
Submitted:
14 June 2024
Posted:
14 June 2024
You are already at the latest version
Abstract
The wide metabolic diversity of microalgae, their fast growth rates, and cost-effective production make these organisms highly promising resources for a variety of biotechnological applications, addressing critical needs in industry, agriculture, and medicine. The utilization of microalgae in consortia with bacteria is proving to be valuable in different biotechnological fields, including treating various types of wastewaters, producing biofertilizers, and extracting various products from their biomass. Monoculture of the microalgae Chlamydomonas has been a prominent research model for many years, extensively utilized in studying photosynthesis, sulfur and phosphorus metabolism, nitrogen metabolism, respiration, and flagella synthesis, among others. Recent re-search has increasingly recognized the potential of Chlamydomonas-bacteria consortia as a bio-technological tool for various applications. Bioremediation of wastewater using Chlamydomonas, and its bacterial consortia presents significant potential for the sustainable reduction of contam-inants, while also facilitating resource recovery and valorization of microalgal biomass. Using Chlamydomonas and its bacterial consortia as biofertilizers can offers several benefits, such as en-hancing crop yield, protecting crops, maintaining soil fertility and stability, aiding in CO2 miti-gation, and contributing to sustainable agriculture practices. Chlamydomonas-bacterial consortia play a significant role in the production of high-value products, particularly in biofuel and en-hancing H2 production. This review aims to achieve a comprehensive understanding of the po-tential of Chlamydomonas monoculture and its bacterial consortia, identifying current applications, and proposing new research and development directions to maximize its potential.
Keywords:
Algal-microbial consortia
; microalga
; Chlamydomonas
; bioremediation
; biofertilization
; high-value added products.
1. Why Chlamydomonas and Why Its Consortia?
Microalgal cells are photosynthetic unicellular organisms that play a crucial role in global ecosystems, contributing to over half of the Earth's total photosynthetic activity and forming the backbone of the food chain [1]. Despite their prevalence in aquatic environments, microalgae thrive in various habitats, including the cold and irradiated poles, the subterranean environments of densely packed rhizospheres, and even within animal tissues in coral reefs [2]. Microalgae share a common evolutionary origin that can be traced back to a primary endosymbiotic event involving a cyanobacterium, which eventually evolved into the plastid. Subsequently, this plastid lineage expanded through secondary and tertiary endosymbiosis [3]. Microalgae play a crucial role in supporting ecosystems; however, they can also disrupt them through algal blooms, presenting substantial ecological, economic, and health risks [4]. Microalgae have demonstrate a wide range of metabolic capabilities, possessing various unique qualities that make them highly valuable for scientific studies [5]. Furthermore, the use of microalgae in diverse biotechnological applications has significantly increased in the last century. Emerging applications of microalgae include the production of biomaterials as alternatives to fossil-based materials, including biofertilizers, biostimulants, biopesticides, and energy sources like biodiesel, bioethanol, and biogas [6]. Microalgae are a source of bioactive compounds, including essential amino acids, polyunsaturated fatty acids, and antioxidants, which have been shown to positively impact nutrition and health [7]. Microalgae are considered a sustainable resource due to their ability to be employed in bioremediation processes, converting waste into valuable products within a circular economy model [8]. Consequently, microalgae are gaining global attention for their significant ecological and economic importance.
Chlamydomonas is a genus of single-celled biflagellated green microalgae that are commonly found in freshwater, but some species can also be found in habitats as diverse as saltwater, soil, and snow. Taxonomically, the genus Chlamydomonas comprises more than 500 species [9]. Chlamydomonas reinhardtii is undoubtedly the most studied species of Chlamydomonas genus. C. reinhardtii was isolated in 1945 from the soil of a potato field in Massachusetts, USA [10]. Since then, it has been utilized to investigate a range of research topics including photosynthesis, respiration, sulfur and phosphorus metabolism, nitrogen metabolism, amino acids and metals metabolism, biosynthetic pathways of starch, carotenoids, lipids, glycerolipids, heme groups, and chlorophyll. Additionally, it has been used to study other fundamental aspects such as the function of chaperones, proteases, flagella biogenesis, thioredoxins, and responses to various stress conditions [9]. The progress in different gene editing techniques like CRISPR-Cas9 in Chlamydomonas represent significant progress in addressing fundamental research inquiries and biotechnological applications [11]. All this demonstrates that Chlamydomonas has been intensively used in basic research; however, the practical application of Chlamydomonas for certain biotechnological purposes is not as numerous. Nevertheless, there has been a noticeable increasing interest for its potential biotech use in recent years [12].
Microalgae have been isolated from their natural habitats, with many proving challenging to cultivate axenically. This reliance on other microorganisms likely stems from the long-term co-evolution between microalgae and their beneficial microbe [13]. In this sense, microalgae have the capability to establish symbiotic associations with a diverse array of organisms, including bacteria, fungi, plants, and animals [14]. The term "phycosphere" refers to the area where algal exudates affect neighboring microorganisms [15]. In mutualistic relationships, microalgae typically provide their partner(s) with fixed carbon and oxygen in exchange for crucial nutrients and essential molecules like CO2, different vitamins, and nitrogen [16]. Moreover, a wide range of molecules can be secreted, detected, and utilized by the interacting partners to stablish a complex molecular dialogue. The comprehension of the complexity and dynamics of many of these symbiotic relationships is still in its early stages [17].
Since its initial discovery, there has been a significant rise in the number of studies exploring the symbiotic relationships between Chlamydomonas and bacteria, mainly focusing on fundamental research [18]. However, as far as we know, a single review compiling the potential of Chlamydomonas-bacteria consortia in crucial biotechnological tasks such as wastewater treatment, bioremediation, biofertilizer production, biomass valorization, and bio-product production has never been undertaken. Therefore, we summarize and categorize these reports here with the aim of highlighting the potential of Chlamydomonas-bacteria consortia to fulfill these tasks.
2. Wastewater Types and Composition
Water can be polluted by various streams, each with varying degrees of intensity. Wastewater can originate from various sources, including households, commercial sites, residential communities, industrial areas, surface runoff, recreational sites, institutional facilities, and agricultural areas (Figure 1). The composition of wastewater varies significantly depending on the source and industrial processes involved, comprising a diverse mixture of organic and inorganic compounds, as well as synthetic substances, with carbohydrates, fats, sugars, and amino acids being among the primary contaminants [19]. Persistent organic pollutants include chlorinated and aromatic compounds, such as organochlorine pesticides, polycyclic aromatic hydrocarbons, and polychlorinated biphenyls [20]. Inorganic constituents found in wastewater include substances such as sodium, calcium, nitrates, potassium, magnesium, sulfur, bicarbonate, arsenic, heavy metals, chlorides, phosphates, and non-metallic salts [21]. Municipal wastewater contains organic matter, nutrients, pathogens, and chemicals. Agricultural wastewater includes organic matter, pesticides, herbicides, and fertilizers. Dairy processing plant wastewater typically shows a very acidic pH, high concentrations of organic compounds, as well as high levels of organic and inorganic phosphate and nitrogen, fats, oils, grease, and detergents. Industrial wastewater may contain heavy metals, organic chemicals, and oils. Synthetic wastewater is a lab-created mimic used for research purposes [22]. Medical wastewater contains antibiotics and antibiotic-resistant genes, organic pollutants (e.g. phenol and its derivatives), refractory micropollutants (e.g., triclosan, ibuprofen, diclofenac) and toxic chemicals (e.g., cyanids, chlorinated lignin, dyes) [23]. Consequently, each type demands specific treatment due to its unique contaminants.
3. Wastewater Treatment Methods
As anthropogenic activities increase, leading to more complex wastewater compositions, it becomes essential to develop wastewater treatment methods that are easy to deploy, effective, and eco-friendly to mitigate water pollution. Traditional wastewater treatment methods involve physical, mechanical, chemical, and biological approaches (Figure 1) [24]. Physical methods include sedimentation, screening, and skimming; mechanical methods use filtration techniques; chemical methods involve processes such as adsorption, neutralization, disinfection, precipitation, and ion exchange [25]; and biological methods use microorganisms to degrade pollutants [26]. Integrating physical-chemical and biological methods offers a sustainable solution by reducing energy and chemical usage, cutting costs, and minimizing environmental impact [27].
Among the biological methods, phycoremediation, derived from the Greek word for algae 'phyco,' is an eco-friendly approach that employs various algae varieties such as microalgae, macroalgae, or cyanobacteria, to cleanse wastewater by eliminating pollutants or extracting products from it (Figure 1). Notable applications of phycoremediation include eliminating nutrients and xenobiotic compounds, reducing excessive nutrients in organic-rich effluents, mitigating CO2, treating effluents with heavy metal ions, and using algae as biosensors to monitor potentially harmful substances [28]. Phycoremediation not only helps in the removal of pollutants but also results in the production of algal biomass, which can be utilized for the production of various valuable products such as food, fertilizers, pharmaceuticals, and biofuels [29]. Chlamydomonas and other algae have been extensively utilized in wastewater treatment [30].
4. Microalgae Cultivation Methods
For the correct and efficient use of Chlamydomonas-bacteria cocultivation for any biotechnological purpose, it is essential to optimize the cultivation conditions and choose the appropriate cultivation method. A perfect microalgal cultivation system should be easy to manage, cost-effective to build, have sufficient light exposure, facilitate efficient gas-liquid transfer, and demonstrate minimal contamination risk. Photobioreactors have emerged as a potentially sustainable method for removing wastewater pollutants while producing microalgal biomass [31]. Typically, reactor configurations used in microalgal-bacteria systems include suspended systems (open reactors and closed reactors) and attached systems (such as biofilm reactors and encapsulated microalgae). Therefore, we will now present the main cultivation methods available for use with microalgae, highlighting the specific characteristics for microalgae-bacteria cocultivation and providing examples of their biotechnological potential in each case.
4.1. Open Reactors
Open reactors include natural ponds and lakes, as well as specially designed high-rate algal ponds (HRAPs) that are tanks or lagoons featuring a paddle wheel that circulates wastewater. HRAPs can be an cost-effective and environmentally friendly approach method for treating wastewater, as microalgae efficiently absorb nutrients such as nitrogen and phosphorus, as well as help remove organic and inorganic contaminants [32]. Unlike indoor laboratory-scale cultivation, outdoor algal cultivation employing HRAPs are heavily influenced by various uncontrollable environmental factors, such as seasonal changes and weather conditions [33]. The following are the generally accepted design parameters for algae-bacteria HRAPs: depths typically range from 0.2 to 1 m, depending on wastewater clarity for light penetration; horizontal water velocities between 0.09 and 0.3 m/s are recommended to ensure good mixing; hydraulic residence times (HRTs) range from 3 to 15 days; and HRAP areas vary from 1,000 to 50,000 m² [34,35]. Numerous studies have reported the use of HRAP in wastewater treatment, primarily focusing on genera such as Scenedesmus and Chlorella [36]. It has been reported that NH4+ was primarily removed through nitrification, followed by assimilation and denitrification, when anaerobically digested swine slurry was treated in algae-bacteria in HRAP [37]. However, very few records exist of applying HRAP with Chlamydomonas. In a pilot-scale experimental wastewater treatment using HRAP with Chlamydomonas sp., a reduction of approximately 90% in biochemical oxygen demand, 65% in chemical oxygen demand (COD), 20% in total phosphorus, and 46% in total nitrogen was reported [38]. A study on the bioremediation of piggery wastewater using HRAP with Chlamydomonas sp. revealed an average COD and total nitrogen removal efficiencies of 76% and 88%, respectively [39]. In another study using HRAP with Chlamydomonas sp. for treating municipal wastewater, the average reductions of volatile suspended solids, biochemical oxygen demand, and total nitrogen were 63%, 98%, and 76%, respectively [40].
