
Submitted:
17 June 2024
Posted:
18 June 2024
You are already at the latest version
Abstract
Background: The study was conducted to investigate if the mRNA vaccine technology could be adapted for the ectothermic vertebrate Atlantic salmon (Salmo salar). The lipid nanoparticle (LNP) technology has been developed and optimized for mRNA vaccines in mammals, stabilizing mRNA and facilitating delivery into cells. However, its utility at the temperature and specific biological environment in ectotherms remains unclear. In addition, it is unknown if modified mRNA containing non-canonical nucleotides can correctly translate in salmonid cells. Methods: We used an mRNA transcript coding for enhanced green fluorescence protein, flanked by the untranslated regions of the hemagglutinin-esterase gene of infectious salmon anemia virus, and a 120-base-long poly(A) tail. The mRNA was generated by in vitro transcription where uridine residues were replaced with N1-methyl-pseudouridines, and then encapsulated in LNPs. Results: When transfected into the salmonid cell line CHH-1, the mRNA-LNP construct induced expression of EGFP. Furthermore, when mRNA-LNPs were injected intramuscularly into salmon, in vivo protein expression was demonstrated by immunohistochemistry. EGFP was observed in cells infiltrating the space between the muscle cells in a focal inflammatory response. Conclusion: The results indicate that the N1-methyl-pseudouridine modified mRNA encapsulated in LNPs can be used to express antigens of interest in salmonid fish.
Keywords:
mRNA vaccination
; lipid nanoparticle technology
; Atlantic salmon
1. Introduction
Infectious diseases represent a major problem for the farming of salmonid fish. Attempts of immunization were done already in the 1930s [1], and the first licensed vaccine, targeting enteric redmouth in rainbow trout (Oncorhynchus mykiss), was launched in 1976 [2]. Most commercial vaccines used in salmonid aquaculture today are inactivated and accompanied by an oil-based adjuvant that may cause adverse side effects [3,4,5]. The first commercial DNA vaccine in any farmed animal was against infectious hemorrhagic necrosis in Atlantic salmon (Salmo salar), a milestone that demonstrated that nucleic acids based vaccines can induce protection in salmonid fish [6]. However, DNA vaccines have some limitations, such as low immunogenicity [7] and the theoretical possibility of DNA integration into the host genome lineage [8].
The success of mRNA vaccines against COVID-19 demonstrated their potential in preventing infectious diseases [9,10]. However, mRNA is inherently unstable under natural conditions [11], and exogenous mRNA entering cells may be recognized by a range of cellular pattern recognition receptors, such as membrane-bound Toll-like receptors and cytosolic RIG-I-like receptors [12,13,14,15]. Consequently, stabilization of the mRNA, protection against degradation, and prevention of adverse immune responses are prerequisites for achieving an efficient response of the mRNA vaccines. The discovery that certain chemical modifications of endogenous nucleotides can reduce the innate immune response [16] led to the advancement of using pseudouridine in exogenous mRNA transcripts. This increased mRNA stability, reduced innate immune responses, and enhanced antigen translation [16,17]. Further enhancement of mRNA stability and translational capacity were achieved through poly(A) tail length optimization, which has been found to be an important factor [18]. However, the inefficient transfer of exogenous mRNA into cells, due to the negative charge of mRNA, and the common presence of extracellular RNases, limited the medical use of naked mRNA [19,20].
Although the use of cationic polymers and lipids has been widely and successfully applied for mRNA studies in cell cultures, their positive charge causes systemic toxicity and has prohibited their use live animals. It was not until the development of so-called ionizable lipids that RNA could be safely applied in vivo in synthetic carriers [21,22]. Ionizable lipids allow for efficient complexation with negatively charged RNA and subsequent LNP encapsulation during LNP assembly at pH 4. These lipids are neutral at physiological pH but regain their charge after entering cells and the intracellular acidic endosomal compartment; this re-charging is considered to promote RNA endosomal escape [23]. In addition to making it possible to achieve efficient antigen production after RNA injection in live animals, mRNA-LNPs were also found to have an adjuvant effect [24,25]. Together, these advancements have made it possible to use mRNA for eliciting a strong immune response to the encoded antigen and thus to use it in vaccines [26]. Implementing RNA-based vaccines against viral diseases in aquaculture could be a useful prophylactic tool and possibly resolve some of the shortcomings of other types of vaccines.
Differences between mammalian and fish immune responses could influence the effect of mRNA vaccines in salmonid fish. The foreign mRNA needs to be transported into cells, which is facilitated by LNPs. LNPs were designed to evade detection by mammalian opsonins and innate immune cells. However, it has been suggested that piscine innate immune molecules have greater diversity than those in mammals [27,28,29]. This raises the question of whether the increased receptor diversity in piscine innate immune molecules is more likely to recognize and degrade mRNA-LNPs prior to cellular uptake. This could potentially result in a reduced circulation time of mRNA-LNP molecules, consequently leading to a decreased antigen production.
Even though the mechanisms of cellular uptake of LNPs, which is dependent on the administration route as well as the size and physicochemical properties of the LNPs, is not yet fully understood, the endocytosis pathway is proposed as the predominant route for cellular entry of these particles [30]. Therefore, differences in cell membrane composition between mammals and fish may alter the efficiency of this uptake process. In vitro studies indicated that phospholipid composition and cholesterol contents of different fish cells was similar to mammalian cells, but membrane tension differed [31]. Similarities in lipid composition between mammalian and salmon leukocytes have also been reported [32]. However, it remains unclear whether potential differences in membrane receptors involved in receptor-mediated endocytosis pathways between mammals and fish will affect the ability of piscine cells to internalize mRNA-LNPs.
Studies on the use of LNP-encapsulated mRNA showed differences in in vivo delivery of mRNA and cellular responses to LNPs [33], even between mammalian species. It thus seems likely that differences between mammals and fish exist. The scarce information about mRNA-based vaccines in fish underscores the importance of further research, particularly considering the essential role the fish industry plays in global economy and as a food source.
The aim of this study was to assess whether the mRNA-LNP technology can induce protein expression in Atlantic salmon, and thus could be a viable strategy for developing vaccines against salmonid pathogens. To do so, N1-methyl-pseudouridine modified-mRNAs encoding enhanced green fluorescence protein (EGFP) were synthesized through in vitro transcription (IVT) and subsequently encapsulated in LNPs. In vitro expression was evaluated by transfection of mRNA-LNPs in a salmonid cell line, while in vivo expression was evaluated by intramuscular injection of mRNA-LNPs in Atlantic salmon. The EGFP protein was detected by fluorescence or immunohistochemical staining.
2. Materials and Methods
Design of DNA Plasmids
A plasmid (pVax1 Vector) was designed with an SP6 promoter and the coding sequence of the EGFP gene, flanked by 5’ and 3’ untranslated regions (UTRs) from the hemagglutinin-esterase (HE) gene of infectious salmon anemia virus (ISAV). Additionally, a 3’ poly(A) of 120 residues was added to the plasmid construct. MluI restriction sites were inserted upstream and downstream of the 5’ UTR and the poly(A) region, respectively. The plasmid encoded a 988 nucleotides (nt) long mRNA construct and was synthesized by GenScript (Piscataway, NJ, USA).
