
Submitted:
11 June 2024
Posted:
12 June 2024
You are already at the latest version
Abstract
This study assessed the impact of inoculation with plant growth-promoting bacteria (PGPB) on stress-resistance and growth-promoting metabolites in Salicornia europaea L. across controlled and field conditions. Salicornia europaea seeds, inoculated with Brevibacterium casei EB3 and Pseudomonas oryzihabitans RL18, were grown in controlled laboratory experiments and in a nat-ural field setting. Metabolite composition was analyzed using GC-MS and UHPLC-MS. Under controlled conditions, PGPB inoculation significantly increased levels of unsaturated fatty acids, sugars, citric acid, acetic acid, chlorogenic acids, and quercetin. Inoculation under field condi-tions increased glucose, fructose, quercetin, and apigenin, alongside decreased unsaturated fatty acids and sterols, indicating a stress response shift in plant metabolism. Under controlled condi-tions, enhanced metabolite profiles induced by inoculation likely contributed to augmented bio-mass production and environmental stress resistance. Field plants revealed distinct phytochemi-cal profiles, suggesting higher stress levels. However, the accumulated metabolites from inocula-tion could enhance the plant's ability to cope with adverse conditions and improving stress tol-erance. This study unveils the intricate metabolic adaptations of Salicornia europaea under con-trolled and field stress conditions, highlighting PGPB's potential to enhance plant stress tolerance. Elevated stress-related metabolites may fortify plant defense mechanisms, laying the ground-work for stress-resistant crop development through PGPB-based inoculants, especially in saline agriculture.
Keywords:
plant-growth-promoting bacteria
; plant metabolites
; stress attenuation
; UHPLC-MS
; GC-MS
; Brevibacterium case
; Pseudomonas oryzihabitans
1. Introduction
Plant growth-promoting halotolerant bacteria have been extensively studied to make agriculture more sustainable, reducing the need for chemical fertilizers and allowing the use and recovery of salinized and contaminated soils. Bacterial plant-growth traits can (i) increase the plant nutrient uptake by phosphate and potassium solubilization, atmospheric nitrogen fixation, and production of siderophores for iron uptake, (ii) control pathogens by releasing antimicrobial compounds, (iii) regulate plant growth through the production of phytohormone-like molecules, and (iv) reduce abiotic stress through the production of enzymes such as 1-aminocyclopropane-1-carboxylate (ACC) deaminase and exopolysaccharides [1]. Plant inoculation with plant growth-promoting bacteria (PGPB) can increase their growth and resilience to abiotic and biotic stresses [2,3,4]. This symbiotic relationship reinforces the plant’s immune system, adding protection against phytopathogens [5]. The halotolerant PGPBs also enhance the phytoremediation potential regarding contaminated and salinized soils [6,7,8].
Inoculation influences the plant’s primary and secondary metabolism [9,10]. Primary metabolites of plants, like carbohydrates, proteins, lipids, and amino acids, play critical roles in biomass acquisition, energy transfer, and signaling [11,12,13]. Secondary metabolites, phenolic acids, flavonoids, terpenes, and sterols, though not essential to survival, increase the plant’s competitive ability [12,14,15].
Stress-protective pathways can be activated by bacterial inoculation that will reshape the plant metabolome and elemental composition [16,17,18,19]. The metabolic enrichment of plant tissues with bioactive compounds due to PGPB inoculation may enhance their economic value for the food and pharmaceutical industries [20]. Additionally, the enrichment patterns may offer insights into the mechanisms of plant-bacteria interactions [21].
It was recently reported that PGPB inoculation of Salicornia europaea boost beneficial bacteria-plant interactions within the rhizosphere [22] and produced a significant impact on the plants’ primary metabolism, favouring biomass production [23]. Thus, we hypothesized that S. europaea inoculation with PGPB might reshape the plants’ secondary biochemical profile to promote both growth and stress resistance.
In this way, this study aimed to investigate the impact of rhizosphere engineering with PGPBs on the expression of stress resistance and growth-promoting secondary metabolites in Salicornia europaea L. under both controlled and field conditions.
2. Results
Gas chromatography-Mass spectrometry (GC-MS) and ultra-high-performance liquid chromatography-mass spectrometry (UHPLC-MS) analyses revealed differences in the metabolic profile between plants inoculated with Brevibacterium casei EB3 and Pseudomonas oryzihabitans RL18 and non-inoculated plants, in both microcosm and field experiments.
2.1. Plant Biochemical Profile – Microcosm Experiments
In the microcosm experiment, considering both GC- and UHPLC-MS analyses, a total of 62 compounds were identified: 55 compounds in non-inoculated plants, and 58 in inoculated plants (Supplementary Tables S2, S4 and S6). Due to a lack of standard, seven compounds detected by GC-MS were not quantified (neophytadiene, oleanitrile, N,N-dimethylglycine, uridine, diacetone alcohol, glyceryl-glycoside, and 7-methylundec-4-ene).
Sugars were the most prevalent compounds, while sterols were the least. Inoculated plants contained more sugars (32.57±0.31 mg.g-1 DW), unsaturated fatty acids (UFA, 3.34±0.86 mg.g-1 DW) and lower amounts of flavonoids (2.69± 0.05 mg.g-1 DW), compared to non-inoculated control plants (15.48±1.26 mg.g-1 DW, 1.60±0.07 mg.g-1 DW and (8.48± 0.45 mg.g-1 DW, respectively) (Figure 1).
Detailed characterisation of the chemical composition (Supplementary Tables S2, S4 and S6) showed that inoculated plants accumulated significantly higher concentrations of some carboxylic and sugar acids, namely acetic, citric, and tartaric acids, compared to control plants. Conversely, the concentration of glyceric and ribonic acids was significantly lower in test plants compared to non-inoculated controls.
The fatty acid profile showed a significant enrichment of the unsaturated oleic acid and the saturated stearic acid in inoculated plants. Inoculation also resulted in an accumulation of the sterols β-sitosterol, detected only in inoculated plants. A wide variety of sugars was detected, among which the most represented were sucrose in inoculated plants and D-glucose in non-inoculated controls. Significant differences were observed only for xylose, absent in inoculated plants, and sucrose, enriched in the latter.
The alcohols glycerol, myo-inositol and butane-1,3-diol were enriched in control plants (p<0.05), whereas xylitol and 2-methylbutane-1,3-diol were detected exclusively in inoculated plants.
The phenolic acid hydroxybenzoic acid and the flavonoids gallocatechin, and catechin were found only in non-inoculated plants (p<0.05) which were also enriched in formononetin. In contrast, the phenolic acids caffeic acid, quinic acid, and the flavonoid quercetin were detected only in inoculated plants, which were also enriched in caffeoylquinic acids.
The fold changes in the concentration of metabolites for which significant differences between inoculated and non-inoculated plants, expressed as Log2 [(EB3+RL18)/NI], were detected are summarized in Figure 2. The heterogeneous response profiles to inoculation are visible within each group of metabolites, were positive and negative responses can be observed. The profile response of sugar acids and flavonoids, especially catechin, gallocatechin and formononetin) was negative, characterized by a decrease of these metabolites with inoculation, whereas the profile response of carboxylic acids (malic, citric and acetic acids), fatty acids (especially unsaturated fatty acids), sucrose, ß-sitosterol, phenolic acids and quercetin was positive, representing an increase of these metabolites in inoculated plants.
2.2. Plant Biochemical Profile – Field Experiments
In the field experiments, 54 distinct compounds were identified in plant material by GC-MS and UHPLC-Ms analysis (Supplementary Tables S3, S5 and S7), 46 in non-inoculated plants and 48 in inoculated plants. Four of the detected compounds, could not be quantified due to the lack of appropriate standards (oleanitrile, glycine, glyceryl-glycoside, and 7-methylundec-4-ene).
In field-grown plants, as in the microcosm experiments, sugars were the compounds that most contributed to the dry weight, and sterols contributed the least. Inoculated plants were enriched in sugars (26.9±2.25 mg.g-1 DW), sugar acids (1.3±0.02 mg.g-1 DW), sterols (0.3±0.04 mg.g-1 DW), phenolic acids (7.7+1.19 mg.g-1 DW ) and flavonoids (6.0±0.98mg.g-1 DW), compared to non-inoculated plants (15.2±1.13, 0.8±0.13, 0.1±0.08, 2.6±0.27 and 1.0±0.12 mg.g-1 DW, respectively) (Figure 3)..
The detailed composition of field plants (Supplementary Tables S3, S5 and S7) revealed that all six carboxylic acids were detected in similar concentrations in inoculated and non-inoculated plants. Ribonic acid (sugar acid) was detected only in the inoculated plants, while glyceric acid was found only in non-inoculated controls. The fatty acid profile showed a significant enrichment in linoleic acid, palmitic and lignoceric acids in inoculated plants. Only two sterols, cholesterol and stigmasterol, were detected. Stigmasterol was found to be enriched in inoculated plants (p<0.05).
