
Submitted:
12 June 2024
Posted:
13 June 2024
You are already at the latest version
Abstract
Pigeonpea (Cajanus cajan) is one of the important grain legume crops cultivated in the semi-arid tropics, playing a crucial role in the economic well-being of subsistence farmers. India is the major producer of pigeonpea, accounting for over 75 per cent of the world's production. Sterility mosaic disease (SMD), caused by Pigeonpea sterility mosaic virus (PPSMV) and transmitted by the eriophyid mite (Aceria cajani), is a major constraint to pigeonpea cultivation in the Indian subcontinent, leading to potential yield losses of up to 100 per cent. The recent characterization of another Emaravirus associated with SMD has further complicated the etiology of this challenging viral disease. This review focuses on critical areas, including the current status of the disease, transmission and host-range, rapid phenotyping techniques, as well as available disease management strategies. The review concludes with insights into the future prospects, offering an overview and direction for further research and management strategies.
Keywords:
Cajanus cajan
; SMD
; PPSMV
; eriophyid mite
; screening
; disease management
1. The Crop
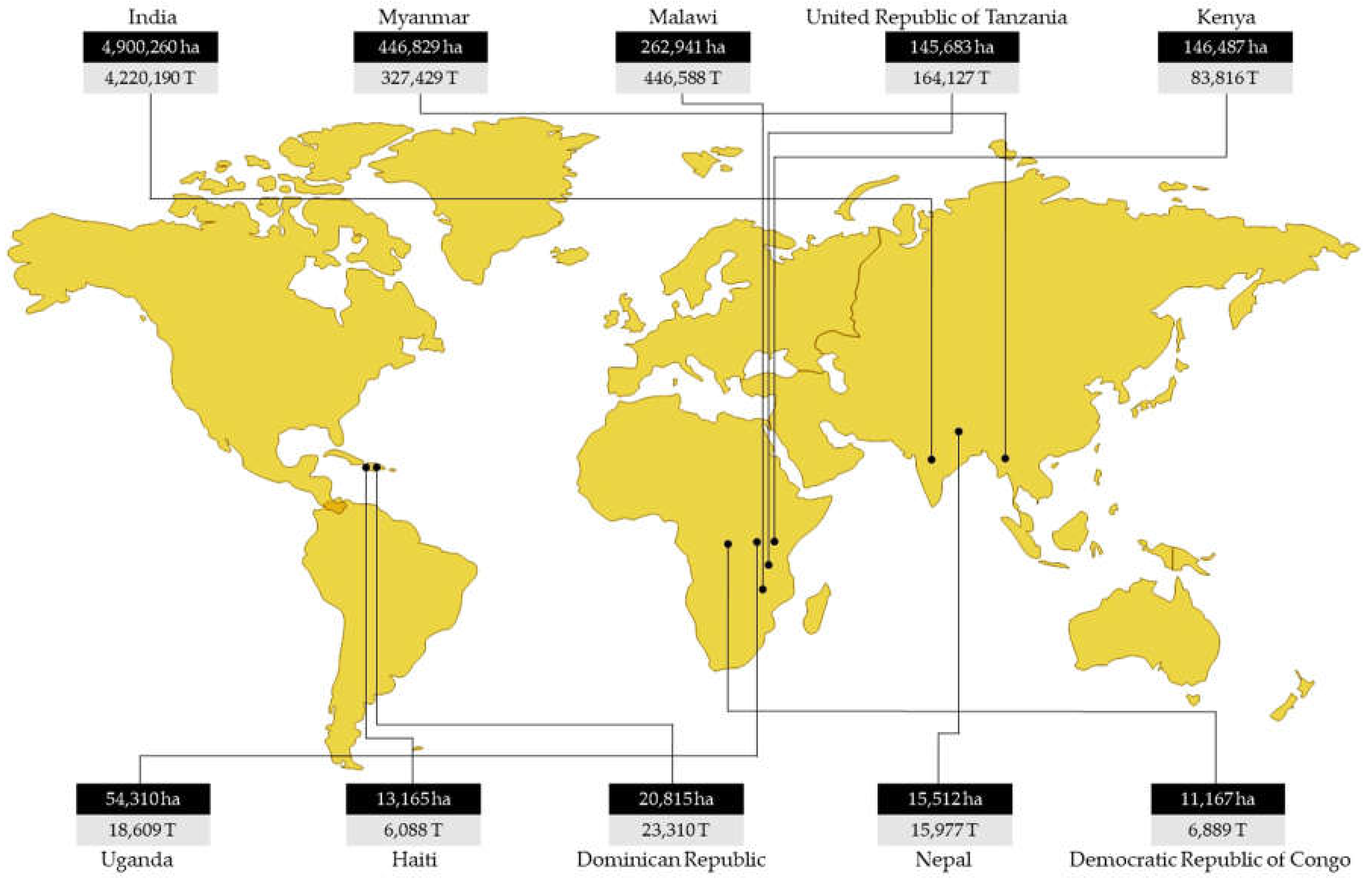
Pigeonpea (Cajanus cajan (L.) Huth) is an important grain legume crop cultivated by small and marginal farmers in semi-arid tropical regions of Asia, Africa and the Americas between 25°N and 35°S [1]. Globally, the pigeonpea crop is harvested from 6.03 million hectares (ha) with a production of about 5.32 million tonnes (MT) (Figure 1) [2]. India is the leading producer, contributing to over 75 per cent of global production [2]. Pigeonpea is primarily grown for its grains, containing 20-30 per cent protein, serving as the principal source of dietary protein for over a billion people [3,4,5,6,7]. Pigeonpea is an important subsistence crop adopted by millions of smallholder farmers and grown as a sole or intercropped with cereals (finger millet, maize, sorghum, pearl millet), legume crops (groundnut, soybean, Indian bean), cotton, chilli etc., under diverse climatic conditions in a rain-fed agricultural system [5,8]. It is a perennial shrub mainly cultivated as an annual, offers cultivars having different durations: extra-early (<130 days), early (131-150 days), mid-early (151-165 days) and medium duration (166-185 days). During the early phase, its slow growth above-ground offers very little competition to the main crop, later its fast-growing nature with deep extensive root system enables the crop to yield in arid conditions with very little available moisture in the soil when no other crop can survive.
The pigeonpea crop has a direct impact on the economic and financial well-being of subsistence farmers in the subcontinent as it is a low-input, rainfed crop that provides economic returns from every part of the plant. Despite a substantial increase in the area and production of pigeonpea over the last two decades, average yields remain low with a meagre 882 kg/ha globally [2] owing to several biotic (Fusarium wilt, sterility mosaic, Phytophthora blight and pod borer complex) and abiotic stresses (drought, salinity and water-logging) encountered at different growth stages. Among the biotic stresses, sterility mosaic disease is the major constraint to pigeonpea production in the Indian subcontinent [5,9,10]. This is a cause of concern as crop yields plateau in the face of rising demand and an increase in the number of mouths to feed.
Figure 1.
Area (hectares (ha)) and production (tonnes (T)) of major pigeonpea producing countries [2].
Figure 1.
Area (hectares (ha)) and production (tonnes (T)) of major pigeonpea producing countries [2].
2. Sterility Mosaic Disease (SMD)
SMD of pigeonpea was initially reported in 1931 by Mitra from Bihar state of India [11] and has since evolved into a serious problem for pigeonpea cultivation in the Indian subcontinent, resulting in an estimated annual loss over US$ 300 million [12]. SMD is endemic to India and has subsequently been reported from Bangladesh, Nepal, Thailand [12,13], Myanmar [14] and Sri Lanka [15] and China but it is not known to occur in Africa or the Americas [12,16]. In India, SMD is reported in various states including, Andhra Pradesh, Bihar, Chhattisgarh, Gujarat, Karnataka, Maharashtra, Punjab, Tamil Nadu, Telangana, Uttar Pradesh, and West Bengal [8,17,18,19]. SMD manifests as yellow mosaic, with bushy pale green plants with excessive vegetative growth, reduced leaf size, stunting, leaf distortion, and partial or complete cessation of reproductive parts [5,8,20,21,22,23,24,25]. Infected plants also show a significant reduction in plant height, the number of branches, flowers per plant and pod length [26].
SMD is referred to as “Green Plague” of pigeonpea, as the infected plants exhibit lush greenery with excessive vegetative growth, without flowers and seed pods. Under favorable conditions, it spreads like a plague in an epidemic form [5,27]. The nature and severity of symptoms depend on the genotype and stage of infection [24]. In susceptible genotypes, early-stage infection (<45-day old plant) results in development of characteristic symptoms and complete cessation of flowering (sterility), while later stage infection (>45-day old plant) results in delayed symptom expression with mild mosaic on few branches or a part of branch, reduced flowering and partial sterility [5,17]. However, in later stage infected plants, new flushes from ratoon crop (severe pruning) exhibit aggressive and clear severe mosaic symptoms [5,24,25,28]. Based on severity, the disease symptoms are categorized into three groups; (i) severe mosaic and sterility, (ii) mild mosaic and partial sterility, and (iii) chlorotic ring spots with no sterility (Figure 2). The virus strain also significantly influences symptom expression. For instance, genotype ICP2376 infected with SMD showed chlorotic ring spots with the Patancheru strain, whereas it exhibited severe mosaic with sterility symptoms in Bengaluru and Coimbatore [12,25,29,30,31]. SMD incidence varies due to pathogen variability. In India, among ten distinct strains of PPSMV identified, strains from Bengaluru, Dholi, Vamban and Varanasi appear to be more virulent than those at Badnapur, Hyderabad, Pantnagar, Kanpur, Ludhiana and Faizabad [32,33,34,35]. Additionally, SMD-infected plants aggravate powdery mildew (Oidiopsis taurica) infection [36] and spider mite (Schizotetranychus cajani) infestation [37] compounding the damage.
2.1. The Vector-Aceria cajani
SMD of pigeonpea is transmitted by an eriophyid mite, Aceria cajani Channabasavanna (Arthropoda: Acari: Arthropoda) [38] (Figure 3) but not through sap, pollen, seed or soil [22,39,40,41,42,43]. Aceria cajani is microscopic and measures about 200-250 µm in length with a short life cycle of about two weeks that includs an egg and two nymphal stages [44,45]. These mites are predominantly found on young symptomatic leaves of SMD-infected plants, residing on the undersurface of leaves and concentrating towards the petiole [28,46,47,48,49] this suggesting a beneficial relationship between the mite vector and the virus [28]. The mites possess a very short stylet (~2.03 µm) generally feed on epidermal and underlying mesophyll cells, acquiring virus only from infected cells [50]. Their direct feeding does not cause any obvious damage to pigeonpea [45]. Aceria cajani is highly host-specific, primarily confined to pigeonpea and few of its wild relatives [35,45]. In nature, these mites can be also seen on Hibiscus panduriformis adjacent to the SMD-infected pigeonpea field [51].