In Vitro Transcription of mRNA
Plasmids were linearized by the MluI restriction enzyme and purified with the NucleoSpin Gel and PCR Clean-up kit (Macherey-Nagel, Düren, Germany). The IVT was carried out using the RiboMAX™ Large Scale RNA Production System – SP6 (Promega, Madison, WI, USA), following the manufacturer’s recommendations. Linearized DNA encoding Firefly luciferase (Fluc) that was included in the kit, was used as a control with an expected mRNA size of 1800 nt. Uridine-5′-triphosphates were replaced with N1-methyl-pseudouridine-5’’-triphosphates (TriLink Biotechnologies, San Diego, CA, USA). The resulting IVT mRNA was purified using the RNeasy mini kit (QIAGEN, Hilden, Germany). Enzymatic capping of the mRNAs was performed post-transcriptionally with the ScriptCap™ Cap 1 Capping System (Cellscript Inc, Madison, WI, USA), and the nucleoside-modified, capped mRNAs were purified using the RNeasy mini kit (QIAGEN) (Figure 1). Finally, RNA quality and integrity were analyzed by capillary gel electrophoresis with the use of a Tapestation 4200 instrument (Agilent, Santa Clara, CA, USA) and an RNA ScreenTape ladder (Agilent). The resulting mRNA was stored at -80 °C until use.
mRNA-LNP Formulation
mRNA-LNPs were formulated using a flow-mixing based approach. Stock solutions of ionizable lipid SM-102, cholesterol, DSPC, and PEG2000-DMG were prepared in EtOH. Lipids were mixed at a molar ratio (%) of 50:38.5:10:1.5 SM-102:cholesterol:DSPC:PEG2000-DMG and the EtOH volume was adjusted to obtain a 6.18 mg/mL (10 mM) total lipid concentration. The mRNA was dissolved in RNase-free 25 mM acetate buffer (pH 4.0) at 39 μg/mL. The lipid mixture in EtOH and the mRNA solution in acetate buffer were placed in syringes on computer operated syringe pumps and mixed in a T-junction mixer at a total flow rate of 12 ml/min and an EtOH:buffer flow rate ration of 1:3 resulting in a 1:30 mRNA:ionizable lipid weight ratio. After formulation, mRNA-LNPs were dialyzed using 10 kDa molecular weight cut-off dialysis cassette (Slide-A-Lyzer™ G2 Dialysis Cassettes, Thermo Scientific™) overnight at room temperature against Tris buffer (10 mM, pH 7.4). After dialysis, mRNA-LNPs solution were adjusted at 10% sucrose, the mRNA concentration was measured (RiboGreen assay, see below) and the mRNA-LNPs were diluted to an mRNA concentration of 10 ug/ml using Tris buffer (10 mM, pH7.4) with 10 weight% sucrose. mRNA-LNPs were stored at -20⁰C in aliquots. On the day of experiments, corresponding aliquots were thawed and used the same day.
mRNA-LNP Characterization
Dynamic light scattering (DLS, Cordouan - VascoKIN) was used to measure mRNA-LNP size (Z-avg) and size distribution (polydispersity index, PDI) after a 40x mRNA-LNP dilution in Tris buffer (10 mM, pH 7.4). Multi-detector field flow fractionation (MD-FFF, AF2000 Multiflow FFF (Postnova Analytics, Germany)) was equipped with a multi-angle light scattering detector (MALS, PN3621) and an UV–Vis absorbance detector (PN3211). MALS was used to assess mRNA-LNP size (radius of gyration, Rg) and size distribution and recovery was determined using the UV absorbance obtained after a direct injection (a measure for total mass present in the sample) and the UV-absorbance from the fraction containing the mRNA-LNPs only. The system was operated according to published methodology [34].
To assess mRNA encapsulation and concentration we used the Quant-it RiboGreen assay (Invitrogen, R11490). Calibration curves (at mRNA concentrations of 0-640 ng/ml) were prepared using the encapsulated mRNA. mRNA-LNPs were assayed at multiple dilutions and both in the presence of 0.5% Triton X (to measure total mRNA concentration, as Triton X disrupts mRNA-LNPs) and in the absence of Triton X (to measure unencapsulated mRNA concentration). Finally, mRNA encapsulation efficiency (EE%) was calculated.
In Vitro Expression
CHH-1 cells, originally established from heart tissue from chum salmon (Oncorhynchus keta) [35], were cultured in Leibovitz’‘s’ L-15 medium (ThermoFisherScientific, Waltham, MA, USA), supplemented with 10% fetal bovine serum and 50 mg/ml gentamycin at 20 °C. For experiments, cells were seeded in 12-well plates, and when 90% cell confluency was reached, 500 ng LNP-encapsulated mRNA encoding EGFP was transfected per well, corresponding to 140 ng of encapsulated mRNA/cm2. Expression of EGFP was monitored during a period of 11 days, using an EVOS Microscope M5000 Imaging System fluorescence microscope (ThermoFisherScientific). Negative controls were either cells transfected with mRNA-LNPs encoding Fluc or cells included with 10 mM TRIS-HCl with 10% sucrose. Images were prepared for publication using brightness and contrast adjustments with ImageJ, with the same settings applied to all images.
In Vitro Cytotoxicity of mRNA-LNPs and mRNA-Lipofectamine
CHH-1 cells were seeded in 24-well plates, approximately 50,000 cells per well. At 24 hours after cell seeding, cells were transfected with 285 or 570 ng EGFP-encoding mRNA encapsulated in LNPs, corresponding to 150 or 300 ng encapsulated mRNA/cm2, respectively. Cytotoxicity was monitored using the CellTiter 96® AQueous One Solution Cell Proliferation Assay (MTS, Promega) at 1, 3 and 5 days after transfection. The medium was removed, washed with PBS, and replaced with fresh medium before the CellTiter 96® AQueous One Solution was added, and incubated for 4 hours at 20 °C. Finally, the amount of formazan, which is produced when metabolically active cells reduce the MTS tetrazolium salt, was recorded at 490 nm with a reference wavelength of 650 nm with a TECAN SPARK spectrophotometer (TECAN, Männedorf, Switzerland).
Injection of mRNA-LNPs in Salmon
The in vivo study was done at VESO Aqualab research facility (Vikan, Norway). Experiments had been approved by the Norwegian Food Safety Authority according to the European Union Directive 2010/63/EU. The fish were unvaccinated Atlantic salmon with an average body weight of 40 g. Fish were seronegative for antibodies against common disease-causing bacteria and tested negative by qPCR for infectious pancreatic necrosis virus, ISAV, salmon pancreas disease virus, piscine orthoreovirus and piscine myocarditis virus before initiation of the study. The fish were kept under 12:12 L:D light regime in 12 °C fresh water. The fish were anesthetized by immersion in benzocaine chloride (5 ± 2 min, 0.5 g / 10 L water) prior to handling, and euthanized using a lethal dose of benzocaine chloride (1 g / 5 L water). The fish were observed at least once per day. No fish died during the trial. Four salmons were injected intramuscularly into white muscle tissue at a fixed injection site near the dorsal fin with 100 µl of the mRNA-LNP solution containing 1 µg of EGFP-encoding mRNA. Two negative controls were used, one injected with 1 µg of Fluc-encoding mRNA-LNPs, and one that did not receive any injection. Muscle tissue samples from the injection site were collected 7 days post injection (dpi), fixed in formalin for 24 hours, and stored in ethanol at 4 °C until paraffin embedding.
