
Submitted:
15 June 2024
Posted:
17 June 2024
You are already at the latest version
Abstract
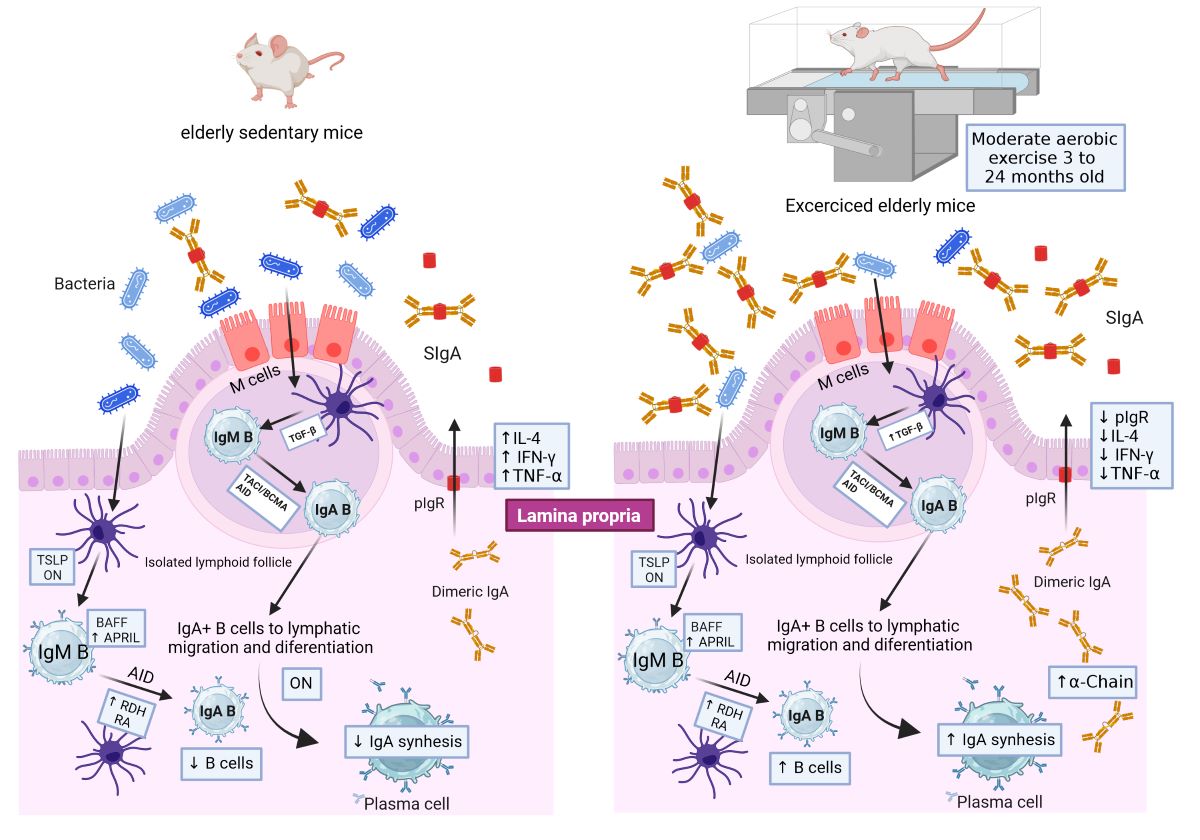
T-cell independent (TI) pathway activated by microbiota results in the generation of low-affinity homeostatic IgA with a critical role in intestinal homeostasis. Moderate aerobic exercise (MAE) provides a beneficial impact on intestinal immunity but the action of MAE on TI-IgA generation under senescence conditions is unknown. This study aimed to determine the effects of long-term MAE on TI-IgA production in young (3 months old) BALB/c mice exercised until adulthood (6 months) or aging (24 months). Lamina propria (LP) from the small intestine was obtained to determine B cells and plasma cells sub-populations by flow cytometry and molecular factors related to class switch recombination (TSLP/APRIL/BAFF/iNOS/RDH) and synthesis of IgA (α-chain/IL-6/IL-21/TGF-β); and epithelial cells to evaluate IgA-transitosis (pIgR/TNFα/IFN-/IL-4), by RT-qPCR technique. Results were compared with data obtained from sedentary age-matched mice. Statistical analysis was computed with ANDOVA and P<0.05 was considered as statistically significant difference. Under senescence conditions MAE promoted the B cell and IgA+B cells and APRIL that may im-prove the intestinal response and ameliorated the inflammatory environment associated pre-sumably with the downmodulation of pIgR. Data suggested that MAE improved the IgA and downmodulate the cytokine pro-inflammatory expression favoring homeostatic conditions in aging.
Keywords:
aerobic moderate exercise
; aging
; intestinal IgA
; T-independent cell pathway
; lamina propria
; epithelial cells
1. Introduction
Immunoglobulin A (IgA) is a key player in mucosal immunity by participating in protection against pathogens that colonize and/or invade the luminal surface and by collaborating in intestinal homeostasis [1]. Synthesis of IgA entails both T-cell dependent (TD) and T-cell independent (TI) pathways [2]. Intestinal IgA synthesis via T-cell dependent and T-cell independent pathways are under the control of intestinal microbiota[3,4] While most colonic commensals induced T-independent IgA generation, atypical commensals such as filamentous bacteria and Mucispirilum induced T-dependent IgA generation [3,4,5].
T-cell independent pathway drives the constitutive IgA synthesis stimulated by luminal bacteria rendering low-affinity IgA and takes place in isolated lymphoid follicles (ILFs), lamina propria, and Peyer´s patches (PP) in absence of germinal center [6]. M cells transport luminal bacteria in the ILFs through a mechanism of transcytosis and then dendritic cells (DC) CD103+ recognize the luminal bacterial antigens to be delivered to B cells via B cell receptor (BCR) in the epithelial dome. This recognition induces signal pathways in näive B cells (IgM+ B cells) leading to the Class Switch Recombination (CSR) of IgM+ B cells to IgA+ B cells under the stimulation of regulatory molecules released by DC CD103+ including B Cell Activating Factor (BAFF), and the A Proliferation-Inducing Ligand (APRIL), and by epithelial cells (EC) such as tumor necrosis factor-α (TNFα), Transforming Growth Factor-β1 (TGF-β1), and Thymic Stromal Lymphopoietin (TSLP). Additionally, T-cell independent CSR factors include retinoic acid (RA) and nitric oxide (NO). Retinoic acid is obtained from the diet, biliary acids, or/and endogenously by retinal dehydrogenase (RDH) expressed in DC CD103+ stimulated with flagellin via TLR5 signal. Nitric oxide is generated by inducible nitric oxide synthase (iNOS) [7].
In lamina propria T-cell independent IgA generation is led by cytokines released from DCs such as TNFα-induced NO production. Nitric oxide enhances the expression of TGF-β1 receptors on B cells to promote IgA synthesis. In addition, EC stimulates the recruitment of B cells to lamina propria through an expression of CCL20 and CCL28. Also, epithelial cells recognize luminal antigens through TLRs promoting the production of TSLP, another CRS stimulatory factor that favors APRIL synthesis from lamina propria DC to be targeted to B cells for IgA synthesis [8].
In addition, others transcription factors are involved in B-cell development and differentiation impacting on IgA production, such as, Core-Binding Factor Alpha 3 (CBFa3/RUNX-3), Upstream Stimulatory Factor (USF) and paired box-5 (Pax5). Furthermore, Pax5 also is critical for activating specific genes in pro-B cells or repressing genes in mature B lymphocytes and terminally differentiated IgA plasma cells as well as IgA class switching [9].
Polymeric IgA produced by plasma cells binds to the polymeric immunoglobulin receptor (pIgR) expressed by epithelial cells [10]. IgA binds to pIgR on the basolateral surface of EC and is internalized in endosomes. It is then0 transported in vesicles to the apical surface. Subsequently, it is proteolytically cleaved, and the extracellular fragment called the secretory component (CS), is released with the IgA ligand. CS is covalently bound to an antibody portion and constitutes an integral part of the secretory IgA (SIgA) complex [4].
Modulation of IgA synthesis by intestinal microbiota is under control of aging and exercise [11] and aging [12]. In humans aging decreased taxon-specific IgA Bifidobacteriaceae and increased pathobionts such as Enterobacteriaceae [12]. In mice, aging increased in potential pathobionts (Desulfovibrio and Bilophila, and decreased IgA-associated and health-promoting bacteria (Akkermansia spp. Lactobacillus spp. and Bifidobacterium spp, [13]. Contrarily, exercise induces the abundance of microbiota members like bifidobacteria and lactobacilli associated with IgA synthesis [14].
