
Submitted:
17 June 2024
Posted:
18 June 2024
You are already at the latest version
Abstract
Photosystem I (PSI) serves as a model system for studying fundamental processes such as electron transfer (ET) and energy conversion, which are not only central to photosynthesis but also have broader implications for bioenergy production and biomimetic device design. In this study, we employed electron paramagnetic resonance (EPR) spectroscopy to investigate key light-induced charge separation steps in PSI isolated from several green algal and cyanobacterial species. Following photoexcitation, rapid, sequential ET occurs through either of two symmetric branches of donor/acceptor cofactors embedded within the protein core, termed the A and B branch. Using high-frequency (130 GHz) time-resolved EPR (TR-EPR) and deuteration techniques to enhance spectral resolution, we observed that at low temperatures prokaryotic PSI exhibits reversible ET in the A branch and irreversible ET in the B branch at low temperatures, while PSI from eukaryotic counterparts displays either reversible ET in both branches or exclusively in the B branch. Furthermore, we observed a notable correlation between low-temperature charge separation to the terminal [4Fe-4S] clusters of PSI, termed FA and FB, as reflected in the measured FA/FB ratio. These findings enhance our understanding of the mechanistic diversity of PSI's ET across different species and underscore the importance of experimental design in resolving these differences. Though further research is necessary to elucidate the underlying mechanisms and the evolutionary significance of these variations in PSI charge separation, this study sets the stage for future investigations into the complex interplay between protein structure, ET pathways, and the environmental adaptations of photosynthetic organisms.
Keywords:
photosynthesis
; EPR
; photosystem I
1. Introduction
Natural photosynthetic conversion of light energy to chemical energy is essential to all life on Earth. The primary energy conversion reactions involve photoinitiated rapid, sequential electron transfer (ET) steps that result in formation of a stabilized, long-lived charge separated state across a biological membrane. These reactions occur in large, integral membrane protein complexes called reaction centers (RCs).[1,2] The oxygenic photosynthesis that occurs in plants, algae, and cyanobacteria utilizes two types of RCs, Photosystem II (PSII) and Photosystem I (PSI).[3,4,5] PSII catalyzes the light-driven oxidation of water whereas PSI catalyzes light-driven transmembrane electron transfer from reduced plastocyanin to oxidized ferredoxin, which subsequently shuttles the reducing equivalents from PSI to various metabolic pathways including CO2 fixation. Each RC has at its core a network of molecular cofactors embedded within a protein matrix which specifically positions each cofactor in optimal geometry and distances along efficient electron transfer pathways.
The first high-resolution structure of PS I has been resolved by X-ray crystallography for the thermophilic cyanobacterium Thermosynechococcus elongatus (previously named Synechococcus elongatus and recently renamed to T. vestitus).[6] Two large subunits (PsaA and PsaB) form the heterodimeric core of PSI which has the same overall for different biological species. These two subunits bind the antenna pigments (about 100 chlorophyll and 20 carotenoid molecules) and the redox cofactors involved in the primary ET processes. Two branches of ET cofactors are arranged in a pseudo two-fold symmetry and referred to as the A and B branches (Figure 1). Following photoexcitation, the primary electron donor P700 (a dimer of chlorophyll molecules) becomes oxidized, transferring one electron to one of the two identical ET branches that each contain two additional chlorophyll molecules (an accessory Chl and the primary acceptor A0), and one phylloquinone (Vitamin K1) termed A1, or secondary acceptor. From the reduced A1-, the electron is transferred to the [4Fe–4S] cluster FX, located at the interface between PsaA and PsaB, and then to the terminal electron acceptors FA and FB, the two [4Fe–4S] clusters housed within an extrinsic protein subunit, PsaC, located on the stromal side of PSI.[3,6,7,8,9,10] Note, that the very first ultrafast steps of charge separation, including the involvement of the accessory Chls, have been extensively investigated by ultrafast optical spectroscopy and different models of the initial steps were proposed.[3,11,12,13,14,15,16] More recently, high resolution crystal and cryo EM structures of PSI (and PSI-LHCI supercomplexes) from different organisms like higher plants,[17] green algae[18,19] and mesophilic cyanobacterium Synechocystis sp. PCC 6803[20] have been determined which allow a structural comparison through an evolutionary lens. Photosynthetic organisms contain the PSI complex in different, but evolutionarily related forms tuned for efficient charge separation with near unity quantum yield.
For a long time, the ET in PSI was presumed to be unidirectional, i.e. only along the A branch by analogy to the Type II purple non-sulfur bacterial photosynthetic RC and Photosystem II, despite no different functionality of the quinones A1A and A1B. According to the primary paradigm, ET in PSI occurs only along the A branch and is blocked at low temperatures beyond A1A quinone. This model is partially in good agreement with our previous time-resolved (TR) electron paramagnetic resonance (EPR) studies of PSI from thermophilic Synechococcus lividus and mesophilic Synechococcus leopoliensis, where only one type of short-lived radical pair (RP), namely, P+A−1A, was observed,[21,22] and in agreement with a number of time-resolved EPR studies.[23,24,25,26,27,28] However, EPR spectra from PSI illuminated at low temperature always reveal the generation of photoinduced strong stable signals of P+ and reduced [4Fe-4S] clusters, which is direct proof of that ET from P to FX, FA, and FB occurs in a fraction of PSI complexes at low temperature. This was originally explained within the unidirectional model as ET occurring via the A branch in partially damaged RCs or RCs in a different conformational state.[21] As an alternative explanation, it was suggested that this ET does not occur via A branch but through the B branch, where the electrons do not stop at the quinone as the RP P+A−1B even at low temperatures but rather proceeds further to FX, FA, and FB. At low temperature, the [4Fe-4S] clusters FA and FB act as deep traps for electrons, and thus electron recombination to the oxidized primary donor P+ is largely suppressed. Therefore, at low temperatures, where most of the TR-EPR experiments are done, the transient RP from the A branch of PSI is the only one observed. Because the charge-separated state P+A−1A is short lived (≤200 μs) due to charge recombination, this ET is often called “reversible” or “cyclic”. In analogy, the ET is called “irreversible” if the electron can proceed beyond A1 and becomes trapped on [4Fe-4S] clusters at these low temperatures.[9,29]
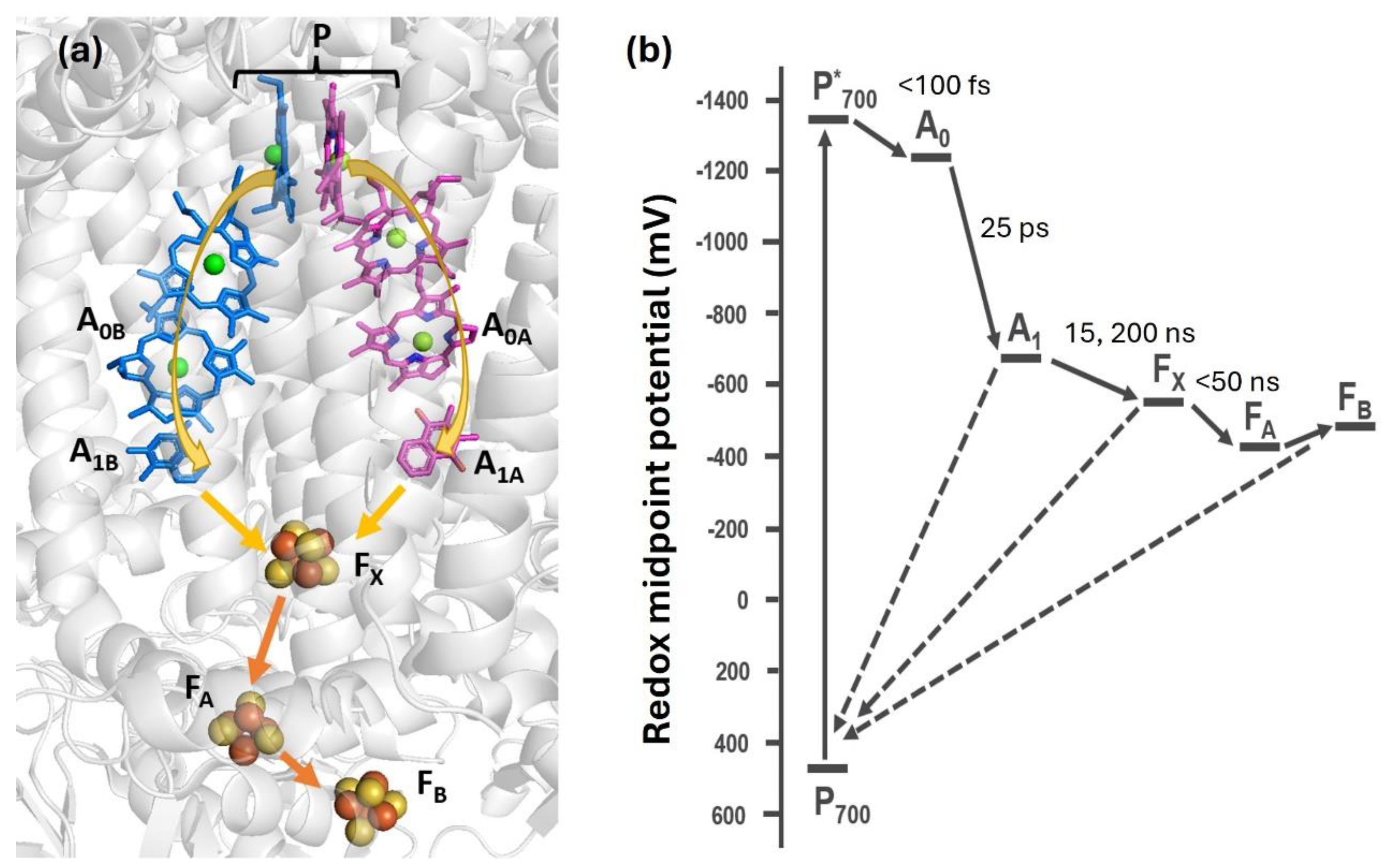
Figure 1.
