
Submitted:
14 June 2024
Posted:
14 June 2024
You are already at the latest version
Abstract
Probiotics are becoming more and more popular in the growing of fin and shellfish because these species are consumed worldwide and because different techniques are employed to maximize the productivity and efficiency of fin and shellfish. Probiotic Bacillus is one of the many types of probiotics that are used, but it stands out due to its superior qualities. For instance, because of its ability to sporulate, it can survive in harsh environments, produce antimicrobial substances, and is safe to feed to fish. The different species of Bacillus used in the farming of fin and shell fish were discussed in this review, along with the benefits of using Bacillus spp. as a good substitute in sustainable aquaculture to improve feed utilization, immune system response, stress adaptation, resistance to infectious diseases, tissue preservation integrity, and water quality. Additionally, we discussed the possible risks to safety that Bacillus poses when raising these significant species. For the purpose of future research and development regarding Bacillus application in aquaculture, a summary of the findings of recent studies about the advantages of applying Bacillus to enhance fin and shellfish aquatic animal culture has been provided.
Keywords:
Bacillus
; finfishes
; shellfishes
; antibiotics
; aquaculture
; bacteria
; probiotics
1. Introduction
A significant obstacle to the cultivation and rearing of fin and shellfish is the rise in illnesses among aquaculture species brought on by high stocking levels to meet the enormous demand for fish [1]. Due to the negative effects of antibiotics, such as bacterial resistance to common antimicrobials resulting from changes in the microbiota of aquaculture systems, which in turn affects the naturally occurring beneficial bacteria flora, probiotics have become a viable alternative to antibiotics [2].
The aquaculture of fin and shellfishes has expanded quickly to become a significant global sector that employs hundreds of thousands of trained and unskilled individuals in addition to generating economic revenue and high-quality food products [3]. According to projections, there will be nine billion people on the earth by the year 2050, and aquaculture will play a significant part in meeting the growing demand for food [4]. Finding long-lasting, ecologically acceptable treatments to fin and shellfish disease is a top priority for researchers [5]. As a result of this, over time, a variety of probiotics have been found and used to improve the growth and immunity of aquaculture species. These probiotics include Arthrobacter, Enterococcus, Bacillus, Lactobacillus, Micrococcus, Lactococcus, Aeromonas, Pediococcus, Enterobacter, Burkholderia, Vibrio, Rhodopseudomonas, Pseudomonas, Shewanella and Roseobacter [6]. Probiotics are used as a safe supplement in aquaculture to enhance the host's health through growth promotion, nutrient provision, microbial colonization modulation, immune response enhancement, feed utilization improvement, increased digestibility and activity of digestive enzymes, improved water quality, and disease control [7].
Bacillus species have a twofold advantage in terms of survival (heat tolerance and longer shelf life) in a variety of environments: their capacity for sporulation, which prolongs their period of effectiveness, and their ability to produce antimicrobial substances that are effective against a wide range of microbes and are non-pathogenic and non-toxic [8]. Bacillus species have been shown to boost the activity of antioxidant and digestive enzymes, as well as the expression of genes linked to stress and immunity, enhancing fish's resistance to pathogenic bacteria [9]. Additionally, Bacillus species improve fish feed utilization, which increases growth rate [9,10]. The goal of this review is to gather data on the role of Bacillus species in regulating digestive enzymes, antioxidant enzymes, immune, stress, and other related gene expression, hepatic indices, disease resistance, feed utilization and growth, and future prospects of Bacillus species in fin fish and shellfish aquaculture, as Figure 1 illustrates.
2. Characteristics of Bacillus spp.
One of the most prevalent groups of microorganisms in nature is the Bacillus bacteria, they are found in soil, water, and the atmosphere [11]. Figure 2 shows the characteristics of Bacillus species that make them viable probiotic options for sustainable growth of fin and shellfish. The ability of Bacillus, a diverse group of rod-shaped, Gram-positive bacteria, to produce a potent spore sets them apart. Bacillus species are rod-shaped, gram-positive, chemoheterotrophic, aerobic, or facultatively anaerobic, catalase-positive bacteria that are usually motile by peritrichous flagella and do not have capsules [12]. Bacillus produce spores that can be kidney-shaped, round, oval, or cylindrical and are more resistant to heat, drying, and disinfectants than their vegetative cells [3] thus continue to exist for a long time. Each cell contains one spore, and exposure to air has no effect on sporulation. Bacillus species are often beta-hemolytic and grow in large, flat colonies on non-selective media. Clistridia and sporolactobacillus are not catalase-negative and aerobic, which distinguishes the few genius Bacillus species from each other [9]. The capacity of numerous Bacillus species to produce antibiotics and other compounds with antagonistic effects on harmful germs makes them significant [4]. Due to their capacity to create antibiotics and other compounds with antagonistic effects on harmful microbes, many Bacillus species are significant [16].
Table 1 illustrates how some commercial Bacillus products have been used to generate a broad spectrum of metabolites with antimicrobial action [17]. They are employed in fin and shellfish production as growth boosters, disease-resistant organisms, and to improve water quality indicators [18]. Over the years, they have been used in experiments to decrease harmful bacteria in fish since they are more affordable, more effective sources of antibiotics, and sometimes even non-toxic and non-pathogenic [16,19]. Similar to other probiotics, Bacillus species possess characteristics such as the capacity to inhibit infections by competing for adhesion sites to halt their growth and the capacity to generate antibiotics [16,20] and bacteriocins [20], quorum quenching: the process of inhibiting the expression of virulence genes and producing lytic enzymes that break down the cell walls of pathogenic microorganisms, like cellulases, proteases, chitinases, and β-1,3-glucanases [2]. The provision of nutrients and enzymatic digestion, which promotes development by secreting digestive enzymes, is another property of Bacillus [4]. Additionally, the immune-stimulating properties of Bacillus species and their encouragement of healthy gut flora enhance the host's innate and adaptive defenses [5]. Bacillus licheniformis, B. subtilis, B. amyloliquefaciens, and B. pumilus are the majority of the Bacillus species that are used as probiotics in fish [26].
3. Utilizing Bacillus in Shellfish and Finfish
3.1. Application of Bacillus in Fin fishes
Numerous investigations have discovered counts of perhaps Bacillus, but no additional identification has been carried out. In this section, the following, bacteria identified as B. aerophilus, B. aerius, B. amyloliquefaciens, B. aryabhattai, B. altitudinis, B. atrophaeus, B. circulans, B. cereus, B. clausii, B. flexus, B. coagulans, B. megaterium, B. stratosphericus, B. licheniformis, B. thermoamylovorans, B. methylotrophicus, B. sonorensis, B. pumilus, B. tequilensis, B. subtilis, B. thuringiensis, and Solibacillus silvestris isolated from the finfish GI tract are discussed.
3.1.1. Bacillus aerius
B. aerius was proposed as a new species within the genus Bacillus by [28]; it was isolated from a cryogenic tube used for sampling air from high altitudes [29]. The strain was able to form endospores, stained Gram-positive, and contained peptidoglycans in its cell walls, with DL-diaminobutyric acid serving as the diamine [29]. The strain's variations in genotypic and phenotypic characteristics allowed it to be identified from strains of the closest related species [29,30]. Few research, as far as we know, have isolated this bacterium from finfish's gastrointestinal tract. [31] found autochthonous B. aerius in the distal intestine (DI) catla (Catla catla) in a study that examined the probiotic properties of exoenzyme-producing bacteria.
Another study used B. aerius strain B81e, which was chosen for its probiotic qualities both in vitro and in vivo after being carefully isolated from the gut of healthy catfish. This bacterium produced a substance resembling bacteriocin and inhibited both Gram-positive and Gram-negative bacteria, including the fish diseases Aeromonas hydrophila and Streptococcus agalactiae. It also demonstrated broad-spectrum antibacterial action [32].
Another study examined the positive effects of mixed probiotics (B. aerius B81e þ L. paraplantarum L34b-2) on fish growth performance, innate immunity, and illness resistance. According to the study, Pangasius fish showed notable probiotic benefits when Bacillus aerius from the Bacillus genus was combined with other bacteria [33].
3.1.2. Bacillus aerophilus
This species possesses traits common to Bacillus species: it is Gram-positive stained, capable of producing endospores, and contains DL-diaminobutyric acid-containing cell-wall peptidoglycans [12]. [28] proposed B. aerophilus as a new species in the genus Bacillus based on isolates from single strains kept in cryogenic tubes that were used to gather air samples at high elevations. [34] reported Bacillus aerophilus, a bacterium with probiotic potential based on good growth in intestinal mucus, resistance to diluted bile juice (2–20%), safety for the target fish, and production of bacteriocin in a study evaluating the autochthonous microbiota of Indian major carp (Cirrhinus mrigala) by cultivation.
In a different investigation, B. aerophilus was identified and employed to strengthen Labeo rohita's immune system as well as assess the impacts against Aeromonas hydrophila [35].
3.1.3. Bacillus amyloliquefaciens
Bacillus amyloliquefaciens was initially isolated from soil in 1943 by Juichiro Fukumoto, a scientist from Japan. The species' unusual characteristic led to the naming of the species because it generated a liquefying α-amylase (amylo) [36,37].
Gram-positive, nonpathogenic, endospore-forming B. amyloliquefaciens is a member of a class of free-living soil bacteria that has several characteristics, such as promoting plant growth, producing metabolites that are antifungal and antibacterial, and producing industrially significant enzymes [38]. In eight finfish research involving eight different fish species, this bacterium has been isolated; in seven of those studies, culture-based techniques were employed. The majority of strains had the ability to produce bacteriocin, enzyme production and antagonism, and probiotic potential. More recently, the intestines of flounders have been used to identify this specific species of Bacillus [39], southern flounder (Paralichthys lethostigma) [40], rohu (Labeo rohita) [41] and Indian major carp [42] employing techniques based on culture, and these investigations showed encouraging aspects.
3.1.4. Bacillus altitudinis
The rod-shaped, gram-positive aerobic bacterium B. altitudinis belongs to the phylum Firmicutes. The first report of its isolation came from samples of extremely UV-stressed air taken in the stratosphere [28]. Since then, reports of B. altitudinis have been made in a variety of habitats, such as the deep freshwater of Manasbal Lake [44], the southern Indian Ocean [43], soil [45], and silt [46].
It has been reported by the culture in the DI of mrigal [47], proximal intestine (PI) of walking catfish (Clarias batrachus) [48] and stinging catfish (Heteropneustes fossilis). In a study, the bacteria B. altitudinis AP-MSU was isolated from the stomach of marine fish Sardinella longiceps and found to be capable of producing esterase at a reasonable cost by using fish processing waste [49].
3.1.5. Bacillus aryabhattai
Bacillus aryabhattai are widely distributed in nature but were only discovered in 2009 [50]. It has been established that this isolate is a long rod, spore-forming, motile, strictly aerobic Gram-positive bacteria that grows best at 30 °C and is non-pigmented on agar-containing media [51].
This species was found in a study that evaluated the probiotic value of bacilli isolated from the intestines of Rhynchocypris lagowskii [52].
3.1.6. Bacillus atrophaeus
Migula initially identified and isolated B. atrophaeus as B. globigii in 1900 [54]. After observing pigment formation when cultured in medium containing tyrosine, it was reexamined and reclassified as B. subitilis var niger [55]. Like other endospore-forming bacteria, B. atrophaeus has three distinct stages in its life cycle: vegetative growth, sporulation, and germination [56]. Spores have a remarkable resistance to environmental damages, including heat, radiation, toxic chemicals, and pH extremes, and they can remain dormant for extended periods of time [54]. Spore germination and outgrowth is the process by which a spore breaks its dormancy and resumes growth in the presence of favorable environmental conditions [54]. This indigenous bacterium has been identified through culturing in the proximal intestine (PI) of walking catfish (Clarias batrachus) [48], stinging catfish (Heteropneustes fossilis) [53], and the DI of mrigal [47]. The primary characteristic of these bacilli is their capacity to manufacture enzymes, and Bacillus atrophaeus is well-known for producing antibiotic substances [54].
3.1.7. Bacillus cereus
The B. cereus group comprises Gram-positive bacteria with low GC content, which belong to the phylum Firmicutes [12]. At least eight closely related species of rod-shaped, facultatively anaerobic, aerobic, spore-forming bacteria are included in this group: B. anthracis, B. thuringiensis, B. cereus, B. mycoides, B. weihenstephanensis, B. pseudomycoides, B. toyonensis and B. cytotoxicus[12]. The genomes of the B. cereus group species, with sizes ranging from 5.2 to 5.9 Mb and relatively comparable 16S rRNA gene sequences, are substantially conserved, with the exception of B. cytotoxicus, which is the most diverged of the group with a chromosome of 4.085 Mb (2) [12]. The mrigal PI and DI contained this amylase, cellulase, and protease-producing bacterium [21]. Cultivable native Bacillus cereus was discovered in the DI of wild olive flounder (Paralichthys olivaceus) fish in a study [22] evaluating the microbial diversity in the guts of farmed and wild fish. When the bacterial population in DI of Atlantic salmon (Salmo salar) was analyzed by culture, B. cereus was found to be a component of the community [57]. In a study by [42], B. cereus was discovered from the intestines of an Indian big carp using a culture-based approach.
3.1.8. Bacillus circulans
Gram-positive B. circulans is a rod that is motile by peritrichous flagella and has a size range of 2.0-4.2 x 0.5-0.8 μm [58]. Colonies that are cultivated at 30°C have irregular margins, an opaque cream color, and a diameter of 1-3 mm [58]. On nutrient agar, the growth is thin and can spread quickly in certain strains [58]. One type of facultative anaerobe is B. circulans [58]. Endospores are produced by this bacterium and can be ellipsoidal, subterminal, or terminal [59]. When living conditions are unfavorable, spores enable bacteria to lie dormant for prolonged periods of time. However, the endospore can reactivate itself into its vegetative stage when favorable conditions arise again [59].
In four culturing tests, B. circulans was discovered in the GI tract of rainbow trout fed different antibiotics [60]. Research on the intestinal microbiota of rohu [61], common carp [62], and mossambicus tilapia [63] also revealed thisbacteria. The strains exhibited enzyme-producing activity in the rohu, common carp, and tilapia tests.
B. circulans was isolated from Catla catla's intestinal tract and supplemented on C. catla fingerlings in a study. Supplementation had favorable effects on growth performance, immunological response, and feed utilization efficiency [64].
3.1.9. Bacillus clausii
Even in the presence of antibiotics, B. clausii, an aerobic, gram-positive, rod-shaped bacterium, can produce spores and endure passage through the stomach's acidic environment to colonize the intestine [65]. This Bacillus species was isolated by [66] from the gut of an orange-spotted grouper (Epinephelus coioides), and it showed amazing resistance to mimicking the GI environment as well as antagonistic activity against certain potentially harmful bacteria. In a different study, this species of Bacillus was isolated from the intestinal tract of groupers (Epinephelus coioides), and it showed benefits for the growth and general well-being of Japanese flounder fish when compared to other diets [67].
3.1.10. Bacillus coagulans
A gram-positive, spore-forming, microaerophilic bacillus that produces lactic acid is called B. coagulans. It was first identified as Lactobacillus sporogenes by Horowitz and Wlassowa in 1932 after it was isolated [68]. Based on its biochemical characteristics, the organism was reclassified in 1957 in Bergey's Manual of Determinative Bacteriology; as a result, Bacillus coagulans is the current correct nomenclature [69]. Among probiotics, B. coagulans is special because it has a protein coating that resembles a spore and protects it from stomach acid. This coating enables B. coagulans to enter the small intestine, germinate, and grow [68]. [21] identified autochthonous bacteria that produce amylase, cellulase, and protease in the posterior and distal intestines of three major Indian carp species: catla, mrigal, and rohu. [70] in their study isolated and identified the potential gut adherent B. coagulans from the stomach of Catla catla and to assess how supplementing that B. coagulans affects the experimental fish's ability to grow and retain nutrients. Common carp is the source of B. coagulans, which is utilized as a dietary probiotic. Growth performance, the capacity to elicit a favorable immunological response, and meat quality all showed improvements [71].
3.1.11. Bacillus flexus
Oxidase-positive, rod-shaped, aerobic, Gram-variable B. flexus is a bacterium that forms endospores [72]. The ellipsoidal endospores are found in unswollen, central/paracentral sporangia [58]. When cultured for 24–72 hours at 30 ± 2°C, it yields opaque, cream-colored, raised-margin colonies in the lab [58]. This bacterium species has only been identified in two investigations that looked at the GI tract of mrigal [73] and walking catfish (Clarias batrachus) [48].
3.1.12. Bacillus licheniformis
A spore-forming, Gram-positive soil bacterium is called B. licheniformis [58]. It can survive under favorable conditions in a vegetative state or in a dormant spore form to withstand harsh environments [58]. This species produces a variety of extracellular enzymes that may aid in the natural cycling of nutrients and is closely linked to the extensively researched model organism B. subtilis [58]. Numerous species of warm- and cold-water finfish have been shown to have B. licheniformis, varying in their ability to suppress pathogens and produce enzymes. Using culture-based techniques, [74] and [75] isolated this species of Bacillus from the posterior intestines of Rohu and Bata (Labeo bata). B. licheniformis was found in the posterior and distal intestines of Atlantic salmon through a culture method investigation conducted by [57]. However, the strain did not show any encouraging exo-enzyme activities or in vitro growth inhibition towards the four pathogens that were tested. Later, B. licheniformis was shown to be a member of the autochthonous enzyme-producing bacteria isolated from PI and DI of two species of Indian air-breathing fish, stinging catfish and murrel (Channa punctatus), using a standard culture approach [76]. B. licheniformis, an exo-enzyme-producing culturable autochthonous organism, was found in the DI of farmed olive flounder [22]. [77] discovered anomalies in exo-enzyme activity after isolating a strain of B. licheniformis from the mid-intestitine (MI) of long whiskers catfish (Mystus gulio). In three studies, Bacillus licheniformis was isolated from the guts of Mrigal [34], Rohu intestine [34], and Nile tilapia [78], and the results showed potential probiotic and pathogen inhibition properties.
3.1.13. Bacillus megaterium
Bacillus megaterium is a rod-shaped, aerobic, Gram-positive bacterium that forms spores and is found in a wide range of environments [79]. Its cells are relatively large for bacteria, measuring up to 100 µm in length and 0.1 µm in diameter [80]. Polysaccharides on the cell walls bind the cells together, which is why the cells frequently occur in pairs and chains [79]. Temperatures between 3 and 45 °C are ideal for B. megaterium growth, with an average of 30 °C. It was discovered that certain isolates from an Antarctic geothermal lake could grow at 63 °C [58]. [81] assessed the gut microbiota in the DI of Atlantic salmon and reported no notable features. [63] and [47] discovered that the guts of Mossambicus tilapia and PI of grass carp (Cetenopharyngodon idellus) were producing antagonistic Bacillus megaterium and exo-enzymes.