The sugars fructose, psicose, mannose, allose, glucose, and sucrose were detected in higher concentrations in inoculated plants. In contrast, ribose was only detected in the controls. Similarly, to the microcosm-grown plants, sucrose was the predominant sugar in both experimental conditions. Of the five alcohols detected by GC-MS, docosan-1-ol was overexpressed in non-inoculated plants. The remaining alcohols were represented in similar concentrations in either experimental condition. In the amide pool, only octanamide was detected in inoculated plants while only dodecanamide was identified in control plants. Contrary to the microcosm experiment, terpenoids were not detected. In the field experiment, the amino acid glycine was detected exclusively in inoculated plants.
The phenolic acids 3-p-coumaroylquinic acid and p-coumaric acid were detected only in non-inoculated plants. Hydroxybenzoic acid, quercetin, and apigenin were detected only in inoculated plants.
The fold changes in the concentration of metabolites for which significant differences between inoculated and non-inoculated plants, expressed as Log2 [(EB3+RL18)/NI], were detected are represented in Figure 4. As in the microcosm experiments, inoculation resulted in both negative and positive response profiles, within each group of metabolites. The profile response of flavonoids (especially quercetin, apigenin and formononetin) and the phenolic acids caffeic, quinic and hydroxybenzoic acids was positive, characterized by an increase of these metabolites in inoculated plants. Similarly, fructose and myo-inositol also showed a substantial increase, as well as lignoceric and oleic acids. Interesting positive changes were also observed in the content of carboxylic acids, namely lactic, succinic, malic and citric acids.
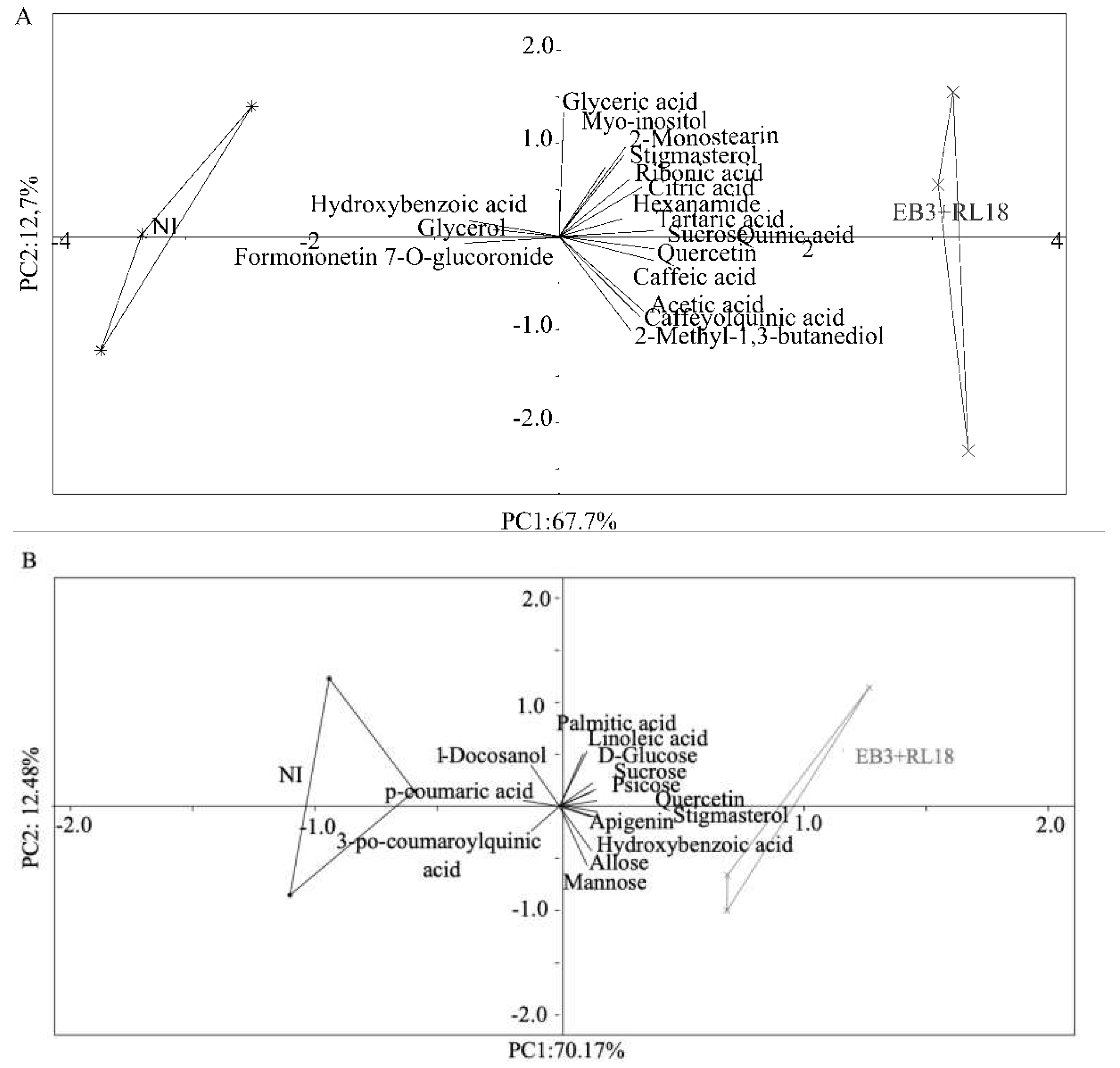
A Principal Component Analysis (PCA) indicated that the 2 first axes explained 80.4% (Figure 5A) and 82.65% (Figure 5B) of total data variability of microcosm and the field experiments, respectively. In each experiment, experimental conditions (inoculated tests and non-inoculated controls) clearly separate along axis 1, which explains 67.7% (microcosm experiment) and 70.17% (field experiment) of the total variability.
Metabolites sucrose, quercetin, tartaric acid, and caffeic acid were strongly correlated with inoculated plants both in the microcosm experiment, while quercetin, apigenin, psicose, and stigmasterol were more correlated with inoculated filed plants. In contrast, hydroxybenzoic acid and formononetin showed the highest correlation with non-inoculated plants from the microcosm experiment.
3. Discussion
The halophyte Salicornia europaea is known for its potential as a versatile plant and model halophyte crop for saline soils and aquaponic systems. It also holds promise for unique interesting secondary metabolic compounds [23,24,25]. These unique characteristics have sparked considerable interest in the plant, leading to a growing number of studies exploring its biochemical potential, particularly for the pharmaceutical industry [26,27,28]. Brevibacterium casei EB3 and Pseudomonas oryzihabitans RL18 have been identified as effective agents in enhancing the growth of S. europaea. Their influence is associated with a restructuring of primary metabolic pathways, favouring biomass production [23]. Given the insights gained from primary metabolism studies, significant alterations in secondary metabolism are anticipated following inoculation. However, the impact of bacterial inoculation on the profile of secondary metabolites remains largely unexplored, which prompted the undertaking of the present study. The objective was to investigate the influence of growth conditions and co-inoculation with PGPB, specifically B. casei EB3 and P. oryzihabitans RL18, on the metabolite profiles of Salicornia europaea.
The phytochemical profile was characterised by GC-MS and UHPLC-MS analyses of extracts from the plant’s aerial part. As a general trend, in microcosm experiments, inoculated plants exhibited enhanced sugar content, and unsaturated fatty acids compared to non-inoculated counterparts. Interestingly, these plants had lower amounts of flavonoids. In field conditions, in addition to containing more flavonoids, inoculated plants were also enriched in phenolic acids, sugar acids, sterols, and sugars.
3.1. The Impact of Inoculation on Metabolite Profile and Plant Growth
PGPB are associated with several positive effects on plant health and phytochemical composition. In this work, both microcosm-grown and field plants inoculated with the selected PGPB, exhibited enriched sugar content. Sugars, including sucrose, glucose, fructose, and trehalose, play diverse roles beyond energy provision, acting as signalling molecules that interact with phytohormones to enhance plant defence mechanisms and promote growth, sometimes prioritizing defence against stress over growth [29,30,31,32,33]. Elevated sucrose levels have been linked to delayed flowering, extending the vegetative phase and prolonging the harvest period for human consumption [34]. Additionally, sucrose exudation from plant roots serves as an osmoprotectant and regulates beneficial rhizobacterial communities by alleviating water and salt stress [35,36,37].
Similarly, fructose, which also accumulated significantly in field-inoculated plants, enhances the plant’s ability to tolerate long-term stress, including chronic drought and high salinity, by activating genes involved in defence mechanisms [38,39].
Elevated glucose levels, observed in inoculated field plants, have been associated with an extended juvenile phase, with reduced growth rates [40], which can help explain the lack of growth enhancement published in a previous study [23], concerning the same plants, where no significant difference in growth was observed between field-inoculated and control plants.
Despite inoculation caused field plants to accumulate psicose, allose, mannose, glucose and sucrose, microcosm plants only showed significant sucrose accumulation. Nonetheless, a metabolic shift towards monosaccharide production was apparent in all inoculated plants. While sucrose showed a strong correlation with microcosm-grown inoculated plants, psicose exhibited a similar relationship with field-inoculated plants.
Studies on strawberries and rice have shown that inoculation led to increased plant growth and total phenolic content [18,19]. In particular, Pseudomonas sp. has revealed positive effects on plant growth and stress tolerance, resulting in alterations in the plant metabolite profile, both in greenhouse and in field conditions [41,42,43,44,45,46].