(a) Schematic structure and ET pathways in cyanobacterial Photosystem I. Following photoexcitation, the primary donor P becomes oxidized, transferring one electron to one of two almost identical chains of electron transfer cofactors: chlorophyll A0, phylloquinone A1, and converging at the three [4Fe-4S] clusters, FX, FA, and FB. Photoinduced ET in PSI is bidirectional at ambient temperature, proceeding through both the A and B branches of cofactors as indicated by arrows. (b) Energy diagram and time constants of forward electron transfer reactions in PSI. Information was taken from refs. 9,10. .
Figure 1.
(a) Schematic structure and ET pathways in cyanobacterial Photosystem I. Following photoexcitation, the primary donor P becomes oxidized, transferring one electron to one of two almost identical chains of electron transfer cofactors: chlorophyll A0, phylloquinone A1, and converging at the three [4Fe-4S] clusters, FX, FA, and FB. Photoinduced ET in PSI is bidirectional at ambient temperature, proceeding through both the A and B branches of cofactors as indicated by arrows. (b) Energy diagram and time constants of forward electron transfer reactions in PSI. Information was taken from refs. 9,10. .
High frequency/high field D-band (130 GHz) TR-EPR experiments enabled the first direct detection of the transient radical pairs P+A−1A and P+A−1B in cyanobacterial PSI, which are clearly resolved at this frequency.[21,26] The geometric parameters of the two distinct donor/acceptor pairs correspond to the charge-separated states along the A and B branches and are in excellent agreement the X-ray crystal structure of PSI.[21] These experiments clearly demonstrated bidirectional ET at low temperature (≤100 K) under strongly reducing conditions for the thermophilic S. lividus PSI. Another crucial set of experiments that examined the involvement of the A and B branches in ET was performed on PSI from mesophilic S. leopoliensis where the [4Fe-4S] clusters FX, FA, and FB were removed to prevent forward ET beyond the quinones. TR-EPR spectra of these biochemically modified PSI complexes at 100 K are comprised of two overlapping signals: one from transient radical pair in the A branch, P+A−1A, and another from transient pair formed in the B branch, P+A−1B, with an almost equal ratio of ET through A and B branches.[22]
These results were possible due to two essential features of the experimental design: deuteration of PSI, which improves the spectral resolution by decreasing the EPR line width, and the high resolution afforded by high-frequency (HF) TR-EPR at 130 GHz/4.6 T. The deuteration of photosynthetic microorganisms by adaption and growth of bacteria, cyanobacteria and algae in heavy water (99.7% D2O) was pioneered over 60 years ago.[30,31,32] EPR studies of RCs isolated from deuterated non-sulfur purple bacteria and cyanobacteria have helped resolve the cofactors and local protein environments involved in light-induced charge separation events.[21,23,33,34,35,36,37,38,39] In contrast, advanced EPR studies of green algal PSI are lacking. Now we report the first EPR studies of deuterated PSI isolated from the green alga Chlorella vulgaris and Scenedesmus obliquus grown in 99.6% D2O. This allows us to obtain high resolution TR-EPR of the transient radical pair P+A1- at D-band (130 GHz) and interrogate ET to the terminal [4Fe-4S] acceptors with X-band EPR studies.
The bidirectional nature of ET in PSI at ambient temperature is nowadays well accepted.[21,22,33,36,39,40,41,42,43] At the same time a very important question remains: how does electron transfer differ from species to species and is there any relation to the electronic properties of [4Fe-4S] clusters? While there have been investigations of PSI isolated from various species, they typically used different methodologies and approaches which make direct comparisons challenging. In this publication, we report a comparative study of isolated PSI from five biological species: Synechococcus S. lividus, Synechococcus S. leopoliensis, Thermosynechococcus elongatus, Chlorella vulgaris and Scenedesmus obliquus. The former three species are either mesophilic or thermophilic cyanobacteria and thus prokaryotes, while the latter two are green algae and thus eukaryotes (like higher plants). RC charge separation has been shown to be sensitive to dynamic protein conformational substates and local heterogeneous protein environments that surround the cofactors.[33,44,45,46,47,48] The comparison of species-dependent spectral signatures thus can provide insight in evolutionary adjustments of PSI protein matrices and charge-separation to different environmental conditions and provide valuable insight into the mechanisms employed by nature to fine tune ET in RCs.
2. Materials and Methods
2.1. Sample Preparation
Synechococcus lividus PSI. PSI RCs were prepared from whole cells of the cyanobacterium Synechococcus lividus as described previously.[49] The final buffer was 20 mM HEPES, pH 8, 0.03% β-DM (n-dodecyl-β-D-maltopyranoside, Anatrace). Sodium ascorbate was added to a final concentration of 5 mM from a concentrated stock solution of 0.64 M sodium ascorbate in 50mM MES, pH 6. For EPR measurements, PSI was concentrated with 50 kDa MWCO microconcentrators (Millipore) to the desired concentration. All samples were kept in the dark and on ice until used for EPR.
Synechococcus leopoliensis PSI. PSI RCs were isolated from whole cells of the cyanobacterium Synechococcus leopoliensis which were grown either in H2O or in D2O (99.6%). Purified PSI was prepared in 20 mM HEPES, pH 8, and 0.03% β-DM. 5 mM sodium ascorbate was added prior to EPR measurement. The [4Fe−4S] clusters (FX, FA, FB) were removed by established preparations. PSI (0.2 mg of chlorophyll/mL) was incubated in a buffer containing 6.8 M urea, 62 mM Tris, and 76 mM glycine−NaOH, pH 10, for 1 h, to remove FA/FB as previously described.[50] The PSI sample was then dialyzed overnight against 50 mM Tris-HCl, pH 8.3. To remove FX, the sample was further treated with 3 M urea, 5mM K3FeCN6, and 50 mM Tris-HCl, pH 8.0, for 4.5 h.[51] The PSI sample was dialyzed overnight against 50 mM Tris-HCl, pH 8.3, and 5 mM 4,5-dihydroxy-1,3-benzene disulfonic acid (disodium salt) and then again overnight against 2 changes of 50 mM Tris-HCl, pH 8.0, 0.03%β-DM. The sample, analyzed by ICP-AES, showed a ratio of ∼1 Fe/PSI monomer after urea treatment, confirming quantitative removal of the three [4Fe−4S] clusters. For EPR measurements, 5 mM sodium ascorbate or 50 mM Tricine-NaOH, pH 8.0, and 10 mM sodium hydrosulfite were added prior to freezing in liquid nitrogen.
Thermosynechococcus elongatus PSI (previously named Synechococcus elongatus and recently renamed Thermosynechococcus vestitus). Trimeric photosystem I was extracted from whole cells of the cyanobacterium Thermosynechococcus vestitus which were cultivated as previously described.[52] After separating PSI from other solubilized protein using a DEAE650 anion exchanger column, the PSI fractions were pooled and purified using a SP Sepharose column with buffer A (5 mM Mes, pH 5.5; 0.013%C12E8(octaethyleneglycol monododecyl ether, Anatrace)) and B (5 mM Mes, pH5.5; 500 mM NaCl; 0.013%C12E8). The purified PSI fractions were pooled again and washed with storage buffer containing 5mM Mes, pH 6; 30 mM MgSO4, and 0.013% C12E8 using an Amicon concentrator with 100 kDa cutoff. Once a concentration of 7.5 mM chlorophyll was reached, trimeric PSI was diluted with buffer containing 5 mM MES pH 6 and 0.013% C12E8 to start crystallization. The crystallization took place overnight and resulted in rectangular shaped crystals of about 40μm in the longest dimension. Afterward, the crystals were pelleted, and after removal of the supernatant, storage buffer was added to dissolve the crystals and yield a solution of PSI at the desired chlorophyll concentration.
