
Submitted:
13 June 2024
Posted:
13 June 2024
You are already at the latest version
Abstract
The Motile Aeromonad Septicemia (MAS) caused by Aeromonas species has been a serious problem in fish health management particularly in Nile tilapia (Orechromis niloticus). This study characterized an Aeromonas species isolated from farmed tilapia fingerlings in Binangonan, Rizal, Philippines, and tested for its pathogenicity in tank trials. We designated this isolate as Aeromonas veronii DFR01 (Diseased Fish Rizal) based on 16S rRNA phylogenetic analysis, 16S rRNA homology and MALDI-TOF MS identification with its biochemical profile generated from API and BiOLOG Gen III systems. A median lethal dose of A. veronii DFR01 was determined at 107 CFU/ml in tank trials and was utilized as whole cell inactivated antigen for oral vaccine development. The immunized tilapia fingerlings produced elevated levels of IgM in the blood as determined by enzyme-linked immunosorbent assay (ELISA). The increase in IgM levels is significantly higher 14 days post vaccination. The relative IgM gene expression as determined by quantitative polymerase chain reaction (qPCR) showed increasing levels after vaccination until 38 days of culture. The vaccinated fish showed 25-35% cumulative mortalities compared to the non-vaccinated-challenged group. Results showing improved relative percent survival (RPS) rate in tilapia fingerlings and induced immune response after vaccination indicate the potential use of the oral vaccine in cultured tilapia.
Keywords:
aeromonas
; MAS
; pathogenicity
; tilapia
; fish oral vaccine
1. Introduction
Increasing reports of fish morbidity and mortality associated with the motile aeromonad septicemia (MAS) (Monir et al., 2020) have been attributed to various Aeromonas species such as A. hydrophila (Monir et al., 2020), A. sobria (Li and Cai, 2010), A. veronii (Dong et al., 2017), and others (Dong et al., 2017; Soto-Rodriguez et al., 2018). It has affected a wide variety of freshwater fish species and occasionally marine fish (Larsen and Jensen, 1977) including humans (Dong et al., 2017; Daily et al., 1981; Janda and Abbott, 2010; Pessoa et al., 2022). A. hydrophila was reported from a diseased Oreochromis niloticus collected from intensive aquaculture farms in Luzon Island in the Philippines (Yambot, 1998; Pakingking et al., 2015). In our aim to develop a fish oral vaccine against bacterial pathogens of Nile tilapia, we isolated and characterized an Aeromonas species that caused serious morbidities and mortalities with signs of MAS. The study progressed into the development of an oral vaccine against A. veronii and potentially against other aeromonads.
Motile Aeromonad septicemia or fatal hemorrhagic septicemia has been identified as one of the major diseases of aquatic organisms particularly in tilapia (Dong et al., 2017; Haenen et al., 2022). The diseases caused by Aeromonas ranged from rapidly fatal septicemia to latent infections and has been referred to as hemorrhagic septicemia or MAS (Dong et al., 2017; Haenen et al., 2022). The fatal hemorrhagic septicemia has caused high mortality in reared Oreochromis mossambicus, O. niloticus and Tilapia zillii, (Lio-Po et al., 1983). The clinical signs of MAS are furunculosis or external ulcerations, abdominal swelling, and hemorrhagic septicemia (Haenen et al., 2022). The course of the disease usually progresses in an acute manner. Clinical conditions associated with systemic infection result in mortality within 24–48 hours.
The pathogen caused serious disease outbreaks resulting to high economic losses in aquaculture farms. Prolonged use of antibiotics for treatment can result to proliferation of antimicrobial resistance (AMR) in bacteria. Aeromonas spp. resistant to ampicillin, penicillin, rifampicin, cephalosporin, sulfonamide, and erythromycin have been reported (Dubey et al., 2022).
Vaccines are preventive measures to stimulate the fish adaptive and innate immune responses. The oral route of immunization is a non-invasive, stress- and cold-chain free method for mass vaccination of fish of all sizes. Most of the oral vaccines against bacteria are currently at the experimental stages (Kayansamruaj et al., 2020; Mutoloki et al., 2015). Attempts at oral vaccination of fish against MAS, vibriosis, yersiniosis, and furunculosis have either yielded mild and short-lived or inadequate responses (Leal et al., 2010). A major problem in the poor response to oral vaccination is the digestive degradation of antigens in the foregut before the vaccine reaches the immune responsive areas in the hind gut and other lymphoid organs (Mutoloki et al., 2015; Embregts and Forlenza, 2016). Strategies that have been explored for improving oral vaccination have included protected antigens such as encapsulated antigens (Mutoloki et al., 2015; Embregts and Forlenza, 2016) and use of biofilm of A. hydrophila for oral vaccination of carp to induce significantly higher antibody titers and protection (Azad et al., 2000; Steefaan et al., 2004). Stefaan et al. (2004) used commercial feed pellets as carrier for vaccine application wherein the feed pellets were coated with V. anguillarum bacterin suspension.
Other studies revealed that the oral or mucosal route of fish immunization conferred 80% to full protection (Palm et al., 1998; Esteve et al., 2004) with higher systemic immune response compared to other routes of administration (Akhlagi, 2000; Rodrigues et al., 2003). The oral vaccination of fish could be successful when the antigens reach the second gut of the intestine in sufficient quantities (Mutoloki et al., 2015).
The application of nanomaterials in drug delivery systems has gained interest and was used in this study for vaccination. Nanoparticles, when incorporated in the cell delivery vehicle, provide a controlled biochemical and mechanical microenvironment to the cells (Wan and Ying, 2010). A variety of naturally occurring and synthetic polymers are used in the encapsulation of antigens, usually through formation of covalent bonds. Nanomaterials have been shown to be more efficient compared to microparticles, due to the latter’s inconsistency in size with every alteration in production condition (Sinyakov et al., 2006) Several researchers have previously explored the possibility of a naturally occurring phyllosilicate material as a drug carrier due to its special intercalation activity. Drug and phyllosilicate interaction slows down drug release and absorption into the body, which may be beneficial when a controlled release is necessary for proper therapeutic action. In addition, due to the clay minerals’ high specific area, adsorption capacity, rheology, chemical inertness, and low toxicity, they have been used in many pharmaceutical formulations beneficial to human health (Carretero, 2002).
This study isolated and characterized an Aeromonas species from tilapia showing symptoms of fatal motile aeromonad septicemia, determined its pathogenicity and developed an oral vaccine using this isolate. This study also detected immunoglobulin (IgM) levels in pre- and post-vaccinated fishes challenged with the organism, and oral vaccine efficacy in tank trials.
2. Materials and Methods
Bacterial Isolation, Characterization and Identification
Isolation
The Aeromonas sp. DFR01 was isolated from Nile tilapia (O. niloticus) fingerlings from a fish farm in Binangonan, Rizal using Alkaline Peptone Salt Water (APSW) as enrichment medium and Starch-Ampicillin Agar (Palumbo, Maxino et al. 1985) as selective medium. White to yellowish colored colonies, 3 to 5 mm in diameter and amylase-positive (with clear zone surrounding the colony) were considered as presumptive Aeromonas. Nutrient Agar and Glutamine-Starch-Phenol Red agar medium (Sigma-Aldrich, USA) were used for maintenance of the organism.
Basic morphological characterization of the isolate was done by Gram staining method using the Hucker staining method (Murray et al., 1994); oxidase, catalase and hanging drop motility of the isolate were performed (Collins and Lyne, 1976).
Characterization Using API and BIOLOG
After the purification of the isolate by repeated streak plating, biochemical characterization was employed using the Analytical Profile Index - API 20E and API 20 NE system (Biomerieux, France) following the manufacturer’s instructions. Appropriate incubation temperatures and periods were observed (API 20E strip at 35°C for 24 hours and 20 NE strip at 29°C for 48 hours). The profiles of the organism were compared to the database for identification.
BiOLOG GEN III microplate (BiOLOG Inc., Hayward, CA, USA) was also used to generate characteristics of the isolate which include the following: utilization of carbon sources, resistance to inhibitory chemicals, growth at pH 5 and 6, and NaCl concentrations of 1%, 4% and 8%. Positive results are indicated by color development in a 96-well microtiter plate. The metabolic profile of the isolate is compared with the BIOLOG database for identification using the SIM index which refers to the similarity of the isolate to the other strains deposited in the BIOLOG database and DIST which refers to the mismatch (es) between the tested isolate and the database pattern. The SIM value at > 0.5 gives a strong ID and DIST should be < 5.00 and >2 when compared to the next matched ID (BiOLOG Microstation™ System/MicroLog User’s Guide).
Molecular Identification (16S rRNA Gene)
The genomic DNA was isolated using phenol chloroform method. Briefly, the bacterial cells were aseptically weighed, mixed with 200 µl 0.1 N NaOH and boiled to lyse the cells. About 100 µl sterile distilled water and 300 µl chloroform-isoamyl alcohol (24:1) were added. The cell suspension was vortexed, centrifuged at 13,000 rpm. The pellet was washed with 500 µl 70% ice cold ethanol and centrifuged at 13,000 rpm to remove salts and small organic molecules followed by careful removal of ethanol. The resulting DNA pellets was air-dried for a maximum of 15 minutes at 45 °C, re-suspended in DNAse/RNAse free water and boiled at 100 °C for 15 minutes. The DNA extract was stored in – 20 °C freezer.
PCRs were run in 20 µl volume reaction using Promega GoTaq® Flexi DNA polymerase (Promega, Fitchburg, WI), according to manufacturer’s specifications. PCR mixture contained 0.2 µl of 10 mM each dNTP, 1 µl of 25 mM MgCl2, 100-300 ng genomic DNA, 2 µl of 5x GoTaq Flexi buffer, 0.5 µl of 10 µm of each primer and 0.05 µl GoTaq DNA polymerase (5 U/µl). Cycling conditions were 96 °C for 5 min followed by 30 x 96 °C 1 min/ 55 °C 1 min/ 74 °C 1 min. Primers used to amplify the 16S rDNA gene were Aero I forward 5’-TAATGGCTCACCAAGGCGACGATCC-3’ and reverse 5’-CGTGCTGGCAACAAAGGACAG-3’ and Aero II forward 5’-CTTCGGGCCTTGCGCGATTGGATA-3’ and reverse 5’-GACGGGCGGTGTGTACAA-3’. The PCR products were checked using electrophoresis in a 1.5% TAE/agarose gel and visualized under UV after ethidium bromide staining.
