
Submitted:
14 June 2024
Posted:
18 June 2024
You are already at the latest version
Abstract
Pseudomonas aeruginosa is an opportunistic Gram-negative bacterium that remains a prevalent clinical and environmental challenge. Quorum sensing (QS) molecules are effective biomarkers in pinpointing the presence of P. aeruginosa. This study aimed to develop a convenient-to-use whole-cell biosensor using P. aeruginosa reporters individually encapsulated within alginate-poly-L-lysine (alginate-PLL) microbeads to specifically detect the presence of bacterial autoinducers. The PLL-reinforced microbeads were prepared using a two-step method involving ionic crosslinking and subsequent coating with thin layers of PLL. The alginate-PLL beads showed good stability in the presence of a known cation scavenger (sodium citrate), which typically limits the widespread applications of calcium alginate. In media containing synthetic autoinducers, such as N-(3-oxo dodecanoyl) homoserine lactone (3-oxo-C12-HSL) and N-butanoyl-L-homoserine lactone (C4-HSL) or cell-free supernatants of planktonic or flow cell biofilm effluent of wild P. aeruginosa (PAO1), the encapsulated bacteria enabled a dose-dependent detection of the presence of these QS molecules. The prepared bioreporter beads remained stable during prolonged storage at 4 and -80 °C and were ready for on-the-spot sensing without the need for recovery. This whole-cell biosensor demonstrates the practicality of the encapsulated bioreporter for bacterial detection based on specific QS molecules.
Keywords:
Alginate
; Hydrogels
; quorum sensing
; C4-HSL
; whole-cell biosensors
; Furanone C-30
; autoinducers
; Pseudomonas aeruginosa
; bioencapsulation
1. Introduction
Pseudomonas aeruginosa is an increasingly prevalent opportunistic Gram-negative bacterium causing nosocomial and life-threatening infections of immunocompromised patients [1]. Patients with cystic fibrosis are especially disposed to P. aeruginosa infections, and for these persons, the bacterium is responsible for high rates of morbidity and mortality [2]. P. aeruginosa can colonize implanted devices, catheters, heart valves, or dental implants [3]. Many Gram-negative bacteria regulate the expression of specialized gene sets in response to their population density by autoinduction, in which small molecules called autoinducers (AIs) are produced, and can diffuse freely across the bacterial cell wall. By reacting to the concentration of AIs, individual cells can sense how many neighboring cells surround them and are, in this way, able to determine whether the population density is high enough to initiate the expression of a particular phenotype. This type of gene regulation is termed quorum sensing (QS).
The study of QS has emerged as a key research area in the past two decades due to its involvement in various health and environmental issues. Particular attention is drawn to understanding QS-mediated bacterial communication, as QS has been implicated in regulating a myriad of microbial activities in a population-density-dependent manner. For instance, several bacterial behaviors such as swarming and motility, sporulation, biofilm formation, bioluminescence, and production of virulence factors [4] are regulated by QS. Moreover, there is an increasing body of knowledge supporting the involvement of QS in the pathogenesis of bacteria-related disorders, whereby about 80% of microbial infections have been associated with biofilm formation [5]; and the contribution of QS molecules in biofouling [6], a major environmental concern, is well established. Molecules such as N-acyl homoserine lactones (AHLs) are used by P. aeruginosa as autoinducers for quorum sensing. These molecules consist of a homoserine lactone and a fatty acyl group. The length of the fatty acyl group varies from 4 to 14 carbons, and the group can be either fully reduced, have a 3-oxo or 3-hydroxyl group, or have an unsaturated bond. AHLs have different roles in bacterial infections. P. aeruginosa employs acyl homoserine lactones AHLs as signal molecules [7], and N-butyryl-homoserine lactone (C4-HSL) [8] and N-3-oxo-dodecanoyl-homoserine lactone (3OC12-HSL) [9] have been detected in clinical samples of cystic fibrosis patients [8,10,11,12]. Their presence is positively correlated with the infection rate [13].
The determination and quantification of AHLs in biological and environmental samples have been a subject of significant scientific interest as diagnostic biomarkers for the presence of Gram-negative bacteria. The need for analytical techniques that are sensitive and selective in detecting and quantifying each AHLs is still imperative. Over the years, numerous analytical methods have been developed to precisely detect AHLs in bacterial culture supernatants. These methods are bacterial whole-cell sensing systems [14] or conventional physical-chemical techniques such as colorimetry, liquid and gas chromatography with UV, mass spectrometry, and diode array detection.[15,16]. Highly sensitive detection of AHLs can be achieved by biosensors [17], LC-MS/MS, [18] and GC-MS/MS [18,19]. Although physical-chemical techniques prove to be sensitive, their widespread use is hampered by the need for solvent extraction, derivatization of the analytes, and the cost of the instrument, as well as the requirement for trained personnel. Consequently, biosensors have been shown to be viable alternatives due to their affordable cost, fast analysis time, and high sensitivity, making them ideal for routine testing and screening of samples [19].
With the recent advances in synthetic biology, engineered whole-cell biosensors have emerged as a new platform for various environmental and biomedical applications, and they hold promise to provide novel diagnostic devices for easily deployed point-of-care testing [20]. Whole-cell sensors are typically constructed using genetically modified cells designed to express a regulatory protein that can identify a specific compound. In response, these cells produce a reporter molecule, the concentration of which can be measured using signals such as fluorescence, chemo- or bioluminescence, electrochemistry, or colorimetry. [14,21] The recognition elements, as well as the signal-generating molecules, are proteins whose genes are encoded by a plasmid harbored within the cell [17]. Various reporters can be employed to monitor the induction of an operon, depending on the desired detection system. These constructs have a sensitivity that matches the physiological concentrations of AHLs in the context of QS. The mutations that are introduced into the bioreporter bacteria strain to detect quorum-sensing molecules render them incapable of producing AHLs themselves but maintaining their capability of sensing their presence. Such mutations are usually introduced upstream of a LuxR-controlled promoter, fused to a gene coding for an easily detectable output signal. For details on the construction of bacterial lux-biosensors, readers are directed to the review work of Bazhenov et al. [22]. Various reporter strains have been constructed, with different specificity and sensitivity for QS signal molecules produced by other bacterial species, exploiting diverse quorum sensing systems, such as the lux [10], las [23], rhl [17], ahl [24], and agr [25] QS systems.
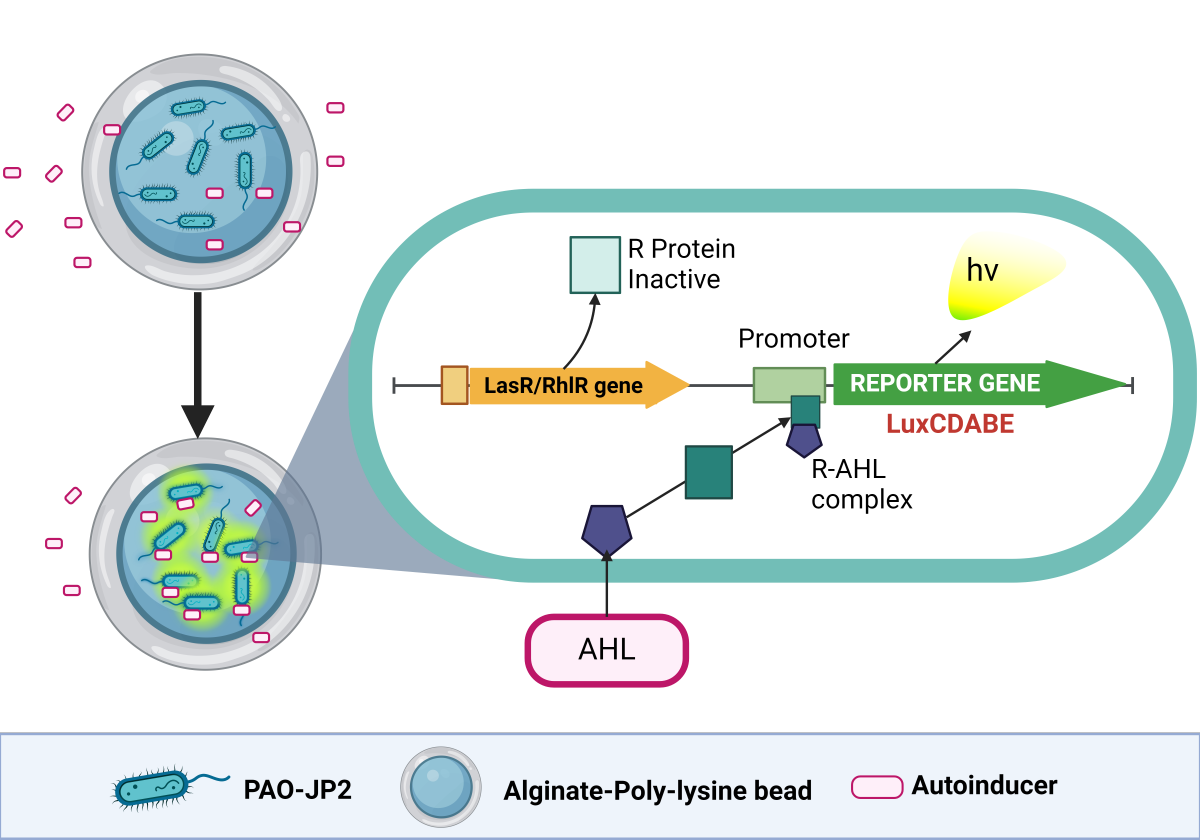
In this work, the bioluminescent reporter gene (luxCDABE) is fused downstream of the luxR-promoter. Among cell-based biosensors, the luxCDABE reporter is reputed for the lowest detection limits (down to 10-18 mol) it offers [26]. The luxA and luxB genes within the luxCDABE gene cassette encode for bacterial luciferase, while the luxC, luxD, and luxE genes in the same cassette encode for the enzymes required for the synthesis and recycling of the luciferase substrate. This setup eliminates the need for any external addition of the substrate [22], enabling continuous real-time monitoring with low background interferences [22]. Transcription is initiated by binding the AHL-luxR complex to the promoter, leading to concomitant luminescent production (Figure 1). The amount of luminescence generated can be correlated with the concentration of the standard AHLs from which the calibration curve is generated. This can be used to quantitatively determine the presence of the quorum-sensing molecules in test/unknown samples. Our research advanced the development of whole-cell sensing systems by immobilizing them through encapsulation in reinforced alginate beads. These beads can be utilized as reporter microbeads for monitoring AHLs in biological and environmental samples, including culture supernatants and biofilm flowthroughs. Whole-cell biosensing systems possess attributes that make them ideal for portable field kits. These attributes include the capacity to withstand a wide range of environmental conditions (such as various temperatures, pH levels, and ionic strengths), providing information on analyte bioavailability, require minimal or no sample pretreatment, are highly sensitive, and selective, are easily prepared, provide rapid detection, are cost-effective, are amenable to high-throughput screening, and miniaturization [27].
Hydrogels are water-retaining and water-insoluble three-dimensional crosslinked natural or synthetic polymeric networks. Hydrogel capsules have been found to have extensive applications in the bioencapsulation of living cells to retain viability and functionality. Sodium alginates are naturally occurring polyanionic polysaccharides comprising a linear copolymer of 1 → 4 linked β-D-mannuronic acid (M units) and α-L-guluronic acid (G units). Sodium alginate possesses a unique ability to chelate with multivalent cations, such as the carboxylic acid groups of the G-G blocks. They can chelate with divalent ions like Ca2+ and Ba2+ in solution and form insoluble calcium alginate with an “egg-box” molecular structure [28]. Therefore, ionic crosslinked alginate hydrogels with various structures can be prepared easily. Owing to this rapid and nontoxic encapsulation approach, alginate has emerged as a choice biomaterial for various encapsulation applications [29,30]. Calcium ionic crosslinked alginate hydrogel stability is challenged in physiological solutions and the presence of cation scavengers such as citrate, ethylene diamine tetra acetic acid, and phosphates. The calcium ions can, therefore, be exchanged by other ions, which means a loss of cross-links from the alginate gels. This makes the gels structurally and mechanically weaker, leading to uncontrollable permeability [31] and eventual loss of stable 3D structure [28]. This characteristic has been limiting the biotechnological applications of calcium alginate hydrogels. Therefore, to address the above problem, efforts have been made to increase the chemical and physical durability of alginate gels by coating them with cationic and anionic polymers such as chitosan [32] and by exploiting several covalent crosslinking strategies and a composite-based approach [28,33]. To that effect, in this study, poly-lysine-reinforced calcium alginate was used to encapsulate the bioreporter in a two-step approach. Although Strand et al. (date) employed Confocal Laser Scanning Microscopy (CLSM) to characterize microcapsules of alginate-poly-L-lysine in terms of layer formation, stability, and distribution [30], the biotechnological applications of these capsules have been sparingly employed in drug release experiments [31] and cell encapsulation studies [34].