In both environmental contexts, inoculation shifted the plant metabolism towards fatty acid production, influencing total fatty acid content in field-grown plants, particularly impacting unsaturated fatty acids (UFAs) in microcosm conditions. UFAs play a crucial role as a carbon and energy source [47,48], while also protecting the photosynthetic machinery of plants that grow in saline conditions, thereby increasing stress resistance through membrane modification. Additionally, UFAs serve as precursors and intermediates in the biosynthesis of various bioactive compounds [49]. In inoculated microcosm-grown plants, UFAs, particularly oleic acid, were significantly elevated. However, in field-grown plants, total UFAs content did not differ from non-inoculated plants, although an enrichment in linoleic acid was noted. Despite this, the overall unsaturated fatty acid content and the ratio between unsaturated and saturated fatty acids were reduced in field-inoculated plants, suggesting potential lipid peroxidation due to oxidative stress [50,51]. In the microcosm experiment, where stress was absent, UFAs likely played a more significant role as growth enhancers, whereas in the more stressed field conditions, their contribution to stress resistance enhancement might have been more pronounced.
The absence of β-sitosterol and the presence of more saturated sterols, such as stigmasterol and cholesterol, suggest increased β-oxidation under stressful field conditions [51]. This observation reinforces the challenging conditions experienced by plants in the field.
Field-inoculated plants grown were also enriched in phenolic compounds. The accumulation of molecules such as hydroxybenzoic acid, quercetin and apigenin, alongside a metabolic shift towards the production of also formononetin, caffeic acid and caffeoylquinic acid, not only suggests that field plants were exposed to stressful conditions, but also supports the hypothesis that inoculation induced a beneficial metabolic shift, providing the plants with a broader spectrum and increased amounts of protective compounds, consistent with the activation and diversification of stress protection mechanisms [15,52,53]. Quercetin, a powerful antioxidant, enhances plant photosynthesis, alters soil chemistry, and attracts beneficial microbial populations, ultimately promoting plant growth [54] as it can modulate auxin transport following inoculation, resulting in the accumulation of this phytohormone in certain plant organs, impacting plant growth and development processes [55,56]. The increase in apigenin levels in inoculated field plants further reinforces their antioxidant and anti-inflammatory potential, showing promise as a chemo preventive agent [57]. Soil/sediment characteristics at the growing field - high values of electrical conductivity of both sediment (129 dS.m-1) and pore water (113 dS.m-1), SAR (13.3) and ESP (17) [58] - sustain the scenario of harsher conditions in the field setting. The registered high salinity coupled with high sodicity creates a stressful environment even for halophytes, and plants must undoubtably strengthen their antioxidant and osmoprotective molecular machinery to survive [59]. The increase observed in lignoceric acid in field-inoculated plants, a precursor of cell waxes and suberin which act as a barrier of defence against environmental factors such as dehydration and UV-induced stress [60], indicates inoculated plants are more effectively prepared to handle these stressors.
The metabolite profile of field plants additionally indicates a higher vulnerability to biotic stress, a conclusion further supported by our study’s exploration of phenolic acids. These compounds may elucidate the observed lignified appearance in the lower portion of shoots of field-grown plants compared to those grown in pots (Supplementary Figure S2). Phenolic acids, integral to processes such as plant-microbe symbiosis, allelopathy, and lignin cross-linking, may contribute to fortifying cell walls against pathogen invasion [61]. Previous findings from Ferreira et al. (2023b) regarding the presence of azelaic acid and the enrichment of their microbiome with pathogenesis-related orthologs in crop plants from the same location provide additional context for understanding the susceptibility of field plants to biotic stress.
Inoculated microcosm-grown plants, despite the absence of visible stress symptoms, also accumulated some specific antioxidant compounds like caffeic acid, quinic acid, caffeoylquinic acids, and quercetin. This indicates that a similar, although more subtill, metabolic shift may have occurred in microcosm conditions, equipping the plants to rapid responses to oxidative and or saline stress. Under controlled conditions, inoculated plants exhibited higher concentrations of some organic acids, like acetic acid, which can be utilized as an alternative carbon source by plant tissues [63]. Acetic and citric acids contribute to increased chlorophyll content and improved stress tolerance, ultimately promoting plant growth [64]. Moreover, inoculation with PGPB can affect the plant’s carboxylic acid profile and recruit beneficial bacteria for the root-associated microbiome [65]. Our analysis supports the hypothesis that in both microcosm and field plants, a metabolic shift occurred promoting the synthesis of these TCA cycle products, that underlies growth promotion.
3.2. Biostimulation of Salicornia europaea with PGPB: Implications on Economic Value
Halophytes like Salicornia europaea, have been gaining recognition as valuable sources of bioactive compounds such as polyphenols and flavonoids [66,67]. These compounds not only bolster plant resilience but also offer potential health benefits for humans. Stress-induced accumulation of phenolic compounds enhances the nutritional and nutraceutical value of these plants [68,69]. Inoculation resulted in an increase in rare sugars, like psicose and allose, known for their anti-obesity activity (Chen et al. 2019, 2018). Other compounds that were enriched in inoculated plants, like caffeoylquinic acids, are beneficial for human sugar metabolism [70]. Moreover, inoculated plants contain more essential fatty acids like linoleic acid (ω-6) and alpha-linolenic acid (ω-3), which have various health benefits, including the regulation of cholesterol levels, and have antibacterial and anti-inflammatory effects [71,72,73,74,75,76].
The phenolic content and sterol content of inoculated field-grown plants increased significantly compared to non-inoculated controls. Sterols and phenolic compounds, such as caffeoylquinic acids and the flavonoids formononetin, quercetin, and apigenin, enhance the anti-inflammatory and anticancer potential of field-inoculated plants [70,77,78]. Notably, the presence of specific metabolites like ergost-25-ene-3,5,6,12-tetrol and phenolic acids such as caffeoylquinic acids, caffeic acid, and formononetin may enhance antimicrobial and neuroprotective properties of this plant [70,77,79].
4. Materials and Methods
4.1. Reagents
For GC-MS analysis, palmitic acid, oleic acid, linoleic acid, malic acid, succinic acid, β-sitosterol, cholesterol, eicosanol, glycerol, galactose, hexatriacontane, N,O-bis(trimethylsilyl)trifluoroacetamide (BSTFA) and trimethylsilyl chloride (TMSCl) were purchased from Sigma-Aldrich® (Steinheim, Germany). Pyridine was purchased from Panreac Quimica SLU (Barcelona, Spain).
For UHPLC-MS analysis, ethanol and methanol were purchased from Sigma-Aldrich® (Steinheim, Germany). Quercetin, (+)-catechin, isorhamnetin, luteolin, chlorogenic acid, kaempferol, and cinnamic acid were purchased from Extrasynthese (Lyon, France).
All reagents were p.a. grade.
4.2. Plant Inoculation and Growth
The methodologies used for seed bacterization, germination, and cultivation were previously described by [23]. In brief, Salicornia europaea seeds collected in the production fields owned by a private company (Horta dos Peixinhos, Aveiro, Portugal) were co-inoculated with Brevibacterium casei EB3 and Pseudomonas oryzihabitans RL18 (EB3+RL18), which display several plant growth-promoting traits (Supplementary Table 1). These bacterial strains were isolated from the roots (rhizosphere and endosphere) of Salicornia europaea specimens from the Portuguese coast [58].
Individual strains were grown in Tryptic Soy Broth (TSB; Liofilchem, Roseto degli Abruzzi) supplemented with 25 g L-1 NaCl for 48h in a rotary shaker (150rpm, 30ºC±2ºC). After centrifugation, cells were collected and washed twice in sterile saline solution (NaCl 9gL-1). A third resuspension in sterile saline was adjusted to 108 CFU mL−1 (OD600∼1). The inoculum was prepared using equal volume of each bacterial suspension (1:1v/v). Seeds were surface disinfected (2 minutes immersion in 1mL solution of 1:1 proportion of hydrogen peroxide (30%) and ethanol (96%), followed by three times rinse with sterile distilled water), submersed in the inoculum solution for 2h in a rotary shaker (150rpm, 30ºC±2ºC), and then pelleted by low-speed centrifugation (5000g; 2 min). Non inoculated seeds (NI, control) were immersed in sterile saline solution under the same conditions.
Simultaneous trials were conducted using both inoculated and control seeds, encompassing a microcosm (under controlled conditions) and a field (under natural conditions) experiment. For the microcosm experiments, seeds were dried in a laminar flow chamber and germinated in 1% agar plates in a Sanyo MLR 350 H Versatile Environmental Test Chamber (Moriguchi, Osaka, Japan) programmed for a 16/8 h light/dark regime for 15 days at 24ºC. Germinated plantlets were transplanted to plastic pots (5.5 cm highX5.5 cm diameter) containing similar weight of a 1:1 mixture of three-time sterilized saltmarsh sediment (from the same field plot used for the field experiment) and perlite. Five pots from each condition (inoculated and control plants) were placed in shallow trays containing 20% Hoagland’s solution modified by addition of 10gL-1 marine salt. For two months, plants were maintained in a removable outdoor plastic greenhouse, under natural temperature and sunlight and, after that time, all pots were placed in a growth chamber (Fitoclima D1200, Aralab, Sintra, Portugal), under controlled conditions (16/8h photoperiod, 20-25ºC temperature, 40% relative humidity and 500μmol m-2 s-1 photon flux density, for 35 days, until the end of the experiment (Supplementary Figure S1A,B).