Immunohistochemistry
Immunohistochemistry was performed with MACH 1 Universal HRP-Polymer Detection kit (Biocare Medical, Pacheco, CA, USA). Tissue from mouse expressing EGFP was included for method validation and was kindly gifted by Guttorm Haraldsen (University of Oslo and Oslo University Hospital, Norway). Sections were dewaxed and subjected to heat-mediated antigen retrieval in DIVA solution at 100 °C for 10 min. Endogenous peroxidase was blocked by adding Peroxidazed 1 and incubated for 10 min at room temperature (RT). Furthermore, blocking of unspecific binding was performed by incubation of the sections with Background Sniper for 10 min at RT. Anti-GFP antibody (A-6455, Invitrogen, ThermoFisherScientific) was diluted 1:1000 in Da Vinci Diluent, and the tissue sections were incubated with the antibody at RT for 1 hour in a humidified chamber. The tissue sections were then incubated with MACH 1 Universal HRP-Polymer for 30 min at RT, and 3, 3’’-diaminobenzidine (DAB) was used to visualize the immunoreaction. Tissue sections were counterstained by hematoxylin before dehydration in 100% ethanol, clearing in xylene, and mounting. A Nikon Eclipse Ni-U microscope (Nikon, Tokyo, Japan) was used to visualize the muscle tissue samples.
3. Results
Production of mRNA and LNP Encapsulation:
mRNAs were in vitro transcribed from plasmids encoding EGFP or Fluc, and size and integrity was evaluated by capillary gel electrophoresis. We confirmed the size of the EGFP construct to be about 1000 nt and the Fluc mRNA to be 1800 nt (Figure 2A). The constructs were encapsulated in LNPs with a lipid composition very similar to the Moderna COVID-19 vaccine. The obtained mRNA-LNPs had a z-average diameter below 90 nm and a PDI below 0.2 as measured by DLS, indicating successful formulation. MD-FFF analysis confirmed a high quality of the formulations. The sharp single peaks in the elution profiles (fractograms, Figure 2B and 2C) demonstrated a well-defined, single population of mRNA-LNPs. The high recovery (> 96%) indicated that nearly all material in the formulation was present as mRNA-LNPs. Similarly, the high encapsulation efficiency (>94%) as measured with the RiboGreen assay demonstrated efficient mRNA encapsulation. See Figure 2D for a summary of mRNA-LNP characteristics.
In Vitro Expression of mRNA-LNPs in CHH-1 Cells
CHH-1 cells were transfected with mRNA-LNPs encoding EGFP to evaluate the capability of LNPs to transport in vitro transcribed mRNAs and to confirm that the N1-methyl-pseuoduridine modified mRNA induced protein expression in salmonid cells. The CHH-1 cells were monitored over 11 days post transfection (dpt) and revealed protein expression already at 1 dpt and 3 dpt, although with a weak EGFP intensity. Expression of EGFP peaked between 5 dpt and 7 dpt, and fluorescence microscopy indicated that most cells expressed EGFP. The production of EGFP started to decline at 9 dpt, which continued until 11 dpt (Figure 3).
Evaluation of mRNA-LNP Cytotoxicity
An MTS assay was used to assess the cytotoxic effects when transfecting 285 or 570 ng of EGFP-encoding mRNA-LNPs in CHH-1 cells. The assay revealed 73% and 82% viability at 1 dpt in cells treated with mRNA-LNPs containing 285 ng or 570 ng mRNA, respectively, when compared to non-transfected control cells. The viability decreased at both mRNA-LNP concentrations at 3 dpt, with cells transfected with 285 ng mRNA-LNPs showing 64% viability compared to control cells, and cells transfected with 570 ng mRNA-LNPs showing 60% viability. A further decrease in viability in mRNA-LNP transfected cells was observed at 5 dpt, where cells transfected with 285 ng or 570 ng of mRNA-LNPs showed 54% or 45% viability compared to control cells, respectively (Figure 4).
In Vivo Protein Expression
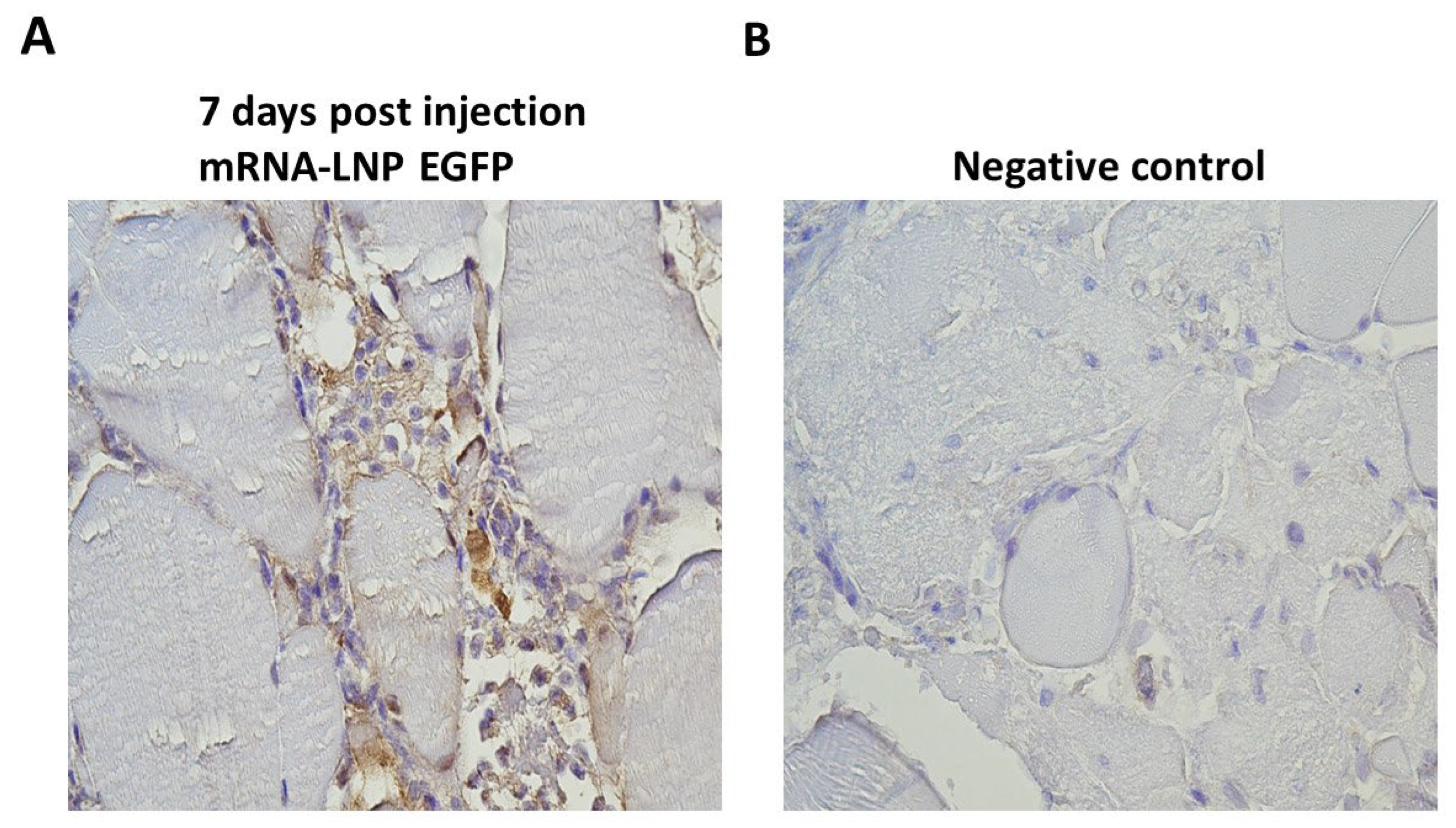
100 µl of mRNA-LNP suspensions containing 1 µg of mRNA encoding EGFP or Fluc were injected intramuscularly into salmon, and muscle tissue samples were collected at 7 dpi, based on the peak of protein expression in vitro. Immunohistochemistry was performed to assess EGFP expression. The muscle tissues collected from the site of injection exhibited signal after staining with anti-GFP antibodies, confirming the presence of EGFP protein (Figure 5A). The signal was observed in cells infiltrating the space between the muscle cells. A focal immuno-positive inflammatory response was found in the white skeletal musculature. Some leukocytes exhibited distinct EGFP signal, while others showed no signal. The myocytes appeared intact and showed no signal for EGFP. Taken together, this demonstrated the LNP technology’s capability to transport mRNAs into salmon cells effectively in vivo.