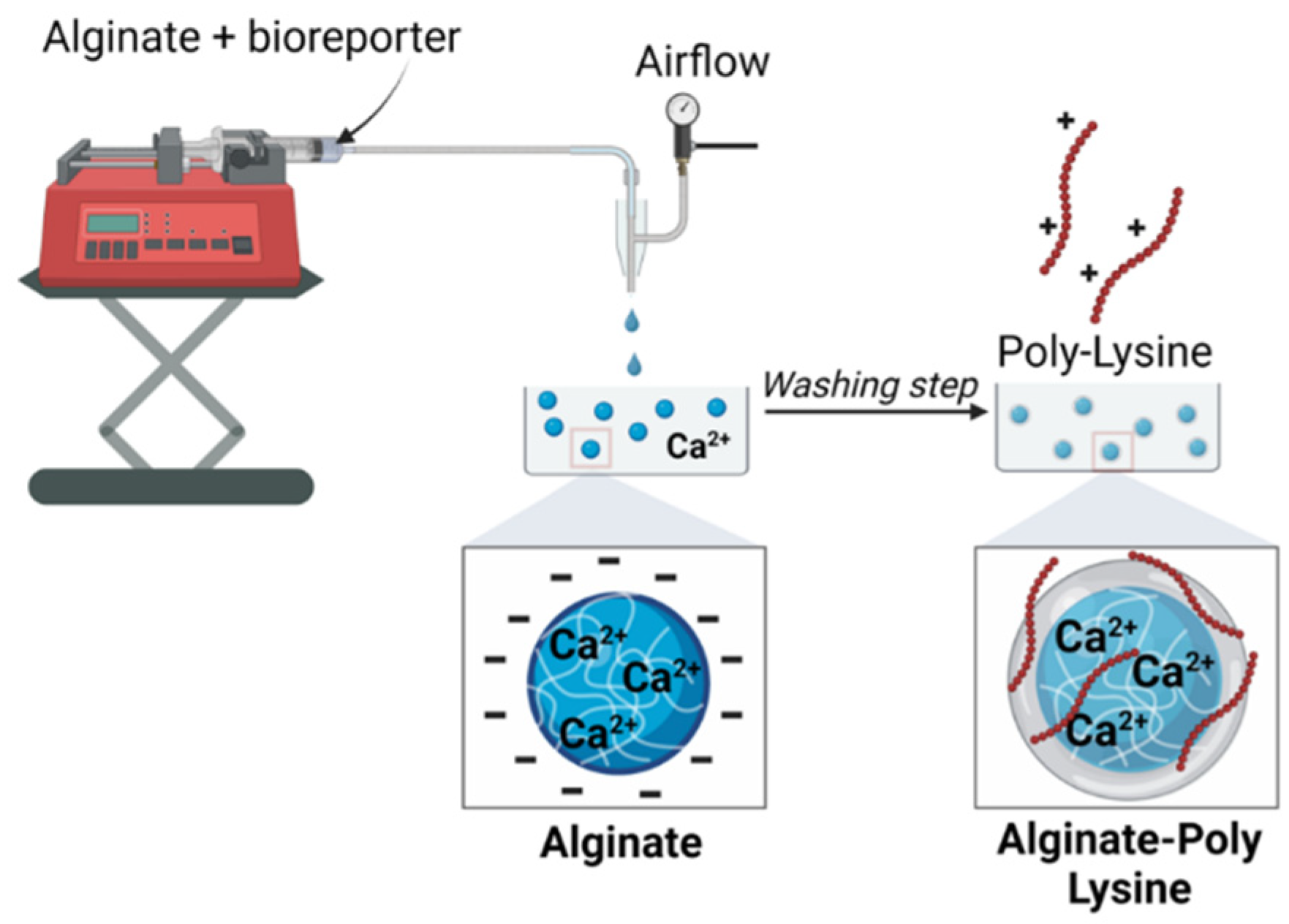
This work aimed to develop an alginate-based bioreporter encapsulation for detecting quorum-sensing molecules with a rationally designed coating developed through the poly-lysine assembly. In brief, our approach involves sending a feed solution of alginate and engineered P. aeruginosa (PAO-JP2) strains through a microcapillary (connected to an airstream) into a reservoir containing Ca2+ to form calcium alginate microbeads (Figure 3). The resulting PAO-JP2-alginate microbeads were incubated in a PLL solution, resulting in a core-shell structure with a core of alginate cross-linked by Ca2+ and a thin shell of PLL-that formed electrostatic interactions between the carboxyl of alginate and e-amino groups of PLL. The thin polymer shell around the alginate core stabilizes the overall microcapsule. Even if the alginate core became degraded due to ion exchange or chelation (in citrate or other solution), the microcapsule remains intact thanks to the PLL membrane.
The procedure described above was used to prepare microbeads to encapsulate bioreporter bacteria and allow the free passage of exogenously added small quorum-sensing (QS) molecules. This elicited a dose-dependent biochemical response from the bioreporter (Figure 10). The prepared bioreporter beads demonstrated biosensing ability towards both synthetic and secreted autoinducers. Furthermore, the bioreporter beads effectively detected the presence of a synthetic quorum-sensing inhibitor, furanone C-30. These beads exhibited excellent storability in all tested conditions, requiring no additional preparation before use. This makes the microbeads suitable as an on-demand capsule for potential wide application in monitoring and diagnostics.
2. Materials and Methods
2.1. Materials
Sodium alginate (low viscosity, A2158), trimethoprim, poly-l-lysine, poly-D-lysine, calcium chloride, Luria-Bertani medium (LB broth), and LB agar were purchased from Germany. Acyl-homoserine lactone (acyl-HSL) 3-oxo-C12-HSL, C4-HSL, C6-HSL, 3-oxo-C8-HSL, 3-oxo-C6-HSL, and (as in Figure 2), and the stock (5 mM) solutions were stored at -20 °C until required, and QS inhibitor - furanone C-30 were purchased from Sigma-Aldrich (St. Louis, MO, USA). Difco Luria-Bertani (LB) Broth, Miller (10 g L−1 tryptone; 5 g L−1 yeast extract; 10 g L−1 NaCl) and Difco LB agar, Miller (10 g L−1 tryptone; 5 g L−1 yeast extract; 10 g L−1 NaCl; 15 g L−1 agar) obtained from Becton (Dickinson & Company, France), LIVE/DEADTM BacLightTM Bacterial Viability Kit (Invitrogen, Thermo Fisher Scientific, USA).
2.1.1. Bacteria Strains
LasR and RhlR reporters- P. aeruginosa PAO-JP2-luxCDABE and PAO-JP2 (pKD-rhlA) QS reporters
PAO-JP2 (pKD201-lasI) is a double mutant (lasI/rhlI-deleted) strain of P. aeruginosa PAO1. It harbors a plasmid pKD201 with a lasI promoter coupled upstream to the luxCDABE luminescence system operon, which is responsive to 3OC12-HSL for LasR activity detection, as described previously by Ganin et al. [35]. PAO-JP2 (pKD-rhlA) is a lasI-rhlI double mutant of Pseudomonas aeruginosa PAO1 that harbors a pKD vector with a rhlA promoter coupled upstream to the luxCDABE operon. K. Duan et al. [36] developed the reporter strain responsive to C4-HSL (Figure S1). The scheme of the plasmid is presented in supplementary materials, Figure S1.
2.2. Synthesis of Microcapsules
The microbeads synthesis device was fabricated similarly to what Ahn et al. describe [37]. The feed solution was 2% (w/v), and the reservoir was made of 150 mM CaCl2 in water. The feed was loaded into a syringe and fed through the capillary device mentioned above, with the feed flow rate typically being 30 ml/hr. The feed solution was converted into droplets using an air-driven droplet generator (Figure 3). As previously described, Droplets were sheared off from the capillary tip by pulses of compressed air into an unstirred reservoir solution; after that, they were crosslinked into capsules. The capsules were allowed to crosslink for 10 minutes, after which they were washed and stored in DDW for subsequent steps.
Figure 3.
Schematics of Microcapsules synthesis. The size of the beads can be controlled by changing the flow rate of the feed solution (alginate + bioreporter) and the air pressure.
Figure 3.
Schematics of Microcapsules synthesis. The size of the beads can be controlled by changing the flow rate of the feed solution (alginate + bioreporter) and the air pressure.
2.3.1. Bacteria Strain Growth Conditions
All cells involved in encapsulation and characterization experiments were PAO-JP2 (pKD201-lasI) and PAO-JP2 (pKD-rhlA), abbreviated as LasR and RhlR, respectively. The glycerol stocks of reporter strains PAO-JP2 (pKD201-lasI) or PAO-JP2 (pKD-rhlA) were stored at -80 °C. The stocks were streaked onto an LB agar plate containing 300 µg/ml trimethoprim and incubated at 37 °C overnight in an incubator (Binder, Camarillo, CA, USA) to form colonies and stored in the refrigerator for less than a month. Starter cultures of LasR or RhlR were prepared by individually introducing a single colony into 10 ml of LB (Difco Lenox medium, BD, France) broth supplemented with 300 µg ml-1 of TMP (Sigma-Aldrich, MO, USA) and grown overnight in a shaking incubator (New Brunswick™ Innova® 44/44R Shaker) at 37 °C and 220 rpm shaking to attain exponential growth phase. The fast-growing bacteria were reinoculated into fresh LB broth in a 1:50 dilution and grown to an approximate optical density of 0.6-0.9 at 600 nm (OD600). Then, a determined volume of the bacteria culture was pelleted in a sterile 50 ml tube by centrifugation at 4 °C and 3000 rpm for 15 min. The supernatant was discarded while the cell pellet was resuspended in 1000-2000 µL of LB and refrigerated until use.
2.3.2. Encapsulation of Bacteria in Single/Multiple-Layer Microcapsules
Poly-L/D-Lysine stabilized bacteria-loaded alginate microbeads were prepared using a simple two-step approach similar to the method [37]. First, 1 ml of the diluted bacteria was added to a 4 ml portion of 2.5% w/v alginate solution (resulting in 2% w/v alginate solution) and mixed well using a revolver tube rotator. The alginate bacteria solution was extruded into 150 mM CaCl2 under a controlled extrusion flow rate and air pressure using the above-described capillary setup (Figure 3), and the resulting microbeads were allowed to harden for 10 min and rinsed with sterile double-distilled water (DDW). The microbeads were incubated in 2 ml of 0.05% w/v solution of PLL (or PDL) for 10 mins to allow the formation of a thin layer around the microbead, followed by washing with DDW. The resulting bacteria-alginate/PLL microbeads were subjected to 10 min incubation in 0.05% w/v alginate to form an outer layer on the capsules and to neutralize the non-reacted PLL, creating a negatively charged surface, and then rinsed again. This standard encapsulation approach was adopted in all subsequent experiments unless otherwise stated.
For double-membrane microbeads, the same procedure was followed as for single-membrane microbeads, except that the PLL incubation time was 7.5 min. After the alginate step, the microcapsules were incubated for 7.5 mins in 2 ml 0.05% PLL, rinsed, and incubated in 0.05% alginate for 10 min, and the rinse resulting in double membrane microbeads.
Three-membrane microbeads were prepared in the same way as the double membrane. After the 0.05% w/v alginate step, the double membrane microbeads were incubated in 2 ml of 0.05% w/v PLL for 5 min and washed. The microbeads were then coated again with a 0.05% w/v alginate solution. The size (diameter) of the beads was determined using a standard bright-field microscope, a 2.5x objective lens, and ImageJ. Alginate-polylysine and alginate-alone beads were tested for stability in the presence of a Ca2+ ion chelator, sodium citrate.
2.4. X-ray Photoelectron Spectroscopy (XPS) Analysis
X-ray photoelectron spectroscopy (XPS) analysis was conducted on the dehydrated microcapsules, beads, alginate, and PLL films using an "ESCALAB Xi+" instrument by Thermo-Fisher Scientific. The ionization energies of the various elements present in the samples were measured. The analysis was performed under an ambient pressure of <1·10-10 mbar. Aluminum Kα radiation source (1486.68eV) was used for photoelectron emission at room temperature, and the spectra were collected at a 90˚ angle from the X-ray source. A low-energy electron flood gun was employed to minimize surface charging, and the measurements were taken with a spot diameter of 650 µm. Spectral analysis was carried out using the Avantage software version 6.6.0, provided by Thermo-Fisher Scientific. High-resolution spectra of C1s, O1s, N1s, and Ca1s peaks and a survey spectrum at 20 eV pass energy were obtained.
2.5. Bioluminescent Assay
The bioluminescent response of the alginate-poly-lysine immobilized bioreporter - PAO-JP2 (pKD201-lasI) and PAO-JP2 (pKD-rhlA) strains were acquired throughout 20-30 h using the Luminoskan Ascent Luminometer (Thermo Fisher Scientific, Waltham, MA, USA) maintained at 37 °C. The luminometer was set to a kinetic mode at 5- or 10-minute intervals between readings. White (non)-transparent 96-well flat-bottom microplate (Nunc, Denmark) was used in all experiments. The experiments were carried out in three/four biological replicates with at least two independent experiments. The maximum luminescence values (expressed in the relative luminescent unit, RLU) were extracted using Microsoft Office Excel, from where the induction factor was calculated as a ratio of the maximum RLU of the test to that of the negative control (in the absence of inducer).
2.5.1. Viability Assay of the Immobilized Bacterial Strains
Microcapsules containing LasR and RhlR strains were placed in a sterile 24-well plate containing LB-broth and were mildly shaken for about 25 hours in an orbital shaker in a hot room maintained at 30 °C. For live/dead staining, capsules were washed with saline solution (0.9% w/v NaCl) three times. After washing, capsules were incubated at room temperature for 15 min in a saline solution containing propidium iodide (dead) and SYTO™ 9 (live) stain washed with saline and transferred to the ibid 12-well plate for confocal microscope imaging.
2.5.2. The Detection of Synthetic AHLs (3-Oxo-C12-HSL and C4-HSL)
The ability of the immobilized reporter bacteria to detect the sub-nanomolar concentration of the inducer was assessed by incubating the biosensor microbeads in the presence of a known concentration of the inducers.
Typically, 3-10 microbeads were added to each well in a white non-transparent 96-well flat-bottom microplate (Nunc, Roskilde, Denmark), followed by the addition of 190 µl LB broth (containing 300 µg/ml TMP) incubated at 37 °C with/without shaking at 220 rpm for a specified period. Dedicated calibration curves were obtained by adding 10 µl of the QS molecules to the well containing the microbeads at a range of final concentrations, 5 µM-500 pM of 3-oxo-C12-HSL (for the LasR reporter), and 10 µM-100 pM of C4-HSL (for RhlR reporter). The plate was incubated for 20-30 h at 37 °C, and during this time, luminescence measurements were performed at 5- or 10-min intervals using a luminometer Luminoskan ascent multi-mode reader (Thermo Scientific, Shanghai, China) luminometer (Luminoskan, Thermo Fisher Scientific, MA, USA). The equations derived from the calibration curves were used to calculate the concentration of 3OC12 -HSL and C4-HSL present in both culture supernatant and biofilm flow-through samples. All the experiments were carried out in three or four replicates and repeated at least twice.
2.5.3. Detection of AHLs in the Supernatant of Wild Cultures of PAO1 Strains
The biosensor was employed to detect the presence of the QS molecules in the supernatants of the culture of a wild strain of P. aeruginosa. The overnight culture of PAO1 was briefly reinoculated and grown overnight in a shaking incubator (New Brunswick™ Innova® 44/44R Shaker) at 37 °C and 220 rpm shaking until OD600 was about 0.9. Then, a determined volume of the bacteria culture was pelleted by centrifugation at 4 °C and 10,000 xg for 10 min. The supernatant was filtered through a 0.22 µm membrane filter and used as a source of AHLs for the biosensor testing.
2.5.4. The Detection of AHLs in the Biofilm Effluent Obtained in the Flow Cell Experiment
The biofilm experiment was carried out under the flow conditions using a standard method [38], as shown in the following supplementary scheme (Protocol S1 and Figure S7). The flow was allowed to continue undisturbed for 12, 24, 48, and 72 h, and the flow through (effluent) at the determined time was collected, filtered using 0.22 µm membrane filter, and stored at -80 °C until required. The flow-through was used as the source of AHLs for the biosensors testing. As stated in the supplementary section (Protocol S2), the resulting biofilms were macroscopically visualized using CLSM and 3D processed by IMARIS software.