The amplified DNA was sent to 1st Base Team (Singapore) for sequencing. The sequence chromatograms were checked using FinchTV 1.4.0. Multiple sequence alignments were performed using ClustalW (Larkin et al., 2007). Basic Local Alignment Search Tool (BLAST) was used to align the sequence with homologous sequence in the Genbank.
The final sequence alignment of 1,043 bp 16S rRNA gene was generated using MEGA X version 10.1.7 (Kumar et al., 2018). Vibrio alginolyticus ATCC 17749 (NR119049.1) was selected as an outgroup. The phylogenetic tree was calculated using the Hasegawa-Kishino-Yano +I+G model which is selected by jModelTest 2 (Darriba et al., 2012). The Bayesian Inference (BI) was used to reconstruct the tree using MrBayes v.3.2.7a (Ronquist et al., 2012). Statistical support for each node of the BI tree was based on two Markov Chain Monte Carlo (MCMC) runs (nchains = 4) for 20,000,000 generations with sampling every 20,000 generations, after discarding 30% of the resulting trees (i.e., the trees below the convergence, with a standard deviation of split frequencies above 0.01) as ‘burn-in’. In addition, Maximum Likelihood (ML) phylogenies were generated using MEGA X version 10.1.7 (Kumar et al., 2018). The phylogenetic trees were visualized using FigTree.v1.4.4 (http://tree.bio.ed.ac.uk/software/figtree/).
MALDI-TOF MS Identification
MALDI-TOF MS (EXS 2600 Zybio) was used to clarify the identity of the bacterial samples according to the manufacturer’s protocol (Zybio, China). Briefly, the spectrometer was calibrated with the Zybio microbiology calibrator solvent. Blank matrix solution (alpha-cyano-hydroxycinnamic acid) was prepared as control. Triplicates of each sample were prepared for analysis.
A direct smear method was used on bacterial colonies from agar plate culture. Fresh 24 hr culture of bacterial isolates were prepared using this method. About 1 µL of the matrix solution was dropped onto the bacterial smear and air-dried. For the extraction method, about 10 µL of bacterial cells was placed into a microcentrifuge tube and mixed with 300 µL distilled water and 900 µL ethanol. The tube was then centrifuged at 12,000 rpm. The supernatant was discarded, and the pellet was dried at room temperature. After drying, 20 µL of E. coli cell lysate was used as a standard and mixed well. The mixture was incubated for 5 minutes at room temperature before adding 20 µL reference lysate II. The tube was centrifuged again for 2 mins at 12,000 rpm and dropped with 1 µL supernatant onto the sample plate. Once dried, 1 µL of the matrix solution was added onto the sample plate and air-dried prior to MALDI-TOF MS.
LD50 Bacterial Challenge Experiment
To evaluate the pathogenicity of A. veronii DFR01, a total of 370 healthy Nile tilapia (O. niloticus) fingerlings, each with an average body weight of 6 grams were used. These fish were produced at the hatchery of the Binangonan Freshwater Station, Southeast Asian Fisheries Development Center (SEAFDEC). The fingerlings were acclimated for 14 days before the experiment. To ensure the fish were free of bacteria, 10 fish were randomly selected and their kidneys were aseptically dissected for bacterial isolation. All tested fish were confirmed to be bacteria-free. Five bacterial concentrations of A. veronii DFR01 were prepared: 109.8 CFU per fish, 108.8 CFU per fish, 107.8 CFU per fish, 106.8 CFU per fish, and 105.8 CFU per fish. Each group, consisting of twenty fish with three replicates per group, was subjected to intraperitoneal injection with the respective bacterial concentrations. The control group was injected with saline (NSS buffer only). Mortalities and signs of disease were monitored for 15 days. To confirm infection, bacteria were re-isolated from the kidneys of the experimentally infected fish.
Oral Vaccine Preparation, Vaccination and Challenge Experiment
Aeromonas veronii DFR01 was grown in Nutrient Broth (HiMedia Laboratories, India) for 24 h at 28 °C. The broth was then used to inoculate GSP (Glutamate Starch Phenol Red Agar; Sigma-Aldrich, USA) agar plates which were incubated for 24 h at 28°C. The cells were harvested into sterile tubes, resuspended in phosphate buffered saline (PBS, pH 7.2), pelleted at 14,000 rpm at 10°C and washed with PBS. The other steps in the preparation of the oral vaccine were based on the proprietary fish oral vaccine methods (Argayosa, 2017). Briefly, the cells were freeze-dried and inactivated. Inactivated cells were microencapsulated with a phyllosilicate carrier and added by mixing evenly with the feeds in a biosafety cabinet (BSL2) and air dried for at least three hours under the UV light to prevent contamination.
A total of 240 healthy tilapia fingerlings were used for the vaccination and challenge experiments. The fish were acclimated in a holding tank for 14 days before the start of the vaccination phase. They were then randomly divided into four groups, each containing 20 fish maintained in a 100-liter aquarium, with three replicates per group. The first group, called the Vaccinated group, was fed a SEAFDEC/AQD formulated diet (FD) supplemented with microencapsulated inactivated A. veronii DFR01 at 3.3 x 109 cells per fish. The second group, the Clay group, received FD supplemented with clay. The third group, the Bacterial Control, and the fourth group, the Negative Control, were both fed FD. Each group received their respective diets daily for 30 days at 2.5% of their body weight. On the 30th day post-immunization, the fish in the Vaccinated, Clay, and Bacterial Control groups were intraperitoneally challenged with A. veronii DFR01. This challenge involved anesthetizing the fish with MS222 (Tricaine Methanesulfonate) before injecting them with A. veronii DFR01 at a dose of 9.2 x 106 CFU/fish. The fish in the Negative Control group were injected with a normal saline solution (NSS) to serve as a control. After the injections, the fish were returned to their respective aquariums, and mortality rates were monitored periodically over the next 14 days. Freshly dead fish were collected for bacterial re-isolation from their kidneys. At the end of the experiment, the kidneys of all surviving and freshly dead fish were subjected to bacterial re-isolation using GSP (Glutamate Starch Phenol Red Agar; Sigma-Aldrich, USA). This procedure aimed to determine the efficacy of the vaccination by assessing the presence of A. veronii in the kidney tissues of the fish. The relative percent survival (RPS) was calculated as follows:
| RPS = 1- | (% mortality of vaccinated treatment | X 100 |
| % mortality of non vaccinated treatment) |
IgM determination by Enzyme-Linked Immunosorbent Assay (ELISA)
To determine the immune response of the experimental fish at different time points, IgM levels were measured in fish blood sera obtained at pre-treatment (prior to administration of vaccine), 14 days post-booster vaccination, and 14 days post infection. ELISA test was used to determine the relative IgM levels of the different treatments. Fish blood sera were diluted tenfold with PBS and added to sulfhydryl-bind strip well plates at a volume of 100 µL well-1 and blocked with 3% casein. After washing the casein, diluted (1:50) tilapia IgM antibody-HRP (C4-HRP Aquatic Diagnostics, Ltd., UK) was loaded into the wells and washed off using degassed PBS. The wells were then allowed to color react with the substrate at 100 µL well-1. The color reaction was arrested with 5M H2SO4 and absorbances read at 450 nm – 490 nm (StatFax 2100 Microplate Reader).
IgM Gene Expression Analysis by RT-PCR
To determine the IgM expression in different tissues of vaccinated and unvaccinated Total RNA was extracted from spleen of pre-vaccinated and vaccinated surviving fish 22 days post-challenge using Trizol reagent (Life Technologies) according to manufacturer’s protocol. The RNA template was used to synthesize the first strand cDNA using RT Maxime premix kit (Intron, Seoul, South Korea) according to manufacturer’s instruction. The reverse-transcript (RT) products were used for qPCR. RT-PCRs were run in 20 µl volume reaction with Biotium EVA Green using Promega GoTaq® Flexi DNA polymerase (Promega, Fitchburg, WI), according to manufacturer’s specifications. Primers used were IgM forward 5’-GCAGCAAGTTTTCTCACAGTA-3’ and reverse 5’-CCTCAAAGGCTCAATCAAGTC-3’, expected product is 485 bp. ß-actin was used as standard, ßa forward 5’- CGTGACATCAAGGAGAAGC-3’ and ßa reverse 5’- ACATCTGCTGGAAGGTGGAC-3’ with expected product of 321 bp (Harms et al., 2003). Cycling conditions were 96°C for 5 min followed by 30 x 96°C 1 min/ 60°C 1 min/ 74°C 1 min. Quantitative PCR was performed using Eco™ Real-Time PCR system (Illumina, San Diego, CA). The multiple of change was calculated relative to the non-vaccinated control after normalization to a housekeeping gene (beta-actin) using 2-∆∆Ct, where ∆Ct is (IgM Ct)- (beta-actin Ct), and ∆∆Ct is (∆Ct treated) – (∆Ct control) (Pfaffl, 2001; Schmittgen and Livak, 2008).
Statistical Analysis
Absorbance readings were subjected to statistical analysis to determine the significant differences from each treatment. Data were analyzed using one-way Analysis of Variance between treatments using GraphPad Prism version 6.01 for Windows, GraphPad Software, La Jolla, California, USA and Tukey Honest Significant Difference post-hoc tests were also employed.