In the field experiment, seeds were sowed immediately after bacterization in the crop area (40º39′2’’N/8º38′42’’W), a former saltpan which has been repurposed for Salicornia europaea crop cultivation since 2015 (Supplementary Figure S1C,D). Seeds were placed inside plastic rims (15 cm diameter) in rows of 10, separated by 50 cm. Sediment salinity was approximately 129 dS m-1 [58].
Reinoculation of microcosm (pot) and field plants was performed by the application of 20 mL of either the bacterial solution mixture (inoculated plants) or sterile saline solution (control plants), every 15 days until the end of the experiment (5 months of growth).
4.3. Sample Preparation for Phytochemical Analyses
After five months of growth, three specimens (aboveground biomass) from each inoculation condition were collected for further analysis (Supplementary Figure S2). Plant material was dried at 60 ºC until constant weight and ground to a fine powder, using liquid nitrogen and a household electric grinder (Krups).
4.4. Silylation and Gas Chromatography-Mass Spectometry (GC-MS) Analysis
Direct silylation was performed to increase the sensitivity of the GC-MS analysis. In a screw-cap glass tube, 10 mg of dried powder of each sample and replicate were mixed with 125 μL of pyridine, 125 μL of BSTFA, 25 μL of TMSCl and 90 μL of the internal standard solution (hexatriacontane 1 mg mL-1). Dichloromethane was added up to a final volume of 1 mL. The mixture was incubated in a water bath at 70ºC for 40 min with continuous magnetic stirring. After cooling, the mixture was filtered through a 0.45 μm nylon filter (Membrane Solutions) and injected in a GC/MS QP2010 Ultra Shimadzu with an A ZB- 5 ms J & W (30 m×0.25 mm×0.25 μm) capillary column. The injector temperature was set at 320 °C, and the transfer line temperature was set at 200ºC. Samples were injected with a split ratio of 1:10, and helium was used as the carrier gas with a flux of 1.19 mL min-1. The temperature of the column was maintained at 70ºC for 5 min and then increased 4by °C per minute until 250ºC, followed by an increase of 2ºC per minute until 300ºC, which was maintained for 5 min, totalling 80 min. The mass spectrometer was operated in electronic impact (EI) mode with an energy of 70 eV and data were collected at a rate of 1 scan s-1 over a range of m/z 50-1000. Identification of the components was carried out using direct comparison with the library entries of the lant spectra database (NIST14 Mass spectral and WILEY RegistryTM of Mass Spectra Data). All reagents used were of high-grade quality, and the equipment was previously calibrated.
The internal standard method was applied, and the amount of each metabolite present in each replicate (3 biological replicates, each analyzed twice) was obtained from calibration curves acquired by injection of known concentration solutions of each standard or with its TMS derivatives. The standard concentrations were chosen to guarantee the quantification of each compound in the samples by interpolating the calibration curve.
4.5. Extraction and Ultra-High-Performance Liquid Chromatography-Mass Spectometry (UHPLC-MS) Analysis
Ethanol extracts were obtained by mixing 10 mg of dry-weight powder (3 replicates) with ethanol p.a. (2 mL, two cycles of 24h each) at room temperature. After the extracting solution was filtered twice with hydrophilic cotton, the solvent was evaporated using a HyperVAC VC2200 at 40ºC until the complete dryness of the mixture was obtained. Each dried ethanol fraction was weighed, and methanol p.a. was added to dilute the samples (final concentration 10 mg/mL). The solutions were filtered through a 0.2 μm nylon membrane (Whatman). Unlike the GC analysis, it was not necessary to add a standard solution since the components are in a liquid state; gas dissipation does not occur in this analysis. Each biological replicate was then injected into the UHPLC- MS apparatus.
UHPLC-MS analysis was performed using a Thermo Scientific Ultimate 3000RSLC (Dionex, Sunnyvale, CA, USA) equipped with a Dionex UltiMate 3000 RS diode array detector and coupled to a mass spectrometer. The column used was a thermos scientific Hypersil gold column (100 mm × 2.1 mm) with a particle size of 1.9 mm, and its temperature was maintained at 30ºC. The mobile phase comprised 0.1% formic acid (v/v), degassed and filtered before use. The flow rate was 0.2 mL/min. The solvent gradient started with 5% solvent over 14 min, followed by 40% solvent for 2 min, 100% over 7 min, and 5% over 10 min. The injection volume was 2 μL. UV–vis spectral data were gathered in a range of 190-700 nm, and the chromatographic profiles were documented at 320 nm. The mass spectrometer was an LTQ XL linear ion trap 2D equipped with an orthogonal electrospray ion source (ESI). The equipment was operated in negative-ion mode with an electrospray ionization source of 5.00 kV and an ESI capillarity temperature of 275ºC. The full scan covered a mass range of 50–2000 m/z. Collision-induced dissociation MS/MS and MS2 experiments were simultaneously acquired for precursor ions. Each sample was analyzed separately but under the same chromatography conditions.
The amount of each metabolite was calculated with calibration curves acquired by injection of known concentration solutions of each standard diluted in methanol p.a. The concentration of the standards was chosen to guarantee the quantification of each compound in the samples by interpolation.
4.6. Statistical Analyses
Normality was assessed by the Shapiro-Wilks test. Differences between experimental conditions were evaluated using a t-test whenever normal distribution was verified. Non normal distributions were evaluated by the Mann-Whitney test. Significant differences were always considered at p < 0.05.
All the above-mentioned statistical analyses were performed using IBM SPSS statistics, version 28.0.1.1 (14) and Infostat version 2020e [80]. Principal component analysis was performed with Past4 .11 [81].
Log2 fold change of two groups’ means was calculated using Microsoft® Excel for Mac (version 16.83).
5. Conclusions
Inoculation of Salicornia europaea seeds with PGPB Brevibacterium casei EB3 and Pseudomonas oryzihabitans RL18 induced lasting effects in the vegetative phase of the plant’s life cycle and in the plants’ metabolome.
In microcosm experiments, the accumulation of sugars, unsaturated fatty acids, citric and acetic acids, sterols, phenolic acids, and flavonoids due to inoculation likely allowed plants to produce biomass and overcome potential environmental stresses.
The phytochemical profile of field-grown plants indicates that they experienced higher levels of stress. This is evidenced by the production of stigmasterol and cholesterol, as well as the decrease in the UFA/SFA ratio. However, inoculation in field-grown plants enhanced their ability to withstand adverse environmental conditions, by the accumulation of metabolites such as sucrose, unsaturated fatty acids, malic acid, phenolic acids, and flavonoids.
The accumulation of important bioactive compounds induced by PGPB-inoculation also enhanced the nutritional and nutraceutical value of the plant.
The results indicate that plant responses are influenced by several factors, including environmental conditions and that inoculation differently redirected the plant metabolism affecting the production of compounds that most benefited biomass production and the reinforcement of the plant’s defence system.
Customized approaches to biofertilization may, therefore, be necessary under different scenarios. A combination of plant metabolomics and a comprehensive analysis of the plant microbiome may elucidate the mechanisms underlying stress attenuation and growth promotion.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: title; Table S1: title; Video S1: title.
Author Contributions
Conceptualization, M.J.F., I.N.S-G., D.C..G.A.P and A.C.; Methodology, M.J.F., A.V.; Validation and formal analysis, M.J.F., C.G., A.V., D.C.G.A.P. and A.C.; Investigation, M.J.F. and A.V.; Resources, D.C.G.A.P and A.C.; Data curation, M.J.F., A.V and A.C.; Writing— original draft preparation, M.J., D.C.G.A.P, and A.C.; writing—review and editing, M.J.F., I.N.S.- G, A.V., C.G, D.C.G.A.P., J.C, H.S. and A.C.; visualization, M.J.F.; supervision, J.C., A.C. and H.S.; project administration, A.C. and H.S.; funding acquisition, A.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by projects Rhizomis PTDC/BIA-MIC/29736/2017 and Halius PTDC/BIA-MIC/3157/2020 (http://doi.org/10.54499/PTDC/BIA-MIC/3157/2020) financed by Fundação para a Ciência e a Tecnologia (FCT) through the Regional Operational Program of the Center (02/SAICT/2017) with FEDER funds (European Regional Development Fund, FNR and OE) and by FCT through CESAM (UIDP/50017/2020 + UIDB/50017/2020), LAQV-REQUIMTE (UIDB/50006/2020 + UIDP/50006/2020)). We also acknowledge FCT/FSE for the financial support to Maria João Ferreira through a PhD grant (PD/BD/150363/2019).