2.5.5. Quorum Sensing Inhibition Assay
The ability of the biosensor to detect quorum sensing inhibitor was evaluated by incubating the biosensor microbeads with various concentrations of a synthetic quorum sensing inhibitor (furanone-C-30) in the presence of a known concentration of autoinducer or cell-free supernatant of wild P. aeruginosa strain (PAO1). The bioluminescent measurement was carried out as usual, and the percent inhibition was determined according to the following equation:
2.6. Statistical Analysis
A two-way ANOVA followed by Tukey’s multiple comparisons test or one-way ANOVA was performed using GraphPad Prism version 8.0.2 for Windows (GraphPad Software, San Diego, California, USA, www.graphpad.com).
3. Results
3.1. Encapsulation and Optimization
3.1.1. Optimization of Microbeads Formation
Using the experimental setup described earlier, it was possible to control the bead's size by controlling the microfluidic pump's air pressure and flow rate. At a constant flow rate of 10 ml/h, higher air pressure (valve opening) produced correspondingly smaller-sized beads until 40 psi (Figure S2). Much smaller beads were obtained when the flow rate was tripled (30 ml/h) at a constant air pressure of 40 psi. Therefore, a 30 ml/h flow rate, 40 psi pressure, and 15 units aperture were adopted in all the subsequent experiments.
3.1.2. Effect of Encapsulation Methods
Next, our research investigated the encapsulation of bacteria in the microbeads. The process of encapsulating bacteria within alginate presents challenges due to potential gel degradation from ion exchange or rapid bacteria growth, which could lead to bacteria escaping from the gel matrix [39]. To address this issue, the polymer shell surrounding the alginate core can serve to protect the cells and maintain them within the core. It is important to note that the polymer shell allowed for the diffusion of small molecules in and out of the core, which is crucial for maintaining cell viability.
Two approaches were investigated to prepare a poly-lysine-reinforced alginate microbead encapsulation system. The first approach involved preparing an alginate inner sphere followed by stepwise incubation in a poly-lysine solution to create a thin membrane around the alginate (Figure 3). In the second approach, the microbeads were dropped into a calcium-poly-lysine solution, followed by two additional incubations in a PLL solution (Figure S3). Both approaches led to reinforced alginate microbeads that are stable when incubated in the presence of 5% sodium citrate.
3.1.3. XPS Data
XPS was applied to quantitatively determine the elemental and chemical composition of the outermost portion of the alginate-poly-lysine microcapsules. This technique is surface-sensitive in that only a sample's outermost (about 10 nm) is analyzed. High-resolution spectra were recorded for oxygen, nitrogen, and carbon. Table 1 and Figure 4 shows that the samples' surface elemental compositions (expressed as relative atomic percentages) were computed from the XPS spectra recorded in survey mode. Nitrogen, however, is only a minor component of not more than 10.7% of the total PLL molecule [40]. The actual difference between alginate and alginate-polylysine can be calculated by dividing the N/C ratio of PLL by the N/C ratio of the capsule membrane. This calculation shows that alginate-PLL membranes comprise 56% PLL and 65% PDL, respectively (Figure 4).
3.1.4. Microbead Stability and Swelling
The main reason for adding a polylysine shell around the alginate gels was to prevent their degradation when they came in contact with certain ions or chelators. So, the microbeads of bare alginate and alginate–polylysine were placed in 5% sodium citrate, a well-known chelator of Ca2+ ions. Figure 5 shows the result, which depicts the scheme of the alginate and PLL-coated beads incubated in citrate solution. As expected, the bare alginate microgels completely degraded within 60 min (Figure 6).
As seen in the schematics, the degradation occurred because citrate removed Ca2+ cross-links, leaving linear alginate chains no longer part of a 3-D network (Figure 5A). In contrast, alginate–PLL and alginate–PDL microbeads retained their sphericality in the citrate solution (Figure 5B). This result shows that regardless of any loss of cross-links from the alginate core, the polymer shells ensure that the microcapsules preserve their structural integrity, although the resulting alginate-PLL microbeads treated with citrate were weaker than the untreated counterparts.
The beads' morphological appearance and size were determined under various treatment conditions, such as freezing, drying, and rehydrating, as shown in Figure 6. Poly-L-lysine and poly-D-lysine coated alginate beads behaved alike in all the conditions tested (frozen, dried, and rehydrated). The beads shrank into irregular, smaller shapes upon drying, and then full sphericity was restored after rehydration in about 30 minutes (as shown in the upper frame in Figure 6).
3.2. Biological Activity Tests
3.2.1. Viability and Bioluminescence
The live/dead bacterial viability stain is a fluorescence-based approach used to monitor the bacteria population based on the integrity of the cell membrane. Cells with compromised membranes that are dead or dying are permeable to propidium iodide, which will stain red with propidium iodide. In contrast, cells with an intact membrane will stain green with SYTO™ 9. Figure S4 confirms the viability of the immobilized bacteria strains alive within the alginate-PLL beads. Also, the encapsulated bioreporters demonstrated a dose-dependent bioluminescent response to the exogenously added autoinducers (Figure 7), showing that the bacteria were not only alive but also functional and that the encapsulating microbeads were permeable to the added AIs. The bioluminescent response was not different (p < 0.05) between the bacteria cells encapsulated within microbeads coated with PDL or PLL, showing that the microbead was permeable towards the probed autoinducers. (Figure 8).
3.2.2. Bioluminescent Response of Alginate Reinforced with L and D Isomers of Poly-Lysine.
A possible alternative to PLL is the use of poly-D-lysine (PDL), a chiral form of α–-polylysine, which is widely used in myriad biomedical applications [41]. While PLL is susceptible to proteolytic degradation [41], PDL is not, because most biological enzymes do not recognize the D configuration on the D amino acid isomers [42,43,44]. This experiment is of interest in an application where what is being immobilized involves proteases of secretes protease as part of the physiological metabolism or in a scenario where the bioreporter microbeads might encounter protease in any unknown clinical or environmental samples. As a result, both isomers were independently used to reinforce alginate and tested for suitability in the bioassay tests. The results showed no significant difference (p < 0.05) between PLL and PDL regarding the physical features (Figure 4) and biosensor responses (Figure 8).
3.2.3. Effect of Activation
One significant advantage of immobilizing bacteria through encapsulation is the capability to store them in a ready-to-use form, typically at lower temperatures. We aimed to investigate whether the bacteria would require a recovery period after being stored in the refrigerator for an extended duration. As depicted in Figure 9, an incubation period of 15-30 minutes at 37 °C with shaking at 220 rpm seemed optimal for recording bioluminescent readings. Even the sample without pre-incubation exhibited a noticeable response. A significant difference was observed in the time taken for the measurements to peak.
(A) Bioluminescent response of LasR strain (B) Bioluminescent response of RhlR strain. The stored microspheres were analyzed with or without pre-incubation to determine the effect of pre-incubation on the biosensor performance of the reporter bacteria strains. Experiments were conducted with 0, 15, 30, 60, 90, and 120 mins of incubation and shaking at 37 °C and 220 rpm. Each curve represents a mean of four replicate experiments
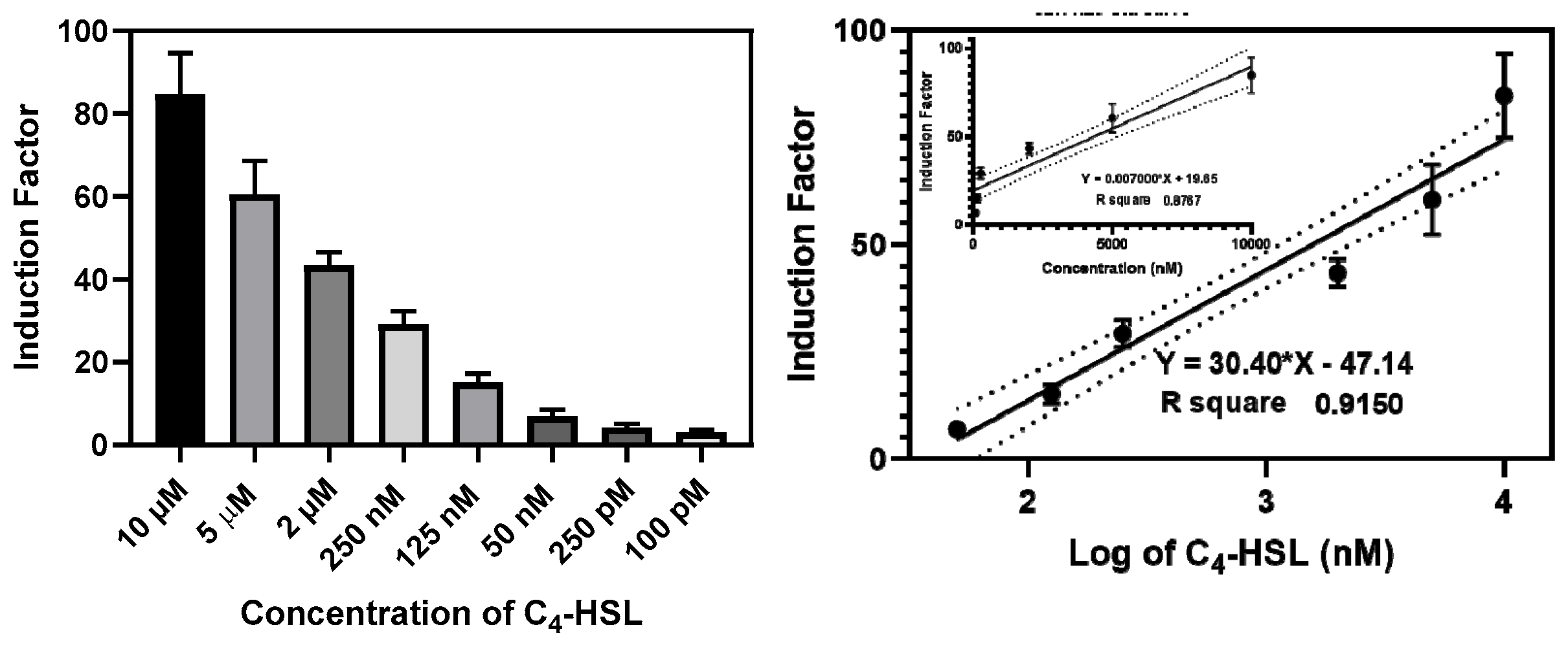
3.2.4. The Construction of the Calibration Curves
The bioluminescent responses of the bioreporter beads were measured at various concentrations of the autoinducers for 20-24 h. The induction factor (I.F) was calculated for each well, and the average I.F. of the replicates was plotted against the corresponding concentrations expressed in nanomolar or against the log of the concentrations (Figure 10 and Figure S10). The linear regression equation was generated to determine the 3-oxo-C12-HSL and C4-HSL present in the cell-free culture supernatant and biofilm effluent. The detection limit was set as the minimum autoinducer (AIs) concentrations that elicit a luminescent response ≥ 20% of the maximum obtained in an experiment without adding AIs.
Figure 10.
Calibration curve of RlhR bioreporter beads with various concentrations of C4-HSL. The left panel shows the luminescence produced by the reporter strain grown in LB in microtiter plates at 37 °C in the presence of increasing concentrations of exogenously added C4-HSL. The right panel shows the functions describing the dose-response relationships inferred from the data in the left panel. Regression lines were drawn on logarithmic (upper insert) and linear scale. The equation and the R2 value for each regression line are shown. Data are the mean ± SD of three independent experiments.
Figure 10.
Calibration curve of RlhR bioreporter beads with various concentrations of C4-HSL. The left panel shows the luminescence produced by the reporter strain grown in LB in microtiter plates at 37 °C in the presence of increasing concentrations of exogenously added C4-HSL. The right panel shows the functions describing the dose-response relationships inferred from the data in the left panel. Regression lines were drawn on logarithmic (upper insert) and linear scale. The equation and the R2 value for each regression line are shown. Data are the mean ± SD of three independent experiments.
3.3. Selectivity and Inhibition Studies
3.3.1. Selectivity of the Bioreporters to the Synthetic and Secreted QS Molecules
The bioluminescent response of the bioreporter was tested in the presence of synthetic autoinducers (Figure 11A and B) and cell-free supernatant obtained from Pseudomonas aeruginosa, Acinetobacter baumannii, Escherichia coli (Figure 11 C and D). Each strain of the reporter bacteria in this study distinctly responded to their cognate autoinducers. The bioreporter selectively responded to the cell-free samples of P. aeruginosa but not E. coli or A. baumannii, as shown in Figure 13. This is desirable because the biosensor can selectively detect P. aeruginosa-specific autoinducer in the presence of other bacteria in the clinical and environmental samples.
3.3.2. Determination of AIs in the flow-Cell Biofilm Effluent
Biofilm formation was followed microscopically and as a function of autoinducer secretion and confirmed by luminescence assays with RlhR and LasR bioreporters (Figure S9). The effluents of flow cell biofilm were collected at 12, 24, 48, and 72-hour intervals and filtered. The filtered effluents were used as the source of autoinducers, and the bioluminescence responses were recorded as usual. The bioluminescence was interpolated from the standard plot.
3.3.3. Inhibition Studies
The biosensor's ability to detect the presence of an inhibitor was tested by incubating it with either synthetic autoinducers (Figure 12) or cell-free supernatants of the wild PAO1 type in the presence of varying concentrations of QSI (Figure S11 and S12).