Chlorella vulgaris and Scenedesmus obliquus thylakoid preparation.Chlorella vulgaris and Scenedesmus obliquus were grown in 99.6 % heavy water as described.[53] Algal cells (5 g) were resuspended in 30 mM Tricine-NaOH, pH 8.0, 300 mM sucrose, and 15 mM NaCl. The cell suspension was placed in a pre-chilled Bead-Beater (BioSpec Products, Inc) with 1 mm glass beads. The sample was beaten for 5 x 1 min bursts, with 5 min rest in between with cooling in a surrounding ice bath. The solution was decanted and spun at 2000 rpm for 2 min in a Beckman Coulter Avanti J-26 XP with a JLA 16.25 rotor to remove glass beads. Unbroken cells and starch were removed by centrifugation at 7000 rpm for 10 min in the JLA 16.25 rotor. The supernatant was spun at 45,000 rpm for 2 hours in a Beckman L-60 ultrafuge with a 60 Ti rotor. The pellets were resuspending in 30 mM Tricine-NaOH, pH 8.0, 300 mM sucrose, and 150 mM NaCl and incubated on ice for 30 min. The sample was pelleted by ultracentrifugation at 45,000 rpm for 2 hours. The pellet was resuspended in 30 mM Tricine-NaOH, pH 8.0, 300 mM sucrose, and 15 mM NaCl at a concentration ~ 2 mg/ml Chl and stored at -80 C.
Chlorella vulgaris and Scenedesmus obliquus PSI isolation. PSI was extracted from thylakoid membranes diluted to 1 mg/ml Chl by addition of 2% β-DM. Following 30 min incubation on ice, insoluble material was removed by centrifugation at 45,000 for 30 min in a Beckman L-60 ultrafuge with a 60 Ti rotor. The supernatant was loaded onto a Toyopearl DEAE 650-C column equilibrated with 30 mM Tricine, pH 8, 15 mM NaCl and 0.2 % β-DM. Protein was eluted from the column with a linear NaCl gradient (15 -250 NaCl) in 30 mM Tricine-NaOH pH 8.0, 0.2% β-DM. The middle dark green fractions were pooled and precipitated with 10% PEG3350. The sample was immediately centrifuged at 5000 rpm in a Beckman Coulter Avanti JA30.50 rotor for 5 min. The green pellet was resuspended in 30 mM Tricine-NaOH, pH 8.0, 0.05% β-DM. The sample was loaded onto a 15-40% sucrose density gradient prepared in 30 mM Tricine-NaOH, pH 8.0, 0.05% β-DM and 15 mM NaCl and spun overnight at 40,000 rpm in a 50Ti rotor at 4 C. The lower band containing PSI was collected. The upper green band contained LHCs. The PSI was repeatedly washed to remove the sucrose using 50 kDa MWCO microconcentrators (Millipore). PSI was stored at -80 C until thawed for EPR use. Samples were further concentrated, and 5 mM sodium ascorbate was added prior to EPR measurements.
2.2. EPR Spectroscopy
X-band: Continuous wave (cw) X-band (9.5 GHz) EPR measurements were carried out with a Bruker ELEXSYS II E500 EPR spectrometer (Bruker Biospin Corp, Ettlingen, Germany) equipped with a TE102 rectangular EPR resonator (Bruker ER 4102ST) and helium gas-flow cryostat (ICE Oxford, UK). Temperature control was provided by an ITC (Oxford Instruments, UK). The cw EPR experiments used field modulation with phase sensitive lock-in detection. This type of detection results in the first derivative-type EPR spectra. Microwave power is given as attenuation (in dB) with respect to 200 mW. EPR samples were prepared in a N2 box, placed in 4 mm o.d. quartz EPR tubes, capped, and frozen in liquid N2 prior to placement in the pre-cooled EPR resonator. Light excitation of the sample was achieved with a Solis-3c “Day Light White” LED (Thorlabs, Newton, NJ).
D-band: EPR measurements were performed on a pulsed/continuous-wave high-frequency (HF) D-band (130 GHz/4.6 T) EPR spectrometer[26,54] with a single mode TE011 cylindrical resonator. Pulsed EPR spectra of stable radical species were recorded by monitoring the electron spin echo (ESE) intensity from a two-microwave pulse sequence as a function of magnetic field. Pulsed TR-EPR spectra were recorded in a similar way by initial photoexcitation of the protein by a short (<10 ns) laser pulse followed by the microwave pulses (π/2−τ−π−τ−echo) at a fixed delay after flash (DAF) time. The duration of the π/2 microwave pulse was 40−60ns. Light excitation of the sample was achieved with an optical parametric oscillator (OPO; basiScan, GWU Laser technik, Germany) pumped by a Nd: YAG laser (Quanta-Ray INDI, Spectra-Physics), the output of which was coupled to an optical fiber. The optical fiber allows delivery of up to 1 mJ/pulse to the sample. The excitation wavelength was 550nm. The samples were loaded into quartz tubes, dark-adapted, and placed in the precooled microwave cavity. The cavity was mounted in an Oxford flow cryostat, and temperature was controlled by an Oxford temperature control system (ITC503).
3. Results and Discussion
3.1. Spin Correlated Radical Pair (SCRP)
To obtain the signatures of the ET pathways in PSI from the different species we performed time-resolved EPR (TR-EPR) experiments at high magnetic fields. Optical excitation of PSI initiates ET leading to the generation of one or more sequential radical-ion pairs (RPs), including the so-called secondary pair P700+ A1– (earlier RPs are too short-lived to be detected by EPR). The weakly interacting spins of the radical pair are initially entangled or correlated and are known as spin-correlated radical pairs (SCRPs).[23,33,34,55] Since the SCRPs are created by rapid ET from the photoexcited singlet state of the primary donor P700 , initially only those states in the four-level system which have singlet character are populated.[55,56] This strong non-Boltzmann electron spin polarization results in line shapes different from radical pair (RP) spectra in thermal equilibrium, creating of a series of alternating emissive and absorptive lines (“antiphase doublets”). These SCRPs are exceptionally sensitive to weak magnetic interactions, structure, and heterogeneous local protein environments and thus can be used as highly sensitive sensors for any changes, e.g. in mutual orientation of the paramagnetic cofactors P700+ and A1–. [24,25,57,58,59,60,61,62] The line shape of the SCRP is especially informative when recorded with high spectral resolution at HF EPR.
Figure 2a shows HF (130 GHz) pulsed EPR spectra of the P700+A−1A radical pair from fully deuterated cyanobacterium S. leopoliensis in thermal equilibrium (purple) and in spin-polarized SCRP state (red) recorded at 100 K. Green and blue spectra in are the simulations for EPR spectra in thermal equilibrium of A1A− and P700+, respectively. The low-field part of the spectra is dominated by the signal from the quinone acceptor A1–, while the high-field part of the spectra is dominated by signals from the primary donor P700+. The gx and gy components of the rhombic g-tensor of A1A− are clearly resolved and well-separated from P700+ signal. Note, that the combination of HF 130 GHz EPR and fully deuterated PSI allows to clearly resolve the rhombic g-tensors of both the quinone anion radical A1A− and the chlorophyll cation radical P700+. No hyperfine structure is visible under these conditions. The TR-EPR signal shows the derivative type of lines characteristic of the SCRP spectrum of P700+ A1– (Figure 2). It is important to mention that the shape and the phase of the lines in SCRP depends on the mutual orientation of the g-tensors of P700+ and A1– and thus the respective molecular orientation. As a consequence, the SCRP spectra are sensitive to whether the SCRP in the A or B branches are detected, i.e. the SCRP P700+ A1A– and P700+ A1B– give different EPR spectra and can thus be distinguished from each other. A detailed technical discussion of this topic is presented in previous publications.[21,22,33].
As discussed above, to observe the difference in the line shapes for the transient SCRP P+A1- from A and B branches, we employed established methods for blocking ET beyond the secondary acceptor A1 by (photo)chemical reduction of the later electron acceptors in PSI. The light-induced HF high frequency TR-EPR signal observed from the dark-adapted non-reduced deuterated S. leopoliensis PSI sample containing the mild reductant sodium ascorbate (Figure 2b, red) is due to ET through the A branch where the ET beyond A1A is blocked at low temperatures like 100 K and below,[29] and thus the P+A1A- SCRP is observed. This SCRP recombines and the next laser flash will again generate the P700+ A1A- SCRP. Hence, this SCRP is referred to as “reversible” or “cyclic”. For this type of sample, the SCRP P700+ A1B- does not contribute to the spectrum because in this case, ET along the B chain proceeds beyond A1B to generate long-lived or stable P700+FX-, P700+FA-, and P700+FB- states.