4.2. Closed Photobioreactors
Closed photobioreactors (PBR) are enclosed systems utilized for the cultivation of microalgae and other phototrophic microorganisms. PBRs can be operated in any open space and provide excellent control over culture conditions with minimal risk of contamination. However, PBRs are costlier to build and necessitate the use of transparent materials like glass and acrylic for their construction. PBRs are characterized by a narrow light path and a large illuminated surface-to-volume ratio, which maximizes light energy capture and conversion [41]. PBR modules can be organized in various configurations, such as horizontal, inclined, vertical, or spiral arrangements. Flat plate and tubular photobioreactors are prevalent designs because of their extensive illuminated surfaces [42]. There are numerous studies utilizing PBR with Chlamydomonas, with many focusing on basic science research. Next, we will present some recent studies that have led PBR applications with significant biotechnological potential. The utilization of the phototactic response of C. reinhardtii to induce biomixing in PBR has been investigated. By exploiting this phototactic mechanism, C. reinhardtii can be stimulated to swim in opposite directions, providing mixing and ensuring access to nutrients without additional energetic costs. This approach could potentially replace mechanical agitation in certain circumstances, enhancing energy efficiency [43]. The multi-scale modular PBR, Antares I, has been tested with C. reinhardtii and has shown improved biomass production compared to traditional flask systems. With Antares I, the estimated doubling time for Chlamydomonas culture was almost half of that reported for culture using similar media and light conditions [44]. A drawback of monoculture microalgae in PBRs is that dissolved oxygen accumulation can reach up to 400% of air saturation, which is detrimental to microalgae growth [45]. However, the high oxygen consumption by bacteria in co-cultured algal-bacteria systems has been shown to reduce these negative effects by decreasing the amount of dissolved oxygen available [46]. When municipal wastewater was treated in a stirred algal-bacterial PBR, it was estimated that 40-53% and 17-20% of NH3 removal were attributed to bacterial assimilation and nitrification, respectively [47].
4.3. Biofilm Reactors
In biofilm reactors, microalgae are immobilized on a surface that acts as a support, resulting in the formation of a continuous layer. This approach provides benefits such as increased cell concentration per volume of medium, simplified harvesting, and decreased or minimal presence of cells in the effluent [48]. Algal-bacteria biofilms can be categorized as either stationary or mobile, based on the movement of the supporting materials. One of the primary advantages of microalgal biofilm reactors compared to other techniques is that the extraction and dewatering of algae cells from biofilms are simplified. This is due to the ease with which the attached cells can be separated from their surrounding growth medium, eliminating the need for costly separation methods like filtration, centrifugation, flocculation, or settling/floating the biomass. In these systems, the biomass needs to be collected by removing it from the support medium through scraping [49]. So far, the basic principles governing the colonization of surfaces by motile, photosynthetic microorganisms remain largely unexplored. The regulation of the adhesion properties of Chlamydomonas could significantly enhance the efficiency of biofilm reactors by controlling surface colonization and biofilm formation. Research has shown that the surface adhesion of C. reinhardtii is flagella-mediated and largely substrate-independent, enabling it to adhere to any type of surface [50]. However, it has been shown that the biofilm adhesion of C. reinhardtii is controlled by the type of light, being activated in blue light and deactivated under red light [51]. The phosphate-hyperaccumulating strain Chlamydomonas pulvinata TCF-48g has been tested using the biofilm system to recover phosphate from municipal wastewater, demonstrating a high phosphorus removal rate of 70%. [52]. With Chlamydomonas sp. JSC4, this technique has been successfully optimized in the removal of phosphorus, nitrogen, and copper from swine wastewater [53].
In biofilm reactors, the medium passes through the bioreactor while the biomass remains attached to a stationary support medium; therefore, the residence time of the algae and bacteria is much longer. This allows algal-bacteria biofilm reactors to be operated at higher organic loading rates and shorter hydraulic retention times than suspended growth systems because communities with slow growth rates are retained in the reactor. Numerous experiments, both small-scale and large-scale, have been conducted using algal-bacteria biofilm reactors for wastewater treatment [54]. Algal-bacteria biofilm reactors can be flushed with CO2 to enhance biomass productivity or integrated with other treatment processes to enhance the efficiency of wastewater treatment. A biomass productivity of 60 g/m²/day has been reported in algal-bacteria biofilm reactors using synthetic wastewater and purged with 0.5% CO2 enriched air [55]. Utilizing an algal-bacteria biofilm reactor in conjunction with additional reactors offers the potential to refine secondary wastewater effluent, resulting in a further reduction of total suspended solids concentrations to below 0.5 mg/L [56].
4.4. Encapsulation
The encapsulation of microalgae involves trapping the microalgae within coating materials, resulting in the formation of beads. This process offers various advantages, such as promoting controlled release, protecting the formation of bioactive compounds, enhancing bioavailability, and improving solubility [57]. Various materials such as alginate, chitosan, carrageenan, and polyvinyl, have been used for the immobilization of microalgae [58]. With Chlamydomonas, alginate has been the most extensively used for its low cost, biocompatibility, transparency, and permeability, which facilitate the diffusion of nutrients and light [59]. Additionally, the preparation of alginate beads is a rapid and straightforward process that can be easily scaled up [60]. The encapsulation of C. reinhardtii in alginate has been successfully carried out to remove various types of contaminants such as nitrogen, phosphorus, cadmium, lead, and mercury [61] or even phenol [62]. In C. reinhardtii alginate beads, pore size has been shown to be critical for contaminant removal, with the highest removal efficiency obtained with a gel bead pore size of 3.5 mm [63]. Silica hydrogels have been utilized to entrap C. reinhardtii cells, offering some advantages over alginate such as higher transparency and greater stability against ions and microbial attacks. Silica hydrogels encapsulating Chlamydomonas have demonstrated potential for applications such as hydrogen production [64]. One drawback of alginate encapsulation is its high porosity, which can lead to the release of large molecules. Nevertheless, it has been noted in C. reinhardtii that the combination of alginate and silica to create hybrid beads can offer enhanced properties that surpass this limitation [65]. Single-cell encapsulation involves coating individual cells with metal-phenolic networks to create a mechanical barrier. With microalgae, this method was first employed with C. reinhardtii, finding that this approach helped delay the proliferation of the coated cells and effectively promoted flocculation [66]. One advantage of encapsulation is that it can be performed with multiple organisms simultaneously, a process known as co-immobilization. The co-immobilization of C. reinhardtii with the acetate-producing cyanobacteria Synechococcus sp. PCC 7002 increases the biomass content [67]. Co-immobilization of Chlamydomonas and the cyanobacteria Lyngbya sp. on silica hydrogel exhibited a 92.5% removal of Pb²⁺ from wastewater [68]. Studies have also shown that in the co-immobilization of Chlamydomonas with the nitrogen-fixing bacterium Azospirillum brasilense, there is a mutualistic relationship supported by the exchange of tryptophan and indole-3-acetic acid (IAA), respectively. This relationship increases microalgal CO2 fixation and biomass production [69].
5. Main Mechanisms and Molecules Bioremediated by Chlamydomonas
Chlamydomonas has demonstrated remarkable capacity and diversity in bioremediating various molecules. Next, we will outline the different mechanisms and types of molecules that can be bioremediated by microalgae, particularly Chlamydomonas. Microalgae have the capacity to absorb and degrade pollutants like heavy metals, hydrocarbons, and pesticides using mechanisms such as biosorption, bioaccumulation, and biotransformation [70] (Figure 1).
5.1. Biosorption
Biosorption is a passive process where microalgae act as a biological sorbent to bind and concentrate pollutants. Microalgae utilize their cell wall and various chemical groups to attract and retain metallic and organic contaminants. The pollutants adhere to the algal membrane and are separated due to the presence of receptors that can bind and attract them [71]. Biosorption is indeed a well-studied mechanism for removing heavy metals from wastewater. C. reinhardtii has demonstrated a high capacity to remove through biosorption, arsenic [72], copper, boron and manganese [73], nickel [74], uranium [75], zinc and cadmium [76].
5.2. Bioaccumulation
Microalgae can eliminate contaminants through the process known as bioaccumulation. The difference between biosorption and bioaccumulation processes lies in the fact that biosorption is a passive process in which microorganisms utilize their cellular structure to trap pollutants, while bioaccumulation is an active process characterized by the buildup of pollutants in the biomass of microalgae, achieved through either accumulation or uptake into intracellular spaces [77]. C. reinhardtii has been shown to bioaccumulated several compounds as o-nitrophenol [78], Prometryne a herbicide [79], and Chlamydomonas mexicana carbamazepine, an antiepileptic agent [80].
5.3. Biotransformation
Biotransformation involves the degradation of pollutants, either within or outside cells, aided by enzymes [81]. While biosorption and bioaccumulation do not raise significant concerns, biotransformation presents greater challenges due to the potential for its byproducts to be more toxic than the original compounds. Some of the pollutants removed by biotransformation by C. reinhardtii include polystyrene [82], polycyclic aromatic hydrocarbons such as benz(a)anthracene [83], organophosphorus pesticide such as trichlorfon [84], microplastics as bisphenol A [85], anti-inflammatory as ibuprofen [86], and antibiotics as sulfadiazine [87]. Genetically modified C. reinhardtii expressing the cyanase gene from Synechococcus elongatus showed the ability to remediate high levels of potassium cyanide, up to 150 mg/L [88]. These findings underscore the ability of Chlamydomonas to eliminate different substances, highlighting their role in environmental processes and potential applications in bioremediation.
6. Chlamydomonas-Bacterial Consortia for Bioremediation
The use of microalgae-bacteria consortia in bioremediation is known as a promising approach for wastewater treatment [89]. These consortia utilize the synergistic relationship between microalgae and bacteria to efficiently degrade organic matter, remove inorganic compounds, enhance biomass production, or improve influent quality, among other benefits, [90]. In fact, the use of microalgae-bacteria consortia can provide several advantages over the use of just microalgae monocultures for the bioremediation treatment of different wastewater such as improve the nutrient or antibiotics removal as well as to reduce the contamination risk. The combined metabolic activities of microalgae and bacteria reduce the risk of contamination compared to microalgae monocultures, as the diverse microbial community is more resilient to environmental changes and potential invaders [91]. A key interaction between microalgae and bacteria involves the exchange of CO2 and O2. Aerobic bacteria consume the oxygen generated by algal photosynthesis and, in turn, produce CO2 which supports algal growth. The bacteria can highly contribute to break down complex organic matter, making it more readily available for microalgal uptake [92]. Microalgae-bacteria consortia have shown superior performance in removing veterinary antibiotics from synthetic wastewater and swine wastewater in pilot-scale photobioreactors [93].
While there are numerous studies supporting the use of microalgae-bacteria consortia in different wastewater bioremediation [94], the use of Chlamydomonas-bacteria based consortia is quite unexplored topic with promising results. Different types of bacteria have shown the ability to enhance the potential of Chlamydomonas to phycoremediate. The potential of C. reinhardtii in consortia with three different bacterial strains (Stenotrophomonas maltophilia, Microbacterium paraoxydans, and Paenibacillus lactis) for bioremediating wastewater contaminated with phenol has been investigated. The results demonstrate that the consortium of M. paraoxydans and C. reinhardtii was very effective in phenol removal due to the synergistic interactions between the microalgae and bacteria, which enhance algal growth [95]. Furthermore, a cooperative consortium between C. reinhardtii and the bacterium Methylobacterium oryzae was found to lead an increased biomass generation and inorganic nitrogen removal when grown in urban wastewater [96]. The collaboration between microalgae and bacteria in wastewater treatment significantly enhances the efficiency of nitrogen removal. Bacteria play crucial roles in the nitrification and denitrification processes essential for completely eliminating nitrogen as nitrogen gas, and both algae and bacteria also contribute to nitrogen removal through assimilation [97]. As mentioned earlier, under illumination, microalgae produce oxygen, which aids in bacterial nitrification while reducing the energy required for culture aeration. C. reinhardtii in consortia with bacteria has been shown to remove nitrogen species from municipal wastewater, achieving up to 80% removal of dissolved organic nitrogen (DON) [98]. In a study using animal wastewater collected from two different sources, an animal feedlot wastewater storage tank and a sheep wastewater storage lagoon, the results showed that between 36-79% of initial DON was eliminated by the Chlamydomonas-bacteria consortia [99]. Acidithiobacillus ferrooxidans and Acidithiobacillus thiooxidans in consortia with Chlamydomonas have been shown to reduce the contamination of sediments containing heavy metals (Cu, Pb, Zn, Mn, Cd, As) [100]. The co-culture of C. reinhardtii with different bacterial strains has allowed the identification of bacterial species such as Sphingobium yanoikuyae, capable of degrading hydrocarbons and aromatic compounds [101], Acidovorax sp. A16OP12, which can utilize common environmental pollutants as a carbon source [102], and Microbacterium sp., capable of removing heavy metals and antibiotics [103].