3.4. Storability Assessments
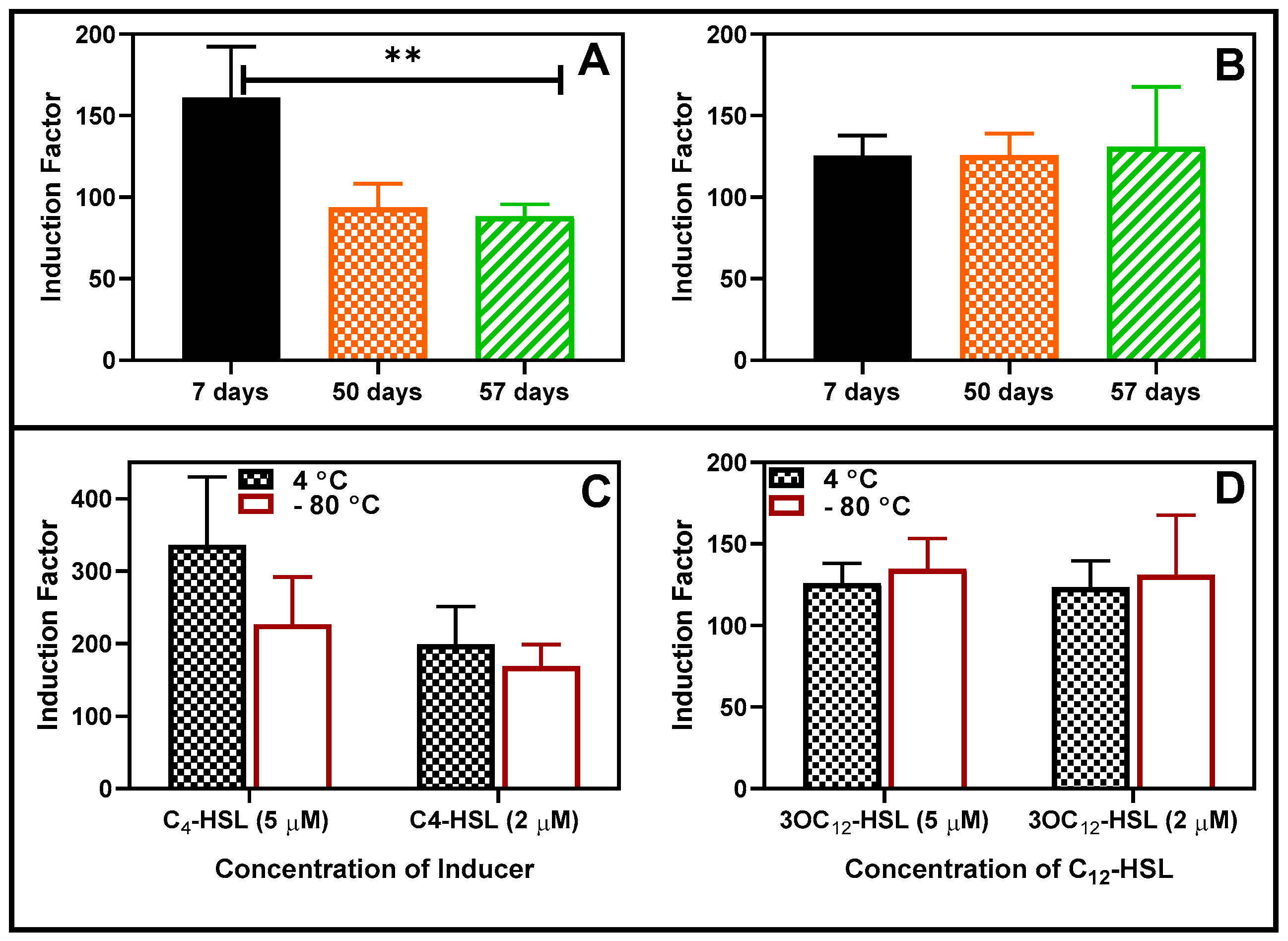
The bioreporter beads were tested for long storability under different temperatures. First, the bacterial beads were stored at -80 °C for 48 hours, after which their activities were tested and compared with the corresponding beads stored at 4 °C (Figure S6). There was no statistically significant difference between the responses to the added autoinducers. The bioreporters were further stored at 4 °C for at least 57 days, and the activities were tested at intervals (Figure 13 A and B). After that, the beads were stored at -80 °C and 4 °C for 30 days, after which the activities were tested and compared as usual and found to be the same at p < 0.05 (Figure 13 C and D).
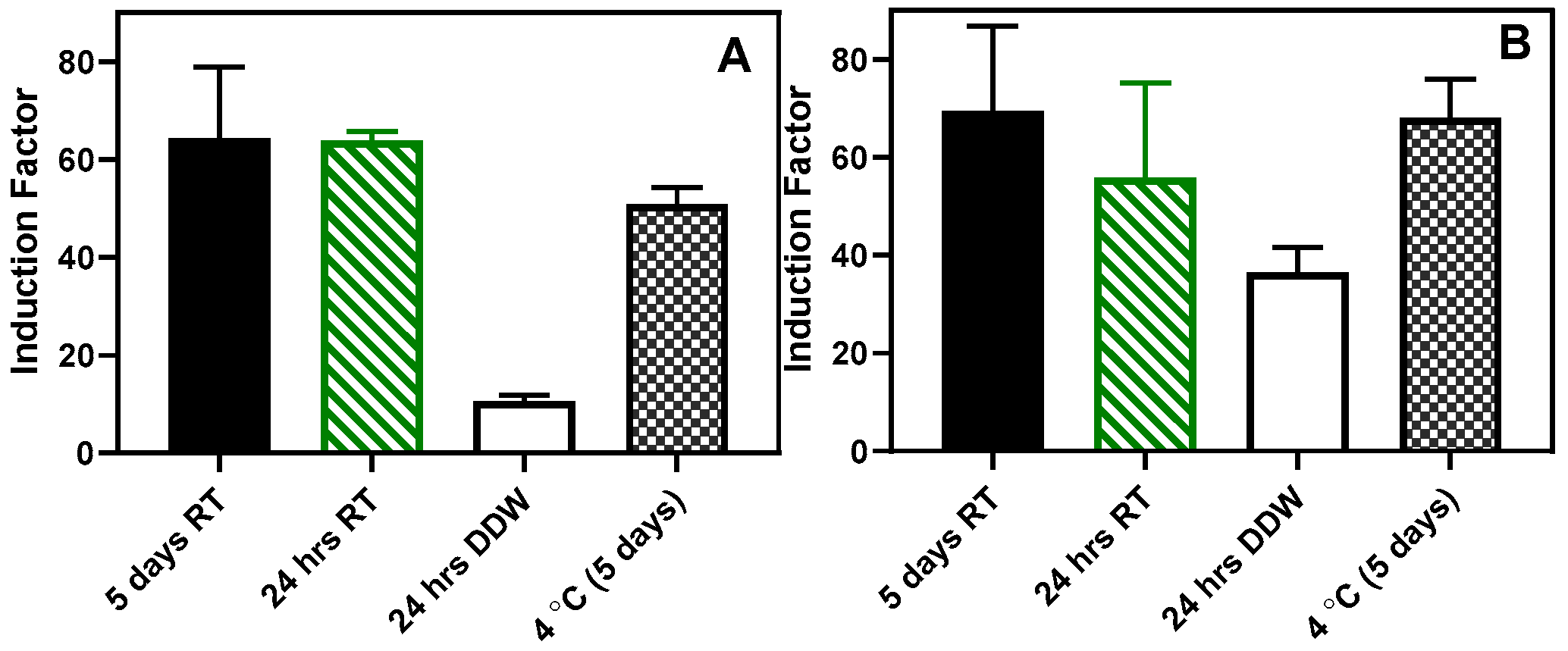
Also, an attempt was made to store the beads at room temperature in dry and wet conditions. As shown in Figure 14, the beads stored in a dried state for 24 hrs and 5 days retained the sensing ability to the autoinducer without statistically significant difference (Figure 14). However, in both RhlR and LasR strains, when the beads were stored in sterile DDW for 24 hours at room temperature, there was a significant (P < 0.05) loss in the sensing ability of the immobilized bioreporters. Similarly, when the beads were lyophilized, there was a considerable loss in the residual bioluminescent response (Figure S13)
Figure 13.
Storability of the bioreporter beads in the refrigerator condition. (A) RhlR (B) LasR 2 µM inducers. RhlR strain experienced a significant (P < 0.05) reduction in activities between days 7 and 50 and remained fairly constant thereafter. Comparison of the Effect of Storage at 4 and -80 degrees (for 30 days) on the luminescence. The residual activity of RhlR (C) and LasR (panel D) was compared after storage in the refrigerator and freezer at -80°C for a minimum of 30 days. Two concentrations of each AHL were tested, and the results are reported as mean ± SD, n=4. Statistical analysis revealed no significant differences in the readings at p < 0.05.
Figure 13.
Storability of the bioreporter beads in the refrigerator condition. (A) RhlR (B) LasR 2 µM inducers. RhlR strain experienced a significant (P < 0.05) reduction in activities between days 7 and 50 and remained fairly constant thereafter. Comparison of the Effect of Storage at 4 and -80 degrees (for 30 days) on the luminescence. The residual activity of RhlR (C) and LasR (panel D) was compared after storage in the refrigerator and freezer at -80°C for a minimum of 30 days. Two concentrations of each AHL were tested, and the results are reported as mean ± SD, n=4. Statistical analysis revealed no significant differences in the readings at p < 0.05.
4. Discussion
Monitoring quorum sensing molecules (QSMs) has become a crucial analytical tool in researching bacteria-related disturbances. The levels of these signaling molecules can potentially serve as biomarkers for disease and environmental monitoring. Therefore, there is an increasing demand for analytical methods that can sensitively, quantitatively, rapidly, and cost-effectively detect bacterial QSMs. Whole-cell biosensing systems have been shown to detect QSMs directly with little or no sample pretreatment. Moreover, microorganisms are tolerant of suboptimal assay conditions and can be prepared in unlimited quantities relatively inexpensively, making them candidates of choice for high-throughput screening, miniaturization, and automation [45].
In this study, we postulated that stable immobilization of bioreporter bacteria within reinforced alginate, could offer robust whole-cell sensing systems for monitoring the levels of AHLs. First, polylysine-coated alginate beads were prepared, which remained intact in the presence of the known Ca2+ chelator citrate (Figure 5). The bacteria were retained within the beads, and the bioluminescent assays demonstrated the passage of autoinducers and quorum-sensing inhibitors into the core of the beads.
The alginate-poly-lysine capsules utilized in this research exhibited similar physical properties regardless of the type of poly-lysine (L and D) isomers and identical permeability to the QS molecules. This suggests that either L or D isomers of poly-lysine can be employed to reinforce alginate based on specific applications. For instance, the D-isomer is known to be less susceptible to proteolysis, making it suitable for encapsulation applications that are likely to encounter protease. For this study, the L-isomer of poly-lysine was utilized in all experiments. The physicochemical analyses presented clear evidence of the presence of alginate and PLL or PDL in the membrane of the APA microcapsules. The X-ray photoelectron spectroscopy (XPS) analysis of the microcapsules' elemental composition indicated that relatively high amounts of PLL were very close to the surface, within the outermost 10 nm. Tam et al. [46] and De Vos et al., [40,47] They obtained similar surface composition using XPS, implying that these results were predictable. Furthermore, the spectroscopic analyses indicated that both alginate and the PLL (or PDL) were present at the surface, suggesting that at least some of the polysaccharide bound to the membrane PLL during the capsule's final incubation in dilute alginate. We excluded the possibility that the alginate at the surface originated from the gel core since XPS detected no calcium. Thu et al. [48] also demonstrated, using radiolabeling techniques, that the coating alginate binds to the microcapsule's PLL layer. Furthermore, the confocal microscopic study of the binding of PLL to the alginate reveals the formation of a polyanion/polycation outer thin shell-like membrane [49]. The three-dimensional image of alginate-PLL capsules showed no visible holes in the PLL membranes, indicating that PLL covers the capsule completely. It was further demonstrated that the PLL-alginate interaction was unchanged when stored for two weeks and two years [49]. This shows the stability of the additional support provided by the PLL.
It was observed in this study that the PLL-coated alginate beads were protected from the effects of a known metal chelator, sodium citrate. Chelators such as EDTA and sodium citrate can induce complete degradation of alginate gels, while the gels may undergo partial degradation when immersed in buffers like phosphate-buffered saline (PBS). This occurs as some Ca2+ cross-links are exchanged with Na+, reducing mechanical rigidity and cross-link density and ultimately weakening the gel. When coated and uncoated alginate beads were incubated in citrate 2.5-5% w/v), the uncoated alginate capsule was completely dissolved. In contrast, PLL-coated alginate beads as well as the dual layer having the interpenetrating network, persisted in 5% citrate solution for over 14 hours at room temperature (Figure 5 and Figure S3). This shows the protective effect of the PLL layers on the materials encapsulated within the alginate beads. Thus, regardless of any loss of cross-links from the alginate core, the poly-lysine layers will ensure that the microcapsules preserve their structural integrity, similar to the report of Gugerli et al. [37]. Additionally, the dried beads regain their sphericity upon rehydration.
Pseudomonas aeruginosa contains two transcription regulators (LasR and RhlR) that, when complexed with their specific autoinducers (3-oxo-C12-HSl and C4-HSL, respectively), activate transcription of different virulence-associated traits and enzymes involved in rhamnolipid biosynthesis [50]. In this study, mutant strains of Pseudomonas aeruginosa were utilized. These strains have a knockout on the gene expressing 3-oxo-C12-HSL and C4-HSL autoinducers, necessitating the exogenous addition of the autoinducer for the activation of the transcription of the promoter-controlled bacterial genes fused to the reporter (luciferase) gene. Consequently, the expression of the reporter genes in the mutant bioreporter can be correlated with the amount of exogenously added autoinducers in the media, forming the basis for detecting QS molecules (autoinducers) using bioreporter bacterial strains.
Two mutant strains of P. aeruginosa (LasR and RhlR) were genetically engineered to express bioluminescence in response to specific quorum sensing (QS) molecules. These strains were individually encapsulated within alginate-PLL beads, following the procedure shown in Figure 3. LasR and RhlR could quantitatively respond to 3-oxo-C12-HSL and C4-HSL, respectively. Increased bioluminescence intensity was observed with increasing concentrations of the AHL compounds used. Upon preparation, the bioreporter-loaded microcapsules were incubated in culture media overnight (20 hours) at 30°C and 150 rpm. Subsequently, viability testing was performed, and confocal imaging confirmed the viability of the encapsulated bioreporter. The bioreporter-loaded beads were subjected to various concentrations (including absence) of autoinducers, and the resulting bioluminescence response was recorded. The obtained results provide evidence that the bacteria remain viable and exhibit their genetically programmed response, thus retaining the sensing functionality.