3.1.14. Bacillus methylotrophicus
Gram-positive Bacillus methylotrophicus is a rod-shaped, specifically aerobic, mobile bacterium that also produces endospores [82]. It is reported to grow well at 30°C with a pH of 7.0 in ammonium mineral salts (AMS) medium containing 600 mM methanol [82]. It was discovered by [22] that B. methylotrophicus is a part of the microbial diversity in the DI of wild and farmed olive flounder. Additionally, B. methylotrophicus was isolated from the EI of channel catfish [83] and PI of Indian major carp [84]. More recently, [41] and [85] isolated Bacillus methylotrophicus, which may have probiotic qualities, from the PI of rohu and the EI content of rainbow trout.
3.1.15. Bacillus nealsonii
Bacillus nealsonii, a spore-former belonging to the genus Bacillus, was isolated from a spacecraft assembly facility. It is defined by its phenotypic traits, 16S rDNA sequence analysis, and DNA-DNA hybridization studies [86]. Endospores are produced by this rod-shaped, facultatively anaerobic, gram-positive eubacterium [86]. This new species of bacteria has spores that are resistant to UV, gamma, H2O2, and desiccation [87]. B. nealsonii was discovered in the intestine of rainbow trout when antibiotics were used against both significant and non-pathogenic fish pathogens [88].
3.1.16. Bacillus pumilus
A common Gram-positive spore-forming bacterium found in a variety of settings, such as soil, deep-sea sediments, and marine water, is B. pumilus [89]. Significant resistance to environmental stressors, such as drought, low or no nutrient availability, irradiation, UV radiation, chemical disinfectants, or oxidizing enzymes, is exhibited by this species [90]. The first study, carried out by [91], identified B. pumilus in rohu fish intestines. The bacterial isolate generated extracellular protease, amylase, and cellulase, leading the researchers to speculate that this specie of bacteria might be essential for rohu fingerling nutrition. [92] identified an allochthonous B. pumilus that may have probiotic qualities from the EI of channel catfish. [83] discovered B. pumilus in the gut of orange-spotted grouper. Based on cultivation, B. pumilus was revealed as a member of the bacterial population derived from the fish distal intestine (DI) in [93]'s study of the gut microbiota of Salmo trutta. Furthermore, [73] recovered this autochthonous bacilli species with chitinae-producing characteristics from the silver carp (Hypophthalmichthys molitrix) DI. When rainbow trout (Onchorhynchus mykiss) were served linseed oil, autochthonous B. pumilus was discovered in their diet, according to a review published in [94]. The bacteria, however, was absent in the DI of fish that were fed marine, rapeseed, or sunflower oil. From the PI of mrigal and rohu, species of the enzyme-producing B. pumilus exhibiting an unfavorable trait were sequestered [47]. With an excellent adhesion feature, this bacilli species was recently isolated from the EI rainbow trout by [85].
3.1.17. Bacillus sonorensis
Gram-positive aerobic bacterium B. sonorensis belongs to the Bacillus subtilis group of microorganisms and forms endospores [95]. It was originally isolated from soil in the Sonoran Desert, and it shares many characteristics with B. licheniformis, including the ability to be facultatively anaerobic, with it [96]. [97] discovered strains of antagonistic autochthonous B. sonorensis with the ability to produce enzymes in the DI of mrigal. The author suggested that more in vivo studies be conducted to clarify their effects on growth performance and health in light of their findings.
3.1.18. Bacillus subtilis
B. subtilis is a Gram-positive, catalase-positive bacterium that is found in soil, the gastrointestinal tracts of humans, ruminants, and marine sponges. It is also referred to as the hay bacillus or grass bacillus [98]. B. subtilis is a rod-shaped member of the genus Bacillus that can withstand harsh environmental conditions by forming a hard, protective endospore [30]. Despite evidence that B. subtilis is a facultative anaerobe, it has historically been categorized as an obligate aerobe [58]. Several studies on finfish have revealed this species in their GI tracts [99,100,101,102]. It was identified as autochthonous in Atlantic salmon PI and DI by [57], and further research revealed similar results by [81]. [103] investigated the microbial communities present in the digestive tracts of three distinct fish species. From the intestine of catfish, a B. subtilis strain which was isolated exhibited exceptional antimicrobial activity against Edwardsiella ictaluri [83]. [76] used a base-culture technique to recover B. subtilis via three fish species. Additionally, this sort has been derived using culture techniques from cyprinid species with DI content [104], southern flounder intestines [39], Nile tilapia PI and DI [105], and Indian major carp [42]. It is important to note that autochthonous Bacillus subtilis was isolated from MI of mrigal [106] and that this revealed high phytase activity. This information was then utilized in a solid-state fermentation study [107] when discussing the presence of B. subtilis. An autochthonous B. subtilis that may be a probiotic was isolated from DI Indian major carp [84]. The authors proposed that the bacterium could be a bio-control agent, but further research into in vivo studies is warranted.
3.1.19. Bacillus tequilensis
B. tequilensis is a rod-shaped, single-celled, motile, Gram-positive bacterium. In terms of biochemistry, B. tequilensis and B. subtilis are fairly similar; however, positive arginine hydrolases, lysine decarboxylase, ornithine decarboxylase, and the ability to produce acid from rhamnose can distinguish the two species [108]. Only one recent study has shown that this species is present in the PI of silver carp, indicating that it is rarely isolated from finfish [47]. In comparison to the other derived from the fish group, the isolate showed high activities of cellulase and xylanase. Furthermore, strain HMF6X exhibited anti-Aeromonas salmonicida activity.
3.1.20. Bacillus thermoamylovorans
A facultative anaerobic organism, B. thermoamylovorans can ferment in the absence of oxygen and produce ATP in the presence of oxygen [58]. Spores produced by B. thermoamylovorans have a high degree of heat resistance, and they have been shown to withstand industrial food sterilization procedures [109]. The organism can grow between 40°C and 58°C and is facultatively anaerobic [110].
To our knowledge, only one finfish study [111] investigated contents of Salvelinus alpinus and found B. themoamylovorans However, neither the probiotic potential nor the production of extracellular enzymes were further evaluated; these are topics that demand more investigation.
3.1.21. Bacillus thuringiensis
The entomocidal parasporal crystal proteins that B. thuringiensis produces, its capacity to dwell in an environment free of other Gram-positive spore-forming bacilli, and its ability to survive in an exclusive environmental niche and in the gut of organisms are just a few of its many unique characteristics [112]. [57] showing in their research a native in Atlantic salmon indicated Bacillus derived inhibited the growth of pathogens. Subsequently, [88] revealed B. thuringiensis in rainbow trout gut.
3.1.22. Bacillus silvestris
Round endospore-forming, rod-shaped, peritrichously flagellated, aerobic, Gram-positive bacteria is called B. silvestris [58]. In a swollen sporangium, the spore position is terminal [114]. Members of Bacillus RNA group 2 are related to it [115]. The primary structure of the 16s rDNA and its phenotypic characteristics allow the isolate to be identified from other species within this group [115]. In a study carried out by [116] the probiotic B. silvestris was initially isolated from forest soil. According to [115], the bacteria was reclassified as B. silvestris. The presence of B. silvestris in fish gastrointestinal tracts was studied in Indian large carp [34].
4. Effects of Bacillus spp on Fin Fish and Shellfish
Bacillus spp has been proven to be a viable, safe and reliable probiotic in finfish and shellfish culture [7]. It has been shown that certain Bacillus species increase the activity of digestive and antioxidant enzymes, improve feed utilization for faster growth, express genes related to immunity and stress, and, most importantly, strengthen the fin's defenses against harmful microorganisms [24,117,118]. This section elaborates on the effects on finfish and shellfish culture.
4.1. Impact of Bacillus Species on Finfish
Over the past ten years, considerable work has been done to evaluate different bacterial species as probiotics in aquaculture [119,120]. Bacillus sp. has a positive impact on the growth performance and disease resistance of several fish species [27,121]. It appears that B. subtilis was the primary focus of the investigations conducted on Bacillus sp [122]. The impacts of several Bacillus species on fin fish species are outlined and discussed in this section.
4.1.1. The Effects of Bacillus on Immunological Parameters and Disease Resistance in Finfish
Bacterial infections are the most frequent cause of disease issues in aquaculture [123]. Streptococcus agalactiae is one of the bacterial pathogens [124]. According to reports, and Aeromonas hydrophila [125] can result in significant financial losses for fish farms. Drug-resistant microorganisms have emerged posing a threat to the environment and compromising food safety [126,127]. Additionally, vaccinations cannot be used as a universal control agent because they are only effective against a limited subset of pathogenic bacteria [128]. Probiotics have therefore been presented as an alternative to improve fish health and manage illnesses [129,130]. Probiotics are thought to work by generating antimicrobial compounds, which drive out pathogens and compete with them for nutrients and space [131,132]. They also cause the host's nonspecific or specific immune system to become more active, which strengthens the host's defense against pathogens. An increasing amount of research has specifically addressed the application of Bacillus species to the management of bacterial diseases in aquaculture. It appears that after adding Bacillus to the diet, there was an increase in resistance to S. iniae [133], A. hydrophila [134], Acinetobacter sp. and Acinetobacter tandoii [135], as well as Aeromonas salmonicida, S. agalactiae, Lactococcus garvieae, and Vibrio parahemolyticus [136]. Additionally, it has been documented that dietary Bacillus subtilis administration improves disease resistance in a variety of aquatic species, including rainbow trout [137], tilapia [138], and white shrimp [139]. As a result, Bacillus species have proven to be useful as substitutes for antibiotics. The probiotic Bacillus sp. reduced the abundances of Aeromonas and Pseudomonas sp. in the tilapia gut microbiota [140]. The Nile tilapia's resistance to disease and general health were improved by Bacillus pumilus and P. fluorescens [141]. Diets enriched with Bacillus subtilis exhibit enhanced prophylactic qualities against the pathogenic microorganism Streptococcus agalactiae [142].
4.1.2. Effects of Bacillus as a Growth Promoter in Fin Fishes
It is now widely acknowledged that a significant amount of farmers' expenses—roughly 50–60%—are related to diet [143]. As a result, numerous researchers are developing various strategies to cut costs, particularly through the use of various growth parameter kinds [144,145]. Probiotics have exhibited growth properties in variety of fish species if administered through diets [146]. Probiotics are thought to do this by improving intestinal physiology and generating exogenous enzymes [147].
In a study conducted by [148], B. subtilis was fed to rohu at three different levels (0.5, 1.0, and 1.5 ×107 CFU g−1) for a period of 15 days, and a noteworthy rise in weight gain was noted. Afterwards, [149] added different concentrations (4.8×108, 1.2×109, 2.01×109, 3.8×109, and 6.1×109 CFU g-1) of commercial B. subtilis to the feed of rainbow trout fry. Fish fed probiotics demonstrated a discernible increase in growth indices after administration (13 days). Additionally, in a 28-day feeding trial, [150] examined the effects of oral B. subtilis E20 (104, 106, and 108 CFU g-1) administration on growth performance parameters of orange-spotted grouper. The results showed that feeding efficiency and weight gain were significantly increased by dietary probiotics [150,151]. [152] fed grass carp with a different strain (B. subtilis Ch9). After 56 days of feeding, fish treated with varying amounts of probiotics (1.0×109, 3.0×109, and 5.0×109 CFU kg-1) showed significantly higher weight increase, SGR, and FCR. Additionally, the authors observed that fish administered probiotics had improved intestinal morphology and a notable increase in the activity of digestive enzymes. In a comparable way, feeding olive flounder with B. subtilis boosted their ultimate weight, FCR, and protein efficiency ratio much more than feeding them the control [153]. Moreover, adding 5×106 CFU g-1 of B. subtilis to the diet of Nile tilapia considerably enhanced growth performance metrics [154]. When Nile tilapia (65 5 g) were fed B subtilis for two months, the fish's weight gain and survival rate increased dramatically [155]. Recently, [156] revealed in a two-month supplementation of B. subtilis efficiently improving growth, intestinal probiotic recovery and enzyme activities in O. niloticus. Moreover, in feeding Larimichthys crocea supplemented diets of B. subtilis for averagely two months enhanced growth was observed [157]. Oral feeding of Nile tilapia for 10 weeks showed similar outcomes [158]. [158] revealed that high dosage of Bacillus improve wellbeing of fish intestines compared to the control and low dosages. However, [159] In a study with catfish (Pangasius hypophthalmus), smaller dosages of B. licheniformis were more beneficial on growth than higher dosages, showing the host and probiotic specific species [160].
Moreover, aside B. subtilis, investigations of other Bacillus species have been undertaken to evaluate effects on the growth factors of fish. [64] investigated the effects of B. circulans extracted from Catla catla's gut as a feed supplement. After a two-month feeding experiment, the scientists found that fish fed bacillus-supplemented diets had improved growth performance measures. Additionally, [161] gave a distinct dosage of dual types of Bacilli in an orange-spotted grouper diet throughout a 30-day study. Probiotic strain from orange-spotted grouper's intestines was provided by the authors. These probionts, in contrast to other research using Bacilli probiotics, were incapable of altering development metrics. On the contrary, the impact of B. amyloliquifaciens were assessed in a three-month diet treatment with O. niloticus [162]. The findings showed that B. amyloliquifaciens had no meaningful effect on O. niloticus. A two-month dietary delivery of B. amyloliquefaciens in turbot showed a moderate boost in fish growth metrics, but enzymes from the gut showed a considerable increase compared to the control fish [40]. In another study it was realized that silver carp given B. latrospores and B. licheniformis saw a superior growth performance compared to the control [163]. Moreover, a two months’ oral application of bacillus improved growth of Athlantic salmon in recirculatory aquaculture [164]. In another study, the researcher observed a combined multi-species administration of Bacillus showed beneficial effects on fish compared to single administrations. Other research agreed [165] when B. subtilis in combination with P. aeruginosa + L. plantarum was employed in Labeo rohita with a good outcome in enhancing of growth parameters than using only B. subtilis. On a contrary to a study conducted [166] no significant difference was observed when Tilapia was fed a single form or combination with B. velezensis for a month.
Besides the various ways of administering Bacillus a few has studies has evaluated the effects of bath applications. For example, in an investigation with O. niloticus, [167] used Bacillus coagulans as water probiotic. After a month, the researchers found that fish in the probiotic water bath improved in their growth metrics. Furthermore, 56-day administration of B. subtilis as a water addition resulted in a significant increase in overall weight and specific growth weight in the probiotic-treated fish. [167].

Table 2.
Effects of Bacillus spp on growth of fin fishes.
| Bacillus spp. | Fish species | Initial Weight | Application of Bacillus spp | Observation on Growth | Conclusion | References |
|---|---|---|---|---|---|---|
| B. subtilis | Oreochromis niloticus | 14.82 ± 0.42 g | Dietary application for 50 days | Fish given probiotics showed noticeably improved growth results as compared to the control group, and their digestive enzyme activity also increased noticeably | In tilapia housed in a biofloc system, B. subtilis efficiently increases fish output, immunity, and defense against LPS-induced damages | [168] |
| (a mix of B. subtilis and B. licheniformis) | Oreochromis niloticus | 53.01 ± 1.0 g | Dietary application for 50 days | All probiotic BS enriched groups showed improved weight gain, specific growth rate, and feed conversion ratio | Application of probiotic BS at 10 gkg-1 (BS10) may be taken into consideration to enhance tilapia farming growth | [169] |
| B. amyloliquefaciens and B. pumilus isolated from striped catfish (P. hypophthalmus) | (Pangasianodon hypophthalmus) | 25.2 ± 1.3 g | dietary supplementation | Growth improvement in fish given a combination of probiotics | Thus, striped catfish health and growth rate can be enhanced by dietary supplementation of a blend of B. amyloliquefaciens and B. pumilus at 5 × 108 CFU g−1 | [170] |
| Two species of Bacillus (B. licheniformis and B. subtilis) | Asian Sea Bass, L. calcarifer | 1.5 ± 0.2 g | During 8 weeks of dietary supplementaion | Compared to Asian sea bass fed the basal food (control), those supplemented with probiotic Bacillus (B. licheniformis and B. subtilis) had noticeably improved growth. | Considering that the optimal outcome is obtained when 1 ×106 CFU g−1 of Bacillus is supplemented in the diet | [160] |
| (B. subtilis and B. licheniformis | Oreochromis niloticus | mean weight of around 150 g | 30 days of dietary supplementation | The fish fed with diets containing 0.04% and 0.08% of probiotics presented higher weight gain than the control group | The establishment of a beneficial microorganism population may improve host health | [102] |
| Mixed probiotic containing (B. licheniformis and B. subtilis and Ferroin solution) | kutum, (Rutilus frisii kutum) | 0.4 ± 0.1 g | 60 days of supplemented diets in ratios | Fish receiving diets supplemented with probiotics and Ferroin solution showed significantly better growth than those fed the basal diet (control) | These results indicate that the combination of probiotic and Ferroin solution represents an effective dietary supplement for growth performance | [171] |
| B. subtilis | grass carp, Ctenopharyngodon idella | 50 ± 2.5 g | 56 days dietary feeding | Significantly higher SGR and lower FCR than those fed the control diet | An optimum dose of B. subtilis Ch9 could induce digestive and potentially promote the digestion and absorption of nutrients, as well as improve the growth performance of grass carp significantly. | [152] |
| B. subtilis | Oreochromis niloticus | 16.5 ± 0.2 g | 21-d growth trial as feed additives | There was no discernible difference in growth performance when any probiotic-added diet was used | Given the short study period, these outcomes are not shocking | [99] |
| B. pumilus | Oreochromis niloticus | 3.62 ± 0.06 g | 84 days of dietary supplementation | The study showed fish fed a pro-enzyme diet had improved feed consumption and growth performance | Pro-enzyme supplementation enhanced the growth performance | [172] |
| B. circulans | Catla catla | 6.48 ± 0.43 g | Feeding of diets supplemented for 60 day | Compared to other treatments, C. catla given feed probiotic showed higher growth performance in terms of live weight gain and specific growth rate | The study's findings support the use of Bacillus circulans PB 7, a probiotic, for improved growth and appropriate nutrient use | [64] |
| B. licheniformis | triangular bream (Megalobrama terminalis) | 30.5 ± 0.5 g | 8-week feeding trial, | Fish fed B. licheniformis showed a considerably higher growth parameters | B. licheniformis, either by itself or in combination, can greatly enhance triangular bream growth performance | [173] |
| B. clausii | Japanese flounder Paralichthys olivaceus | Average weight of 21 g | Dietary supplementation for 56 days | Fish fed B. clausii gained more weight than the control group | The growth performance and health benefits of the Japanese flounder were enhanced by B. clausii | [67] |
4.1.3. Bacillus spp Effects on Liver Health of Fin Fishes
Recently, there has been speculation that the accumulation of pollutants may be the reason behind gall syndrome and liver enlargement in many farmed fish [174]. A rise in the levels of the enzymes aspartate transaminase (AST) and alanine transaminase (ALT), which interconvert amino acids with other metabolic intermediates and are involved in a variety of biochemical events in metabolism, can be an indication of tissue damage, as in the case of chronic liver disease [175,176]. According to [177], AST and ALT are sensitive biomarkers used in the detection of liver damage because they are cytoplasmic in origin and are released into circulation (blood) after cellular injury. In vertebrates, AST is present in both mitochondrial and cytoplasmic forms; the tissues containing the greatest amounts of AST are the heart, liver, muscle, and kidney, in that order [175]. Fish rely heavily on the activity of the enzymes AST and ALT because they can be used to identify tissue damage caused by toxicants (found in feed given or in the environment) [174]. Bacillus spp. have been found to influence AST and ALT in fish; fish fed a diet enriched with probiotic B. licheniformis and B. subtilis, for example, had decreased AST and ALT [142]. On the contrary, in a study [178], no changes were estimated in the levels of glucose, hepatic (alanine aminotransferase (ALT) and aspartate aminotransferase (AST), and renal biomarkers (Urea, Creatinine, and Uric acid) in the serum of fish-fed bacillus supplemented diets. In another research [179], assessing the effects of bacillus on nile tilapia it was observed after 12 weeks that levels of aspartate aminotransferase and alanine aminotransferase reduced compared to the control. After challenge test, aspartate transaminase (AST), alanine transaminase (ALT) activity, and liver malondialdehyde level increased significantly in control groups; however, level of these parameters were considerably lower in fish fed with probiotic supplemented diets when B. amyloliquefaciens CCF7 was added to L. rohita [180].