7. Chlamydomonas-Bacterial Consortia for Biomass and Bio-Product Generation
7.1. Biomass
Microalgae biomass is highly valued in a bio-based economy, being utilized for the sustainable production of various products such as fuel, food, energy, pharmaceuticals, among others [104]. Microalgae have various advantages in biomass production; they are more productive per unit of land area than any plant system, do not compete for arable lands, and can be cultivated throughout the year [105]. Microalgae biomass is a versatile resource that offers a wide range of applications through the extraction of specific compounds for various purposes. The composition of biomass, including proteins, lipids, and carbohydrates, as well as active compounds, is significantly influenced by the strains of microalgae and the cultivation conditions they are exposed to [106]. Various approaches have been explored to obtain microalgae biomass enriched in specific biomolecules. Studies on increasing biomass production through the co-cultivation of Chlamydomonas and bacteria have shown promising results. Next, we will present studies in which an increase in biomass production is achieved through the interaction of bacteria and Chlamydomonas. The Chlamydomonas vitamin B12 auxotroph strain was able to increase its biomass when co-cultured with B12-producing bacteria, including the rhizobium Mesorhizobium loti or even an E. coli strain engineered to produce and release B12 [107]. Methylobacterium spp. have excellent biotechnological potential in agriculture because they produce phytohormones, promote plant growth through N2 fixation, and provide protection against pathogens and pollutants [108]. The cocultivation of Mesorhizobium oryzae and C. reinhardtii in ethanol-containing media significantly increases biomass production, with a potential increase of up to 700%. The crucial metabolic aspect of this association depended on the bacterial conversion of ethanol into acetate, which supported C. reinhardtii heterotrophic growth [96]. Methylobacterium aquaticum has been shown to enhance the C. reinhardtii biomass production in a proline-rich medium by transferring a portion of the NH4+ obtained through its metabolization to the alga. In turn, C. reinhardtii donates part of its fixed carbon in the form of glycerol to M. aquaticum, which uses it as its carbon source [109].
Diazotrophs are microorganisms that have the capability to convert atmospheric nitrogen into bioavailable forms like ammonia. The microalgae, including Chlamydomonas, have shown notable interactions with various types of nitrogen-fixing organisms [110]. Azotobacter spp., which are free-living symbiotic diazotrophs, have been widely employed as biofertilizers, effectively boosting the yields of various crop plants [111]. Different Azotobacter species, including A. chroococcum, A. beljerinckii, A. agilis, and A. vinelandii, have been demonstrated to enhance C. reinhardtii biomass production through the transfer of a portion of the fixed nitrogen to Chlamydomonas [112]. On the other hand, researchers have also observed that C. reinhardtii can support Azotobacter growth by transferring some of the carbon fixed through photosynthesis; however, the specific compounds involved in this process remain unidentified [113]. This Chlamydomonas-Azotobacter mutualism could have great biotechnological importance, as it would allow for the production of a large amount of biomass using only N2 and CO2 from the air, without the need to add organic nitrogen and carbon sources, which would decrease the economic profitability of the process.
7.2. Biofuels
Biofuels originate from renewable biological sources such as plants or plant-derived materials and include biodiesel, bioethanol, biogas, and biohydrogen. First-generation biofuels are made from food crops, second-generation biofuels come from non-food sources like waste and third-generation biofuels are produced from sources that do not compete with arable land. Biofuels produced from algae are third-generation biofuels [114]. These biofuels can be produced through transesterification, photosynthesis-mediated microbial fuel production, and other thermochemical and biochemical conversions [115]. Several approaches have been undertaken to increase biofuel production through the co-cultivation of microalgae and bacteria [116], with studies mainly focusing on biohydrogen production in Chlamydomonas.
7.2.1. Biohydrogen
Biohydrogen is the term used to refer to hydrogen produced by living organisms such as bacteria, cyanobacteria, and algae [117]. The first indications suggesting that Chlamydomonas had the capability to produce H2 were noted in experiments involving C. moewusii under anaerobic conditions [118]. In biological processes, hydrogenases are the enzymes responsible for hydrogen production [119]. Chlamydomonas has two distinct hydrogenases that have been the subject of extensive research, with the primary goal of exploring ways to enhance the efficiency of hydrogen production [120]. Hydrogenases can utilize two processes to obtain the energy for the reduction of protons into H2: they can either use the energy from light, a process known as biophotolysis, or oxidize organic compounds, such as starch, a process known as dark fermentation. The primary challenge in utilizing C. reinhardtii for hydrogen production is the rapid inactivation of both hydrogenases by oxygen, especially given that oxygen is produced during photosynthesis. Therefore, with C. reinhardtii, the first evidence of its capacity to produce H2 was reported under anaerobic conditions [121]. The use of sulfur-deprived C. reinhardtii was the first successful strategy demonstrating significant and consistent hydrogen production under aerobic conditions [122]. This occurs because the lack of sulfur inhibits protein synthesis, consequently disrupting photosynthesis and the generation of oxygen.
Chlamydomonas-bacteria consortia have opened a new window to improve hydrogen production. Several studies have successfully employed different consortia between C. reinhardtii and various bacterial partners, and analyzed the factors behind the improved hydrogen production [123]. One of the main reasons why the co-cultivation of Chlamydomonas and bacteria enhances hydrogen production is because the bacteria consume oxygen, thus preventing the inhibition of hydrogenase. This has been demonstrated in the co-cultivation of Chlamydomonas with A. chroococcum [124] or with Bradyrhizobium japonicum [125]. In the co-culture of Chlamydomonas with P. putida, P. stutzeri, Rhizobium etli, and E. coli, an increase in Chlamydomonas hydrogen production was also observed. This enhancement was attributed not only to the bacterial consumption of oxygen but also to the bacterial production of acetic acid from sugars, which Chlamydomonas utilizes as a carbon source [126,127]. In the coculture of C. reinhardtii and Methylobacterium oryzae, the improved hydrogen capacity of Chlamydomonas was due to the ability of M. oryzae to oxidize ethanol to acetate , which supports the heterotrophic growth of Chlamydomonas [96]. In the co-cultivation of C. reinhardtii and Mesorhizobium sangaii, the addition of various nitrogen compounds, such as NaNO2, NaNO3, and NH4Cl, led to enhanced photobiological hydrogen generation compared to monocultures. Specifically, the addition of 3 g/L NaNO2 resulted in the maximum H2 production of 226.98 μmol/mg chlorophyll, which was 5.2 times higher than that of the pure algal culture [128].
A very promising approach of Chlamydomonas-bacterial consortia is to achieve hydrogen production from wastewater (Figure 2). Although producing hydrogen from wastewater is very challenging, research on hydrogen production from waste degradation presents an intriguing avenue with significant scale-up potential. Considering that Chlamydomonas serves as a model microalga for biohydrogen production, and bacteria contribute to enhancing hydrogen generation, it is logical to explore the capacity of Chlamydomonas-bacteria consortia for biohydrogen production alongside wastewater bioremediation [129]. In fact, the biohydrogen production by three microalgae, including C. reinhardtii, has led to a novel approach for autotrophic denitrification using hydrogen-consuming denitrifiers [130]. Nitrate contamination in drinking water is a significant environmental concern. Autotrophic denitrification, which converts nitrate into harmless gases using microorganisms, is a promising alternative but an electron donor is needed to sustain this process. In this sense, the hydrogen produced by Chlamydomonas, in co-culture with hydrogen-consuming denitrifiers, serves as an electron donor, providing a sustainable, innovative, and eco-friendly solution for addressing nitrate contamination [130]. Mixed culture of Chlamydomonas-Rhizobium consortia has also been shown for coupled hydrogen and biogas production [131]. After the hydrogen production stage, the Chlamydomonas-Rhizobium biomass was used for biogas generation in the second stage of the process. This study demonstrates that the Chlamydomonas-Rhizobium consortium can produce hydrogen without the need for sulphur deprivation, which is typically required for biohydrogen production by Chlamydomonas monoculture.
7.2.2. Lipids
Triacylglycerols (TAG) are crucial lipids in microalgae for biofuel production. Oleaginous microalgae, rich in TAG, can be converted into biodiesel through transesterification, a process that transforms TAG into fatty acid methyl esters (FAME), the key components of biodiesel [105]. Given that biodiesel production is closely linked to the quantity of lipids and TAGs, various strategies have been explored to enhance their production in Chlamydomonas-bacterial consortia. High lipid content was found when a consortium of Chlamydomonas and cyanobacteria was cultured in wastewater from a dairy farm. The consortium was found to be capable of removing more than 98% of nutrients from wastewater, reaching a high biomass production and algal lipid content which could be converted into biodiesel [132]. It has been observed that immobilizing a co-culture of C. reinhardtii with the acetate-producing cyanobacteria Synechococcus sp. PCC 7002 increases the microalgal lipid yield [67]. The addition of A. chroococcum to C. reinhardtii culture significantly boosted lipid accumulation and productivity compared to the traditional nitrogen deprivation condition for increasing lipid accumulation, making it an efficient and economical strategy for enhancing lipid production [133]. Co-culturing Chlamydomonas sp. in the presence of the floc-forming bacterium Bacillus infantis showed that the co-cultures had higher microalgae biomass and lipid content compared to the axenic culture [134]. The co-culture of C. reinhardtii in municipal and swine wastewater effluents exhibited enhanced algal growth, biomass production, and lipid accumulation when indigenous bacteria were present [135].
7.3. Biofertilizers
Microalgae serve as biofertilizers and biostimulants, enhancing crop growth and enriching soil nutrients, which in turn reduces the need for chemical fertilizers and supports sustainable agriculture [136]. However, despite their abundance in natural soil ecosystems, Chlamydomonas species have been largely overlooked and underutilized in agricultural practices. In this context, lyophilized powders obtained from C. reinhardtii have been found to have a beneficial impact on maize plant growth, producing bioactive substances that act as biostimulants, improving plant growth, yield, quality, and crop performance [137]. It has been shown that Chlamydomonas sajao can enhance soil physical characteristics, such as aggregation and stability, thereby aiding in the improvement of soil structure and the retention of nutrients [138]. Research investigating the impact of Chlamydomonas applanata M9V as a biofertilizer on wheat growth revealed that both live and dead forms of the microalgae performed even better than a specific quantity of chemical fertilizer in terms of fresh weight, plant height, carotenoid and chlorophyll content [139]. Applying live Chlamydomonas cells through the soil drench method significantly enhanced leaf size, fresh weight, shoot length, pigment content, and the number of flowers in Medicago truncatula [140]. The acid-hydrolyzed dry biomass of C. reinhardtii enhanced the nitrogen, phosphorus, and carotenoid levels in Solanum lycopersicum [141]. Extracts from the biomass of Chlamydomonas sp. demonstrated auxin-like activity, leading to an increased number of roots in cucumber plants [142].
The significance of microalgae in the plant microbiome has only begun to be recognized recently, and their application for enhancing soil fertility, conserving water, and promoting plant growth is now emerging as a promising strategy for sustainable agriculture [143]. The plant hormone IAA, known as auxin, is a key signaling molecule involved in regulating plant growth and physiology [144]. Microalgae can release L-tryptophan, which can be converted by bacteria into IAA to stimulate algal growth [145]. In this regard, Chlamydomonas has the ability to release L-tryptophan, which serves as a suitable substrate for symbiotic microbes to produce IAA [146]. Moreover, it has been shown that Chlamydomonas itself can also synthesize IAA from L-tryptophan, an activity mediated by the enzyme L-amino acid oxidase (LAO1) [147]. Interestingly, elevated levels of IAA inhibit Chlamydomonas cell proliferation and chlorophyll degradation. However, in a consortium of C. reinhardtii with the plant-growth-promoting bacterium Methylobacterium aquaticum, these inhibitory effects were alleviated. These results may have significant agricultural implications, as Chlamydomonas and Methylobacterium spp. coexist in the plant rhizosphere [148]. Their ability to regulate IAA levels could influence plant health and may be utilized for enhancing crops in sustainable agriculture practices.
