Submitted:
17 June 2024
Posted:
20 June 2024
You are already at the latest version
Abstract
Nanovaccines are able to deliver antigen(s) and immunomodulator(s) directly to antigen-presenting cells (APCs) of the lymphatic system. Positive charges improve their uptake by APCs and often drive a Th-1 biased immune response so necessary against infectious diseases caused by intracellular pathogens and cancers. However, cationic compounds often display a dose-dependent toxicity so that systematic evaluation of cytotoxicity against cells in culture is required. After the rapid evolution of nanovaccines against SARS-Covid 2, mRNA vaccines gained much strength and have been designed against several infectious diseases, often relying on cationic components for mRNA protection. Once established the limiting concentration for the cationic compound, cationic nanovaccines perform well eliciting the desired Th-1 improved immune response in most cases. Other valuable approach found in the literature has been the construction of biocompatible nanoparticles (NPs) carrying cationic lipids, polymers or surfactants so that minimization of the concentration for the cationic component led to absence of toxicity. Against cancer, recent constructions of nanovaccines in situ after cancer cell disruption in the presence of adjuvants led to systemic presentation of multiple tumoral antigens yielding, in many instances, prevention of metastasis appearance and conversion of tumors non treatable by immunotherapy into treatable ones.
Keywords:
Cationic lipids
; cationic polymers
; cationic nanostructures
; Th1 improved responses
; dose-dependent cytotoxicity of cationic assemblies
; biocompatible polymers
; combinatory approaches in cancer nanovaccines
1. Why Nanovaccines?
Both microparticles (MPs) and nanoparticles (NPs) in efficient vaccines display an appropriate targeting of the innate immune system, mainly via prolonged delivery of antigens and immunomodulators to professional antigen-presenting cells (APCs) in the lymphatic system [1].
Particles traffic to the draining lymph-nodes (LN) depends on particle size; microparticles (500-2000 nm) mostly become associated with dendritic cells (DC) from the injection site whereas NPs (20-200 nm) including virus-like particles VLP (30 nm) were found both in LN-resident DC and macrophages; in vivo imaging studies in mice demonstrated that DC are strictly required for transport of MPs from the injection site to the LN in contrast to the free drainage of NPs to the LN [2,3,4,5,6].
Biodegradable MPs of poly (lactide-co-glycolide) with a cationic surface improved the delivery of adsorbed DNA into antigen-presenting cells after intramuscular (i.m.) injection [7]; MPs released intact and functional DNA over two weeks in vitro and higher levels of marker gene expression in vivo; after i.m. immunization, MPs not only significantly enhanced serum antibody responses in comparison to naked DNA but also the cationic MPs elicited potent cytotoxic T lymphocyte responses at a low dose [8,9]. The delivery of DNA vaccines using biodegradable polymeric magnetic NPs as opposed to MPs evidenced that DNA vaccines with NPs as carriers were preferentially taken up by dendritic cells inducing maximal levels of combined humoral and cellular immunity[10].
The comparison between MPs and NPs was recently reviewed for NPs and MPs of aluminium salts [11]. Aluminium (Al) compounds are well tolerated as adjuvants in human and veterinary prophylactic vaccines; their MPs are very polydisperse in water dispersion (sizes ranging from ~0.5 to 10 µm) generally inducing type 2 (Th2)-biased immune responses; importantly, nanometric sizes were shown to change these adjuvants to yield type 1 (Th1) responses, which can potentially be exploited for the development of vaccines in essential need of cellular Th1 immune responses [12,13]. For cancer immunotherapy, Al-NPs can elicit a robust and durable balanced (Th1/Th2) immune response and increase the number of cytotoxic T cells compared to Al-MPs, thereby inducing lysis of the cancer cells [11].