4. Discussion
To our knowledge, no studies have investigated the use of mRNAs delivered by LNPs as a tool for generating antigens in salmonid fish. The high need for improved vaccines in the salmon farming industry combined with the demonstrated and increasing potential of mRNA-LNP vaccines motivated us to assess whether this technology can be translated to salmon. Our study aimed to determine the effectiveness of LNPs in transfecting mRNA into salmon cells and induce expression of corresponding proteins. Our findings indicate that the technology may be adopted in vaccine strategies against piscine pathogens. While previous studies have shown the ability of mRNA-LNP technology to successfully express proteins in zebrafish [36], important biological and environmental disparities exist between zebrafish and salmonids. For example, these species have adapted to very different temperatures, approximately 25-30 °C for zebrafish and 5-15 °C for salmonids, and variations in temperature might influence the functionality of LNPs [37].
In the present study, mRNAs were synthesized by IVT from a DNA plasmid containing UTRs from the ISAV HE glycoprotein sequence and the coding sequence for EGFP. The DNA plasmid also included a 120-base-long poly(A) tail, which ensures the reproducibility of IVT mRNA with a fixed poly(A) length, as opposed to enzymatic post-transcriptional addition of the poly(A) tail [18]. Additionally, the mRNA transcript was modified with N1-methyl-pseudouridine that enhance protein translation and mRNA stability [16], suppress innate immune responses [38,39,40] and reduce the formation of double-stranded RNA contaminants, which is a powerful inducer of innate immune response [41]. LNPs were engineered to enhance the in vivo transfection efficiency of mRNA, increasing subsequent protein expression. Different modifications have been implemented to refine LNP characteristics, including the utilization of ionizable cationic lipids [42]. Herein, we employed LNPs with ionizable cationic lipids to evaluate their ability in delivering mRNAs encoding EGFP into salmonid cells.
First, to determine the ability of LNP to effectively transport mRNA into salmonid cells, the mRNA-LNP encoding EGFP was transfected into CHH-1 cells. EGFP expression was observed already at 1 dpt, and the intensity of EGFP fluorescence activity steadily increased until 7 dpt. Notably, EGFP activity persisted even after 11 dpt, indicating the sustained protein expression achieved through mRNA-LNP transfection. The formulation of the LNPs we used were originally developed for use in mammals; thus, their functionality has been tailored to this specific biological environment, maintained at 37 °C. Therefore, it is important to establish that the mRNA-LNPs are internalized in salmonid cells and can induce protein translation from the encapsulated mRNA. In addition, the N1-pseudouridine-modification raised some questions regarding functionality in salmonid cells. The presence of pseudouridine synthase has been detected in Chinook salmon (Oncorhynchus tshawytscha) [43], which indicates the natural appearance of pseudouridine in RNAs in salmonid fish. The production of functional EGFP in CHH-1 cells showed that N1-methylated pseudouridine-modified mRNA can instruct for correct translation of functional proteins in salmonid cells.
LNPs have previously been shown to possess adjuvant activity, which is important for achieving a robust adaptive immune response to vaccination [44].
The viability assay revealed a decrease in cellular metabolic activity in cells transfected with mRNA-LNPs as early as 1 dpt, compared to non-transfected control cells, and it was more pronounced at 3 and 5 dpt. The decreased cellular metabolic activity indicated that the mRNA-LNPs have potential to cause cytotoxicity in salmon cells, at least in vitro. This was also observed microscopically, where a significant decrease in cell density occurred after washing. This indicated decreased cell viability and ability to stick to the well surface compared to the control cells where the density remained the same after washing. It should be kept in mind that the cytotoxic effect of the mRNA-LNPs might differ between different cell lines [45].
To assess the in vivo performance of the mRNA-LNPs, salmon were injected intramuscularly with mRNA-LNP encoding EGFP, and muscle tissue samples were harvested at 7 dpi, which is within the time frame of high in vitro protein expression in CHH-1 cells. The muscle tissue samples were subjected to IHC analysis, which visualized the presence of EGFP. Importantly, for the in vivo analysis, salmon were kept at their natural temperature, ensuring that the experiment was conducted in a biologically relevant environment. The presence of EGFP indicated that LNPs can be taken up by cells in salmon muscle tissue and induce correct translation of the encapsulated mRNA. EGFP expression was mostly observed in infiltrating phagocytes in the space between the muscle cells, while there was no expression in the muscle fiber that could be visually observed. This is in accordance with localization of protein expression after intramuscular administration of mRNA-LNPs in rodents [46]. The accumulation of phagocytes indicates that the LNPs are recognized as foreign material and induces an inflammatory response. The phagocytic cells recognize and internalize foreign material such as the administered mRNA-LNP and demonstrates the functionality of LNPs beyond their original application in mammals, suggesting their potential as efficient delivery vehicles for mRNA in salmonid fish.
It should be noted that the current work was performed with an LNP composition developed for use in humans. An optimalization of the delivery system to the physiology and immunology of salmon, and concomitant quantitative comparison of the efficacy of different LNPs, could be interesting for further development.
5. Conclusions
This study demonstrates the successful use of LNPs for mRNA delivery in salmonids, ensuring protein expression in salmonid cell cultures in vitro and in salmonid fish in vivo. The production of EGFP in response to mRNA-LNP injection in salmon suggests that this vaccine technology has promise for immunizing antigen production to counteract pathogens infecting salmonids and potentially other fish species.