Aging of the immune system, or immunesenescence, leads to systemic deterioration with changes in the level of immunoglobulins and cytokines, which decreases the ability to react appropriately against pathogens, making us more susceptible to infections and other health problems [15,16]. Aging has subtle and complex effects on SIgA levels: average SIgA levels tend to increase with age until 60 years, and then decrease slightly [17].
Scarce evidence of aging on intestinal IgA production has shown that total IgA concentration in intraluminal secretions of the small intestine from old rats is similar to adult rats [18]. Furthermore, aging increases monomeric IgA without changes in dimeric IgA in the intestinal lumen, but the binding capacity of dimeric IgA to bacteria is also reduced, which could be reflected in the quality and functionality of intestinal IgA in aging [19]. In addition, a lower specific IgA response in jejunum and ileum from elderly rats immunized with choleric toxin has been identified [20]. Another study in mice immunized with cholera holotoxin also showed a higher number of IgA secreting-cells in lamina propria in both immunized and no immunized old mice in comparison to young mice; however, serum-specific IgA concentration was lower in old rats [21]. Despite the last results suggesting an effect of aging on IgA production by the T-dependent pathway, there is no evidence of aging-induced changes in IgA production by the TI pathway.
Regarding the effect of aging on transcytosis of IgA in the intestinal epithelium, showing that a low level of urine IgA in ddYHigh mice is caused by a deficiency of transcytosis of IgA in the intestinal mucosa. Therefore, the age-dependent changes in the pIgR expression but not in the IgA structure seem to be a possible cause for the elevation of serum IgA [22].
Currently, exercise has been promoted as a strategy to strengthen immunity against diseases associated with aging caused, in part, by the functional imbalance of the immune system, a process known as immune senescence [23,24,25]. The contributions regarding the effects of exercise on the intestinal immune response made by our working group showed that moderate exercise upmodulated the intestinal levels of IgA in the small intestine of adult mice [26,27]. Other trials showed that moderate exercise plus a final session of intense acute exercise promotes the increase of IgA however reduces the transcriptional expression of most mediators that promote its generation and transport in young adult mice (6 months of age) [28]. A mechanism through the exercise that can impact mucosa immunity is by activation of the hypothalamus-hypophysis-adrenal axis. For that, serum cortisol concentration was assayed observing that exercise significantly increased cortisol concentration in adult mice but significantly decreased in elderly mice from this study (data previously published in Sierra-Ramírez, et al. 2022) [29]. Last result let us consider exercise as a good strategy to decrease the impact of aging on overall health.
Although such findings indicate that exercise is an important modulator of intestinal immunity, the cellular and molecular mechanisms underlying its role under conditions of senescence remain unclear. This study aims, for the first time, to assess the effect of performing MAE from young age to aging on markers of the T cell-independent pathway involved in intestinal immunity. Findings may support the impact of exercise to promote and strengthen intestinal protection and homeostasis and to ameliorate the development of several diseases in the elderly.
2. Results
The results represent the effect of long-term MAE performed from young age (3 months old) to adulthood (6 months old) or senile (24 months old). All data were compared in regard young group considered as baseline parameters. The effect of age was analyzed by comparing elderly versus young and adult groups; the effect of exercise was seen by comparing exercised vs sedentary groups.
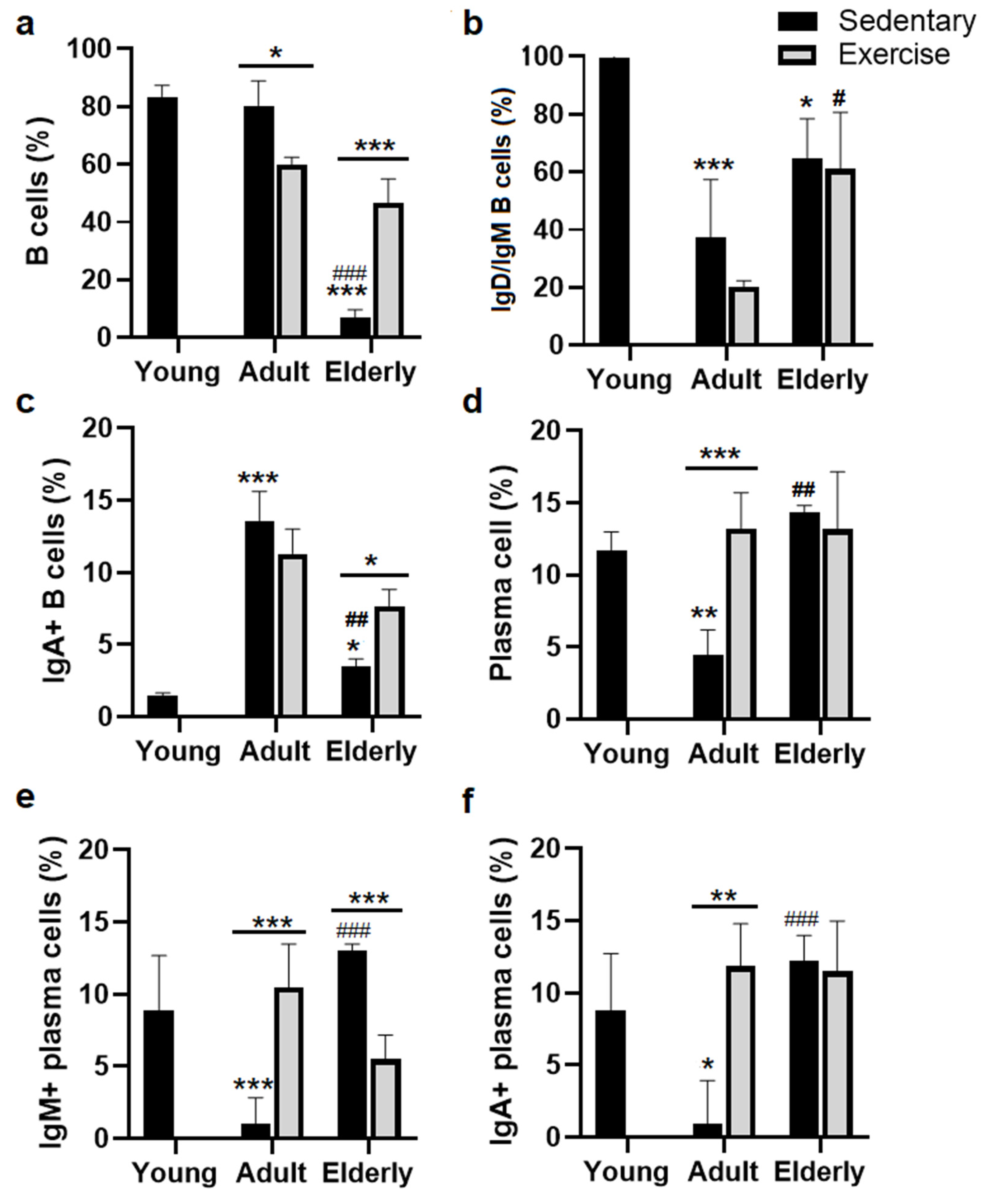
2.1. Exercise Increased total B Cells in Senile Mice.
Comparisons among the sedentary groups total B cell percentage (%) was decreased in the elderly than in young mice and adult mice (both p<0.001, see Figure 1a). Concerning their respective age-matched sedentary group, B cell % was either lower in adult exercised mice or higher in the senile exercised group (adult p<0.05, elderly p<0.001, Figure 1a). Within the sedentary groups, IgM+ B cell % was lower in adults and the elderly than in young mice (adult p<0.001, elderly p<0.05). Within the exercised groups, IgM+ B cell % was greater in elderly versus adult mice (p<0.05, Figure 1b).
2.2. Exercise Increased the IgA+ B Cells in Senile Mice
Analysis among the sedentary mice indicated that IgA+ B cell % was found greater in adult and elderly than young mice (adult p<0.001, elderly p<0.05, Figure 1c) and lower in elderly than adult mice (p<0.01, Figure 1c). Comparisons within the age-matched groups evidenced that IgA+ cell % was greater in the senile exercised than in the senile sedentary group (p<0.05, Figure 1c).
Analysis within sedentary groups indicated that plasma cell % was lower in adults than young mice or greater in senile than adult mice (both p<0.01, Figure 1d). Within the age-matched groups, plasma cell % was greater in adults exercised than in adult sedentary mice (p<0.001, Figure 1d).
