenney, A. Myocardial Injury from Tranylcypromine-Induced Hypertensive Crisis Secondary to Excessive Tyramine Intake. Cardiovasc. Toxicol. 2018, 18, 583–586. [Google Scholar] [CrossRef]
- Windarsih, A.; Rohman, A.; Irnawati; Riyanto, S. The Combination of Vibrational Spectroscopy and Chemometrics for Analysis of Milk Products Adulteration. Int. J. Food Sci. 2021, 2021, 1–15. [Google Scholar] [CrossRef]
- Xu, L.; Yan, S.-M.; Cai, C.-B.; Wang, Z.-J.; Yu, X.-P. The Feasibility of Using Near-Infrared Spectroscopy and Chemometrics for Untargeted Detection of Protein Adulteration in Yogurt: Removing Unwanted Variations in Pure Yogurt. J. Anal. Methods Chem. 2013, 2013, 1–9. [Google Scholar] [CrossRef]
- Gorla, G.; Fumagalli, S.; Jansen, J.J.; Giussani, B. Acquisition Strategies for Fermentation Processes with a Low-Cost Miniaturized NIR-Spectrometer from Scratch: Issues and Challenges. Microchem. J. 2022, 183, 108035. [Google Scholar] [CrossRef]
- Grassi, S.; Alamprese, C.; Bono, V.; Picozzi, C.; Foschino, R.; Casiraghi, E. Monitoring of Lactic Acid Fermentation Process Using Fourier Transform near Infrared Spectroscopy. J. Infrared Spectrosc. 2013, 21, 417–425. [Google Scholar] [CrossRef]
- Ntsame Affane, A.L.; Fox, G.P.; Sigge, G.O.; Manley, M.; Britz, T.J. Quantitative Analysis of DL-Lactic Acid and Acetic Acid in Kefir Using near Infrared Reflectance Spectroscopy. J. Infrared Spectrosc. 2009, 17, 255–264. [Google Scholar] [CrossRef]
- Ntsame Affane, A.L.; Fox, G.P.; Sigge, G.O.; Manley, M.; Britz, T.J. Simultaneous Prediction of Acidity Parameters (pH and Titratable Acidity) in Kefir Using near Infrared Reflectance Spectroscopy. Int. Dairy J. 2011, 21, 896–900. [Google Scholar] [CrossRef]
- Shao, Y.; He, Y. Measurement of Soluble Solids Content and pH of Yogurt Using Visible/Near Infrared Spectroscopy and Chemometrics. Food Bioprocess Technol. 2009, 2, 229–233. [Google Scholar] [CrossRef]
Figure 1.
Score plots of three-dimensional principal component analysis (PCA, panel A) and discriminant analysis of principal components (DAPC, panel B) of all the metabolites identified using different metabolomics platforms in goat milk kefir.
Figure 1.
Score plots of three-dimensional principal component analysis (PCA, panel A) and discriminant analysis of principal components (DAPC, panel B) of all the metabolites identified using different metabolomics platforms in goat milk kefir.