Author Contributions
ER launched the project idea and participated in the overall design and coordination of the study and interpretation of data. LOSD and ER drafted the manuscript. SB and LOSD performed sample preparation, RT-qPCR and mRNA production. SH, AM. FR and JP performed the LNP production. HB performed morphological analyses of the in vivo expression. SH, SEB, JHF and ØW participated in the coordination of the study, interpretation of data, and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The Norwegian Seafood Research Fund (FHF), Grant #901746, supported this study financially.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Snieszko, S.F. A bacterial disease of carp in central Europe. The Progressive Fish-Culturist 1940, 7, 12–15. [Google Scholar] [CrossRef]
- Gudding, R.; Van Muiswinkel, W.B. A history of fish vaccination: Science-based disease prevention in aquaculture. Fish Shellfish Immunol 2013, 35, 1683–1688. [Google Scholar] [CrossRef]
- Brudeseth, B.E.; Wiulsrød, R.; Fredriksen, B.N.; Lindmo, K.; Løkling, K.E.; Bordevik, M.; Steine, N.; Klevan, A.; Gravningen, K. Status and future perspectives of vaccines for industrialised fin-fish farming. Fish Shellfish Immunol 2013, 35, 1759–1768. [Google Scholar] [CrossRef]
- Midtlyng, P.J.; Reitan, L.J.; Lillehaug, A.; Ramstad, A. Protection, immune responses and side effects in atlantic salmon (salmo salarl.) vaccinated against furunculosis by different procedures. Fish & Shellfish Immunology 1996, 6, 599–613. [Google Scholar]
- Ma, J.; Bruce, T.J.; Jones, E.M.; Cain, K.D. A review of fish vaccine development strategies: Conventional methods and modern biotechnological approaches. Microorganisms 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Hølvold, L.B.; Myhr, A.I.; Dalmo, R.A. Strategies and hurdles using DNA vaccines to fish. Veterinary Research 2014, 45, 21. [Google Scholar] [CrossRef] [PubMed]
- Lu, S. Immunogenicity of DNA vaccines in humans: It takes two to tango. Hum Vaccin 2008, 4, 449–452. [Google Scholar] [CrossRef]
- Khan, F.H. The elements of immunology. Pearson Education India: 2009.
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B. , et al. Efficacy and safety of the mrna-1273 sars-cov-2 vaccine. N Engl J Med 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C. , et al. Safety and efficacy of the bnt162b2 mrna covid-19 vaccine. N Engl J Med 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Houseley, J.; Tollervey, D. The many pathways of rna degradation. Cell 2009, 136, 763–776. [Google Scholar] [CrossRef]
- Scheel, B.; Braedel, S.; Probst, J.; Carralot, J.P.; Wagner, H.; Schild, H.; Jung, G.; Rammensee, H.G.; Pascolo, S. Immunostimulating capacities of stabilized rna molecules. Eur J Immunol 2004, 34, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Karikó, K.; Ni, H.; Capodici, J.; Lamphier, M.; Weissman, D. Mrna is an endogenous ligand for toll-like receptor 3. J Biol Chem 2004, 279, 12542–12550. [Google Scholar] [CrossRef] [PubMed]
- Heil, F.; Hemmi, H.; Hochrein, H.; Ampenberger, F.; Kirschning, C.; Akira, S.; Lipford, G.; Wagner, H.; Bauer, S. Species-specific recognition of single-stranded rna via toll-like receptor 7 and 8. Science 2004, 303, 1526–1529. [Google Scholar] [CrossRef] [PubMed]
- Pichlmair, A.; Schulz, O.; Tan, C.P.; Näslund, T.I.; Liljeström, P.; Weber, F.; Reis e Sousa, C. Rig-i-mediated antiviral responses to single-stranded rna bearing 5’’-phosphates. Science 2006, 314, 997–1001. [Google Scholar] [CrossRef] [PubMed]
- Karikó, K.; Muramatsu, H.; Welsh, F.A.; Ludwig, J.; Kato, H.; Akira, S.; Weissman, D. Incorporation of pseudouridine into mrna yields superior nonimmunogenic vector with increased translational capacity and biological stability. Mol Ther 2008, 16, 1833–1840. [Google Scholar] [CrossRef]
- Karikó, K.; Buckstein, M.; Ni, H.; Weissman, D. Suppression of rna recognition by toll-like receptors: The impact of nucleoside modification and the evolutionary origin of rna. Immunity 2005, 23, 165–175. [Google Scholar] [CrossRef]
- Holtkamp, S.; Kreiter, S.; Selmi, A.; Simon, P.; Koslowski, M.; Huber, C.; Türeci, O.; Sahin, U. Modification of antigen-encoding rna increases stability, translational efficacy, and t-cell stimulatory capacity of dendritic cells. Blood 2006, 108, 4009–4017. [Google Scholar] [CrossRef]
- Wadhwa, A.; Aljabbari, A.; Lokras, A.; Foged, C.; Thakur, A. Opportunities and challenges in the delivery of mrna-based vaccines. Pharmaceutics 2020, 12. [Google Scholar] [CrossRef]
- Dowdy, S.F. Overcoming cellular barriers for rna therapeutics. Nat Biotechnol 2017, 35, 222–229. [Google Scholar] [CrossRef]
- Kulkarni, J.A.; Witzigmann, D.; Chen, S.; Cullis, P.R.; van der Meel, R. Lipid nanoparticle technology for clinical translation of sirna therapeutics. Acc Chem Res 2019, 52, 2435–2444. [Google Scholar] [CrossRef]
- Jeeva, S.; Kim, K.H.; Shin, C.H.; Wang, B.Z.; Kang, S.M. An update on mrna-based viral vaccines. Vaccines (Basel) 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Schlich, M.; Palomba, R.; Costabile, G.; Mizrahy, S.; Pannuzzo, M.; Peer, D.; Decuzzi, P. Cytosolic delivery of nucleic acids: The case of ionizable lipid nanoparticles. Bioeng Transl Med 2021, 6, e10213. [Google Scholar] [CrossRef] [PubMed]
- Fenton, O.S.; Kauffman, K.J.; McClellan, R.L.; Appel, E.A.; Dorkin, J.R.; Tibbitt, M.W.; Heartlein, M.W.; DeRosa, F.; Langer, R.; Anderson, D.G. Bioinspired alkenyl amino alcohol ionizable lipid materials for highly potent in vivo mrna delivery. Adv Mater 2016, 28, 2939–2943. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Zaks, T.; Langer, R.; Dong, Y. Lipid nanoparticles for mrna delivery. Nat Rev Mater 2021, 6, 1078–1094. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.J.; Naradikian, M.S.; Parkhouse, K.; Cain, D.W.; Jones, L.; Moody, M.A.; Verkerke, H.P.; Myles, A.; Willis, E. , et al. Nucleoside-modified mrna vaccines induce potent t follicular helper and germinal center b cell responses. J Exp Med 2018, 215, 1571–1588. [Google Scholar] [CrossRef] [PubMed]