3. Results
3.1. Isolation and Biochemical Characterization/Identification
The bacterial isolate was characterized as Gram-negative motile rods ranging from 3-5 μm in length as observed under the microscope (Figure 1). Bacterial colonies exhibited yellowish clearing zones in GSP agar plates and whitish colonies in modified GSP medium without phenol red and starch added with glucose, peptone and beef extract. Motile rods were observed under a microscope (100x).
The biochemical characterization using the API 20E and API 20 NE systems showed the following characteristics of the DFR01: production of β-galactosidase (ONPG), arginine dihydrolase (ADH), lysine decarboxylase (LDC), tryptophanase (IND), gelatinase, acetoin (butyl glycol pathway for D-Glc fermentation) (VP), fermentation of glucose and sucrose, reduction of nitrates to nitrites, assimilation of mannose, mannitol, N-Acetyl-Glucosamine, maltose, potassium gluconate, capric acid, malate and citrate). The isolate, however, was negative for ornithine decarboxylase (ODC), H2S production, and urease activity (Tables S1 and S2).
When compared to the API database, the biochemical profile of the DFR01 in API 20E showed good identification as Aeromonas hydrophila/caviae/sobria 2 (96.4%) and in API 20NE showed very good identification (99.2%) as Aeromonas sobria.
The BiOLOG system, molecular identification using 16S rRNA genes and MALDI-TOF MS were also done to clarify the identification.
The biochemical profile of the DFR01 in Biolog Gen III can be seen in Table S3. Results showed positive growth in the following substrates: alpha-D-Glc, D-Man, D-Fruc, D-Gal, glycerol, L-arginine, L-Glutamic acid (Table S3). It can grow in pH 5-6 and can tolerate up to 1% NaCl. It is sensitive to vancomycin and rifampicin antibiotics.
The Biolog Gen III was able to identify the isolate only to the genus level but listed A. veronii/sobria (DNAgroup 8) (SIM, 0.429, DIST. 4.590).
3.2. Molecular and MALDI-TOF Identification of the Aeromonas DFR01 Isolate
Phylogenetic tree was used to understand the relationship between the DFR01 isolate and representative Aeromonas species. Using 16S rRNA sequence, the DFR01 isolate grouped with a cluster of known species of A. veronii strains and A. sobria. (Figure 2).
Based on 16S rRNA homology using BLAST, DFR01 showed 99.14 % homology with A. veronii strain JCM 7375, A.veronii bv. veronii strain 35624 16S ribosomal RNA partial sequence and 98.67 % homology with A. sobria strain JCM 2139, ATCC 43979.
MALDI-TOF analysis of the isolate revealed A. veronii identity based on the cut-off score of above 2.0 generated from the mass-to-charge (m/z) ratio of the ribosomal protein fragments. The scores generated an average identification score of 2.32 +/- 0.12, for A. veronii. One of the representative spectra of the peaks of the m/z of the fragments generated is shown (Figure 3, Table S4) with a score of 2.42 for sample S16_2.
3.3. Experimental Infection
The mortality of fish infected with the DFR01 isolate is shown in Figure 4 and summarized in Table S5. The cumulative mortality in three groups infected with 1 × 109.8, 1 × 108.8 and 1 × 107.8 CFU/mL bacterial suspension are 95%, 75% and 65% respectively, while the other groups are 20% and 10%. The LD50 was calculated as 1.31 × 107 CFU/mL.
Gross clinical signs of MAS infection were observed such as ulceration, swollen belly, opaque to bulging eyes, fin rot, skin discoloration (Figure 5A). Mortalities were seen within 18h after injection. Gross internal clinical pathologies of the liver such as greenish coloration, hemorrhagic spots, ruptured abdomens were observed, possibly due to the congestion of internal organs. Infected fish died within 2 days post infection (Figure 5B). Bacterial isolates were re-isolated from the infected fish, the re-isolation rate from the kidney is indicated in Table S5.
3.4. Efficacy of Vaccine
Vaccinated groups demonstrated lower mortality rates after the bacterial challenge compared with the control groups. The relative percent survival (RPS) for the vaccinated group was 67% (Replicate 1) and 53% (Replicate 2) (Table S6, Figure 6). The cumulative mortality of the vaccine group was only 25%. All the dead fish were found positive for the organism upon bacterial re-isolation. This bacterium was also detected from surviving fish. The re-isolation of the bacteria in the pathogenicity tests confirm Aeromonas DFR01 as the causative agent of the motile aeromonad septicemia according to the Koch’s postulate.
At the end of the challenge test, all the unvaccinated survivors displayed clinical signs of MAS disease such as darker coloration of the skin and abnormal swimming behavior. While none of the vaccinated fish challenged exhibited any clinical signs.
3.5. Antibody Response by ELISA and Real Time PCR
Systemic IgM antibody levels at pre-vaccination (pre-treatment), 14 days post booster vaccination and 14 days post infection, as indicated by optical density (OD) at 450-490 nm, were determined by ELISA (Figure 7). The vaccinated fingerlings showed elevated levels of circulating IgM in the blood. The increase in IgM levels was significantly higher (p <0.05) 14 days post vaccination. The required second dose (before any booster) sustained the IgM compared to the lower levels in non-vaccinated fingerlings one week after the second dose. The levels continued to increase after two weeks post infection. Surviving non-vaccinated fingerlings showed lower IgM levels.
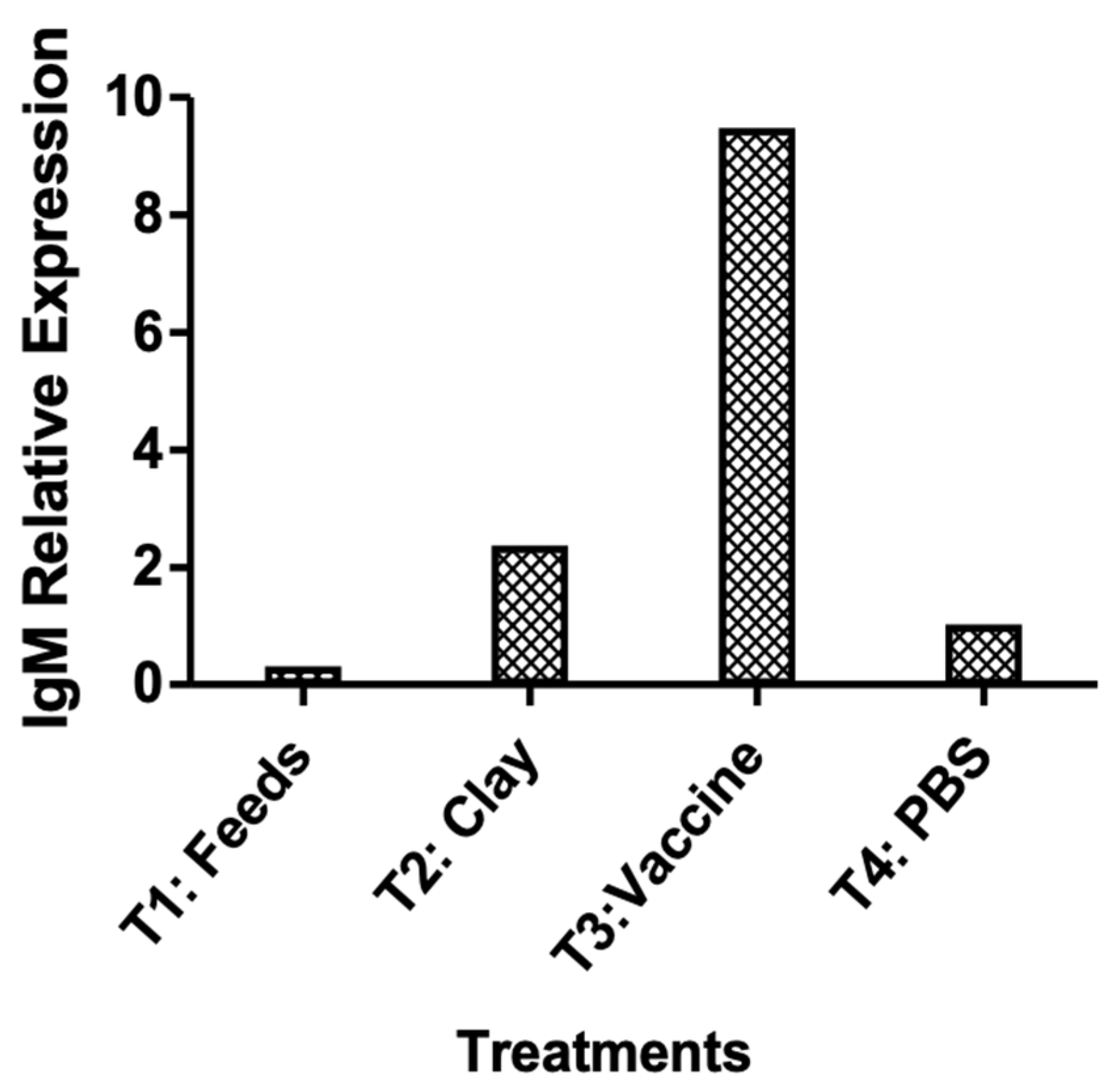
The relative IgM gene expression by RT-PCR suggests that the oral immunization effectively increased the IgM levels which conferred protection against bacterial challenge (Figure 8).
4. Discussion
The isolation and characterization of A. veronii from apparently healthy tilapia fingerlings which incurred high mortality after transport to the laboratory was meaningful to the successful development of the vaccine. This can be an effective strategy in isolating fish bacterial pathogens that might be present in the nursery culture that may later progress into fatal septicemia (MAS) infections in grow-out farms. The study was benchmarked to A. hydrophila reports where a vaccine was first reported to yield significant protection at 53-61% RPS (Ruangpan et al., 1986). A known A. hydrophila (BIOTECH 10089) reference strain and another A. hydrophila isolate (IDF2) used in the initial challenge tests (data not shown) failed to give high mortality rates after the challenge. Interestingly, we were able to isolate and characterize an A. veronii strain which showed mortality rates of the fish upon challenge.