Regarding NPs charge, the cationic ones electrostatically combine with oppositely charged antigens or immunostimulants of interest such as peptides, proteins, nucleic acids, oligonucleotides, etc [14,15,16,17,18,19,20,21,22]. At a molecular level, cationic compounds such as lipids, surfactants and polymers [14,23,24,25,26,27] have been used to build cationic nanostructures and can elicit different immune responses depending on the affinity between the antigen and the adjuvant. For cationic NPs adsorbing the model antigen ovalbumin (OVA), four different cationic nanostructures were evaluated regarding the occurrence of humoral and cellular responses as illustrated in Figure 1. This figure was reproduced from reference [14]. The cationic NPs were based on 4 different compositions: 1) cationic lipid and ovalbumin [21]; 2) cationic polymer and ovalbumin [20]; 3) biocompatible neutral polymer, cationic lipid, cationic polymer, and ovalbumin [16]; 4) biocompatible neutral polymer, cationic lipid, and ovalbumin [15]. Combined with OVA, the cationic bilayer fragments and the cationic polymer elicited Th1 and Th2 immune responses, respectively. The other two cationic nanostructures yielded balanced Th1/Th2 responses (Figure 1). The affinity between OVA and the cationic polymer yielded entangled nanostructures from which OVA could not easily be released to the DC cytosol for presentation via major histocompatibility complex one (MHC-1); OVA degradation in the endosome would be required in order to expose OVA derived peptides by MHC-2. On the other hand, OVA loosely attached to cationic bilayer fragments possibly bypassed the endosome easily detaching from the adjuvant in the cytosol for presentation by MHC-1. High affinity between antigen and adjuvant implied in Th-2 response whereas low affinity implied in Th-1 response. Intermediate affinities in the other two cationic nanostructures shown in Figure 1 would yield both responses [16]. The main properties for novel adjuvants seem to be the nanometric size, the cationic character, and the biocompatibility, even if it is achieved in a low dose-dependent manner [14]. NPs shape has also been shown to be important [18,25,26,28,29,30,31]. For example, whereas micrometric liposomes of the cationic lipid dioctadecyldimethylammonium bromide (DODAB) as adjuvants carrying a recombinant protein of Mycobacterium leprae induced a large cellular (Th-1) immune response and poor humoral response [26], the use of nanometric DODAB bilayer fragments (DODAB BF) as adjuvants to carry outer membrane antigens of Neisseria meningitidis produced balanced cellular (Th-1) and humoral response (Th-2)[32].
In this review, the adjuvant properties of cationic nanostructures are overviewed in the context of vaccines for infectious diseases and cancer. In the next section, the role of size and positive charge in vaccines design is discussed.
2. How Particle Size and Positive Charge Drive the Immune Response?
NPs size significantly influences DC uptake, which is crucial for effective vaccine delivery and the initiation of an immune response. Endocytosis was defined as the main mechanism for cell uptake of NPs smaller than 0.5 μm in size whereas phagocytosis accounted for the uptake of particles larger than 0.5 μm [31]. In the lungs, alveolar macrophages ingested more efficiently liposomes of 0.65 -2 μm than smaller ones [33,34].
DC are able to promote differentiation of CD4+ T cells toward Th1 or Th2 subsets according to the different invaders of the host organism [35]. Th1 cells secrete pro-inflammatory cytokines, e.g. IFN-γ and TNF-α destined to promote elimination of invaders by cytolysis. Th2 cells secrete preferentially anti-inflammatory cytokines, such as interleukins IL-4, IL-5, IL-6 and IL-13 for activating the humoral immune responses destined to avert the pathogen. Gold NPs about 2 nm in diameter do not activate DCs due to their diffusion into cells and their non-membranous intracellular localization whereas NPs 12 nm in size enter DCs by endocytosis and elicit Th1 response [36]. OVA presented by 50 nm polymeric NPs contrasted to micrometric MPs for OVA presentation regarding the immune responses elicited in mice: Th1 for the nanometric particles [3,37] and Th2 for the micrometric ones [38,39].