4.1.4. Bacillus Species-Related Effects on Finfish Gene Expression
It has been reported that certain Bacillus species increase the expression of genes linked to growth metabolism, inflammation, digestion, the cytoskeleton, transport of proteins, junction complex protein encoding genes, and antioxidant genes [181,182]. There aren't as many studies on Bacillus and the expression of the aforementioned genes as there are on the bacteria's role in regulating growth, feed utilization, etc. [183]. By reducing the expression of the ++HSP70 gene, a combination of Bacillus species has been shown to lessen the severity of cellular stress in sea bream larvae, improving the fish's tolerance to rearing conditions [184]. In a different study, it was found that fish fed Bacillus diets had significantly higher gene expression levels of heat shock protein 70 (HSP70), interleukin (IL-1β), interferon-gamma (IFN-γ), and tumour necrosis factor (TNF-α) than fish fed the control diet [100]. The liver tissue showed significant modulation of growth-related genes, such as insulin-like growth factor genes (IGF-1 and IGF-2) and growth hormone receptor genes (GHR-1 and GHR-2) in an investigation on the potential of Bacillus on Tilapia. The IGF-1 gene was significantly more expressed in muscle tissue [185]. Other evidence exists, including the altered expression of mucosal genes in gilthead Sea breams [186], immune-related genes expressed in the head kidney of Carassius auratus [187], pro-inflammatory cytokines (IL-8 and IL-1β), TLR5, and TGF-β1 expressed in the intestine and head kidney of E. coioides [188], and pro-inflammatory cytokines expressed in the intestine of Nile tilapia [100] following probiotic Bacillus administration.
4.2. Effects of Bacillus spp on Shellfish
Bacillus is one of the many potential probiotics in shrimp aquaculture that has produced positive outcomes [189]. Bacillus has been utilized to boost immunological response, treat illness, and increase growth performance [156,190]. Furthermore, it’s known that Bacillus species secrete a variety of extracellular materials and antimicrobial peptides that enhance water quality, aid in the digestion and absorption of feed, strengthen shrimp immunity, encourage growth and reproduction, and increase shrimp's ability to withstand pathogenic microorganisms [191,192,193,194]. This section explains in detail how Bacillus, a probiotic, affects disease resistance, immune responses, and growth performance in cultured shrimp.
4.2.1. Enhancement of Immune Response Resistance to Diseases in Shell Fishes
The invertebrate's only defense against harmful pathogens is its non-specific immune system, which has been shown to be enhanced by probiotics [195]. According to [196], Bacillus sp. (strain S11) can be used to protect tiger shrimp (P. monodon) from disease by stimulating their humoral and cellular immune systems. Transduction signaling molecules produced by Bacillus can warn the immune system about pathogenic agent attacks [197] and certain diseases like edema of the gut [198]. Numerous research studies have demonstrated that probiotics, whether commercial or derived from other sources, can enhance the cellular and humoral components of the innate immune system in a variety of fish and shellfish species [196,197,199,200,201,202,203,204,205,206].
Probiotics that increase shrimp immunity to diseases have garnered a lot of attention during the last ten years [18,207]. It is believed that Bacillus species are harmless probiotic bacteria that improve the wellbeing of their hosts by boosting natural defenses and strengthening the body's defenses against harmful microbial infections [16,27]. Prior research utilizing Bacillus indicated that the probiotic bacteria could augment and confer resistance in shrimp species (P. monodon) upon encountering V. harveyi [208]. But the mode of action wasn't fully understood until a few authors began explaining the underlying mechanism, which included the immune response and whether or not invertebrates, like shrimps, could mount an immune response that shared some characteristics with vertebrates' immune systems. [196] noted that Bacillus probiotics, such as Bacillus S11, enhance survival and disease resistance against V. harveyi by inducing phenoloxidase, phagocytosis, and antimicrobial activity in P. monodon hemolymph, thereby stimulating the immune response. Furthermore, it has been observed that diets supplemented with Bacillus to shrimp species enhanced the natural defensive system, increasing their resistance to infections [209]. For example, it has been reported that B. subtilis E20 increases the resistance of white shrimp (L. vannamei) to V. alginolyticus by increasing phenoloxidase and phagocytic activity. Additionally, the administering of Bacillus to white leg shrimp enhances the immune response (phagocytosis, phenoloxidase, etc.) shielding this specie against diseases (WSSV) [210,211]. The amount of Vibrio and total viable counts of bacteria in the shrimp gut were reduced when L. vannamei was fed Bacillus probiotics [211]. B.fusiformis also administered to L. vannamei daily or at intervals increased survival and prevented disease as immune system was boosted.[212]. On the other hand, B. subtilis increased Litopenaeus vannamei's antioxidant capacity and phenoloxidase activities [213]. Numerous immunological factors, such as lysozyme, respiratory burst, phenoloxidase, and bactericidal activity in L. vanamei, were increased by B. cereus [214]. The improved immune status was validated by the increased survival rate of the treated shrimp following V. harveyi challenge. Similar to this, feed supplemented with Bacillus spp. at 1 × 107 and 1 × 109 CFU/kg diet and fed continuously to shrimp for 5 weeks effectively enhanced growth, as demonstrated by feed conversion ratios, average daily growth, final weight gain, and specific growth rates. Furthermore, this probiotic improved the expression of the prophenoloxidase, lysozyme, and anti-lipopolysaccharide factor genes and markedly increased immune responses through phagocytic activity and clearance efficiency [209].
A two-month trial was conducted to assess the impact of different feed probiotic supplements on young L. vannamei. The lysozyme activity of shrimp fed probiotics and over-the-counter diets demonstrated significant improvement when compared to the control diet. Furthermore, compared to shrimp fed control and over-the-counter diets, immune-related gene expression in shrimp fed BS8, PP8, and LL8 diets was found to be significantly higher [215].
In addition to reducing Vibrio population abundance, B. aryabhattai supplementation to Pacific white shrimp (Litopenaeus vannamei) also changed the bacterial community within the shrimp's gastrointestinal tract. B. aryabhattai was found to activate antioxidant and innate immune responses in shrimp [216]. Furthermore, B. subtilis E20 has been shown to enhance a number of immune response factors, in addition to survivability and resilience to stress in white shrimp, which includes salinity, temperature, and nitrite-N [217].
When L. vannamei was fed B. subtilis orally, it was observed that they were more resilient to ammonia oxygen shortage and had a higher survival rate than the control group [218]. This could be because the B. subtilis probiotic converted and transformed ammonia. [219] assessed how Bacillus protected juvenile Marsupenaeus japonicus against temperature stress by acting as an immunomodulator and protective agent. In a different investigation, demonstrating that adding Bacillus (such as B. subtilis and B. licheniformis) to L. vannamei culture water improved immune response, as evidenced by the increase in prophenoloxidase (ProPO), peroxinectin (PE), and other markers [220,221]. Additionally, probiotic bacteria increase juvenile white shrimps' resistance to disease from the pathogenic V. harveyi [220,222]. A strain of Bacillus, isolated from the gut of Fenneropenaeus chinensis, was shown in 2016 by [192] to enhance defense mechanisms, protecting L. vannamei against infection by the white spot syndrome virus. Some authors have reported an intriguing discovery: Some few species of Bacillus have been found to contain PHB helps aquatic animals by boosting their immune systems, both specific and non-specific [223,224]. Additionally, a small number of researchers indicated Bacillus for giving immune abilities to shrimp species against pathogenic microorganisms [225]. According to [193], P. mondon postlarvae can be stimulated to mount an innate immune response by a Bacillus strain containing PHB. More recently, shrimp fed Bacillus alone or in combination, for 21 days at 28ºC have been shown to improve the natural defense and stimulate appetite in V. harveyi [226].
4.2.2. Modifying Digestive Enzymes and Encouraging Shellfish Development
It has been discovered that Bacillus species offer encouraging development and survival in a system for raising L. vannamei shrimp without water exchange [227]. For aquaculture species, probiotics can therefore be thought of as growth promoters in addition to their many other advantages [7,228].
Probiotic-based microbiota manipulation in shrimp aquaculture has been shown to be an effective strategy for controlling or inhibiting pathogenic bacteria, enhancing host immune response to pathogenic infection or physical stress, and improving growth performance and digestive enzyme activity [215,229]. One anticipated benefit of Bacillus as probiotics is that it directly promotes development by inducing digestive enzymes, such as amylase and protease, which in turn increases the host's natural digestive enzyme activity [24,230]. It has been noted that Bacillus sp. improves P. monodon postlarvae growth and survival without requiring water exchange, and it helps the host feed shrimp, especially by supplying fatty acids and vitamins [191,197,231]. Bacillus probiotics are said to enhance the growth of shrimp and the activity of digestive enzymes [189,220]. The administration of B. subtilis to L. vannamei was found to enhance the activity of the digestive enzymes protease and amylase, which in turn promotes the growth of juvenile shrimp [189,220]. [232] and [221] evaluated the impact of commercial Bacillus probiotics on L. vannamei rearing performance in terms of growth, bacterial count, feed efficiency, and body composition. According to the study, L. vannamei experimental tanks supplemented with Bacillus had a much greater influence on growth than the control group. Furthermore, the probiotic facilitated an increase in the amount of feed that L. vannamei postlarvae consumed [233]. It was also reported that the addition of Bacillus to in white shrimp (L. vannamei) water improved growth features significantly. Moreover, isolated bacteria from shrimp showed promising effects on growth of L. vannamei [234] and [192]. The outcomes demonstrated that Bacillus probiotics enhanced growth performance and decreased the risks associated with stressful conditions or factors in shrimp culture. Probiotic species selection is a significant factor, as evidenced by a study by [235] that found that B. subtilis given to L. vannamei orally outgrew both non probiotic diets and B. megaterium groups in terms of weight gain and food-conversion ratio. When applied through diets alone or in combination, both species of Bacillus improved the growth of shrimp and improved conditions of their environment [226].
Under ideal circumstances, Bacillus, which are frequently obtained from shrimp has been demonstrated to hoard PHB in the range of 11% to more than 50% on cell dry weight [236]. Bacillus bacteria isolated shrimp culture ponds has been seen to hoard PHB which improves the growth and reducing stress in P. monodon [225].
Table 3.
Effects of Bacillus spp on growth of Shell fishes.
| Bacillus spp. | Fish species | Initial Weight | Application of Bacillus spp | Observation on Growth | Conclusion | References |
|---|---|---|---|---|---|---|
| B. cereus | L. vannamei | 4.55 g | 28 days dietary administration | Findings demonstrated that following a 28-day probiotic feeding regimen, shrimp growth was enhanced in comparison to the control group | The study showed that probiotics might be used to enhance white shrimp growth | [237] |
| B. licheniformis | Haliotis discus hannai Ino | 4.17 ± 0.32 | 8-week culture experiment | In comparison to the control group, the supplemented diet group exhibited a significantly greater specific growth rate of shell length, food intake, and food conversion rate | The study suggests B. licheniformis diets enhanced abalones food intake and growth. | [238] |
| B. amyloliquefaciens | H. discus hannai | 4.28 ± 0.23 g | 8-week culture experiment | Diets supplemented with Bacillus showed significantly increased body weight, specific growth rate, and food conversion efficiency compared to the control group | The experimental meal that contained 105 CFU/g of B. amyloliquefaciens promoted abalone growth and food intake | [239] |
| B. subtilis | (L. vannamei) | 12.03 ± 2.76 g | 1 month feeding of supplemented diets | Effective enhancement of growth | A notable rise in growth metrics suggested that Bacillus was the best option | [209] |
| B. subtilis and B. licheniformis | (L. vannamei) | 1 ± 0.1 g | Dietary supplementation for a month | Shrimp's specific growth rate was much higher in the supplemented diet than in the (control) treatment, and the treated groups' ultimate growth was comparable | Shrimp growth parameters were effectively increased by the Bacilli probiotic combination | [240] |
| B. licheniformis | H. discus hannai | 4.91 ± 0.34 g | 70 days of beimg fed dietary supplemented diets | The abalone in the supplemented diet exhibited a considerably greater feed conversion efficiency and specific growth rate compared to the control group | Maximum growth benefits of the probiotic were noted | [241] |
| B. subtilis | (L. vannamei) | 0.67 ± 0.06g | Dietary application for 8 weeks | The final weight, weight gain, and digestive enzyme activity of the shrimp fed Bacillus diets were significantly higher than those of the untreated control group | B. subtilis treatment can enhance shrimp growth performance | [189] |
| B. cereus | (P. monodon) | 0.204 ± 0.004 g | Dietary appliacation for 90 days | The group that received a supplemented meals achieved a higher FCR of 1.27 ± 0.081, a maximum production of 10.45 ± 0.275 g, and an SGR of 4.40 ± 0.179%, according to the overall growth responses | B. cereus, a lyophilized probiotic, was effective in boosting shrimp growth when added to feed at a concentration of 0.4%/100 g | [191] |
| B. pumilus | (Macrobrachium rosenbergii) | 1.81 ± 0.01 g | 2 months dietary application | The experimental groups exhibited significantly increased final weight, weight gain rate (WGR), and specific growth rate (SGR) compared to the control group | Adding B. pumilus to feed at a dosage of 1 × 108 CFU/g would enhance M. rosenbergii's growth and digestive enzymes | [242] |
4.2.3. Enhancing Shrimp Culture Water Quality Factors
In a study, [243] showed that using native Bacillus species in the pond waters used for P. monodon culture rearing could preserve the water quality and reduce Vibrio count. When [244] examined the impact two available products of bacillus on P. monodon, they found that ponds treated with a combination of Sacchromyces and Bacillus spp. exhibited relatively lower concentrations of biological oxygen demand, TAN, nitrate, and COD in beginning stages when compared to groups treated with other additives. Utilizing a blend of probiotics including Bacillus enhanced density of advantageous bacteria, dissolved oxygen content, and water transparency in the existing water environment of L. vannamei ponds. However, it decreased the levels of total inorganic nitrogen, phosphate, and COD [245]. Additionally, the pH value of the probiotic-treated group did not considerably change, but the control group's pH level significantly increased following a notable decline during the trial's first 40 days. Additionally, the application of Bacillus greatly increased dissolved oxygen while lowering COD [246,247]. When [248] added a Bacillus sp. probiotic to shrimp pond aerators, they observed a decrease in COD along with an increase in final production. In hatcheries of P. monodon and L. vannamei, the use of a combined strains of commercial Bacillus, was able to lower the density of Vibrio bacteria in the water column and improve the water quality [249]. In a study by [250], M. resenbergii was cultured and Bacillus was employed to treat the water. The results showed lower values of ammonia, nitrite, and pH over the course of the 60-day cultivation period when compared to the control. The ability of some native strains of Bacillus to reduce TAN by bioremediation was assessed after they were extracted from sea water and soil samples [231]. Additionally, B. pumilus was assessed on L. vannamei in a recirculatory system and it was realized phytoplankton and bacterial concentration was regulated [251]. The once-weekly application of an indigenous probiotic B. subtilis improved the water quality values two days after the bacterial community of L. vannamei culture was raised. As a result, there was an increase in COD and chlorophyll density and a decrease in pH, nitrite, water transparency, and soluble reactive phosphorus [252]. The pH, ammonia, and nitrite levels of the water are improved when encapsulated probiotics are added to the water column. This appears to favor the good bacteria present in shrimp and culture water. This was corroborated by [232], who found that when two mixtures of Bacillus species were used in the rearing water of L. vannamei as microencapsulated probiotics using Artemia, the levels of pH, ammonia, and nitrite in the treated shrimp were lower than in the controls. Moreover, compared to controls, the treated groups' shrimp and water had greater densities of Bacillus count and total heterotrophic bacteria. Additionally, some water parameters (hardness, pH and Alkalinity) increased with a reduction in other parameters when combined strains of Bacillus was added into water of L. vannamei. [220] in his study showed incorporating Bacillus resulted in significant improvement in water quality parameters. These parameters included salinity, bicarbonates, carbonates, Mg hardness, total alkalinity, total hardness, Ca hardness, pH and ammonia [227].
Biological methods are widely used to reduce the amount of harmful chemicals in fish farming, and the use of certain bacteria, like Bacillus, has been shown to be helpful in the process of converting organic wastes that are hazardous to the environment into compounds that are safe for human consumption [254]. By offering an ideal environment, Bacillus probiotics can improve the rearing water quality parameters [255]. This would improve the health status of aquatic animals as their environment have massive impacts on their well-being. Additionally, through bacterial competition, Bacillus probiotics reduce the density of potentially harmful microbes, reducing stress in culture conditions and enhancing the immunological-physiological balance of animals [6]. It is important to note, nevertheless, that Bacillus species are not particularly effective at removing ammonia, and no Bacillus strain has ever been shown to remove more than 90% of ammonia [256,257]. Therefore, a nitrifying probiotic could be a mix of a high cleaner ammonia bacterium and a Bacillus species could be employed to remove ammonia and nitrite from aquaculture rearing waters at the same time, as [226] demonstrated a blend of Nitrosomonas sp. and Bacillus sp. proved most effective in eliminating large amounts of TAN. A three weeks’ oral use of B. licheniformis and B. flexus on L. vannamei showed a substantial decrease in some water parameters in shrimp rearing water, suggesting that oral application may improve the water quality conditions, even though the majority of researchers incorporating Bacillus did not measure the parameters of water [226].