The plasmids used in this study were constructed by taking advantage of the RhlI/RhlR and LasI/LasR quorum sensing systems of the Gram-negative bacterium P. aeruginosa. C4-HSL and 3-oxo-C12-HSL are known to be the respective cognate activator molecules, i.e., the natural ligands that P. aeruginosa synthesize, sense, and respond to [51]. In this study we observed increased bioluminescence intensity with increasing concentrations of C4-HSL and 3-oxo-C12-HSL. The RhlR-based bioreporter beads demonstrate a dynamic range and limit of detection of 10 µM-0.1 nM and 50 pM, respectively, for C4-HSL. In contrast, the LasR-based bioreporter showed a dynamic range and a detection limit of 0.1 nM for 3-oxo-C12-HSL. A calibration curve was generated with either C4-HSL or 3-oxo-C12-HSL at known concentrations, which can be used to calculate the concentration of C4-HSL or 3oxo-C12-HSL activity in each sample. The molecule C4-HSL, at a concentration of 10 µM, induced the maximum response in the RhlR system carrying the plasmid pKD-rhlA. Meanwhile, 2 µM of the 3-oxo-C12-HSL molecule exhibited a maximum response in the bioreporter containing the plasmid pKD201 lasl. These findings align with previous studies, indicating that long-chain AHLs induce the LasR regulatory system to a greater extent, whereas short-chain AHLs efficiently activate the RhlR regulatory system [52]. Additionally, the limit of detection (LOD) was defined as the lowest concentration of AHL capable of inducing a 20% increase in luminescence by the biosensor without any added molecule.
The wide range of response and the high sensitivity of these bioreporters suggest that the bacteria strain in this study could be used for the direct determination of 3-oxo-C12-HSL and C4-HSL levels in P. aeruginosa laboratory cultures and clinical samples where P. aeruginosa is implicated. According to previous reports, the highest concentration of AHLs in laboratory cultures and clinical samples (cystic fibrosis sputa) is ≤ 28 μM and ≤ 21 nM, respectively [12,53].
After assay optimization, our whole-cell sensing systems were employed to detect the presence of AHLs in samples collected from cell-free biofilm and planktonic bacteria cultures. We also evaluated our whole-cell sensing systems for their ability to detect the presence of quorum sensing inhibitors (QSI) by measuring the residual QS activity in the presence of autoinducers and QSI. To this end, we tested our system's response to furanone C-30 (FC30), a well-known QSI molecule that interferes with RhlR and LasR QS signaling [54]. The system exhibited a similar inhibition response to FC30 when tested in the presence of either synthetic autoinducers (Figure 12) or cell-free culture of the PAO1 strain of P. aeruginosa (Figure S11). Furthermore, the system maintained its sensing ability when stored at 4°C for over 60 days and frozen at -80°C. Lyophilization resulted in a significant loss of bioluminescence response, but simple air-drying of the bioreporter did not affect its sensing capability. The biosensor strains did not require special treatment to revive them after long storage; they existed as ready-to-deploy microcapsules. Overall, the bioreporter encapsulated in alginate-PLL microbeads has the potential to be a sensitive, easy-to-prepare, and easy-to-deploy analytical unit suitable for various biotechnological applications in both developed and low-income countries.
5. Conclusions
We have developed hydrogel-immobilized bioreporters as on-demand whole-cell biosensing systems for the sensitive, selective, and rapid detection of AHLs in spiked samples and laboratory cultures. The method is simple, sensitive, reproducible, and amenable for point-of-need deployment without extensive sample preparation. The RhlR-based bioreporter beads demonstrate a dynamic range and limit of detection of 10 µM-0.1 nM and 50 pM, respectively, for C4-HSL, while the LasR-based bioreporter showed a dynamic range and a limit of detection of 0.1 nM for 3-oxo-C12-HSL.
The immobilized bioreporters can be stored stably and refrigerated as ready-to-use microbeads for over 60 days. Furthermore, the method developed is cost-effective and robust, characterized by high throughput and adaptability for screening QS inhibitors. We believe this novel biosensing system could be employed in the diagnosis and environmental monitoring. Additionally, we believe that these biosensing systems could be successfully employed to study the effect of various agonist and antagonist molecules on the quorum sensing mechanism, providing a valuable tool for the screening of novel QSI-based drugs active against P. aeruginosa infection and a variety of diseases that involve QS in their pathogenesis.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, AAP. and RSM; methodology, APP and YSK; validation, RSM and AK; formal analysis, AAP; investigation, AAP, and YSK; writing—original draft preparation, AAP; writing—review and editing, AAP; YSK, AK, and RSM; supervision, RSM and AK; project administration, RSM and AK; All authors have read and agreed to the published version of the manuscript.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article and the supplementary materials; further inquiries can be directed to the corresponding author (rsmarks@bgu.ac.il)
Acknowledgements
The project is partially funded by the European Union’s Horizon 2020 research and innovation program under the Marie Skłodowska-Curie grant agreement No 872662, IPANEMA project. Furthermore, the authors thank the Avram and Stella Goldstein-Goren fund for partial support. We also thank Dr. Victor Multanen from the Department of Civil and Environmental Engineering for his help during microscopy, Ms. Kathelina Kristollari for her help with biorender illustrations, and Dr. Guillaume Le Saux from the Department of Materials Engineering for his kind gift of poly-l-lysine.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Van Delden, C.; Iglewski, B.H.J.E.i.d. Cell-to-cell signaling and Pseudomonas aeruginosa infections. 1998, 4, 551. [CrossRef]
- Lyczak, J.B.; Cannon, C.L.; Pier, G.B.J.C.m.r. Lung infections associated with cystic fibrosis. 2002, 15, 194-222. [CrossRef] [PubMed]
- De Kievit, T.R.; Iglewski, B.H.J.I.; immunity. Bacterial quorum sensing in pathogenic relationships. 2000, 68, 4839-4849. [CrossRef]
- Winson, M.K.; Camara, M.; Latifi, A.; Foglino, M.; Chhabra, S.R.; Daykin, M.; Bally, M.; Chapon, V.; Salmond, G.; Bycroft, B.W.J.P.o.t.N.A.o.S. Multiple N-acyl-L-homoserine lactone signal molecules regulate production of virulence determinants and secondary metabolites in Pseudomonas aeruginosa. 1995, 92, 9427-9431. [CrossRef] [PubMed]
- Davies, D.J.N.r.D.d. Understanding biofilm resistance to antibacterial agents. 2003, 2, 114-122. [CrossRef]
- Hu, J.Y.; Fan, Y.; Lin, Y.-H.; Zhang, H.-B.; Ong, S.L.; Dong, N.; Xu, J.-L.; Ng, W.J.; Zhang, L.-H.J.R.i.m. Microbial diversity and prevalence of virulent pathogens in biofilms developed in a water reclamation system. 2003, 154, 623-629. [CrossRef] [PubMed]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P.J.J.o.b. Quorum sensing in bacteria: the LuxR-LuxI family of cell density-responsive transcriptional regulators. 1994, 176, 269-275.
- Erickson, D.L.; Endersby, R.; Kirkham, A.; Stuber, K.; Vollman, D.D.; Rabin, H.R.; Mitchell, I.; Storey, D.G.J.I.; immunity. Pseudomonas aeruginosa quorum-sensing systems may control virulence factor expression in the lungs of patients with cystic fibrosis. 2002, 70, 1783-1790. [CrossRef] [PubMed]
- Pearson, J.P.; Gray, K.M.; Passador, L.; Tucker, K.D.; Eberhard, A.; Iglewski, B.H.; Greenberg, E.P.J.P.o.t.N.A.o.S. Structure of the autoinducer required for expression of Pseudomonas aeruginosa virulence genes. 1994, 91, 197-201. [CrossRef] [PubMed]
- Middleton, B.; Rodgers, H.C.; Cámara, M.; Knox, A.J.; Williams, P.; Hardman, A.J.F.m.l. Direct detection of N-acylhomoserine lactones in cystic fibrosis sputum. 2002, 207, 1-7. [CrossRef]
- Favre-Bonté, S.; Pache, J.-C.; Robert, J.; Blanc, D.; Pechère, J.-C.; van Delden, C.J.M.p. Detection of Pseudomonas aeruginosa cell-to-cell signals in lung tissue of cystic fibrosis patients. 2002, 32, 143-147. [CrossRef] [PubMed]
- Chambers, C.E.; Visser, M.B.; Schwab, U.; Sokol, P.A.J.F.m.l. Identification of N-acylhomoserine lactones in mucopurulent respiratory secretions from cystic fibrosis patients. 2005, 244, 297-304. [CrossRef]
- Barr, H.L.; Halliday, N.; Cámara, M.; Barrett, D.A.; Williams, P.; Forrester, D.L.; Simms, R.; Smyth, A.R.; Honeybourne, D.; Whitehouse, J.L.J.E.R.J. Pseudomonas aeruginosa quorum sensing molecules correlate with clinical status in cystic fibrosis. 2015, 46, 1046-1054. [CrossRef]
- Massai, F.; Imperi, F.; Quattrucci, S.; Zennaro, E.; Visca, P.; Leoni, L.J.B.; Bioelectronics. A multitask biosensor for micro-volumetric detection of N-3-oxo-dodecanoyl-homoserine lactone quorum sensing signal. 2011, 26, 3444-3449. [CrossRef]
- Ortori, C.A.; Atkinson, S.; Chhabra, S.R.; Cámara, M.; Williams, P.; Barrett, D.A.J.A.; chemistry, b. Comprehensive profiling of N-acylhomoserine lactones produced by Yersinia pseudotuberculosis using liquid chromatography coupled to hybrid quadrupole–linear ion trap mass spectrometry. 2007, 387, 497-511. [CrossRef]
- McLean, R.J.; Pierson III, L.S.; Fuqua, C.J.J.o.m.m. A simple screening protocol for the identification of quorum signal antagonists. 2004, 58, 351-360. [CrossRef]
- Kumari, A.; Pasini, P.; Deo, S.K.; Flomenhoft, D.; Shashidhar, H.; Daunert, S.J.A.c. Biosensing systems for the detection of bacterial quorum signaling molecules. 2006, 78, 7603-7609. [CrossRef] [PubMed]
- Kumari, A.; Pasini, P.; Daunert, S.J.A.; chemistry, b. Detection of bacterial quorum sensing N-acyl homoserine lactones in clinical samples. 2008, 391, 1619-1627. [CrossRef]
- Horáček, O.; Portillo, A.E.; Dhaubhadel, U.; Sung, Y.-S.; Readel, E.R.; Kučera, R.; Armstrong, D.W.J.T. Comprehensive chiral GC-MS/MS and LC-MS/MS methods for identification and determination of N-acyl homoserine lactones. 2023, 253, 123957. [CrossRef] [PubMed]
- Checa, S.K.; Zurbriggen, M.D.; Soncini, F.C.J.C.o.i.b. Bacterial signaling systems as platforms for rational design of new generations of biosensors. 2012, 23, 766-772. [CrossRef]
- Wang, Q.; Tang, S.-Y.; Yang, S.J.F.o.C.S.; Engineering. Genetic biosensors for small-molecule products: Design and applications in high-throughput screening. 2017, 11, 15-26. [CrossRef]
- Bazhenov, S.; Novoyatlova, U.; Scheglova, E.; Prazdnova, E.; Mazanko, M.; Kessenikh, A.; Kononchuk, O.; Gnuchikh, E.; Liu, Y.; Al Ebrahim, R.J.B.; et al. Bacterial lux-biosensors: Constructing, applications, and prospects. 2023, 13, 100323. [CrossRef]
- Struss, A.; Pasini, P.; Ensor, C.M.; Raut, N.; Daunert, S.J.A.c. Paper strip whole cell biosensors: a portable test for the semiquantitative detection of bacterial quorum signaling molecules. 2010, 82, 4457-4463. [CrossRef]
- Deng, X.; Zhuang, G.; Ma, A.; Yu, Q.; Zhuang, X.J.J.o.E.S. Construction of a dual fluorescence whole-cell biosensor to detect N-acyl homoserine lactones. 2014, 26, 415-422. [CrossRef] [PubMed]
- Lubkowicz, D.; Ho, C.L.; Hwang, I.Y.; Yew, W.S.; Lee, Y.S.; Chang, M.W.J.A.s.b. Reprogramming probiotic Lactobacillus reuteri as a biosensor for Staphylococcus aureus derived AIP-I detection. 2018, 7, 1229-1237. [CrossRef]
- Van Dyk, T.K.; Rosson, R.A.J.B.M.; Protocols. Photorhabdus luminescens luxCDABE promoter probe vectors. 1998, 85-95. [CrossRef]
- Daunert, S.; Barrett, G.; Feliciano, J.S.; Shetty, R.S.; Shrestha, S.; Smith-Spencer, W.J.C.r. Genetically engineered whole-cell sensing systems: coupling biological recognition with reporter genes. 2000, 100, 2705-2738. [CrossRef]
- Gao, Y.; Jin, X.J.M.d. Dual crosslinked methacrylated alginate hydrogel micron fibers and tissue constructs for cell biology. 2019, 17, 557. [CrossRef]
- Paul, A.A.; Markus, V.; Kristollari, K.; Marks, R.S. Alginate-Based Applications in Biotechnology with a Special Mention to Biosensors. 2023.
- Mehrotra, T.; Dev, S.; Banerjee, A.; Chatterjee, A.; Singh, R.; Aggarwal, S.J.J.o.E.C.E. Use of immobilized bacteria for environmental bioremediation: a review. 2021, 9, 105920. [CrossRef]
- Gómez-Mascaraque, L.G.; Martínez-Sanz, M.; Hogan, S.A.; López-Rubio, A.; Brodkorb, A.J.C.P. Nano-and microstructural evolution of alginate beads in simulated gastrointestinal fluids. Impact of M/G ratio, molecular weight and pH. 2019, 223, 115121. [CrossRef]
- Gupta, A.; Terrell, J.L.; Fernandes, R.; Dowling, M.B.; Payne, G.F.; Raghavan, S.R.; Bentley, W.E.J.B.; bioengineering. Encapsulated fusion protein confers “sense and respond” activity to chitosan–alginate capsules to manipulate bacterial quorum sensing. 2013, 110, 552-562. [CrossRef]
- Budianto, E.; Saepudin, E.; Nasir, M. The encapsulation of Lactobacillus casei probiotic bacteria based on sodium alginate and chitosan. In Proceedings of the IOP Conference Series: Earth and Environmental Science, 2020; p. 012043. [CrossRef]
- Liu, X.Y.; Nothias, J.-M.; Scavone, A.; Garfinkel, M.; Millis, J.M.J.A.j. Biocompatibility investigation of polyethylene glycol and alginate-poly-L-lysine for islet encapsulation. 2010, 56, 241-245. [CrossRef] [PubMed]
- Ganin, H.; Tang, X.; Meijler, M.M.J.B.; letters, m.c. Inhibition of Pseudomonas aeruginosa quorum sensing by AI-2 analogs. 2009, 19, 3941-3944. [CrossRef] [PubMed]
- Duan, K.; Surette, M.G.J.J.o.b. Environmental regulation of Pseudomonas aeruginosa PAO1 Las and Rhl quorum-sensing systems. 2007, 189, 4827-4836. [CrossRef]
- Gugerli, R.; Cantana, E.; Heinzen, C.; Stockar, U.v.; Marison, I.J.J.o.m. Quantitative study of the production and properties of alginate/poly-L-lysine microcapsules. 2002, 19, 571-590. [CrossRef] [PubMed]
- Golberg, K.; Markus, V.; Kagan, B.-e.; Barzanizan, S.; Yaniv, K.; Teralı, K.; Kramarsky-Winter, E.; Marks, R.S.; Kushmaro, A.J.P. Anti-Virulence Activity of 3, 3′-Diindolylmethane (DIM): A Bioactive Cruciferous Phytochemical with Accelerated Wound Healing Benefits. 2022, 14, 967. [CrossRef]
- Hunt, N.; Smith, A.M.; Gbureck, U.; Shelton, R.; Grover, L.J.A.b. Encapsulation of fibroblasts causes accelerated alginate hydrogel degradation. 2010, 6, 3649-3656. [CrossRef]
- de Vos, P.; Hoogmoed, C.G.; Busscher, H.J.J.J.o.B.M.R.A.O.J.o.T.S.f.B., The Japanese Society for Biomaterials,; Biomaterials, T.A.S.f.; Biomaterials, t.K.S.f. Chemistry and biocompatibility of alginate-PLL capsules for immunoprotection of mammalian cells. 2002, 60, 252-259. [CrossRef]
- French, T.; So, P.; Weaver, J., DJ; Coelho-Sampaio, T.; Gratton, E.; Voss, J., EW; Carrero, J.J.J.o.m. Two-photon fluorescence lifetime imaging microscopy of macrophage-mediated antigen processing. 1997, 185, 339-353. [CrossRef]
- Creighton, T.E. Proteins: structures and molecular properties; Macmillan: 1993.