Aceria cajani dispersal is passive, primarily aided by wind currents, with the mites blown up to 2 km from source of inoculums [52]. A Mean temperature of 20-30 °C being favorable for mite proliferation, while higher temperatures and heavy rains are unfavorable [53,54]. Aceria cajani transmits PPSMV in a semi-persistent manner, with a single mite having a transmission efficiency up to 40 per cent; however, more than 5 mites are required for 100 per cent transmission. It requires a minimum acquisition access period (AAP) of 15 minutes and an inoculation access period (IAP) of 90 minutes for the successful transmission of the PPSMV. The mites are virulent for about 6-13 hours after virus acquisition, and the virus is neither propagative nor transovarially transmitted [43,44]. Accurate identification of eriophyid mite based on morphological characters is difficult due to their small size. A diverse range of molecular markers has been used to analyze different strains (biotype) within a species. Analysis of ribosomal DNA internal transcribed spacer (rDNA-ITS) sequence of Aceria cajani collected from different locations in India, Nepal and Myanmar revealed little to no sequence divergence amongst them [48,55]. This probably indicates that there is a single biotype of Aceria cajani in transmitting PPSMV and that did not differ much in their transmission.
2.2. SMD Transmission
Experimentally, PPSMV can be transmitted by leaf stapling method onto pigeonpea, Wild Cajanus species, Phaseolus vulgaris cvs. Top crop, Kintoki and Bountiful, as well as Chrozophora rottleri thus aiding in mite transfer to test genotypes and their feeding transfers [35,43,56]. Mechanical sap inoculation on to Nicotiana benthamiana, N. clevalandi, was successful. However, attempts to sap transmit PPSMV on to pigeonpea was not successful [35,57]. Even purified PPSMV preparation were not infectious [58]. However, PPSMV can be experimentally transmitted on to pigeonpea by grafting [23,59]. Hibiscus panduriformins, Oxalis corniculata, Canavis sativa may act as refuge for mite survival in transit, thereby aiding the spread of SMD [51,53]. Though PPSMV can infect plants outside the genus Cajanus, due to host specificity of its mite vector to Cajanus cajan and few of its wild relatives restricts its potential source of SMD inoculum under field conditions. The perennial, volunteer and ratooned pigeonpea may provide a pool for mite vector and virus, thus serves as primary source of inoculum for SMD [60].
2.3. SMD Etiology and Detection
The causal agent of SMD has remained elusive for several decades. Despite continuous efforts by various laboratories, the identification of etiology has been unsuccessful, posing a significant challenge to the scientific community [27,31,61]. Based on symptoms and mode of transmission, the causal agent of SMD suggested the likely involvement of a virus or virus-like agent [21,61]. Supporting this hypothesis, several studies have demonstrated that there is no involvement of mite toxemia or fungi, bacteria, nematode, Phytoplasma or viroid in causing SMD in pigeonpea [27,32,41,42,61].
Though SMD of pigeonpea was first identified in 1931 [11], the etiology remained a major conundrum until a breakthrough in 1999 by Kumar and his coworkers [29,30,62] leading to the identification of the causal organism provisionally named Pigeonpea sterility mosaic virus (PPSMV), a tenui-like, highly flexuous and branched filaments virus-like particles (VLPs) measuring about 3-10 nm diameter. The purified virus preparation contains a 32 kDa major protein and up to seven segmented RNA species of size 6.8-1.1 kb [58,62,63]. Ultra-structural studies of thin sections of SMD-infected pigeonpea cultivars (ICP8863 and ICP2376) revealed two types of inclusions: (i) quasi-spherical membrane-bound bodies (MBB’s) of 100-150 nm diameter associated with amorphous electron-dense material (EDM) and (ii) fibrous inclusions (FIs) found adjacent to the cell nucleus. In PPSMV-infected Nicotiana benthamiana sections, only MBB’s and EDM were observed with no FIs. These structures bear a resemblance to those infected by eriophyid mite-transmitted High plains virus (HPV) [58,63].
Polyclonal antibodies developed in rabbits against purified PPSMV virus-like particles (VLPs) proved effective in detecting PPSMV in plants through direct antigen coating (DAC)- ELISA and double antibody sandwich (DAS)- ELISA [35,55,58,63,64]. In the ELISA test, samples from hundreds of healthy or uninoculated pigeonpea plants consistently yielded negative results, providing circumstantial evidence that PPSMV is the causal agent of SMD of pigeonpea. Based on the above distinctive characters, PPSMV is presumed to form a separate genus in virus taxonomy [58,63]. Detection of PPSMV was successful in SMD-affected pigeonpea plants infected experimentally by A. cajani, and through grafting, and naturally in the field at several different locations in India. Additionally, PPSMV was detected in infected accessions of wild pigeonpea, Cajanus scarabaeoides. Detection was also accomplished in N. benthamiana and N. clevelandii infected by mechanical sap inoculation [57]. Viruliferous mite extracts were found positive for PPSMV by DAS-ELISA and dot immunobinding assay (DIBA) [43,55] Polyclonal antisera, developed to unique peptide sequences specific to nucleocapsid of PPSMV-1 (PPI5-20 MPSKTPFSNMPAASKK*) and PPSMV-2 (PPII299-314 *STFLPALEADRLASLP) (peptide conjugated to KLH at cysteine residue i.e “*”position) were utilized to detect PPSMV-1 and PPSMV-2 [25]. Next-generation sequencing using Illumina technology of double stranded (ds)-RNAs recovered from pigeonpea leaves infected with SMD revealed that PPSMV comprises a five segmented RNA genome encoding five peptides. These peptides show homology to the polypeptides encoded by corresponding RNA segments of Emaravirus and known members of Bunyaviridae, demonstrating a consistent relationship with all Emaravirus and in particular Fig mosaic virus (FMV) and Rose rosette virus (RRV) [65].
The earlier observation of 5-7 RNA segments in the PPSMV genome [58] was inconclusive as it was based on analysis of dsRNA, this was likely compromised by the presence of mixed infection with other PPSMV strain or another virus. During a survey of pigeonpea fields in Hyderabad, some plants showing typical SMD symptoms tested negative for PPSMV by RT-PCR [65,66]. Further, testing of SMD- infected samples collected from the Chevella region, near Hyderabad revealed the presence of more than six dsRNA segments. This finding suggested the potential association of yet another mite-transmitted virus with SMD in pigeonpea [25].
Deep sequencing of dsRNAs recovered from pigeonpea leaves affected with mosaic disease revealed the presence of another Emaravirus, Pigeonpea sterility mosaic virus-2 (PPSMV-2) (PPSMV will be referred to as PPSMV-1 hereafter), a six-segmented RNA virus. Phylogenic analysis of nucleotide and amino acid sequence of PPSMV-2 showed the highest identity with FMV and RRV and distinct from PPSMV-1. The separate branching of PPSMV-1 and PPSMV-2 in the phylogenic tree suggests that these two Emaraviruses infecting pigeonpea have followed independent evolutionary paths. Based on the molecular and morphological features, both the viruses were placed under the family Fimoviridae of the genus Emaravirus [25,66,67].
2.4. PPSMV Genome Organization and Genetic Relationship
Oligonucleotide primers, SM-1 (5'-ACATAGTTCAATCCTTGAGTGCG-3') and SM-2 (5'-ATATTTTAATACACTGATAGGA-3') derived from the nucleotide sequence of RNA-5 segment, specifically amplified a 321 bp product from purified PPSMV RNA preparation and total RNA extracts of PPSMV-infected pigeonpea leaves and N. benthamiana plants by reverse transcription-polymerase chain reaction (RT-PCR) [63,68]. However, subsequent investigations revealed that the above primers (SM-1 and SM-2) were amplifying a partial sequence of RNA-3 segment of PPSMV [25,67].
The novel Emaravirus, PPSMV associated with SMD of pigeonpea composed of five large RNA fragments, RNA-1 (7022 nt) encodes for RNA-dependent RNA polymerase (RdRP, p1), exhibiting 37 to 54 per cent amino acid sequence similarity with RdRp of other Emaraviruses. RNA-2 (2223 nt), encodes for a glycoprotein precursor (GP, p2), and shares 31 to 45 per cent identity at the amino acid level with GPs of Fig mosaic virus (FMV), Rose rosette virus (RRV), Raspberry leaf blotch virus (RLBV) and European mountain ash ringspot-associated virus (EMARaV). RNA-3 (1442 nt), encodes for nucleocapsid protein (NC, p3), showing 25 to 44 per cent identity at amino acid level with NC proteins of RRV, FMV, RRV, EMARaV, RLBV, and Maize red stripe virus (MRSV). RNA-4 (1563 nt, p4), encodes the movement protein (MP, p4), sharing 41 per cent amino acid similarity with MP encoded by RRV. RNA-5 (1801 nt), encodes a polypeptide (p5) of unknown function, which shares 33 per cent of sequence identity with RNA-5 encoded protein of FMV (Elbeaino et al., 2014). A phylogenetic tree, constructed with the complete amino acid sequence of PPSMV RNA particles and the homologous proteins encoded by corresponding RNA segments of the family Bunyaviridae, showed a consistent relationship of PPSMV with all Emaravirus, particularly with FMV and RRV [25,65,67].
Subsequently, another eriophyid mite transmitted Emaravirus has been characterized from pigeonpea leaves associated with SMD and provisionally named as PPSMV-2 which consists of six RNA fragments. Like PPSMV-1, corresponding RNA particle codes for similar polypeptides [66]. The two polypeptides (p5 and p6) with unknown functions could be involved in the virus life cycle through different roles including the transmission by eriophyid mites [69]. The phylogenetic trees developed showed Emaraviruses clustered in two discrete clades: one containing WMoV and RLBV, and the second comprising all other Emaraviruses. PPSMV-1 and PPSMV-2 originate from a common ancestor branched into different clades, distinct from each other, and diversifying as two distinct viruses infecting a common host. Closely related PPSMV-2 and FMV are present in the same clade, reflecting an evolutionary relationship, while PPSMV-1 and RRV formed as closely associated taxa in a separate clade with a common ancestor from other Emaraviruses [8,25,66].