5. Pathogenic Bacillus in Fin and Shellfish
There are few studies on how Bacillus pathogenesis affects fish and shellfish [258]. B. mycoides was found by [259] to be the cause of a superficial epizootic disease in commercial channel catfish in Alabama, USA. The dorsal surface of the infected fish showed pale patches or ulcers, focal necrosis of the epaxial muscle, and chains of Gram-positive bacilli that were determined to be B. mycoides. Upon administering 1.6 × 104 cfu/fish of the isolated bacterium subcutaneously or intramuscularly to healthy catfish, lesions resembling those observed in a spontaneous epizootic occurred. The toxins produced by these bacilli bacteria could be the reason for the fish's congestion and lack of bleeding. In 2000, [260] discovered a novel bacterial white spot syndrome in Malaysian shrimp farming that was brought on by B. subtilis in cultivated tiger shrimp. The infected shrimp exhibited white spots resembling those of the white spot viral disease (WSVD), but they continued to grow and were active, with no appreciable death or disability. Evaluations using microscopy showed that the epicuticle's cuticle had degenerated and become discolored, and that underlying cuticular layers had been found. It was proposed that frequent use of B. subtilis probiotics in shrimp ponds could be linked to this kind of illness. Enzymes of protease, amylase, glucanase, and lipase can be excreted by B. subtilis [261] indicating the probiotics ability to cause dissolution in the shrimp epidermis makeup [262].
In an experimental study by [263] expression of B. cereus hemolysin II in B. subtilis rendered the bacterium being pathogenic for the crustacean Daphnia magna when it was challenged with the expressed B. subtilis at 04-106 cfu L-1 at water temperature 20 ± 5˚C. The lethal concentrations 50% (LC50) on the fifth day of the experiment for the expressed B. subtilis and B. cereus were 5.4 × 105 and 4.5 × 105 cfu mL-1, respectively.
More recently, in 2015, reports of a novel bacterial white patch disease caused by B. cereus were received from various L. vannamei aquaculture sites in India [264]. The disease resulted in a persistent morbidity and mortality rate. The afflicted shrimps displayed necrosis, pale white muscles, whitish blue pigmentation, loss of appetite, and white opaque spots in their carapace. It can also release enzymes such as lipase, glucanase, amylase, and protease [265], Therefore, it could potentially be able to enter inhibited aquatic organisms under severe environmental conditions that are conducive to expressing the bacterium's virulence genes [266].
6. Fish and Shellfish Safety with Bacillus
The possible use of Bacillus as probiotics in aquatic animal feed or as a supplement to rearing water that enters the human food chain is a significant public health concern [16]. However, there is currently no information on the potential for Bacillus used in aquatic animals to contaminate food intended for human consumption [267]. The spread of antibiotic resistance brought on by transmissible antibiotic resistance genes present in certain probiotic bacteria is one of the main risk factors [268]. AFurthermore, the risks of probiotic infections and the existence of enterotoxins and emetic toxins in probiotic bacteria must be closely monitored by the aquaculture sector [269]. The majority of published information on Bacillus supplementation in fish farming, however, relates to their effectiveness rather than their safety [270]. Furthermore, it's critical to remember that safety evaluations and data regarding a specific Bacillus strain probiotic shouldn't be compared to those of similar bacterial strains; rather, each probiotic's safety and risk assessment should be assessed separately [271]. The extent to which a particular probiotic can be harmful depends on how susceptible the target aquatic animal-such as freshly hatched fish larvae-is to immune-physiological conditions [272]. As a result, it's possible that a probiotic strain of Bacillus that is deemed harmless in some circumstances is harmful in others. It is possible to claim that, similar to antibiotics, there is no particular probiotic that is 100% safe [273].
Another significant safety and quality concern with probiotics is the potential presence of undesirable or contaminated microorganisms or the harmful compounds they produce. Occasionally, these contaminants could pose a greater risk than the particular quality of the probiotics [274]. Probiotics, including those found in Bacillus species, are currently regarded as safe when added to aquatic animal feed or water cultures in recent reports [27,228,275]. Nevertheless, some bacterial probiotic species increase the likelihood that certain pathogenic bacteria may develop antibiotic resistance or that their enterotoxins will be produced [276].
7. Conclusions and Future Perspectives
It is concerning how important Bacillus is to the rearing of fin and shellfish. This summary makes it clear that Bacillus has a lot of potential to support the continued cultivating of fish and shellfish by preserving the overall health of cultured fish, which includes improving growth, feed utilization, immune response, defense against infections, specifically bacterial infections, and water quality. But in order to advance studies and the use of probiotic Bacillus in fin and shellfish culture, we recommend the following. Future studies on the effects of probiotics should include high throughput assays for transcriptome and proteome analysis in addition to the several genome-sequencing technologies already in use. Furthermore, a detailed documentation of the transcriptome and proteome profiling of the gut microbiota is necessary to understand the various mechanisms of action of various probiotic organisms. Therefore, future research on probiotics should be highly prioritized, and a standard criterion for evaluating their impact on fish nutrition and health should include analysis at the molecular level.
Author Contributions
Samuel Ayeh Osei: Writing-Original Draft, Review of Manuscript. Christian Larbi Ayisi: Writing-Original Draft, Conceptualization, Review of Manuscript.
Conflicts of Interest
The Authors declare no Conflict of Interest.
References
- Buchmann, K. Control of parasitic diseases in aquaculture. Parasitology 2022, 149, 1985–1997. [Google Scholar] [CrossRef] [PubMed]
- Arsène, M.M.; Davares, A.K.; Andreevna, S.L.; Vladimirovich, E.A.; Carime, B.Z.; Marouf, R.; Khelifi, I. The use of probiotics in animal feeding for safe production and as potential alternatives to antibiotics. Vet. World 2021, 14, 319. [Google Scholar] [CrossRef] [PubMed]
- Scanes, C.G. Animal agriculture: Livestock, poultry, and fish aquaculture. In Animals and Human Society; Academic Press: Cambridge, MA, USA, 2018; pp. 133–179. [Google Scholar]
- Chan, C.Y.; Tran, N.; Cheong, K.C.; Sulser, T.B.; Cohen, P.J.; Wiebe, K.; Nasr-Allah, A. M, The future of fish in Africa: Employment and investment opportunities. PloS ONE 2021, 16, e0261615. [Google Scholar] [CrossRef] [PubMed]
- Flores-Kossack, C.; Montero, R.; Köllner, B.; Maisey, K. Chilean aquaculture and the new challenges: Pathogens, immune response, vaccination and fish diversification. Fish Shellfish. Immunol. 2020, 98, 52–67. [Google Scholar] [CrossRef] [PubMed]
- Ringø, E, Probiotics in shellfish aquaculture. Aquac. Fish. 2020, 5, 1–27. [CrossRef]
- El-Saadony, M.T.; Alagawany, M.; Patra, A.K.; Kar, I.; Tiwari, R.; Dawood, M.A.; Abdel-Latif, H.M. The functionality of probiotics in aquaculture: An overview. Fish Shellfish. Immunol. 2021, 117, 36–52. [Google Scholar]
- Luise, D.; Bosi, P.; Raff, L.; Amatucci, L.; Virdis, S.; Trevisi, P. Bacillus spp. probiotic strains as a potential tool for limiting the use of antibiotics, and improving the growth and health of pigs and chickens. Frontiers in Microbiology 2022, 13, 801827. [Google Scholar]
- Panase, A.; Thirabunyanon, M.; Promya, J. ; Chitmanat, C, Influences of Bacillus subtilis and fructooligosaccharide on growth performances, immune responses, and disease resistance of Nile tilapia, Oreochromis niloticus. Frontiers in Veterinary Science 2023, 9, 1094681. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, A.; Hussain, Z.; Akram, A.M.; Alorabi, M.; Sarwar, N.; Rehman, R.A.; Alkafafy, M. Impact of Bacillus subtilis supplemented feed on growth and biochemical constituents in Labeo rohita fingerlings. Journal of King Saud University-Science 2021, 33, 101668. [Google Scholar]
- Lorenzo, J.M.; Munekata, P.E.; Dominguez, R.; Pateiro, M.; Saraiva, J.A. ; Franco, D, Main groups of microorganisms of relevance for food safety and stability: General aspects and overall description. In Innovative Technologies for Food Preservation. Academic Press: Cambridge, MA, USA, 2018; pp. 53-107.
- Ehling-Schulz, M.; Lereclus, D.; Koehler, T.M. The Bacillus cereus group: Bacillus species with pathogenic potential. Microbiol. Spectr. 2019, 7, 10–1128. [Google Scholar] [CrossRef]
- Bahaddad, S.A.; Almalki, M.H.; Alghamdi, O.A.; Sohrab, S.S.; Yasir, M.; Azhar, E.I. ; Chouayekh, H, Bacillus Species as direct-fed microbial antibiotic alternatives for monogastric production. Probiotics Antimicrob. Proteins 2023, 15, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Tran, C.; Cock, I.E.; Chen, X. ; Feng, Y, Antimicrobial Bacillus: metabolites and their mode of action. Antibiotics 2022, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Butt, U.D.; Lin, N.; Akhter, N.; Siddiqui, T.; Li, S. ; Wu, B, Overview of the latest developments in the role of probiotics, prebiotics and synbiotics in shrimp aquaculture. Fish Shellfish. Immunol. 2021, 114, 263–281. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Hong, B.; Luo, K.; Li, Y.; Fu, H. ; Wang, J, Isolation of Bacillus subtilis and Bacillus pumilus with Anti-Vibrio parahaemolyticus Activity and Identification of the Anti-Vibrio parahaemolyticus Substance. Microorganisms 2023, 11, 1667. [Google Scholar] [CrossRef] [PubMed]
- Ogbuewu, I.P.; Mabelebele, M.; Sebola, N.A. ; Mbajiorgu, C, Bacillus probiotics as alternatives to in-feed antibiotics and its influence on growth, serum chemistry, antioxidant status, intestinal histomorphology, and lesion scores in disease-challenged broiler chickens. Front. Vet. Sci. 2022, 9, 876725. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.K.; Roy, T.; Mondal, S. ; Ringø, E, Identification of gut-associated amylase, cellulase and protease-producing bacteria in three species of Indian major carps. Aquac. Res. 2010, 41, 1462–1469. [Google Scholar]
- Kim, D.H.; Kim, D.Y. Microbial diversity in the intestine of olive flounder (Paralichthys olivaceus). Aquaculture 2013, 414, 103–108. [Google Scholar] [CrossRef]
- Jlidi, M.; Akremi, I.; Ibrahim, A.H.; Brabra, W.; Ali, M.B.; Ali, M. B, Probiotic properties of Bacillus strains isolated from the gastrointestinal tract against pathogenic Vibriosis. Front. Mar. Sci. 2022, 9, 884244. [Google Scholar] [CrossRef]
- Nayak, S. K, Multifaceted applications of probiotic Bacillus species in aquaculture with special reference to Bacillus subtilis. Rev. Aquac. 2021, 13, 862–906. [Google Scholar] [CrossRef]
- Shivaji, S.; Chaturvedi, P.; Suresh, K.; Reddy, G.S.N.; Dutt, C.B.S.; Wainwright, M. . & Bhargava, P. M.; Bacillus aerius sp. nov.; Bacillus aerophilus sp. nov.; Bacillus stratosphericus sp. nov. and Bacillus altitudinis sp. nov.; isolated from cryogenic tubes used for collecting air samples from high altitudes. Int. J. Syst. Evol. Microbiol. 2006, 56, 1465–1473. [Google Scholar]
- Dunlap, C. A, The status of the species Bacillus aerius. Request for an Opinion. Int. J. Syst. Evol. Microbiol. 2015, 65, 2341–2341. [Google Scholar] [CrossRef] [PubMed]
- Zeigler, D.R.; Perkins, J.B. The genus bacillus. In Practical handbook of microbiology; CRC Press: Boca Raton, FL, USA, 2021; pp. 249–278. [Google Scholar]
- Dutta, D.; Banerjee, S.; Mukherjee, A. ; Ghosh, K, Selection and probiotic characterization of exoenzyme-producing bacteria isolated from the gut of Catla catla (Actinopterygii: Cypriniformes: Cyprinidae). Acta Ichthyologica et Piscatoria 2015.
- Meidong, R.; Khotchanalekha, K.; Doolgindachbaporn, S.; Nagasawa, T.; Nakao, M.; Sakai, K.; Tongpim, S. Evaluation of probiotic Bacillus aerius B81e isolated from healthy hybrid catfish on growth, disease resistance and innate immunity of Pla-mong Pangasius bocourti. Fish Shellfish. Immunol. 2018, 73, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Meidong, R.; Nakao, M.; Sakai, K. ; Tongpim, S, Lactobacillus paraplantarum L34b-2 derived from fermented food improves the growth, disease resistance and innate immunity in Pangasius bocourti. Aquaculture 2021, 531, 735878. [Google Scholar] [CrossRef]
- Mukherjee, A.; Dutta, D.; Banerjee, S.; Ringø, E.; Breines, E.M.; Hareide, E. . & Ghosh, K, Potential probiotics from Indian major carp, Cirrhinus mrigala. Characterization, pathogen inhibitory activity, partial characterization of bacteriocin and production of exoenzymes. Res. Vet. Sci. 2016, 108, 76–84. [Google Scholar] [PubMed]
- Ramesh, D.; Souissi, S.; Ahamed, T. S, Effects of the potential probiotics Bacillus aerophilus KADR3 in inducing immunity and disease resistance in Labeo rohita. Fish Shellfish. Immunol. 2017, 70, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Priest, F.G.; Goodfellow, M.; Shute, L.A.; Berkeley, R.C.W. Bacillus amyloliquefaciens sp. nov. Nom. Rev. Int. J. Syst. Evol. Microbiol. 1987, 37, 69–71. [Google Scholar]
- Fukumoto, J, Studies on the production of bacterial amylase. I. Isolation of bacteria secreting potent amylases and their distribution. Nippon. Nogeikagaku Kaishi 1943, 19, 487–503.
- Liu, H.; Prajapati, V.; Prajapati, S.; Bais, H. ; Lu, J, Comparative genome analysis of Bacillus amyloliquefaciens focusing on phylogenomics, functional Traits, and prevalence of antimicrobial and virulence genes. Front. Genet. 2021, 12, 724217. [Google Scholar]
- Chen, Y.; Li, J.; Xiao, P.; Li, G.Y.; Yue, S.; Huang, J. . & Mo, Z. L, Isolation and characterization of Bacillus spp. M 001 for potential application in turbot (Scophthalmus maximus L.) against V ibrio anguillarum. Aquac. Nutr. 2016, 22, 374–381. [Google Scholar]
- Chen, Y.; Li, J.; Xiao, P.; Zhu, W.; Mo, Z, The ability of marine Bacillus spp. isolated from fish gastrointestinal tract and culture pond sediment to inhibit growth of aquatic pathogenic bacteria. Iran. J. Fish. Sci. 2016, 15, 701–714. [Google Scholar]
- Mukherjee, A.; Dutta, D.; Banerjee, S.; Ringø, E.; Breines, E.M.; Hareide, E. . & Ghosh, K, Culturable autochthonous gut bacteria in rohu, Labeo rohita. In vitro growth inhibition against pathogenic Aeromonas spp.; stability in gut, bio-safety and identification by 16S rRNA gene sequencing. Symbiosis 2017, 73, 165–177. [Google Scholar]
- Kavitha, M.; Raja, M. ; Perumal, P, Evaluation of probiotic potential of Bacillus spp. isolated from the digestive tract of freshwater fish Labeo calbasu (Hamilton, 1822). Aquaculture Reports 2018, 11, 59–69. [Google Scholar]
- Halder U, Banerjee A, Chaudhry V, Varshney RK, Mantri S, Bandopadhyay R, Draft genome report of Bacillus altitudinis SORB11, isolated from the Indian sector of the Southern Ocean. Genome Announc 2017, 5, e00339-17.
- Shafi S, Kamili AN, Shah MA, Bandh SA, Dar R, Dynamics of bacterial class Bacilli in the deepest valley lake of Kashmir-the Manasbal Lake. Microb Pathog 2017, 104, 78–83. [CrossRef] [PubMed]
- Vijay Kumar E, Srijana M, Kiran Kumar K, Harikrishna N, Reddy G, A novel serine alkaline protease from Bacillus altitudinis GVC11 and its application as a dehairing agent. Bioprocess Biosyst Eng 2011, 34, 403–409. [CrossRef] [PubMed]
- Mao S, Lu Z, Zhang C, Lu F, Bie X, Purification, characterization, and heterologous expression of a thermostable β-1,3–1,4-glucanase from Bacillus altitudinis YC-9. Appl. Biochem. Biotechnol. 2013, 169, 960–975. [CrossRef] [PubMed]
- Banerjee, S.; Mukherjee, A.; Dutta, D.; Ghosh, K. Non-Starch Polysaccharide Degrading Gut Bacteria in Indian Major Carps and Exotic Carps. Jordan J. Biol. Sci. 2016, 9. [Google Scholar] [CrossRef]
- Dey, A.; Ghosh, K.; Hazra, N, Evaluation of extracellular enzyme-producing autochthonous gut bacteria in walking catfish, Clarias batrachus (L. ). J. Fish. 2016, 4, 345–352. [Google Scholar] [CrossRef]
- Esakkiraj, P.; Usha, R.; Palavesam, A. ; Immanuel, G, Solid-state production of esterase using fish processing wastes by Bacillus altitudinis AP-MSU. Food Bioprod. Process. 2012, 90, 370–376. [Google Scholar] [CrossRef]
- Shivaji, S.; Chaturvedi, P.; Begum, Z.; Pindi, P.K.; Manorama, R.; Padmanaban, D.A. . & Narlikar, J. V, Janibacter hoylei sp. nov.; Bacillus isronensis sp. nov. and Bacillus aryabhattai sp. nov.; isolated from cryotubes used for collecting air from the upper atmosphere. Int. J. Syst. Evol. Microbiol. 2009, 12, 2977–2986. [Google Scholar]
- Elarabi, N.I.; Abdelhadi, A.A.; Ahmed, R.H.; Saleh, I.; Arif, I.A.; Osman, G.; Ahmed, D. S, Bacillus aryabhattai FACU: A promising bacterial strain capable of manipulate the glyphosate herbicide residues. Saudi J. Biol. Sci. 2020, 27, 2207–2214. [Google Scholar] [CrossRef] [PubMed]
- Elsadek, M.M.; Zhu, W.; Wang, S.; Wang, X.; Guo, Z.; Lin, L. . & Zhang, D, Beneficial effects of indigenous Bacillus spp. on growth, antioxidants, immunity and disease resistance of Rhynchocypris lagowskii. Fish Shellfish. Immunol. 2023, 141, 109047. [Google Scholar] [PubMed]
- Khan, A. ; Ghosh, K, Characterization and identification of gut-associated phytase-producing bacteria in some fresh water fish cultured in ponds. Acta Ichthyol. Et Piscat. 2012, 42, 37–45. [Google Scholar] [CrossRef]
- Sella, S.R.; Vandenberghe, L.P.; Soccol, C. R, Bacillus atrophaeus: main characteristics and biotechnological applications–a review. Crit. Rev. Biotechnol. 2015, 35, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, H.S.; Broomall, S.M.; McNew, L.A.; Daligault, H.; Chapman, C.; Bruce, D. ;... & Skowronski, E. W, Genomic signatures of strain selection and enhancement in Bacillus atrophaeus var. globigii, a historical biowarfare simulant. PLoS ONE.