Similar associations between size and type of immune response were also found for fungi [40]. The respiratory tree maintains sterilizing immunity against human fungal pathogens. Humans usually inhale filamentous molds and fungal pathogens that form small airborne conidia so that the respiratory tract maintains sterilizing immunity against them; the smaller conidia of fungus drives responses toward Th1 in contrast to hyphae that induce Th2 [41]. Curiously, nano-spikes on TiO2 MPs yielded Th1 responses in spite of the micrometric size of the particles [42].
NPs around 20-50 nm are particularly effective for receptor-mediated endocytosis, which is a selective and efficient method for internalizing materials at the cellular level. This size range allows NPs to interact optimally with cell surface receptors, which facilitates the encapsulation of the NPs within cellular vesicles and their subsequent transport into the cytoplasm. This pathway named cross-presentation is essential for delivering antigens directly to the cytoplasm of antigen-presenting cells, such as DC, where they can be processed and presented to T cells [43,44].
Particles within the nanoscale are readily internalized by cells via endocytosis, a process that encompasses various mechanisms including phagocytosis, macropinocytosis, and receptor-mediated endocytosis. The efficiency and pathway of uptake are highly dependent on the NPs size [31,43,45].
Larger NPs typically those over 100 nm, tend to be internalized via phagocytosis or macropinocytosis. These pathways are less selective and involve the engulfment of particles along with extracellular fluid, which can lead to less efficient antigen presentation. The larger size of these particles may result in their accumulation within endosomal or lysosomal compartments, potentially leading to degradation of the encapsulated antigens rather than their processing and presentation on MHC molecules [46,47].
Conversely, smaller NPs, particularly those under 20 nm, can diffuse more freely across cellular membranes but might bypass the endocytic pathway, leading to suboptimal delivery of antigens to the immune system. This can reduce the immunogenicity of the vaccine, since antigens can fail to reach the cellular compartments where they are needed to elicit a strong immune response [47,48].
Optimizing NPs size for vaccine delivery ensures antigen internalization into the cytoplasm of target cells, processing, and presentation in a manner that maximizes the immune response. This optimization directly influences the efficacy of vaccines, particularly those designed to induce cellular immunity, critical for fighting intracellular pathogens and cancer cells.
Once delivered into the cytoplasm, antigens are processed into peptides through the proteasome degradation pathway. These peptides are then transported to the endoplasmic reticulum, where they bind to MHC class I molecules and are presented on the cell surface. The exposed peptides on MHC class I molecules are recognized by T-cell receptors on cytotoxic T lymphocytes (CTLs), which, upon activation, proliferate and differentiate into effector cells, CD8+, capable of targeting and destroying cells presenting the same antigenic peptides [1,49,50].
NPs can be engineered to release antigens slowly, promoting sustained immune engagement and the development of long-lasting immune memory. This controlled release, combined with the particles' ability to mimic the size and shape of pathogens, enhances their uptake by DC, leading to more effective T cell activation [51]. This approach not only maximizes the vaccine's immunological impact but also facilitates dose sparing, important in scenarios where vaccine availability is limited.
The lymphatic vessels, primarily responsible for transporting fluids, cells, and other substances, have an endothelial lining that allows particles of certain sizes to pass through more effectively [52]. Particles in the range of 20-100 nm are ideal because they are small enough to enter the lymphatic capillaries, yet large enough to avoid rapid clearance by renal filtration. NPs sized 20-100 nm are optimally designed for lymph node targeting, as their dimensions allow them to enter the lymphatic system through interstitial spaces and travel efficiently to lymph nodes [48,53]. The NPs' ability to target lymph nodes and facilitate DC maturation and antigen presentation is illustrated by the development of a manganese-based nanodriver. This delivery system efficiently activates the stimulator of interferon genes (STING) pathway directly within the cytoplasm, promoting the production of type I interferon and enhancing the immune response against glioblastoma; the incorporation of doxorubicin in the NPs further amplifies this effect, demonstrating a potent synergistic interaction between the drug and the antigen that enhances tumor suppression [54].