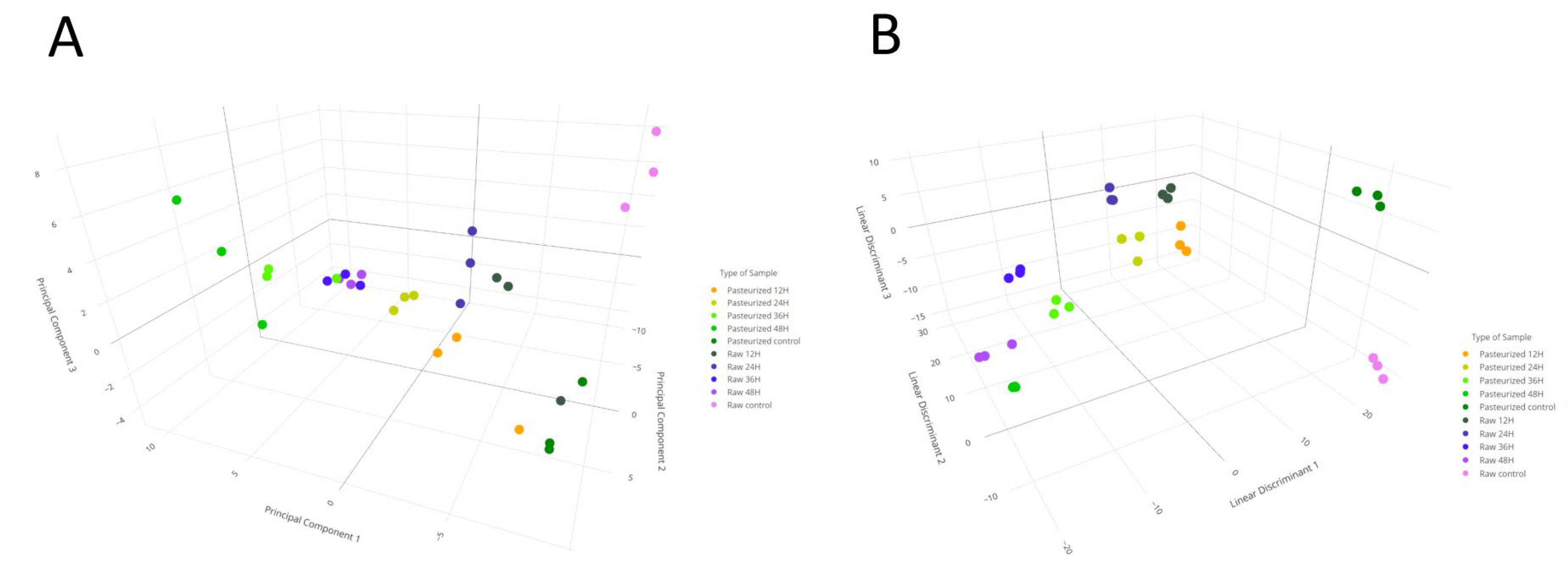
Figure 2.
Heatmap visualizations of differences in metabolites identified in goat milk kefir with the different platforms: A) GC-FID for fatty acids; B) Headspace GC-QqQ-MS for volatile compounds; C) UHPLC-QToF-MS for non-volatile compounds using commercial standards for identification; D) GC-ToF-MS for non-volatile compounds using commercial standards for identification; and E) GC-ToF-MS for non-volatile compounds using spectra libraries for identification.
Figure 2.
Heatmap visualizations of differences in metabolites identified in goat milk kefir with the different platforms: A) GC-FID for fatty acids; B) Headspace GC-QqQ-MS for volatile compounds; C) UHPLC-QToF-MS for non-volatile compounds using commercial standards for identification; D) GC-ToF-MS for non-volatile compounds using commercial standards for identification; and E) GC-ToF-MS for non-volatile compounds using spectra libraries for identification.