- Vasta, G.R.; Nita-Lazar, M.; Giomarelli, B.; Ahmed, H.; Du, S.; Cammarata, M.; Parrinello, N.; Bianchet, M.A.; Amzel, L.M. Structural and functional diversity of the lectin repertoire in teleost fish: Relevance to innate and adaptive immunity. Dev Comp Immunol 2011, 35, 1388–1399. [Google Scholar] [CrossRef] [PubMed]
- Yoder, J.A.; Litman, G.W. The phylogenetic origins of natural killer receptors and recognition: Relationships, possibilities, and realities. Immunogenetics 2011, 63, 123–141. [Google Scholar] [CrossRef] [PubMed]
- Sunyer, J.O.; Zarkadis, I.K.; Lambris, J.D. Complement diversity: A mechanism for generating immune diversity? Immunol Today 1998, 19, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Sousa de Almeida, M.; Susnik, E.; Drasler, B.; Taladriz-Blanco, P.; Petri-Fink, A.; Rothen-Rutishauser, B. Understanding nanoparticle endocytosis to improve targeting strategies in nanomedicine. Chem Soc Rev 2021, 50, 5397–5434. [Google Scholar] [CrossRef]
- Shiomi, A.; Nagao, K.; Kasai, H.; Hara, Y.; Umeda, M. Changes in the physicochemical properties of fish cell membranes during cellular senescence. Bioscience, Biotechnology, and Biochemistry 2020, 84, 583–593. [Google Scholar] [CrossRef]
- Thompson, K.D.; Henderson, R.J.; Tatner, M.F. A comparison of the lipid composition of peripheral blood cells and head kidney leucocytes of atlantic salmon (salmo salar l.). Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology 1995, 112, 83–92. [Google Scholar] [CrossRef]
- Hatit, M.Z.C.; Lokugamage, M.P.; Dobrowolski, C.N.; Paunovska, K.; Ni, H.; Zhao, K.; Vanover, D.; Beyersdorf, J.; Peck, H.E.; Loughrey, D. , et al. Species-dependent in vivo mrna delivery and cellular responses to nanoparticles. Nature Nanotechnology 2022, 17, 310–318. [Google Scholar] [CrossRef]
- Parot, J.; Mehn, D.; Jankevics, H.; Markova, N.; Carboni, M.; Olaisen, C.; Hoel, A.D.; Sigfúsdóttir, M.S.; Meier, F.; Drexel, R. , et al. Quality assessment of lnp-rna therapeutics with orthogonal analytical techniques. Journal of Controlled Release 2024, 367, 385–401. [Google Scholar] [CrossRef]
- Lannan, C.N.; Winton, J.R.; Fryer, J.L. Fish cell lines: Establishment and characterization of nine cell lines from salmonids. In Vitro 1984, 20, 671–676. [Google Scholar] [CrossRef]
- Linares-Fernández, S.; Moreno, J.; Lambert, E.; Mercier-Gouy, P.; Vachez, L.; Verrier, B.; Exposito, J.Y. Combining an optimized mrna template with a double purification process allows strong expression of in vitro transcribed mrna. Mol Ther Nucleic Acids 2021, 26, 945–956. [Google Scholar] [CrossRef]
- Yu, M.; Song, W.; Tian, F.; Dai, Z.; Zhu, Q.; Ahmad, E.; Guo, S.; Zhu, C.; Zhong, H.; Yuan, Y. , et al. Temperature- and rigidity-mediated rapid transport of lipid nanovesicles in hydrogels. Proc Natl Acad Sci U S A 2019, 116, 5362–5369. [Google Scholar] [CrossRef]
- Anderson, B.R.; Muramatsu, H.; Jha, B.K.; Silverman, R.H.; Weissman, D.; Karikó, K. Nucleoside modifications in rna limit activation of 2’-5’-oligoadenylate synthetase and increase resistance to cleavage by rnase l. Nucleic Acids Res 2011, 39, 9329–9338. [Google Scholar] [CrossRef]
- Andries, O.; Mc Cafferty, S.; De Smedt, S.C.; Weiss, R.; Sanders, N.N.; Kitada, T. N1-methylpseudouridine-incorporated mrna outperforms pseudouridine-incorporated mrna by providing enhanced protein expression and reduced immunogenicity in mammalian cell lines and mice. Journal of Controlled Release 2015, 217, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Svitkin, Y.V.; Cheng, Y.M.; Chakraborty, T.; Presnyak, V.; John, M.; Sonenberg, N. N1-methyl-pseudouridine in mrna enhances translation through eif2α-dependent and independent mechanisms by increasing ribosome density. Nucleic Acids Res 2017, 45, 6023–6036. [Google Scholar] [CrossRef] [PubMed]
- Baiersdörfer, M.; Boros, G.; Muramatsu, H.; Mahiny, A.; Vlatkovic, I.; Sahin, U.; Karikó, K. A facile method for the removal of dsrna contaminant from in vitro-transcribed mrna. Mol Ther Nucleic Acids 2019, 15, 26–35. [Google Scholar] [CrossRef]
- Chen, J.; Ye, Z.; Huang, C.; Qiu, M.; Song, D.; Li, Y.; Xu, Q. Lipid nanoparticle-mediated lymph node-targeting delivery of mrna cancer vaccine elicits robust cd8(+) t cell response. Proc Natl Acad Sci U S A 2022, 119, e2207841119. [Google Scholar] [CrossRef] [PubMed]
- Bowen, L.; von Biela, V.R.; McCormick, S.D.; Regish, A.M.; Waters, S.C.; Durbin-Johnson, B.; Britton, M.; Settles, M.L.; Donnelly, D.S.; Laske, S.M. , et al. Transcriptomic response to elevated water temperatures in adult migrating yukon river chinook salmon (oncorhynchus tshawytscha). Conserv Physiol 2020, 8, coaa084. [Google Scholar] [CrossRef] [PubMed]
- Alameh, M.G.; Tombácz, I.; Bettini, E.; Lederer, K.; Sittplangkoon, C.; Wilmore, J.R.; Gaudette, B.T.; Soliman, O.Y.; Pine, M.; Hicks, P. , et al. Lipid nanoparticles enhance the efficacy of mrna and protein subunit vaccines by inducing robust t follicular helper cell and humoral responses. Immunity 2021, 54, 2877–2892.e2877. [Google Scholar] [CrossRef] [PubMed]
- Naderi Sohi, A.; Kiani, J.; Arefian, E.; Khosrojerdi, A.; Fekrirad, Z.; Ghaemi, S.; Zim, M.K.; Jalili, A.; Bostanshirin, N.; Soleimani, M. Development of an mrna-lnp vaccine against sars-cov-2: Evaluation of immune response in mouse and rhesus macaque. Vaccines (Basel) 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Hassett, K.J.; Rajlic, I.L.; Bahl, K.; White, R.; Cowens, K.; Jacquinet, E.; Burke, K.E. Mrna vaccine trafficking and resulting protein expression after intramuscular administration. Mol Ther Nucleic Acids 2024, 35, 102083. [Google Scholar] [CrossRef]
Figure 1.
Overview of mRNA construct. The mRNA was generated from a plasmid encoding EGFP with untranslated regions from the hemagglutinin-esterase gene and a 120 residues poly(A) tail. In addition, uridine residues were substituted with N1-methyl-pseudouridines, and a 5’ cap was enzymatically added post-transcriptionally. Created with BioRender.com.
Figure 1.