- Wu, W.J. ; Chang, J, Effect of oxygen on the germination and culturability of Bacillus atrophaeus spores. Int. Microbiol. 2022, 1–11. [Google Scholar]
- Askarian, F.; Zhou, Z.; Olsen, R.E.; Sperstad, S.; Ringø, E, Culturable autochthonous gut bacteria in Atlantic salmon (Salmo salar L. ) fed diets with or without chitin. Characterization by 16S rRNA gene sequencing, ability to produce enzymes and in vitro growth inhibition of four fish pathogens. Aquaculture 2012, 326, 1–8. [Google Scholar] [CrossRef]
- Logan, N.A.; Vos, P. D. ; Bacillus. Bergey's manual of systematics of archaea and bacteria 2015, 1-163.
- Priest, F. G, Aerobic Endospore-Forming. Bacillus 2013, 2, 27. [Google Scholar]
- Austin, B.; Al-Zahrani, A.M. J, The effect of antimicrobial compounds on the gastrointestinal microflora of rainbow trout, Salmo gairdneri Richardson. J. Fish Biol. 1988, 33, 1–14. [Google Scholar] [CrossRef]
- Ghosh, K.; Sen, S.K.; Ray, A. K, Growth and survival of rohu, Labeo rohita (Hamilton) spawn fed diets supplemented with fish intestinal microflora. Acta Ichthyologica et piscatorial 2002, 1, 83–92. [Google Scholar] [CrossRef]
- Ray, A.K.; Bairagi, A.; Ghosh, K.S.; Sen, S. K, Optimization of fermentation conditions for cellulase production by Bacillus subtilis CY5 and Bacillus circulans TP3 isolated from fish gut. Acta Ichthyologica et Piscatoria 2007, 37, 47–53. [Google Scholar] [CrossRef]
- Saha, S.; Roy, R.N.; Sen, S.K.; Ray, A. K, Characterization of cellulase-producing bacteria from the digestive tract of tilapia, Oreochromis mossambica (Peters) and grass carp, Ctenopharyngodon idella (Valenciennes). Aquaculture Research 2006, 37, 380–388. [Google Scholar] [CrossRef]
- Bandyopadhyay, P.; Das Mohapatra, P. K, Effect of a probiotic bacterium Bacillus circulans PB7 in the formulated diets: on growth, nutritional quality and immunity of Catla catla (Ham. ). Fish physiology and biochemistry 2009, 35, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Ianiro, G.; Rizzatti, G.; Plomer, M.; Lopetuso, L.; Scaldaferri, F.; Franceschi, F. ; .. & Gasbarrini, A, Bacillus clausii for the treatment of acute diarrhea in children: a systematic review and meta-analysis of randomized controlled trials. Nutrients 2018, 10, 1074. [Google Scholar] [PubMed]
- Wang, J.; Yang, H.L.; Xia, H.Q.; Ye, J.D.; Lu, K.L.; Hu, X. ; .. & Sun, Y.Z. Supplementation of heat-inactivated Bacillus clausii DE 5 in diets for grouper, Epinephelus coioides, improves feed utilization, intestinal and systemic immune responses and not growth performance. Aquaculture Nutrition 2018, 24, 821–831. [Google Scholar]
- Ye, J.D.; Wang, K.; Li, F.D.; Sun, Y.Z. Single or combined effects of fructo-and mannan oligosaccharide supplements and Bacillus clausii on the growth, feed utilization, body composition, digestive enzyme activity, innate immune response and lipid metabolism of the Japanese flounder Paralichthys olivaceus. Aquaculture nutrition 2011, 17, e902–e911. [Google Scholar]
- Jurenka, J.S. Bacillus coagulans. Alternative medicine review 2012, 17, 76–82. [Google Scholar] [PubMed]
- Breed, R.S.; Murray, E.G.D.; Smith, N.R. Bergey's manual of determinative bacteriology. Bergey's Manual of Determinative Bacteriology; (7th Edition) (1957).
- Bhatnagar, A.; Raparia, S. ; Kumari, S, Influence of isolated Bacillus coagulans on growth performance and digestive enzyme activities of Catla catla. Journal of Nature Science and Sustainable Technology 2012, 6, 225. [Google Scholar]
- Xu, Y.; Wang, Y.; Lin, J. Use of Bacillus coagulans as a Dietary Probiotic for the Common Carp, Cyprinus carpio. Journal of the World Aquaculture society 2014, 45, 403–411. [Google Scholar] [CrossRef]
- Obidi, O.F.; Soyinka, S.O.; Kamoru, T.A. Morphological, Biochemical and Molecular Characterisations of Bacteria Isolated from Water and Submerged Painted Boat Hulls in Badagry Lagoon, Lagos State, Nigeria. Journal of Applied Sciences and Environmental Management 2023, 27, 1579–1589. [Google Scholar]
- Banerjee, S.; Mukherjee, A.; Dutta, D.; Ghosh, K. Evaluation of chitinolytic gut microbiota in some carps and optimization of culture conditions for chitinase production by the selected bacteria. The Journal of Microbiology, Biotechnology and Food Sciences 2015, 5, 12. [Google Scholar] [CrossRef]
- Ghosh, K.; Roy, M.; Kar, N.; RingØ, E. Gastrointestinal bacteria in rohu, Labeo rohita (Actinopterygii: Cypriniformes: Cyprinidae): scanning electron microscopy and bacteriological study. Acta Ichthyologica et Piscatoria 2010, 40, 129–135. [Google Scholar] [CrossRef]
- Mondal, S.; Roy, T.; Ray, A.K. Characterization and identification of enzyme-producing bacteria isolated from the digestive tract of bata, Labeo bata. Journal of the World Aquaculture Society 2010, 41, 369–377. [Google Scholar] [CrossRef]
- Banerjee, G.; Ray, A.K.; Askarian, F.; Ringø, E. Characterisation and identification of enzyme-producing autochthonous bacteria from the gastrointestinal tract of two Indian air-breathing fish. Beneficial Microbes 2013, 4, 277–284. [Google Scholar] [CrossRef]
- Das, P.; Mandal, S.; Khan, A.; Manna, S.K.; Ghosh, K. Distribution of extracellular enzyme-producing bacteria in the digestive tracts of 4 brackish water fish species. Turkish Journal of Zoology 2014, 38, 79–88. [Google Scholar] [CrossRef]
- Ghosh, K.; Banerjee, S.; Moon, U.M.; Khan, H.A.; Dutta, D. Evaluation of gut associated extracellular enzyme-producing and pathogen inhibitory microbial community as potential probiotics in Nile tilapia, Oreochromis niloticus. Int. J. Aquac. 2017, 7. [Google Scholar]
- Guzmán-Moreno, J.; García-Ortega, L.F.; Torres-Saucedo, L.; Rivas-Noriega, P.; Ramírez-Santoyo, R.M.; Sánchez-Calderón, L. ;... & Vidales-Rodríguez, L. E, Bacillus megaterium HgT21: A Promising Metal Multiresistant Plant Growth-Promoting Bacteria for Soil Biorestoration. Microbiol. Spectr. 2022; 10. [Google Scholar]
- David, F.; Hebeisen, M.; Schade, G.; Franco-Lara, E.; Di Berardino, M. Viability and membrane potential analysis of Bacillus megaterium cells by impedance flow cytometry. Biotechnology and bioengineering 2012, 109, 483–492. [Google Scholar] [CrossRef]
- Green, T.J.; Smullen, R.; Barnes, A. C, Dietary soybean protein concentrate-induced intestinal disorder in marine farmed Atlantic salmon, Salmo salar is associated with alterations in gut microbiota. Vet. Microbiol. 2013, 166, 286–292. [Google Scholar] [CrossRef]
- Kumar, M.; Yadav, A.N.; Saxena, R.; Rai, P.K.; Paul, D.; Tomar, R. S Novel, methanotrophic and methanogenic bacterial communities from diverse ecosystems and their impact on environment. Biocatal. Agric. Biotechnol. 2021, 33. [Google Scholar] [CrossRef]
- Ran, C.; Carrias, A.; Williams, M.A.; Capps, N.; Dan, B.C.; Newton, J.C. ;... & Liles, M. R, Identification of Bacillus strains for biological control of catfish pathogens. 2012.
- Mukherjee, A.; Ghosh, K. Antagonism against fish pathogens by cellular components and verification of probiotic properties in autochthonous bacteria isolated from the gut of an I ndian major carp, Catla catla (Hamilton). Aquaculture Research 2016, 47, 2243–2255. [Google Scholar] [CrossRef]
- Ramírez-Torrez, J.A.; Monroy-Dosta, M.D.C.; Hernández-Hernández, L.H.; Castro-Mejía, J.; Bustos-Martínez, J.A.; Hamdan-Partida, A. Presumptive probiotic isolated from Oncorhynchus mykiss (Walbaum, 1792), cultivated in Mexico. Int. J. Aquatic Sci. 2018, 9, 3–12. [Google Scholar]
- Venkateswaran, K.; Kempf, M.; Chen, F.; Satomi, M.; Nicholson, W. ; Kern, R, Bacillus nealsonii sp. nov.; isolated from a spacecraft-assembly facility, whose spores are γ-radiation resistant. International Journal of Systematic and Evolutionary Microbiology 2003, 53, 165–172. [Google Scholar] [PubMed]
- Zammuto, V.; Fuchs, F.M.; Fiebrandt, M.; Stapelmann, K.; Ulrich, N.J.; Maugeri, T.L. ; .. & Moeller, R, Comparing spore resistance of Bacillus strains isolated from hydrothermal vents and spacecraft assembly facilities to environmental stressors and decontamination treatments. Astrobiology 2018, 18, 1425–1434. [Google Scholar] [PubMed]
- Yilmaz, S.; Ergün, S.; Yıgıt, M. Effects of dietary FARMARIN® XP supplement on immunological responses and disease resistance of rainbow trout (Oncorhynchus mykiss). Aquaculture 2018, 496, 211–220. [Google Scholar] [CrossRef]
- Fu, X.; Gong, L.; Liu, Y.; Lai, Q.; Li, G. ; Shao, Z, Bacillus pumilus group comparative genomics: toward pangenome features, diversity, and marine environmental adaptation. Front. Microbiol. 2021, 12, 571212. [Google Scholar] [CrossRef] [PubMed]
- Dobrzyński, J.; Jakubowska, Z.; Dybek, B. Potential of Bacillus pumilus to directly promote plant growth. Front. Microbiol. 2022, 13, 1069053. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, K.; Sen, S.K.; Ray, A.K. Characterization of Bacilli isolated from the gut of rohu, Labeo rohita, fingerlings and its significance in digestion. J. Appl. Aquac. 2002, 12, 33–42. [Google Scholar] [CrossRef]
- Yang, H.L.; Sun, Y.Z.; Hu, X.; Ye, J.D.; Lu, K.L.; Hu, L.H.; Zhang, J.J. Bacillus pumilus SE5 originated PG and LTA tuned the intestinal TLRs/MyD88 signaling and microbiota in grouper (Epinephelus coioides). Fish Shellfish Immunol. 2019, 88, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Al-Hisnawi, A.; Ringø, E.; Davies, S.J.; Waines, P.; Bradley, G.; Merrifield, D.L. First report on the autochthonous gut microbiota of brown trout (S almo trutta Linnaeus). Aquac. Res. 2015, 46, 2962–2971. [Google Scholar] [CrossRef]
- Ringø, E.Z.Z.V.; Zhou, Z.; Vecino, J.G.; Wadsworth, S.; Romero, J.; Krogdahl, Å. . & Merrifield, D.L. Effect of dietary components on the gut microbiota of aquatic animals. A never-ending story? Aquac. Nutr. 2016, 22, 219–282. [Google Scholar]
- Fritze, D. Taxonomy of the genus Bacillus and related genera: the aerobic endospore-forming bacteria. Phytopathology 2004, 94, 1245–1248. [Google Scholar] [CrossRef] [PubMed]
- Adimpong, D.B.; Sørensen, K.I.; Nielsen, D.S.; Thorsen, L.; Rasmussen, T.B.; Derkx, P.M.; Jespersen, L. Draft whole-genome sequence of Bacillus sonorensis strain L12, a source of nonribosomal lipopeptides. Genome Announc. 2013, 1, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Dutta, D.; Ghosh, K. Screening of extracellular enzyme-producing and pathogen inhibitory gut bacteria as putative probiotics in mrigal, Cirrhinus mrigala (Hamilton, 1822). International Journal of Fisheries and Aquatic Studies 2015, 2, 310–318. [Google Scholar]
- EL-Mongy, M.; Hamouda, R.; Ali, S.; Sedeek, E.; Mahmoud, E. Antibacterial, antioxidant and anticancer of fermentation by Bacillus subtilis on bagasse and wheat bran. Curr. Chem. Lett. 2022, 11, 383–392. [Google Scholar] [CrossRef]
- Addo, S.; Carrias, A.A.; Williams, M.A.; Liles, M.R.; Terhune, J.S.; Davis, D.A. Effects of Bacillus subtilis strains on growth, immune parameters, and Streptococcus iniae susceptibility in Nile tilapia, Oreochromis niloticus. J. World Aquac. Soc. 2017, 48, 257–267. [Google Scholar] [CrossRef]
- Won, S.; Hamidoghli, A.; Choi, W.; Park, Y.; Jang, W.J.; Kong, I.S.; Bai, S.C. Effects of Bacillus subtilis WB60 and Lactococcus lactis on growth, immune responses, histology and gene expression in Nile tilapia, Oreochromis niloticus. Microorganisms v, 8(1).
- Romanova, E.; Spirina, E.; Romanov, V.; Lyubomirova, V.; Shadyeva, L. Effects of Bacillus subtilis and Bacillus licheniformis on catfish in industrial aquaculture. In E3S Web of Conferences. EDP Sci. 2020, 175, 02013. [Google Scholar]
- Tachibana, L.; Telli, G.S.; Dias, D.D.C.; Goncalves, G.S.; Guimaraes, M.C.; Ishikawa, C.M. . & Ranzani-Paiva, M.J.T. Bacillus subtilis and Bacillus licheniformis in diets for Nile tilapia (Oreochromis niloticus): Effects on growth performance, gut microbiota modulation and innate immunology. Aquac. Res. 2021, 52, 1630–1642. [Google Scholar]
- He, S.; Wu, Z.; Liu, Y.; Wu, N.; Tao, Y.; Xu, L. . & Ringø, E. Effects of dietary 60 g kg− 1 dried distiller's grains in least-cost practical diets on production and gut allochthonous bacterial composition of cage-cultured fish: comparison among fish species with different natural food habits. Aquac. Nutr. 2013, 19, 765–772. [Google Scholar]
- Li, Z.; Xu, L.; Liu, W.; Liu, Y.; Ringø, E.; Du, Z.; Zhou, Z. Protein replacement in practical diets altered gut allochthonous bacteria of cultured cyprinid species with different food habits. Aquac. Int. 2015, 23, 913–928. [Google Scholar] [CrossRef]
- Talukdar, S.; Ringø, E.; Ghosh, K. Extracellular tannase-producing bacteria detected in the digestive tracts of freshwater fishes (Actinopterygii: Cyprinidae and Cichlidae), 2016.