Comparative analysis with the sedentary groups showed IgM+ plasma cell % was lower in adult versus young mice or higher in senile versus adult mice (both p<0.001, Figure 1e). Analysis within the age-matched groups indicated that IgM+ plasma cell % was higher in adult exercised versus adult sedentary mice or lower in aged exercised than aged sedentary mice (both p<0.001, Figure 1e)
Analysis within the sedentary mice evidenced that IgA+ plasma cell % was lower in adult than young mice or higher in senile than adult mice (adult p<0.05, senile p<0.001, Figure 1f). Within the age-matched groups, IgA+ plasma cell % was greater in adults exercised than in adult sedentary mice (p<0.01, Figure 1f).
Figure 1.
Lymphoid subpopulations in lamina propria from small intestine of sedentary and exercised mice at different ages. Data represent the mean and standard deviation of (a) total B cells; (b) membrane IgM+/IgD+ B cells; (c) membrane IgA+ B cells; (d) plasma cells; (e) cytoplasmic IgM+ plasma cells; (f) cytoplasmic IgA+ plasma cells. (*) upon bar versus young mice; (#) upon bar versus respective adult control; asterisks upon line on bars, versus respective sedentary control. *p<0.05, **p<0.01 and ***p<0.001, #p<0.05, ##p<0.01 and ###p<0.001.
Figure 1.
Lymphoid subpopulations in lamina propria from small intestine of sedentary and exercised mice at different ages. Data represent the mean and standard deviation of (a) total B cells; (b) membrane IgM+/IgD+ B cells; (c) membrane IgA+ B cells; (d) plasma cells; (e) cytoplasmic IgM+ plasma cells; (f) cytoplasmic IgA+ plasma cells. (*) upon bar versus young mice; (#) upon bar versus respective adult control; asterisks upon line on bars, versus respective sedentary control. *p<0.05, **p<0.01 and ***p<0.001, #p<0.05, ##p<0.01 and ###p<0.001.
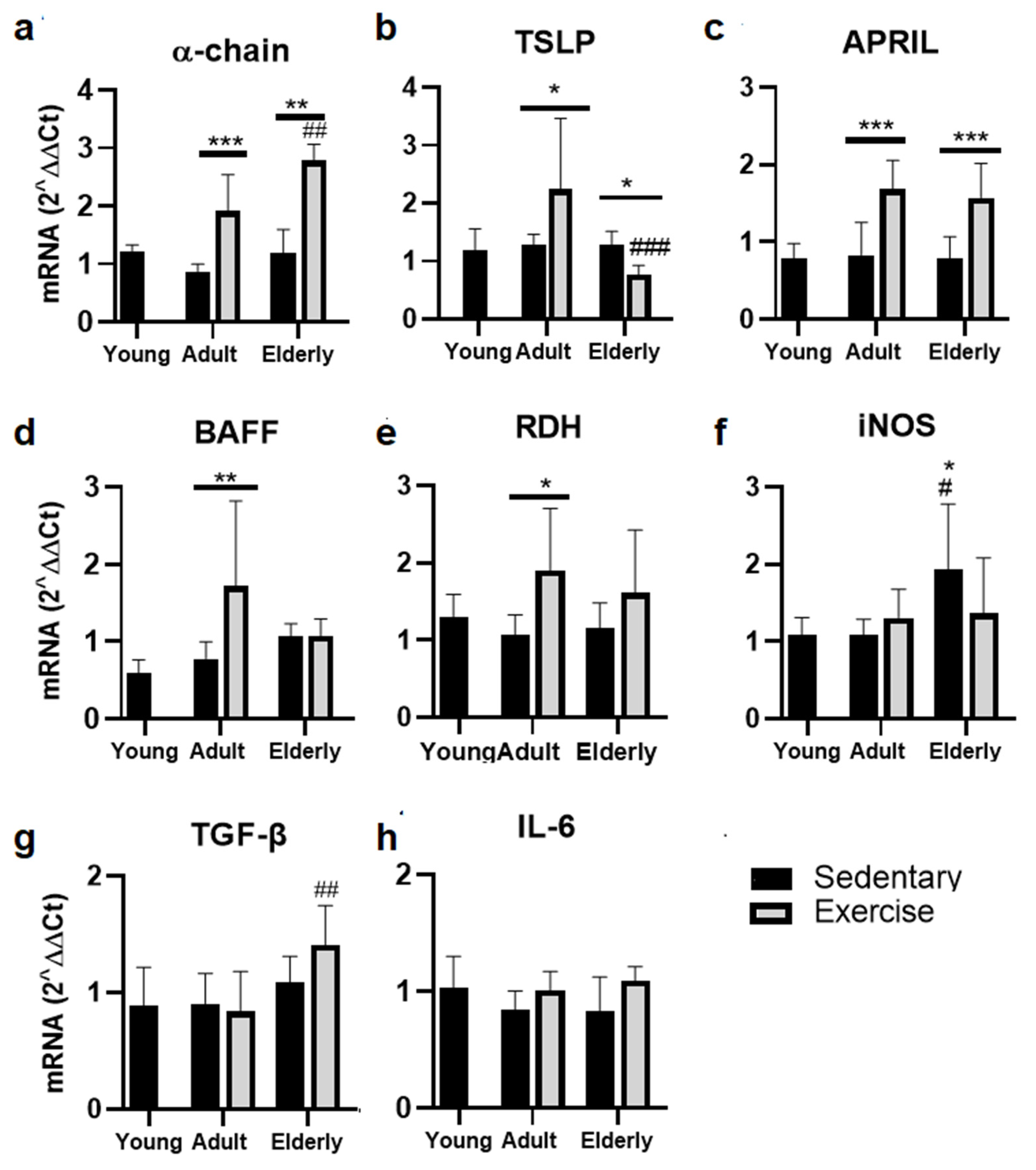
2.3. Exercise Increased the α-Chain and APRIL mRNA in Adult and Senile Mice
Within aged-paired groups, α-chain mRNA was found greater in the adult and senile exercised mice than their respective sedentary control groups (adult p<0.001, senile p<0.01, Figure 2a). Moreover, α-chain mRNA level was greater in senile exercised than adult exercised mice (p<0.01, Figure 2a)
Comparative analysis of TSLP mRNA evidenced a greater level in exercised than sedentary adult mice and in sedentary aged versus sedentary adult mice (both p<0.05); lower TSLP mRNA level was found in senile exercised than adult exercised mice (p<0.001, Figure 2b).
Analysis of APRIL mRNA evidenced that it was greater in adult and senile exercised mice versus their respective sedentary control groups (both p<0.001, Figure 2c).
Comparative analysis of BAFF and RDH mRNA within the adult mice evidenced greater levels in exercised than sedentary mice (BAFF p<0.01, Figure 2d; RDH p<0.05, Figure 2e). Within the sedentary groups, iNOS mRNA level was found greater in elderly versus young and adult mice (both p<0.05, Figure 2f). Within the exercised groups, TGF-β1 mRNA was seen higher in elderly than adult mice (p<0.01, Figure 2g). Regarding IL-6 mRNA no significant differences were seen (Figure 2h)
Figure 2.
Gene expression of factors related to production of IgA in lamina propria of small intestine from sedentary or exercise mice at different ages. Data represent the mean and standard deviation of (a) α-chain, (b) Thymic Stromal Lymphopoietin (TSLP), (c) A Proliferation-Inducing Ligand (APRIL), (d) B-cell Activating Factor (BAFF), (e) inducible nitric oxide synthase (iNOS), (f) retinol dehydrogenase (RDH), (g) transforming growth factor beta (TGF-β), (h) interleukin (IL)-6. (*) upon bar versus respective young mice; (#) upon bar versus respective adult control; asterisks upon line on bars, versus respective sedentary control. *p<0.05, **p<0.01 and ***p<0.001, #p<0.05, ##p<0.01 and ###p<0.001.
Figure 2.
Gene expression of factors related to production of IgA in lamina propria of small intestine from sedentary or exercise mice at different ages. Data represent the mean and standard deviation of (a) α-chain, (b) Thymic Stromal Lymphopoietin (TSLP), (c) A Proliferation-Inducing Ligand (APRIL), (d) B-cell Activating Factor (BAFF), (e) inducible nitric oxide synthase (iNOS), (f) retinol dehydrogenase (RDH), (g) transforming growth factor beta (TGF-β), (h) interleukin (IL)-6. (*) upon bar versus respective young mice; (#) upon bar versus respective adult control; asterisks upon line on bars, versus respective sedentary control. *p<0.05, **p<0.01 and ***p<0.001, #p<0.05, ##p<0.01 and ###p<0.001.
2.4. Factors Related to B cells Maduration and Diferentiation
Within age-paired groups, CBFa3 mRNA levels were significantly increased in senile exercised mice versus senile sedentary mice (p<0.001, Figure 3a). In USF-1 mRNA level, no significant differences were seen at 6 or 24 months old (Figure 3b). However, Pax5 mRNA levels was significantly decreased in senile exercise mice versus sedentary senile mice (p<0.001, Figure 3c). Regarding aging, CBFa3, Pax5, and USF mRNA levels no significant differences were seen in senile sedentary mice versus young mice (Figure 3).