To observe the signature of B branch SCRP, PSI samples containing sodium hydrosulfite were prereduced by illumination at 205-245 K and then the temperature lowered to 100 K. This so-called photoaccumulation procedure allows to reduce FA, FB, FX, and A1A but not A1B. The light-induced high frequencyHF TR-EPR spectrum after this treatment is shown in Figure 2b, blue. This is a characteristic spectrum of SCRP in B branch, P+A1B- (Figure 2b, blue). The different line shapes of the two SCRP spectra of P700+A1A- and P700+A1B-, particularly at the high-field portions where P+ is dominating, are caused by the different directions of the P700+ - A1A- and P700+ - A1B- interspin vectors in the g-tensor principal axes system of P700+. Differences in the A1- contribution to the spectrum are much more subtle since the two interspin vectors are very comparable in the g-tensor principal axis system of either quinone. Small changes in the gx value and linewidth of the two quinones have been reported.[21,22,33]
SCRPs in both A and B branches can be detected simultaneously in PSI where the three Fe-S clusters FX, FA and FB were removed to prevent forward ET from the quinones to the [4Fe-4S] clusters (Figure 2b, green). In this case the TR-EPR spectrum at 100 K is comprised of two overlapping spectra: one from SCRP in the A branch, P+A−1A, and another one from SCRP formed in the B branch, P+A−1B. This observed spectrum can be modeled as the sum of the SCRP in A and B branches with an almost equal ratio. This is direct evidence that in native PSI from the cyanobacterium S. leopoliensis both branches are equally active at low temperature.
The pulsed high frequencyHF TR-EPR spectra of SCRPs were recorded for five biological species: S. lividus, S. leopoliensis, T. vestitus, C. vulgaris and S. obliquus are shown in Figure 3. All spectra (except some S. leopoliensis samples) were recorded on PSI complexes with a small amount the mild reductant sodium ascorbate added. added which ensures that in dark-adapted PSI the primary donor is reduced. This reductant is not reducing enough to change the oxidation state of the [4Fe-4S] clusters FX, FA and FB. When available, spectra were recorded both for fully protonated and fully deuterated PSI.
The SCRP spectra recorded for protonated PSI are very similar, indicating that no major changes in the g-tensors of either P700+ or A1-, nor in the mutual orientation of these spin-carrying cofactors occurred (Figure 3). Thus, the overall surrounding of these two cofactors and the overall structural arrangement is comparable in all protonated samples from the different species. While there are minor differences in the spectral lineshape in P700+ region, these differences are hard to interpret. In contrast, clear differences can be observed in the lineshapes of SCRP from deuterated proteins, where the linewidth is significantly reduced due to the reduction of hyperfine interaction (Figure 3). In particular, the P700+ region of the spectra recorded for deuterated PSI are different. The lineshape of SCRP spectrum in S. leopoliensis and S. lividus is typical for A branch SCRP, P700+ A−1A (compared with red spectrum in Figure 2b). In the case of C. vulgaris the line shape is different and resembles the line shape of the SCRP in S. leopoliensis with FA, FB, and FX clusters removed, where both SCRPs in A and B branches are visible (Figure 2, green). For the ease of comparison, the SCRP of deuterated S. leopoliensis with FA, FB, and FX clusters removed were recorded under similar experimental conditions and a good agreement with the C. vulgaris spectrum was observed (Figure 3). For PSI from S. obliquus the differences of the SCRP with the A branch SCRP are even more pronounced and instead it resembles the blue spectrum shown in Figure 2b, where only SCRP in the B branch, P700+ A1B– , is observed.
The straightforward interpretation of these data is the follows. In C. vulgaris at low temperatures ET is not proceeding beyond acceptor A1 in both branches, thus both SCRP, P700+ A−1A and P700+ A−1B are visible in the TR-EPR spectra with almost equal amounts. In Scenedesmus obliquus only SCRP in B-branch, P+A−1B, is visible. The plausible explanation that, contrary to S. leopoliensis and S. lividus, at low temperatures in Scenedesmus obliquus ET beyond A1 is blocked in the B-branch but can proceed to the Fe-S cluster via the A-branch.
These are remarkable results indicating that low temperature ET pathways in green algae PSI are different from those in cyanobacteria. In previous studies, differences in the directionality of electron ET between prokaryotic and eukaryotic PSIs were observed, with the fraction of the B branch generally being larger and having a longer lifetime in eukaryotic PSI compared to prokaryotic PSI.[63]
The difference of the low temperature ET in PSI and relative activity of A and B branches was explained by relative redox potential of acceptors A1A and A1B in respect to Fx (see Scheme 1). If the redox potential of A1A is higher (less reducing) compared to the redox potential of Fx, then at low temperatures cyclic ET is observed in A-branch. If redox potential of A1B is higher (less reducing) than the redox potential of Fx, then ET in B-branch is cyclic/reversable. In the case when Fx potential lower than redox potentials of both acceptors A1A and A1B, cyclic ET can be observed in both branches with the ratio determined by relative potential of A1A to A1B.[38,40]
Importantly, in 1999, Joliot and Joliot observed at ambient temperature biphasic kinetics of oxidation of reduced phylloquinone and attributed these two phases to oxidation A1A and A1B , respectively. Since the amplitudes of both phases were comparable, they interpreted this as a sign of equal ET activity of both branches in green algae Chlorella sorokiniana.[64] This is an indication of the similar microenvironment and, as consequence, similar redox potentials of A1A and A1B acceptors. At low temperature this might result in cyclic/reversible ET both in the A- and B-branches in Chlorella sorokiniana PSI, similar to what we observed for C. vulgaris.
3.2. Iron Sulfur Clusters
Iron sulfur [4Fe-4S] clusters within PSI reaction centers play a vital role in electron transfer. The three [4Fe-4S] clusters FX, FA and FB shuttle electrons form the site of initial charge separation to the stromal side where mobile electron transfer proteins ferredoxin or flavodoxin are facilitating further ET.[7] As pointed out above, the relative redox potentials of these complexes determine whether one or another branch of ET in PSI is active and photoinduced electron transfer can proceed through a quinone acceptor to the final acceptors. EPR spectroscopy is the most informative technique for detecting and resolving signals from individual [4Fe-4S]clusters, offering structural insights otherwise challenging to obtain. To compare if there are substantial differences between electronic characteristic of [4Fe-4S] complexes in the proteins under study and test the relationships between these properties and directionality of ET, we analyzed EPR spectra of the reduced [4Fe-4S] complexes. The cw X-band EPR spectra of the reduced [4Fe-4S] complexes (Figure 4) were recorded for the same five biological species as above: S. lividus, S. leopoliensis, T. elongatus, C. vulgaris and S. obliquus.
In the oxidized state, the [4Fe-4S]2+ are diamagnetic and thus give no EPR signal. In the reduced state, [4Fe-4S]1+, one ferric and three ferrous iron atoms within the clusters are magnetically coupled, resulting in an effective total spin of S = ½. EPR signals from [4Fe-4S] clusters have extremely short relaxation times, thus the measurements were done at 10 K. Freezing a PSI complex in the dark, with subsequent illumination at low temperature allows promotion of only one electron from P700 to either FA or FB, but not both, in a given PSI complex. Consequently, the resulting EPR spectra represent a sum of signals from reduced centers with three distinct g-values, reflecting 'rhombic' symmetry of the g-tensor. The redox potential of the [4Fe-4S] cluster FX is too low and thus cannot be stabilized under our experimental conditions. To obtain g-tensor parameters gx, gy, and gz of both FA and FB centers and their relative contribution to the experimental spectra (FA/FB ratio), the spectra were simulated (see Figure S1). The simulation parameters are summarized in Table 1.
The g-tensor principal values are in the typical range for reduced FA and FB clusters reported in the literature. While these values are slightly different for each type of PSI, there is no clear trend of this dependence from one to another species. Within the experimental error, no differences in the g-values or FA/FB ratio between protonated and deuterated proteins were detected.
Surprisingly, we observed that the FA/FB ratios in PSI of green algae differ from those PSI complexes from cyanobacteria. For the thermophilic cyanobacteria T. e longatus and S. lividus, the ratio of FA to FB is two to one. In the mesophilic S. leopoliensis, this ratio increases to four to one. In contrast, for the two green algae, C. vulgaris and Scenedesmus obliquus the FA to FB ratio is approximately equal. This can be correlated to the activity of A or B branches at low temperatures and, as a consequence, to the redox potentials of A1A and A1B relative to FX.