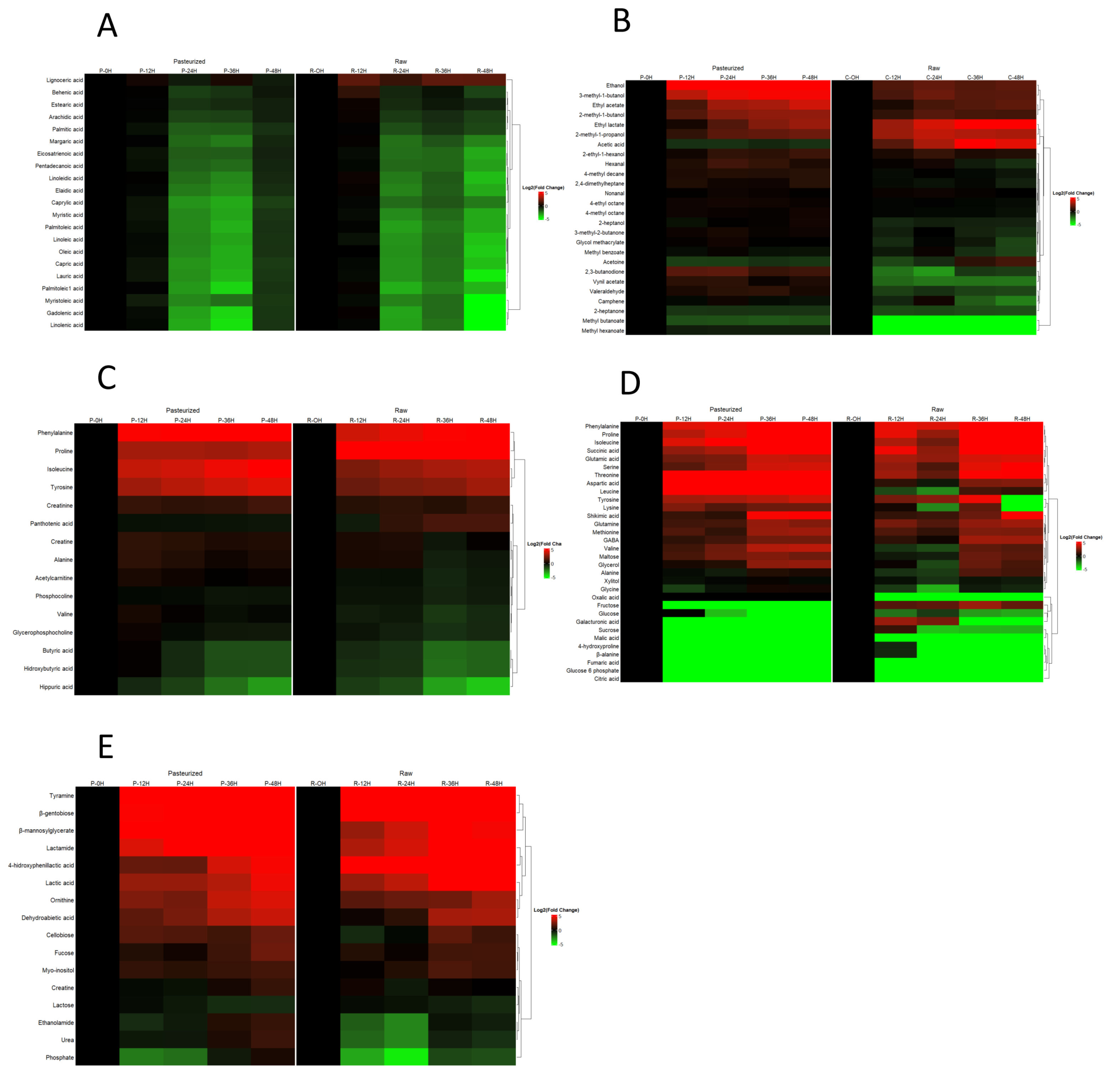
Figure 3.
Time-course evolution of selected metabolites in raw (panels A, C and E) and pasteurized goat milk kefir (panels B, D and F).
Figure 3.
Time-course evolution of selected metabolites in raw (panels A, C and E) and pasteurized goat milk kefir (panels B, D and F).
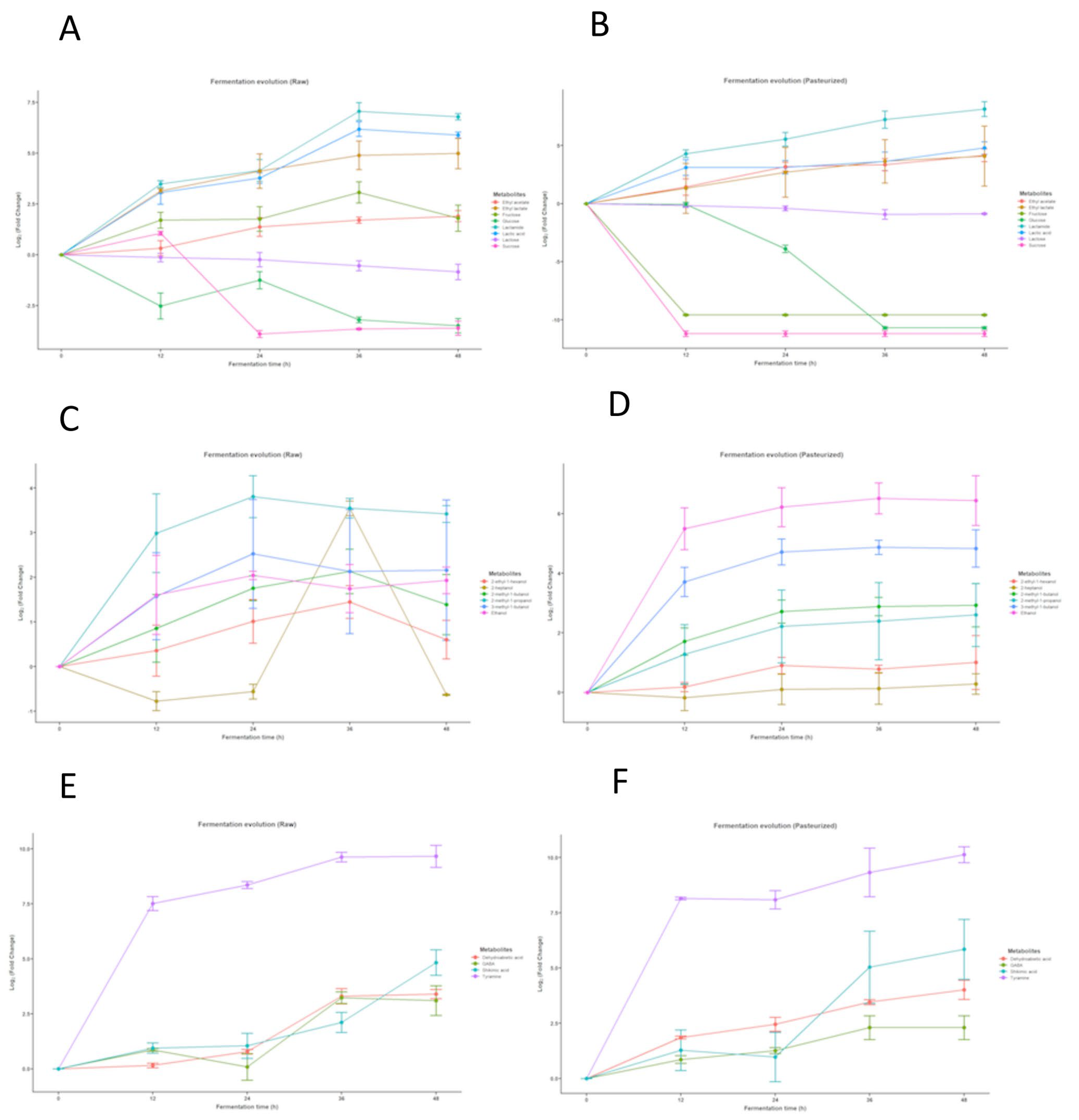
Figure 4.
Score plots of two-dimensional principal component analysis (PCA, panel A) and discriminant analysis of principal components (DAPC, panel B) of NIRS data in goat milk kefir.
Figure 4.
Score plots of two-dimensional principal component analysis (PCA, panel A) and discriminant analysis of principal components (DAPC, panel B) of NIRS data in goat milk kefir.
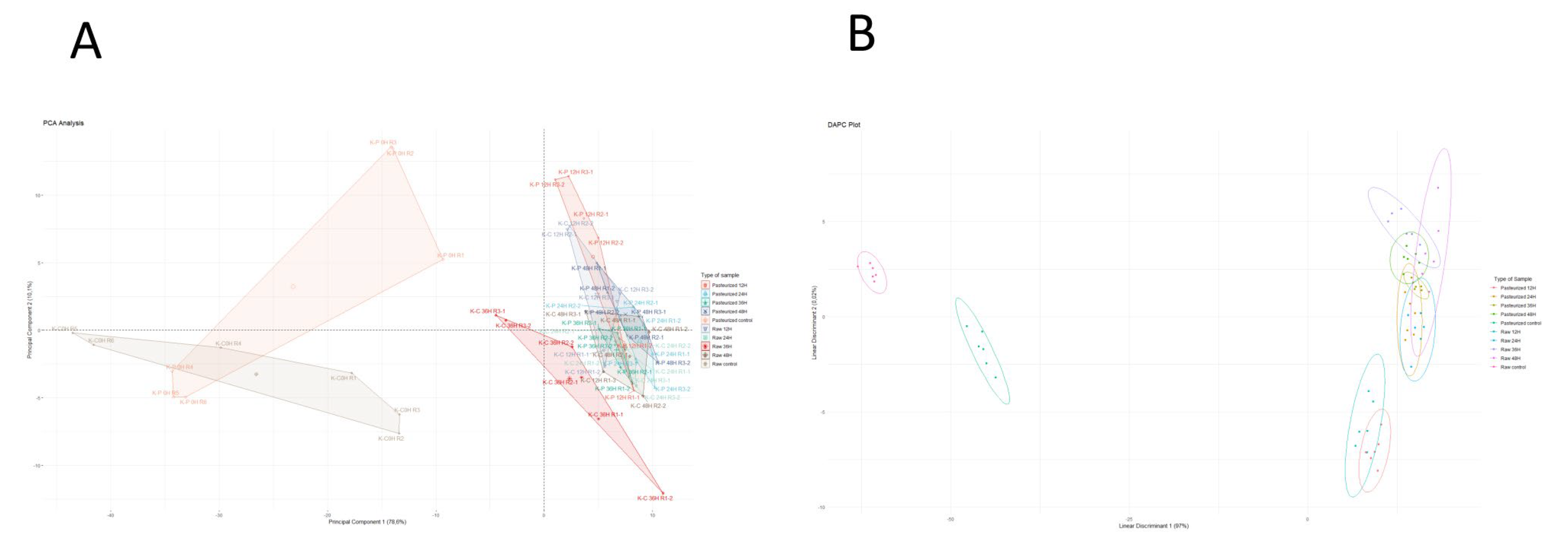
Figure 5.