8. Future Perspective
This review aims to highlight the great potential of Chlamydomonas in monoculture and in co-culture with bacterial partners across various aspects, including the bioremediation of wastewater, biomass production, biofuels, and biofertilizers. As mentioned, one of the main products derived from the cultivation of microalgae-bacteria is their biomass, which is used as a raw material for obtaining other derived bioproducts. It would be highly beneficial economically to use the biomass resulting from the bioremediation process for bioproduct purification (Figure 2). However, the utilization of biomass and the water derived from wastewater treatment faces several inherent challenges. These challenges include the presence of xenobiotic residues and heavy metals within the biomass, the scalability of biomass production, as well as contamination with bacteria, fungi, and viruses, all of which limit their extensive application. Achieving future improvement and optimization of these processes to avoid these drawbacks is one of the most promising future perspectives, which would allow establishing a relationship with the circular economy by reducing waste and reusing materials, thus minimizing environmental and economic impact.
Author Contributions
AL original idea, conceptualization, and preparation of the first draft; AL, C B-P and MJ T-P wrote the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by Gobierno de España, Ministerio de Ciencia e Innovacion (Grant PID2020-118398GB-I00), Junta de Andalucía (Grant ProyExcel_00483), the “Plan Propio” from University of Cordoba, and a grant awarded by the Torres-Gutierrez foundation.
Data Availability Statement
All data required to evaluate the conclusions of this paper are included in the main text.
Acknowledgments
This paper is dedicated to Emilio Fernandez Reyes, who has recently retired after almost 40 years of studying Chlamydomonas reinhardtii as a reference organism. He was the driving force that promoted our research on Chlamydomonas, the pillar that allowed its advancement, and our great teacher whom we will never be able to repay for all the learnings received. We also thank Maribel Macias for her constant technical support.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Rani, S.; Gunjyal, N.; Ojha, C.S.P.; Singh, R.P. Review of Challenges for Algae-Based Wastewater Treatment: Strain Selection, Wastewater Characteristics, Abiotic, and Biotic Factors. J. Hazardous, Toxic, Radioact. Waste 2021, 25, 03120004. [Google Scholar] [CrossRef]
- Görgényi, J.; T-Krasznai, E.; Lukács, Á.; Kókai, Z.; B-Béres, V.; Várbíró, G.; Ács, É.; Kiss, K.T.; Tóthmérész, B.; Borics, G. Functional Properties of Planktic Microalgae Determine Their Habitat Selection. Hydrobiologia 2024, 851, 801–821. [Google Scholar] [CrossRef]
- Ochoa de Alda, J.A.G.; Esteban, R.; Diago, M.L.; Houmard, J. The Plastid Ancestor Originated among One of the Major Cyanobacterial Lineages. Nat. Commun. 2014, 5, 4937. [Google Scholar] [CrossRef] [PubMed]
- Kazmi, S.S.U.H.; Yapa, N.; Karunarathna, S.C.; Suwannarach, N. Perceived Intensification in Harmful Algal Blooms Is a Wave of Cumulative Threat to the Aquatic Ecosystems. Biology (Basel). 2022, 11, 852. [Google Scholar] [CrossRef] [PubMed]
- Gerotto, C.; Norici, A.; Giordano, M. Toward Enhanced Fixation of CO2 in Aquatic Biomass: Focus on Microalgae. Front. Energy Res. 2020, 8, 213. [Google Scholar] [CrossRef]
- Olguín, E.J.; Sánchez-Galván, G.; Arias-Olguín, I.I.; Melo, F.J.; González-Portela, R.E.; Cruz, L.; De Philippis, R.; Adessi, A. Microalgae-Based Biorefineries: Challenges and Future Trends to Produce Carbohydrate Enriched Biomass, High-Added Value Products and Bioactive Compounds. Biology (Basel). 2022, 11, 1146. [Google Scholar] [CrossRef] [PubMed]
- Mavrommatis, A.; Tsiplakou, E.; Zerva, A.; Pantiora, P.D.; Georgakis, N.D.; Tsintzou, G.P.; Madesis, P.; Labrou, N.E. Microalgae as a Sustainable Source of Antioxidants in Animal Nutrition, Health and Livestock Development. Antioxidants 2023, 12, 1882. [Google Scholar] [CrossRef]
- Satya, A.D.M.; Cheah, W.Y.; Yazdi, S.K.; Cheng, Y.S.; Khoo, K.S.; Vo, D.V.N.; Bui, X.D.; Vithanage, M.; Show, P.L. Progress on Microalgae Cultivation in Wastewater for Bioremediation and Circular Bioeconomy. Environ. Res. 2023, 218, 114948. [Google Scholar] [CrossRef]
- Sasso, S.; Stibor, H.; Mittag, M.; Grossman, A.R. From Molecular Manipulation of Domesticated Chlamydomonas Reinhardtii to Survival in Nature. Elife 2018, 7, e39233. [Google Scholar] [CrossRef]
- Merchant, S.S.; Prochnik, S.E.; Vallon, O.; Harris, E.H.; Karpowicz, S.J.; Witman, G.B.; Terry, A.; Salamov, A.; Fritz-Laylin, L.K.; Maréchal-Drouard, L.; et al. The Chlamydomonas Genome Reveals the Evolution of Key Animal and Plant Functions. Science 2007, 318, 245–250. [Google Scholar] [CrossRef]
- Kelterborn, S.; Boehning, F.; Sizova, I.; Baidukova, O.; Evers, H.; Hegemann, P. Gene Editing in Green Alga Chlamydomonas Reinhardtii via CRISPR-Cas9 Ribonucleoproteins. Plant Synth. Biol. Methods Mol. Biol. 2022, 2379, 45–65. [Google Scholar] [CrossRef]
- Schroda, M.; Remacle, C. Molecular Advancements Establishing Chlamydomonas as a Host for Biotechnological Exploitation. Front. Plant Sci. 2022, 13, 911483. [Google Scholar] [CrossRef] [PubMed]
- Ramanan, R.; Kim, B.-H.; Cho, D.-H.; Oh, H.-M.; Kim, H.-S. Algae-Bacteria Interactions: Evolution, Ecology and Emerging Applications. Biotechnol. Adv. 2016, 34, 14–29. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.; Nayak, M.; Ghosh, A. A Review on Co-Culturing of Microalgae: A Greener Strategy towards Sustainable Biofuels Production. Sci. Total Environ. 2022, 802, 149765. [Google Scholar] [CrossRef] [PubMed]
- Bell, W.; Mitchell, R. Chemotactic and Growth Responses of Marine Bacteria to Algal Extracellular Products. Biol. Bull. 1972, 143, 265–277. [Google Scholar] [CrossRef]
- Hasnain, M.; Zainab, R.; Ali, F.; Abideen, Z.; Yong, J.W.H.; El-Keblawy, A.; Hashmi, S.; Radicetti, E. Utilization of Microalgal-Bacterial Energy Nexus Improves CO2 Sequestration and Remediation of Wastewater Pollutants for Beneficial Environmental Services. Ecotoxicol. Environ. Saf. 2023, 267, 115646. [Google Scholar] [CrossRef] [PubMed]
- Venkataram, S.; Kuo, H.-Y.; Hom, E.F.Y.; Kryazhimskiy, S. Mutualism-Enhancing Mutations Dominate Early Adaptation in a Two-Species Microbial Community. Nat. Ecol. Evol. 2023, 7, 143–154. [Google Scholar] [CrossRef]
- Calatrava, V.; Tejada-Jimenez, M.; Sanz-Luque, E.; Fernandez, E.; Galvan, A.; Llamas, A. Chlamydomonas Reinhardtii, a Reference Organism to Study Algal–Microbial Interactions: Why Can’t They Be Friends? Plants 2023, 12, 788. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, Q.; Zhou, R.; Feng, J.; Zhang, K.; Li, X.; Ma, X.; Dietrich, A.M. Occurrence of Free Amino Acids in the Source Waters of Zhejiang Province, China, and Their Removal and Transformation in Drinking Water Systems. Water (Switzerland) 2020, 12, 73. [Google Scholar] [CrossRef]
- Gaur, N.; Dutta, D.; Singh, A.; Dubey, R.; Kamboj, D.V. Recent Advances in the Elimination of Persistent Organic Pollutants by Photocatalysis. Front. Environ. Sci. 2022, 10, 872514. [Google Scholar] [CrossRef]
- Ahamed, A.; Ge, L.; Zhao, K.; Veksha, A.; Bobacka, J.; Lisak, G. Environmental Footprint of Voltammetric Sensors Based on Screen-Printed Electrodes: An Assessment towards “Green” Sensor Manufacturing. Chemosphere 2021, 278, 130462. [Google Scholar] [CrossRef] [PubMed]
- Kargol, A.K.; Burrell, S.R.; Chakraborty, I.; Gough, H.L. Synthetic Wastewater Prepared from Readily Available Materials: Characteristics and Economics. PLOS Water 2023, 2, e0000178. [Google Scholar] [CrossRef]
- Ali, I.; Naz, I.; Peng, C.; Abd-Elsalam, K.A.; Khan, Z.M.; Islam, T.; Pervez, R.; Amjed, M.A.; Tehrim, A.; Perveen, I.; et al. Sources, Classifications, Constituents, and Available Treatment Technologies for Various Types of Wastewater: An Overview. Aquananotechnology 2021, 11–46. [Google Scholar] [CrossRef]
- Krishnan, R.Y.; Manikandan, S.; Subbaiya, R.; Biruntha, M.; Govarthanan, M.; Karmegam, N. Removal of Emerging Micropollutants Originating from Pharmaceuticals and Personal Care Products (PPCPs) in Water and Wastewater by Advanced Oxidation Processes: A Review. Environ. Technol. Innov. 2021, 23, 101757. [Google Scholar] [CrossRef]
- Marlina, E. ; Purwanto Electro-Fenton for Industrial Wastewater Treatment: A Review. E3S Web Conf. 2019, 125, 03003. [Google Scholar]
- Talukdar, A.; Kundu, P.; Bhattacharya, S.; Dutta, N. Microplastic Contamination in Wastewater: Sources, Distribution, Detection and Remediation through Physical and Chemical-Biological Methods. Sci. Total Environ. 2024, 916, 170254. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.A. Wastewater Treatment and Reuse for Sustainable Water Resources Management: A Systematic Literature Review. Sustainability 2023, 15, 10940. [Google Scholar] [CrossRef]
- Yadav, G.; Shanmugam, S.; Sivaramakrishnan, R.; Kumar, D.; Mathimani, T.; Brindhadevi, K.; Pugazhendhi, A.; Rajendran, K. Mechanism and Challenges behind Algae as a Wastewater Treatment Choice for Bioenergy Production and Beyond. Fuel 2021, 285, 119093. [Google Scholar] [CrossRef]
- Razaviarani, V.; Arab, G.; Lerdwanawattana, N.; Gadia, Y. Algal Biomass Dual Roles in Phycoremediation of Wastewater and Production of Bioenergy and Value-Added Products. Int. J. Environ. Sci. Technol. 2023, 20, 8199–8216. [Google Scholar] [CrossRef]
- Dayana Priyadharshini, S.; Suresh Babu, P.; Manikandan, S.; Subbaiya, R.; Govarthanan, M.; Karmegam, N. Phycoremediation of Wastewater for Pollutant Removal: A Green Approach to Environmental Protection and Long-Term Remediation. Environ. Pollut. 2021, 290, 117989. [Google Scholar] [CrossRef]