Acknowledgments
We are grateful to Horta dos Peixinhos for their help and support during seed collection and provision of space for the field experiment. We also thank Glória Pinto for the use of the climate chambers during the final months of the experiment, and Enrique Mateos-Naranjo and Jennifer Mesa-Marín of the Departamento de Biología Vegetal y Ecología of the University of Sevilla, for their ad- vice on the cultivation of Salicornia plants in laboratory and greenhouse conditions.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Olanrewaju, O.S.; Glick, B.R.; Babalola, O.O. Mechanisms of action of plant growth promoting bacteria. World J. Microbiol. Biotechnol. 2017, 33, 197. [Google Scholar] [CrossRef]
- Alexander, A.; Singh, V.K.; Mishra, A. Halotolerant PGPR Stenotrophomonas maltophilia BJ01 Induces Salt Tolerance by Modulating Physiology and Biochemical Activities of Arachis hypogaea. Front. Microbiol. 2020, 11, 568289. [Google Scholar] [CrossRef]
- Fadiji, A.E.; Santoyo, G.; Yadav, A.N.; Babalola, O.O. Efforts towards overcoming drought stress in crops: Revisiting the mechanisms employed by plant growth-promoting bacteria. Front. Microbiol. 2022, 13, 962427. [Google Scholar] [CrossRef]
- Khumairah, F.H.; Setiawati, M.R.; Fitriatin, B.N.; Simarmata, T.; Alfaraj, S.; Ansari, M.J.; El Enshasy, H.A.; Sayyed, R.Z.; Najafi, S. Halotolerant Plant Growth-Promoting Rhizobacteria Isolated From Saline Soil Improve Nitrogen Fixation and Alleviate Salt Stress in Rice Plants. Front. Microbiol. 2022, 13, 905210. [Google Scholar] [CrossRef]
- Glick, B.R. Plant Growth-Promoting Bacteria: Mechanisms and Applications. Scientifica 2012, 2012, 963401. [Google Scholar] [CrossRef]
- Poria, V.; Dębiec-Andrzejewska, K.; Fiodor, A.; Lyzohub, M.; Ajijah, N.; Singh, S.; Pranaw, K. Plant Growth-Promoting Bacteria (PGPB) integrated phytotechnology: A sustainable approach for remediation of marginal lands. Front. Plant Sci. 2022, 13, 999866. [Google Scholar] [CrossRef]
- Mokrani, S.; Nabti, E.-H.; Cruz, C. Current Advances in Plant Growth Promoting Bacteria Alleviating Salt Stress for Sustainable Agriculture. Appl. Sci. 2020, 10, 7025. [Google Scholar] [CrossRef]
- Rajkumar, M.; Freitas, H. Effects of inoculation of plant-growth promoting bacteria on Ni uptake by Indian mustard. Bioresour. Technol. 2007, 99, 3491–3498. [Google Scholar] [CrossRef]
- Alves, N.S.F.; Inoue, S.G.K.; Carneiro, A.R.; Albino, U.B.; Setzer, W.N.; Maia, J.G.; Andrade, E.H.; da Silva, J.K.R. Variation in Peperomia pellucida growth and secondary metabolism after rhizobacteria inoculation. PLOS ONE 2022, 17, e0262794. [Google Scholar] [CrossRef]
- Wang, J.; Zhao, H.; Chen, T.; Lin, W.; Lin, S. Effect of Burkholderia ambifaria LK-P4 inoculation on the plant growth characteristics, metabolism, and pharmacological activity of Anoectochilus roxburghii. Front. Plant Sci. 2022, 13, 1043042. [Google Scholar] [CrossRef]
- Maeda, H.A. Evolutionary Diversification of Primary Metabolism and Its Contribution to Plant Chemical Diversity. Front. Plant Sci. 2019, 10, 881. [Google Scholar] [CrossRef]
- Butnariu, M.; Bocso, N.-S. The biological role of primary and secondary plants metabolites. Nutr. Food Process. 2022, 5, 01–07. [Google Scholar] [CrossRef]
- Zaynab, M.; Fatima, M.; Sharif, Y.; Zafar, M.H.; Ali, H.; Khan, K.A. Role of primary metabolites in plant defense against pathogens. Microb. Pathog. 2019, 137, 103728. [Google Scholar] [CrossRef]
- DMisra; Dutta, W.; Jha, G.; Ray, P. Interactions and Regulatory Functions of Phenolics in Soil-Plant-Climate Nexus. Agronomy, vol. 13, no. 2. 2023.
- Laoué, J.; Fernandez, C.; Ormeño, E. Plant Flavonoids in Mediterranean Species: A Focus on Flavonols as Protective Metabolites under Climate Stress. Plants 2022, 11, 172. [Google Scholar] [CrossRef]
- Zuluaga, M.Y.A.; Milani, K.M.L.; Miras-Moreno, M.B.; Lucini, L.; Valentinuzzi, F.; Mimmo, T.; Pii, Y.; Cesco, S.; Rodrigues, E.P.; de Oliveira, A.L.M. The adaptive metabolomic profile and functional activity of tomato rhizosphere are revealed upon PGPB inoculation under saline stress. Environ. Exp. Bot. 2021, 189, 104552. [Google Scholar] [CrossRef]
- Vaishnav, A.; Kumari, S.; Jain, S.; Varma, A.; Tuteja, N.; Choudhary, D.K. PGPR-mediated expression of salt tolerance gene in soybean through volatiles under sodium nitroprusside. J. Basic Microbiol. 2016, 56, 1274–1288. [Google Scholar] [CrossRef]
- Rahman, M. Plant probiotic bacteria Bacillus and Paraburkholderia improve growth, yield and content of antioxidants in strawberry fruit. Sci. Rep. 2018; 8. [Google Scholar]
- Chamam, A.; Sanguin, H.; Bellvert, F.; Meiffren, G.; Comte, G.; Wisniewski-Dyé, F.; Bertrand, C.; Prigent-Combaret, C. Plant secondary metabolite profiling evidences strain-dependent effect in the Azospirillum–Oryza sativa association. Phytochemistry 2013, 87, 65–77. [Google Scholar] [CrossRef]
- Pang, Z.; Chen, J.; Wang, T.; Gao, C.; Li, Z.; Guo, L.; Xu, J.; Cheng, Y. Linking Plant Secondary Metabolites and Plant Microbiomes: A Review. Front. Plant Sci. 2021, 12. [Google Scholar] [CrossRef]
- Gupta, S.; Schillaci, M.; Roessner, U. Metabolomics as an emerging tool to study plant–microbe interactions. Emerg. Top. Life Sci. 2022, 6, 175–183. [Google Scholar] [CrossRef]
- Sierra-García, I.N.; Ferreira, M.J.; Torres-Ballesteros, A.; Louvado, A.; Gomes, N.; Cunha, A. Brevibacterium EB3 inoculation enhances rhizobacterial community interactions leading to improved growth of Salicornia europaea. Appl. Soil Ecol. 2024, 196. [Google Scholar] [CrossRef]
- Ferreira, M.J. Biostimulation of Salicornia europaea L. crops with plant-growth-promoting bacteria in laboratory and field conditions: effects on growth and metabolite profile. J. Appl. Microbiol. 2023; lxad036. [Google Scholar]
- Jerónimo, D.; Lillebø, A.I.; Cremades, J.; Cartaxana, P.; Calado, R. Recovering wasted nutrients from shrimp farming through the combined culture of polychaetes and halophytes. Sci. Rep. 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Kim, S.; Lee, E.-Y.; Hillman, P.F.; Ko, J.; Yang, I.; Nam, S.-J. Chemical Structure and Biological Activities of Secondary Metabolites from Salicornia europaea L. Molecules 2021, 26, 2252. [Google Scholar] [CrossRef]
- Hulkko, L.S.S.; Rocha, R.M.; Trentin, R.; Fredsgaard, M.; Chaturvedi, T.; Custódio, L.; Thomsen, M.H. Bioactive Extracts from Salicornia ramosissima J. Woods Biorefinery as a Source of Ingredients for High-Value Industries. Plants 2023, 12, 1251. [Google Scholar] [CrossRef]
- Giordano, R.; Aliotta, G.E.; Johannesen, A.S.; Voetmann-Jensen, D.; Laustsen, F.H.; Andersen, L.A.; Rezai, A.; Fredsgaard, M.; Vecchio, S.L.; Arendt-Nielsen, L.; et al. Effects of Salicornia-Based Skin Cream Application on Healthy Humans’ Experimental Model of Pain and Itching. Pharmaceuticals 2022, 15, 150. [Google Scholar] [CrossRef]