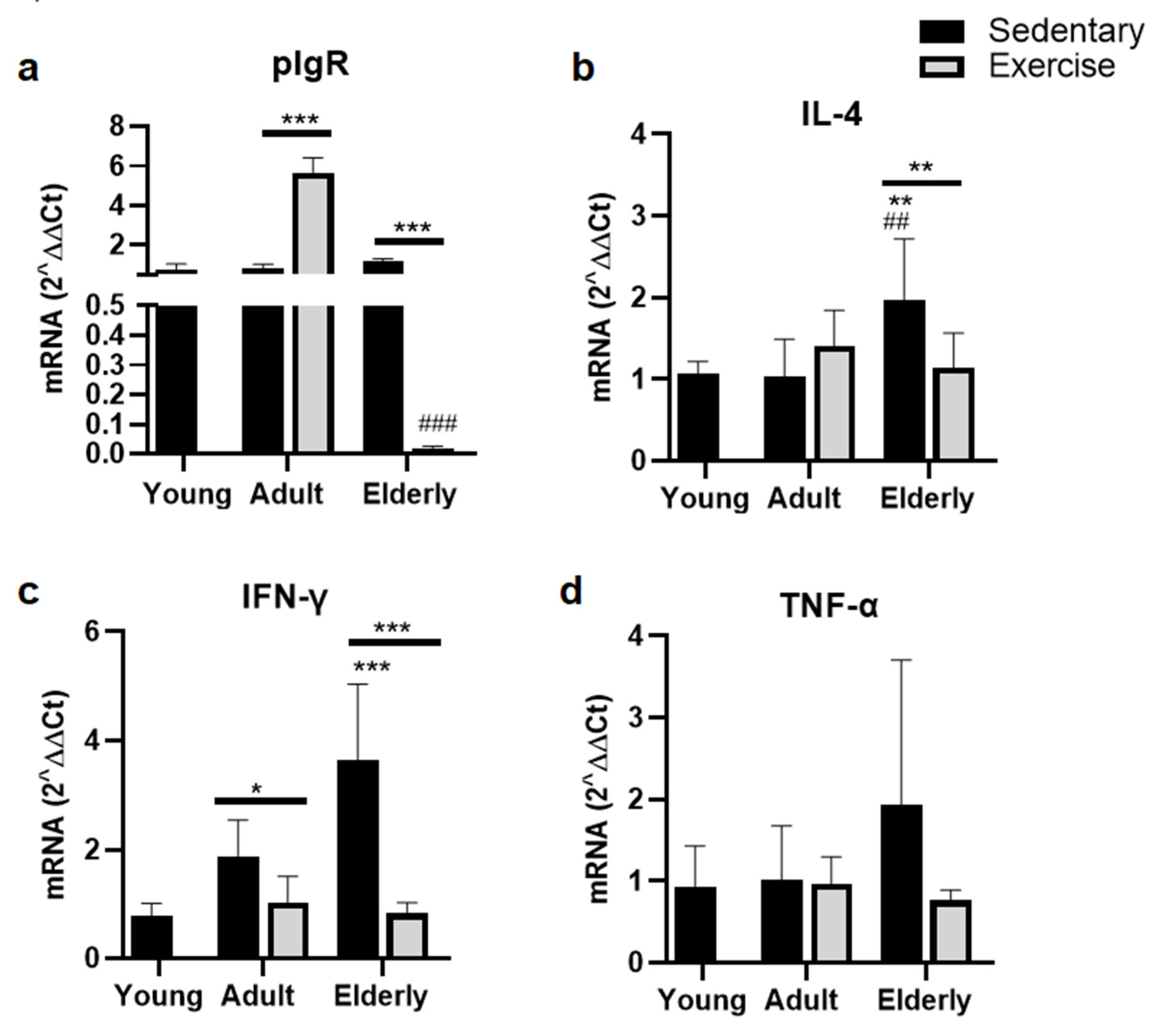
2.5. Exercise Suppressed Parameters of pIgR for IgA-Transcytosis
Within aged-paired groups, pIgR mRNA level was found either greater in the adult exercised versus adult sedentary group or lower in the senile exercised versus senile sedentary group (both p<0.001, Figure 4a). Within exercised groups, pIgR mRNA level was significantly lower in senile versus adult mice p<0.001, Figure 4a).
Comparisons within sedentary groups indicated that IL-4 mRNA level was seen higher in elderly versus young and adult mice (both p<0.01, Figure 4b). Moreover, within the senile mice, IL-4 mRNA was seen lower in exercised than sedentary group (p<0.01, Figure 4b).
Within the sedentary mice, interferon-γ (IFN-γ) mRNA level was found higher in elderly and adult versus young mice (adult p<0.05; senile p<0.001, Figure 3d). Within the senile mice, IFN-γ mRNA was seen lower in exercised than sedentary group (all p<0.001, Figure 4c). Regarding TNFα RNA no significant differences were seen (Figure 4d).
Figure 4.
Gene expression of polymeric immunoglobulin receptor in epithelial cells (a) and cytokines related with its expression in lamina propria (b-c) of small intestine from sedentary or exercise mice at different ages. Data represent the mean and standard deviation of (a) pIgR, (b) interleukin (IL)-4, (c) interferon (IFN)-γ, and (d) tumor necrosis factor (TNF)-α. (*) upon bar versus respective young mice; (#) upon bar versus respective adult control; asterisks upon line on bars versus respective sedentary control. *p<0.05, **p<0.01 and ***p<0.001, ##p<0.01 and ###p<0.001.
Figure 4.
Gene expression of polymeric immunoglobulin receptor in epithelial cells (a) and cytokines related with its expression in lamina propria (b-c) of small intestine from sedentary or exercise mice at different ages. Data represent the mean and standard deviation of (a) pIgR, (b) interleukin (IL)-4, (c) interferon (IFN)-γ, and (d) tumor necrosis factor (TNF)-α. (*) upon bar versus respective young mice; (#) upon bar versus respective adult control; asterisks upon line on bars versus respective sedentary control. *p<0.05, **p<0.01 and ***p<0.001, ##p<0.01 and ###p<0.001.
3. Discussion
It is well-recognized that physical activity has a substantial impact on components of intestinal immunity, microbiota, and IgA production in aging [30,31]. At present the evidence about the T-cell pathways involved in the production of IgA in the small intestine of elderly mice is minimal. In the current assay conducted in the same experimental mice groups, the impact of MAE was analyzed, for the first time, in terms of a T-independent pathway.
Comparative analysis between exercised versus sedentary age-matched groups indicated that MAE upmodulated α-chain mRNA in both senile and adult mice although favored the response of immature cells (B cells and IgA+ B cells) in senile mice and the response of mature cells (including plasma cells, IgM+ and IgA+ plasma cells) in adult mice. It is known that aging downmodulates the B cell populations but paradoxically, increases the antibody production attributable to dysregulation of the mucosal immune system [32]. Thus, data suggested that MAE drives divergent B cell responses in aged and adult mice according to B cell maturity. A recent contribution from our group of study [33], indicated that MAE increased the intestinal IgA antibody concentration via T-cell dependent only in adult mice. In senile mice, MAE induced the increase of homeostatic IgA presumably via microbiota modulation [33]. In the current assay, interestingly, MAE only enhanced APRIL mRNA levels in senile mice whereas elicited TSLP, APRIL, BAFF, and RDH mRNA levels in adult mice. Data suggested that MAE enhanced the APRIL mRNA associated with the T-cell independent pathway of intestinal immune response of IgA. The last findings are very important in aging because it has been described that homeostatic IgA can eliminate some pathogens such as rotavirus and Salmonella typhimurium before the pathogen-specific high-affinity IgA is secreted within the intestinal lumen. As claimed by others, T-cell independent pathway temporarily replaces the T-cell dependent pathway during the early phase of infection [2]. The impact of MAE on the up-modulation of specific IgA antibody response with protective outcome against the infection was reported in the model of Salmonella typhimurium infection in 29-week-old BALB/c mice [34]. Human trials and experimental settings indicate that aging blunts the CSR that drives the B cells committed to antibody production and immunologic memory as described at the systemic compartment [35]. These findings suggested that MAE improved the intestinal response by enhancing the APRIL mRNA level associated with the T-cell independent pathway of B cell response committed for an increase of IgA generation. Findings support the impact of MAE on the increase (twofold) of intestinal IgA concentration in aged mice by comparison with senile sedentary mice [33].