- Barboza-Rodríguez, R.; Rodríguez-Jasso, R.M.; Rosero-Chasoy, G.; Rosales Aguado, M.L.; Ruiz, H.A. Photobioreactor Configurations in Cultivating Microalgae Biomass for Biorefinery. Bioresour. Technol. 2024, 394, 130208. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, F.G.G.; Alonso Gómez, V.; Muñoz Torre, R.; de Godos Crespo, I. Scale-down of High-Rate Algae Ponds Systems for Urban Wastewater Reuse. J. Water Process Eng. 2023, 56, 104342. [Google Scholar] [CrossRef]
- Jebali, A.; Acién, F.G.; Rodriguez Barradas, E.; Olguín, E.J.; Sayadi, S.; Molina Grima, E. Pilot-Scale Outdoor Production of Scenedesmus Sp. in Raceways Using Flue Gases and Centrate from Anaerobic Digestion as the Sole Culture Medium. Bioresour. Technol. 2018, 262, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Keeley, R.; Zalivina, N.; Halfhide, T.; Scott, K.; Zhang, Q.; van der Steen, P.; Ergas, S.J. Advances in Algal-Prokaryotic Wastewater Treatment: A Review of Nitrogen Transformations, Reactor Configurations and Molecular Tools. J. Environ. Manage. 2018, 217, 845–857. [Google Scholar] [CrossRef] [PubMed]
- de Godos, I.; Arbib, Z.; Lara, E.; Rogalla, F. Evaluation of High Rate Algae Ponds for Treatment of Anaerobically Digested Wastewater: Effect of CO2 Addition and Modification of Dilution Rate. Bioresour. Technol. 2016, 220, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Leong, Y.K.; Huang, C.Y.; Chang, J.S. Pollution Prevention and Waste Phycoremediation by Algal-Based Wastewater Treatment Technologies: The Applications of High-Rate Algal Ponds (HRAPs) and Algal Turf Scrubber (ATS). J. Environ. Manage. 2021, 296, 113193. [Google Scholar] [CrossRef] [PubMed]
- González-Fernández, C.; Molinuevo-Salces, B.; García-González, M.C. Nitrogen Transformations under Different Conditions in Open Ponds by Means of Microalgae-Bacteria Consortium Treating Pig Slurry. Bioresour. Technol. 2011, 102, 960–966. [Google Scholar] [CrossRef] [PubMed]
- Grönlund, E.; Hanaeus, J.; Johansson, E.; Falk, S. Performance of an Experimental Wastewater Treatment High-Rate Algal Pond in Subarctic Climate. Water Environ. Res. a Res. Publ. Water Environ. Fed. 2010, 82, 830–839. [Google Scholar] [CrossRef] [PubMed]
- Godos, I. de; Blanco, S.; García-Encina, P.A.; Becares, E.; Muñoz, R. Long-Term Operation of High Rate Algal Ponds for the Bioremediation of Piggery Wastewaters at High Loading Rates. Bioresour. Technol. 2009, 100, 4332–4339. [Google Scholar] [CrossRef]
- Bohutskyi, P.; Phan, D.; Spierling, R.E.; Kopachevsky, A.M.; Bouwer, E.J.; Lundquist, T.J.; Betenbaugh, M.J. Production of Lipid-Containing Algal-Bacterial Polyculture in Wastewater and Biomethanation of Lipid Extracted Residues: Enhancing Methane Yield through Hydrothermal Pretreatment and Relieving Solvent Toxicity through Co-Digestion. Sci. Total Environ. 2019, 653, 1377–1394. [Google Scholar] [CrossRef]
- Villalba, M.R.; Cervera, R.; Sánchez, J. Green Solutions for Urban Sustainability: Photobioreactors for Algae Cultivation on Façades and Artificial Trees. Buildings 2023, 13, 1541. [Google Scholar] [CrossRef]
- Ugwu, C.U.; Aoyagi, H.; Uchiyama, H. Photobioreactors for Mass Cultivation of Algae. Bioresour. Technol. 2008, 99, 4021–4028. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Carvajal, G.D.; Taidi, B.; Jarrahi, M. Towards a Low Energy, Stirless Photobioreactor Using Photosynthetic Motile Microalgae. Algal Res. 2024, 77, 103350. [Google Scholar] [CrossRef]
- Rodríguez-Bolaños, M.; Vargas-Romero, G.; Jaguer-García, G.; Aguilar-Gonzalez, Z.I.; Lagos-Romero, V.; Miranda-Astudillo, H. V. Antares I: A Modular Photobioreactor Suitable for Photosynthesis and Bioenergetics Research. Appl. Biochem. Biotechnol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Molina, E.; Fernández, J.; Acién, F.G.; Chisti, Y. Tubular Photobioreactor Design for Algal Cultures. J. Biotechnol. 2001, 92, 113–131. [Google Scholar] [CrossRef]
- Bilanovic, D.; Holland, M.; Starosvetsky, J.; Armon, R. Co-Cultivation of Microalgae and Nitrifiers for Higher Biomass Production and Better Carbon Capture. Bioresour. Technol. 2016, 220, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Mennerich, A.; Urban, B. Municipal Wastewater Treatment and Biomass Accumulation with a Wastewater-Born and Settleable Algal-Bacterial Culture. Water Res. 2011, 45, 3351–3358. [Google Scholar] [CrossRef] [PubMed]
- Moreno Osorio, J.H.; Pollio, A.; Frunzo, L.; Lens, P.N.L.; Esposito, G. A Review of Microalgal Biofilm Technologies: Definition, Applications, Settings and Analysis. Front. Chem. Eng. 2021, 3, 737710. [Google Scholar] [CrossRef]
- Kesaano, M.; Sims, R.C. Algal Biofilm Based Technology for Wastewater Treatment. Algal Res. 2014, 5, 231–240. [Google Scholar] [CrossRef]
- Kreis, C.T.; Grangier, A.; Bäumchen, O. In Vivo Adhesion Force Measurements of Chlamydomonas on Model Substrates. Soft Matter 2019, 15, 3027–3035. [Google Scholar] [CrossRef]
- Catalan, R.E.; Fragkopoulos, A.A.; von Trott, N.; Kelterborn, S.; Baidukova, O.; Hegemann, P.; Bäumchen, O. Light-Regulated Adsorption and Desorption of Chlamydomonas Cells at Surfaces. Soft Matter 2023, 19, 306. [Google Scholar] [CrossRef] [PubMed]
- Schaedig, E.; Cantrell, M.; Urban, C.; Zhao, X.; Greene, D.; Dancer, J.; Gross, M.; Sebesta, J.; Chou, K.J.; Grabowy, J.; et al. Isolation of Phosphorus-Hyperaccumulating Microalgae from Revolving Algal Biofilm (RAB) Wastewater Treatment Systems. Front. Microbiol. 2023, 14, 1219318. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Yu, T.; Xie, Y.; Chen, J.; Ho, S.H.; Wang, Y.; Huang, F. Attached Culture of Chlamydomonas Sp. JSC4 for Biofilm Production and TN/TP/Cu(II) Removal. Biochem. Eng. J. 2019, 141, 1–9. [Google Scholar] [CrossRef]
- Posadas, E.; García-Encina, P.A.; Soltau, A.; Domínguez, A.; Díaz, I.; Muñoz, R. Carbon and Nutrient Removal from Centrates and Domestic Wastewater Using Algal-Bacterial Biofilm Bioreactors. Bioresour. Technol. 2013, 139, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, L.; Wang, J.; Chen, Y.; Gao, X.; Zhang, Z.; Liu, T. Attached Cultivation for Improving the Biomass Productivity of Spirulina Platensis. Bioresour. Technol. 2015, 181, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Yang, Z.H.; Li, C.; Zeng, G.M.; Ma, D.H.; Zhou, L. A Novel Algal Biofilm Membrane Photobioreactor for Attached Microalgae Growth and Nutrients Removal from Secondary Effluent. Bioresour. Technol. 2015, 179, 8–12. [Google Scholar] [CrossRef]
- Vieira, M. V.; Pastrana, L.M.; Fuciños, P. Microalgae Encapsulation Systems for Food, Pharmaceutical and Cosmetics Applications. Mar. Drugs 2020, 18, 644. [Google Scholar] [CrossRef]
- Han, M.; Zhang, C.; Ho, S.H. Immobilized Microalgal System: An Achievable Idea for Upgrading Current Microalgal Wastewater Treatment. Environ. Sci. Ecotechnology 2023, 14, 100227. [Google Scholar] [CrossRef]
- Han, M.; Xie, P.; Ren, N.; Ho, S.H. Cytoprotective Alginate Microcapsule Serves as a Shield for Microalgal Encapsulation Defensing Sulfamethoxazole Threats and Safeguarding Nutrient Recovery. J. Hazard. Mater. 2024, 465, 133454. [Google Scholar] [CrossRef]
- Weng, Y.; Yang, G.; Li, Y.; Xu, L.; Chen, X.; Song, H.; Zhao, C.X. Alginate-Based Materials for Enzyme Encapsulation. Adv. Colloid Interface Sci. 2023, 318, 102957. [Google Scholar] [CrossRef]
- de-Bashan, L.E.; Bashan, Y. Immobilized Microalgae for Removing Pollutants: Review of Practical Aspects. Bioresour. Technol. 2010, 101, 1611–1627. [Google Scholar] [CrossRef]
- Nazos, T.T.; Ghanotakis, D.F. Biodegradation of Phenol by Alginate Immobilized Chlamydomonas Reinhardtii Cells. Arch. Microbiol. 2021, 203, 5805–5816. [Google Scholar] [CrossRef]
- Lee, H.; Jeong, D.; Im, S.J.; Jang, A. Optimization of Alginate Bead Size Immobilized with Chlorella Vulgaris and Chlamydomonas Reinhardtii for Nutrient Removal. Bioresour. Technol. 2020, 302, 122891. [Google Scholar] [CrossRef]
- Homburg, S.V.; Kruse, O.; Patel, A. V. Growth and Photosynthetic Activity of Chlamydomonas Reinhardtii Entrapped in Lens-Shaped Silica Hydrogels. J. Biotechnol. 2019, 302, 58–66. [Google Scholar] [CrossRef]
- Zhang, B.B.; Wang, L.; Charles, V.; Rooke, J.C.; Su, B.L. Robust and Biocompatible Hybrid Matrix with Controllable Permeability for Microalgae Encapsulation. ACS Appl. Mater. Interfaces 2016, 8, 8939–8946. [Google Scholar] [CrossRef]
- Mandsberg, N.K.; Liao, W.; Yamanouchi, Y.A.; Boisen, A.; Ejima, H. Encapsulation of Chlamydomonas Reinhardtii into a Metal-Phenolic Network. Algal Res. 2022, 61, 102569. [Google Scholar] [CrossRef]
- Therien, J.B.; Zadvornyy, O.A.; Posewitz, M.C.; Bryant, D.A.; Peters, J.W. Growth of Chlamydomonas Reinhardtii in Acetate-Free Medium When Co-Cultured with Alginate-Encapsulated, Acetate-Producing Strains of Synechococcus Sp. PCC 7002. Biotechnol. Biofuels 2014, 7, 154. [Google Scholar] [CrossRef]
- Kumar, R.; Goyal, D. Comparative Biosorption of Pb 2+ by Live Algal Consortium and Immobilized Dead Biomass from Aqueous Solution. Indian J. Exp. Biol. 2009, 46, 690–694. [Google Scholar]
- Choix, F.J.; Guadalupe López-Cisneros, C.; Oscar Méndez-Acosta, H. Azospirillum Brasilense Increases CO2 Fixation on Microalgae Scenedesmus Obliquus, Chlorella Vulgaris, and Chlamydomonas Reinhardtii Cultured on High CO2 Concentrations. Microb. Ecol. 2018, 76, 430–442. [Google Scholar] [CrossRef]
- Bhatia, S.K.; Mehariya, S.; Bhatia, R.K.; Kumar, M.; Pugazhendhi, A.; Awasthi, M.K.; Atabani, A.E.; Kumar, G.; Kim, W.; Seo, S.O.; et al. Wastewater Based Microalgal Biorefinery for Bioenergy Production: Progress and Challenges. Sci. Total Environ. 2021, 751, 141599. [Google Scholar] [CrossRef]
- Leong, Y.K.; Chang, J.S. Bioremediation of Heavy Metals Using Microalgae: Recent Advances and Mechanisms. Bioresour. Technol. 2020, 303, 122886. [Google Scholar] [CrossRef]