- Giordano, R.; Saii, Z.; Fredsgaard, M.; Hulkko, L.S.S.; Poulsen, T.B.G.; Thomsen, M.E.; Henneberg, N.; Zucolotto, S.M.; Arendt-Nielsen, L.; Papenbrock, J.; et al. Pharmacological Insights into Halophyte Bioactive Extract Action on Anti-Inflammatory, Pain Relief and Antibiotics-Type Mechanisms. Molecules 2021, 26, 3140. [Google Scholar] [CrossRef]
- Morkunas, I.; Ratajczak, L. The role of sugar signaling in plant defense responses against fungal pathogens. Acta Physiol. Plant. 2014, 36, 1607–1619. [Google Scholar] [CrossRef]
- Gómez-Ariza, J.; Campo, S.; Rufat, M.; Estopà, M.; Messeguer, J.; Segundo, B.S.; Coca, M.; Lenz, R.; Louie, K.; Sondreli, K.; et al. Sucrose-Mediated Priming of Plant Defense Responses and Broad-Spectrum Disease Resistance by Overexpression of the Maize Pathogenesis-Related PRms Protein in Rice Plants. Mol. Plant-Microbe Interactions® 2007, 20, 832–842. [Google Scholar] [CrossRef]
- Lacrampe, N.; Lopez-Lauri, F.; Lugan, R.; Colombié, S.; Olivares, J.; Nicot, P.C.; Lecompte, F. Regulation of sugar metabolism genes in the nitrogen-dependent susceptibility of tomato stems toBotrytis cinerea. Ann. Bot. 2020, 127, 143–154. [Google Scholar] [CrossRef]
- Stein, O.; Granot, D. An Overview of Sucrose Synthases in Plants. Front. Plant Sci. 2019, 10, 95. [Google Scholar] [CrossRef]
- Jeandet, P.; Formela-Luboińska, M.; Labudda, M.; Morkunas, I. The Role of Sugars in Plant Responses to Stress and Their Regulatory Function during Development. Int. J. Mol. Sci. 2022, 23, 5161. [Google Scholar] [CrossRef]
- Ohto, M.-A.; Onai, K.; Furukawa, Y.; Aoki, E.; Araki, T.; Nakamura, K. Effects of Sugar on Vegetative Development and Floral Transition in Arabidopsis. Plant Physiol. 2001, 127, 252–261. [Google Scholar] [CrossRef]
- Gouffi, K.; Pica, N.; Pichereau, V.; Blanco, C. Disaccharides as a New Class of Nonaccumulated Osmoprotectants for Sinorhizobium meliloti. Appl. Environ. Microbiol. 1999, 65, 1491–1500. [Google Scholar] [CrossRef]
- O’hara, L.E.; Paul, M.J.; Wingler, A. How Do Sugars Regulate Plant Growth and Development? New Insight into the Role of Trehalose-6-Phosphate. Mol. Plant 2013, 6, 261–274. [Google Scholar] [CrossRef]
- Zhu, G.; Geuns, J.M.C.; Dussert, S.; Swennen, R.; Panis, B. Change in sugar, sterol and fatty acid composition in banana meristems caused by sucrose-induced acclimation and its effects on cryopreservation. Physiol. Plant. 2006, 128, 80–94. [Google Scholar] [CrossRef]
- Morkunas, I.; Bednarski, W.; Kozłowska, M. Response of embryo axes of germinating seeds of yellow lupine to Fusarium oxysporum. Plant Physiol. Biochem. 2004, 42, 493–499. [Google Scholar] [CrossRef]
- Cho, Y.H.; Yoo, S.D. Signaling role of fructose mediated by FINS1/FBP in Arabidopsis thaliana. PLoS Genet., vol. 7, no. 1, 2011.
- Siddiqui, H.; Sami, F.; Hayat, S. Glucose: Sweet or bitter effects in plants-a review on current and future perspective. Carbohydr. Res. 2020, 487, 107884. [Google Scholar] [CrossRef]
- Walker, V.; Couillerot, O.; Von Felten, A.; Bellvert, F.; Jansa, J.; Maurhofer, M.; Bally, R.; Moënne-Loccoz, Y.; Comte, G. Variation of secondary metabolite levels in maize seedling roots induced by inoculation with Azospirillum, Pseudomonas and Glomus consortium under field conditions. Plant Soil 2011, 356, 151–163. [Google Scholar] [CrossRef]
- Walker, V.; Bertrand, C.; Bellvert, F.; Moënne-Loccoz, Y.; Bally, R.; Comte, G. Host plant secondary metabolite profiling shows a complex, strain-dependent response of maize to plant growth-promoting rhizobacteria of the genus Azospirillum. New Phytol., vol. 189, no. 2, pp. 494–506, 2011.
- Han, Q.-Q.; Lã¼, X.-P.; Bai, J.-P.; Qiao, Y.; Parã©, P.W.; Wang, S.-M.; Zhang, J.-L.; Wu, Y.-N.; Pang, X.-P.; Xu, W.-B.; et al. Beneficial soil bacterium Bacillus subtilis (GB03) augments salt tolerance of white clover. Front. Plant Sci. 2014, 5, 525–525. [Google Scholar] [CrossRef]
- Chen, L.; Liu, Y.; Wu, G.; Njeri, K.V.; Shen, Q.; Zhang, N.; Zhang, R. Induced maize salt tolerance by rhizosphere inoculation of Bacillus amyloliquefaciens SQR9. Physiol. Plant. 2016, 158, 34–44. [Google Scholar] [CrossRef]
- Kang, S.-M.; Shahzad, R.; Bilal, S.; Khan, A.L.; Park, Y.-G.; Lee, K.-E.; Asaf, S.; Khan, M.A.; Lee, I.-J. Indole-3-acetic-acid and ACC deaminase producing Leclercia adecarboxylata MO1 improves Solanum lycopersicum L. growth and salinity stress tolerance by endogenous secondary metabolites regulation. BMC Microbiol. 2019, 19, 80. [Google Scholar] [CrossRef]
- Sunita, K.; Mishra, I.; Mishra, J.; Prakash, J.; Arora, N.K. Secondary Metabolites From Halotolerant Plant Growth Promoting Rhizobacteria for Ameliorating Salinity Stress in Plants. Front. Microbiol. 2020, 11, 567768. [Google Scholar] [CrossRef]
- Aluko, O.O.; Li, C.; Wang, Q.; Liu, H. Sucrose Utilization for Improved Crop Yields: A Review Article. Int. J. Mol. Sci. 2021, 22, 4704. [Google Scholar] [CrossRef]
- de Faria, R.T.; Rodrigues, F.N.; Oliveira, L.D.V.; Müller, C. In vitro Dendrobium nobile plant growth and rooting in different sucrose concentrations. Hortic. Bras. 2004, 22, 780–783. [Google Scholar] [CrossRef]
- He, M.; Ding, N.-Z. Plant Unsaturated Fatty Acids: Multiple Roles in Stress Response. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Magni, N.N.; Veríssimo, A.C.S.; Silva, H.; Pinto, D.C.G.A. Metabolomic Profile of Salicornia perennis Plant’s Organs under Diverse In Situ Stress: The Ria de Aveiro Salt Marshes Case. Metabolites 2023, 13, 280. [Google Scholar] [CrossRef]
- Mathesius, U. Flavonoid Functions in Plants and Their Interactions with Other Organisms. Plants 2018, 7, 30. [Google Scholar] [CrossRef]
- Ameixa, O.M.C.C.; Rebelo, J.; Silva, H.; Pinto, D.C.G.A. Gall midge Baldratia salicorniae Kieffer (Diptera: Cecidomyiidae) infestation on Salicornia europaea L. induces the production of specialized metabolites with biotechnological potential. Phytochemistry 2022, 200. [Google Scholar] [CrossRef]
- Singh, P.; Arif, Y.; Bajguz, A.; Hayat, S. The role of quercetin in plants. Plant Physiol. Biochem. 2021, 166, 10–19. [Google Scholar] [CrossRef]
- Zuluaga, M.Y.A.; Milani, K.M.L.; Miras-Moreno, B.; Lucini, L.; Valentinuzzi, F.; Mimmo, T.; Pii, Y.; Cesco, S.; Rodrigues, E.P.; de Oliveira, A.L.M. Inoculation with plant growth-promoting bacteria alters the rhizosphere functioning of tomato plants. Appl. Soil Ecol. 2020, 158, 103784. [Google Scholar] [CrossRef]
- Peer, W.A.; Murphy, A.S. Flavonoids and auxin transport: modulators or regulators? Trends Plant Sci. 2007, 12, 556–563. [Google Scholar] [CrossRef]
- Shankar, E.; Goel, A.; Gupta, K.; Gupta, S. Plant Flavone Apigenin: an Emerging Anticancer Agent. Curr. Pharmacol. Rep. 2017, 3, 423–446. [Google Scholar] [CrossRef]