- Maurer, P.H.J.T.J.o.e.m. Antigenicity of polypeptides (poly alpha amino acids) XIII. Immunological studies with synthetic polymers containing only D-or D-and L-α-amino acids. 1965, 121, 339-349. [CrossRef]
- Sela, M.J.A.i.i. Immunological studies with synthetic polypeptides. 1966, 5, 29-129. [CrossRef]
- Gu, M.B.; Mitchell, R.J.; Kim, B.C.J.B. Whole-cell-based biosensors for environmental biomonitoring and application. 2004, 269-305. [CrossRef]
- Tam, S.K.; Dusseault, J.; Polizu, S.; Ménard, M.; Hallé, J.-P.J.B. Physicochemical model of alginate–poly-l-lysine microcapsules defined at the micrometric/nanometric scale using ATR-FTIR, XPS, and ToF-SIMS. 2005, 26, 6950-6961. [CrossRef]
- de Vos, P.; van Hoogmoed, C.G.; van Zanten, J.; Netter, S.; Strubbe, J.H.; Busscher, H.J.J.B. Long-term biocompatibility, chemistry, and function of microencapsulated pancreatic islets. 2003, 24, 305-312. [CrossRef]
- Thu, B.; Bruheim, P.; Espevik, T.; Smidsrød, O.; Soon-Shiong, P.; Skjåk-Bræk, G.J.B. Alginate polycation microcapsules: I. Interaction between alginate and polycation. 1996, 17, 1031-1040. [CrossRef]
- Strand, B.L.; Mørch, Y.A.; Espevik, T.; Skjåk-Bræk, G.J.B.; bioengineering. Visualization of alginate–poly-L-lysine–alginate microcapsules by confocal laser scanning microscopy. 2003, 82, 386-394. [CrossRef]
- Medina, G.; Juárez, K.; Valderrama, B.; Soberón-Chávez, G.J.J.o.B. Mechanism of Pseudomonas aeruginosa RhlR transcriptional regulation of the rhlAB promoter. 2003, 185, 5976-5983. [CrossRef]
- Miller, M.B.; Bassler, B.L.J.A.R.i.M. Quorum sensing in bacteria. 2001, 55, 165-199. [CrossRef]
- Winson, M.K.; Swift, S.; Fish, L.; Throup, J.P.; Jørgensen, F.; Chhabra, S.R.; Bycroft, B.W.; Williams, P.; Stewart, G.S.J.F.m.l. Construction and analysis of luxCDABE-based plasmid sensors for investigating N-acyl homoserine lactone-mediated quorum sensing. 1998, 163, 185-192. [CrossRef]
- Chugani, S.A.; Whiteley, M.; Lee, K.M.; D'Argenio, D.; Manoil, C.; Greenberg, E.J.P.o.t.n.a.o.s. QscR, a modulator of quorum-sensing signal synthesis and virulence in Pseudomonas aeruginosa. 2001, 98, 2752-2757. [CrossRef]
- Markus, V.; Golberg, K.; Teralı, K.; Ozer, N.; Kramarsky-Winter, E.; Marks, R.S.; Kushmaro, A.J.M. Assessing the molecular targets and mode of action of furanone C-30 on Pseudomonas aeruginosa quorum sensing. 2021, 26, 1620. [CrossRef]
Figure 1.
Schematic of the cellular events that result in the expression of a reporter protein. The cell can easily take up a bioavailable analyte, such as AHL, through its membrane. AHL (3-oxo-C12-HSL and C4-HSL) binds with a regulatory protein called LasR or RhlR, forming a protein-AHL complex. This complex binds to its corresponding promoter, Plasl or Prhl, which triggers the activation of transcription and translation of a reporter gene called luxCDABE. This results in the expression of the reporter protein, luciferase, inducing the bioluminescent reaction characterized by the emission of blue-green light (~490 nm). Created with BioRender.com.
Figure 1.
Schematic of the cellular events that result in the expression of a reporter protein. The cell can easily take up a bioavailable analyte, such as AHL, through its membrane. AHL (3-oxo-C12-HSL and C4-HSL) binds with a regulatory protein called LasR or RhlR, forming a protein-AHL complex. This complex binds to its corresponding promoter, Plasl or Prhl, which triggers the activation of transcription and translation of a reporter gene called luxCDABE. This results in the expression of the reporter protein, luciferase, inducing the bioluminescent reaction characterized by the emission of blue-green light (~490 nm). Created with BioRender.com.
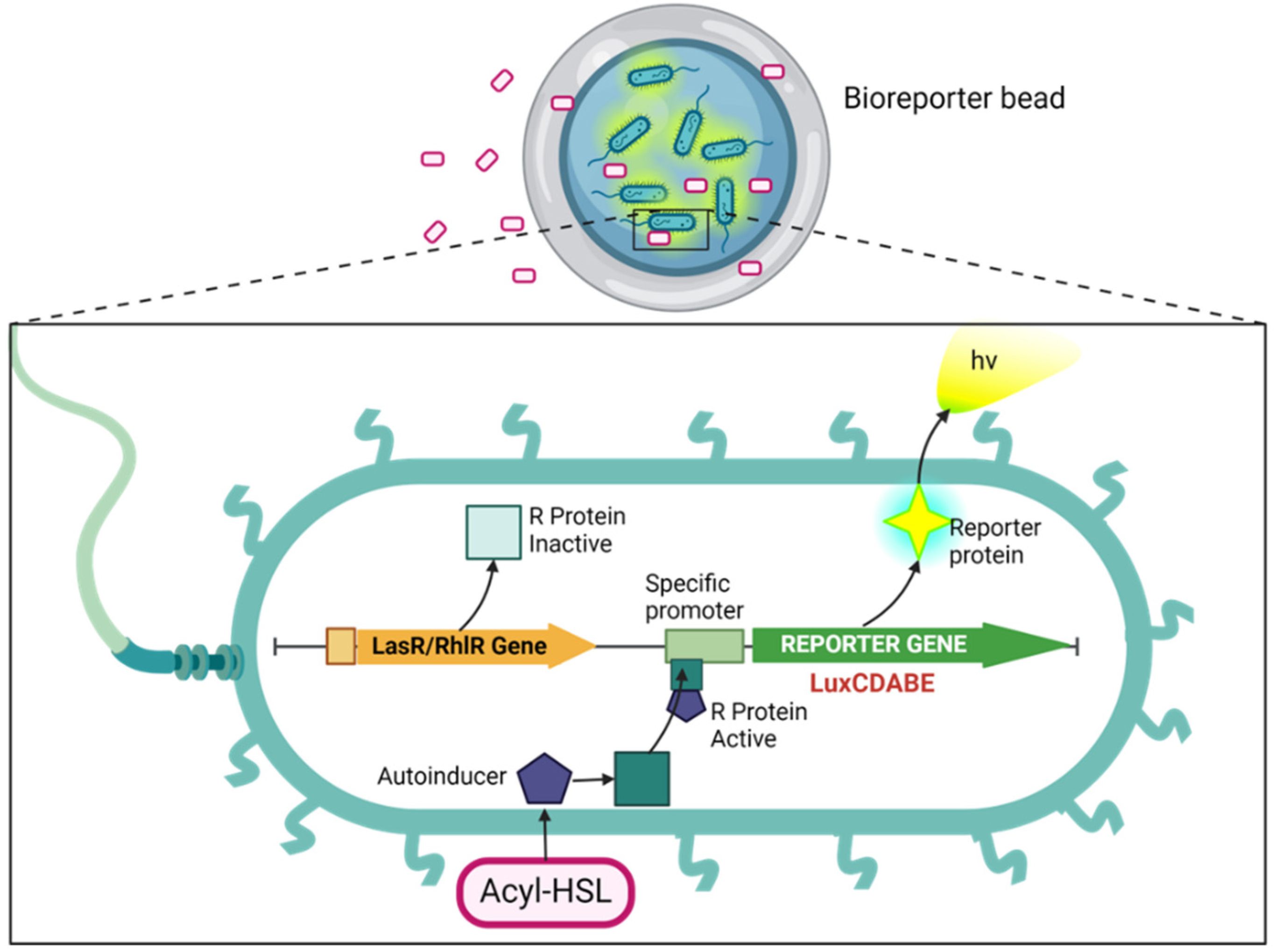
Figure 2.
The structures of QS molecules used in the study. C4-HSL and 3-oxo-C12-HSL are well-characterized P. aeruginosa autoinducers of RhlR and LasR quorum sensing signaling, respectively.
Figure 2.
The structures of QS molecules used in the study. C4-HSL and 3-oxo-C12-HSL are well-characterized P. aeruginosa autoinducers of RhlR and LasR quorum sensing signaling, respectively.
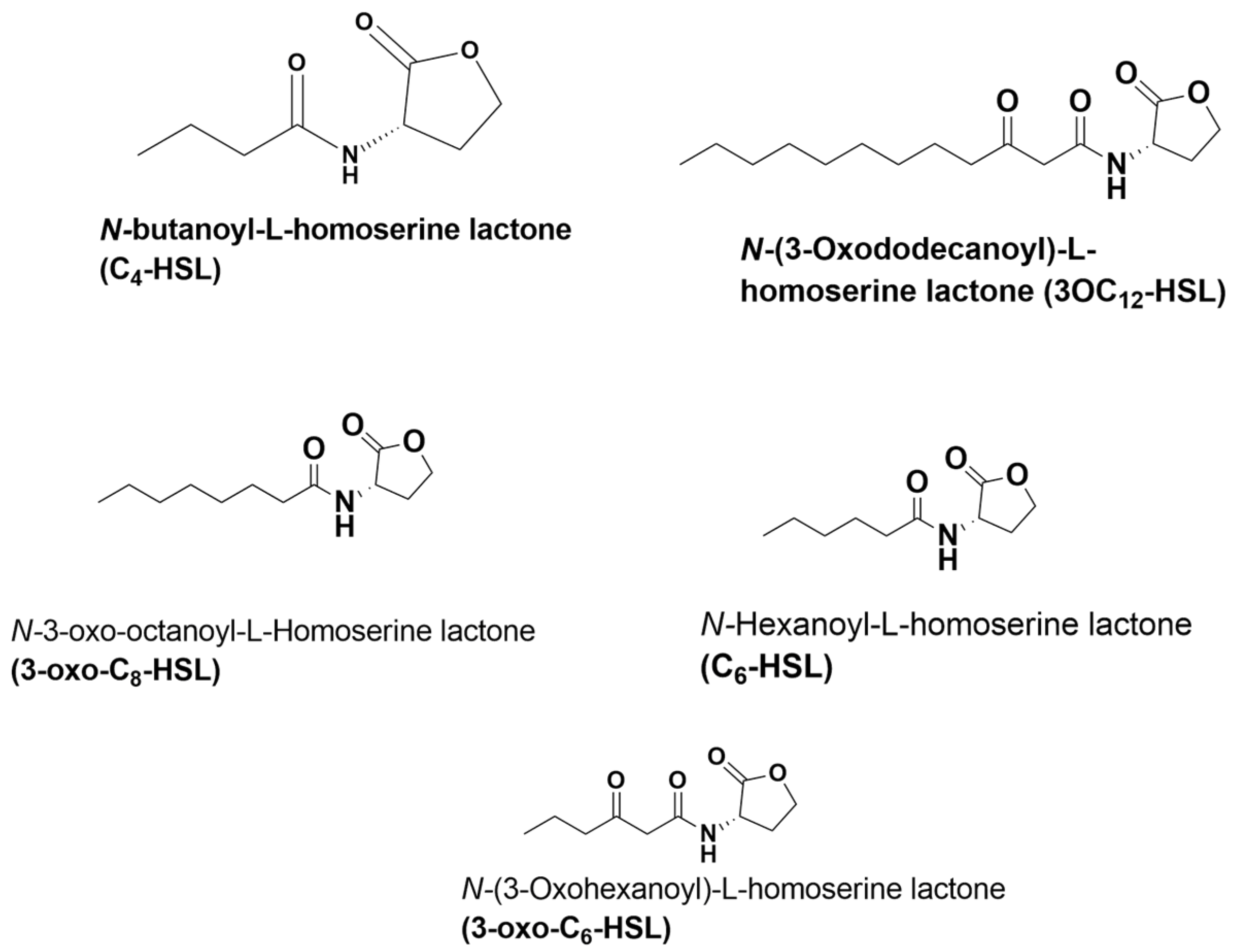
Figure 4.
XPS atomic peak composition of alginate-polylysine beads.