Overview of mRNA construct. The mRNA was generated from a plasmid encoding EGFP with untranslated regions from the hemagglutinin-esterase gene and a 120 residues poly(A) tail. In addition, uridine residues were substituted with N1-methyl-pseudouridines, and a 5’ cap was enzymatically added post-transcriptionally. Created with BioRender.com.
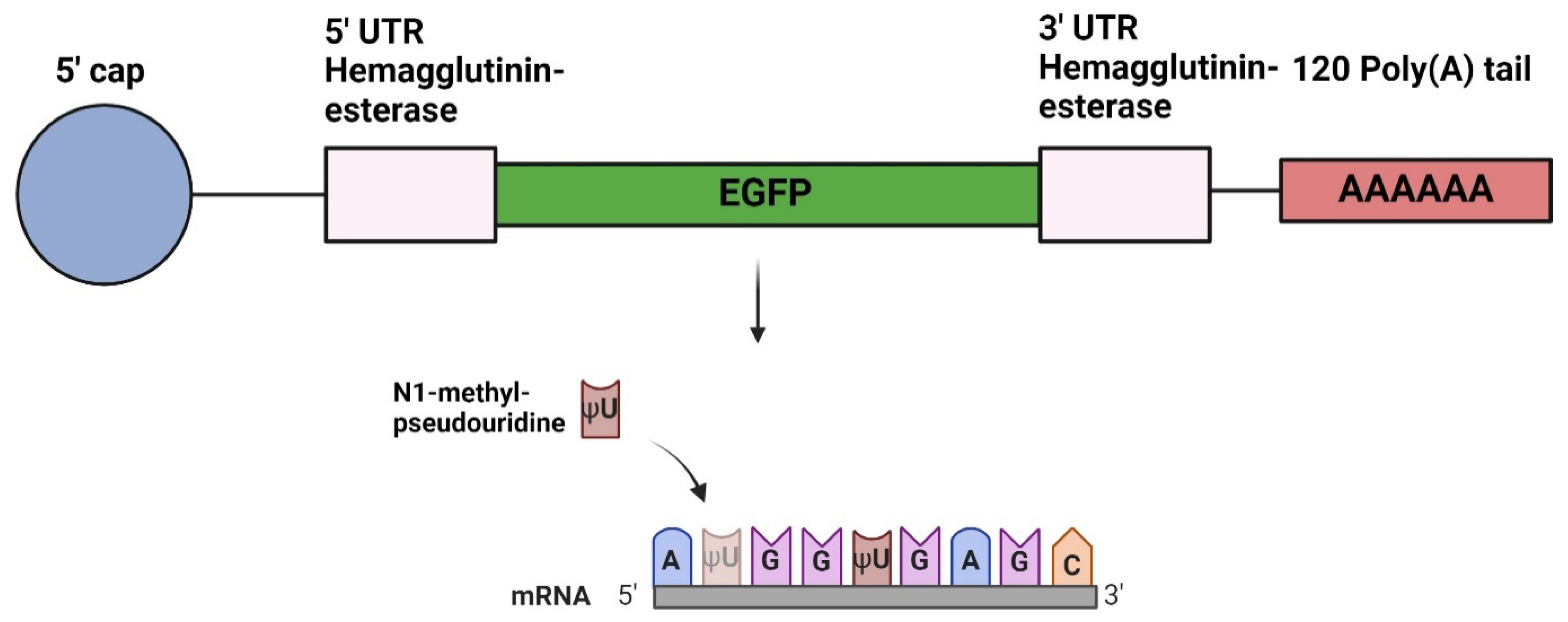
Figure 2.
Generation of EGFP and Fluc mRNA, and LNP encapsulation. (A) Evaluation of mRNA quality and size after IVT of EGFP and Fluc mRNAs with the use of a Tapestation 4200 instrument. (B,C) MD-FFF fractograms for the 2 mRNA-LNP formulations. (D) Summary of mRNA-LNP characteristics obtained with dynamic light scattering (DLS), multi detector field flow fractionation (MD-FFF) and the RiboGreen assay.
Figure 2.
Generation of EGFP and Fluc mRNA, and LNP encapsulation. (A) Evaluation of mRNA quality and size after IVT of EGFP and Fluc mRNAs with the use of a Tapestation 4200 instrument. (B,C) MD-FFF fractograms for the 2 mRNA-LNP formulations. (D) Summary of mRNA-LNP characteristics obtained with dynamic light scattering (DLS), multi detector field flow fractionation (MD-FFF) and the RiboGreen assay.
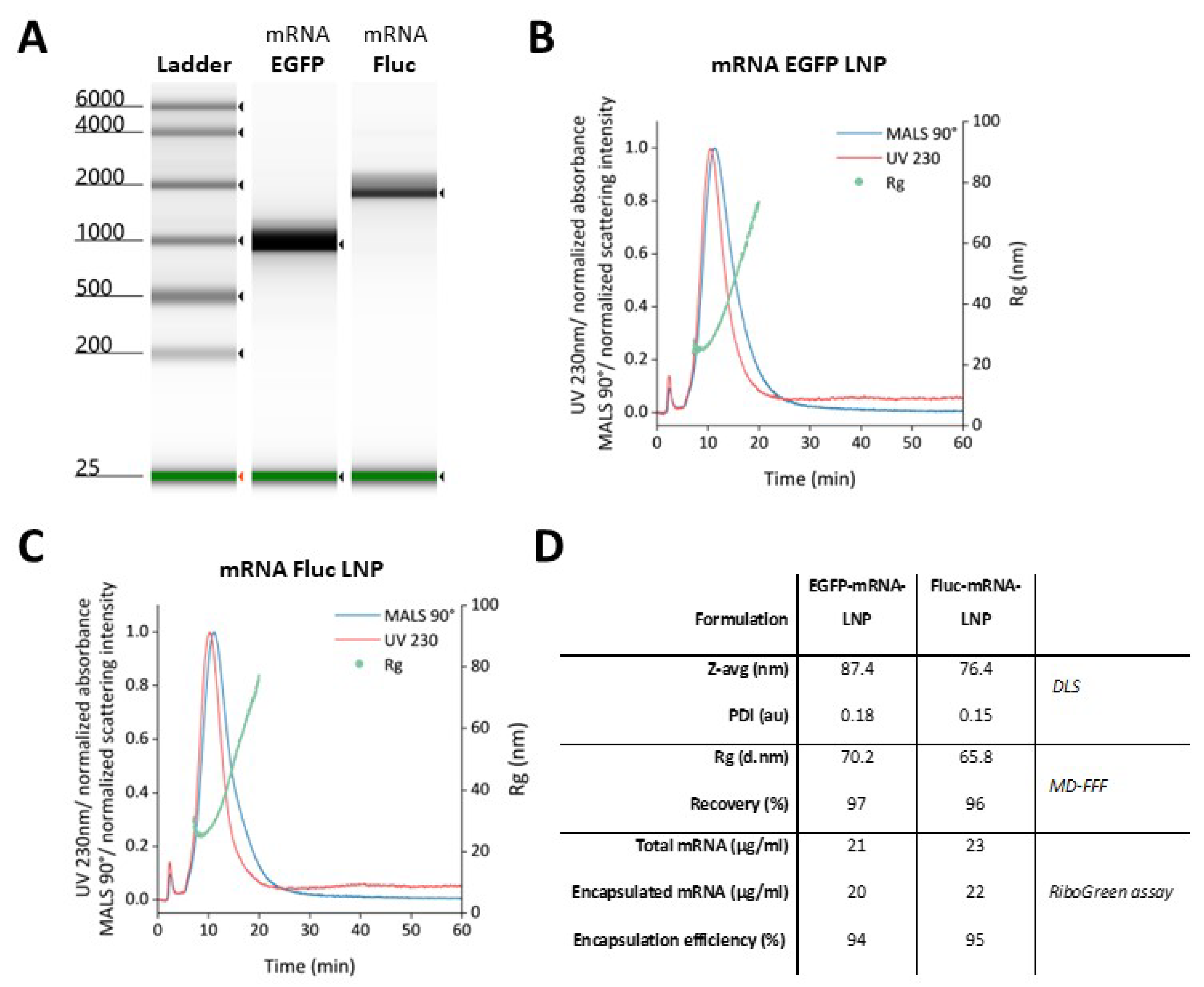
Figure 3.