- Ferreira, M.J.; Cunha, A.; Figueiredo, S.; Faustino, P.; Patinha, C.; Silva, H.; Sierra-Garcia, I.N. The Root Microbiome of Salicornia ramosissima as a Seedbank for Plant-Growth Promoting Halotolerant Bacteria. Appl. Sci. 2021, 11, 2233. [Google Scholar] [CrossRef]
- Balasubramaniam, T.; Shen, G.; Esmaeili, N.; Zhang, H. Plants’ Response Mechanisms to Salinity Stress. Plants 2023, 12, 2253. [Google Scholar] [CrossRef]
- Li, Y.; Beisson, F.; Ohlrogge, J.; Pollard, M. Monoacylglycerols Are Components of Root Waxes and Can Be Produced in the Aerial Cuticle by Ectopic Expression of a Suberin-Associated Acyltransferase. Plant Physiol. 2007, 144, 1267–1277. [Google Scholar] [CrossRef]
- Deng, Y.; Lu, S. Biosynthesis and Regulation of Phenylpropanoids in Plants. Crit. Rev. Plant Sci. 2017, 36, 257–290. [Google Scholar] [CrossRef]
- Ferreira, M.J.; Sierra-Garcia, I.N.; Louvado, A.; Gomes, N.C.M.; Figueiredo, S.; Patinha, C.; A Pinto, D.C.G.; Cremades, J.; Silva, H.; Cunha. Domestication shapes the endophytic microbiome and metabolome of <italic>Salicornia europaea</italic>. J. Appl. Microbiol. 2023, 134. [CrossRef]
- Hann, E.C.; Overa, S.; Harland-Dunaway, M.; Narvaez, A.F.; Le, D.N.; Orozco-Cárdenas, M.L.; Jiao, F.; Jinkerson, R.E. A hybrid inorganic–biological artificial photosynthesis system for energy-efficient food production. Nat. Food 2022, 3, 461–471. [Google Scholar] [CrossRef]
- Chen, H.-C.; Zhang, S.-L.; Wu, K.-J.; Li, R.; He, X.-R.; He, D.-N.; Huang, C.; Wei, H. The effects of exogenous organic acids on the growth, photosynthesis and cellular ultrastructure of Salix variegata Franch. Under Cd stress. Ecotoxicol. Environ. Saf. 2019, 187, 109790. [Google Scholar] [CrossRef]
- Rekha, K.; Baskar, B.; Srinath, S.; Usha, B. Plant-growth-promoting rhizobacteria Bacillus subtilis RR4 isolated from rice rhizosphere induces malic acid biosynthesis in rice roots. Can. J. Microbiol. 2018, 64, 20–27. [Google Scholar] [CrossRef]
- Ferreira, M.J.; Pinto, D.C.G.A.; Cunha, .; Silva, H. Halophytes as Medicinal Plants against Human Infectious Diseases. Appl. Sci. 2022, 12, 7493. [CrossRef]
- Patel, S. Salicornia: evaluating the halophytic extremophile as a food and a pharmaceutical candidate. 3 Biotech 2016, 6, 1–10. [Google Scholar] [CrossRef]
- Sun, W.; Shahrajabian, M.H. Therapeutic Potential of Phenolic Compounds in Medicinal Plants—Natural Health Products for Human Health. Molecules 2023, 28, 1845. [Google Scholar] [CrossRef]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef]
- Liu, W. Current Advances in Naturally Occurring Caffeoylquinic Acids: Structure, Bioactivity, and Synthesis. J. Agric. Food Chem. 2020, 68, 10489–10516. [Google Scholar] [CrossRef]
- Hur, S.J.; Park, G.B.; Joo, S.T. Biological activities of conjugated linoleic acid (CLA) and effects of CLA on animal products. Livest. Sci. 2007, 110, 221–229. [Google Scholar] [CrossRef]
- Natali, F.; Siculella, L.; Salvati, S.; Gnoni, G.V. Oleic acid is a potent inhibitor of fatty acid and cholesterol synthesis in C6 glioma cells. J. Lipid Res. 2007, 48, 1966–1975. [Google Scholar] [CrossRef]
- Dilika, F.; Bremner, P.; Meyer, J. Antibacterial activity of linoleic and oleic acids isolated from Helichrysum pedunculatum: a plant used during circumcision rites. Fitoterapia 2000, 71, 450–452. [Google Scholar] [CrossRef]
- Kusumah, D.; Wakui, M.; Murakami, M.; Xie, X.; Yukihito, K.; Maeda, I. Linoleic acid, α-linolenic acid, and monolinolenins as antibacterial substances in the heat-processed soybean fermented with Rhizopus oligosporus. Biosci. Biotechnol. Biochem. 2020, 84, 1285–1290. [Google Scholar] [CrossRef]
- Zheng, C.J.; Yoo, J.-S.; Lee, T.-G.; Cho, H.-Y.; Kim, Y.-H.; Kim, W.-G. Fatty acid synthesis is a target for antibacterial activity of unsaturated fatty acids. FEBS Lett. 2005, 579, 5157–5162. [Google Scholar] [CrossRef]
- Kolar, M.J.; Konduri, S.; Chang, T.; Wang, H.; McNerlin, C.; Ohlsson, L.; Härröd, M.; Siegel, D.; Saghatelian, A. Linoleic acid esters of hydroxy linoleic acids are anti-inflammatory lipids found in plants and mammals. J. Biol. Chem. 2019, 294, 10698–10707. [Google Scholar] [CrossRef]
- Youssef, A.M.M.; Maaty, D.A.M.; Al-Saraireh, Y.M. Phytochemical Analysis and Profiling of Antitumor Compounds of Leaves and Stems of Calystegia silvatica (Kit.) Griseb. Molecules 2023, 28, 630. [Google Scholar] [CrossRef]
- Novotny, L.; Mahmoud, F.; Abdel-Hamid, M.; Hunakova, L. Anticancer potential of β-sitosterol. Int. J. Clin. Pharmacol. Pharmacother. 2017, 2. [Google Scholar] [CrossRef]
- Tian, J.; Wang, X.-Q.; Tian, Z. Focusing on Formononetin: Recent Perspectives for its Neuroprotective Potentials. Front. Pharmacol. 2022, 13, 905898. [Google Scholar] [CrossRef]
- D. R. J.A., C. F., B. M.G., G. L., M. Tablada, and C. W. Robledo, “Infostat.” 2008.
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. Past: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4. [Google Scholar]
Figure 1.
Phytochemical profile of non-inoculated and inoculated Salicornia europaea in controlled conditions (microcosm experiments), according to chemical families of compounds detected by GC-MS and UHPLC-MS. NI- non-inoculated plants; EB3+RL18 – plants inoculated with Brevibacterium casei EB3 and Pseudomonas oryzihabitans RL18. The columns represent the average of 3 replicates, and the error bars correspond to the standard error. Data obtained were compared by t-test (sugar acids, saturated fatty acids, alcohols, amides and phenolic acids) or Mann-Whitney test (unsaturated fatty acids, sterols, sugars, and flavonoids); significant differences are indicated by **(p=0.004) and ***(p=0.002) between non-inoculated control and test.
Figure 1.
Phytochemical profile of non-inoculated and inoculated Salicornia europaea in controlled conditions (microcosm experiments), according to chemical families of compounds detected by GC-MS and UHPLC-MS. NI- non-inoculated plants; EB3+RL18 – plants inoculated with Brevibacterium casei EB3 and Pseudomonas oryzihabitans RL18. The columns represent the average of 3 replicates, and the error bars correspond to the standard error. Data obtained were compared by t-test (sugar acids, saturated fatty acids, alcohols, amides and phenolic acids) or Mann-Whitney test (unsaturated fatty acids, sterols, sugars, and flavonoids); significant differences are indicated by **(p=0.004) and ***(p=0.002) between non-inoculated control and test.
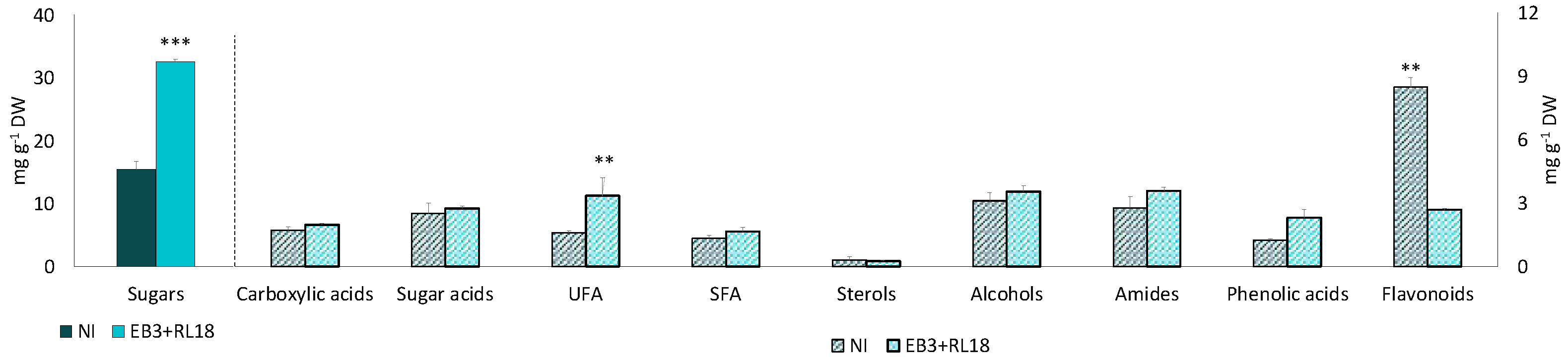
Figure 2.
Response of Salicoria europaea secondary metabolism to inoculation with PGPB Brevibacterium casei EB3 and Pseudomonas oryzihabitans RL18, under controlled microcosm conditions. Fold changes are expressed as Log2 [(EB3+RL18)/NI]. A-Carboxylic acids; B-Sugar acids; C-Sterols; D-Fatty acids; E-Alcohols; F-Amides; G-Sugars; H-Phenolic compounds.
Figure 2.