The conventional techniques used to identify the bacteria, such as morphological, biochemical, and molecular techniques were not straightforward in the identification of Aeromonas at the species level. Aeromonas spp. have been reported and identified as an emerging pathogenic species and its taxonomy continues to change. For example, the A. veronii biogroup sobria was formerly assigned as A. sobria (Joseph et al., 1991). A. veronii was proposed for a separate species due to its ornithine decarboxylase-positive trait which distinguishes it from the other aeromonads (Joseph et al., 1991; Hickman-Brenner et al., 1987). Our MALDI-TOF analysis was consistent for A. veronii. The negative ornithine decarboxylase activity of the isolate yielded A. veronii biovar sobria which is a characteristic present in the DNA/DNA hybridization group or cluster. The biochemical features of the DFR01 isolate were not consistent with the A. veronii type strain ATCC 25604 (Hickman-Brenner et al., 1987), which has a positive reaction to ornithine decarboxylase and negative reaction to arginine dihydrolase.
On the reisolation of A. veronii DFR01 after experimental infection from the water and diseased fish tissues such as kidney, liver, and spleen, the combined biochemical properties demonstrate with high confidence that the motile aeromonad septicemia was caused by the isolate.
The proposed identification of the isolate as A. veronii was based on 16S rRNA phylogenetic analysis, homology and MALDI-TOF MS. MALDI-TOF analysis revealed A. veronii profile based on the ribosomal protein fragment sizes generated and compared to the database. API 20 E, NE and BiOLOG Gen III assays did not give definite identification to the species level. Using biochemical techniques to identify motile Aeromonas isolates at the genospecies level are imprecise because of the absence of a sharp dividing line between species (Paniagua et al., 1990). For example, difficulties in trying to separate A. caviae from A. hydrophila using biochemical tests have been encountered.
Although ornithine decarboxylase may be a key characteristic of the species (Janda and Abbot, 2010; Abbot et al., 2003) and DFR01 showed negative results for this enzyme, it is still possible that the isolate is A. veronii. The isolate was submitted for whole genome sequencing for definite identification and for mining and analyses of genes relevant to its pathogenicity. .
A. hydrophila isolates were found virulent in fish by intramuscular challenge, but the frequency of virulent strains was lower when intraperitoneal injections were used; mortality never exceeded 60%. This suggests that the strains studied lacked the protective surface characteristics as previously described by other authors (Mittal et al., 1980; Janda et al., 1987; Dooley and Trust, 1988) for highly virulent strains, which contributed to overcoming the peritoneal phagocytic barrier and the natural defenses of the host, permitting an easier establishment in the tissues. It is possible that known virulence factors of A. hydrophila for warm-blooded animals may not be attributable to its fish pathogenicity, as previously suggested (Santos et al., 1988). A different mechanism of virulence may be involved in the invasion of the bacteria in poikilothermic and homoeothermic hosts (Paniagua et al., 1990).
The increase of the IgM levels in the blood after immunization with the oral vaccine carrying the inactivated A. veronii DFR01 correlated to lower mortalities after the bacterial challenge. The high antibody titer was apparent two weeks after the booster application and was enhanced 14 days after the challenge. Aside from ELISA this was corroborated by the IgM gene expression analyses from the spleen samples of the immunized survivors. The activated systemic immune response produced higher survival rates after the bacterial challenge. Similarly, hybrid red tilapia treated with an oral vaccine composed of formalin-fixed A. hydrophila (with FIA) gave higher IgM levels than the unvaccinated group after 96 hours of bacterial challenge (Monir et al., 2020). On the other hand, serological analysis may not necessarily correlate with vaccine efficacy wherein Nile tilapia immunized with Flavobacterium columnare oral and parenteral vaccines produced high absorbance values in ELISA for circulating antibodies did not result to an effective protection after vaccination and challenge (Leal, et al., 2010).
Any adverse effect of the vaccine treatment may be detected in the blood chemistry profiles, but the results of the analyses of the ions, liver enzymes, and proteins indicate that the treated fish show healthy blood chemistry values (Table S7). The delivery of the inactivated or killed bacteria encapsulated with the naturally occurring phyllosilicate material is shown safe by these values compared to the untreated fingerlings.
Survival rates in the immunized group were significantly higher compared to the non-immunized fingerlings after an intraperitoneal injection of the pathogen. Considering the exposure of the vaccine through the acidic stomach of the fish, the increase in the relative percent survival of the vaccinated group may be attributed to successful encapsulation of the antigen and release in immune relevant mucosal tissues in the small intestines or second gut segment. The increase in circulating IgM levels is indicative of the efficacy of the oral vaccine for tilapia aquaculture systems. Several weeks to at least a month on the duration of the protection concur with the IgM levels after a month of vaccination and culture. For the oral vaccine technology like this, a convenient application of an oral booster to enhance protection until the fish reaches market size (~250g-1kg/pc) may be done in the field trials or commercial grow-out.
The potency of the vaccine could be dependent on many levels including bacterial strain and virulence factors present. Virulence genes in other A. veronii strains such as aerolysin (aerA), lipase, cytotoxic enterotoxin (alt) among others (Youssef et al., 2023) showed 90-100% mortality in pathogenicity tests. The virulence of the isolate was estimated from the lethal dose concentration determined for the isolate. The application of the isolate as an oral vaccine in aquaculture farms is possible as suspected cross protection between Aeromonas species due to common virulence factors shared by the aeromonads (Youssef et al., 2023).
The LD50 obtained for the isolate was 1.31 × 107 CFU/mL. Highly virulent fish bacterial pathogens can have an LD50 of about 106 CFU/fish while weakly virulent Aeromonas register LD50 of > 107 CFU/fish (Mittal et al., 1980).
Severe ulcerations in the stomach area were observed in the diseased fish, while skin and muscle lesions in discolored moribund fish were typical pathologies of MAS (Monir et al., 2020). Virulence test results may sometimes be variable when conducted in non-optimal conditions which are affected by the temperature, dissolved oxygen, and source. In addition, Aeromonas virulence can also be strain-specific (Rasmussen-Ivey et al., 2016).
The intraperitoneal injection used in the study was reported to be a more direct, reliable, and reproducible experimental method of infection (Angelidis et al., 2006), yet the natural route is through the mouth, gills, skin, gut, and anus. The natural oral route of infection and the corresponding oral vaccination strategy through a carrier on coated feeds might result in better protection after immunization. Oral administration of the antigen has been found to stimulate both systemic and mucosal responses compared to parenteral delivery where gut responses will be almost absent (Mutoloki et al., 2015).
In this study, the efficacy of an oral vaccine coupled with a naturally occurring phyllosilicate material was evaluated in Nile tilapia. Data suggest that the Aeromonas vaccine could activate a specific antibody response through the oral mucosal route. It had sufficiently produced and upregulated levels of the specific antibodies (IgM) which gave a significant protection after the bacterial challenge in tank trials.
The observed 67% RPS in tank trials indicate acceptable application in pre-commercial deployment in fish farms. Addition of a booster vaccination is suggested to increase protection and survival rates. Although the A. veronii DFR01 strain employed has given significant protection, other virulent Aeromonas spp. or strains may be isolated to improve the survival rates especially in grow-out areas showing outbreaks in bacterial infection caused by Aeromonas spp.
5. Conclusions
In conclusion, the A. veronii DFR01 strain isolated from fingerlings with Aeromonas infections showed pathogenicity for Nile Tilapia and was utilized as whole cell inactivated antigen for oral vaccine development. Oral vaccination produced increased IgM levels. The positive RPS in tank trials challenged with the bacterial isolate indicate potential for improvement for large scale deployment in pre-commercial trials in ponds or cages. Oral vaccines are still the most ideal delivery of vaccines for mass immunization of fish for aquaculture. It has promising advantages in mitigating the emergence of antibiotic resistance pathogens, improving fish health management and production for food security and One Health.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Biochemical characteristics of A. veronii DFR01 using API 20E; Table S2: Biochemical characteristics of A. veronii DFR01 using API 20 NE; Table S3: Biochemical characteristics of A. veronii DFR01 in Biolog Gen III; Table S4: MALDI-TOF scores of the A. veronii DFR01 isolated from a diseased fish (O. niloticus) from Binangonan, Rizal. Table S5: Inoculum dose, cumulative mortality and re-isolation rate of A. veronii DFR of experimentally infected O. niloticus juveniles (ABW: 6 g, n=20 fish/inoculum dose). Table S6: Mortality rate, relative percent survival (RPS) and bacterial re-isolation data from A. veronii DFR01-challenged tilapia at different treatments: clay, vaccine and control in two replicates. Table S7: Blood chemistry values for vaccinated and non-vaccinated O. niloticus.
Author Contributions
Conceptualization: AMA; Investigation and Methodology: AMA, MNMS, VBA, RVP, WB, MLS, RET; Funding acquisition, administration and supervision: AMA; Visualization and Validation: AMA, MNMS, VBA; Writing – original draft: AMA, MNMS, VBA Writing – Review and editing: AMA, MNMS, VBA, RVP.
Funding
Financial support for this study came from a research grant to AMA from the Department of Science and Technology (DOST) Philippine Council for Aquatic and Marine Research and Development (PCAMRD), now Philippine Council for Agriculture, Aquatic, and Natural Resources Research and Development (PCAARRD; 2021-01-A2-CRADLE-3044). Many thanks to Trinity University of Asia and Santeh Feeds Corp. for the support and counterpart funding.
Institutional Review Board Statement
All animal experiments were conducted in accordance with the institutional guidelines of Southeast Asian Fisheries Development Center Aquaculture Department (SEAFDEC/AQD), Tigbauan, Iloilo.
Data Availability Statement
The original data presented in the study are included in the article and Supplementary material, further inquiries can be directed to the corresponding author.
Acknowledgments
The authors wish to thank Rodolph Charles N. Rosel, Ariel Principe for the lab assistance, RainPhil for the use of the MALDI-TOF equipment, Philippine Nuclear Research Institute (PNRI) for bacterial inactivation, E. Gitalada for the financial reports and Chelo S. Pascua as co-inventor in the patent. Many thanks also to Alpha Rae M. Espigar for her assistance in the PCAMRD Year 1 Project.