Lipid nanoparticle-mediated lymph node-targeting delivery of mRNA vaccines showcases significant advancements in cancer immunotherapy, particularly through the enhancement of CD8+ T cell responses. Novel lipid NPs, named 113-O12B, were designed to target lymph nodes directly without the need for active targeting ligands. NPs demonstrated a superior ability to deliver mRNA encoding full-length ovalbumin (OVA) and TRP-2 peptide antigens directly to lymph nodes, leading to significantly enhanced CD8+ T cell activation compared to conventional lipid NPs. NPs' targeting capability resulted in potent therapeutic and protective effects against melanoma models such as B16F10 melanoma. Importantly, the study found that 113-O12B NPs, when used in conjunction with anti-PD-1 therapy, showed a complete response in a substantial proportion of cases, substantially improving cancer vaccine efficacy. The effective delivery of mRNA to DCs in lymph nodes not only minimized side effects associated with non-targeted delivery systems but also maximized the immunogenic potential of the vaccines by enhancing antigen presentation directly where it is most effective for initiating robust immune responses [55].
Once antigens are captured by DCs, the DCs degrade the antigens into peptide fragments. The antigen peptides are carried to the surface of DCs, and thus presented to naïve helper CD4+ T cells and naïve cytotoxic CD8+ T cells in lymph nodes. The naïve T cells differentiate into effector T cells. The effector helper CD4+ T cells comprise different subsets, such as Th1, Th2 and Treg. Th1 cells mainly produce interleukin (IL)-2, IL-12, interferon (IFN)-γ, tumor necrosis factor (TNF)-α, and others, which trigger cellular immunity by activating cytotoxic CD8+ T cells. Th2 cells mainly secrete IL-4, IL-5, IL-6, IL-10, IL-13, and other cytokines, which generally induce humoral immunity by activating B cells to produce antibodies. These cytokines tend to activate their own T cell subset and suppress the other T cell subsets. For instance, IFN-γ secreted by Th1 cells simultaneously stimulates the Th1 cell development and inhibits Th2 cell production. Treg cells generally release transforming growth factor (TGF)-β, IL-10, and other factors, which downregulate effector T cells, maintain tolerance to self-antigens, and prevent autoimmune diseases. Thus, different cytokine profiles secreted by effector helper CD4+ T cells determine the directions of the subsequent adaptive immunity, which include stimulating B cells to produce antibodies, activating cytotoxic CD8+ T cells and inducing immune tolerance [49].
The positive charge on cationic NPs is typically conferred by the presence of amine groups [56,57,58] or quaternary ammonium moieties [59,60,61,62]. Particularly important are the cationic lipids or surfactants such as DODAB and cetyltrimethylammonium bromide (CTAB) [63,64,65,66]. Synthetic or natural polymers such as PDDA, PEI, poly(lactide-co-glycolide) acid (PLGA) and chitosan [67,68,69,70,71,72] allow a broad range of sizes for producing cationic NPs in combinations with DODAB; silica and DODAB or polymers and DODAB can also yield a range of controllable nanometric sizes [73,74]. The cationic nature of these NPs enhances their interaction with negatively charged biological molecules and cellular membranes, making them particularly effective for delivering genetic material and antigens into cells. Their positive charge facilitates strong interactions with negatively charged cellular membranes, which improves cellular uptake of the vaccine components such as antigens and other immune stimulators [75]. Furthermore, loaded mRNA and protein antigens become protected from degradation in the extracellular environment [76,77]. This protective effect increases the likelihood that a higher quantity of intact antigen reaches the target cells, enhancing the potential for a strong immune response [78]. Combinations between cationic lipids and polymers also allow interesting developments such as the cationic supported bilayers on polystyrene sulfate nanoparticles [79] or the hybrid cationic lipid-biocompatible polymer NPs with the polymer selected as PLGA or PMMA [65,80,81].