Confusion matrix using LOOCV to evaluate the performance of the model classifying goat milk kefir samples according to fermentation time and thermal treatment of milk. The numbers within each square represents the assigned class by the model. The sample is correctly classified if the real class corresponds with the predicted class (diagonal). A) Confusion matrix for metabolomics data; B) Confusion matrix for NIRS data.
Figure 5.
Confusion matrix using LOOCV to evaluate the performance of the model classifying goat milk kefir samples according to fermentation time and thermal treatment of milk. The numbers within each square represents the assigned class by the model. The sample is correctly classified if the real class corresponds with the predicted class (diagonal). A) Confusion matrix for metabolomics data; B) Confusion matrix for NIRS data.
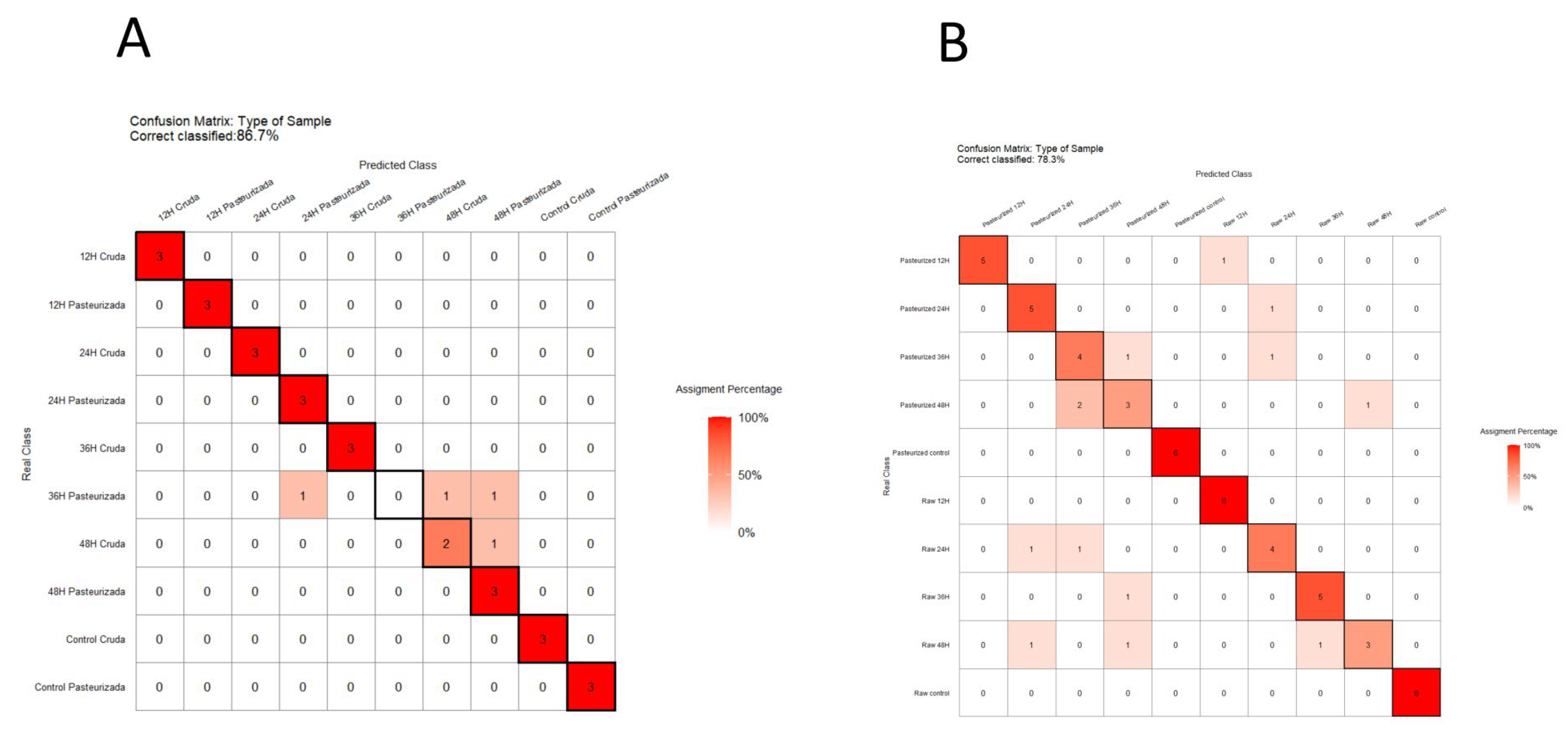

Table 1.