Conflicts of Interest
The authors declare that they have no competing interests or personal relationships that may have influenced this paper.
References
- Monir, W.; et al. Pomegranate peel and moringa-based diets enhanced biochemical and immune parameters of Nile tilapia against bacterial infection by Aeromonas hydrophila. Microb Pathog 2020, 145, 104202. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cai, S.H. Identification and pathogenicity of Aeromonas sobria on tail-rot disease in juvenile tilapia Oreochromis niloticus. Curr Microbiol 2010, 62, 623–627. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.T.; et al. Aeromonas jandaei and Aeromonas veronii caused disease and mortality in Nile tilapia, Oreochromis niloticus (L). J Fish Dis 2017, 40, 1395–1403. [Google Scholar] [CrossRef] [PubMed]
- Soto-Rodriguez, S.A.; et al. Virulence of the fish pathogen Aeromonas dhakensis: genes involved, characterization and histopathology of experimentally infected hybrid tilapia. Dis Aquat Organ 2018, 129, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.L.; Jensen, N.J. An Aeromonas species implicated in ulcer-disease of the cod (Gadus morhua). Nord Vet Med 1977, 29, 199–211. [Google Scholar]
- Daily, O.P.; et al. Association of Aeromonas sobria with human infection. J Clin Microbiol 1981, 13, 769–777. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.M.; Abbott, S.L. The genus Aeromonas: taxonomy, pathogenicity, and infection. Clin Microbiol Rev 2010, 23, 35–73. [Google Scholar] [CrossRef]
- Pessoa, R.B.G.; et al. Aeromonas and Human Health Disorders: Clinical Approaches. Front Microbiol 2022, 13, 868890. [Google Scholar] [CrossRef]
- Yambot, A. Isolation of Aeromonas hydrophila from Oreochromis niloticus during Fish Disease Outbreaks in the Philippines. Asian Fisheries Science 1998, 10, 347–354. [Google Scholar] [CrossRef]
- Pakingking, R., Jr.; Palma, P.; Usero, R. Quantitative and qualitative analyses of the bacterial microbiota of tilapia (Oreochromis niloticus) cultured in earthen ponds in the Philippines. World J Microbiol Biotechnol 2015, 31, 265–275. [Google Scholar] [CrossRef]
- Haenen, O.; Dong, H.T.; Hoai, T.D.; Crumlish, M.; Karunasagar, I.; Barkham, T.; Chen, S.L.; Zadoks, R.; Kiermeier, A.; Wang, B.; Gamarro, E.G.; Takeuchi, M.; Azmai, M.N.A.; Fouz, B.; Pakingking, R., Jr.; Wei, Z.W.; Bondad-Reantaso, M. Bacterial diseases of tilapia, their zoonotic potential and risk of antimicrobial resistance. Reviews in Aquaculture 2022, 15, 154–185. [Google Scholar] [CrossRef]
- Lio-Po, G.; Pascual, J.P.; Santos, J.G. Fish quarantine and fish diseases in South East Asia; Jakarta, Indonesia; IDRC: Ottawa Ontario, 1983. [Google Scholar]
- Dubey, S.; et al. Aeromonas species isolated from aquatic organisms, insects, chicken, and humans in India show similar antimicrobial resistance profiles. Front Microbiol 2022, 13, 1008870. [Google Scholar] [CrossRef] [PubMed]
- Kayansamruaj, P.; Areechon, N.; Unajak, S. Development of fish vaccine in Southeast Asia: A challenge for the sustainability of SE Asia aquaculture. Fish Shellfish Immunol 2020, 103, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Mutoloki, S.; Munang’andu, H.M.; Evensen, O. Oral Vaccination of Fish - Antigen Preparations, Uptake, and Immune Induction. Front Immunol 2015, 6, 519. [Google Scholar] [CrossRef] [PubMed]
- Leal, C.A.G.; Carvalho-Castro, G.A.; Sacchetin, P.S.C.; Lopes, C.O.; Moraes, A.M.; Figueiredo, H.C.P. Oral and parenteral vaccines against Flavobacterium columnare: evaluation of humoral immune response by ELISA and in vivo efficiency in Nile tilapia (Oreochromis niloticus). Aquacult. Int. 2010, 18, 657–666. [Google Scholar] [CrossRef]
- Embregts, C.W.; Forlenza, M. Oral vaccination of fish: Lessons from humans and veterinary species. Dev Comp Immunol 2016, 64, 118–137. [Google Scholar] [CrossRef]
- Azad, I.S.; Shankar, K.M.; Mohan, C.V.; Kilata, B. Protective response of common carp vaccinated with biofilm and free cells of Aeromonas hydrophila. J. Aquaculture 2000, 15, 65–70. [Google Scholar]
- Stefaan, V.; Frans, O.; Renaar, K.; Armand, M. Oral vaccination of African catfish with Vibrio anguillarum O2: effect on antigen uptake and immune response by absorption enhancers in lag time coated pellets. Fish and Shellfish Immunology 2004, 16, 407–414. [Google Scholar]
- Palm, R.C.; Landolt, M.; Busch, R.A. Route of vaccine administration effects on the specific humoral response in rainbow trout. Diseases of Aquatic Organisms 1998, 33, 157–166. [Google Scholar]
- Esteve, G.; Barrera, R.; Amaro, C. Vaccination of market-size eels against vibriosis due to Vibrio vulnificus serovar E. Aquaculture 2004, 241, 9–19. [Google Scholar]
- Akhlagi, M. Immunogenicity of Aeromonas hydrophila in common carp. Journal of Veterinary Medicine Tehran 2000, 55, 56–62. [Google Scholar]
- Rodrigues, P.; Henrique, C.P.; Priscila, V.R.; Angela, M. Encapsulation of Aeromonas hydrophila in alginate microspheres for fish oral vaccine delivery. 2003. [Google Scholar]
- Wan, A.C.; Ying, J.Y. Nanomaterials for in situ cell delivery and tissue regeneration. Advanced Drug Delivery Reviews 2010, 62, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Sinyakov, M.S.; Dror, M.; Tennenbaum, T.L.; Saizberg, S.; Margel, S.; Avtalion, R.A. Nano- and microparticles as adjuvants in vaccine design: Success and failure is related to host natural antibodies. Vaccine 2006, 24, 6534–6541. [Google Scholar] [CrossRef] [PubMed]
- Carretero, M.I. Clay minerals and their beneficial effects upon human health. Applied Clay Science 2002, 21, 155–163. [Google Scholar]
- Palumbo, S.A.; Maxino, F.; Williams, A.C.; Buchanan, R.L.; Thayer, W.G. Starch-Ampicillin Agar for the Quantitative Detection of Aeromonas hydrophila. Appl Environ Microbiol 1985, 50, 1027–1030. [Google Scholar] [CrossRef] [PubMed]
- Murray, R.G.E.; Doetsch, R.N.; Robinow, C.F. Determinative and Cytological Light Microscopy. In Methods in General and Molecular Bacteriology; Gerhardt, P., Murray, R.G.E., Wood, W.A., Krieg, N.R., Eds.; American Society for Microbiology: USA, 1994. [Google Scholar]
- Collins, C.H.; Lyne, P.M. Microbiological methods, 4th ed.; Butterworths: London, 1976. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Clustal W and Clustal X version 2.0. Bioinformatics. 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol Biol 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: more models, new heuristics and parallel computing. Nat Methods 2012, 9, 772–772. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. 2012 Mrbayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Argayosa, A.M.; Pascua, C.S.; Sumera, F.; Yason, J.A.D.L.; Espigar, A.R. IPOPHIL (1/2013/000256), p. 18. https://onlineservices.ipophil.gov.ph/patgazette/IPASJournal/V2ON110_INV_2nd.pdf 2017.
- Harms, G.; Layton, A.C.; Dionisi, H.M.; Gregory, I.R.; Garrett, V.M.; Hawkins, S.A.; Robinson, K.G.; Sayler, G.S. Real-time PCR quantification of nitrifying bacteria in a municipal wastewater treatment plant. Environ Sci Technol 2003, 37, 343–351. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res 2001, 29, e45. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Ruangpan, L.; Kitao, T.; Yoshida, T. Protective efficacy of Aeromonas hydrophila vaccines in nile tilapia. Vet Immunol Immunopathol 1986, 12, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.W.; Carnahan, A.M.; Braytonl, P.R.; Fanning, G.R.; Almazan, R.; Drabick, C.; Trudo, E.W., Jr.; Colwell, R.R. Aeromonas jandaei and Aeromonas veronii dual infection of a human wound following aquatic exposure. J Clin Microbiol 1991, 565–569. [Google Scholar] [CrossRef]
- Hickman-Brenner, F.W.; et al. Aeromonas veronii, a new ornithine decarboxylase-positive species that may cause diarrhea. J Clin Microbiol 1987, 25, 900–906. [Google Scholar] [CrossRef] [PubMed]
- Paniagua, C.; Rivero, O.; Anguita, J.; Naharro, G. Pathogenicity factors and virulence for rainbow trout (Salmo gairdneri) of motile Aeromonas spp. isolated from a river. J Clin Microbiol 1990, 28, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Abbott, S.L.; Cheung, W.K.; Janda, J.M. The genus Aeromonas: biochemical characteristics, atypical reactions, and phenotypic identification schemes. J Clin Microbiol 2003, 41, 2348–2357. [Google Scholar] [CrossRef] [PubMed]
- Mittal, K.R.; Lalonde, G.; Leblanc, D.; Olivier, G.; Lallier, R. Aeromonas hydrophila in rainbow trout: relation between virulence and surface characteristics. Can J Microbiol 1980, 26, 1501–1503. [Google Scholar] [CrossRef] [PubMed]
- Janda, J.M.; Oshiro, L.S.; Abbott, S.L.; Duffey, P.S. Virulence markers of mesophilic aeromonads: association of the autoagglutination phenomenon with mouse pathogenicity and the presence of a peripheral cell-associated layer. Infect Immun 1987, 55, 3070–3077. [Google Scholar] [CrossRef] [PubMed]
- Dooley, J.S.; Trust, T.J. Surface protein composition of Aeromonas hydrophila strains virulent for fish: identification of a surface array protein. J Bacteriol 1988, 170, 499–506. [Google Scholar] [CrossRef]
- Santos, Y.; Toranzo, A.E.; Barja, J.L.; Nieto, T.P.; Villa, T.G. Virulence properties and enterotoxin production of Aeromonas strains isolated from fish. Infect Immun 1988, 56, 3285–3293. [Google Scholar] [CrossRef]
- Youssef, H.A.; Soror, H.F.; Matter, A.F. Virulence genes contributing to Aeromonas veronii pathogenicity in Nile tilapia (Oreochromis niloticus): approaching the development of live and inactivated vaccines. Aquaculture International 2023, 31, 1253–1267. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen-Ivey, C.R.; Figueras, M.J.; McGarey, D.; Liles, M.R. Virulence Factors of Aeromonas hydrophila: In the Wake of Reclassification. Front Microbiol 2016, 7, 1337. [Google Scholar] [CrossRef] [PubMed]
- Angelidis, P.; Karagiannis, D.; Crump, E.M. Efficacy of a Listonella anguillarum (syn. Vibrio anguillarum) vaccine for juvenile sea bass Dicentrarchus labrax. Dis Aquat Organ 2006, 71, 19–24. [Google Scholar] [PubMed]
Figure 1.