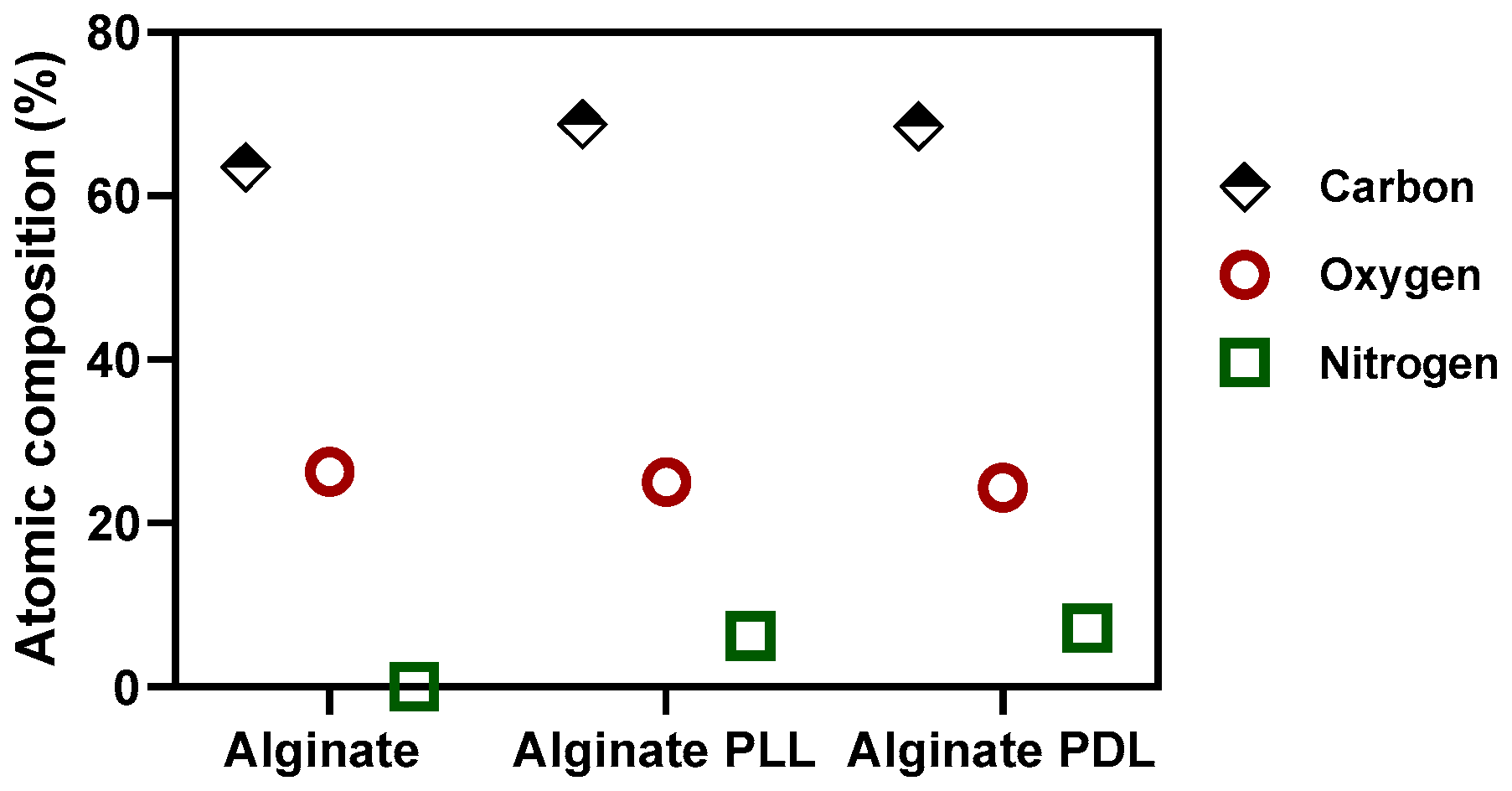
Figure 5.
Stability of alginate-poly-lysine microbeads in the presence of cation scavengers. (A) alginate microbeads (control) are rapidly degraded within 60 minutes when incubated in 5% w/v sodium citrate because of the Ca2+ removal from the hydrogels. (B) Alginate-PLL (C) Alginate-PLL IPN capsules remained stable in a 5% solution of sodium citrate after 14 hours.
Figure 5.
Stability of alginate-poly-lysine microbeads in the presence of cation scavengers. (A) alginate microbeads (control) are rapidly degraded within 60 minutes when incubated in 5% w/v sodium citrate because of the Ca2+ removal from the hydrogels. (B) Alginate-PLL (C) Alginate-PLL IPN capsules remained stable in a 5% solution of sodium citrate after 14 hours.

Figure 6.
Swelling Properties. (A) beads stored in refrigerators (pristine), air-dried, and rehydrated (wet). (B) beads stored at -80 °C (pristine), air-dried and rehydrated (wet).
Figure 6.
Swelling Properties. (A) beads stored in refrigerators (pristine), air-dried, and rehydrated (wet). (B) beads stored at -80 °C (pristine), air-dried and rehydrated (wet).
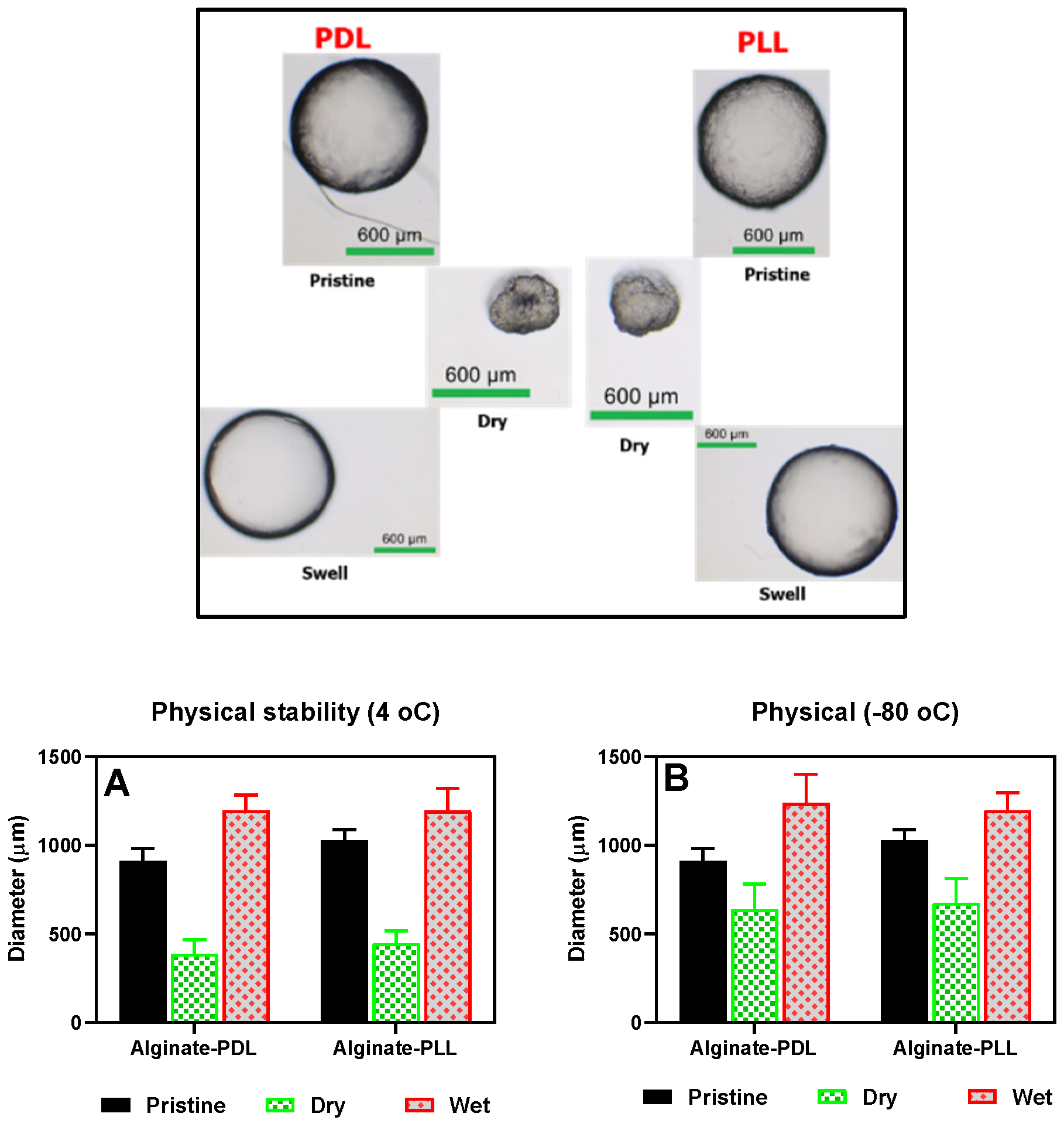
Figure 7.
The bioluminescent response of the immobilized RhlR in different C4-HSL concentrations. The insert illustrates the induction factor calculated using the maximum relative luminescence unit (RLU) derived from the spectral data. The induction factor is the ratio of the test's maximum RLU conducted in the presence of an inducer to the maximum RLU obtained in the absence of added inducers.
Figure 7.
The bioluminescent response of the immobilized RhlR in different C4-HSL concentrations. The insert illustrates the induction factor calculated using the maximum relative luminescence unit (RLU) derived from the spectral data. The induction factor is the ratio of the test's maximum RLU conducted in the presence of an inducer to the maximum RLU obtained in the absence of added inducers.
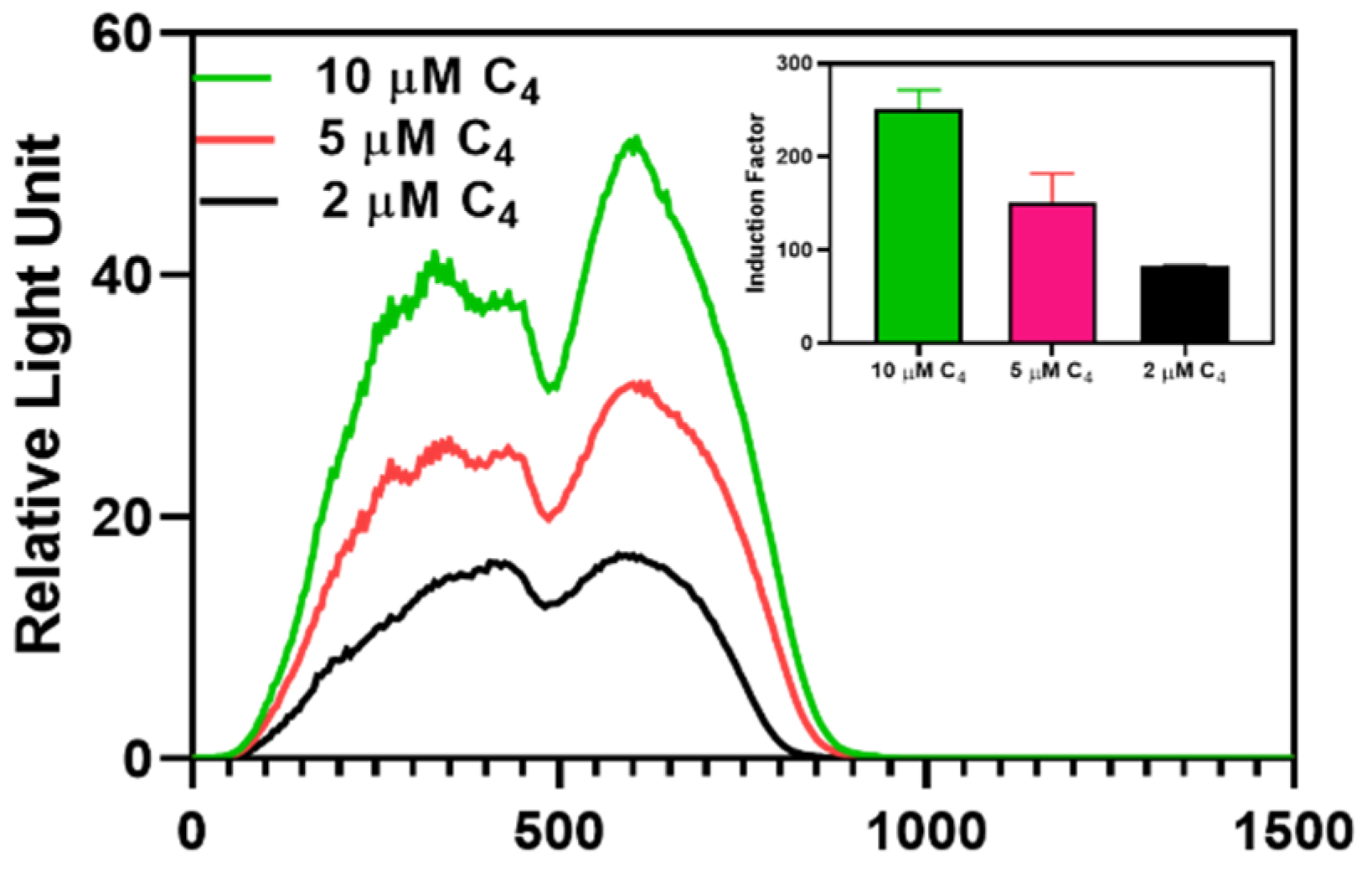
Figure 8.
The bioluminescent Response of PLL and PDL-coated Alginate was Similar. The PDL and PLL coatings were compared regarding the bacteria's response to the added synthetic (C4-HSL) and secreted (by PAO1 wild strain of P. aeruginosa) autoinducers.
Figure 8.
The bioluminescent Response of PLL and PDL-coated Alginate was Similar. The PDL and PLL coatings were compared regarding the bacteria's response to the added synthetic (C4-HSL) and secreted (by PAO1 wild strain of P. aeruginosa) autoinducers.