On other hand, among transcript factor related to efficient IgA class switching, is RUNX3, which is important downstream target of TGF-β signaling to activate Sa germline transcription (GLT) [36]. Results of this study indicates that MAE promotes RUNX3 mRNA expression in adulthood and aging, contributing to IgA CSR.
Cell intrinsic mechanisms of survival of mature plasma cells entail the down-regulation of Bcl6 and Pax5 as well as the up-regulation of the transcription factors IRF4 and Blimp-1 [37]. In addition, the loss of Pax5 levels during terminal differentiation contributes to the plasma cell transcription program to IgA CSR [9]. In spite of the plasma cell number was unchanged by the exercise, in this study increased luminal IgA concentration (previously reported [33] ), and α-chain mRNA level found in exercised mice may result from the decrease of Pax5 mRNA favoring both the IgA plasma cell surviving and IgA CSR.
In this study, MAE increased the pIgR mRNA level in adult mice as also documented in 8-weeks-old age (young) BALB/c mice [27]. In the current assay IL-4, and TNFα were unaffected while IFNγ mRNA was decreased by MAE in adult mice. Previous studies documented that IL-4 and TNFα levels were upmodulated by MAE in the duodenum of the small intestine of young BALB/c mice [27]. Apparent discordances may reflect the impact of age on cytokine-mediated pIgR expression. Although pIgR expression is constitutive, it can be regulated by IL-4 and pro-inflammatory cytokines such as TNFα and IFN-γ [38].
Furthermore, MAE decreased the pIgR mRNA in aged mice along with IL-4 and IFN-γ mRNA. pIgR mRNA expression is controversial given is unaffected by aging as documented in senile male Fisher rats [39] but it is affected by diet in six-week-old male C57BL/6 mice [40]. Studies in differentiated intestinal epithelial cell has been showed that basal expression of pIgR is enhanced by the USF proteins [41], which binding enhanced promoter activity of pIgR gene; in this study, changes in USF1 mRNA levels were not seen in exercised mice suggesting that USF1 activity was no related with the decrease of pIgR expression. The impact of aging on pIgR expression has been evidenced in dYY mice (with high monomeric IgA concentration in serum and low monomeric IgA concentration in urine) associated with defective pIgR-mediated IgA transport [22]. In a previous study, we reported that luminal IgA increased in senile exercise mice [33]; in the current study, pIgR mRNA was found decreased what suggested that luminal IgA concentration increases independent of pIgR transcytosis. Pro-inflammatory cytokines including IFN-γ, TNFα, and IL-4 favor pIgR mRNA expres-sion, thus data suggested then that, MAE under senescence conditions down-modulates these proinflammatory cytokines and up-modulates the anti-inflammatory cytokines such as IL-10 and TGF-β. Therefore, MAE may provide a beneficial impact in aging by both ameliorating the cytokines that participate in the inflammatory process and by in-creasing the luminal IgA via independent pIgR-mediated IgA-transcytosis. Thus, MAE contributing to the regulation of the immune system at the mucosal level and ultimately in the intestinal homeostasis in aging.
The impact of aging on pIgR expression has been evidenced in dYY mice (with high monomeric IgA concentration in serum and low monomeric IgA concentration in urine) associated with defective pIgR-mediated IgA transport [22]. In a previous study luminal IgA increase in senile exercised mice was seen (33); in the current study pIgR mRNA was found decreased what suggested that the luminal IgA concentration increases independent of pIgR-mediated IgA-transcytosis Pro-inflammatory cytokines including IFN-γ, TNFα, and IL-4 favor pIgR mRNA expression, thus data suggested then that, MAE under senescence conditions down-modulates these proinflammatory cytokines and up-modulates the anti-inflammatory cytokines such as IL-10 and TGF-β. Therefore, MAE may provide a beneficial impact in aging by both ameliorating the cytokines that participate in the inflammatory process and by increasing the luminal IgA via independent pIgR-mediated IgA-transcytosis. Thus, MAE contributing to the regulation of the immune system at the mucosal level and ultimately in the intestinal homeostasis in aging.
Comparative analysis between sedentary groups, both adult and elderly mice, indicated that aging impacted the decrease of B cell % and increase of IgA+ B cell % versus young mice whereas upmodulated IgM+ B cells as well total, IgM+ and IgA+ plasma cells regarding the adult group. These findings agreed with the impact of aging on up-modulation of lamina propria plasma cells and the decrease of circulant IgA in senile rats [21,22,23,24,25,26,27,28,29,30,31,32]. In other assays, a decrease of surface IgA (sIgA) cells and an increase of sIgM cells were documented in PP from senile mice [42]. Apparent divergences may be derived from the inductive versus effector site of analysis: PP in the previous study [42] and lamina propria in the current. Furthermore, these discordances may reflect the effect of senescence on the reduction of B cell precursors of IgA-secreting plasma cells and Peyer´s patches' function and cellularity [21]. Moreover, age impacts on development and function of ILFs as potential sites for T-independent IgA intestinal response by decreasing the B-cell population and increasing T cells [32]. In this study, decreased B cell population in lamina propria may result from the decreased mRNA and protein expression of CCL20 and CXCL13 chemokines involved in B cell recruitment into ILFs, as documented previously [32]. In this study aging increased the iNOS and IL-4 mRNA versus young and adult mice and increased IFN-γ mRNA only versus young mice while no changes in pIgR mRNA were seen. Data suggested that aging promoted the pro-inflammatory cytokine levels associated with the increase of pIgR (IL-4 and IFN-γ) and presumably these cytokines could have maintained the pIgR expression in the aged mice to the same extent that young and adult mice. Finally, aging increased the iNOS mRNA level in young and adult mice. It is known, iNOS is a factor associated with the T-cell independent pathway.
In this study, assessment at the luminal level of IgA-bacteria complexes was not assessed to address potential relationships with the mucosal parameters with microbiota with the critical role on IgA responses via T-independent pathways. Despite these limitations we can conclude that MAE improved the T-independent IgA-response, favored the luminal IgA production via independent pIgR-transcytosis and suppressed the inflammatory cytokines expression, favoring a homeostatic environment in aging.
4. Materials and Methods
4.1. Animals
Six weeks old male BALB/c mice (N=40) were obtained from “Unidad de Producción y Experimentación de Animales de Laboratorio, Universidad Autónoma Metropolitana Unidad Xochimilco (UPEAL UAMX). Mice were housed in groups of five per cage with light-dark cycles of 12:12 h (7 AM/7 PM), at a relative humidity of 55% at a temperature 20 °C ± 5 °C. Mice were provided with water and fed with Laboratory Rodent Diet 5001 (LabDiet, Saint Louis MO, USA) ad libitum. Manipulation and the exercise protocol were conducted between 8 AM and 11 AM to avoid the influence of circadian cycles of adrenocorticotropic hormone (ACTH) and corticosterone [43]. The procedures carried out on the animals complied with the requirements that determine the standard: NOM-062-ZOO-1999 "Technical specifications for the production, care, and use of laboratory animals" SAGARPA; and the "Guide for the Care and Use of Laboratory Animals". This protocol was approved by the Institutional Committee for the Care and Use of Laboratory Animals of the Higher School of Medicine of the National Polytechnic Institute, Mexico with register key: ESMCICUAL_04/08-11-2020.
4.2. Experimental Design
Animals were adapted to their environment for 2 weeks before starting the exercise protocol on week 9 of life. Mice were divided into 5 groups: 1) Young sedentary group sacrificed at 3 months of age, 2) sedentary adult group sacrificed at 6 months of age, 3) exercised adult group exposed to the MAE protocol up 6 months of age, 4) senile sedentary group sacrificed at 24 months of age, and 5) exercised senile group exposed to the MAE protocol up to 24 months old.
4.3. Moderate Aerobic Exercise Protocol
Before starting the exercise protocol, mice underwent an adaptation period during weeks 9 and 10 of life. The adaptation period consisted of a 10-minute/day session of exercise in the endless band at a speed of 5 m/h and 0° of inclination with increments of 5 min and 5 m/h every other day until reaching 30 min/day at speed of 25-30 m/h. After that, mice ran in an endless band a speed of 30 m/h at 0° of inclination, during 30 min/day, 5 days/week, and rested 2 days (on weekend) until 6 or 24 months old [29].
4.4. Sampling
At 6 or 24 months of age, sedentary and exercise mice were sacrificed by administering a lethal dose of sodium pentobarbital at 150 mg/kg body weight, by intraperitoneal via. After that, the small intestine was dissected and intestinal lavage with PBS was performed. After intestinal lavage, Peyer patches were discarded and epithelial cells and lamina propria were obtained using different Percoll gradients, as described previously by Resendiz-Albor et al., 2010) [44]. The cells obtained were separated and analyzed by flow cytometry or stored at -70 °C for posterior analysis by RT-qPCR.