There was speculation regarding the number of RNA segments in the PPSMV-1. Hits recognized in deep sequencing analysis, corresponding to RNA-5 was matched to PPSMV-2. This was confirmed by RT-PCR analysis using RNA-5 specific primers, suggesting that PPSMV-1 may not possess RNA-5 as a genomic segment. In further investigations, some pigeonpea plants of cv. ICP 8863, when inoculated with PPSMV-P sub-isolate Chevella, showed the presence of PPSMV-1 alone without PPSMV-2. These plants did not amplify RNA-5 when specific primers, confirming the absence of this segment. Phylogenetic analysis revealed that RNA-5 of PPSMVs formed a single clade with a common ancestor, suggesting that earlier reports of PPSMV-1 containing five genomic RNAs may not be convincing [25]. The Variability in Emaravirus associated with SMD in India indicated the independent existence of PPSMV-1 and PPSMV-2 alongside instances of mixed infections. Phylogenetic analysis of nucleotide and amino acid sequences of RNA-3 and RNA-4 of PPSMV isolates showed significant sequence variability [8,67,70].
3. Epidemiology of SMD
The epidemiology of SMD is complex, involving the virus, mite vector, pigeonpea cultivar, and the unpredictable environmental conditions of the semi-arid tropics. In India, SMD occurs every year in almost all pigeonpea growing regions, but the disease incidence varies widely across different regions and seasons [27]. Conflicting reports exist regarding the influence of climatic conditions on SMD epidemiology [47,53]. Crops grown under irrigated conditions or in proximity to irrigated fields are particularly vulnerable to early SMD infection [71]. The severity of the disease depends on the genotype and stage of infection [24]. In susceptible genotypes, early-stage infection (<45-day old plant) results in sterility, whereas later stage infection (>45-day old plant) results in mild mosaic on few branches or a part of branch, reduced flowering, and partial sterility [5,17].
SMD is not seed or soil-borne; it is solely introduced by its mite vector. Diseased plants left in the field after harvest, on the field bank, in kitchen garden or wild relatives of pigeonpea such as C. platycarpus and C. scarabaeoides serves as reservoir for the mite vector and the virus during the offseason, thus acting as an inoculum for SMD [5,35,60]. In rainfed pigeonpea cultivation, the primary source of inoculum is the stubbles left in the field after harvesting, volunteer plants, or plants near water sources or in the shade as these plants maintain foliage and harbor vector mite as well as virus. After the early rains, these plants produce new flush thus, creating favorable conditions for multiple cycles of the mite vector before spreading to newly planted crops. The spread of the disease within the field depends on pigeonpea cultivar, plant age, climatic factors, and mite population. In regions where one pigeonpea crop is followed by a wide time gap, and volunteer crop is not common, the reappearance of the SMD is unknown. However, in such regions, it is suspected that wild relatives of pigeonpea such as C. platycarpus and C. scarabaeoides serve as the primary source of inoculum, and viruliferous mites assisted by wind currents aid in the spread of disease [5,6]. Weed species such as Hibiscus panduriformins, Oxalis corniculata and Canavis sativa may act as refuges for mite survival in transit and may therefore contribute to the spread of SMD [51,53].
A weather-based forecasting study of SMD was conducted for SK Nagar (Gujarat), Gulbarga/Kalburgi (Karnataka), Rahuri (Maharashtra) and Vamban (Tamil Nadu). The Hybrid models, specifically Autoregressive Integrated Moving Average (ARIMA)- Support Vector Regression (SVR) and ARIMA-Artificial Neural Network (ANN), outperformed individual models in predicting disease incidence at SK Nagar, Gulbarga/Kalburgi, and Vamban. However, at Rahuri, individual models demonstrated better compared to the hybrid model with ARIMA. The use of the ARIMA-SVR hybrid model was found to be particularly applicable under conditions where the seasonal mean severity of SMD exceeds one per cent. Conversely, the SVR model can effectively predict disease incidence when it is less than one per cent [72].
4. Screening Techniques for SMD Resistance
The PPSMV is not mechanically sap transmissible onto pigeonpea [6,35,57]. Screening for SMD in pigeonpea genotypes is commonly performed using two methods: (i) leaf stapling [73,74] and (ii) infector hedge/ row [75,76] facilitating PPSMV transmission through the mite vector. Vector mite transmission of PPSMV occurs if the genotype is susceptible to both the virus and its vector. Failure of virus transmission suggests that the tested genotype could possess resistance to the vector, the virus, or to both. Genotypes resistant to mite inoculation are further assessed for virus resistance using the “grafting technique” [23,59]. PPSMV exhibits four different host responses in pigeonpea germplasm (a) severe mosaic and sterility (ICP8863), (b) mild mosaic and partial sterility (ICP8862), (c) chlorotic ringspot (ICP2376 to Patancheru isolate), and (d) no visible symptoms (ICP7035) [6]. Based on SMD incidence, test genotypes are categorized into four groups, (i) resistant (R) (<10% SMD incidence), (ii) moderately resistant (MR) (10.1-20% SMD incidence), (iii) susceptible (S) (20.1-40% SMD incidence), and highly susceptible (HS) (>40.1% SMD incidence) [10]. The response of host differentials against SMD of pigeonpea was developed and presented in Table 1 [77].
4.1. Leaf Stapling Method
Leaf stapling technique developed by Nene and Reddy [73,74], allows for the rapid screening of pigeonpea genotypes under both field and glasshouse conditions. Young SMD-infected leaflets from susceptible genotype (eg: ICP8863) collected in a moist cloth bag were observed for mite infestation to ensure a minimum of 10 mites per leaf. The mite-infested leaflets were then stapled onto test plants at the two- to three-leaf stage in such a way that the undersurface of the diseased leaflet comes in contact with both surfaces of the test plant by folding, anchoring mites for transfer. The feeding of these mites results in PPSMV transmission onto the test plant (Figure 4). Test genotypes are subsequently observed for initial disease symptom development, and the observations were taken 60 days after planting.
4.2. Infector Hedge/Row Method
This field screening method is designated for large-scale pigeonpea genotypes screening through natural infestation. 4-5 rows of a susceptible genotype (eg.: ICP8863) are planted across the wind direction and upwind one side of the field, one-month prior the actual trial begins (Figure 5a). SMD is maintained in these rows using the leaf stapling method. Subsequently, test genotypes and a susceptible genotype (used as an indicator/infector row to monitor disease spread) are planted in the field at a 10:1 ratio (Figure 5b). The viruliferous mites from infector hedge are carried by the wind, facilitate disease transmission. Successful screening is determined when infector rows between test plants reach 100 per cent infection. Disease incidence is recorded twice: first in the seedling stage (30-40 days after planting) and then at maturity. SMD inoculum can be maintained on pruned foliage of the infector hedge for next crop season [75,76].
4.3. Petiole-Grafting
Genotypes resistant to mite inoculation were evaluated for virus resistance using the petiole-grafting technique [23,59]. Fourteen to sixteen-day-old plants, grown in growth chamber were used for grafting. The process involved using an infected leaflets from the PPSMV-infected ICP8863 genotype as scion. These leaflets were freed from mites through miticide spraying. The terminal end of the test genotype was excised, and a 5-10 mm incision down the center of the stem was made using scalpel blade. The scion was trimmed into wedge shape and inserted into the stem slit of the stock plant, tightly bound with cellophane tape/ grafting clips, and covered with polythene bags to maintain humidity for seven days inside mite-proof growth cabinet. The test genotypes were examined for development of disease symptoms and assayed for PPSMV by DAS-ELISA or RT-PCR [62] at 14, 20, and 35 days after grafting.
5. SMD Management
Various strategies have been explored to mitigate SMD incidence, including the application of pesticides to delay the onset of infection and disease spread, as well as control through cropping methods and host-plant resistance. In field conditions, SMD spreads to pigeonpea exclusively through eriophyid mites, the interference of vector transmission through the application of pesticides such as chinomethionate, dinocap, dicofol, fenazaquin, golecron, sulphur, metasystox, monocrotophos, morestan, profenophos, proporgite or wettable sulfur has proven effective in reducing SMD [5,12,41,60,78,79,80,81,82,83,84,85,86]. Seed treatment with 25 per cent carbofuran or soil application has been shown to protect the crop from SMD infection up to 75 days. Water spray has also demonstrated a 70 per cent reduction in the vector mite infestation [41,52]. Proper detection and monitoring of vector mites are crucial for effective control because usually the presence A. cajani is determined based on SMD symptoms induced by PPSMV.
Though chemical management of SMD is effective, it is an expensive option for subsistence farmers considering pigeonpea cultivation with low-input cost. Studies on different sowing dates, plant density, spacing, intercropping, or border crops have shown no significant effect on SMD incidence [18,53,81,87,88]. Destruction of sources of SMD inoculum prior to the cropping season can reduce SMD incidence and/or delay the early onset of the disease, thereby minimizing its impact. However, such practices are seldom followed due to the farmers’ preoccupation with other revenue-generating activities, lack of resources and labor shortages, in marginal farming conditions where the crop is predominantly cultivated.
Management of SMD through host-plant resistance (HPR) has been prioritized, as it requires no special expertise from growers. HPR is the most reliable and cost-effective approach for the SMD management in pigeonpea. The identification of sources of resistance to SMD begun with the pigeonpea landrace Sabour 2E in India [61,81]. Subsequent efforts for identifying sources of resistance were initiated at International Crops Research Institute for the Semi-Arid Tropics (ICRISAT) in collaboration with National Agricultural Research Systems (NARS). From the global pigeonpea collection at ICRISAT, consisting of 13,015 pigeonpea accessions, 326 resistant and 97 tolerant lines were identified [61]. The selected 326 resistant lines were evaluated at various locations in India, only 10 genotypes exhibited resistance across all locations. The variation in the resistance levels could be attributed to the occurrence of different virus strains. In India, 10 distinct strains of PPSMV have been identified, with strains from Bengaluru, Dholi, Vamban and Varanasi appearing to be more virulent than those at Badnapur, Hyderabad, Pantnagar, Kanpur, Ludhiana and Faizabad [32,33,34,35]. The dynamic nature of the PPSMV has guaranteed the use of strain-specific sources of resistance in crop improvement. Thus, there is a pressing need to identify strain-specific resistance sources for more effective management strategies.