Indeed, in T. elongatus, S. lividus, and S. leopoliensis the FA/FB ratio exceeds 1. In these proteins low temperature ET cyclic in the A branch and irreversible in B branch. This can be explained by the following order of the PSI acceptors redox potentials: A1A > Fx> A1B. The observed ratio FA/FB of 2 and 4 can be accounted for slightly higher redox potential of FA. On the contrary, in Chlorella vulgaris and Scenedesmus obliquus, where FA/FB ratio is equal to 1, B branch is more active in cyclic ET at low temperature. In S. lividus the cyclic ET observed in both A and B branches, indicating that their redox potentials are higher than redox potential of FX. In S. leopoliensis only B branch demonstrates cyclic ET. Equal contribution of FA and FB to the EPR spectrum can be explained by equal redox potential of these complexes.
4. Conclusions
EPR spectroscopy was used to study light-induced charge separation in PSI isolated from both prokaryotes (cyanobacteria) and eukaryotes (green algae). In PSI, primary charge separation occurs through two symmetric branches of redox cofactors, the A and B branch, and the bidirectional ET asymmetry can be differentiated with low temperature high-frequency TR-EPR spectroscopy of deuterated PSI complexes. In this work, fully deuterated green algal PSI was utilized to resolve, for the first time, the SCRPs P700+A−1A and P700+A−1B involved in eukaryotic PSI charge separation. Whereas at low temperatures prokaryotic PSI shows cyclic ET in A branch and irreversible ET in the B branch, green algal PSI shows cyclic ET in both A and B branches or only the B branch. This is a surprising result. Prior to this, B-branch SCRP P700+A−1B had been observed for “special” PSI mutants and cyanobacterial PSI pretreated with strong reducing conditions or upon removal of the [4Fe-4S] clusters FX, FA, and FB. Light-driven low temperature charge separation from the primary donor P700 to FA and FB shows that these clusters are intact in the isolated green algal PSI. Furthermore, we observed a notable species-dependent correlation between low temperature charge separation in the measured FA/FB ratio. As the core molecular cofactor ET chains are the same in eukaryotic and prokaryotic PSI structures, we hypothesize that the dynamic protein matrix plays a role in controlling the relative ratios of A and B branch primary charge separation and charge separation to the [4Fe-4S] clusters. These differences could be reflective of localized heterogeneous protein environments or, perhaps, reflect overall protein oligomeric structures, with algal PSI typically being monomeric and attached to multiple LHCI antenna in PSI-supercomplexes versus cyanobacterial PSI which is trimeric and can be purified without bound LHCI. Though further research is necessary to elucidate the underlying mechanisms to explain these experimental variations in PSI charge separation, this study sets the stage for future investigations into the complex interplay between protein structure, ET pathways, and the environmental species-dependent adaptations of photosynthetic organisms.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supporting Information including simulations of all cw X-band (9.5 GHz) EPR spectra. The Supporting Information is available free of charge at.
Acknowledgments
This material is based upon work supported by the U.S. Department of Energy, Office of Science, Office of Basic Energy Sciences, Division of Chemical Sciences, Geosciences, and Biosciences, under contract number DE-AC02-06CH11357 at Argonne National Laboratory (J.K.B., T.M., M.Y.T., J.N., L.M.U., O.G.P.). M.K. and J.K. acknowledge support by the U.S. Department of Energy, Office of Science, Office of Basic Energy Sciences under contract number DE-AC02-05CH11231.
Abbreviations
RC- reaction center; PSI- Photosystem I; PSII- Photosystem II; HF- high frequency; ET- electron transfer; SCRPs- spin correlated radical pairs; EPR- electron paramagnetic resonance; ESE- electron spin echo; DAF- delay after flash.
References
- Blankenship, R. E. Molecular Mechanisms of Photosynthesis; Blackwell Science Limited 2002.
- Lawlor, D. W. Photosynthesis; BIOS Scientific PublishersLimited, 2001.
- Golbeck, J. H. Photosystem I : the light-driven plastocyanin:ferredoxin oxidoreductase; Springer, 2006.
- Shevela, D.; Kern, J. F.; Govindjee, G.; Messinger, J. Solar energy conversion by photosystem II: principles and structures. Photosynth Res 2023, 156, 279–307. [Google Scholar] [CrossRef]
- Wydrzynski, T. J. S., K.; Freeman, J. A. The Light-Driven Water:Plastoquinone Oxidoreductase; Springer, 2005.
- Jordan, P.; Fromme, P.; Witt, H. T.; Klukas, O.; Saenger, W.; Krauss, N. Three-dimensional structure of cyanobacterial photosystem I at 2.5 Å resolution. Nature 2001, 411, 909–917. [Google Scholar] [CrossRef]
- Vassiliev, I. R.; Antonkine, M. L.; Golbeck, J. H. Iron-sulfur clusters in type I reaction centers. BBA-Bioenergetics 2001, 1507, 139–160. [Google Scholar] [CrossRef]
- Brettel, K. Electron transfer and arrangement of the redox cofactors in photosystem I. BBA-Bioenergetics 1997, 1318, 322–373. [Google Scholar] [CrossRef]
- Brettel, K.; Leibl, W. Electron transfer in photosystem I. BBA-Bioenergetics 2001, 1507, 100–114. [Google Scholar] [CrossRef]
- Mamedov, M.; Govindjee; Nadtochenko, V. ; Semenov, A. Primary electron transfer processes in photosynthetic reaction centers from oxygenic organisms. Photosynth Res 2015, 125, 51–63. [Google Scholar] [CrossRef]
- Cherepanov, D. A.; Shelaev, I. V.; Gostev, F. E.; Nadtochenko, V. A.; Xu, W.; Golbeck, J. H.; Semenov, A. Y. Symmetry breaking in photosystem I: ultrafast optical studies of variants near the accessory chlorophylls in the A and B branches of electron transfer cofactors. Photochem. Photobiol. Sci. 2021, 20, 1209–1227. [Google Scholar] [CrossRef]
- Holzwarth, A. R.; Müller, M. G.; Niklas, J.; Lubitz, W. Charge recombination fluorescence in photosystem I reaction centers from Chlamydomonas reinhardtii. J Phys Chem B 2005, 109, 5903–5911. [Google Scholar] [CrossRef]
- Holzwarth, A. R.; Müller, M. G.; Niklas, J.; Lubitz, W. Ultrafast transient absorption studies on Photosystem I reaction centers from Chlamydomonas reinhardtii. 2: Mutations near the P700 reaction center chlorophylls provide new insight into the nature of the primary electron donor. Biophys. J. 2006, 90, 552–565. [Google Scholar] [CrossRef]