Response of Salicoria europaea secondary metabolism to inoculation with PGPB Brevibacterium casei EB3 and Pseudomonas oryzihabitans RL18, under controlled microcosm conditions. Fold changes are expressed as Log2 [(EB3+RL18)/NI]. A-Carboxylic acids; B-Sugar acids; C-Sterols; D-Fatty acids; E-Alcohols; F-Amides; G-Sugars; H-Phenolic compounds.
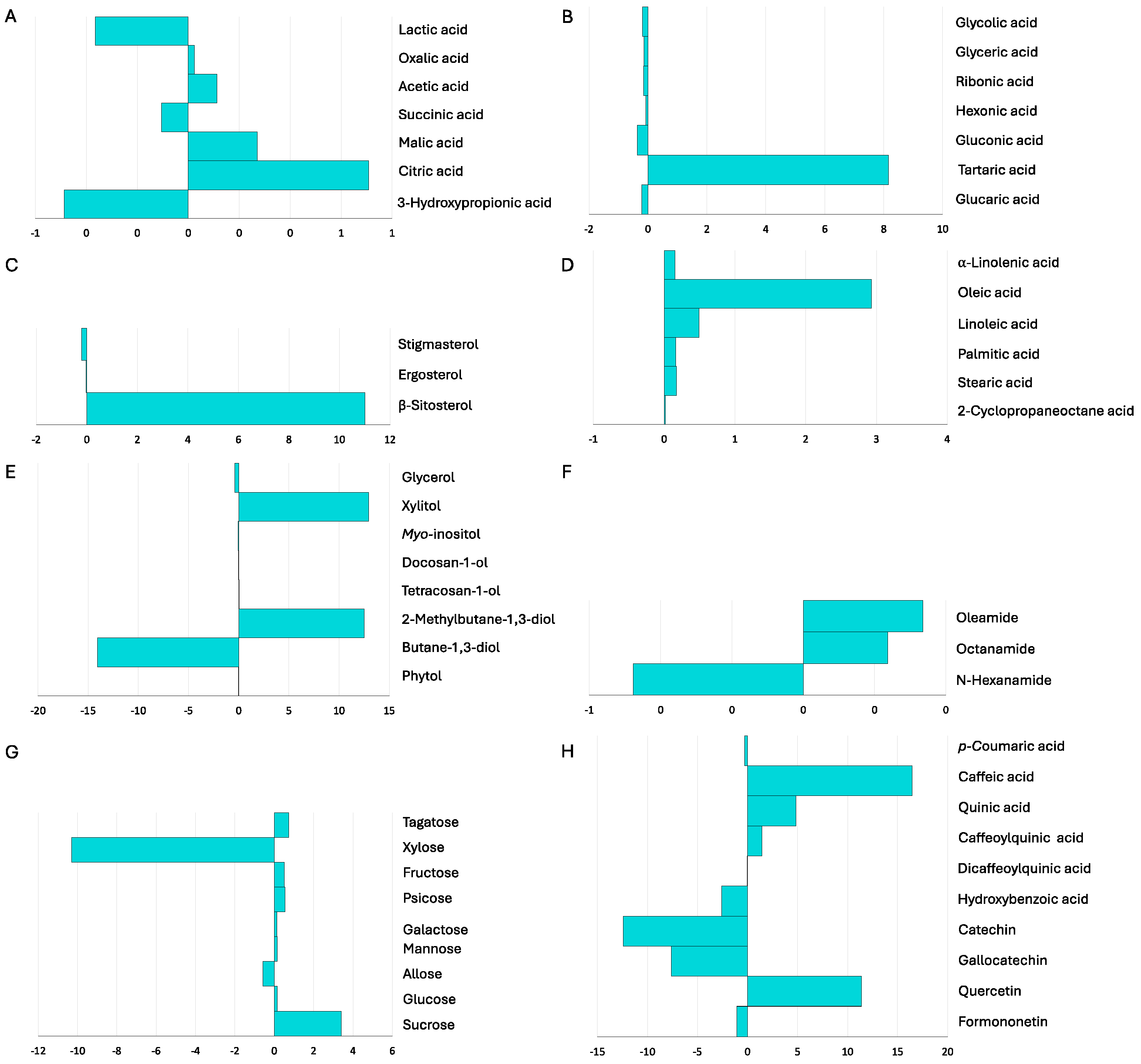
Figure 3.
Phytochemical profile of non-inoculated and inoculated Salicornia europaea in field conditions (microcosm experiments), according to chemical families of compounds detected by GC-MS and UHPLC-MS. NI- non-inoculated plants; EB3+RL18 – plants inoculated with Brevibacterium casei EB3 and Pseudomonas oryzihabitans RL18. The columns represent the average of 3 replicates, and the error bars correspond to the standard error. Data obtained were compared by t-test (carboxylic acids, sugar acids, and amides) and the Mann-Whitney U test (saturated and unsaturated fatty acids, sterols, alcohols, and sugars); * indicates significant differences (p<0.05) between control and test.
Figure 3.
Phytochemical profile of non-inoculated and inoculated Salicornia europaea in field conditions (microcosm experiments), according to chemical families of compounds detected by GC-MS and UHPLC-MS. NI- non-inoculated plants; EB3+RL18 – plants inoculated with Brevibacterium casei EB3 and Pseudomonas oryzihabitans RL18. The columns represent the average of 3 replicates, and the error bars correspond to the standard error. Data obtained were compared by t-test (carboxylic acids, sugar acids, and amides) and the Mann-Whitney U test (saturated and unsaturated fatty acids, sterols, alcohols, and sugars); * indicates significant differences (p<0.05) between control and test.
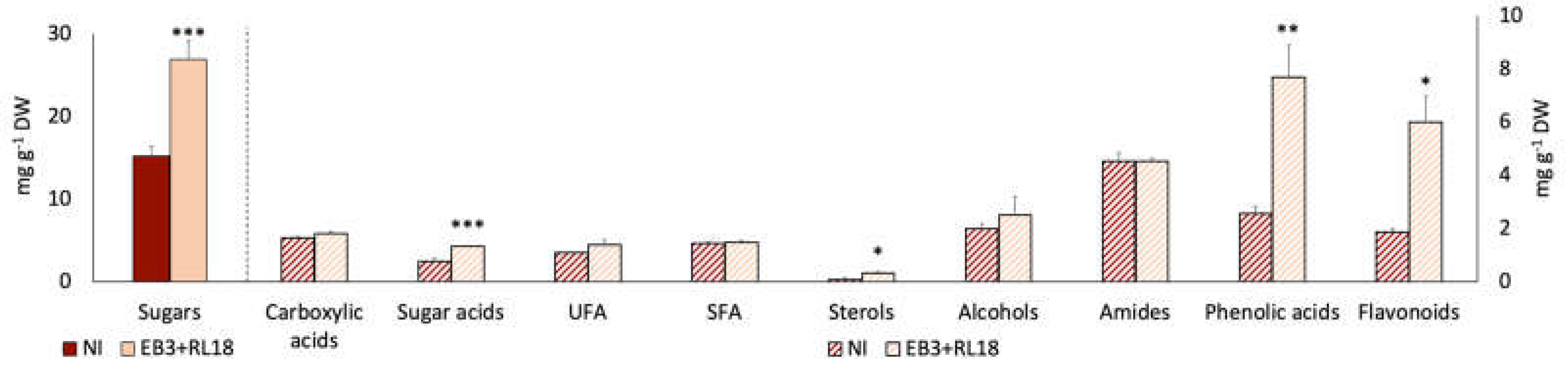
Figure 4.
Response of Salicornia europaea secondary metabolism to inoculation with PGPB Brevibacterium casei EB3 and Pseudomonas oryzihabitans RL18, in field conditions. Fold changes are expressed as Log2 [(EB3+RL18)/NI]. A-Carboxylic acids; B-Sugar acids; C-Sterols; D-Fatty acids; E-Alcohols; F-Amides; G-Sugars; H-Phenolic compounds.
Figure 4.
Response of Salicornia europaea secondary metabolism to inoculation with PGPB Brevibacterium casei EB3 and Pseudomonas oryzihabitans RL18, in field conditions. Fold changes are expressed as Log2 [(EB3+RL18)/NI]. A-Carboxylic acids; B-Sugar acids; C-Sterols; D-Fatty acids; E-Alcohols; F-Amides; G-Sugars; H-Phenolic compounds.
Figure 5.
Principle Component Analysis (PCA) of (A) microcosm experiment and (B) field experiment. Changes in the metabolite composition of Salicornia europaea plants inoculated with Brevibacterium casei EB3 and Pseudomonas oryzihabitans RL18 (EB3+RL18, grey stars) versus the corresponding non-inoculated controls (NI, black asterisks). Each symbol corresponds to an individual analyzed plant.
Figure 5.
Principle Component Analysis (PCA) of (A) microcosm experiment and (B) field experiment. Changes in the metabolite composition of Salicornia europaea plants inoculated with Brevibacterium casei EB3 and Pseudomonas oryzihabitans RL18 (EB3+RL18, grey stars) versus the corresponding non-inoculated controls (NI, black asterisks). Each symbol corresponds to an individual analyzed plant.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