By facilitating the delivery of antigens directly into the cytoplasm [82,83], cationic NPs enable the antigens to be processed and presented on MHC class I molecules [83]. This pathway is essential for the activation of CD8+ cytotoxic T lymphocytes, which are critical for killing infected cells and tumor cells. The ability to enhance this pathway is particularly beneficial in vaccines aimed at eliciting strong cellular immunity, as required against intracellular bacteria and cancer [49]. Additionally, the interaction of cationic NPs with immune cells can lead to the activation of various toll-like receptors (TLRs), which play a critical role in the innate immune system [58,84]. Cationic NPs act similarly to danger signals coming from infective agents such as agonists of molecular patterns recognition receptors (MPRR) as lipopolysaccharides (LPS) from Gram-negative bacteria and lipoteichoic acids from Gram-positive bacteria.
NPs positive charge can induce the activation of the NALP3 inflammasome, a main component of the innate immune system that regulates the production of pro-inflammatory cytokines like IL-1β. This activation can occur via the NPs' interaction with cellular components that trigger the inflammasome pathway [58,85,86]. Moreover, inherent properties of cationic NPs can lead to the rupture of cell membranes [20,83,87,88]. This disruption can act as a danger signal, alerting the immune system to potential threats by releasing intracellular contents that are recognized as damage-associated molecular patterns (DAMPs). These DAMPs then stimulate further immune responses, attracting more immune cells to the site of vaccine administration [83].
Similarly to cationic NPs, assemblies of cationic lipids in vivo are potent inducers of pro inflammatory cytokines such as those required for the cellular immune response and combat of intracellular pathogens [18,21,26,89]. The use of cationic lipids in vivo is limited not only by their pro-inflammatory properties but also by their poor stability [90,91]. In vitro, there is a strong affinity between serum proteins and cationic lipid carriers driven by the electrostatic attraction and the hydrophobic effect [20,92,93]. In vivo, the interaction of the cationic carrier with albumin and immune globulins leads to opsonization and rapid removal of the protein-surrounded cationic carriers from the circulation by the macrophages of the reticule endothelial system (RES)[94]. Thus, most formulations based on cationic nano-carriers are delivered subcutaneously, intradermally, intratumorally or via intraperitoneal routes in order to avoid the inflammatory toxicity and the elimination by RES associated to the cationic carriers administered by the parenteral route [95]. In order to disguise the inflammatory properties of cationic lipid assemblies and their cargo we employed a dual strategy to obtain controllable inflammation: the anti-inflammatory drug indomethacin complexed with the cationic lipid carrier was encapsulated by carboxymethylcellulose (CMC) [96]; this approach still requires further evaluation in vivo for antigens delivery by the parenteral route. Similarly, anti-tubercular drug rifampicin was complexed with cationic lipid nano-assemblies and the complexes surrounded by CMC and delivered in vitro to micobacteria achieving 100% reduction in micobacteria cell viability [97]; these assemblies should be further tested in vivo aiming at simultaneous drug and vaccine delivery against micobacteria.
Cationic NPs can also facilitate the escape of encapsulated antigens from endosomes and lysosomes to the cytosol. By destabilizing the membranous barriers of endosomes, the NPs prevent the degradation of antigens, ensuring their proper processing and presentation on MHC class I molecules, which is essential for the activation of CD8+ T cells [98,99,100]. Lysosomal escape enhances the presentation of antigens to the immune system, leading to a more robust activation of T cells [101]. The ability of cationic NPs to bypass the degradation pathways and present antigens directly to the immune system is a pivotal factor in their effectiveness as vaccine adjuvants [102]. The activation of the immune system by cationic NPs through these mechanisms—NALP3 inflammasome activation, cell membrane rupture, and lysosomal escape—highlights their potential as powerful tools in vaccine development. These properties not only enhance the immediate immune response but also contribute to the long-term efficacy of vaccines by ensuring the generation of memory cells [103,104,105].