Transfection of mRNA-LNPs encoding EGFP in CHH-1 cells. 500 ng of mRNA-LNP encoding EGFP was transfected in CHH-1 cells and protein expression was monitored by fluorescence microscopy analysis over 11 days.
Figure 3.
Transfection of mRNA-LNPs encoding EGFP in CHH-1 cells. 500 ng of mRNA-LNP encoding EGFP was transfected in CHH-1 cells and protein expression was monitored by fluorescence microscopy analysis over 11 days.
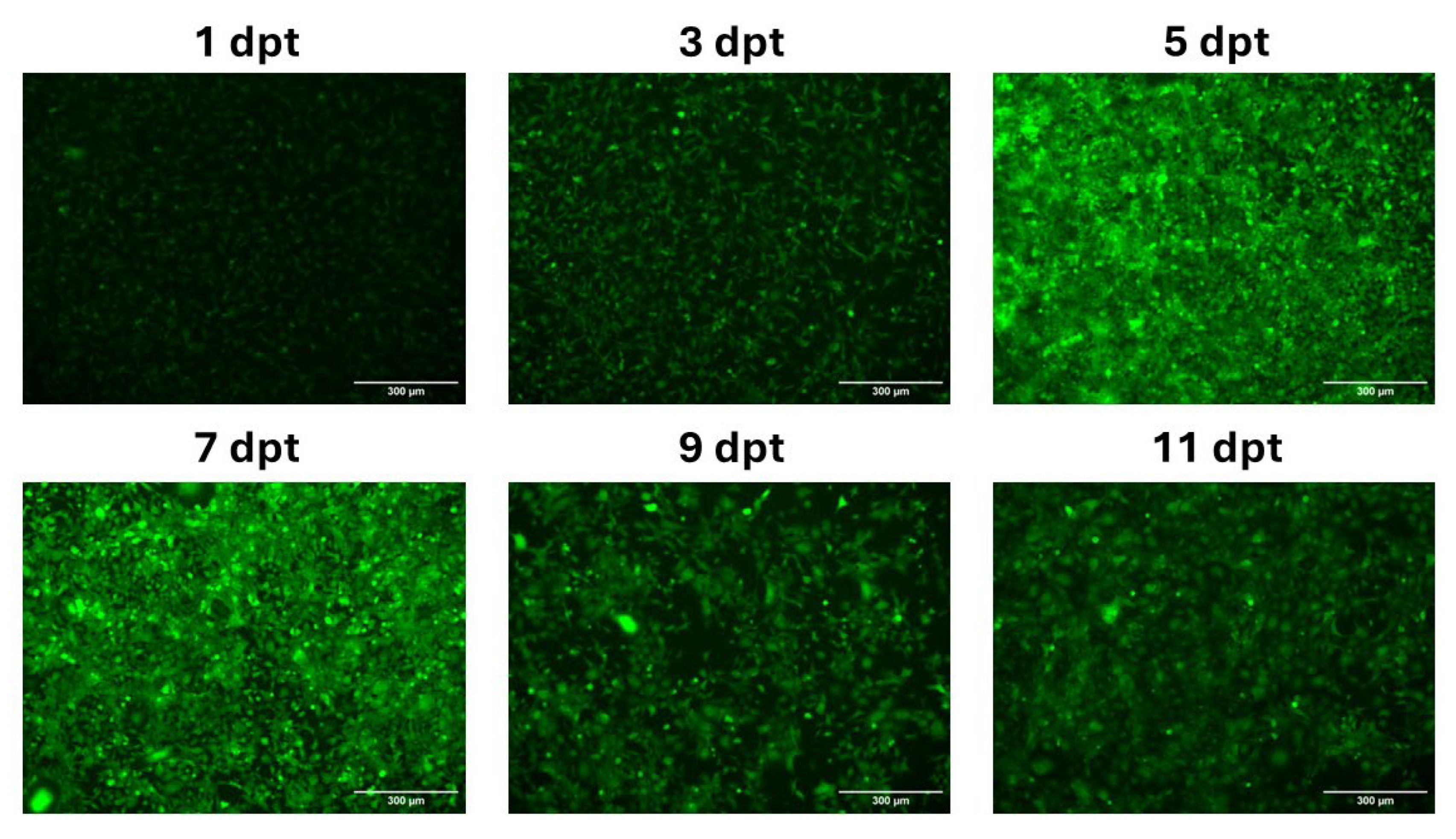
Figure 4.
Evaluation of cell cytotoxicity of mRNA-LNPs. Cell viability were monitored by an MTS assay at 1, 3 and 5 dpt in non-transfected control CHH-1 cells or CHH-1 cells treated with 285 ng or 570 ng of mRNA-LNPs.
Figure 4.
Evaluation of cell cytotoxicity of mRNA-LNPs. Cell viability were monitored by an MTS assay at 1, 3 and 5 dpt in non-transfected control CHH-1 cells or CHH-1 cells treated with 285 ng or 570 ng of mRNA-LNPs.
Figure 5.
Immunohistochemical detection of EGFP expression following mRNA-LNP injection. Atlantic salmon were injected with 1 µg of mRNA-LNP encoding EGFP or Fluc, and tissue samples were collected at 7 dpi. Formalin-fixed paraffin embedded tissue sections were stained with an antibody detecting GFP. (A) Muscle tissue injected with mRNA-LNPs encoding EGFP showed a focal inflammatory response in white skeletal musculature, where some leukocytes exhibit positive signal (brown), while others do not. (B) The negative control tissue was sampled from salmon injected with mRNA-LNPs encoding Fluc and showed no positive signal.
Figure 5.
Immunohistochemical detection of EGFP expression following mRNA-LNP injection. Atlantic salmon were injected with 1 µg of mRNA-LNP encoding EGFP or Fluc, and tissue samples were collected at 7 dpi. Formalin-fixed paraffin embedded tissue sections were stained with an antibody detecting GFP. (A) Muscle tissue injected with mRNA-LNPs encoding EGFP showed a focal inflammatory response in white skeletal musculature, where some leukocytes exhibit positive signal (brown), while others do not. (B) The negative control tissue was sampled from salmon injected with mRNA-LNPs encoding Fluc and showed no positive signal.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