- Xi, Y.; Han, B.; Kong, F.; You, T.; Bi, R.; Zeng, X.; Wang, S.; Jia, Y. Enhancement of Arsenic Uptake and Accumulation in Green Microalga Chlamydomonas Reinhardtii through Heterologous Expression of the Phosphate Transporter DsPht1. J. Hazard. Mater. 2023, 459, 132130. [Google Scholar] [CrossRef]
- Saavedra, R.; Muñoz, R.; Taboada, M.E.; Vega, M.; Bolado, S. Comparative Uptake Study of Arsenic, Boron, Copper, Manganese and Zinc from Water by Different Green Microalgae. Bioresour. Technol. 2018, 263, 49–57. [Google Scholar] [CrossRef]
- Nam, S.H.; Kwak, J. Il; An, Y.J. Assessing Applicability of the Paper-Disc Method Used in Combination with Flow Cytometry to Evaluate Algal Toxicity. Environ. Pollut. 2018, 234, 979–987. [Google Scholar] [CrossRef]
- Baselga-Cervera, B.; García-Balboa, C.; Díaz-Alejo, H.M.; Costas, E.; López-Rodas, V. Rapid Colonization of Uranium Mining-Impacted Waters, the Biodiversity of Successful Lineages of Phytoplankton Extremophiles. Microb. Ecol. 2020, 79, 576–587. [Google Scholar] [CrossRef]
- Ibuot, A.; Webster, R.E.; Williams, L.E.; Pittman, J.K. Increased Metal Tolerance and Bioaccumulation of Zinc and Cadmium in Chlamydomonas Reinhardtii Expressing a AtHMA4 C-Terminal Domain Protein. Biotechnol. Bioeng. 2020, 117, 2996–3005. [Google Scholar] [CrossRef]
- Hoyos, B.S.; Hernandez-Tenorio, F.; Miranda, A.M.; Villanueva-Mejía, D.F.; Sáez, A.A. Systematic Analysis of Genes Related to Selenium Bioaccumulation in Microalgae: A Review. Biology (Basel). 2023, 12, 703. [Google Scholar] [CrossRef]
- Wei, S.; Cao, J.; Ma, X.; Ping, J.; Zhang, C.; Ke, T.; Zhang, Y.; Tao, Y.; Chen, L. The Simultaneous Removal of the Combined Pollutants of Hexavalent Chromium and O-Nitrophenol by Chlamydomonas Reinhardtii. Ecotoxicol. Environ. Saf. 2020, 198, 110648. [Google Scholar] [CrossRef]
- Jin, Z.P.; Luo, K.; Zhang, S.; Zheng, Q.; Yang, H. Bioaccumulation and Catabolism of Prometryne in Green Algae. Chemosphere 2012, 87, 278–284. [Google Scholar] [CrossRef]
- Xiong, J.Q.; Kurade, M.B.; Abou-Shanab, R.A.I.; Ji, M.K.; Choi, J.; Kim, J.O.; Jeon, B.H. Biodegradation of Carbamazepine Using Freshwater Microalgae Chlamydomonas Mexicana and Scenedesmus Obliquus and the Determination of Its Metabolic Fate. Bioresour. Technol. 2016, 205, 183–190. [Google Scholar] [CrossRef]
- Touliabah, H.E.S.; El-Sheekh, M.M.; Ismail, M.M.; El-Kassas, H. A Review of Microalgae-and Cyanobacteria-Based Biodegradation of Organic Pollutants. Molecules 2022, 27. [Google Scholar] [CrossRef]
- Li, S.; Wang, P.; Zhang, C.; Zhou, X.; Yin, Z.; Hu, T.; Hu, D.; Liu, C.; Zhu, L. Influence of Polystyrene Microplastics on the Growth, Photosynthetic Efficiency and Aggregation of Freshwater Microalgae Chlamydomonas Reinhardtii. Sci. Total Environ. 2020, 714, 136767. [Google Scholar] [CrossRef]
- Luo, J.; Deng, J.; Cui, L.; Chang, P.; Dai, X.; Yang, C.; Li, N.; Ren, Z.; Zhang, X. The Potential Assessment of Green Alga Chlamydomonas Reinhardtii CC-503 in the Biodegradation of Benz(a)Anthracene and the Related Mechanism Analysis. Chemosphere 2020, 249, 126097. [Google Scholar] [CrossRef]
- Wan, L.; Wu, Y.; Ding, H.; Zhang, W. Toxicity, Biodegradation, and Metabolic Fate of Organophosphorus Pesticide Trichlorfon on the Freshwater Algae Chlamydomonas Reinhardtii. J. Agric. Food Chem. 2020, 68, 1645–1653. [Google Scholar] [CrossRef]
- Carbó, M.; Chaturvedi, P.; Álvarez, A.; Pineda-Cevallos, D.; Ghatak, A.; González, P.R.; Cañal, M.J.; Weckwerth, W.; Valledor, L. Ferroptosis Is the Key Cellular Process Mediating Bisphenol A Responses in Chlamydomonas and a Promising Target for Enhancing Microalgae-Based Bioremediation. J. Hazard. Mater. 2023, 448, 130997. [Google Scholar] [CrossRef]
- Seoane, M.; Conde-Pérez, K.; Esperanza, M.; Cid, Á.; Rioboo, C. Unravelling Joint Cytotoxicity of Ibuprofen and Oxytetracycline on Chlamydomonas Reinhardtii Using a Programmed Cell Death-Related Biomarkers Panel. Aquat. Toxicol. 2023, 257, 106455. [Google Scholar] [CrossRef]
- Li, Z.; Dong, S.; Huang, F.; Lin, L.; Hu, Z.; Zheng, Y. Toxicological Effects of Microplastics and Sulfadiazine on the Microalgae Chlamydomonas Reinhardtii. Front. Microbiol. 2022, 13, 865768. [Google Scholar] [CrossRef]
- Sobieh, S.S.; Abed El-Gammal, R.; El-Kheir, W.S.A.; El-Sheimy, A.A.; Said, A.A.; El-Ayouty, Y.M. Heterologous Expression of Cyanobacterial Cyanase Gene (CYN) in Microalga Chlamydomonas Reinhardtii for Bioremediation of Cyanide Pollution. Biology (Basel). 2022, 11, 1420. [Google Scholar] [CrossRef]
- Dai, C.; Wang, F. Potential Applications of Microalgae–Bacteria Consortia in Wastewater Treatment and Biorefinery. Bioresour. Technol. 2024, 393, 130019. [Google Scholar] [CrossRef]
- Gururani, P.; Bhatnagar, P.; Kumar, V.; Vlaskin, M.S.; Grigorenko, A. V. Algal Consortiums: A Novel and Integrated Approach for Wastewater Treatment. Water (Switzerland) 2022, 14, 3784. [Google Scholar] [CrossRef]
- Chang, Y.L.; Nagarajan, D.; Chen, J.H.; Yen Chen, C.; Wu, Y.J.; Whang, L.M.; Lee, D.J.; Chang, J.S. Microalgae-Bacteria Consortia for the Treatment of Raw Dairy Manure Wastewater Using a Novel Two-Stage Process: Process Optimization and Bacterial Community Analysis. Chem. Eng. J. 2023, 473, 145388. [Google Scholar] [CrossRef]
- Zambrano, J.; García-Encina, P.A.; Jiménez, J.J.; Ciardi, M.; Bolado-Rodríguez, S.; Irusta-Mata, R. Removal of Veterinary Antibiotics in Swine Manure Wastewater Using Microalgae–Bacteria Consortia in a Pilot Scale Photobioreactor. Environ. Technol. Innov. 2023, 31, 103190. [Google Scholar] [CrossRef]
- Zambrano, J.; García-Encina, P.A.; Hernández, F.; Botero-Coy, A.M.; Jiménez, J.J.; Irusta-Mata, R. Kinetics of the Removal Mechanisms of Veterinary Antibiotics in Synthetic Wastewater Using Microalgae–Bacteria Consortia. Environ. Technol. Innov. 2023, 29, 103031. [Google Scholar] [CrossRef]
- Bhatt, P.; Bhandari, G.; Bhatt, K.; Simsek, H. Microalgae-Based Removal of Pollutants from Wastewaters: Occurrence, Toxicity and Circular Economy. Chemosphere 2022, 306, 135576. [Google Scholar] [CrossRef]
- Mora-Salguero, D.; Vives Florez, M.J.; Husserl Orjuela, J.; Fernández-Niño, M.; González Barrios, A.F. Evaluation of the Phenol Degradation Capacity of Microalgae-Bacteria Consortia from the Bay of Cartagena, Colombia. TecnoLógicas 2019, 22, 149–158. [Google Scholar] [CrossRef]
- Torres, M.; Gonzalez-Ballester, D.; Gomez-Osuna, A.; Galván, A.; Fernandez, E.; Dubini, A. Chlamydomonas-Methylobacterium Oryzae Cooperation Leads to Increased Biomass, Nitrogen Removal, and Hydrogen Production. Bioresour. Technol. 2022, 352, 127088. [Google Scholar] [CrossRef] [PubMed]
- Stein, L.Y.; Klotz, M.G. The Nitrogen Cycle. Curr Biol 2016, 26, R94–8. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Simsek, H. Bioavailability of Wastewater Derived Dissolved Organic Nitrogen to Green Microalgae Selenastrum Capricornutum, Chlamydomonas Reinhardtii, and Chlorella Vulgaris with/without Presence of Bacteria. J. Environ. Sci. 2017, 57, 346–355. [Google Scholar] [CrossRef]
- Sun, J.; Khan, E.; Simsek, S.; Ohm, J.-B.; Simsek, H. Bioavailability of Dissolved Organic Nitrogen (DON) in Wastewaters from Animal Feedlots and Storage Lagoons. Chemosphere 2017, 186, 695–701. [Google Scholar] [CrossRef]
- Yang, Q.; Jie, S.; Lei, P.; Gan, M.; He, P.; Zhu, J.; Zhou, Q. Effect of Anthropogenic Disturbances on the Microbial Relationship during Bioremediation of Heavy Metal-Contaminated Sediment. Microorganisms 2023, 11, 1185. [Google Scholar] [CrossRef]
- Mitra, M.; Nguyen, K.M.A.K.; Box, T.W.; Gilpin, J.S.; Hamby, S.R.; Berry, T.L.; Duckett, E.H. Isolation and Characterization of a Novel Sphingobium Strain Variant That Uses Biohazardous Saturated and Aromatic Compounds as Sole Carbon. F1000Research 2022, 9, 767. [Google Scholar] [CrossRef] [PubMed]
- Mitra, M.; Nguyen, K.M.A.K.; Box, T.W.; Gilpin, J.S.; Hamby, S.R.; Berry, T.L.; Duckett, E.H. Isolation and Characterization of a Novel Bacterial Strain from a Tris-Acetate-Phosphate Agar Medium Plate of the Green Micro-Alga Chlamydomonas Reinhardtii That Can Utilize Common Environmental Pollutants as a Carbon Source. F1000Research 2022, 9, 656. [Google Scholar] [CrossRef] [PubMed]
- Mitra, M.; Nguyen, K.M.-A.-K.; Box, T.W.; Berry, T.L.; Fujita, M. Isolation and Characterization of a Heavy Metal- and Antibiotic-Tolerant Novel Bacterial Strain from a Contaminated Culture Plate of Chlamydomonas Reinhardtii, a Green Micro-Alga. F1000Research 2023, 10, 533. [Google Scholar] [CrossRef]
- Siddiki, S.Y.A.; Mofijur, M.; Kumar, P.S.; Ahmed, S.F.; Inayat, A.; Kusumo, F.; Badruddin, I.A.; Khan, T.M.Y.; Nghiem, L.D.; Ong, H.C.; et al. Microalgae Biomass as a Sustainable Source for Biofuel, Biochemical and Biobased Value-Added Products: An Integrated Biorefinery Concept. Fuel 2022, 307, 121782. [Google Scholar] [CrossRef]
- Sivaramakrishnan, R.; Suresh, S.; Kanwal, S.; Ramadoss, G.; Ramprakash, B.; Incharoensakdi, A. Microalgal Biorefinery Concepts’ Developments for Biofuel and Bioproducts: Current Perspective and Bottlenecks. Int. J. Mol. Sci. 2022, 23, 2623. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; W. Hassan, S.; Banat, F. An Overview of Microalgae Biomass as a Sustainable Aquaculture Feed Ingredient: Food Security and Circular Economy. Bioengineered 2022, 13, 9521–9547. [Google Scholar] [CrossRef] [PubMed]
- Bunbury, F.; Deery, E.; Sayer, A.P.; Bhardwaj, V.; Harrison, E.L.; Warren, M.J.; Smith, A.G. Exploring the Onset of B 12-Based Mutualisms Using a Recently Evolved Chlamydomonas Auxotroph and B12-Producing Bacteria. Environ. Microbiol. 2022, 24, 3134–3147. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, M.-Y.; Khan, N.; Tan, L.-L.; Yang, S.; Naveed, M. Sustainability Potentials, Utilization, and Bioengineering of Plant Growth-Promoting Methylobacterium for Sustainable Agriculture. Sustainability 2021, 13, 3941–12. [Google Scholar] [CrossRef]