During a five-years period from 1978 to 1983, 88 pigeonpea germplasm lines that had shown resistance at Patancheru, Badnapur and Kanpur were further tested at 10 different locations in India (Badnapur, Bengaluru, Dholi, Faizabab, Kanpur, Ludhiana, Pantnagar, Vamban and Varanasi) through joint ICRISAT-Indian Council of Agricultural Research (ICAR) uniform trails for broad-based resistance to pigeonpea sterility mosaic. Among the test lines, ICP 7867, 10976 and 10977 exhibited resistance across all 10 locations, while ICP 11146 showed resistance or tolerance at nine locations except in Dholi, and ICP 10983 shown resistance or tolerance at eight locations, except at Bengaluru and Varanasi [89]. Novel resistance sources in pigeonpea genotypes and released varieties are presented in Table 2 and Table 3.
Adequate levels of SMD resistance is scarce within the cultivated pigeonpea gene pool. However wild relatives of pigeonpea have been shown to possess high level of resistance to various biotic constraints [107]. The global pigeonpea germplasm collection at ICRISAT comprises over 270 accessions of 47 wild species related to genus Cajanus. Among the 115 accessions representing six wild Cajanus species, C. albicans, C. cajanifolius. C. lineatus, C. platycarpus, C. scarabaeoides, were screened for resistance to PPSMV isolates at Patancheru, Bengaluru and Coimbatore. Fifteen accessions, ICP 15164, 15615, 15626, 15684, 15688, 15700, 15701, 15725, 15734, 15736, 15737, 15740, 15924, 15925 and 15926 were identified as posing broad-based resistance. Some of these accessions also showed resistance to pest damage, cyst nematode and wilt. Except for C. platycarpus, the tested species belong to the secondary gene pool, and are inter-fertile by traditional breeding. Therefore, the resistance found in these accessions is transferable to pigeonpea through conventional breeding program. The resistance in few wild Cajanus species accessions such as, C. cajanifolius (IC272730, 552553, 552557, 552601), C. platycarpus (IC525192, 550320, 550321), C. scarabaeoides (IC308694, 308802, 325999, 382113), C. albicans, C. volubilis, C. sericeus and C. lineatus was reconfirmed [98].
6. Application of Molecular Markers
Sequencing-based bulk segregant analysis (Seq-BSA) was used to map resistant genes for Fusarium wilt (FW) and SMD in pigeonpea. Resistance (R) and susceptible (S) from extreme recombinant inbred lines of ICPL 20096 x ICPL 332 were sequenced. Seq-BSA has provided 7 candidate SNPs for FW and SMD resistance in pigeonpea which are useful for the genomics-assisted breeding in pigeonpea [108]. A diagnostic marker kit was developed with 10 markers for identification of SMD resistant lines [109]. A 50 hypervariable pigeonpea specific simple sequence repeat (AHSSR) markers were screened to identify genomic regions associated with resistance to SMD through bulk segregant analysis (BSA) approach in 84 RILs derived from a cross ICP 8863 (S) x BRG 3 (R). Three SSR markers such as, AHSSR 50150, AHSSR 34130, and AHSSR 20125 were differentiated resistant bulks, susceptible bulks and RIL individuals used for constituting bulks. Single marker analysis (SMA) showed these three markers were closely associated with SMD resistance [110].
Bulk segregant analysis (BSA) was employed to identify the RAPD marker linked to SMD and to develop a SCAR for the SMD resistance gene of ICPL 7035. Of the 200 RAPD primers screened, OPA1880 showed polymorphism in resistant and susceptible lines of a cross ICPL 7035 x ICPL 8863 indicating that OPA1880 was associated with SMD resistance in ICPL 7035. A SCAR marker, SCAR 816 (16f/r) developed from end sequences, was present in all generations (parents, F1 and F2) and is found useful to identify the transferred SMD resistance gene to a line [111]. To identify gene(s) or QTLs linked with SMD resistance, two F2:3 populations such as ICP 8863 x ICPL 20097 (segregating for Patancheru SMD isolate) and TTB 7 x ICP 7035 (segregating for Patancheru and Bengaluru SMD isolates) were phenotyped to respective SMD isolates. More than 3000 SSR markers on parental genotypes of each mapping population, intra-specific genetic maps comprising of 11 linkage groups and 120 and 78 SSR loci were developed for ICP 8863 x ICPL 20097 and TTB 7 x ICP 7035 populations respectively. Composite interval mapping (CIM) based QTL analysis by using genetic mapping and phenotypic data provided four QTLs for Patancheru isolate and two for Bengaluru isolate. This identification of different QTLs for resistance to Patancheru and Bengaluru isolates indicates that involvement of different genes conferring the resistance [112]. From the three mapping populations including two recombinant inbred lines PRIL_B (ICPL 20096 × ICPL 332), PRIL_C (ICPL 20097 × ICP 8863), and one F2 (ICP 8863 × ICPL 87119), a total of 212,464 SNPs were identified in PRIL_B, 89,699 in PRIL_C, and 64,798 in the F2 population through genotyping-by-sequencing approach. Composite interval mapping based QTL analysis identified three major QTLs across the three populations. Particularly, one candidate genomic region identified on CcLG11 seems to be promising QTL for molecular breeding in developing superior lines with enhanced resistance to SMD [113].
7. Future Line of Work
The management of SMD of pigeonpea presents a complex challenge. With the recent discovery of a second Emaravirus associated with SMD, the identification and development of pigeonpea varieties resistant to SMD has become a critical focus for future research. The complex nature of the disease is compounded by the diverse virus strains found across India, prompting the need for the development of strain-specific resistant sources to enhance the adaptability of pigeonpea cultivars to varying disease pressure. Reevaluating mini-core and wild Cajanus collections becomes vital to identifying new sources of resistance, considering the evolving dynamics of virus strains over time. Recognizing the socio-economic significance of pigeonpea, there is a pressing need to channel efforts into developing new high-yielding lines. This initiative aligns with the broader goal of increasing agricultural productivity to support the well-being of subsistence farmers who heavily depend on pigeonpea cultivation. In addition, with the challenges posed by SMD, the simultaneous occurrence of Fusarium wilt (FW) underscores the importance of integrated resistance strategies. Future efforts should prioritize the development of pigeonpea varieties that exhibit robust resistance to both SMD and FW, offering a holistic approach to sustainable disease management. Adopting new breeding approaches such as speed breeding, accelerates and enables quicker development of lines which enhances the efficiency of developing SMD resistant lines, aligning with the urgency of addressing disease challenges in pigeonpea cultivation. Moreover, the application of molecular markers in breeding programs is instrumental in developing resistant lines. Markers such as SNP, SSR, and SCAR facilitate the precise identification and selection of pigeonpea genotypes with enhanced resistance traits, streamlining the breeding process. This targeted approach accelerates the development of SMD-resistant pigeonpea varieties, ensuring the efficient deployment of genetic resources contributing to sustainable crop improvement programs. The Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)-Associated (Cas) system provides precise gene editing capabilities and has successfully been employed in managing several plant diseases [114]. This approach offers a complementary strategy to traditional breeding methods, potentially enabling the rapid development of resistant pigeonpea lines to specific virus strains, thus contributing to sustainable disease management in pigeonpea cultivation.
Climate change directly influences the distribution, severity, and emergence of plant diseases. Future research on climate change and its impact on pigeonpea SMD resistance should focus on understanding the ecology and behavior of SMD vectors. Additionally, efforts should be directed towards integrating climate data with genetic information to develop pigeonpea varieties resistant to SMD. Implementing integrated pest management (IPM) strategies adopted to mitigate SMD impact in varying climatic scenarios is essential. Collaborative interdisciplinary research and knowledge sharing among scientists, breeders, and agricultural extension services will be vital for developing sustainable solutions aimed at combating SMD.
Author Contributions
Conceptualization, B.R.S., S.H.K. and P.A.K.; methodology, B.R.S. and K.R.M.; software, H.S.S.; validation, S.H.K., P.A.K., R.C.N.L. and J.B.C.R.; formal analysis, B.R.S., H.S.S. and D.R.S.E.; investigation, B.R.S., K.S.M., M.M. and D.R.S.E.; resources, S.H.K. and N.R.M.; data curation, S.H.K., N.R.M. and R.C.N.L.; writing original manuscript, S.B.R., K.S.M., D.R.S.E. and H.S.S.; visualization, S.B.R., R.C.N.L. and J.B.C.R.; Supervision, S.H.K. and N.R.M. All authors have read and agreed to the published version of the manuscript.
Acknowledgments
We would like to thank Mr. Vinaynath Reddy of World Vegetable Center- South Central Asia for assistance in refining the pictures.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Van der Maesen, L.J.G. Pigeonpea: origin, history, evolution and taxonomy. In The pigeonpea; Nene, Y.L., Hall, S.D., Sheila, V.K., Eds.; CAB International: Wallingford, UK, 1990; pp. 15–46. [Google Scholar]
- Food and Agriculture Organization (FAO) STAT. 2022. Available online: https://www.fao.org/faostat/en/#data/QCL.