Cell and colony morphology of A. veronii DFR01. (A) Gram staining image showing Gram negative rods (bar = 10 microns); (B) Colonial morphology in modified GSP agar medium (bar = 10 mm) and (C) in GSP medium (bar = 10 mm).
Figure 1.
Cell and colony morphology of A. veronii DFR01. (A) Gram staining image showing Gram negative rods (bar = 10 microns); (B) Colonial morphology in modified GSP agar medium (bar = 10 mm) and (C) in GSP medium (bar = 10 mm).
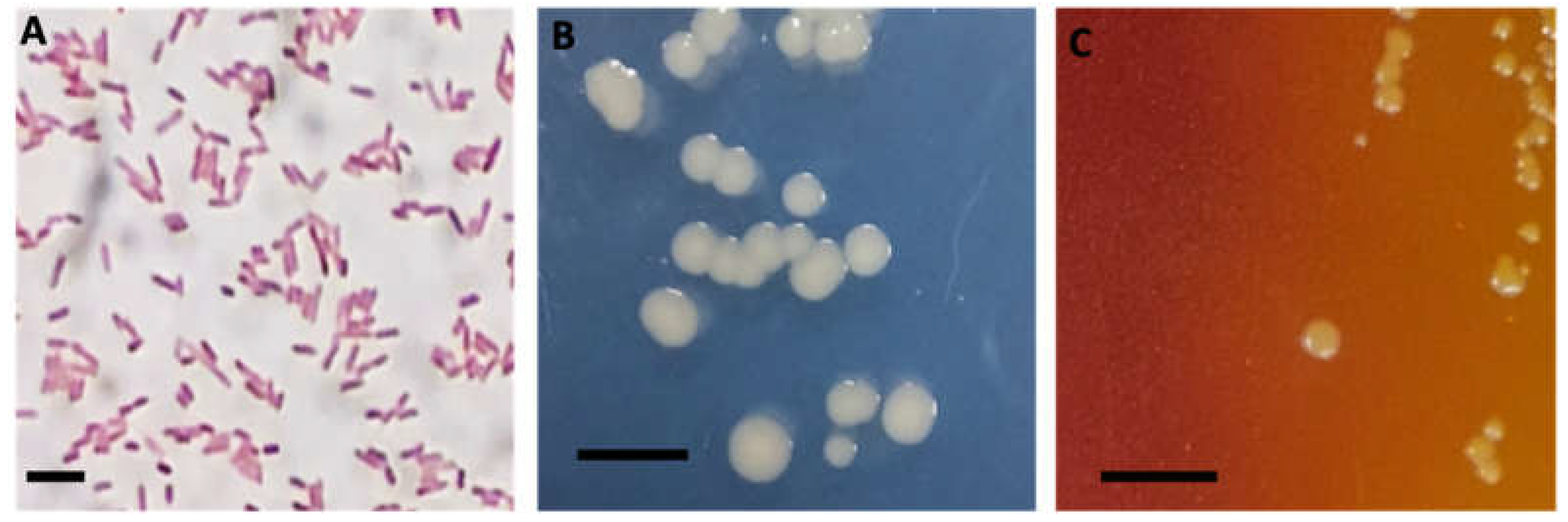
Figure 2.
Bayesian Inference (BI) phylogeny of Aeromonas based on 16S rRNA. Nodal support: posterior probabilities from BI / bootstrap percentage from maximum likelihood. Only posterior probabilities ≥ 0.5 and bootstrap percentages ≥ 50 are shown.
Figure 2.
Bayesian Inference (BI) phylogeny of Aeromonas based on 16S rRNA. Nodal support: posterior probabilities from BI / bootstrap percentage from maximum likelihood. Only posterior probabilities ≥ 0.5 and bootstrap percentages ≥ 50 are shown.
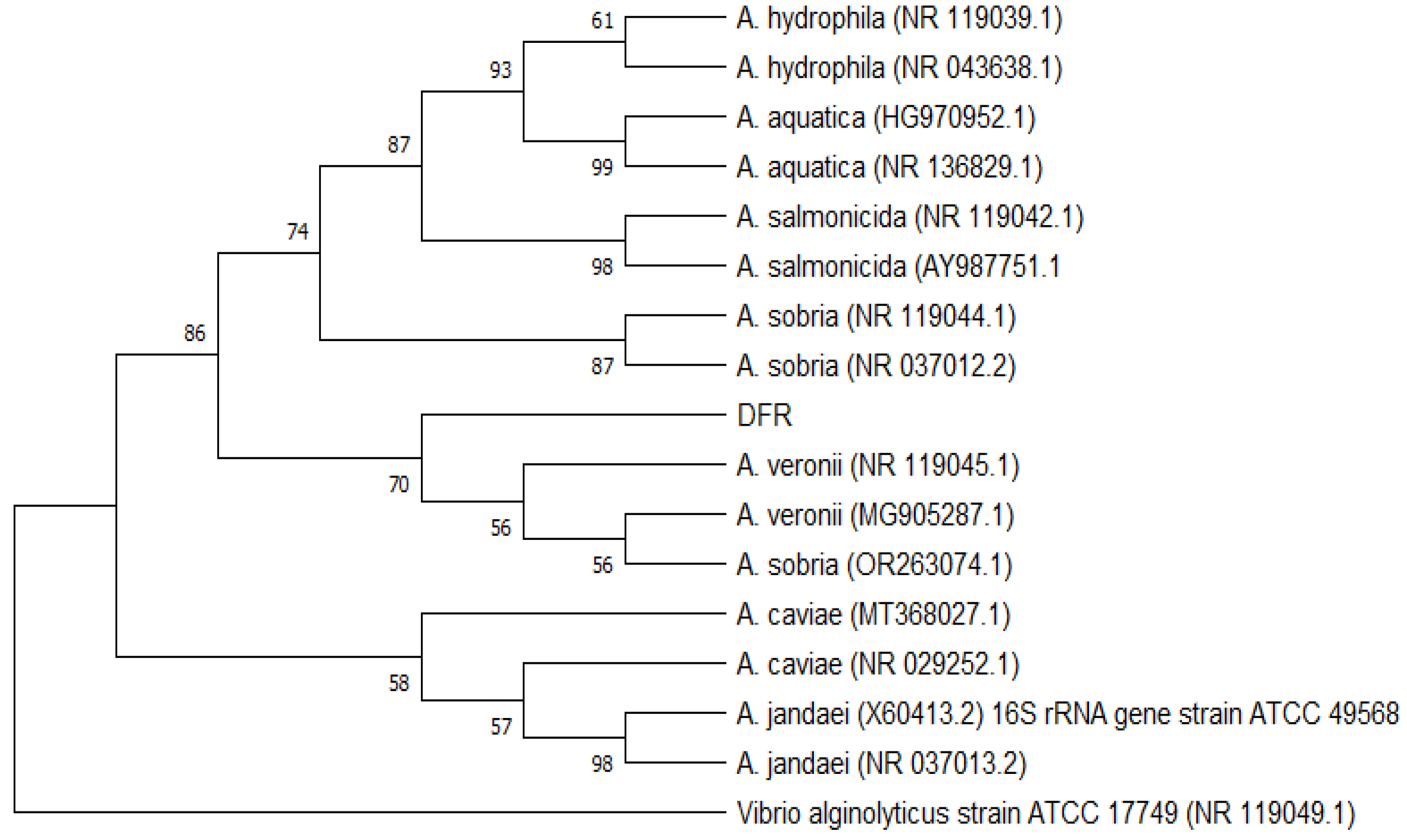
Figure 3.
MALDI-TOF spectrum of the ribosomal protein fragments of the Aeromonas veroni DFR01 mass cultivated in beef extract-peptone-glucose (BPG) broth media. Direct smear method of a bacterial colony was performed. The spectrum of six (6) replicates has an average identification score of 2.32 +/- 0.12.
Figure 3.
MALDI-TOF spectrum of the ribosomal protein fragments of the Aeromonas veroni DFR01 mass cultivated in beef extract-peptone-glucose (BPG) broth media. Direct smear method of a bacterial colony was performed. The spectrum of six (6) replicates has an average identification score of 2.32 +/- 0.12.