- Calatrava, V.; Hom, E.F. .; Llamas, Á.; Fernández, E.; Galván, A. OK, Thanks! A New Mutualism between Chlamydomonas and Methylobacteria Facilitates Growth on Amino Acids and Peptides. FEMS Microbiol. Lett. 2018, 365, fny021. [Google Scholar] [CrossRef]
- Llamas, A.; Leon-Miranda, E.; Tejada-Jimenez, M. Microalgal and Nitrogen-Fixing Bacterial Consortia: From Interaction to Biotechnological Potential. Plants 2023, 12, 2476. [Google Scholar] [CrossRef]
- Romano, I.; Ventorino, V.; Pepe, O. Effectiveness of Plant Beneficial Microbes: Overview of the Methodological Approaches for the Assessment of Root Colonization and Persistence. Front. Plant Sci. 2020, 11, 6. [Google Scholar] [CrossRef] [PubMed]
- Gyurjan, I.; Koranyi, P.; Paless, G.Y. Ultrastructural Analysis of an Artificial Alga-Bacterium Endosymbiosis After Prolonged Cultivation. Symbiosis 1992, 14, 475–468. [Google Scholar]
- Preininger, E.; Ponyi, T.; Sarkadi, L.; Nyitrai, P.; Gyurjan, I. Long-Living Azotobacter-Chlamydomonas Association as a Model System for Plant-Microbe Interactions. Symbiosis 2006, 42, 45–50. [Google Scholar]
- Chaos-Hernández, D.; Reynel-Ávila, H.E.; Bonilla-Petriciolet, A.; Villalobos-Delgado, F.J. Extraction Methods of Algae Oils for the Production of Third Generation Biofuels – A Review. Chemosphere 2023, 341, 139856. [Google Scholar] [CrossRef]
- Daneshvar, E.; Sik Ok, Y.; Tavakoli, S.; Sarkar, B.; Shaheen, S.M.; Hong, H.; Luo, Y.; Rinklebe, J.; Song, H.; Bhatnagar, A. Insights into Upstream Processing of Microalgae: A Review. Bioresour. Technol. 2021, 329, 124870. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Lyu, S.; An, Y.; Lu, J.; Gjermansen, C.; Schramm, A. Microalgae–Bacteria Symbiosis in Microalgal Growth and Biofuel Production: A Review. J. Appl. Microbiol. 2019, 126, 359–368. [Google Scholar] [CrossRef]
- Nirmala, N.; Praveen, G.; AmitKumar, S.; SundarRajan, P.S.; Baskaran, A.; Priyadharsini, P.; SanjayKumar, S.P.; Dawn, S.S.; Pavithra, K.G.; Arun, J.; et al. A Review on Biological Biohydrogen Production: Outlook on Genetic Strain Enhancements, Reactor Model and Techno-Economics Analysis. Sci. Total Environ. 2023, 896, 165143. [Google Scholar] [CrossRef]
- Frenkel, A.W. Hydrogen Evolution by the Flagellate Green Alga, Chlamydomonas Moewusii. Arch Biochem Biophys 1951, 38, 219–230. [Google Scholar] [CrossRef]
- Xu, X.; Zhou, Q.; Yu, D. The Future of Hydrogen Energy: Bio-Hydrogen Production Technology. Int. J. Hydrogen Energy 2022, 47, 33677–33698. [Google Scholar] [CrossRef]
- King, S.J.; Jerkovic, A.; Brown, L.J.; Petroll, K.; Willows, R.D. Synthetic Biology for Improved Hydrogen Production in Chlamydomonas Reinhardtii. Microb. Biotechnol. 2022, 15, 1946–1965. [Google Scholar] [CrossRef]
- Ben-Amotz, A.; Erbes, D.L.; Riederer-Henderson, M.A.; Peavey, D.G.; Gibbs, M. H2 Metabolism in Photosynthetic Organisms: I. Dark H2 Evolution and Uptake by Algae and Mosses. Plant Physiol 1975, 56, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Melis, A.; Zhang, L.; Forestier, M.; Ghirardi, M.L.; Seibert, M. Sustained Photobiological Hydrogen Gas Production upon Reversible Inactivation of Oxygen Evolution in the Green Alga Chlamydomonas Reinhardtii. Plant Physiol. 2000, 122, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Fakhimi, N.; Gonzalez-Ballester, D.; Fernández, E.; Galván, A.; Dubini, A. Algae-Bacteria Consortia as a Strategy to Enhance H2 Production. Cells 2020, 9, 1353. [Google Scholar] [CrossRef]
- Xu, L.; Cheng, X.; Wang, Q. Effect of Co-Cultivation of Chlamydomonas Reinhardtii with Azotobacter Chroococcum on Hydrogen Production. Int. J. Hydrogen Energy 2017, 42, 22713–22719. [Google Scholar] [CrossRef]
- Wu, S.; Li, X.; Yu, J.; Wang, Q. Increased Hydrogen Production in Co-Culture of Chlamydomonas Reinhardtii and Bradyrhizobium Japonicum. Bioresour. Technol. 2012, 123, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Fakhimi, N.; Dubini, A.; Tavakoli, O.; González-Ballester, D. Acetic Acid Is Key for Synergetic Hydrogen Production in Chlamydomonas-Bacteria Co-Cultures. Bioresour. Technol. 2019, 289, 121648. [Google Scholar] [CrossRef] [PubMed]
- Fakhimi, N.; Tavakoli, O.; Marashi, S.A.; Moghimi, H.; Mehrnia, M.R.; Dubini, A.; González-Ballester, D. Acetic Acid Uptake Rate Controls H2 Production in Chlamydomonas-Bacteria Co-Cultures. Algal Res. 2019, 42, 101605. [Google Scholar] [CrossRef]
- Yu, Q.; He, J.; Zhao, Q.; Wang, X.; Zhi, Y.; Li, X.; Li, X.; Li, L.; Ge, B. Regulation of Nitrogen Source for Enhanced Photobiological H2 Production by Co-Culture of Chlamydomonas Reinhardtii and Mesorhizobium Sangaii. Algal Res. 2021, 58, 102422. [Google Scholar] [CrossRef]
- Iqbal, K.; Saxena, A.; Pande, P.; Tiwari, A.; Chandra Joshi, N.; Varma, A.; Mishra, A. Microalgae-Bacterial Granular Consortium: Striding towards Sustainable Production of Biohydrogen Coupled with Wastewater Treatment. Bioresour. Technol. 2022, 354, 127203. [Google Scholar] [CrossRef]
- Rezvani, F.; Sarrafzadeh, M.H. Autotrophic Granulation of Hydrogen Consumer Denitrifiers and Microalgae for Nitrate Removal from Drinking Water Resources at Different Hydraulic Retention Times. J. Environ. Manage. 2020, 268, 110674. [Google Scholar] [CrossRef]
- Wirth, R.; Lakatos, G.; Maróti, G.; Bagi, Z.; Minárovics, J.; Nagy, K.; Kondorosi, É.; Rákhely, G.; Kovács, K.L. Exploitation of Algal-Bacterial Associations in a Two-Stage Biohydrogen and Biogas Generation Process. Biotechnol. Biofuels 2015, 8, 59. [Google Scholar] [CrossRef] [PubMed]
- Hena, S.; Fatimah, S.; Tabassum, S. Cultivation of Algae Consortium in a Dairy Farm Wastewater for Biodiesel Production. Water Resour. Ind. 2015, 10, 1–14. [Google Scholar] [CrossRef]
- Xu, L.; Cheng, X.; Wang, Q. Enhanced Lipid Production in Chlamydomonas Reinhardtii by Co-Culturing With Azotobacter Chroococcum. Front. Plant Sci. 2018, 9, 741. [Google Scholar] [CrossRef]
- Yee, C.S.; Okomoda, V.T.; Hashim, F.; Waiho, K.; Abdullah, S.R.S.; Alamanjo, C.; Hasan, H.A.; Mustafa, E.M.; Kasan, N.A. Marine Microalgae Co-Cultured with Floc-Forming Bacterium: Insight into Growth and Lipid Productivity. PeerJ 2021, 9, 11217. [Google Scholar] [CrossRef]
- Toyama, T.; Kasuya, M.; Hanaoka, T.; Kobayashi, N.; Tanaka, Y.; Inoue, D.; Sei, K.; Morikawa, M.; Mori, K. Growth Promotion of Three Microalgae, Chlamydomonas Reinhardtii, Chlorella Vulgaris and Euglena Gracilis, by in Situ Indigenous Bacteria in Wastewater Effluent. Biotechnol. Biofuels 2018, 11, 176. [Google Scholar] [CrossRef] [PubMed]
- Miranda, A.M.; Hernandez-Tenorio, F.; Villalta, F.; Vargas, G.J.; Sáez, A.A. Advances in the Development of Biofertilizers and Biostimulants from Microalgae. Biology (Basel). 2024, 13. [Google Scholar] [CrossRef]
- Martini, F.; Beghini, G.; Zanin, L.; Varanini, Z.; Zamboni, A.; Ballottari, M. The Potential Use of Chlamydomonas Reinhardtii and Chlorella Sorokiniana as Biostimulants on Maize Plants. Algal Res. 2021, 60, 102515. [Google Scholar] [CrossRef] [PubMed]
- Metting, B. Population Dynamics of Chlamydomonas Sajao and Its Influence on Soil Aggregate Stabilization in the Field. Appl. Environ. Microbiol. 1986, 51, 1161–1164. [Google Scholar] [CrossRef] [PubMed]
- Sido, M.Y.; Tian, Y.; Wang, X.; Wang, X. Application of Microalgae Chlamydomonas Applanata M9V and Chlorella Vulgaris S3 for Wheat Growth Promotion and as Urea Alternatives. Front. Microbiol. 2022, 13, 1035791. [Google Scholar] [CrossRef]
- Gitau, M.M.; Farkas, A.; Balla, B.; Ördög, V.; Futó, Z.; Maróti, G. Strain-Specific Biostimulant Effects of Chlorella and Chlamydomonas Green Microalgae on Medicago Truncatula. Plants 2021, 10, 1060. [Google Scholar] [CrossRef]
- Mutale-Joan, C.; Redouane, B.; Najib, E.; Yassine, K.; Lyamlouli, K.; Laila, S.; Zeroual, Y.; Hicham, E.A. Screening of Microalgae Liquid Extracts for Their Bio Stimulant Properties on Plant Growth, Nutrient Uptake and Metabolite Profile of Solanum Lycopersicum L. Sci. Rep. 2020, 10, 2820. [Google Scholar] [CrossRef] [PubMed]
- Stirk, W.A.; Ördög, V.; Staden, J. Van; Jäger, K. Cytokinin-and Auxin-like Activity in Cyanophyta and Microalgae. J. Appl. Phycol. 2002, 14, 215–221. [Google Scholar] [CrossRef]
- Alvarez, A.L.; Weyers, S.L.; Goemann, H.M.; Peyton, B.M.; Gardner, R.D. Microalgae, Soil and Plants: A Critical Review of Microalgae as Renewable Resources for Agriculture. Algal Res. 2021, 54, 102200. [Google Scholar] [CrossRef]
- Keswani, C.; Satyendra, &; Singh, P. ; Cueto, L.; García-Estrada, C.; Mezaache-Aichour, S.; Glare, T.R.; Borriss, R.; Surya, &; Angel Blázquez, M.; et al. Auxins of Microbial Origin and Their Use in Agriculture. Appl. Microbiol. Biotechnol. 2020, 104, 8549–8565. [Google Scholar] [PubMed]
- Amin, S.A.; Hmelo, L.R.; van Tol, H.M.; Durham, B.P.; Carlson, L.T.; Heal, K.R.; Morales, R.L.; Berthiaume, C.T.; Parker, M.S.; Djunaedi, B.; et al. Interaction and Signalling between a Cosmopolitan Phytoplankton and Associated Bacteria. Nature 2015, 522, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Palombella, A.L.; Dutcher, S.K. Identification of the Gene Encoding the Tryptophan Synthase-Subunit from Chlamydomonas Reinhardtii. Plant Physiol 1998, 117, 455–464. [Google Scholar] [CrossRef]
- Calatrava, V.; Hom, E.F.Y.; Guan, Q.; Llamas, A.; Fernández, E.; Galván, A. Genetic Evidence for Algal Auxin Production in Chlamydomonas and Its Role in Algal-Bacterial Mutualism. iScience 2024, 27, 108762. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The Rhizosphere Microbiome: Significance of Plant Beneficial, Plant Pathogenic, and Human Pathogenic Microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef]
Figure 1.