- Faris, D.G.; Saxena, K.B.; Mazumdar, S.; Singh, U. Vegetable pigeonpea: a compromising crop for India. Patancheru, A.P. India, ICRISAT. 1987. [Google Scholar]
- Saxena, K.B.; Kumar, R.V.; Rao, P.V. Pigeonpea nutrition and its improvement. J. Crop Prod. 2002, 5, 227–260. [Google Scholar] [CrossRef]
- Jones, A.T.; Kumar, P.L.; Saxena, K.B.; Kulkarni, N.K.; Muniyappa, V.; Waliyar, F. Sterility mosaic disease- the “Green Plague” of pigeonpea. Advances in understanding the etiology, transmission and control of a major virus disease. Plant Dis. 2004, 88, 436–445. [Google Scholar] [CrossRef]
- Kumar, P.L.; Jones, A.T.; Waliyar, F. Biology, etiology and management of pigeonpea sterility mosaic disease. Annu. Rev. Plant. Pathol. 2004, 3, 77–100. [Google Scholar]
- Varshney, R.K.; Chen, W.; Li, Y.; Bharti, A.K.; Saxena, R.K.; Schlueter, J.A.; Donoghue, M.T.; Azam, S.; Fan, G.; Whaley, A.M.; Farmer, A.D.; Sheridan Iwata, A.; Tuteja, R.; Penmetsa, R.V.; Wu, W.; Upadhyaya, H.D.; Yang, S.P.; Shah, T.; Saxena, K.B.; Michael, T.; McCombie, W.R.; Yang, B.; Zhang, G.; Yang, H.; Wang, J.; Spillane, C.; Cook, D.R.; May, G.D.; Xu, X.; Jackson, S. Draft genome sequence of pigeonpea (Cajanus cajan), an orphan legume crop of resource-poor farmers. Nat. Biotechnol. 2012, 30, 83–89. [Google Scholar] [CrossRef]
- Sayiprathap, B.R.; Patibanda, A.K.; Prasanna Kumari, V.; Jayalalitha, K.; Srinivasa Rao, V.; Sharma, M.; Sudini, H.K. Prevalence of sterility mosaic disease (SMD) and variability in Pigeonpea sterility mosaic virus (PPSMV) in southern India. Indian Phytopathol. 2020, 73, 741–750. [Google Scholar] [CrossRef]
- Sharma, M.; Rathore, A.; Mangala, U.N.; Ghosh, R.; Sharma, S.; Upadhyay, H.D.; Pande, S. New sources of resistance to Fusarium wilt and sterility mosaic disease in a mini-core collection of pigeonpea germplasm. Eur. J. Plant Pathol. 2012, 133, 707–714. [Google Scholar] [CrossRef]
- Sharma, M.; Telangre, R.; Ghosh, R.; Pande, S. Multi-environment field testing to identify broad, stable resistance to sterility mosaic disease of pigeonpea. J. Gen. Plant Pathol. 2015, 81, 249–259. [Google Scholar] [CrossRef]
- Mitra, M. Report of the Imperial Mycologist. Scientific report of the agricultural research institute, Pusa, New Delhi, India. 19: 58-71. Mosaic of pigeonpea. In Plant Diseases of International Importance: Diseases of Cereals and Pulses; Singh, U.S., Mukhopadhyay, A.N., Kumar, J., Chaube, H.S., Eds.; Prentice Hall, New Jersey, 1931. [Google Scholar]
- Reddy, M.V.; Raju, T.N.; Lenne, J.M. Diseases of pigeonpea. In The pathology of food and pasture legumes; Allen, D.J., Lenne, J.M., Eds.; CAB International: Wallingford; UK, 1998; pp. 517–558. [Google Scholar]
- Nene, Y.L.; Sheila, V.K.; Sharma, S.B. A world list of chickpea (Cicer arietinum L.) and pigeonpea (Cajanus Cajan (L.)) pathogens. Legume Pathology Progress Report-7. ICRISAT Publication: Patancheru, Andhra Pradesh, India, 1996. [Google Scholar]
- Su, U. Plant disease in Burma. Int. Bull. Plant Prot. 1931, 5, 141. [Google Scholar]
- Newton, W.; Peiris, T.W.L. Virus disease of plant in Cylon. Plant Prot. Bull. FAO. 1953, 2, 17–21. [Google Scholar]
- Chaohong, Z.; Saxena, K.B.; Li, Z.; Zong, X.; Yang, S. Pigeonpea diseases in China. Int. Chickpea Pigeonpea Newsl. 2001, 8, 39–40. [Google Scholar]
- Kannaiyan, J.; Nene, Y.L.; Reddy, M.V.; Ryan, J.G.; Raju, T.N. Prevalence of pigeonpea diseases and associated crop losses in Asia, Africa and the Americas. Trop. Pest Manag. 1984, 30, 62–71. [Google Scholar] [CrossRef]
- Zote, K.K.; Mali, VR.; Mayee, C.D.; Kulkarni, S.V.; Mote, T.S. Outbreak of sterility mosaic of pigeonpea In Marathwada region of Maharashtra, India. Int. Pigeonpea Newsl. 1991, 14, 19–21. [Google Scholar]
- Singh, J.; Raghuraman, M. Emerging scenario of important mite pest in north India. Zoosymposia. 2011, 6, 170–179. [Google Scholar] [CrossRef]
- Alam, M. Arhar sterility. In Proceedings of the 20th Annual Meeting of the Indian Science Congress, Poona. 1933, 43, 15–16. [Google Scholar]
- Capoor, S.P. Observations on the sterility disease of pigeonpea. Indian J. Agri. Sci. 1952, 22, 271–274. [Google Scholar]
- Nene, Y.L. A survey of viral diseases of pulse crops in Uttar Pradesh. Experimental Station Bulletin; G. B. Pant University Agriculture and Technology: Pantnagar, Uttar Pradesh, 1972; pp. 4–191. [Google Scholar]
- Reddy, A.S.; Kulkarni, N.K.; Kumar, P.L.; Jones, A.T.; Muniyappa, V.; Reddy, D.V.R. A new graft-inoculation method for screening for resistance to Pigeonpea sterility mosaic virus. Int. Chickpea Pigeonpea Newsl. 2002, 9, 44–46. [Google Scholar]
- Patil, B.P.; Kumar, P.L. Pigeonpea sterility mosaic virus: a legume infecting Emaravirus from South Asia. Mol. Plant Pathol. 2015, 16, 775–786. [Google Scholar] [CrossRef]
- Kumar, S.; Subbarao, B.L.; Vipin, H. Molecular characterization of Emaraviruses associated with pigeonpea sterility mosaic disease. Sci. Rep. 2017, 7, 11831. [Google Scholar] [CrossRef]
- Dahal, G.; Neupane, K.R. Incidence and effect of sterility mosaic disease of pigeonpea in Nepal. Int. Pigeonpea Newsl. 1991, 13, 23–24. [Google Scholar]
- Singh, A.K.; Rathi, Y.P.S.; Agrawal, K.C. Sterility mosaic of pigeonpea: A challenge of 20th Century. Indian J. Virol. 1999, 15, 85–92. [Google Scholar]
- Reddy, M.V.; Nene, Y.L. Influence of sterility mosaic resistant pigeonpeas on multiplication of the mite vector. Indian Phytopathol. 1980, 33, 61–63. [Google Scholar]
- Kumar, P.L.; Jones, A.T.; Sreenivasulu, P.; Reddy, D.V.R. Unfolding the mystery of the causal agent of pigeonpea sterility mosaic disease. In Proceedings of the International Conference on Life Sciences in Next Milleniurnm, 11–14 December 1999; University of Hyderbad: India. [Google Scholar]
- Kumar, P.L.; Jones, A.T.; Sreenivasulu, P.; Fenton, B.; Reddy, D.V.R. Characterization of a virus from pigeonpea with affinities to species in the genus Aureusvirus, Family Tombusviridae. Plant dis. 2001, 85, 208–215. [Google Scholar] [CrossRef]
- Kumar, P.L.; Jones, A.T.; Duccan, G.; Roberts, I.M.; Reddy, D.V.R. Characterization of a novel mite transmitted virus associated with pigeonpea sterility mosaic disease. Phytopathol. 2001, 91, 51. [Google Scholar]
- Reddy, M.V.; Beniwal, S.P.S.; Sheila, V.K.; Sithananthan, S.; Nene, Y.L. Role of eriophyid mite (Aceria cajani) in transmission and spread of sterility mosaic of pigeonpea. In Progress in Acarology; Channbasavanna, G.P., Virakthmath, C.A., Eds.; Oxfard and IEH Publishing Co.: New Dellhi, India, 1989. [Google Scholar]
- Reddy, M.V.; Raju, T.N.; Ghanekar, A.M.; Amin, K.S.; Arjunan, G.; Ashtaputhra, J.V.; Sinha, B.K.; Muniyappa, V.; Reddy, S.V.; Guptha, R.P.; Kausalya, G. Variability in sterility mosaic pathogen of pigeonpea in India. Indian Phytopathol. 1993, 46, 206–212. [Google Scholar]
- Kulkarni, N.K.; Reddy, A.S.; Kumar, P.L.; Vijaynarasimha, J.; Rangaswamy, K.T.; Reddy, L.J.; Saxena, K.B.; Jones, A.T.; Reddy, D.V.R. Broad-based resistance to pigeonpea sterility mosaic disease in the accessions of Cajanus scarabaeoides. Indian J. Plant Prot. 2003, 31, 6–11. [Google Scholar]
- Sayiprathap, B.R.; Patibanda, A.K.; Prasanna Kumari, V.; Jayalalitha, K.; Ramappa, H.K.; Rajeswari, E.; Karthiba, L.; Saratbabu, K.; Sharma, M.; Sudini, H.K. Salient findings on host range, resistance screening, and molecular studies on sterility mosaic disease of pigeonpea induced by Pigeonpea sterility mosaic viruses (PPSMV-I and PPSMV-II). Front. Microbiol. 2022, 13, 838047. [Google Scholar] [CrossRef]
- Reddy, M.V.; Kannaiyan, J.; Nene, Y.L. Increased susceptibility of sterility mosaic infected pigeonpea to powdery mildew. Int. J. Trop. Plant. Dis. 1984, 2, 35–40. [Google Scholar]
- Sithanantham, S.; Reddy, M.V.; Rao, R. Increased damage by the spider mite. Schizotetranychus cajani in pigeonpea plants affected by sterility mosaic. In Progress in Acarology; Channabasavanna, G.P., Viraktamath, C.A., Eds.; Oxford and IHB Publishing Co.: India, 1989; Volume 2, pp. 11–14. [Google Scholar]
- Channabasavanna, G.P.A. Contribution of the knowledge of Indian eriophyid mites (Eriophyoidae: Trombidiformes: Acarina). Thesis, University of Agricultural Sciences, Bangalore, India, 1966. [Google Scholar]
- Kandaswamy, T.K.; Ramakrishnan, K. A epiphytotic of pigeonpea sterility mosaic at Coimbatore. Madras Agric. J. 1960, 47, 440–441. [Google Scholar]