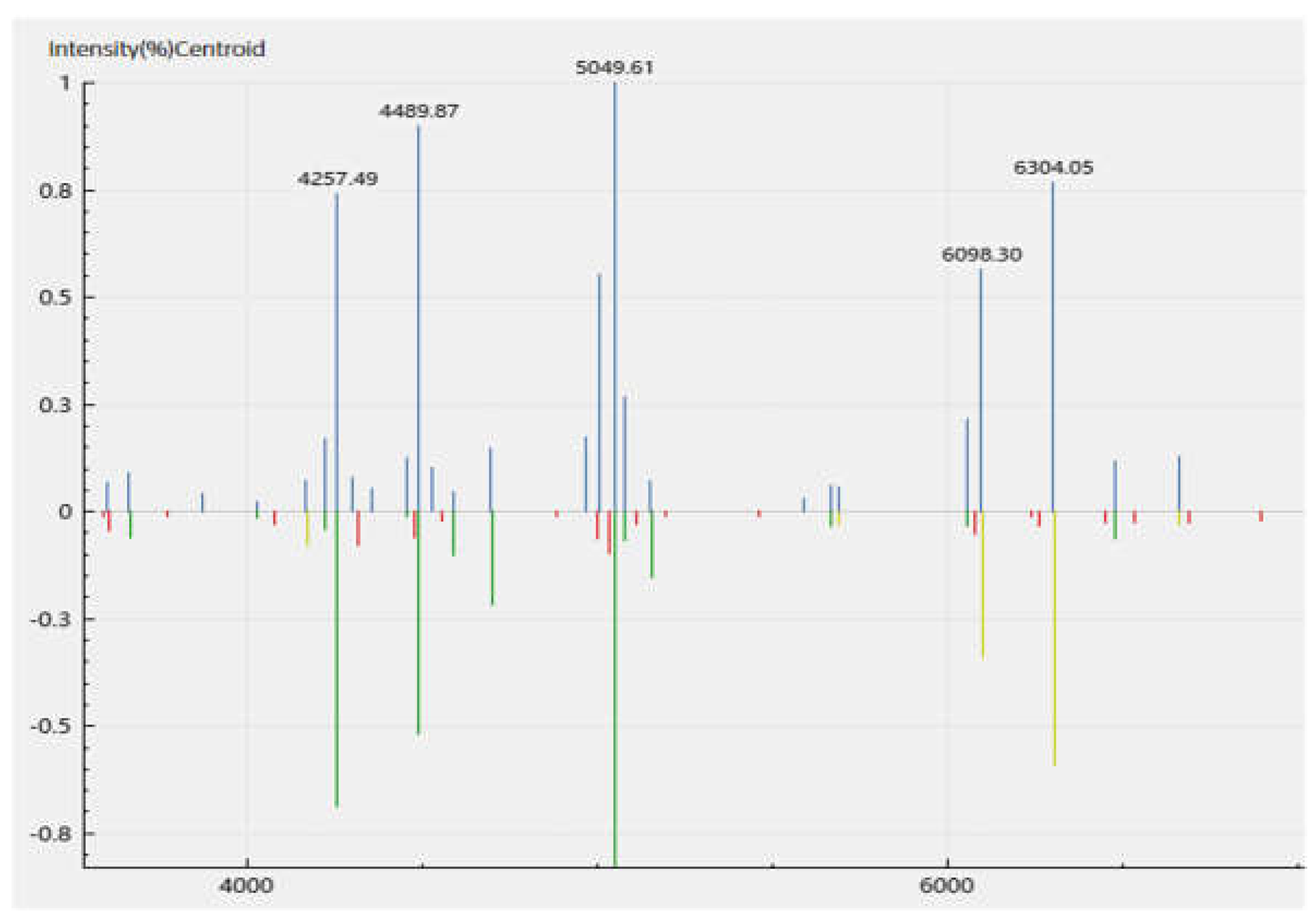
Figure 4.
Cumulative mortalities of tilapia juveniles (MBW:6 g; n=20 fish/ inoculum dose) experimentally infected by intraperitoneal injection with A. veronii DFR01. (■) 109.8 CFU fish-1; (□) 108.8 CFU fish-1; (●) 107.8 CFU fish-1; (○) 106.8 CFU fish-1; (▲) 105.8 CFU fish-1; (◊) control (NSS buffer only).
Figure 4.
Cumulative mortalities of tilapia juveniles (MBW:6 g; n=20 fish/ inoculum dose) experimentally infected by intraperitoneal injection with A. veronii DFR01. (■) 109.8 CFU fish-1; (□) 108.8 CFU fish-1; (●) 107.8 CFU fish-1; (○) 106.8 CFU fish-1; (▲) 105.8 CFU fish-1; (◊) control (NSS buffer only).
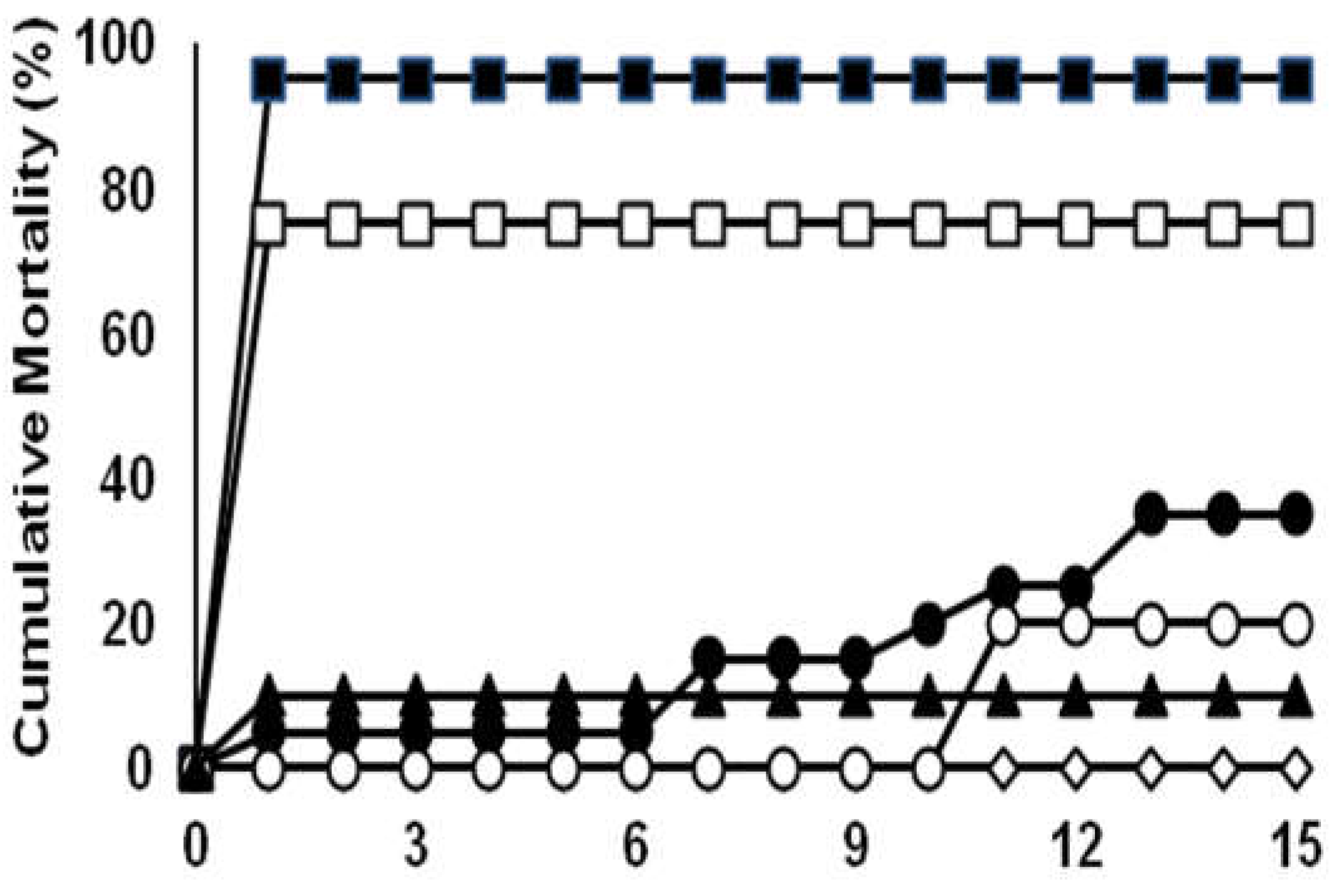
Figure 5.
(A) Gross clinical signs of motile aeromonad septicemia infection observed in diseased Nile tilapia (O. niloticus) infected with A. veronii DFR01 through intraperitoneal injection; (B) Internal signs of MAS in diseased fish showing pale liver, greenish coloration with hemorrhagic spots (A,B), and acute ulceration of abdominal cavity (C).
Figure 5.
(A) Gross clinical signs of motile aeromonad septicemia infection observed in diseased Nile tilapia (O. niloticus) infected with A. veronii DFR01 through intraperitoneal injection; (B) Internal signs of MAS in diseased fish showing pale liver, greenish coloration with hemorrhagic spots (A,B), and acute ulceration of abdominal cavity (C).