- Müller, M. G.; Niklas, J.; Lubitz, W.; Holzwarth, A. R. Ultrafast transient absorption studies on Photosystem I reaction centers from Chlamydomonas reinhardtii. 1. A new interpretation of the energy trapping and early electron transfer steps in Photosystem I. Biophys. J. 2003, 85, 3899–3922. [Google Scholar] [CrossRef]
- Müller, M. G.; Slavov, C.; Luthra, R.; Redding, K. E.; Holzwarth, A. R. Independent initiation of primary electron transfer in the two branches of the photosystem I reaction center. Proc. Natl. Acad. Sci. U. S. A. 2010, 107, 4123–4128. [Google Scholar] [CrossRef]
- Gorka, M.; Baldansuren, A.; Malnati, A.; Gruszecki, E.; Golbeck, J. H.; Lakshmi, K. V. Shedding Light on Primary Donors in Photosynthetic Reaction Centers. Front. Microbiol. 2021, 12, 31. [Google Scholar] [CrossRef]
- Mazor, Y.; Borovikova, A.; Caspy, I.; Nelson, N. Structure of the plant photosystem I supercomplex at 2.6 Å resolution. Nat Plants 2017, 3. [Google Scholar] [CrossRef]
- Qin, X. C.; Pi, X.; Wang, W. D.; Hang, G. Y.; Zhu, L. X.; Liu, M. M.; Cheng, L. P.; Shen, J. R.; Kuang, T. Y.; Sui, S. F. Structure of a green algal photosystem I in complex with a large number of light-harvesting complex I subunits. Nat Plants 2019, 5, 263–272. [Google Scholar] [CrossRef]
- Su, X. D.; Ma, J.; Pan, X. W.; Zhao, X. L.; Chang, W. R.; Liu, Z. F.; Zhang, X. Z.; Li, M. Antenna arrangement and energy transfer pathways of a green algal photosystem-I-LHCI supercomplex. Nat Plants 2019, 5, 273–281. [Google Scholar] [CrossRef]
- Malavath, T.; Caspy, I.; Netzer-El, S. Y.; Klaiman, D.; Nelson, N. Structure and function of wild-type and subunit-depleted photosystem I in. BBA-Bioenergetics 2018, 1859, 645–654. [Google Scholar] [CrossRef]
- Poluektov, O. G.; Paschenko, S. V.; Utschig, L. M.; Lakshmi, K. V.; Thurnauer, M. C. Bidirectional electron transfer in photosystem I: Direct evidence from high-frequency time-resolved EPR spectroscopy. J Am Chem Soc 2005, 127, 11910–11911. [Google Scholar] [CrossRef]
- Poluektov, O. G.; Utschig, L. M. Directionality of Electron Transfer in Type I Reaction Center Proteins: High-Frequency EPR Study of PS I with Removed Iron-Sulfur Centers. J Phys Chem B 2015, 119, 13771–13776. [Google Scholar] [CrossRef]
- vanderEst, A.; Prisner, T.; Bittl, R.; Fromme, P.; Lubitz, W.; Mobius, K.; Stehlik, D. Time-resolved X-, K-, and W-band EPR of the radical pair state P•+700A•-1 of photosystem I in comparison with P•+865Q•- A in bacterial reaction centers. J Phys Chem B 1997, 101, 1437–1443. [Google Scholar] [CrossRef]
- Zech, S. G.; Hofbauer, W.; Kamlowski, A.; Fromme, P.; Stehlik, D.; Lubitz, W.; Bittl, R. A structural model for the charge separated state P•+700A•-1 in photosystem I from the orientation of the magnetic interaction tensors. J Phys Chem B 2000, 104, 9728–9739. [Google Scholar] [CrossRef]
- Poluektov, O. G.; Paschenko, S. V.; Utschig, L. M. Spin-dynamics of the spin-correlated radical pair in photosystem I. Pulsed time-resolved EPR at high magnetic field. Phys Chem Chem Phys 2009, 11, 6750–6756. [Google Scholar] [CrossRef]
- Poluektov, O. G.; Utschig, L. M.; Schlesselman, S. L.; Lakshmi, K. V.; Brudvig, G. W.; Kothe, G.; Thurnauer, M. C. Electronic structure of the P700 special pair from high-frequency electron paramagnetic resonance spectroscopy. J Phys Chem B 2002, 106, 8911–8916. [Google Scholar] [CrossRef]
- Niklas, J.; Epel, B.; Antonkine, M. L.; Sinnecker, S.; Pandelia, M. E.; Lubitz, W. Electronic Structure of the Quinone Radical Anion A•-1 of Photosystem I Investigated by Advanced Pulse EPR and ENDOR Techniques. J Phys Chem B 2009, 113, 10367–10379. [Google Scholar] [CrossRef]
- Savitsky, A.; Niklas, J.; Golbeck, J. H.; Mobius, K.; Lubitz, W. Orientation Resolving Dipolar High-Field EPR Spectroscopy on Disordered Solids: II. Structure of Spin-Correlated Radical Pairs in Photosystem I. J Phys Chem B 2013, 117, 11184–11199. [Google Scholar] [CrossRef]
- Schlodder, E.; Falkenberg, K.; Gergeleit, M.; Brettel, K. Temperature dependence of forward and reverse electron transfer from A, the reduced secondary electron acceptor in photosystem I. Biochemistry-Us 1998, 37, 9466–9476. [Google Scholar] [CrossRef]
- Chorney, W. S., N.; Crespi, H. L.; Katz J., J. The growth of algae in deuterium oxide. Biochem. Biophys. Acta 1960, 37, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Crespi, H. L. A., S.; Katz J., J. Culture of algae and other micro-organisms in deuterium oxide. Nature 1959, 184, 729–730. [Google Scholar] [CrossRef] [PubMed]
- Crespi, H. L. C., S.; Uphaus, R. A.; Katz, J. J. Cultivation of microorganisms in heavy water. Ann. N. Y. Acad. Sci 1960, 84, 648–666. [Google Scholar] [CrossRef] [PubMed]
- Poluektov, O. G.; Niklas, J.; Utschig, L. M. Spin-Correlated Radical Pairs as Quantum Sensors of Bidirectional ET Mechanisms in Photosystem I. J Phys Chem B 2019, 123, 7536–7544. [Google Scholar] [CrossRef]
- Bittl, R.; Weber, S. Transient radical pairs studied by time-resolved EPR. BBA-Bioenergetics 2005, 1707, 117–126. [Google Scholar] [CrossRef]
- Bittl, R.; Zech, S. G. Pulsed EPR spectroscopy on short-lived intermediates in photosystem I. BBA-Bioenergetics 2001, 1507, 194–211. [Google Scholar] [CrossRef]
- Santabarbara, S.; Kuprov, I.; Fairclough, W. V.; Purton, S.; Hore, P. J.; Heathcote, P.; Evans, M. C. W. Bidirectional electron transfer in photosystem I: Determination of two distances between P+700 and A-1 in spin-correlated radical pairs. Biochemistry-Us 2005, 44, 2119–2128. [Google Scholar] [CrossRef]
- Berthold, T.; von Gromoff, E. D.; Santabarbara, S.; Stehle, P.; Link, G.; Poluektov, O. G.; Heathcote, P.; Beck, C. F.; Thurnauer, M. C.; Kothe, G. Exploring the Electron Transfer Pathways in Photosystem I by High-Time-Resolution Electron Paramagnetic Resonance: Observation of the B-Side Radical Pair P+700A-1B in Whole Cells of the Deuterated Green Alga Chlamydomonas reinhardtii at Cryogenic Temperatures. J Am Chem Soc 2012, 134, 5563–5576. [Google Scholar] [CrossRef]
- Santabarbara, S.; Reifschneider, K.; Jasaitis, A.; Gu, F. F.; Agostini, G.; Carbonera, D.; Rappaport, F.; Redding, K. E. Interquinone Electron Transfer in Photosystem I As Evidenced by Altering the Hydrogen Bond Strength to the Phylloquinone(s). J Phys Chem B 2010, 114, 9300–9312. [Google Scholar] [CrossRef]