4.5. Lymphoid Sub-Populations in Lamina Propria by Cytometry
The percentage of the following cell populations was determined: naïve B cells [IgM+/IgD+/ B220+/CD19+], membrane IgA+ B cells [IgA+/B220+/ CD19+], and cytoplasmic IgA plasma cells [IgA+/CD138+] in the lamina propria of the small intestine using the flow cytometry technique. The staining of the lymphocytes was carried out using the next anti-mouse antibodies from Becton Dickinson (BD) Technologies: PECy7-CD19 (cat. 561739, BD Pharmigen), CD45R/B220 (cat. 553089, BD Pharmigen), APC-IgD (Cat. 560868, BD Pharmigen), PerCP-Cy5.5-IgM (cat. 550881, BD Pharmigen), FITC-IgA (cat. 559354, BD Pharmigen) obtained from Pharmingen (San Diego, CA). The protocol provided by the manufacturer was used for surface staining. After staining plasma cells from LP, cells were permeabilized and fixed with cytoperm/cytofix kit (cat. 554722 Becton Dickinson) for cytoplasmic IgA determination. Samples were acquired and analyzed in a BD FACSAria Fusion flow cytometer using FACSDiva software. Ten thousand events in the lymphocyte region in the SSC vs. FSC dot-plot for B cells and 20,000 events for plasma cells were analyzed.
4.6. Relative Expression of mRNA Assay
In the cell suspensions of lamina propria were determined the relative mRNA levels of genes related with IgA-synthesis (α-chain, TSLP, APRIL, BAFF, iNOS, RDH, TGF-β1, and IL-6 mRNA) and B cell differentiation and maturation (CBFa3/RUNX-3, USF-1, and Pax5). In suspensions of epithelial cells were assessed mRNA levels of IL-4, TNFα, IFN-γ and pIgR by RT-qPCR.
Total RNA extraction was done using the protocol described for the fabricant of TRIzol® reagent (cat. 15596018, Invitrogen). The concentration and purity of total RNA were determined by spectrophotometry in the Nanodrop™ 2000/2000c (Cat: ND-2000, USA, Thermo Scientific Wilmington). The ratio of absorbances A260/280 with a value between 2.0-2.2 and > 1.7 was considered an optimal purity. The RNA integrity was analyzed by electrophoresis in the gel of agarose at 1%. Reverse transcription was performed using the Maxima First cDNA Synthesis Kit for RT–qPCR (cat. No. K1642, Thermo Scientific, Lithuania) at 37 °C for 50 min.
Quantitative PCR was subsequently performed by duplicate, using a Maxima Probe/ROX qPCR Master mix Kit (cat. K0261, Thermo Scientific™, Lithuania) in a LightCycler ® Nano Instrument (Roche Diagnostics GmbH, Rotkreuz Switzerland) at an initial denaturing step (at 95 °C for 10 min), followed by 45 cycles of amplification (at 95 °C for 10 s, 60 °C for 35 s, and 72 °C for 1 s) and one cycle of cooling (40 °C for 30 s).
Specific oligonucleotide primers were originally generated by using the online assay design software (ProbeFinder: http://www.universal-probelibrary. com) and the primer sequences for TSLP, APRIL, BAFF, iNOS, RDH, TGF-β1, IL-6, CBFa3/RUNX-3, USF-1, Pax5 and GAPDH are shown in Table 1. A primer sequence of α-chain, pIgR, IFN-γ, IL-4, and TFN-α was previously published by Godinez-Victoria, et al. 2012 [28]. Messenger RNA expression levels were calculated using the comparative parameter quantification cycle method and normalized to the relative expression of GAPDH mRNA [45].
4.7. Statistical Analysis
At the end of the study (at 24 months old), the survival was ~60% in the control group and ~80% in exercised group. For that, data are presented as the mean and standard deviation (SD) of 6–8 animals per group. For the comparison between groups in the different measurement times, two-way ANOVA was used considering as factors the group (sedentary or exercised) and the age of the mice (young, adult, and elderly mice), followed by a Turkey method post hoc test. For all tests, p< 0.05 was considered significant. All data were analyzed using SigmaPlot for Windows version 11.1 (Systat Software Inc. San Jose CA, USA). P<0.05 was considered to indicate a statistically significant difference.
5. Conclusions
This study evidenced that MAE has differential outcomes on factors associated with IgA generation according to the age of mice. MAE enhanced the T-cell-independent pathway in both adult and senile mice. Under senescence conditions, MAE promoted the B cell and IgA+ B cells and APRIL mRNA that may improve the intestinal response. Under senescence conditions, MAE ameliorated the inflammatory environment associated presumably with the downmodulation of pIgR mRNA. The data support the presumably MAE-enhanced IgA [33] via pIgR-independent transcytosis.
Authors contribution
Conceptualization, Maria-Elisa Drago-Serrano and Marycarmen Godinez-Victoria; Data curation, Angel Hernández-Urbán; Formal analysis, Angel Hernández-Urbán, Maria-Elisa Drago-Serrano, Aldo Reséndiz-Albor and Marycarmen Godinez-Victoria; Funding acquisition, Marycarmen Godinez-Victoria; Investigation, José Sierra-Ramírez and Marycarmen Godinez-Victoria; Methodology, Angel Hernández-Urbán, Aldo Reséndiz-Albor, José Sierra-Ramírez, Fabiola Guzmán-Mejia and Rigoberto Oros-Pantoja; Project administration, Marycarmen Godinez-Victoria; Software, Rigoberto Oros-Pantoja; Validation, Maria-Elisa Drago-Serrano, Fabiola Guzmán-Mejia and Marycarmen Godinez-Victoria; Visualization, Maria-Elisa Drago-Serrano, José Sierra-Ramírez, Fabiola Guzmán-Mejia and Rigoberto Oros-Pantoja; Writing – original draft, Maria-Elisa Drago-Serrano; Writing – review & editing, Fabiola Guzmán-Mejia, Rigoberto Oros-Pantoja and Marycarmen Godinez-Victoria.
Funding Statement
Authors should state how the research and publication of their article was funded, by naming financially supporting bodies followed by any associated grant numbers in square brackets. Consejo Nacional de Humanidades, Ciencia y Tecnología (CONAHCyT), México [254492] and Secretaría de Investigación y Posgrado del Instituto Politécnico Nacional [20195200] to Dr. Marycarmen Godínez-Victoria as leader.
Data Availability
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Acknowledgments
Authors honor the memory of working group leader Ph.D. Rafael Campos Rodriguez, who died on July 4th, 2019, and his contributions on the immunomodulatory effects of exercise.
Conflicts of Interest
The authors declare that there is no conflict of interest regarding the publication of this paper.