- Das, P.; Ghosh, K. Evaluation of phytase-producing ability by a fish gut bacterium, Bacillus subtilis subsp. subtilis. J. Biol. Sci. 2013, 13, 691. [Google Scholar] [CrossRef]
- Das, P.; Ghosh, K. Improvement of nutritive value of sesame oil cake in formulated diets for rohu, Labeo rohita (Hamilton) after bio-processing through solid state fermentation by a phytase-producing fish gut bacterium. Int. J. Aquat. Biol. 2015, 3, 89–101. [Google Scholar]
- Shah, Z.; Krumholz, L.; Aktas, D.F.; Hasan, F.; Khattak, M.; Shah, A.A. Degradation of polyester polyurethane by a newly isolated soil bacterium, Bacillus subtilis strain MZA-75. Biodegradation 2013, 24, 865–877. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, E.M.; Krawczyk, A.O.; Klaus, V.; de Jong, A.; Boekhorst, J.; Eijlander, R.T. . & Wells-Bennik, M.H. Bacillus thermoamylovorans spores with very-high-level heat resistance germinate poorly in rich medium despite the presence of ger clusters but efficiently upon exposure to calcium-dipicolinic acid. Appl. Environ. Microbiol. 2015, 81, 7791–7801. [Google Scholar] [PubMed]
- Choonut, A.; Prasertsan, P.; Klomklao, S.; Sangkharak, K. Bacillus thermoamylovorans-related strain isolated from high temperature sites as potential producers of medium-chain-length polyhydroxyalkanoate (mcl-PHA). Curr. Microbiol. 2020, 77, 3044–3056. [Google Scholar] [CrossRef] [PubMed]
- Nyman, A.; Huyben, D.; Lundh, T.; Dicksved, J. Effects of microbe-and mussel-based diets on the gut microbiota in Arctic charr (Salvelinus alpinus). Aquac. Rep. 2017, 5, 34–40. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Griko, N.; Junker, M.; Bulla, L.A. Bacillus thuringiensis: a genomics and proteomics perspective. Bioeng. Bugs 2010, 1, 31–50. [Google Scholar] [CrossRef]
- Peixoto, S.B.; Cladera-Olivera, F.; Daroit, D.J.; Brandelli, A. Cellulase-producing Bacillus strains isolated from the intestine of Amazon basin fish. Aquac. Res. 2011, 42, 887–891. [Google Scholar] [CrossRef]
- Krishnamurthi, S.; Ruckmani, A.; Pukall, R.; Chakrabarti, T. Psychrobacillus gen. nov. and proposal for reclassification of Bacillus insolitus Larkin & Stokes, 1967, B. psychrotolerans Abd-El Rahman et al.; 2002 and B. psychrodurans Abd-El Rahman et al.; 2002 as Psychrobacillus insolitus comb. nov.; Psychrobacillus Psychrotolerans Comb. Nov. Psychrobacillus Psychrodurans Comb. Nov. Syst. Appl. Microbiol. 2010, 33, 367–373. [Google Scholar]
- Krishnamurthi, S.; Chakrabarti, T.; Stackebrandt, E. Re-examination of the taxonomic position of Bacillus silvestris Rheims et al. 1999 and proposal to transfer it to Solibacillus gen. nov. as Solibacillus Silvestris Comb. Nov. Int. J. Syst. Evol. Microbiol. 2009, 59, 1054–1058. [Google Scholar] [CrossRef]
- Rheims, H.; Frühling, A.; Schumann, P.; Rohde, M.; Stackebrandt, E. Bacillus silvestris sp. nov.; a new member of the genus Bacillus That Contain. Lysine Its Cell Wall. Int. J. Syst. Evol. Microbiol. 1999, 49, 795–802. [Google Scholar]
- Abdel-Moneim, A.M.E.; Selim, D.A.; Basuony, H.A.; Sabic, E.M.; Saleh, A.A.; Ebeid, T.A. Effect of dietary supplementation of Bacillus subtilis spores on growth performance, oxidative status, and digestive enzyme activities in Japanese quail birds. Trop. Anim. Health Prod. 2020, 52, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Latif, H.M.; Chaklader, M.R.; Shukry, M.; Ahmed, H.A.; Khallaf, M.A. A multispecies probiotic modulates growth, digestive enzymes, immunity, hepatic antioxidant activity, and disease resistance of Pangasianodon hypophthalmus fingerlings. Aquaculture 2023, 563, 738948. [Google Scholar] [CrossRef]
- Hasan, K.N.; Banerjee, G. Recent studies on probiotics as beneficial mediator in aquaculture: a review. J. Basic Appl. Zool. 2020, 81, 1–16. [Google Scholar] [CrossRef]
- Puvanasundram, P.; Chong, C.M.; Sabri, S.; Yusoff, M.S.; Karim, M. Multi-strain probiotics: Functions, effectiveness and formulations for aquaculture applications. Aquac. Rep. 2021, 21, 100905. [Google Scholar] [CrossRef]
- Cherdkeattipol, K.; Chuchird, N.; Chonudomkul, D.; Yongmanitchai, W.; Pichitkul, P. Effect of partial replacement of fish meal by Bacillus sp-fermented soybean meal on growth performance, immunity, hepatopancreas microbiota and disease resistance in pacific White Shrimp (Litopenaeus vannamei). J. Fish. Environ. 2021, 45, 32–42. [Google Scholar]
- Kaspar, F.; Neubauer, P.; Gimpel, M. Bioactive secondary metabolites from Bacillus subtilis: a comprehensive review. J. Nat. Prod. 2019, 82, 2038–2053. [Google Scholar] [CrossRef]
- Assefa, A.; Abunna, F. Maintenance of fish health in aquaculture: review of epidemiological approaches for prevention and control of infectious disease of fish. Veterinary medicine international 2018, 2018. [Google Scholar] [CrossRef]
- Raabe, V.N.; Shane, A.L. Group B streptococcus (Streptococcus agalactiae). Microbiol. Spectr. 2019, 7, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen-Ivey, C.R.; Figueras, M.J.; McGarey, D.; Liles, M.R. Virulence factors of Aeromonas hydrophila: in the wake of reclassification. Frontiers in microbiology 2016, 1337. [Google Scholar]
- Vivas, R.; Barbosa, A.A.T.; Dolabela, S.S.; Jain, S. Multidrug-resistant bacteria and alternative methods to control them: an overview. Microb. Drug Resist. 2019, 25, 890–908. [Google Scholar] [CrossRef]
- Uddin, T.M.; Chakraborty, A.J.; Khusro, A.; Zidan, B.R.M.; Mitra, S.; Emran, T.B. . & Koirala, N. Antibiotic resistance in microbes: History, mechanisms, therapeutic strategies and future prospects. J. Infect. Public Health 2021, 14, 1750–1766. [Google Scholar]
- Jansen, K.U.; Knirsch, C.; Anderson, A.S. The role of vaccines in preventing bacterial antimicrobial resistance. Nat. Med. 2018, 24, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Singh, R. Probiotics in aquaculture: a promising emerging alternative approach. Symbiosis 2019, 77, 99–113. [Google Scholar] [CrossRef]
- Yilmaz, S.; Yilmaz, E.; Dawood, M.A.; Ringø, E.; Ahmadifar, E.; Abdel-Latif, H.M. Probiotics, prebiotics, and synbiotics used to control vibriosis in fish: A review. Aquaculture 2022, 547, 737514. [Google Scholar] [CrossRef]
- Knipe, H.; Temperton, B.; Lange, A.; Bass, D.; Tyler, C.R. Probiotics and competitive exclusion of pathogens in shrimp aquaculture. Rev. Aquac. 2021, 13, 324–352. [Google Scholar] [CrossRef]
- Ventura, M.T.; Casciaro, M.; Gangemi, S.; Buquicchio, R. Immunosenescence in aging: between immune cells depletion and cytokines up-regulation. Clin. Mol. Allergy 2017, 15, 1–8. [Google Scholar] [CrossRef]
- Chattaraj, S.; Ganguly, A.; Mandal, A.; Das Mohapatra, P.K. A review of the role of probiotics for the control of viral diseases in aquaculture. Aquac. Int. 2022, 30, 2513–2539. [Google Scholar] [CrossRef]
- Cao, L.; Pan, L.; Gong, L.; Yang, Y.; He, H.; Li, Y. . & Xia, L. Interaction of a novel Bacillus velezensis (BvL03) against Aeromonas hydrophila in vitro and in vivo in grass carp. Appl. Microbiol. Biotechnol. 2019, 103, 8987–8999. [Google Scholar] [PubMed]
- Iqtedar, M.; Aslam, M.; Akhyar, M.; Shehzaad, A.; Abdullah, R.; Kaleem, A. Extracellular biosynthesis, characterization, optimization of silver nanoparticles (AgNPs) using Bacillus mojavensis BTCB15 and its antimicrobial activity against multidrug resistant pathogens. Prep. Biochem. Biotechnol. 2019, 49, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Dubey, S.; Diep, D.B.; Evensen, Ø.; Munang’andu, H.M. Garvicin KS, a Broad-Spectrum Bacteriocin Protects Zebrafish Larvae against Lactococcus garvieae Infection. Int. J. Mol. Sci. 2022, 23, 2833. [Google Scholar] [CrossRef]
- Yılmaz, S.; Ergun, S.; Yigit, M.; Çelik, E.Ş. Effect of combination of dietary Bacillus subtilis and trans-cinnamic acid on innate immune responses and resistance of rainbow trout, Oncorhynchus mykiss to Yersinia ruckeri. Aquac. Res. 2020, 51, 441–454. [Google Scholar] [CrossRef]
- Xia, Y.; Wang, M.; Gao, F.; Lu, M.; Chen, G. Effects of dietary probiotic supplementation on the growth, gut health and disease resistance of juvenile Nile tilapia (Oreochromis niloticus). Anim. Nutr. 2020, 6, 69–79. [Google Scholar] [CrossRef]
- Sun, H.; Tang, J.W.; Yao, X.H.; Wu, Y.F.; Wang, X.; Feng, J. Improvement of the Nutritional Quality of Cottonseed Meal by Bacillus subtilis and the Addition of Papain. International Journal of Agriculture & Biology 2012, 14. [Google Scholar]
- Wu, Z.X.; Feng, X.; Xie, L.L.; Peng, X.Y.; Yuan, J.; Chen, X.X. Effect of probiotic Bacillus subtilis Ch9 for grass carp, Ctenopharyngodon idella (Valenciennes, 1844), on growth performance, digestive enzyme activities and intestinal microflora. J. Appl. Ichthyol. 2012, 28, 721–727. [Google Scholar] [CrossRef]
- Cha, J.H.; Rahimnejad, S.; Yang, S.Y.; Kim, K.W.; Lee, K.J. Evaluations of Bacillus spp. as dietary additives on growth performance, innate immunity and disease resistance of olive flounder (Paralichthys olivaceus) against Streptococcus iniae and as water additives. Aquaculture 2013, 402, 50–57. [Google Scholar] [CrossRef]
- Telli, G.S.; Ranzani-Paiva, M.J.T.; de Carla Dias, D.; Sussel, F.R.; Ishikawa, C.M.; Tachibana, L. Dietary administration of Bacillus subtilis on hematology and non-specific immunity of Nile tilapia Oreochromis niloticus raised at different stocking densities. Fish Shellfish Immunol. 2014, 39, 305–311. [Google Scholar] [CrossRef]
- Aly, S.M.; Ahmed, Y.A.G.; Ghareeb, A.A.A.; Mohamed, M.F. Studies on Bacillus subtilis and Lactobacillus acidophilus, as potential probiotics, on the immune response and resistance of Tilapia nilotica (Oreochromis niloticus) to challenge infections. Fish & shellfish immunology.
- Liu, H.; Wang, S.; Cai, Y.; Guo, X.; Cao, Z.; Zhang, Y. . & Zhou, Y. Dietary administration of Bacillus subtilis HAINUP40 enhances growth, digestive enzyme activities, innate immune responses and disease resistance of tilapia, Oreochromis niloticus. Fish Shellfish Immunol. 2017, 60, 326–333. [Google Scholar]
- Ai, Q.; Xu, H.; Mai, K.; Xu, W.; Wang, J.; Zhang, W. Effects of dietary supplementation of Bacillus subtilis and fructooligosaccharide on growth performance, survival, non-specific immune response and disease resistance of juvenile large yellow croaker, Larimichthys crocea. Aquaculture, 2011; 317, 155–161. [Google Scholar]
- Reda, R.M.; Selim, K.M. Evaluation of Bacillus amyloliquefaciens on the growth performance, intestinal morphology, hematology and body composition of Nile tilapia, Oreochromis niloticus. Aquac. Int. 2015, 23, 203–217. [Google Scholar] [CrossRef]
- Gobi, N.; Malaikozhundan, B.; Sekar, V.; Shanthi, S.; Vaseeharan, B.; Jayakumar, R.; Nazar, A.K. GFP tagged Vibrio parahaemolyticus Dahv2 infection and the protective effects of the probiotic Bacillus licheniformis Dahb1 on the growth, immune and antioxidant responses in Pangasius hypophthalmus. Fish Shellfish Immunol. 2016, 52, 230–238. [Google Scholar] [CrossRef]
- Adorian, T.J.; Jamali, H.; Farsani, H.G.; Darvishi, P.; Hasanpour, S.; Bagheri, T.; Roozbehfar, R. Effects of probiotic bacteria Bacillus on growth performance, digestive enzyme activity, and hematological parameters of Asian sea bass, Lates calcarifer (Bloch). Probiotics and antimicrobial proteins 2019, 11, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.Z.; Yang, H.L.; Ma, R.L.; Lin, W.Y. Probiotic applications of two dominant gut Bacillus strains with antagonistic activity improved the growth performance and immune responses of grouper Epinephelus coioides. Fish Shellfish Immunol. 2010, 29, 803–809. [Google Scholar] [CrossRef]
- Silva, T.F.A.; Petrillo, T.R.; Yunis-Aguinaga, J.; Marcusso, P.F.; Claudiano, G.D.S.; Moraes, F.R.D.; Moraes, J.R. Efectos del probiótico Bacillus amyloliquefaciens en el crecimiento, hematología y morfometría intestinal en tilapias del Nilo criadas en balsa jaula. Lat. Am. J. Aquat. Res. 2015, 43, 963–971. [Google Scholar]
- Sahandi, J.; Jafarian, H.; Roozbehfar, R.; Babaei, S.; Dehestani, M. The use of two enrichment forms (Brachionus plicatilis enrichment and rearing water enrichment) with probiotic bacilli spore on growth and survival of Silver carp (Hypophthalmichthys molitrix), 2012.
- Wang, Y.; Chi, L.; Liu, Q.; Xiao, Y.; Ma, D.; Xiao, Z. . & Li, J. Effects of stocking density on the growth and immunity of Atlantic salmon salmo salar reared in recirculating aquaculture system (RAS). J. Oceanol. Limnol. 2019, 37, 350–360. [Google Scholar]
- Giri, A.K.; Sahu, N.P.; Saharan, N.; Dash, G. Effect of dietary supplementation of chromium on growth and biochemical parameters of Labeo rohita (Hamilton) fingerlings. Indian J. Fish. 2014, 61, 73–81. [Google Scholar]
- Van Doan, H.; Hoseinifar, S.H.; Khanongnuch, C.; Kanpiengjai, A.; Unban, K.; Srichaiyo, S. Host-associated probiotics boosted mucosal and serum immunity, disease resistance and growth performance of Nile tilapia (Oreochromis niloticus). Aquaculture 2018, 491, 94–100. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, Y.; Gu, Q.; Li, W. Effect of dietary probiotic, Bacillus coagulans, on growth performance, chemical composition, and meat quality of Guangxi Yellow chicken. Poult. Sci. 2010, 89, 588–593. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, G.; Adorian, T.J.; Rafiee, G. Beneficial effects of Bacillus subtilis on water quality, growth, immune responses, endotoxemia and protection against lipopolysaccharide-induced damages in Oreochromis niloticus under biofloc technology system. Aquac. Nutr. 2020, 26, 1476–1492. [Google Scholar] [CrossRef]
- Abarike, E.D.; Cai, J.; Lu, Y.; Yu, H.; Chen, L.; Jian, J. . & Kuebutornye, F.K. Effects of a commercial probiotic BS containing Bacillus subtilis and Bacillus licheniformis on growth, immune response and disease resistance in Nile tilapia, Oreochromis niloticus. Fish Shellfish Immunol. 2018, 82, 229–238. [Google Scholar] [PubMed]
- Thy, H.T.T.; Tri, N.N.; Quy, O.M.; Fotedar, R.; Kannika, K.; Unajak, S.; Areechon, N. Effects of the dietary supplementation of mixed probiotic spores of Bacillus amyloliquefaciens 54A, and Bacillus pumilus 47B on growth, innate immunity and stress responses of striped catfish (Pangasianodon hypophthalmus). Fish Shellfish Immunol. 2017, 60, 391–399. [Google Scholar]
- Azarin, H.; Aramli, M.S.; Imanpour, M.R.; Rajabpour, M. Effect of a probiotic containing Bacillus licheniformis and Bacillus subtilis and ferroin solution on growth performance, body composition and haematological parameters in Kutum (Rutilus frisii kutum) fry. Probiotics Antimicrob. Proteins 2015, 7, 31–37. [Google Scholar] [CrossRef]
- Hassaan, M.S.; Mohammady, E.Y.; Soaudy, M.R.; Elashry, M.A.; Moustafa, M.M.; Wassel, M.A. . & Elsaied, H.E. Synergistic effects of Bacillus pumilus and exogenous protease on Nile tilapia (Oreochromis niloticus) growth, gut microbes, immune response and gene expression fed plant protein diet. Anim. Feed Sci. Technol. 2021, 275, 114892. [Google Scholar]
- Zhang, C.N.; Li, X.F.; Xu, W.N.; Zhang, D.D.; Lu, K.L.; Wang, L.N. . & Liu, W.B. Combined effects of dietary fructooligosaccharide and Bacillus licheniformis on growth performance, body composition, intestinal enzymes activities and gut histology of triangular bream (Megalobrama terminalis). Aquac. Nutr. 2015, 21, 755–766. [Google Scholar]
- Kunjiappan, S.; Bhattacharjee, C.; Chowdhury, R. In vitro antioxidant and hepatoprotective potential of Azolla microphylla phytochemically synthesized gold nanoparticles on acetaminophen–induced hepatocyte damage in Cyprinus carpio L. Vitr. Cell. Dev. Biol. -Anim. 2015, 51, 630–643. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi-Arpanahi, D.; Soltani, E.; Jafaryan, H.; Soltani, M.; Naderi-Samani, M.; Campa-Córdova, A.I. Efficacy of two commercial and indigenous probiotics, Bacillus subtilis and Bacillus licheniformis on growth performance, immuno-physiology and resistance response of juvenile white shrimp (Litopenaeus vannamei). Aquaculture 2018, 496, 43–49. [Google Scholar] [CrossRef]
- Babazadeh, D.; Vahdatpour, T.; Nikpiran, H.; Jafargholipour, M.A.; Vahdatpour, S. Effects of probiotic, prebiotic and synbiotic intake on blood enzymes and performance of Japanese quails (Coturnix japonica). Indian J. Anim. Sci. 2011, 81, 870. [Google Scholar]
- Chimela, W.; Mesua, N.; Abdulraheem, B.A. Aspartate transaminase (AST) activity in selected tissues & organs of Clarias Gariepinus exposed to different levels of paraquat. J. Environ. Anal. Toxicol. 2014, 4, 2161–0525. [Google Scholar]
- Hassaan, M.S.; Soltan, M.A.; Jarmołowicz, S.; Abdo, H.S. Combined effects of dietary malic acid and B acillus subtilis on growth, gut microbiota and blood parameters of N ile tilapia (Oreochromis niloticus). Aquac. Nutr. 2018, 24, 83–93. [Google Scholar] [CrossRef]
- Sutthi, N.; Thaimuangphol, W.; Rodmongkoldee, M.; Leelapatra, W.; Panase, P. Growth performances, survival rate, and biochemical parameters of Nile tilapia (Oreochromis niloticus) reared in water treated with probiotic. Comp. Clin. Pathol. 2018, 27, 597–603. [Google Scholar] [CrossRef]
- Nandi, A.; Banerjee, G.; Dan, S.K.; Ghosh, K.; Ray, A.K. Evaluation of in vivo probiotic efficiency of Bacillus amyloliquefaciens in Labeo rohita challenged by pathogenic strain of Aeromonas hydrophila MTCC 1739. Probiotics and antimicrobial proteins 2018, 10, 391–398. [Google Scholar] [CrossRef]
- Pérez-Sánchez, J.; Benedito-Palos, L.; Estensoro, I.; Petropoulos, Y.; Calduch-Giner, J.A.; Browdy, C.L.; Sitjà-Bobadilla, A. Effects of dietary NEXT ENHANCE® 150 on growth performance and expression of immune and intestinal integrity related genes in gilthead sea bream (Sparus aurata L. ). Fish & shellfish immunology 2015, 44, 117–128. [Google Scholar]
- Kuebutornye, F.K.; Tang, J.; Cai, J.; Yu, H.; Wang, Z.; Abarike, E.D. . & Afriyie, G. In vivo assessment of the probiotic potentials of three host-associated Bacillus species on growth performance, health status and disease resistance of Oreochromis niloticus against Streptococcus agalactiae. Aquaculture 2020, 527, 735440. [Google Scholar]
- Ghalwash, H.R.; Salah, A.S.; El-Nokrashy, A.M.; Abozeid, A.M.; Zaki, V.H.; Mohamed, R.A. Dietary supplementation with Bacillus species improves growth, intestinal histomorphology, innate immunity, antioxidative status and expression of growth and appetite-regulating genes of Nile tilapia fingerlings. Aquac. Res. 2022, 53, 1378–1394. [Google Scholar] [CrossRef]
- Avella, M.A.; Gioacchini, G.; Decamp, O.; Makridis, P.; Bracciatelli, C.; Carnevali, O. Application of multi-species of Bacillus in sea bream larviculture. Aquaculture 2010, 305, 12–19. [Google Scholar]
- Midhun, S.J.; Neethu, S.; Arun, D.; Vysakh, A.; Divya, L.; Radhakrishnan, E.K.; Jyothis, M. Dietary supplementation of Bacillus licheniformis HGA8B improves growth parameters, enzymatic profile and gene expression of Oreochromis niloticus. Aquaculture 2019, 505, 289–296. [Google Scholar] [CrossRef]
- Vallejos-Vidal, E.; Sanz-Milián, B.; Teles, M.; Reyes-Cerpa, S.; Mancera, J.M.; Tort, L.; Reyes-Lopez, F.E. The gene expression profile of the glucocorticoid receptor 1 (gr1) but not gr2 is modulated in mucosal tissues of gilthead sea bream (Sparus aurata) exposed to acute air-exposure stress. Front. Mar. Sci. 2022, 9, 977719. [Google Scholar] [CrossRef]
- Yi, Y.; Zhang, Z.; Zhao, F.; Liu, H.; Yu, L.; Zha, J.; Wang, G. Probiotic potential of Bacillus velezensis JW: antimicrobial activity against fish pathogenic bacteria and immune enhancement effects on Carassius auratus. Fish Shellfish Immunol. 2018, 78, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.Z.; Xia, H.Q.; Yang, H.L.; Wang, Y.L.; Zou, W.C. TLR2 signaling may play a key role in the probiotic modulation of intestinal microbiota in grouper Epinephelus coioides. Aquaculture 2014, 430, 50–56. [Google Scholar] [CrossRef]
- Zokaeifar, H.; Balcázar, J.L.; Saad, C.R.; Kamarudin, M.S.; Sijam, K.; Arshad, A.; Nejat, N. Effects of Bacillus subtilis on the growth performance, digestive enzymes, immune gene expression and disease resistance of white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2012, 33, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Keysami, M.A.; Mohammadpour, M.; Saad, C.R. Probiotic activity of Bacillus subtilis in juvenile freshwater prawn, Macrobrachium rosenbergii (de Man) at different methods of administration to the feed. Aquac. Int. 2012, 20, 499–511. [Google Scholar] [CrossRef]
- NavinChandran, M.; Iyapparaj, P.; Moovendhan, S.; Ramasubburayan, R.; Prakash, S.; Immanuel, G.; Palavesam, A. Influence of probiotic bacterium Bacillus cereus isolated from the gut of wild shrimp Penaeus monodon in turn as a potent growth promoter and immune enhancer in P. monodon. Fish Shellfish Immunol. 2014, 36, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Chai, P.C.; Song, X.L.; Chen, G.F.; Xu, H.; Huang, J. Dietary supplementation of probiotic Bacillus PC465 isolated from the gut of Fenneropenaeus chinensis improves the health status and resistance of Litopenaeus vannamei against white spot syndrome virus. Fish Shellfish Immunol. 2016, 54, 602–611. [Google Scholar] [CrossRef] [PubMed]
- Laranja, J. Amorphous poly-β-hydroxybutyrate (PHB)-accumulating Bacillus spp. as biocontrol agents in crustacean culture (Doctoral dissertation, Ghent University), 2017.