- Santabarbara, S.; Kuprov, I.; Hore, P. J.; Casal, A.; Heathcote, P.; Evans, M. C. W. Analysis of the spin-polarized electron spin echo of the [P+700A-1] radical pair of photosystem I indicates that both reaction center Subunits are competent in electron transfer in cyanobacteria, green algae, and higher plants. Biochemistry-Us 2006, 45, 7389–7403. [Google Scholar] [CrossRef]
- Santabarbara, S.; Kuprov, I.; Poluektov, O.; Casal, A.; Russell, C. A.; Purton, S.; Evans, M. C. W. Directionality of Electron-Transfer Reactions in Photosystem I of Prokaryotes: Universality of the Bidirectional Electron-Transfer Model. J Phys Chem B 2010, 114, 15158–15171. [Google Scholar] [CrossRef]
- Santabarbara, S.; Galuppini, L.; Casazza, A. P. Bidirectional Electron Transfer in the Reaction Centre of Photosystem I. J Integr Plant Biol 2010, 52, 735–749. [Google Scholar] [CrossRef]
- Santabarbara, S.; Heathcote, P.; Evans, M. C. W. Modelling of the electron transfer reactions in Photosystem I by electron tunnelling theory: The phylloquinones bound to the PsaA and the PsaB reaction centre subunits of PSI are almost isoenergetic to the iron-sulfur cluster Fx. BBA-Bioenergetics 2005, 1708, 283–310. [Google Scholar] [CrossRef]
- Muhiuddin, I. P.; Heathcote, P.; Carter, S.; Purton, S.; Rigby, S. E. J.; Evans, M. C. W. Evidence from time resolved studies of the P700/A radical pair for photosynthetic electron transfer on both the PsaA and PsaB branches of the photosystem I reaction centre. Febs Lett 2001, 503, 56–60. [Google Scholar] [CrossRef]
- Utschig, L. M.; Thurnauner, M. C.; Tiede, D. M.; Poluektov, O. G. Low-temperature interquinone electron transfer in photosynthetic reaction centers from Rhodobacter sphaeroides and Blastochloris viridis: Characterization of QB- states by high-frequency electron paramagnetic resonance (EPR) and electron-nuclear double resonance (ENDOR). Biochemistry-Us 2005, 44, 14131–14142. [Google Scholar] [CrossRef]
- Poluektov, O. G.; Utschig, L. M.; Dubinskij, A. A.; Thurnauer, M. C. Electron transfer pathways and protein response to charge separation in photosynthetic reaction centers:: Time-resolved high-field ENDOR of the spin-correlated radical pair P865+QA-. J Am Chem Soc 2005, 127, 4049–4059. [Google Scholar] [CrossRef]
- Poluektov, O. G.; Utschig, L. M.; Dalosto, S.; Thurnauer, M. C. Probing local dynamics of the photosynthetic bacterial reaction center with a cysteine specific spin label. J Phys Chem B 2003, 107, 6239–6244. [Google Scholar] [CrossRef]
- Utschig, L. M.; Ohigashi, Y.; Thurnauer, M. C.; Tiede, D. M. A new metal-binding site in photosynthetic bacterial reaction centers that modulates QA to QB electron transfer. Biochemistry-Us 1998, 37, 8278–8281. [Google Scholar] [CrossRef]
- Utschig, L. M.; Greenfield, S. R.; Tang, J.; Laible, P. D.; Thurnauer, M. C. Influence of iron-removal procedures on sequential electron transfer in photosynthetic bacterial reaction centers studied by transient EPR spectroscopy. Biochemistry-Us 1997, 36, 8548–8558. [Google Scholar] [CrossRef]
- Utschig, L. M.; Chen, L. X.; Poluektov, O. G. Discovery of native metal ion sites located on the ferredoxin docking side of photosystem I. Biochemistry-Us 2008, 47, 3671–3676. [Google Scholar] [CrossRef]
- Parrett, K. G.; Mehari, T.; Warren, P. G.; Golbeck, J. H. Purification and Properties of the Intact P-700 and Fx-Containing Photosystem-I Core Protein. Biochim Biophys Acta 1989, 973, 324–332. [Google Scholar] [CrossRef]
- Warren, P. V.; Parrett, K. G.; Warden, J. T.; Golbeck, J. H. Characterization of a Photosystem-I Core Containing P700 and Intermediate Electron Acceptor-A1. Biochemistry-Us 1990, 29, 6545–6550. [Google Scholar] [CrossRef]
- Hellmich, J.; Bommer, M.; Burkhardt, A.; Ibrahim, M.; Kern, J.; Meents, A.; Muh, F.; Dobbek, H.; Zouni, A. Native-like Photosystem II Superstructure at 2.44 Å Resolution through Detergent Extraction from the Protein Crystal. Structure 2014, 22, 1607–1615. [Google Scholar] [CrossRef]
- Daboll, H. F. C. L.; Katz, J. J.. Mass cultivation of algae in pure heavy water. Biotechnol. Bioeng. 1962, 4, 281–297. [Google Scholar] [CrossRef]
- Bresgunov, A. Y.; Dubinskii, A. A.; Krimov, V. N.; Petrov, Y. G.; Poluektov, O. G.; Lebedev, Y. S. Pulsed EPR in 2-mm Band. Appl Magn Reson 1991, 2, 715–728. [Google Scholar] [CrossRef]
- Harvey, S. M.; Wasielewski, M. R. Photogenerated Spin-Correlated Radical Pairs: From Photosynthetic Energy Transduction to Quantum Information Science. J Am Chem Soc 2021, 143, 15508–15529. [Google Scholar] [CrossRef]
- Closs, G. L.; Forbes, M. D. E.; Norris, J. R. Spin-Polarized Electron-Paramagnetic Resonance-Spectra of Radical Pairs in Micelles - Observation of Electron-Spin Spin Interactions. J Phys Chem-Us 1987, 91, 3592–3599. [Google Scholar] [CrossRef]
- Poluektov, O. G.; Utschig, L. M. Quantum Sensing of Electron Transfer Pathways in Natural Photosynthesis Using Time-Resolved High-Field Electron Paramagnetic Resonance/Electron-Nuclear Double Resonance Spectroscopy. J Phys Chem B 2021, 125, 4025–4030. [Google Scholar] [CrossRef]
- Epel, B.; Niklas, J.; Antonkine, M. L.; Lubitz, W. Absolute signs of hyperfine coupling constants as determined by pulse ENDOR of polarized radical pairs. Appl Magn Reson 2006, 30, 311–327. [Google Scholar] [CrossRef]
- Hore, P. J.; Mouritsen, H. The Radical-Pair Mechanism of Magnetoreception. Annu Rev Biophys 2016, 45, 299–344. [Google Scholar] [CrossRef]
- Wiltschko, R.; Wiltschko, W. Magnetoreception in birds. J R Soc Interface 2019, 16. [Google Scholar]
- Bindra, J. K.; Niklas, J.; Jeong, Y.; Jasper, A. W.; Kretzschmar, M.; Kern, J.; Utschig, L. M.; Poluektov, O. G. Coherences of Photoinduced Electron Spin Qubit Pair States in Photosystem I. J Phys Chem B 2023, 127, 10108–10117. [Google Scholar] [CrossRef]
- Jeong, Y.; Bindra, J. K.; Niklas, J.; Utschig, L. M.; Poluektov, O. G.; Jasper, A. W. Theoretical examination of nuclear spin diffusion in light-induced spin coherences in photosystem I. Appl Phys Lett 2024, 124. [Google Scholar]
- Redding, K. E. van der Est, A. The directionality of electron transport in Photosystem I. In Photosystem I: The Light-Driven Plastocyanin:Ferredoxin Oxidoreductase, Golbeck, J. H. Ed.; Vol. 24; Springer, 2006; pp 413-437.
- Joliot, P.; Joliot, A. In vivo analysis of the electron transfer within Photosystem I: Are the two phylloquinones involved? Biochemistry-Us 1999, 38, 11130–11136. [Google Scholar] [CrossRef]
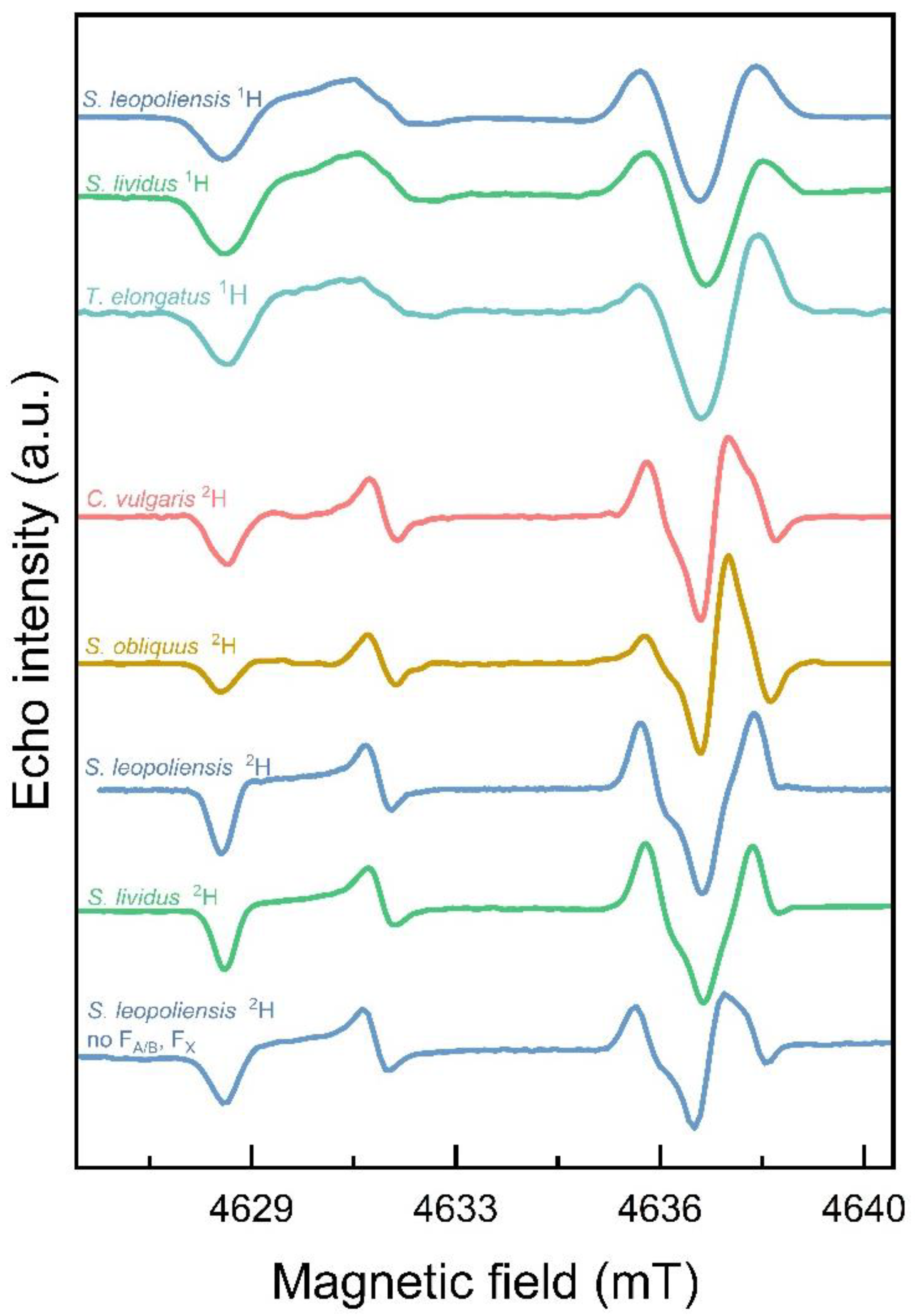
Figure 3.