Metabolites detected in goat milk kefir with all the metabolomics techniques.
| Technique | Metabolites detected | Name |
|---|---|---|
| GC-FID | 21 | Arachidic acid, behenic acid, capric acid, caprylic acid, eicosatrienoic acid, elaidic acid, estearic acid, gadolenic acid, lauric acid, lignoceric acid, linoleic acid, linoleidic acid, linolenic acid, margaric acid, myristic acid, myristoleic acid, oleic acid, palmitic acid, palmitoleic acid, palmitoleic1 acid, pentadecanoic acid |
| UHPLC-MS (QToF) | 15 | Acetylcarnitine, alanine, butyric acid, creatine, creatinine, Glycerophosphocholine, hippuric acid, hidroxybutyric acid, isoleucine, panthotenic acid, fenilalanine, tyrosine, proline, phosphocoline, valine |
| GC-MS (QqQ) | 26 | 2-methyl-1-butanol,2-methyl-1-propanol, 3-methyl-1-butanol, 2,3-butanodione, 2,4-dimethylheptane, 2-ethyl-1-hexanol, 2-heptanol, 2-heptanone, 3-methyl-2-butanone, 4-ethyl octane, 4-methyl decane, 4-methyl octane, ethanol, ethyl acetate, vynil acetate, acetoine, acetic acid, methyl benzoate, methyl butanoate, camphene, glycol methacrylate, hexanal, mehtyl hexanoate, ethyl lactate, nonanal, valeraldehyde |
| GC-MS (Tof) | 32 | 4-hidroxyproline, aspartic acid, citric acid, fumaric acid, galacturonic acid, glutamic acid, malic acid, oxalic acid, shikimic acid, succinic acid, alanine, phenylalanine, fructose, GABA, glycerol, glycine, glucose, glucose-6-phosphate, glutamine, isoleucine, leucine, lysine, maltose, methionine, proline, sucrose, serine, tyrosine, threonine, valine, xylitol, β-alanine |
| 16 | 4-hidroxyphenillactic acid, dehidroabietic acid, lactic acid, celobiose, creatine, ethanolamide, phosphate, fucose, lactamide, lactose, myo-inositol, ornithine, tyramine, urea, β-gentobiose, β-mannosylglycerate |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