Schematic overview of the wastewater treatment process using Chlamydomonas-bacterial consortia. Wastewater treatments from various sources such as agricultural, industrial, medical, and municipal activities involve, physical, chemical, mechanical and biological processes including phytoremediation with microalgae. Consortia between Chlamydomonas and bacteria can be cultivated in open reactors, photobioreactors (PBR), biofilms or being encapsulated in beads. The final stage is the harvest of the biomass which results in bioremediated treated water.
Figure 1.
Schematic overview of the wastewater treatment process using Chlamydomonas-bacterial consortia. Wastewater treatments from various sources such as agricultural, industrial, medical, and municipal activities involve, physical, chemical, mechanical and biological processes including phytoremediation with microalgae. Consortia between Chlamydomonas and bacteria can be cultivated in open reactors, photobioreactors (PBR), biofilms or being encapsulated in beads. The final stage is the harvest of the biomass which results in bioremediated treated water.
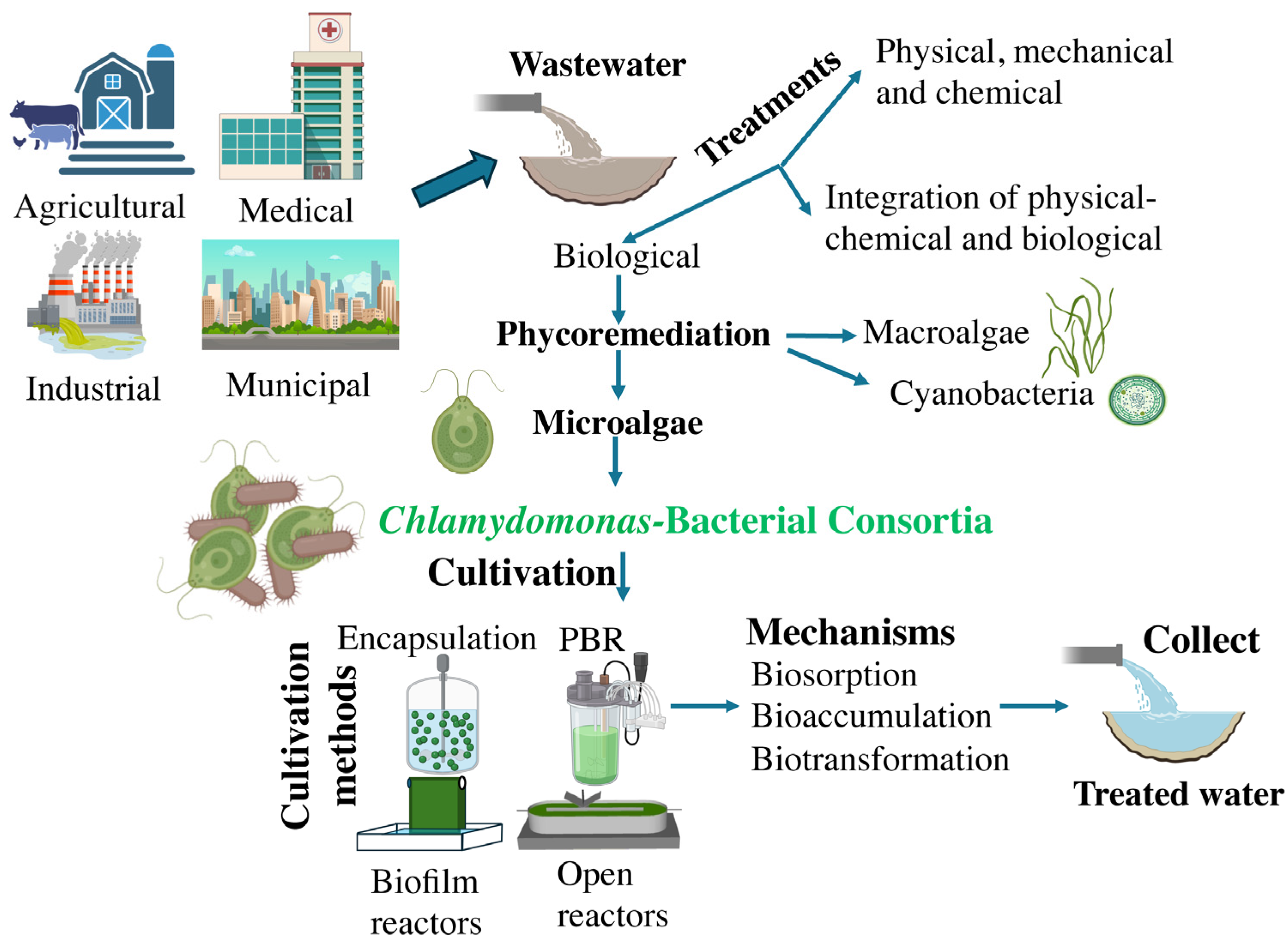
Figure 2.
Schematic overview of a hypothetical model based on the concept of a circular economy using Chlamydomonas-bacteria consortia in wastewater treatment plants. Different wastewater sources are treated in wastewater treatment plants utilizing Chlamydomonas-bacteria consortia. This treatment results in algal biomass, which can be converted into bio-hydrogen production, biofuel through lipid accumulation and biofertilizers. The clean, bioremediated water produced exhibits reduced COD (Chemical Oxygen Demand), low inorganic nitrogen levels, and low lipid and protein content, rendering it suitable for reuse in various applications, including the irrigation of agricultural crops, among others.
Figure 2.
Schematic overview of a hypothetical model based on the concept of a circular economy using Chlamydomonas-bacteria consortia in wastewater treatment plants. Different wastewater sources are treated in wastewater treatment plants utilizing Chlamydomonas-bacteria consortia. This treatment results in algal biomass, which can be converted into bio-hydrogen production, biofuel through lipid accumulation and biofertilizers. The clean, bioremediated water produced exhibits reduced COD (Chemical Oxygen Demand), low inorganic nitrogen levels, and low lipid and protein content, rendering it suitable for reuse in various applications, including the irrigation of agricultural crops, among others.
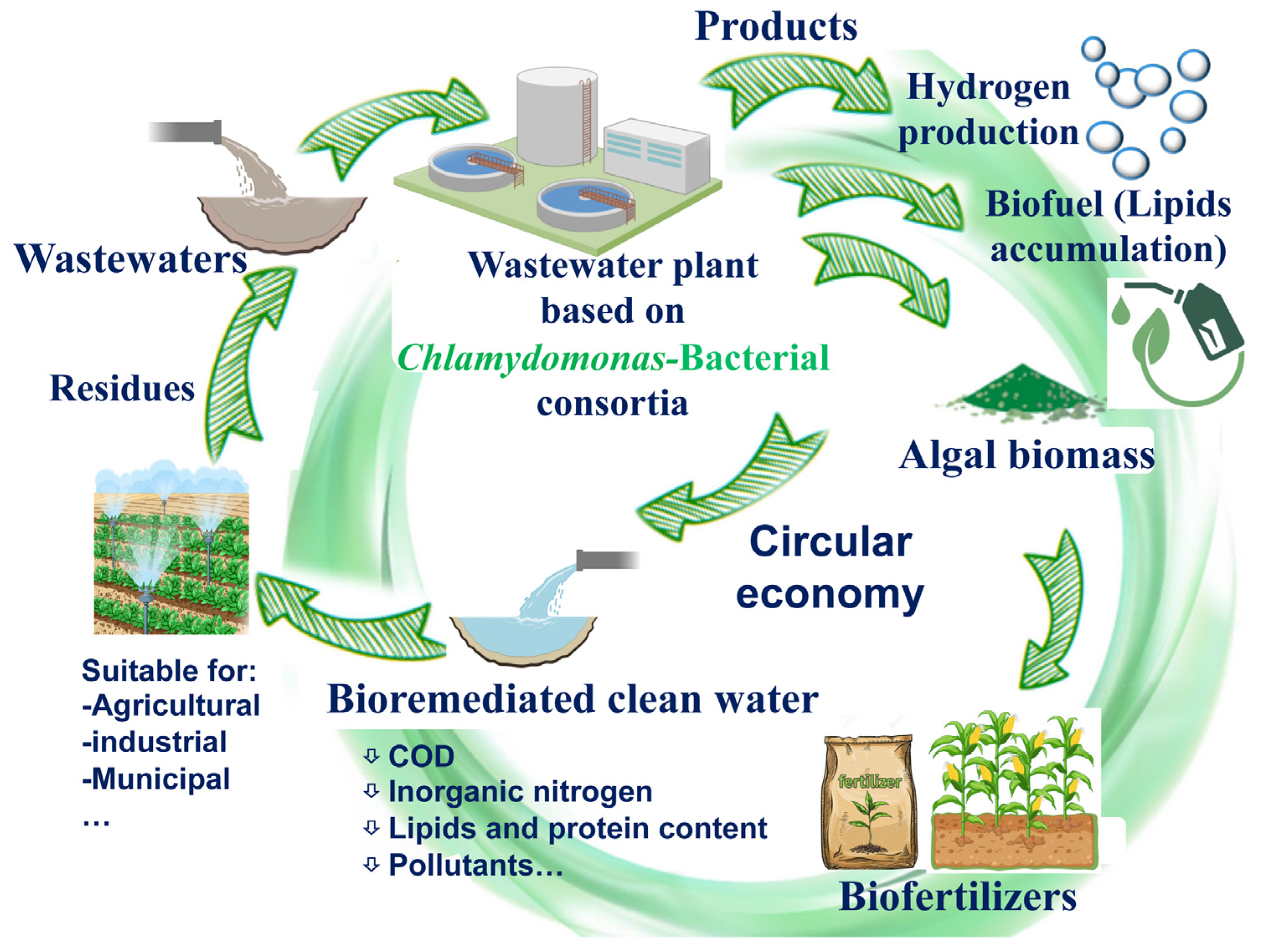
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