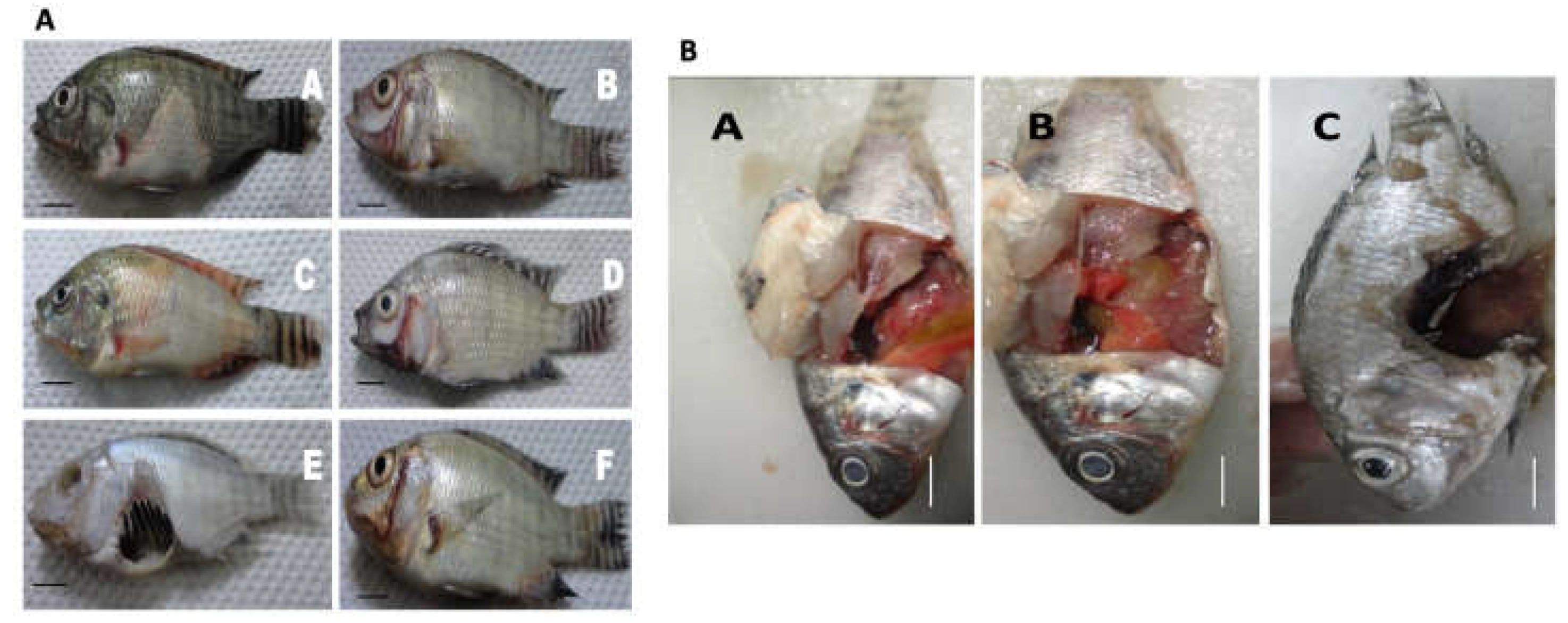
Figure 6.
Cumulative mortalities of the different groups of tilapia juveniles intraperitoneally challenged with A. veronii DFR01 at a dose of 1.31 × 107 CFU/mL. Clay (carrier) group: 45%; Vaccine group: 25%; Control (Non-vaccinated) - challenge group: 75%; Control (NSS) –mock challenge: 0%.
Figure 6.
Cumulative mortalities of the different groups of tilapia juveniles intraperitoneally challenged with A. veronii DFR01 at a dose of 1.31 × 107 CFU/mL. Clay (carrier) group: 45%; Vaccine group: 25%; Control (Non-vaccinated) - challenge group: 75%; Control (NSS) –mock challenge: 0%.
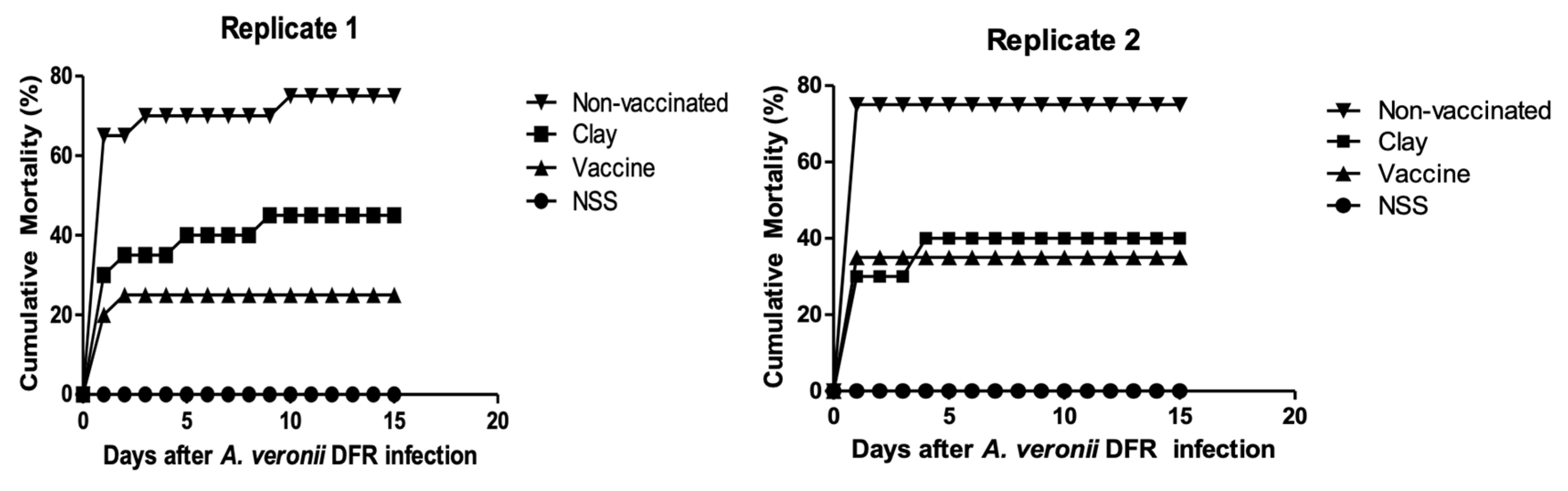
Figure 7.
IgM levels of the experimental O. niloticus samples during the laboratory trial of the vaccine, detected by ELISA.
Figure 7.
IgM levels of the experimental O. niloticus samples during the laboratory trial of the vaccine, detected by ELISA.
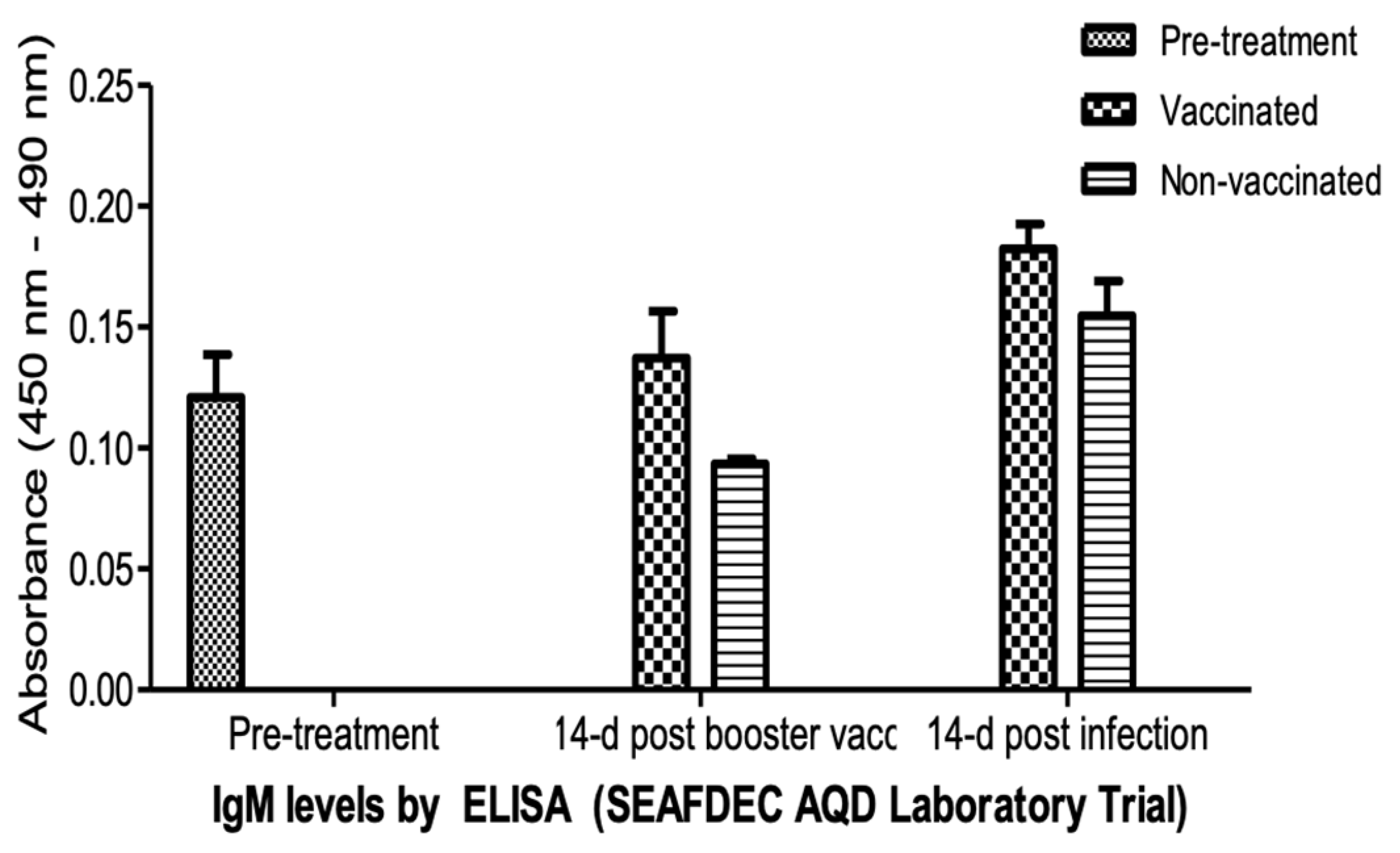
Figure 8.
qRT-PCR Relative IgM expression analysis in pre-vaccinated and vaccinated survivors. Gene expression in pooled spleen samples were normalized to β-actin and calibrated to the pre-vaccinated group.
Figure 8.
qRT-PCR Relative IgM expression analysis in pre-vaccinated and vaccinated survivors. Gene expression in pooled spleen samples were normalized to β-actin and calibrated to the pre-vaccinated group.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