- Adilah, R.N.; Chiu, S.T.; Hu, S.Y.; Ballantyne, R.; Happy, N.; Cheng, A.C.; Liu, C.H. Improvement in the probiotic efficacy of Bacillus subtilis E20-stimulates growth and health status of white shrimp, Litopenaeus vannamei via encapsulation in alginate and coated with chitosan. Fish Shellfish Immunol. 2022, 125, 74–83. [Google Scholar] [CrossRef]
- Liu, H.; Li, Z.; Tan, B.; Lao, Y.; Duan, Z.; Sun, W.; Dong, X. Isolation of a putative probiotic strain S12 and its effect on growth performance, non-specific immunity and disease-resistance of white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2014, 41, 300–307. [Google Scholar] [CrossRef]
- El-Son, M.A.; Elshopakey, G.E.; Rezk, S.; Eldessouki, E.A.; Elbahnaswy, S. Dietary mixed Bacillus strains promoted the growth indices, enzymatic profile, intestinal immunity, and liver and intestinal histomorphology of Nile tilapia, Oreochromis niloticus. Aquac. Rep. 2022, 27, 101385. [Google Scholar] [CrossRef]
- Hassaan, M.S.; Soltan, M.A.; Ghonemy, M.M.R. Effect of synbiotics between Bacillus licheniformis and yeast extract on growth, hematological and biochemical indices of the Nile tilapia (Oreochromis niloticus). Egypt. J. Aquat. Res. 2014, 40, 199–208. [Google Scholar] [CrossRef]
- C De, B.; Meena, D.K.; Behera, B.K.; Das, P.; Das Mohapatra, P.K.; Sharma, A.P. Probiotics in fish and shellfish culture: immunomodulatory and ecophysiological responses. Fish Physiol. Biochem. 2014, 40, 921–971. [Google Scholar] [CrossRef]
- Gullian, M.; Thompson, F.; Rodriguez, J. Selection of probiotic bacteria and study of their immunostimulatory effect in Penaeus vannamei. Aquaculture 2004, 233, 1–14. [Google Scholar] [CrossRef]
- Rodríguez, J.; Espinosa, Y.; Echeverría, F.; Cárdenas, G.; Román, R.; Stern, S. Exposure to probiotics and β-1, 3/1, 6-glucans in larviculture modifies the immune response of Penaeus vannamei juveniles and both the survival to White Spot Syndrome Virus challenge and pond culture. Aquaculture 2007, 273, 405–415. [Google Scholar] [CrossRef]
- Balcázar, J.L.; Rojas-Luna, T.; Cunningham, D.P. Effect of the addition of four potential probiotic strains on the survival of pacific white shrimp (Litopenaeus vannamei) following immersion challenge with Vibrio parahaemolyticus. J. Invertebr. Pathol. 2007, 96, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.T.; Maita, M.; Futami, K.; Endo, M.; Katagiri, T. Effects of a probiotic bacterial Lactobacillus rhamnosus dietary supplement on the crowding stress response of juvenile Nile tilapia Oreochromis niloticus. Fish. Sci. 2011, 77, 633–642. [Google Scholar] [CrossRef]
- Cerezuela, R.; Fumanal, M.; Tapia-Paniagua, S.T.; Meseguer, J.; Moriñigo, M.Á.; Esteban, M.Á. Changes in intestinal morphology and microbiota caused by dietary administration of inulin and Bacillus subtilis in gilthead sea bream (Sparus aurata L.) specimens. Fish Shellfish Immunol. 2013, 34, 1063–1070. [Google Scholar] [CrossRef]
- Meena, D.K.; Das, P.; Kumar, S.; Mandal, S.C.; Prusty, A.K.; Singh, S.K. . & Mukherjee, S.C. Beta-glucan: an ideal immunostimulant in aquaculture (a review). Fish Physiol. Biochem. 2013, 39, 431–457. [Google Scholar] [PubMed]
- Hoseinifar, S.H.; Sun, Y.Z.; Wang, A.; Zhou, Z. Probiotics as means of diseases control in aquaculture, a review of current knowledge and future perspectives. Front. Microbiol. 2018, 9, 2429. [Google Scholar] [CrossRef]
- Ghosh, A.K. Functionality of probiotics on the resistance capacity of shrimp against white spot syndrome virus (WSSV). 2023, Fish & shellfish immunology 2023, 108942. [Google Scholar]
- El-Saadony, M.T.; Swelum, A.A.; Ghanima, M.M.A.; Shukry, M.; Omar, A.A.; Taha, A.E. . & Abd El-Hack, M.E. Shrimp production, the most important diseases that threaten it, and the role of probiotics in confronting these diseases: a review. Res. Vet. Sci. 2022, 144, 126–140. [Google Scholar]
- Rengpipat, S.; Phianphak, W.; Piyatiratitivorakul, S.; Menasveta, P. Effects of a probiotic bacterium on black tiger shrimp Penaeus monodon survival and growth. Aquaculture 1998, 167, 301–313. [Google Scholar] [CrossRef]
- Kewcharoen, W.; Srisapoome, P. Probiotic effects of Bacillus spp. from Pacific white shrimp (Litopenaeus vannamei) on water quality and shrimp growth, immune responses, and resistance to Vibrio parahaemolyticus (AHPND strains). Fish Shellfish Immunol. 2019, 94, 175–189. [Google Scholar] [CrossRef]
- Tseng, D.Y.; Ho, P.L.; Huang, S.Y.; Cheng, S.C.; Shiu, Y.L.; Chiu, C.S.; Liu, C.H. Enhancement of immunity and disease resistance in the white shrimp, Litopenaeus vannamei, by the probiotic, Bacillus subtilis E20. Fish & shellfish immunology 2009, 26, 339–344. [Google Scholar]
- Li, J.; Tan, B.; Mai, K. Dietary probiotic Bacillus OJ and isomaltooligosaccharides influence the intestine microbial populations, immune responses and resistance to white spot syndrome virus in shrimp (Litopenaeus vannamei). Aquaculture 2009, 291, 35–40. [Google Scholar] [CrossRef]
- Guo, J.J.; Liu, K.F.; Cheng, S.H.; Chang, C.I.; Lay, J.J.; Hsu, Y.O. . & Chen, T.I. Selection of probiotic bacteria for use in shrimp larviculture. Aquac. Res. 2009, 40, 609–618. [Google Scholar]
- Shen, W.Y.; Fu, L.L.; Li, W.F.; Zhu, Y.R. Effect of dietary supplementation with Bacillus subtilis on the growth, performance, immune response and antioxidant activities of the shrimp (Litopenaeus vannamei). Aquac. Res. 2010, 41, 1691–1698. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Khalil, R.H.; Nour, A.M.; Elkhayat, B.K.; Khalifa, E.; Abdel-Latif, H.M. Effects of Bacillus subtilis-fermented rice bran on water quality, performance, antioxidants/oxidants, and immunity biomarkers of White leg shrimp (Litopenaeus vannamei) reared at different salinities with zero water exchange. J. Appl. Aquac. 2022, 34, 332–357. [Google Scholar] [CrossRef]
- Won, S.; Hamidoghli, A.; Choi, W.; Bae, J.; Jang, W.J.; Lee, S.; Bai, S.C. Evaluation of potential probiotics Bacillus subtilis WB60, Pediococcus pentosaceus, and Lactococcus lactis on growth performance, immune response, gut histology and immune-related genes in whiteleg shrimp, Litopenaeus vannamei. Microorganisms 2020, 8, 281. [Google Scholar] [CrossRef]
- Tepaamorndech, S.; Chantarasakha, K.; Kingcha, Y.; Chaiyapechara, S.; Phromson, M.; Sriariyanun, M. . & Visessanguan, W. Effects of Bacillus aryabhattai TBRC8450 on vibriosis resistance and immune enhancement in Pacific white shrimp, Litopenaeus vannamei. Fish Shellfish. Immunol. 2019, 86, 4–13. [Google Scholar]
- Liu, K.F.; Chiu, C.H.; Shiu, Y.L.; Cheng, W.; Liu, C.H. Effects of the probiotic, Bacillus subtilis E20, on the survival, development, stress tolerance, and immune status of white shrimp, Litopenaeus vannamei larvae. Fish Shellfish Immunol. 2010, 28, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Olmos, J.; Acosta, M.; Mendoza, G.; Pitones, V. Bacillus subtilis, an ideal probiotic bacterium to shrimp and fish aquaculture that increase feed digestibility, prevent microbial diseases, and avoid water pollution. Arch. Microbiol. 2020, 202, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Dong, Hong-biao, Yong-quan Su, Yong Mao, Xin-xin You, Shao-xiong Ding, and Jun Wang. "Dietary supplementation with Bacillus can improve the growth and survival of the kuruma shrimp Marsupenaeus japonicus in high-temperature environments." Aquaculture international 2014, 22, 607–617.
- Zokaeifar, H.; Babaei, N.; Saad, C.R.; Kamarudin, M.S.; Sijam, K.; Balcazar, J.L. Administration of Bacillus subtilis strains in the rearing water enhances the water quality, growth performance, immune response, and resistance against Vibrio harveyi infection in juvenile white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2014, 36, 68–74. [Google Scholar] [CrossRef]
- Sadat Hoseini Madani, N.; Adorian, T.J.; Ghafari Farsani, H.; Hoseinifar, S.H. The effects of dietary probiotic Bacilli (Bacillus subtilis and Bacillus licheniformis) on growth performance, feed efficiency, body composition and immune parameters of whiteleg shrimp (Litopenaeus vannamei) postlarvae. Aquac. Res. 2018, 49, 1926–1933. [Google Scholar] [CrossRef]
- Nimrat, S.; Khaopong, W.; Sangsong, J.; Boonthai, T.; Vuthiphandchai, V. Improvement of growth performance, water quality and disease resistance against Vibrio harveyi of postlarval whiteleg shrimp (Litopenaeus vannamei) by administration of mixed microencapsulated Bacillus probiotics. Aquac. Nutr. 2020, 26, 1407–1418. [Google Scholar] [CrossRef]
- Suguna, P.; Binuramesh, C.; Abirami, P.; Saranya, V.; Poornima, K.; Rajeswari, V.; Shenbagarathai, R. Immunostimulation by poly-β hydroxybutyrate–hydroxyvalerate (PHB–HV) from Bacillus thuringiensis in Oreochromis mossambicus. Fish Shellfish Immunol. 2014, 36, 90–97. [Google Scholar] [CrossRef]
- Hassan, M.A.; Bakhiet, E.K.; Ali, S.G.; Hussien, H.R. Production and characterization of polyhydroxybutyrate (PHB) produced by Bacillus sp. isolated from Egypt. J. Appl. Pharm. Sci. 2016, 6, 046–051. [Google Scholar] [CrossRef]
- Laranja, J.L.Q.; Ludevese-Pascual, G.L.; Amar, E.C.; Sorgeloos, P.; Bossier, P.; De Schryver, P. Poly-β-hydroxybutyrate (PHB) accumulating Bacillus spp. improve the survival, growth and robustness of Penaeus monodon () postlarvae. Vet. Microbiol. 2014, 173, 310–317. [Google Scholar] [CrossRef]
- Cai, Y.; Yuan, W.; Wang, S.; Guo, W.; Li, A.; Wu, Y. . & Zhou, Y. In vitro screening of putative probiotics and their dual beneficial effects: to white shrimp (Litopenaeus vannamei) postlarvae and to the rearing water. Aquaculture 2019, 498, 61–71. [Google Scholar]
- Monier, M.N.; Kabary, H.; Elfeky, A.; Saadony, S.; El-Hamed, N.N.A.; Eissa, M.E.; Eissa, E.S.H. The effects of Bacillus species probiotics (Bacillus subtilis and B. licheniformis) on the water quality, immune responses, and resistance of whiteleg shrimp (Litopenaeus vannamei) against Fusarium solani infection. Aquaculture International 2023, 1–19. [Google Scholar]
- Dawood, M.A.; Koshio, S.; Abdel-Daim, M.M.; Van Doan, H. Probiotic application for sustainable aquaculture. Rev. Aquac. 2019, 11, 907–924. [Google Scholar] [CrossRef]
- Xie, J.J.; Liu, Q.Q.; Liao, S.; Fang, H.H.; Yin, P.; Xie, S.W. . & Niu, J. Effects of dietary mixed probiotics on growth, non-specific immunity, intestinal morphology and microbiota of juvenile pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 90, 456–465. [Google Scholar] [PubMed]
- Bhatnagar, A.; Lamba, R. Antimicrobial ability and growth promoting effects of feed supplemented with probiotic bacterium isolated from gut microflora of Cirrhinus mrigala. J. Integr. Agric. 2015, 14, 583–592. [Google Scholar] [CrossRef]
- Devaraja, T.; Banerjee, S.; Yusoff, F.; Shariff, M.; Khatoon, H. A holistic approach for selection of Bacillus spp. as a bioremediator for shrimp postlarvae culture. Turk. J. Biol. 2013, 37, 92–100. [Google Scholar] [CrossRef]
- Nimrat, S.; Suksawat, S.; Boonthai, T.; Vuthiphandchai, V. Potential Bacillus probiotics enhance bacterial numbers, water quality and growth during early development of white shrimp (Litopenaeus vannamei). Vet. Microbiol. 2012, 159, 443–450. [Google Scholar] [CrossRef]
- Bachruddin, M.; Sholichah, M.; Istiqomah, S.; Supriyanto, A. Effect of probiotic culture water on growth, mortality, and feed conversion ratio of Vaname shrimp (Litopenaeus vannamei Boone). In IOP Conference Series: Earth and Environmental Science; IOP Publishing: 2018; Vol. 137, p. 0120. [Google Scholar]
- Kongnum, K.; Hongpattarakere, T. Effect of Lactobacillus plantarum isolated from digestive tract of wild shrimp on growth and survival of white shrimp (Litopenaeus vannamei) challenged with Vibrio harveyi. Fish Shellfish Immunol. 2012, 32, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Tamilarasu, A.; Ahilan, B.; Gopalakannan, A.; Somu Sunder Lingam, R. Evaluation of probiotic potential of Bacillus strains on growth performance and physiological responses in Penaeus vannamei. Aquac. Res. 2021, 52, 3124–3136. [Google Scholar] [CrossRef]
- Kaynar, P.; Beyatli, Y. Determination of poly-β-hydroxybutyrate production by Bacillus spp. isolated from the intestines of various fishes. Fish. Sci. 2009, 75, 439–443. [Google Scholar] [CrossRef]
- Hao, K.; Liu, J.Y.; Ling, F.; Liu, X.L.; Lu, L.; Xia, L.; Wang, G.X. Effects of dietary administration of Shewanella haliotis D4, Bacillus cereus D7 and Aeromonas bivalvium D15, single or combined, on the growth, innate immunity and disease resistance of shrimp, Litopenaeus vannamei. Aquaculture 2014, 428, 141–149. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, M.; Li, X.; Han, Y.; Wu, F.; Liu, Y. Effects of a probiotic (Bacillus licheniformis) on the growth, immunity, and disease resistance of Haliotis discus hannai Ino. Fish Shellfish Immunol. 2018, 76, 143–152. [Google Scholar] [CrossRef]
- Xiaolong, Gao, Ke Caihuan, Zhang Mo, Li Xian, Wu Fucun, and Liu Ying. "Effects of the probiotic Bacillus amyloliquefaciens on the growth, immunity, and disease resistance of Haliotis discus hannai." Fish & shellfish immunology 2019, 94, 617–627.