- Seth, M.L. Transmission of pigeonpea sterility by an eriophyid mite. Indian Phytopathol. 1962, 15, 225–227. [Google Scholar]
- Ghanekar, A.M.; Sheila, V.K.; Beniwal, S.P.S.; Reddy, M.V.; Nene, Y.L. Sterility mosaic of pigeonpea. In Plant Diseases of International Importance: Diseases of Cereals and Pulses; Singh, U.S., Mukhopadhyay, A.N., Kumar, J., Chaube., H.S., Eds.; Prentice Hall, New Jersey, 1992; Volume 1, pp. 415–428. [Google Scholar]
- Reddy, M.V.; Reddy, D.V.R.; Sacks, W.R. Epidemiology and management of sterility mosaic disease of pigeonpea. Proceedings of International Symposium. Rose Rosette and other Eriophyid Mite-Transmitted Plant Disease Agents of Uncertain Etiology, 19–21 May 1994; Iowa State University: United States. [Google Scholar]
- Kulkarni, N.K.; Kumar, P.L.; Muniyappa, V.; Jones, A.T.; Reddy, D.V.R. Transmission of Pigeonpea sterility mosaic virus by the eriophyid mite, Aceria cajani (Acari: Arthropoda). Plant Dis. 2002, 86, 1297–1302. [Google Scholar] [CrossRef]
- Oldfield, G.N.; Reddy, M.V.; Nene, Y.L.; Reed, W. Preliminary studies of the eriophyid vector of sterility mosaic. Int. Chickpea Pigeonpea Newsl. 1981, 1, 25–27. [Google Scholar]
- Sheila, V.K.; Manohar, S.K.; Nene, Y.L. Biology and morphology of Aceria cajani. Int. Pigeonpea Newsl. 1988, 7, 28. [Google Scholar]
- Muniyappa, V.; Nangia, N. Pigeonpea cultivars and selections for resistance to sterility mosaic in relation to prevalence of eriophyid mite Aceria cajani Channabasavanna. Trop. Grain Legume Bull. 1982, 25, 28–30. [Google Scholar]
- Dhar, V.; Rathore, Y.S. Pattern of population distribution of Aceria cajani on pigeonpea plant. Indian J. Pulses Res. 1995, 7, 137–143. [Google Scholar]
- Kumar, P.L.; Fenton, B.; Duncan, G.; Jones, A.T.; Sreenivasulu, P.; Reddy, D.V.R. Assessment of variation in Aceria cajani (Acari: Eriophyidae) using analysis of nuclear rDNA ITS regions and scanning electron microscopy: Implications for the variability observed in host plant resistance to pigeonpea sterility mosaic disease. Ann. Appl. Biol. 2001, 139, 61–73. [Google Scholar] [CrossRef]
- Pallavi, M.S.; Ramappa, H.K. Population dynomics of pigronpea sterility mosaic virus vector Aceria cajani. Mysore J. Agric. Sci. 2014, 48, 394–399. [Google Scholar]
- Reddy, M.V.; Sheila, V.K.; Murthy, A.K.; Padma, P. Mechanism of resistance to Aceria cajani in pigeonpea. Int. J. Trop. Plant Dis. 1995, 13, 51–57. [Google Scholar]
- Kulkarni, N.K.; Kumar, P.L.; Muniyappa, V.; Jones, A.T.; Reddy, D.V.R. Studies on the natural and experimental host range of Pigeonpea sterility mosaic virus. J. Mycol. Plant Pathol. 2003, 33, 141–145. [Google Scholar]
- Reddy, M.V.; Arjunan, N.; Muniyappa, V. Survival of pigeonpea sterility mosaic pathogen and its vector during summer in southern India. Int. Pigeonpea newsl. 1990, 11, 16–17. [Google Scholar]
- Singh, A.K.; Rathi, Y.P.S. Epidemiology of vector of Pigeonpea sterility mosaic virus. Indian J. Virol. 1997, 13, 143–145. [Google Scholar]
- Kaushik, D.; Srivastava, S.; Nath, B.C.; Chauhan, V.B.; Singh, R. Correlation between mite population (Aceria cajani) and environmental factors causing sterility mosaic disease of pigeon pea. Int. J. Life Sci. 2013, 1, 228–232. [Google Scholar]
- Latha, T.K.S.; Doraiswamy, S. Detection of Pigeonpea sterility mosaic virus, the causal agent of sterility mosaic disease of pigeonpea in viruliferous mite vector by DAS-ELISA and DIBA. Arch. Phytopathol. Plant Prot. 2008, 41, 537–542. [Google Scholar] [CrossRef]
- Kumar, P.L.; Latha, T.K.S.; Kulkarni, N.K.; Raghavendra, N.; Saxena, K.B.; Waliyar, F.; Rangaswamy, K.T.; Muniyappa, V.; Doriswamy, S.; Jones, A.T. Broad-based resistance to pigeonpea sterility mosaic disease in wild relatives of pigeonpea (Cajanus: Phaseoleae). Ann. Appl. Biol. 2005, 146, 371–379. [Google Scholar] [CrossRef]
- Kumar, P.L.; Duncan, G.H.; Roberts, I.M.; Jones, A.T.; Reddy, D.V.R. Cytopathology of Pigeonpea sterility mosaic virus in pigeonpea and Nicotiana benthamiana: Similarities with those of eriophyid mite-borne agents of undefined aetiology. Ann. Appl. Biol. 2002, 140, 87–96. [Google Scholar] [CrossRef]
- Kumar, P.L.; Jones, A.T.; Reddy, D.V.R. Mechanical transmission of Pigeonpea sterility mosaic virus. J. Mycol. Plant Pathol. 2002, 32, 88–89. [Google Scholar]
- Jones, A.T. Experimental transmission of viruses in diagnosis. In Diagnosis of plant virus diseases; Matthews, R.E.F., Ed.; CRC Press: Boca Raton, Florida, USA, 1993; pp. 49–72. [Google Scholar]
- Manjunatha, L. Detection and characterization of Pigeonpea sterility mosaic virus (PPSMV) and its management. Ph. D. Thesis, University of Agricultural Sciences, Bengaluru, India, 2012. [Google Scholar]
- Nene, Y.L. Sterility mosaic of pigeonpea: the challenge continues. Indian J. Mycol. Plant Pathol. 1995, 25, 1–11. [Google Scholar]
- Kumar, P.L.; Jones, A.T.; Reddy, D.V.R. ELISA for the detection of pigeonpea sterility mosaic virus. Patancheru 502 324, Andhra Pradesh, India: International Crops Research Institute for the Semi-Arid Tropics. 2000. Available online: http://www.icrisat.org/text/research/grep/homepage/archives_cd/frame.htm.
- Kumar, P.L.; Jones, A.T.; Reddy, D.V.R. A novel mite-transmitted virus with a divided RNA genome closely associated with pigeonpea sterility mosaic disease. Phytopathol. 2003, 93, 71–81. [Google Scholar]
- Kumar, P.L.; Jones, A.T.; Waliyar, F. Serological and nucleic acid based methods for detection of plant viruses: Methods manual. ICRISAT: Patancheru, A.P., India, 2004. [Google Scholar]
- Elbeaino, T.; Digiaro, M.; Uppala, M.; Sudini, H.K. Deep sequencing of Pigeonpea sterility mosaic virus discloses five RNA segments related to Emaraviruses. Virus Res. 2014, 188, 27–31. [Google Scholar] [CrossRef]
- Elbeaino, T.; Digiaro, M.; Uppala, M.; Sudini, H.K. Deep sequencing of dsRNAs recovered from mosaic-diseased pigeonpea reveals the presence of a novel Emaravirus, Pigeonpea sterility mosaic virus 2. Arch. Virol. 2015, 160, 2019–2029. [Google Scholar]
- Patil, B.P.; Meenakshi, D.; Ritesh, M. Variability of Emaravirus species associated with sterility mosaic disease of pigeonpea in India provides evidence of segment reassortment. Viruses 2017, 9, 183. [Google Scholar] [CrossRef]
- Latha, T.K.S.; Kuamr, P.L. Preliminary characterization of Coimbatore isolate of Pigeonpea sterility mosaic virus. Madras Agri. J. 2015, 102, 344–347. [Google Scholar]
- Tatineni, S.; McMechan, A.J.; Wosula, E.N.; Wegulo, S.N.; Graybosch, R.A.; French, R.; Hein, G.L. An eriophyid mite-transmitted plant virus contains eight genomic RNA segments with unusual heterogenicity in the nucleocapsid protein. J. Virol. 2014, 88, 11834–11845. [Google Scholar] [CrossRef]
- Sailaja, K. Biological and molecular characterization of Pigeonpea sterility mosaic virus. M.Sc Thesis, Acharya N.G. Ranga Agricultural University, Lam, Guntur, India, 2016. [Google Scholar]
- Padule, D.N.; Mewase, A.G.; Patel, B.P. Survey for the incidence of Fusarium wilt and other diseases of pigeonpea in central part of western Maharashtra. J. Maharashtra Agri. University. 1982, 118, 57–70. [Google Scholar]
- Paul, R.K.; Vennila, S.; Yadav, S.K.; Bhat, M.N.; Kumar, M.; Chandra, P.; Paul, A.K.; Prabhakar, M. Weather based forecasting of sterility mosaic disease in pigeonpea (Cajanus cajan) using machina learning techniques and hybrid models. Indian J. Agric. Sci. 2020, 90, 1952–1958. [Google Scholar] [CrossRef]
- Nene, Y.L.; Reddy, M.V. A new technique to screen pigeonpea for resistance to sterility mosaic. Trop. Grain Legume Bull. 1976, 5, 23–24. [Google Scholar]
- Nene, Y.L.; Reddy, M.V. Screening for resistance to sterility mosaic of pigeonpea. Plant. Dis. Rep. 1976, 66, 1034–1036. [Google Scholar]
- Nene, Y.L.; Kannaiyan, J.; Haware, M.P.; Reddy, M.V. Pigeonpea diseases: Resistance-screening techniques. Information Bulletin. ICRISAT, Patancheru, Andhra Pradesh, India. 1981, 1, 1–14. [Google Scholar]
- Pandey, S.; Sharma, M.; Guvvala, G.; Telangre, R. High throughput phenotyping of pigeonpea diseases: stepwise identification of host plant resistance. Information Bulletin; ICRISAT: Patancheru, Andhra Pradesh, India, 2012; p. 93. [Google Scholar]
- AICRP on Pigeonpea Annual Report 2020-21, All India Coordinated Research Project on Pigeonpea, ICAR-Indian Institute of Pulses Research, Kanpur-208024, pp. 406.