References
- J. M. Woof & M. A. Kerr, The function of immunoglobulin A in immunity. The Journal of Pathology, 208 (2006) 270–282. [CrossRef]
- H. Tezuka & T. Ohteki, Regulation of IgA Production by Intestinal Dendritic Cells and Related Cells. Frontiers in Immunology, 10 (2019) 1891. [CrossRef]
- J. J. Bunker, S. A. Erickson, T. M. Flynn, C. Henry, J. C. Koval, M. Meisel, B. Jabri, D. A. Antonopoulos, P. C. Wilson, & A. Bendelac, Natural polyreactive IgA antibodies coat the intestinal microbiota. Science (New York, N.Y.), 358 (2017) eaan6619. [CrossRef]
- E. D. León & M. P. Francino, Roles of Secretory Immunoglobulin A in Host-Microbiota Interactions in the Gut Ecosystem. Frontiers in Microbiology, 13 (2022) 880484. [CrossRef]
- C. Lindner, B. Wahl, L. Föhse, S. Suerbaum, A. J. Macpherson, I. Prinz, & O. Pabst, Age, microbiota, and T cells shape diverse individual IgA repertoires in the intestine. The Journal of Experimental Medicine, 209 (2012) 365–377. [CrossRef]
- F. Barone, P. Patel, J. D. Sanderson, & J. Spencer, Gut-associated lymphoid tissue contains the molecular machinery to support T-cell-dependent and T-cell-independent class switch recombination. Mucosal Immunology, 2 (2009) 495–503. [CrossRef]
- S. Uematsu, K. Fujimoto, M. H. Jang, B.-G. Yang, Y.-J. Jung, M. Nishiyama, S. Sato, T. Tsujimura, M. Yamamoto, Y. Yokota, H. Kiyono, M. Miyasaka, K. J. Ishii, & S. Akira, Regulation of humoral and cellular gut immunity by lamina propria dendritic cells expressing Toll-like receptor 5. Nature Immunology, 9 (2008) 769–776. [CrossRef]
- Y.-J. Liu, TSLP in epithelial cell and dendritic cell cross talk. Advances in Immunology, 101 (2009) 1–25. [CrossRef]
- A. Delogu, A. Schebesta, Q. Sun, K. Aschenbrenner, T. Perlot, & M. Busslinger, Gene repression by Pax5 in B cells is essential for blood cell homeostasis and is reversed in plasma cells. Immunity, 24 (2006) 269–281. [CrossRef]
- Pabst & E. Slack, IgA and the intestinal microbiota: the importance of being specific. Mucosal Immunology, 13 (2020) 12–21. [CrossRef]
- S. Yuan, J. Yang, Y. Jian, Y. Lei, S. Yao, Z. Hu, X. Liu, C. Tang, & W. Liu, Treadmill Exercise Modulates Intestinal Microbes and Suppresses LPS Displacement to Alleviate Neuroinflammation in the Brains of APP/PS1 Mice. Nutrients, 14 (2022) 4134. [CrossRef]
- H. Sugahara, S. Okai, T. Odamaki, C. B. Wong, K. Kato, E. Mitsuyama, J.-Z. Xiao, & R. Shinkura, Decreased Taxon-Specific IgA Response in Relation to the Changes of Gut Microbiota Composition in the Elderly. Frontiers in Microbiology, 8 (2017) 1757. [CrossRef]
- B. van der Lugt, A. A. van Beek, S. Aalvink, B. Meijer, B. Sovran, W. P. Vermeij, R. M. C. Brandt, W. M. de Vos, H. F. J. Savelkoul, W. T. Steegenga, & C. Belzer, Akkermansia muciniphila ameliorates the age-related decline in colonic mucus thickness and attenuates immune activation in accelerated aging Ercc1 -/Δ7 mice. Immunity & Ageing: I & A, 16 (2019) 6. [CrossRef]
- W. Yang, Y. Liu, G. Yang, B. Meng, Z. Yi, G. Yang, M. Chen, P. Hou, H. Wang, & X. Xu, Moderate-Intensity Physical Exercise Affects the Exercise Performance and Gut Microbiota of Mice. Frontiers in Cellular and Infection Microbiology, 11 (2021) 712381. [CrossRef]
- A. Aiello, F. Farzaneh, G. Candore, C. Caruso, S. Davinelli, C. M. Gambino, M. E. Ligotti, N. Zareian, & G. Accardi, Immunosenescence and Its Hallmarks: How to Oppose Aging Strategically? A Review of Potential Options for Therapeutic Intervention. Frontiers in Immunology, 10 (2019) 2247. [CrossRef]
- M. Sellami, N. L. Bragazzi, B. Aboghaba, & M. A. Elrayess, The Impact of Acute and Chronic Exercise on Immunoglobulins and Cytokines in Elderly: Insights From a Critical Review of the Literature. Frontiers in Immunology, 12 (2021) 631873. [CrossRef]
- A. Jafarzadeh, M. Sadeghi, G. A. Karam, & R. Vazirinejad, Salivary IgA and IgE levels in healthy subjects: relation to age and gender. Brazilian Oral Research, 24 (2010) 21–27. [CrossRef]
- D. A. Sullivan, J. P. Vaerman, & C. Soo, Influence of severe protein malnutrition on rat lacrimal, salivary and gastrointestinal immune expression during development, adulthood and ageing. Immunology, 78 (1993) 308–317.
- S. Senda, E. Cheng, & H. Kawanishi, Aging-associated changes in murine intestinal immunoglobulin A and M secretions. Scandinavian Journal of Immunology, 27 (1988) 157–164. [CrossRef]
- D. L. Schmucker, C. K. Daniels, R. K. Wang, & K. Smith, Mucosal immune response to cholera toxin in ageing rats. I. Antibody and antibody-containing cell response. Immunology, 64 (1988) 691–695.
- K. Thoreux, R. L. Owen, & D. L. Schmucker, Intestinal lymphocyte number, migration and antibody secretion in young and old rats. Immunology, 101 (2000) 161–167. [CrossRef]
- T. Yanagihara, Y. Kumagai, Y. Norose, I. Moro, M. Nanno, M. Murakami, & H. Takahashi, Age-dependent decrease of polymeric Ig receptor expression and IgA elevation in ddY mice: a possible cause of IgA nephropathy. Laboratory Investigation; a Journal of Technical Methods and Pathology, 84 (2004) 63–70. [CrossRef]
- D. Wilson, T. Jackson, E. Sapey, & J. M. Lord, Frailty and sarcopenia: The potential role of an aged immune system. Ageing Research Reviews, 36 (2017) 1–10. [CrossRef]
- A. L. de Araújo, L. C. R. Silva, J. R. Fernandes, & G. Benard, Preventing or reversing immunosenescence: can exercise be an immunotherapy? Immunotherapy, 5 (2013) 879–893. [CrossRef]
- R. J. Simpson, T. W. Lowder, G. Spielmann, A. B. Bigley, E. C. LaVoy, & H. Kunz, Exercise and the aging immune system. Ageing Research Reviews, 11 (2012) 404–420. [CrossRef]
- M.-E. Drago-Serrano, M. Godínez-Victoria, E. Lara-Padilla, A. A. Resendiz-Albor, H. Reyna-Garfias, I. M. Arciniega-Martínez, A. Kormanovski-Kovsova, & R. Campos-Rodriguez, Moderate exercise enhances expression of SIgA in mouse ileum. International Journal of Sports Medicine, 33 (2012) 1020–1025. [CrossRef]
- M. Viloria, E. Lara-Padilla, R. Campos-Rodríguez, A. Jarillo-Luna, H. Reyna-Garfias, P. López-Sánchez, V. Rivera-Aguilar, A. Salas-Casas, F. J. Berral de la Rosa, & E. García-Latorre, Effect of moderate exercise on IgA levels and lymphocyte count in mouse intestine. Immunological Investigations, 40 (2011) 640–656. [CrossRef]