High-frequency D-band (130 GHz) pulsed EPR spectra of the SCRPs recorded in PSI from different biological species. T=100 K, DAF = 1 μs.
Figure 3.
High-frequency D-band (130 GHz) pulsed EPR spectra of the SCRPs recorded in PSI from different biological species. T=100 K, DAF = 1 μs.
Scheme 1.
Schematic presentation of relative redox potential of acceptors A1A and A1B in respect to Fx. Note, that lower redox midpoint potential means a more reducing potential. Dashed arrow indicates recombination reaction to P700+. Left: cyclic electron transfer in the A branch. Middle: cyclic electron transfer in the B branch. Right: cyclic electron transfer in both A and B branches.
Scheme 1.
Schematic presentation of relative redox potential of acceptors A1A and A1B in respect to Fx. Note, that lower redox midpoint potential means a more reducing potential. Dashed arrow indicates recombination reaction to P700+. Left: cyclic electron transfer in the A branch. Middle: cyclic electron transfer in the B branch. Right: cyclic electron transfer in both A and B branches.
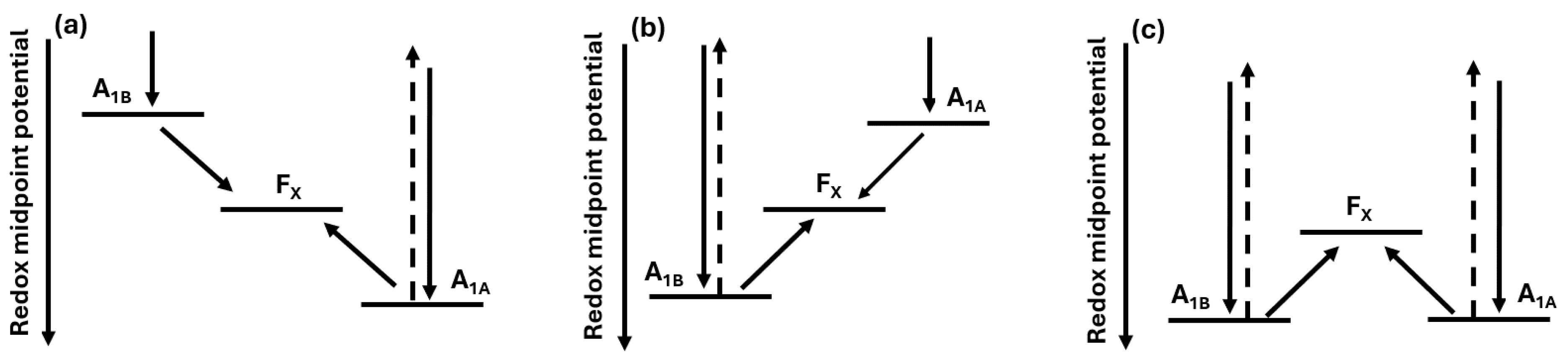
Figure 4.
Continuous wave (cw) X-band (9.5 GHz) EPR spectra of various PSI samples. Samples were frozen in the dark, cooled down to 10 K in the cavity of the EPR spectrometer and subsequently illuminated with a white light LED. Microwave power 3 mW, modulation amplitude 1.2 mT.
Figure 4.
Continuous wave (cw) X-band (9.5 GHz) EPR spectra of various PSI samples. Samples were frozen in the dark, cooled down to 10 K in the cavity of the EPR spectrometer and subsequently illuminated with a white light LED. Microwave power 3 mW, modulation amplitude 1.2 mT.
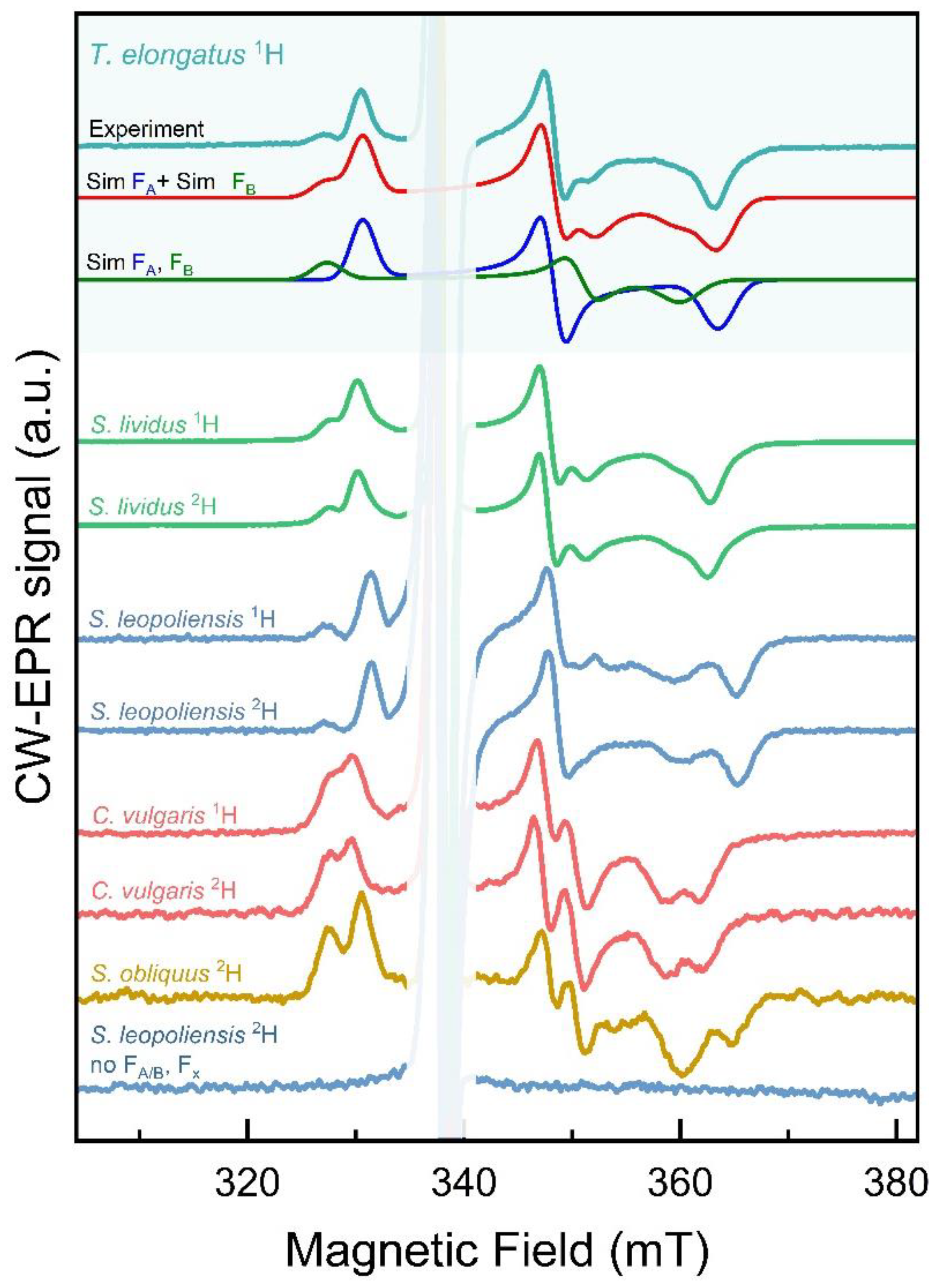

Table 1.
g-values of the [4Fe-4S] cluster FA and FB, and their relative contribution to the EPR spectrum FA/FB.
Table 1.
g-values of the [4Fe-4S] cluster FA and FB, and their relative contribution to the EPR spectrum FA/FB.
| FA | FB | FA/Fb | |||||
|---|---|---|---|---|---|---|---|
| gx | gy | gz | gx | gy | gz | ||
| T. elongatus1H | 2.048 | 1.945 | 1.863 | 2.069 | 1.930 | 1.881 | 2:1 |
| S. lividus1H | 2.050 | 1.946 | 1.865 | 2.068 | 1.933 | 1.879 | 2:1 |
| S. lividus2H | 2.050 | 1.947 | 1.867 | 2.069 | 1.933 | 1.879 | 2:1 |
| S. leopoliensis1H | 2.044 | 1.942 | 1.854 | 2.070 | 1.921 | 1.880 | 4:1 |
| S. leopoliensis2H | 2.044 | 1.942 | 1.854 | 2.070 | 1.921 | 1.880 | 4:1 |
| C. vulgaris1H | 2.052 | 1.948 | 1.869 | 2.067 | 1.933 | 1.886 | 1:1 |
| C. vulgaris2H | 2.054 | 1.949 | 1.868 | 2.068 | 1.934 | 1.886 | 1:1 |
| S. obliquus2H | 2.046 | 1.945 | 1.853 | 2.067 | 1.932 | 1.880 | 1:1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