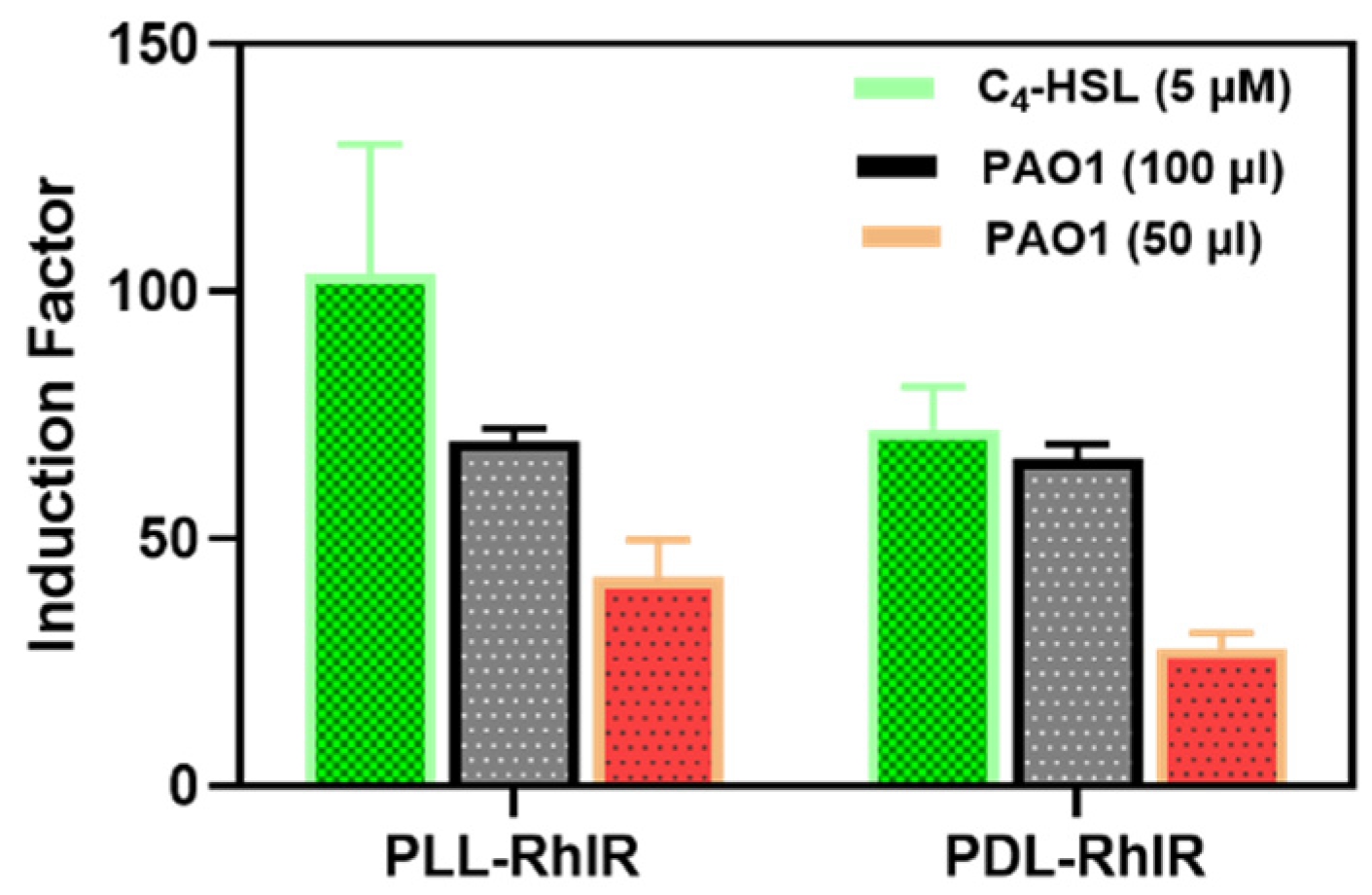
Figure 9.
The effect of pre-incubation and shaking before testing.
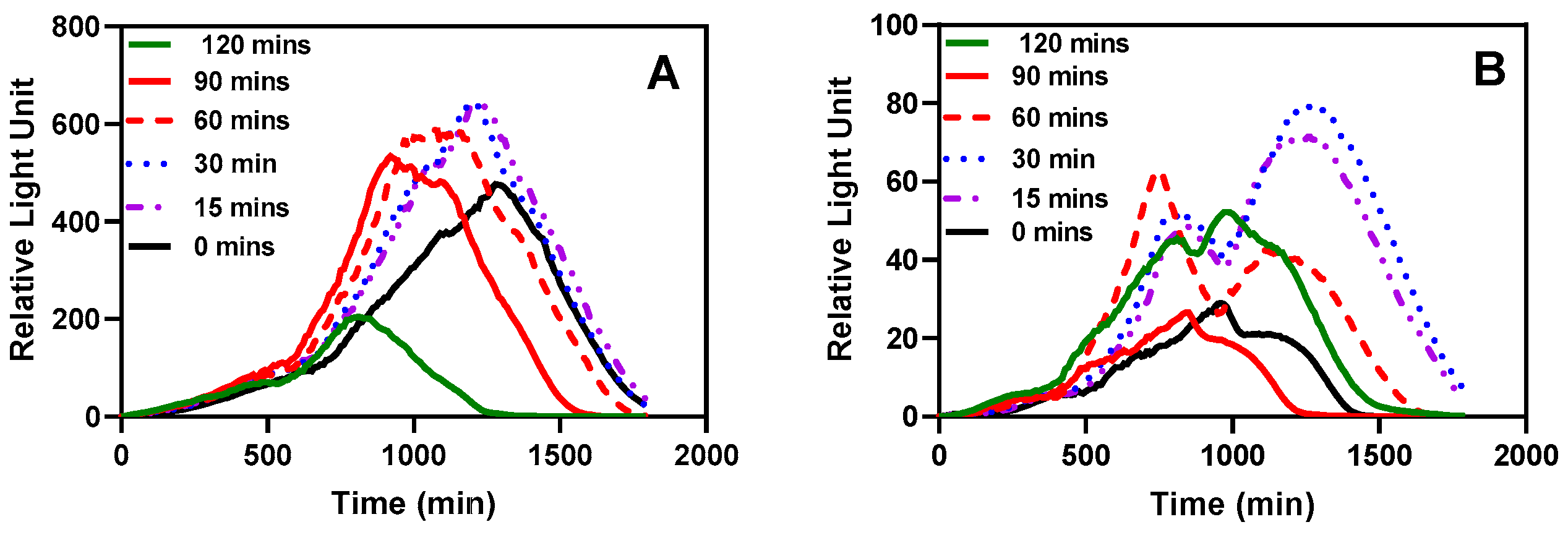
Figure 11.
The bioreporters’ specificity test. RhlR (A) and LasR (B) response towards synthetic QS molecules respectively The second experiment was conducted with either 20 µl of each gram-negative bacterium such as Escherichia coli and Acinetobacter baumannii (72 h biofilm set up) or 5 µM of C4-HSL (C) or 3-oxo-C12-HSL (D). The biosensor performance was tested on other gram-negative bacteria, such as.
Figure 11.
The bioreporters’ specificity test. RhlR (A) and LasR (B) response towards synthetic QS molecules respectively The second experiment was conducted with either 20 µl of each gram-negative bacterium such as Escherichia coli and Acinetobacter baumannii (72 h biofilm set up) or 5 µM of C4-HSL (C) or 3-oxo-C12-HSL (D). The biosensor performance was tested on other gram-negative bacteria, such as.
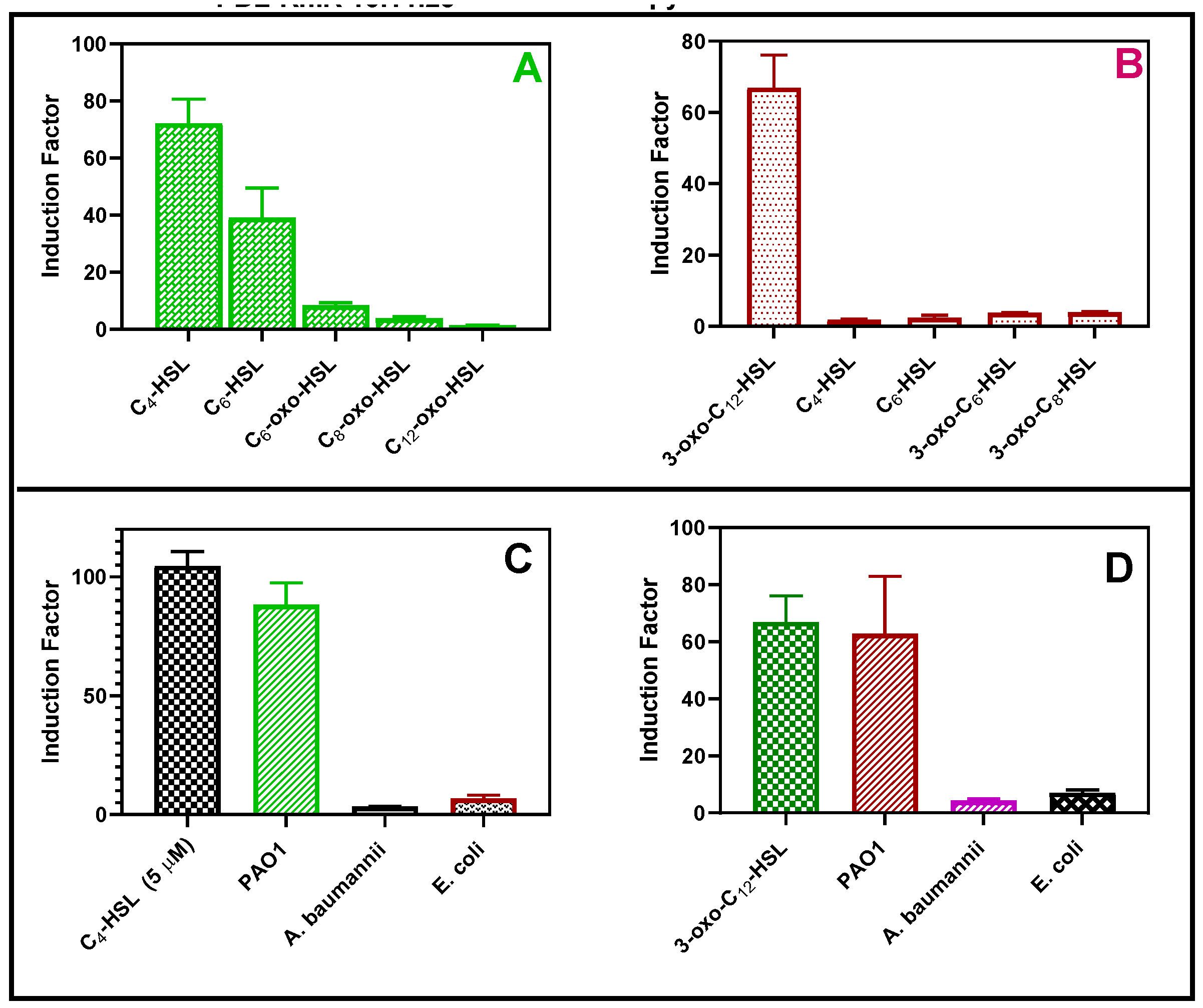
Figure 12.
QS Inhibitory effect of furanone C-30 in the presence of Synthetic AHLs. (A) Inhibition of RhlR signaling in the presence of 5 µM C4-HSL and (B) Inhibition of LasR bioluminescence system in the presence of 5 µM 3-OC12-HSL. Luminescent readings were taken for 10 hours, and the maximum relative light unit (RLU) was plotted. The results are the mean ± SD of biological triplicates.
Figure 12.
QS Inhibitory effect of furanone C-30 in the presence of Synthetic AHLs. (A) Inhibition of RhlR signaling in the presence of 5 µM C4-HSL and (B) Inhibition of LasR bioluminescence system in the presence of 5 µM 3-OC12-HSL. Luminescent readings were taken for 10 hours, and the maximum relative light unit (RLU) was plotted. The results are the mean ± SD of biological triplicates.
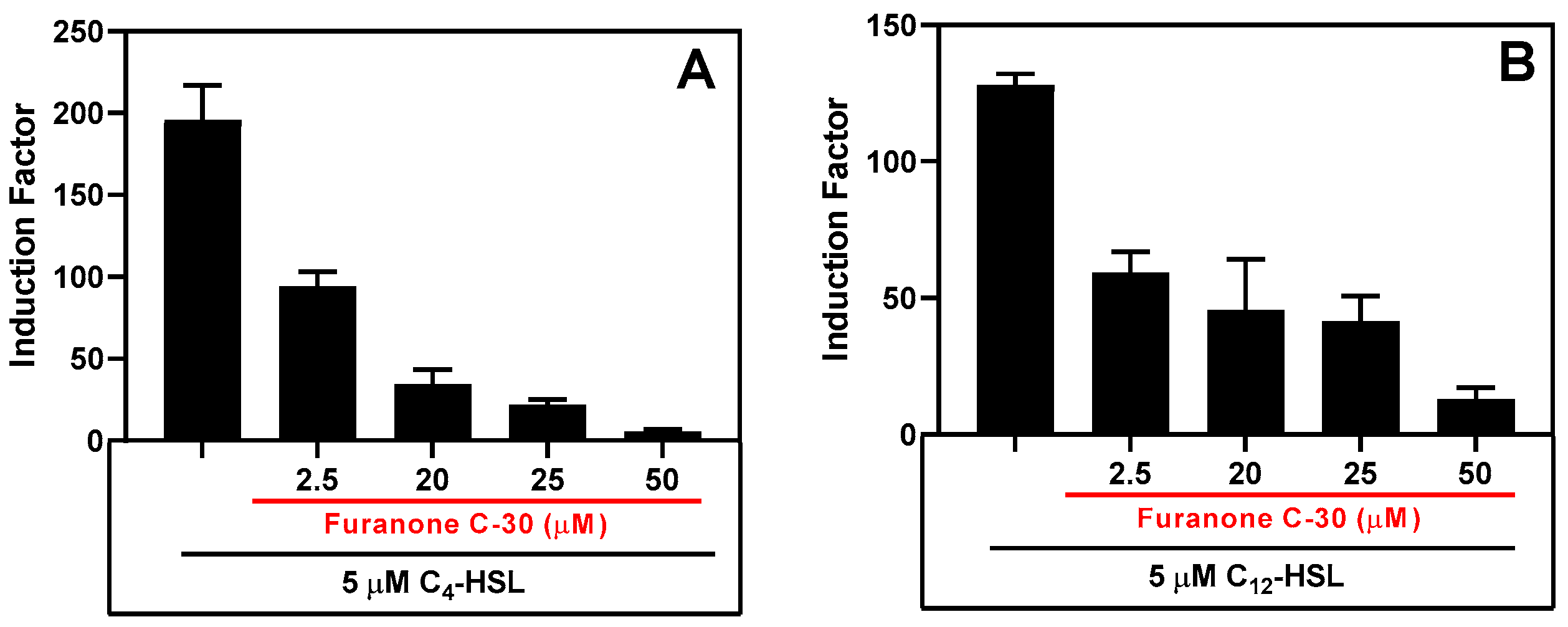
Figure 14.
Storage at room temperature. (A) shows RhlR, and (B) shows the results obtained for the LasR. Result is presented as mean ± SD, (n = 4).
Figure 14.
Storage at room temperature. (A) shows RhlR, and (B) shows the results obtained for the LasR. Result is presented as mean ± SD, (n = 4).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