- Singh, A.K. Sterility mosaic of pigeonpea: etiology, epidemiology and management. Ph.D. Thesis, G. B. Pant University of Agriculture and Technology, Uttar Pradesh, India, 1992. [Google Scholar]
- De, R.K.; Dhar, V.; Rathore, Y.S. Control of mosaic in pigeonpea with insecticides and acaricides. Indian J. Pulses Res. 1996, 11, 87–89. [Google Scholar]
- Rathi, Y.P.S. Temik treatment of pigeonpea seeds for prevention of sterility mosaic. Acta Bot. Indica. 1997, 90–91. [Google Scholar]
- Ramakrishnan, K.; Kandaswamy, T.K. Investigation on virus disease of pulse crops in Tamil Nadu. Final Technical Report; Tamil Nadu Agricultural University: Coimbatore, Tamil Nadu, India, 1972. [Google Scholar]
- Narayanaswamy, C.A. Studies on Aceria cajani (Acari: Eriophyidae), mite vector of sterility mosaic disease with special reference to varietal screening and its interplant distribution. M.Sc Thesis, University of Agricultural Sciences, Banglore, India, 2004. [Google Scholar]
- Manjunatha, L.; Ramappa, H.K.; Mahantesha, S.R.V.; Gowda, M.B.; Rajappa, P.V.; Kavitha, T.R. Management of sterility mosaic disease (SMD) of pigeonpea. Plant Arch. 2012, 12, 1007–1012. [Google Scholar]
- Sudharani, S.; Amaresh, Y.S.; Naik, M.K.; Arunkumar, H.; Sunkad, G. Management of sterility mosaic disease through vector management. Indian J. Agric. Res. 2017, 51, 498–501. [Google Scholar]
- Rajeswari, E.; Smitha, K.P.; Kamalakannan, A.; Alice, D.; Kannan Bapu, J.R. Management of pigeonpea sterility mosaic disease. Legume Res. 2016, 39, 648–650. [Google Scholar] [CrossRef]
- Maurya, R.K.; Kumar, B.; Kumar, R.; Singh, M. Transmission of Pigeonpea sterility mosaic virus and management of sterility mosaic disease of pigeonpea by different acaricides under middle IGP of Bihar. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 3711–3716. [Google Scholar] [CrossRef]
- Gurha, S.N.; Singh, D.N.; Gopal, L.K. Effect of population density on the incidence of sterility and yellow mosaic disease of pigeonpea. Int. Pigeonpea Newsl. 1983, 2, 47–48. [Google Scholar]
- Bhatnagar, V.S.; Nene, Y.L.; Jadhaw, D.R. Sterility mosaic disease of pigeonpea in sorghum-based cropping system. Int. Pigeonpea Newsl. 1984, 3, 37–38. [Google Scholar]
- Nene, Y.L.; Reddy, M.V.; Beniwal, S.P.S.; Mahmood, M.; Zote, K.K.; Singh, R.N.; Sivaprakasam, K. Multilocational testing of pigeonpea for broad-based resistance sterility mosaic in India. Indian Phytopathol. 1989, 42, 444–448. [Google Scholar]
- Subramanian, K.S.; Samuel, G.S.; Janarthanan, R.; Kandaswamy, T.K. Studies on the varietal resistance of pigeonpea (Cajanus cajan) to sterility mosaic disease. Madras Agric. J. 1973, 60, 38–40. [Google Scholar]
- Rathi, Y.P.S. Studies on sterility mosaic disease of pigeonpea. Final technical report; GBPU &AT: Pantanagar, U.P., India, 1983. [Google Scholar]
- Singh, A.B.; Singh, D.P.; Singh, N.P.; Kumar, R. Evaluation of pigeonpea germplasm for resistance to sterility mosaic. Int. Chickpea Pigeonpea Newsl. 1995, 2, 69–70. [Google Scholar]
- Amin, K.S.; Reddy, M.V.; Nene, Y.L.; Raju, T.N.; Shukla, P.; Zote, K.K.; Arjunan, G.; Bendre, J.N.; Rathi, Y.P.S.; Sinha, B.K.; Gupta, R.P.; Anilkumar, T.B.; Chauhan, V.B.; Singh, G.; Jha, D.K.; Gangadharan, K. Multi-location evaluation of pigeonpea (Cajanus cajan) for broad based resistance to sterility mosaic disease in India. Indian J. Agri. Sci. 1993, 63, 542–546. [Google Scholar]
- Reddy, M.V.; Jain, K.C.; Chauhan, Y.S.; Singh, L. Wilt and sterility mosaic disease resistant pigeonpea genotype ICPL 87119 benefits farmers in Medak district of Andhra Pradesh, India. Int. Chickpea Pigeonpea Newsl. 1995, 2, 71–72. [Google Scholar]
- Rangaswamy, K.T.; Sulladmath, V.V.; Byre, G.M. Reaction of certain pigeonpea entries and varieties to sterility mosaic disease. Abs. Symposium on economically important diseases of crop plants, IPS southern chapter, University of Agricultural Sciences, Bangalore. 1997, 18–20. [Google Scholar]
- Nagaraj, K.M.; Chikkadevaiah, Muniyappa, V.; Rangaswamy, K.T.; Kumar, P.L. Evaluation of pigeonpea genotypes for resistance to Pigeonpea sterility mosaic virus-B isolate. Indian J. Plant Prot. 2006, 34, 216–220. [Google Scholar]
- Sharma, M.; Rathore, A.; Mangala, U.N.; Ghosh, R.; Sharma, S.; Upadhyay, H.D.; Pande, S. New source of resistance to fusarium wilt and sterility mosaic disease in a mini-core collection of pigeonpea germplasm. Eur J. Plant Pathol. 2012, 133, 707–714. [Google Scholar] [CrossRef]
- Singh, N.; Tyagi, R.K.; Pandey, C. Genetic resources of pigeonpea (Cajanus cajan): conservation for use. National Bureau of Plant Genetic Resources, New Delhi. 2013. [Google Scholar]
- Jaggal, L.G.; Patil, B.R.; Salimath, P.M.; Madhusudhan, K.; Patil, M.S.; Udikeri, S.S. Evaluation of mini core accessions of pigeonpea (Cajanus cajan (L.) Millsp.) against sterility mosaic disease and fusarium wilt. Karnataka J. Agric. Sci, 2014; 27, 337–339. [Google Scholar]
- Bhaskar, A.V. Screening of pigeonpea genotypes against wilt and sterility mosaic disease in Telangana state, India. Indian J. Agric. Res. 2016, 50, 172–176. [Google Scholar] [CrossRef]
- Tharageshwari, L.M.; Hemavathy, A.T.; Jayamani, P.; Karthiba, L. Evaluation of pigeonpea (Cajanus cajan) genotypes against pigeonpea sterility mosaic disease. Electron. J. Plant Breed. 2019, 10, 727–731. [Google Scholar] [CrossRef]
- International Crops Research Institute for the Semi-Arid Tropics (ICRISAT). 2021. Available online: https://www.icrisat.org/new-high-yielding-pigeonpea-variety-set-for-release-in-india-more-to-come/.
- Dhanushasree, M.A.; Thanga, H.; Gnanamalar, R.P.; Karthiba, L. Evaluation of Pigeonpea Genotypes against Sterility Mosaic Virus (PPSMV) Disease. Biol. Forum Int. J. 2022, 14, 1263–1268. [Google Scholar]
- Mediga, K.R.; Sunkad, G.; Kulkarni, S.; Chandran, U.S.S.; Ghosh, R.; Kshirsagar, D.; Sonnappa, M.; Katravath, S.; Parthasarathy, A.; Sharma, M. Multi-environment testing based G×E interactions reveal stable host-plant resistance against sterility mosaic disease in pigeonpea. Agron 2023, 13, 2859. [Google Scholar] [CrossRef]
- Seednet India Portal, Ministry of Agriculture and Farmers Welfare. Available online: https://seednet.gov.in/.
- Project coordinator’s report, Annual group meet on pigeonpea, 2015-16, ICAR, IIPR, Kanpur, pp. 272.
- Remanandan, P. The wild gene pool of Cajanus at ICRISAT: present and future. Proc. Int. Workshop on Pigeonpea, 15–19 December 1980; ICRISAT: Patancheru 502324 AP, India. [Google Scholar]
- Singh, V.K.; Khan, A.W.; Saxena, R.K.; Kumar, V.; Kale, S.M.; Sinha, P.; Chitikineni, A.; Pazhamala, L.T.; Garg, V.; Sharma, M.; Sameer Kumar, C.V.; Parupalli, P.; Vechalapu, S.; Patil, S.; Muniswamy, S.; Ghanta, A.; Yamini, K.N.; Dharmaraj, P.S.; Varshney, R.K. Next-generation sequencing for identification of candidate genes for Fusarium wilt and sterility mosaic disease in pigeonpea (Cajanus cajan). Plant Biotechnol. J. 2016, 14, 1183–1194. [Google Scholar] [CrossRef]
- Saxena, R.K.; Hake, A.; Bohra, A.; Khan, A.W.; Hingane, H.; Sultana, R.; Singh, I.P.; Naik, S.S.J.; Varshney, R.K. A diagnostic marker kit for Fusarium wilt and sterility mosaic diseases resistance in pigeonpea. Theor. Appl. Genet. 2021, 134, 367–379. [Google Scholar] [CrossRef]
- Patil, P.G.; Byregowda, M.; Patil, B.R.; Das, A.; Reena, M.G.A.; Sowjanya, M.S.; Shashidhar, H.E. Microsatellite markers linked to sterility mosaic disease resistance in pigeonpea (Cajanus cajan (L.) Millsp.). Legume Genom. Genet. 2016, 7, 1–9. [Google Scholar] [CrossRef]
- Prasanthi, L.R.; Reddy, B.V.B.; Rani, K.R.; Sivaprasad, Y.; Rajeswari, T.; Reddy, K.R. RAPD and SCAR markers linked to the sterility mosaic disease resistance gene in pigeonpea (Cajanus cajan (L.) Millisp.). Asian Australasian J. Plant Sci. Biotecnol 2009, 16–20. [Google Scholar]
- Gnanesh, B.N.; Bohra, A.; Sharma, M.; Byregowda, M.; Pande, S.; Wesley, V.; Saxena, R.K.; Saxena, K.B.; Kavi Kishor, P.B.; Varshney, R.K. Genetic mapping and quantitative trait locus analysis of resistance to sterility mosaic disease in pigeonpea (Cajanus cajan (L.) Milisp.). Field Crops Res. 2011, 123, 53–61. [Google Scholar] [CrossRef]
- Saxena, R.K.; Sandip, M.K.; Kumar, V.; Parupali, S.; Joshi, S.; Singh, V.; Garg, V.; Das, R.R.; Sharma, M.; Yamini, K.N.; Ghanta, A.; Rathore, A.; Sameerkumar, C.V.; Saxena, K.B.; Varshney, R.K. Genotyping-by-sequencing of three mapping populations for identification of candidate genomic regions for resistance to sterility mosaic disease in pigeonpea. Sci. Rep. 2017, 7, 1813. [Google Scholar] [CrossRef]
- Cao, Y.; Zhou, H.; Zhou, X.; Li, F. Control of plant viruses by CRISPR/Cas system-mediated adaptive immunity. Front. Microbiol. 2020, 11, 593700. [Google Scholar] [CrossRef]
Figure 2.