- Sánchez-Ortiz, A.C.; Angulo, C.; Luna-González, A.; Álvarez-Ruiz, P.; Mazón-Suástegui, J.M.; Campa-Córdova, Á.I. Effect of mixed-Bacillus spp isolated from pustulose ark Anadara tuberculosa on growth, survival, viral prevalence and immune-related gene expression in shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2016, 59, 95–102. [Google Scholar] [CrossRef]
- Xiaolong, G.; Caihuan, K.; Fucun, W.; Xian, L.; Ying, L. Effects of Bacillus lincheniformis feeding frequency on the growth, digestion and immunity of Haliotis discus hannai. Fish Shellfish Immunol. 2020, 96, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zhu, J.; Hu, J.; Dong, X.; Sun, L.; Zhang, X.; Miao, S. Effects of dietary Bacillus pumilus on growth performance, innate immunity and digestive enzymes of giant freshwater prawns (Macrobrachium rosenbergii). Aquac. Nutr. 2019, 25, 712–720. [Google Scholar] [CrossRef]
- Dalmin, G.; Kathiresan, K.; Purushothaman, A. Effect of probiotics on bacterial population and health status of shrimp in culture pond ecosystem, 2001.
- Matias, H.B.; Yusoff, F.M.; Shariff, M.; Azhar, O. Effects of commercial microbial products on water quality in tropical shrimp culture ponds. Asian Fish. Sci. 2002, 15, 239–248. [Google Scholar] [CrossRef]
- Wang, Y.B.; Xu, Z.R.; Xia, M.S. The effectiveness of commercial probiotics in northern white shrimp Penaeus vannamei ponds. Fish. Sci. 2005, 71, 1036–1041. [Google Scholar] [CrossRef]
- Reddy, K.V.; Reddy, A.V.K.; Babu, B.S.; Lakshmi, T.V. Applications of Bacillus sp in aquaculture waste water treatment. Int JS Res Sci. Tech 2018, 4, 1806–1812. [Google Scholar]
- Hu, Xiaojuan, Yu Xu, Haochang Su, Wujie Xu, Guoliang Wen, Chuangwen Xu, Keng Yang, Song Zhang, and Yucheng Cao. "Effect of a Bacillus Probiotic Compound on Penaeus vannamei Survival, Water Quality, and Microbial Communities." Fishes 2023, 8, 362.
- Phung, L.T.; Phung, N.K.; Phuong, T.T.M.; Nicolas, M.; Vincent, M.; Sandra, A.; Philippe, D.S.J. Effect of Bacillus sp. as probiotic on the treatment of environment in brackish water shrimp aquaculture. In IOP Conference Series: Materials Science and Engineering; IOP Publishing: 2020; Volume 991, p. 012052.
- Decamp, O.; Moriarty, D.J.; Lavens, P. Probiotics for shrimp larviculture: review of field data from Asia and Latin America. Aquac. Res. 2008, 39, 334–338. [Google Scholar] [CrossRef]
- Mujeeb Rahiman, K.M.; Jesmi, Y.; Thomas, A.P.; Mohamed Hatha, A.A. Probiotic effect of Bacillus NL110 and Vibrio NE17 on the survival, growth performance and immune response of Macrobrachium rosenbergii (de Man). Aquac. Res. 2010, 41, e120–e134. [Google Scholar] [CrossRef]
- de Paiva-Maia, E.; Alves-Modesto, G.; Otavio-Brito, L.; Vasconcelos-Gesteira, T.C.; Olivera, A. Effect of a commercial probiotic on bacterial and phytoplankton concentration in intensive shrimp farming (Litopenaeus vannamei) recirculation systems. Lat. Am. J. Aquat. Res. 2013, 41, 126–137. [Google Scholar] [CrossRef]
- Wu, D.X.; Zhao, S.M.; Peng, N.; Xu, C.P.; Wang, J.; Liang, Y.X. Effects of a probiotic (B acillus subtilis FY99-01) on the bacterial community structure and composition of shrimp (L itopenaeus vannamei, B oone) culture water assessed by denaturing gradient gel electrophoresis and high-throughput sequencing. Aquac. Res. 2016, 47, 857–869. [Google Scholar] [CrossRef]
- Hoang, H.G.; Thuy, B.T.P.; Lin, C.; Vo, D.V.N.; Tran, H.T.; Bahari, M.B.; Vu, C.T. The nitrogen cycle and mitigation strategies for nitrogen loss during organic waste composting: A review. Chemosphere 2022, 300, 134514. [Google Scholar] [CrossRef]
- Kamilya, D.; Devi, W.M. Bacillus probiotics and bioremediation: an aquaculture perspective. In Bacilli in Agrobiotechnology: Plant Stress Tolerance, Bioremediation, and Bioprospecting; Springer International Publishing: Cham, Switzerland, 2022. [Google Scholar]
- Sineva, E.V.; Andreeva-Kovalevskaya, Z.I.; Shadrin, A.M.; Gerasimov, Y.L.; Ternovsky, V.I.; Teplova, V.V. . & Solonin, A.S. Expression of Bacillus cereus hemolysin II in Bacillus subtilis renders the bacteria pathogenic for the crustacean Daphnia magna. FEMS Microbiol. Lett. 2009, 299, 110–119. [Google Scholar]
- Velmurugan, S.; Palanikumar, P.; Velayuthani, P.; Donio, M.B.S.; Babu, M.M.; Lelin, C. . & Citarasu, T. Bacterial white patch disease caused by Bacillus cereus, a new emerging disease in semi-intensive culture of Litopenaeus vannamei. Aquaculture 2015, 444, 49–54. [Google Scholar]
- Hendriksen, N.B.; Hansen, B.M.; Johansen, J.E. Occurrence and pathogenic potential of Bacillus cereus group bacteria in a sandy loam. Antonie Van Leeuwenhoek 2006, 89, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Andreeva, Z.I.; Nesterenko, V.F.; Fomkina, M.G.; Ternovsky, V.I.; Suzina, N.E.; Bakulina, A.Y. . & Sineva, E.V. The properties of Bacillus cereus hemolysin II pores depend on environmental conditions. Biochim. Et Biophys. Acta (BBA)-Biomembr. 2007, 1768, 253–263. [Google Scholar]
- Elshaghabee, F.M.; Rokana, N.; Gulhane, R.D.; Sharma, C.; Panwar, H. Bacillus as potential probiotics: status, concerns, and future perspectives. Front. Microbiol. 2017, 8, 1490. [Google Scholar] [CrossRef] [PubMed]
- Ouwehand, A.C.; Forssten, S.; Hibberd, A.A.; Lyra, A.; Stahl, B. Probiotic approach to prevent antibiotic resistance. Ann. Med. 2016, 48, 246–255. [Google Scholar] [CrossRef]
- Bhogoju, S.; Nahashon, S. Recent advances in probiotic application in animal health and nutrition: A review. Agriculture 2022, 12, 304. [Google Scholar] [CrossRef]
- Anokyewaa, M.A.; Amoah, K.; Li, Y.; Lu, Y.; Kuebutornye, F.K.; Asiedu, B.; Seidu, I. Prevalence of virulence genes and antibiotic susceptibility of Bacillus used in commercial aquaculture probiotics in China. Aquaculture Reports 2021, 21, 100784. [Google Scholar] [CrossRef]
- Roe, A.L.; Boyte, M.E.; Elkins, C.A.; Goldman, V.S.; Heimbach, J.; Madden, E. ;... & Smith, A. Considerations for determining safety of probiotics: A USP perspective. Regulatory Toxicology and Pharmacology 2022, 105266. [Google Scholar]
- Doan, H.V.; Soltani, E.; Ingelbrecht, J.; Soltani, M. Medicinal herbs and plants: Potential treatment of monogenean infections in fish. Rev. Fish. Sci. Aquac. 2020, 28, 260–282. [Google Scholar] [CrossRef]
- Anadón, A.; Ares, I.; Martínez-Larrañaga, M.R.; Martínez, M.A. Probiotics: safety and toxicity considerations. In Nutraceuticals; Academic Press: Cambridge, MA, USA, 2021; pp. 1081–1105. [Google Scholar]
- Jackson, S.A.; Schoeni, J.L.; Vegge, C.; Pane, M.; Stahl, B.; Bradley, M. ;... & Sanders, M.E. Improving end-user trust in the quality of commercial probiotic products. Front. Microbiol. 2019, 739. [Google Scholar]
- James, G.; Das, B.C.; Jose, S.; VJ, R.K. Bacillus as an aquaculture friendly microbe. Aquac. Int. 2021, 29, 323–353. [Google Scholar] [CrossRef]
- Mathipa, M.G.; Thantsha, M.S. Probiotic engineering: towards development of robust probiotic strains with enhanced functional properties and for targeted control of enteric pathogens. Gut Pathog. 2017, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Probiotic Bacillus effects on the environment and fin and shell fish Adopted from [7].
Figure 1.
Probiotic Bacillus effects on the environment and fin and shell fish Adopted from [7].
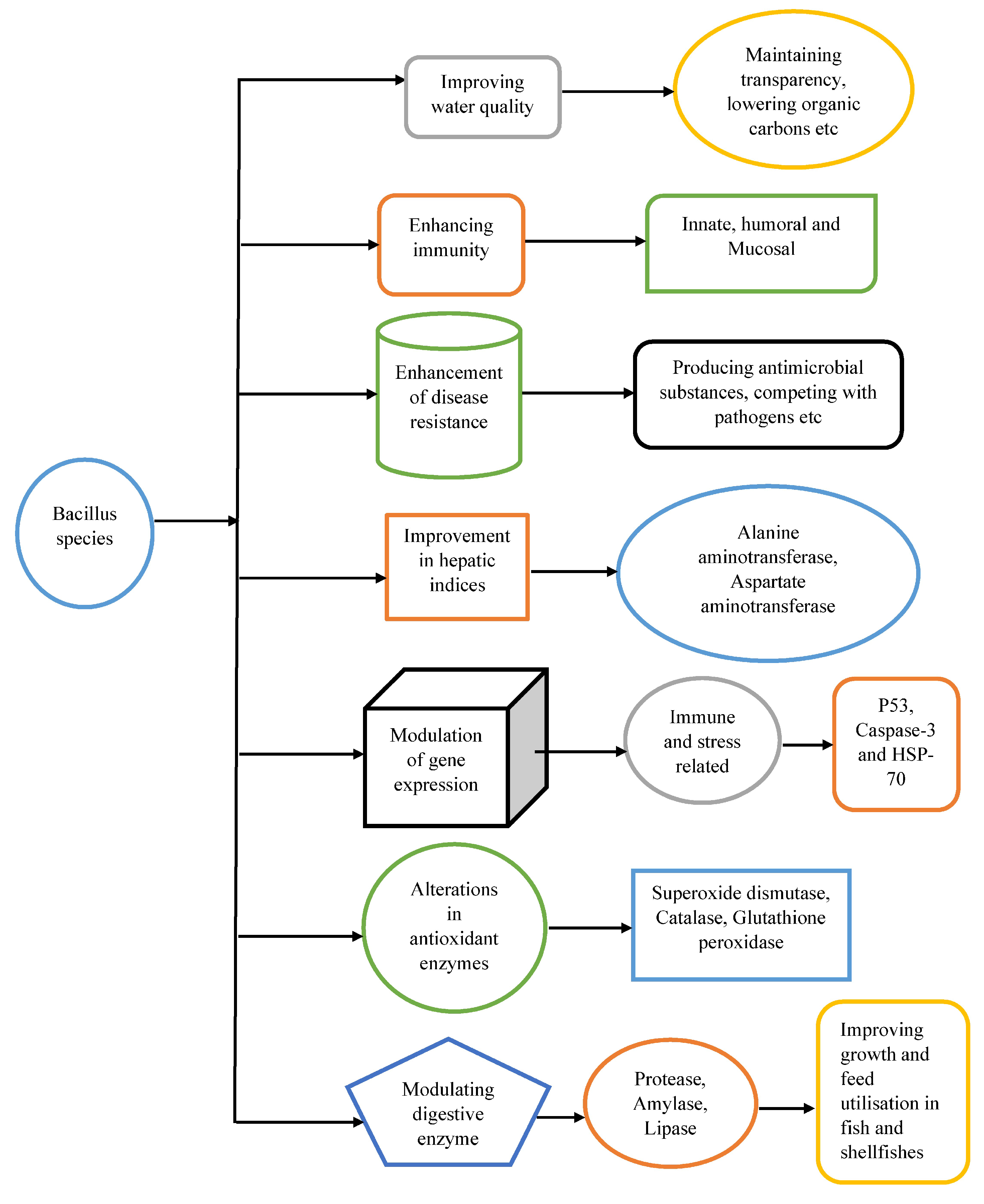
Figure 2.
Features of Bacillus to enhance immunity and health in fin and shellfish as well as commercial production of feed. Adopted from [27].
Figure 2.
Features of Bacillus to enhance immunity and health in fin and shellfish as well as commercial production of feed. Adopted from [27].
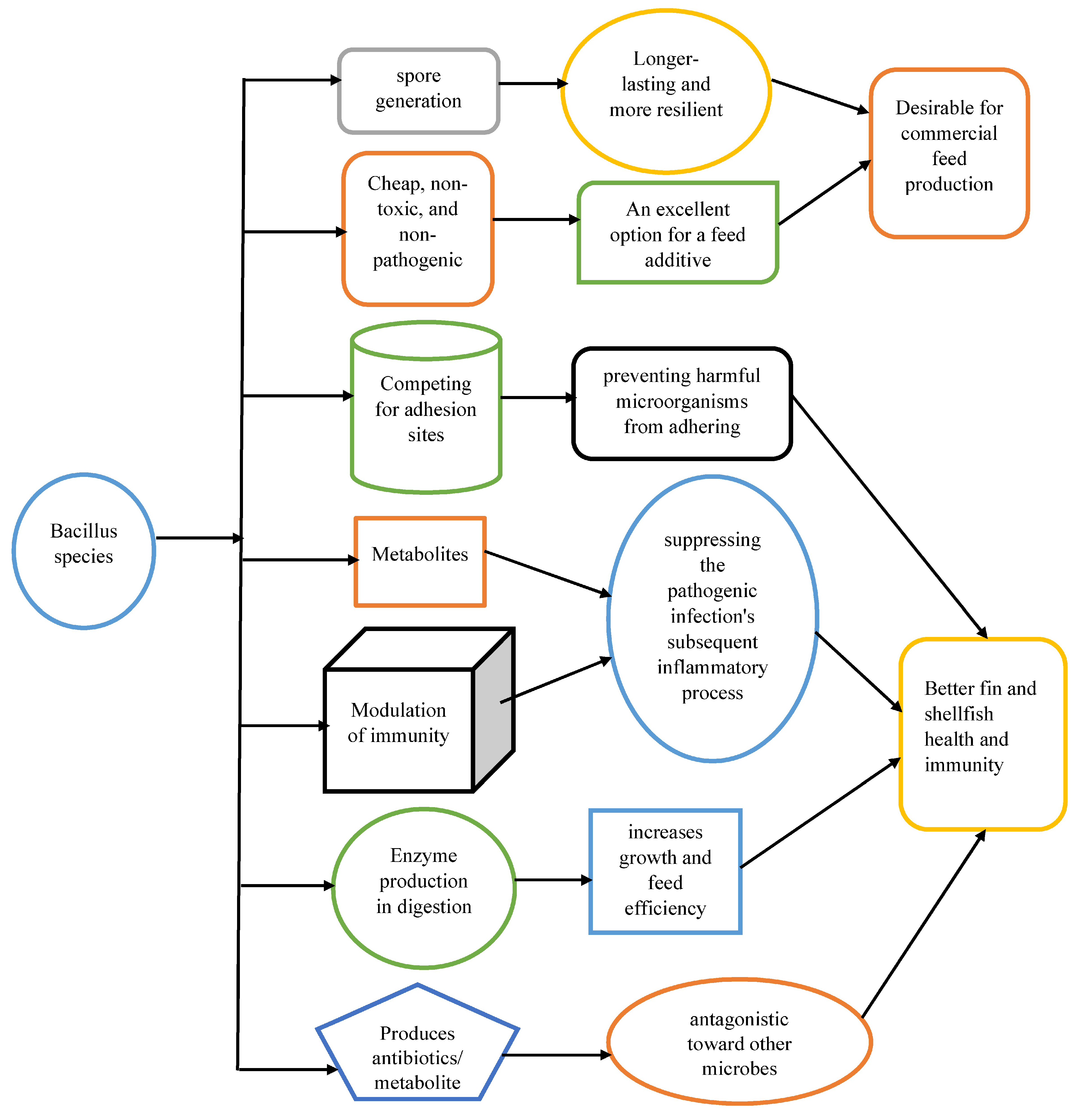
Table 1.
Commercial products of Bacillus for fin and shell fish.
| Bacillus probiotic for Finfishes aquaculture | ||
|---|---|---|
| Brand | Manufacturer | Comments |
| Naturalle Bacillus subtilis | Wuhan Nature´s Favour Bioengineering Co.; Ltd, Wuhan City, China http://www.wuhannature.com | Bacillus subtilis (2x1010 CFU/g). |
| Biozyme | Bio-Pharmachemie Joint-Venture Company, Ho Chi Minh, Vietnam http://www.biopharmachemie.com | Bacillus subtilis and Saccaromyces cerevisiae. |
| Fubon B. subtilis | Angel Yeast Co.; Ltd. Hubei, China http://www.angelyeast.com | Bacillus subtilis (≥ 20 billion CFU/g). |
| Bioron | American Pharma International, India http://www.americanpharmainternational.com | Each kg contains: B. subtilis (4.5x108 CFU), B. licheniformis, B. megaterium, Lactobacillus lactis, L. helveticus, Nitrosomonas sp. Nitrobacter sp. Saccharomyces cerevisiae and Aspergillus oryzae. |
| Lactomin | American Pharma International, India http://www.americanpharmainternational.com | Each kg contains: B. subtilis (45,000 million CFU), B. licheniformis, Lactobacillus acidophilus, L. sporogenes and Saccharomyces cerevisiae. |
| Bacillus probiotic for shellfishes aquaculture | ||
| Brand | Composition | Dose |
| Aqua photo | Bacillus subtilis and Rhodopseudomonas | 50–70 ml/100 dec. Control unwanted gas, sediment and increase growth of plankton |
| Bio-zyme | Bacillus subtilis, Saccharomyces cerevisiae | 500 g/100 kg feed |
| Eco marine | Bacillus subtilis, Bacillus pumilis, Bacillus amyloliquefaciens, Bacillus megaterium | 3–4 tablet/acre |
| Golden Bac | Yeast,Bacillus subtilis, Lactobacillus sp. | 1.5–2 kg/acre |
| pH fixer | Bacillus sp. | 1–2 kg/acre |
| Procon-PS | Bacillus sp. Rhodococcus, and Rhodobacter | 5 L/hac (l m depth) |
| Super Biotic | Bacillus sp. | 1–2 kg/acre |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