- M. Godínez-Victoria, M. E. Drago-Serrano, H. Reyna-Garfias, M. Viloria, E. Lara-Padilla, A. A. Resendiz-Albor, L. E. Sánchez-Torres, T. R. Cruz-Hernández, & R. Campos-Rodriguez, Effects on secretory IgA levels in small intestine of mice that underwent moderate exercise training followed by a bout of strenuous swimming exercise. Brain, Behavior, and Immunity, 26 (2012) 1300–1309. [CrossRef]
- J. A. Sierra-Ramírez, L. Saucedo-Bueno, A. L. García-Hernández, A. Martínez-Dávalos, C. Rodríguez-López, M. E. Drago-Serrano, & M. Godínez-Victoria, Moderate aerobic exercise on bone quality changes associated with aging and oxidative stress in BALB/c mice. Journal of Biomechanics, 135 (2022) 111035. [CrossRef]
- M. D. Cook, J. M. Allen, B. D. Pence, M. A. Wallig, H. R. Gaskins, B. A. White, & J. A. Woods, Exercise and gut immune function: evidence of alterations in colon immune cell homeostasis and microbiome characteristics with exercise training. Immunology and Cell Biology, 94 (2016) 158–163. [CrossRef]
- R. Divella, G. DE Palma, A. Tufaro, G. Pelagio, G. Gadaleta-Caldarola, R. Bringiotti, & A. Paradiso, Diet, Probiotics and Physical Activity: The Right Allies for a Healthy Microbiota. Anticancer Research, 41 (2021) 2759–2772. [CrossRef]
- K. G. McDonald, M. R. Leach, C. Huang, C. Wang, & R. D. Newberry, Aging impacts isolated lymphoid follicle development and function. Immunity & Ageing: I & A, 8 (2011) 1. [CrossRef]
- A. J. Hernández-Urbán, M. E. Drago-Serrano, A. Cruz-Baquero, A. L. García-Hernández, I. M. Arciniega-Martínez, J. Pacheco-Yépez, F. Guzmán-Mejía, & M. Godínez-Victoria, Exercise improves intestinal IgA production by T-dependent cell pathway in adults but not in aged mice. Frontiers in Endocrinology, 14 (2023) 1190547. [CrossRef]
- M. Godínez-Victoria, R. Campos-Rodriguez, V. Rivera-Aguilar, E. Lara-Padilla, J. Pacheco-Yepez, R. A. Jarillo-Luna, & M. E. Drago-Serrano, Intermittent fasting promotes bacterial clearance and intestinal IgA production in Salmonella typhimurium-infected mice. Scandinavian Journal of Immunology, 79 (2014) 315–324. [CrossRef]
- D. Frasca & B. B. Blomberg, Inflammaging decreases adaptive and innate immune responses in mice and humans. Biogerontology, 17 (2016) 7–19. [CrossRef]
- B. Zhao, Z. Xia, B. Yang, Y. Guo, R. Zhou, M. Gu, M. Liu, Q. Li, W. Bai, J. Huang, X. Zhang, C. Zhu, K. T. Leung, C. Chen, & J. Dong, USP7 promotes IgA class switching through stabilizing RUNX3 for germline transcription activation. Cell Reports, 43 (2024) 114194. [CrossRef]
- J. L. Gommerman, O. L. Rojas, & J. H. Fritz, Re-thinking the functions of IgA(+) plasma cells. Gut Microbes, 5 (2014) 652–662. [CrossRef]
- F.-E. Johansen & P. Brandtzaeg, Transcriptional regulation of the mucosal IgA system. Trends in Immunology, 25 (2004) 150–157. [CrossRef]
- C. K. Daniels, D. L. Schmucker, H. Bazin, & A. L. Jones, Immunoglobulin A receptor of rat small intestinal enterocytes is unaffected by aging. Gastroenterology, 94 (1988) 1432–1440. [CrossRef]
- T. Shibuya, T. Kaburagi, R. Nagai, & S. Oshiro, The effects of moderate exercise on secretory IgA production in mice depends on dietary carbohydrate intake. Journal of Clinical Biochemistry and Nutrition, 57 (2015) 44–49. [CrossRef]
- M. E. C. Bruno, R. B. West, T. A. Schneeman, E. H. Bresnick, & C. S. Kaetzel, Upstream stimulatory factor but not c-Myc enhances transcription of the human polymeric immunoglobulin receptor gene. Molecular Immunology, 40 (2004) 695–708. [CrossRef]
- H. Kawanishi & J. Kiely, Immune-related alterations in aged gut-associated lymphoid tissues in mice. Digestive Diseases and Sciences, 34 (1989) 175–184. [CrossRef]
- A. E. Gaffey, C. S. Bergeman, L. A. Clark, & M. M. Wirth, Aging and the HPA axis: Stress and resilience in older adults. Neuroscience and Biobehavioral Reviews, 68 (2016) 928–945. [CrossRef]
- A. Reséndiz-Albor, H. Reina-Garfias, S. Rojas-Hernández, A. Jarillo-Luna, V. Rivera-Aguilar, A. Miliar-García, & R. Campos-Rodríguez, Regionalization of pIgR expression in the mucosa of mouse small intestine. Immunology Letters, 128 (2010) 59–67. [CrossRef]
- K. J. Livak & T. D. Schmittgen, Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods (San Diego, Calif.), 25 (2001) 402–408. [CrossRef]
Figure 3.
Gene expression of transcription factors involved in B-cell development and differentiation impacting on IgA production. Data represent the mean and standard deviation of (a) Core-Binding Factor Alpha 3 (CBFa3/RUNX-3), (b) Upstream Stimulatory Factor-1 (USF), and (c) paired box5 (Pax5). (***) upon bar versus young mice; or asterisks upon line on bars versus respective sedentary control. *p<0.05 and ***p<0.001.
Figure 3.
Gene expression of transcription factors involved in B-cell development and differentiation impacting on IgA production. Data represent the mean and standard deviation of (a) Core-Binding Factor Alpha 3 (CBFa3/RUNX-3), (b) Upstream Stimulatory Factor-1 (USF), and (c) paired box5 (Pax5). (***) upon bar versus young mice; or asterisks upon line on bars versus respective sedentary control. *p<0.05 and ***p<0.001.
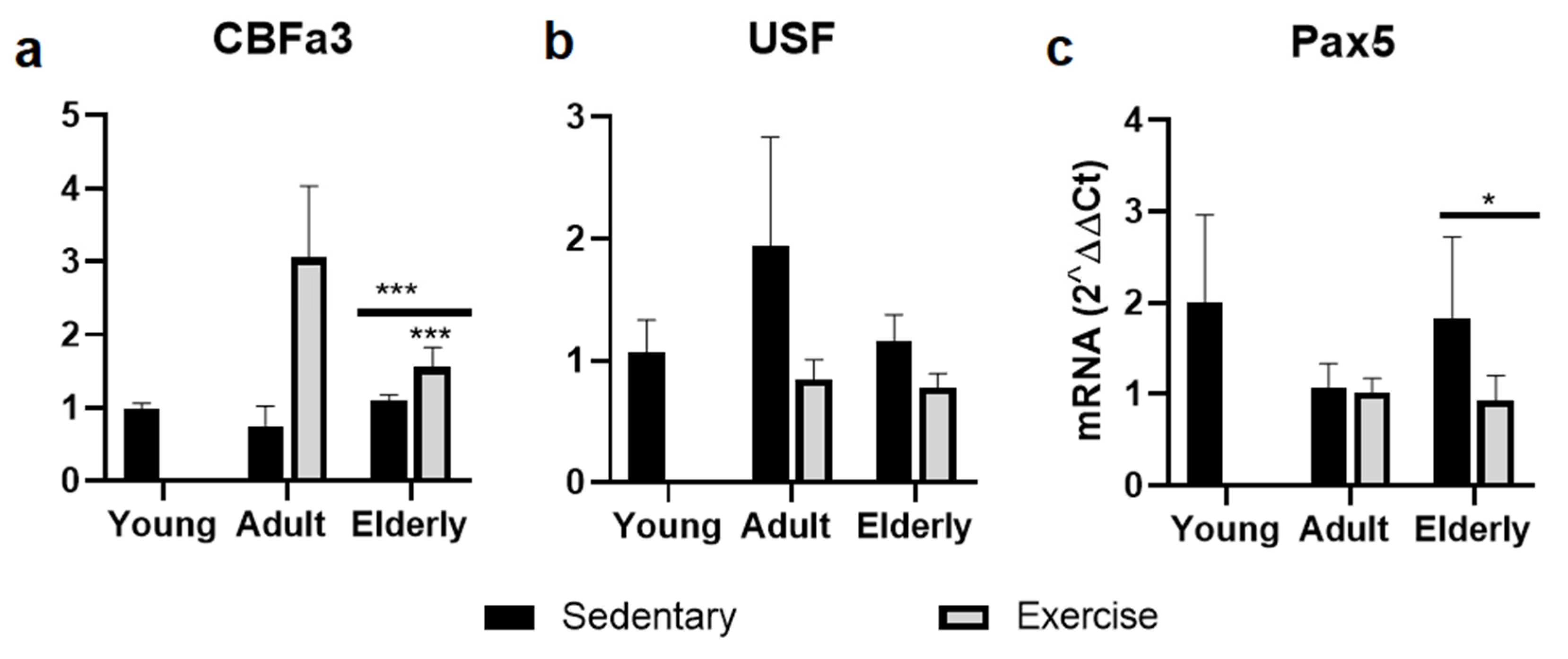

Table 1.
Forward and reverse primers for real-time PCR assays designed according to the ensemble transcript ID of the Universal ProbeLibrary.
Table 1.
Forward and reverse primers for real-time PCR assays designed according to the ensemble transcript ID of the Universal ProbeLibrary.
| Gen | Sequence 5´→3´ | ID* | |
|---|---|---|---|
| Forward | Reverse | ||
| TSLP | cag ctt gtc tcc tga aaa tcg | aaa tgt ttt gtc ggg gag tg | NM_021367.2 |
| BAFF | atg cgg aag gca gat tga | tgc atc ttt tgc tac cct ga | NM_001347309.1 |
| APRIL | agc tgg gca ctg agc ttt ac | aag ttg gcc tcg aac tca tc | NM_023517.2 |
| iNOS | ctt ttc cta tgg ggc aaa aa | ctg gaa ctc tgg gct gtc a | NM_010927.4 |
| RDH11 | tgt act tgg tca cgc caa aa | ccg gga agc tga aca tta ga | NM_021557.5 |
| TGF-β1 | tgg agc aac atg tgg aac tc | gtc agc agc cgg tta cca | NM_011577.2 |
| IL-6 | gct acc aaa ctg gat ata atc agg a | cca ggt agc tat ggt act cca gaa | NM_031168.2 |
| CBFa3 | gct ctc tca gca cca cga g | tca ggt ctg agg agc ctt g | NM_019732.2 |
| USF-1 | tca aga ggt ggg aaa gga tg | cat tgg gcc ccc ttc tac | NM_001305676.1 |
| Pax5 | cct ggg agt gaa ttt tct gg | tgg gga acc tcc aag aat c | NM_008782.2 |
| GAPDH | aag agg gat gct gcc ctt ac | cca ttt tgt cta cgg gac ga | NM_001289726.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