Pigeonpea sterility mosaic disease symptoms (a) mosaic; (b) chlorotic ringspots; (c) partial sterility; (d) complete sterile pigeon field.
Figure 2.
Pigeonpea sterility mosaic disease symptoms (a) mosaic; (b) chlorotic ringspots; (c) partial sterility; (d) complete sterile pigeon field.

Figure 3.
Scanning electron microscopic (SEM) picture of Aceria cajani, mite vector of pigeonpea sterility mosaic disease.
Figure 3.
Scanning electron microscopic (SEM) picture of Aceria cajani, mite vector of pigeonpea sterility mosaic disease.

Figure 4.
Leaf-stapling method of screening for sterility mosaic disease resistance.

Figure 5.
(a) Infector-hedge method (4-5 rows) of screening for sterility mosaic disease resistance. (b) Infector-row method (10:1) of screening for sterility mosaic disease resistance.
Figure 5.
(a) Infector-hedge method (4-5 rows) of screening for sterility mosaic disease resistance. (b) Infector-row method (10:1) of screening for sterility mosaic disease resistance.


Table 1.
The response of pigeonpea host-differentials to Pigeonpea sterility mosaic virus (PPSMV) isolates in India.
Table 1.
The response of pigeonpea host-differentials to Pigeonpea sterility mosaic virus (PPSMV) isolates in India.
| Sl. No. | Variety | Patancheru | Dholi | Varanasi | Badnapur | Rahuri | Bengaluru | Coimbatore | Dharwad | Tirupati | Vijayapura | Warangal |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Bahar | R | R | R | R | R | R | R | R | HS | R | R |
| 2 | BRG 1 | R | R | R | R | R | R | MR | R | R | MR | R |
| 3 | BRG 2 | R | R | R | R | R | S | MR | R | MR | MR | R |
| 4 | BRG 3 | R | R | R | R | R | R | R | R | R | MR | R |
| 5 | BRG 5 | * | R | R | R | MR | S | R | MR | R | MR | R |
| 6 | BSMR 736 | R | R | MR | R | R | R | MR | R | R | R | R |
| 7 | ICP 7035 | R | MR | R | R | R | R | R | R | HS | R | S |
| 8 | IPA 8F | R | R | R | R | HS | R | R | R | R | R | R |
| 9 | Purple 1 | MR | R | HS | R | R | S | R | MR | HS | R | S |
| 10 | ICP 2376 | R | R | R | R | R | MR | R | S | HS | R | S |
| 11 | ICP 8863 | HS | HS | HS | HS | HS | HS | S | HS | HS | MR | HS |
R-resistant (<10% SMD incidence); MR-moderately resistant (10.1-20% SMD incidence); S-susceptible (20.1-40% SMD incidence); and HS-highly susceptible (>40.1% SMD incidence) [10]. * not tested.
Table 2.
Available pigeonpea resistant sources against sterility mosaic disease (SMD) in India.
| Sl. No. | Genotypes/Cultivars | Reference |
|---|---|---|
| 1. | NP (WR) 15, P-435, P-1100, P-1289, P-1778 and P-2621 | [81] |
| 2. | L-3 and P-4875 | [90] |
| 3. | ICRISAT-3783, 6986, 1137, 2719, 7119, HY-3c, ICP-7035 and Atylosia lineata | [74] |
| 4. | ICRISAT-3784, 5449, 6497, 7035, 7119, Pant 8-76, 8-77, E-41, P-4785 and L-26 | [91] |
| 5. | ICP-7035, 3782, 6986, 6997, 7119, 7197, 7867, 7942 and 8136 | [28] |
| 6. | ICP 7378, 2S2 | [46] |
| 7. | Bahar, ICPLC-88046, DA-35, K32-1, Pusa-14, 19, Gant-9005, DA-I 1, 32, 33 | [92] |
| 8. | ICP 7035 | [93] |
| 9. | ICPL 787119 | [94] |
| 10. | ICP 7035 and HY3C | [95] |
| 11. | ICP 7035, MAL 14, and MAL 19 | [96] |
| 12. | ICP’s 7869, 9045, 11015, 11230, 11281, 11910, and 14976. Combined resistance to FW and SMD-ICP 6739, 8860, 11015, 13304, 14638, and 14819 | [97] |
| 13. | BRG3, IPA8F, IPA 15-F, GT 101, ICP 7035, ICPL 87091 and JKM 189 | [60] |
| 14. | ICPL 85010, IC245198, IC45768, IC73313, IC73336, IC73342, IC73340, IC73347, IC73727, IC73731, IC73735, IC73739, IC73841, IC73879, IC73880, IC73914, IC73925, IC73747, IC245198, IC45768, IC73332, IC74126, IC74084, IC74107, IC74123, Sehore, DPPA84-61-3, DPPA84-8-3, ICP786 (IC306500), ICP4395, ICP6997, ICP10976, CP10977, ICP7035 (IC306496), ICP8862, MA97, Rampur, Amar, Bahar, Bageshwari, PantA3, PantA104, PantA8505, PantA8508, Narendra arhar 1, comp 1-ESR6, Hy 3C, Maruti, BSMR1, BSMR2, BSMR604, BSMR736, Bhawanisagar 1, NPRR-1, ICP6997, MA3, LRG36, S1-3, ICPL88034(IC245153), DPPA85-3(IC527890), DPPA85-14(IC527891), ICPL7197, ICPL7264, PUSA14, USA15, PUSA17, PUSA18, Purple-1, PR 5149, PI 397430, KA-32-1, ICPL342, ICPL88034, DPPA85-2, DPPA85-15, BWR159, MA3, MA6, KPL44, AWR1,KAWR2, KAWR7, KAWR9, KAWR73, | [98] |
| 15. | ICP772, ICP939, ICP995, ICP1071, ICP1126, ICP1156, ICP1273, ICP1279, ICP2577, ICP2698, ICP2746, ICP3451, ICP3576, ICP4029, ICP4167, ICP4307, ICP7426, ICP4575, ICP4715, ICP6128, ICP6370, ICP6668, ICP6739, ICP6845, ICP6859, ICP6929, ICP7076, ICP7148, ICP7223, ICP6049, ICP7803, ICP7869, ICP8012, ICP8255, ICP8266, ICP8602, ICP8793, ICP8860, ICP8949, ICP9045, ICP9336, ICP9655, ICP10094, ICP10228, ICP11321, ICP10447, ICP11015, ICP11059, ICP11230, ICP11281, ICP11627, ICP11823, ICP11910, ICP12105, ICP12123, ICP12410, ICP12654, ICP13011, ICP13139, ICP13191, ICP13244, ICP13270, ICP13304, ICP13359, ICP13431, ICP13577, ICP13579, ICP13633, ICP13662, ICP13884, ICP14116, ICP14120, ICP14147, ICP14155, ICP14545, ICP14569, ICP14638, ICP14701, ICP14722, ICP14801, ICP14819, ICP14832, ICP14903, ICP14976, ICP15049, ICP15068, ICP15107, ICP15161, ICP15185, ICP12142, ICP16264 and ICP163 | [99] |
| 16. | ICPL 20094, ICPL 20106, ICPL 20098 and ICPL 20115 | [10] |
| 17. | ICPL-87119, ICPL-2376, BDN-2, PT-4-307, CORG-9701, BSMR-736, GRG-811 and BSMR-853 | [100] |
| 18. | DPP 2-89, DPP 3-182, IC 22557 and ICP 3666 | [101] |
| 19. | TDRG 59 (ICPL 99050) and Bheema | [102] |
| 20. | BWR 153 and CRG 16-07 | [103] |
| 21. | ICPL-16086 and ICPL-16087 | [35] |
| 22. | NAM 2082, NAM 2088, NAM 2089, NAM 2162, GRG 11, TS-3R-58-53-2, GRG 152, KRG 33, BSMR 736, AND ICPL 87119 | [104] |
| State | Varieties |
|---|---|
| Andhra Pradesh | Laxmi, LRG-41, LRG-38, WRG-27, WRG-53, Bahar, Pusa-9, NDA 1, WRG 65, Surya (MRG 1004) |
| Bihar | MA-6, Ajad, DA-11, IPA-203, Bahar, Pusa-9, Narendra Arhar-2 |
| Madhya Pradesh | JKM-189, TJT-501, JKM-7, TT-401, BSMR-175, ICPL-87119, BSMR-736 |
| Chhattisgarh | Rajiv Lochan, MA-3, ICPL-87119, Vipula, BSMR-853 |
| Gujarat | GT-100, GT-101, Banas, BDN-2, BSMR-853, AGT 2 |
| Haryana | Paras, Pusa-992, UPAS-120, AL-201, Manak, Pusa-855, PAU-881 |
| Karnataka | Vamban-3, CORG-9701, ICPL-84031, BRG-2, Maruti (ICP-8863), WRP-1, Asha (ICPL 87119), TS-3, KM 7 |
| Maharashtra | BDN-711, BSMR-736, AKT-8811, PKV Tara, Vipula, BDN-708, Asha, BSMR 175, Vaishali (BSMR 853) |
| Punjab | AL-201, PAU-881, Pusa-992, Upas-120 |
| Uttar Pradesh | Pradesh Bahar, NDA-1, NDA-2, Amar, MA-6, MAL-13, IPA-203, UPAS 120 |
| Rajasthan | UPAS-120, PA-291, Pusa-992, Asha (ICPL-87119), VLA-1 |
| Tamil Nadu | Co-6, CORG-9701, Vamban-3, ICPL-151, Vamban 1, Vamban 2 |
| Jharkhand | Bahar, Asha, MA-3 |
| Uttarakhand | VLA-1, PA-291, UPAS 120 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