The physicochemical properties of cationic NPs—such as colloidal stability, spontaneous interaction with negatively charged antigens, enhanced cellular uptake, and adjuvant effects—make them exceptionally useful in the development of efficient and effective vaccine formulations. Stability is relevant for effective immunization strategies. Firstly, the colloidal stability of these nanoparticles ensures that they remain dispersed in solution, preventing aggregation that could diminish their efficacy and safety. This stability is particularly important for maintaining the integrity and uniformity of vaccine formulations during storage and transport [15,20,21]. Furthermore, the versatility of cationic nanoparticles allows for the co-delivery of antigens and adjuvants within the same particle. This co-delivery can synchronize antigen and adjuvant presentation to the immune system, thus maximizing the immune response [21,106].
Figure 2 illustrates the routes followed by cationic NPs upon their interaction with dendritic cells (DC) that culminate in antigen presentation to the immune system.
In our laboratory we have been developing biomimetic cationic NPs and nanostructures over the last 30 years; these cationic nanostructures still need further testing as adjuvants for vaccines. Among them some examples are the supported cationic bilayers on silica [22,107,108,109] or polystyrene sulfate [19,64,73,74,79], the hybrid lipid-polymer NPs based on the layer-by-layer approach [110,111] or on the interaction forces driving formation of the cationic lipid-biocompatible polymer nanostructure [15,81,112] or cationic polymer-biocompatible polymer NPs [16,81,113] or on the electrostatic attraction between cationic polymer and antigen [20]. The bilayer fragments (BF) made of synthetic lipids such as the anionic sodium dihexadecyl phosphate (DHP)[114] or the cationic DODAB [115] were first described in the eighties and nineties and used in drug and vaccine delivery since then [18,96,116,117]. DODAB BFs are one of the most explored nanostructures (60-80 nm) to carry a variety of antigens [21,24,28,118]. In this review, we discuss seminal and recent advances in applications of cationic nanostructures as immunoadjuvants for vaccine design against infectious diseases and cancer. Emphasis will be placed on nanoadjuvants able to elicit Th1 responses.
3. Cationic Nanostructures in Vaccine Design Against Infections
Biomimetic NPs [109,118,119] and virus-like NPs (VLP) used in the design of vaccines mimic native viruses but lack viral genome being similarly taken up by the immune system [120]. Figure 3 illustrates the processing of VLPs by the immune system [120].
DNA manipulation using recombinant DNA technology has been intimately associated to our understanding of microorganisms including the pathogens that cause many infectious diseases in humans [121]. Scaling-up of antigen production for subunit vaccines design can readily be achieved from techniques that enable us to read, copy and manipulate DNA sequences.
A good example is the development of vaccines against the human papilloma virus (HPV). Three prophylactic HPV vaccines based on the recombinant DNA technology were approved; purified L1 protein self-assembled as HPV empty shells and induced production of specific neutralizing antibodies [122]. One should notice that not all antigens self-assemble to yield NPs in dispersion. In many cases the vaccine subunit design requires also adjuvant(s) and/or immunostimulator(s).
Figure 4.
The amplification of L1 coding region of human papilloma virus (HPV) using recombinant DNA technology for production of prophylactic vaccines. Eukaryotic systems include Cervarix, Gardasil, and Gardasil-9 vaccines whereas prokaryotic systems include Cecolin and Gelcolin vaccines. One should notice that the self-assembly of the L1 recombinant protein generated the virus like nanoparticle (HPV- VLP) [122].
Figure 4.
The amplification of L1 coding region of human papilloma virus (HPV) using recombinant DNA technology for production of prophylactic vaccines. Eukaryotic systems include Cervarix, Gardasil, and Gardasil-9 vaccines whereas prokaryotic systems include Cecolin and Gelcolin vaccines. One should notice that the self-assembly of the L1 recombinant protein generated the virus like nanoparticle (HPV- VLP) [122].
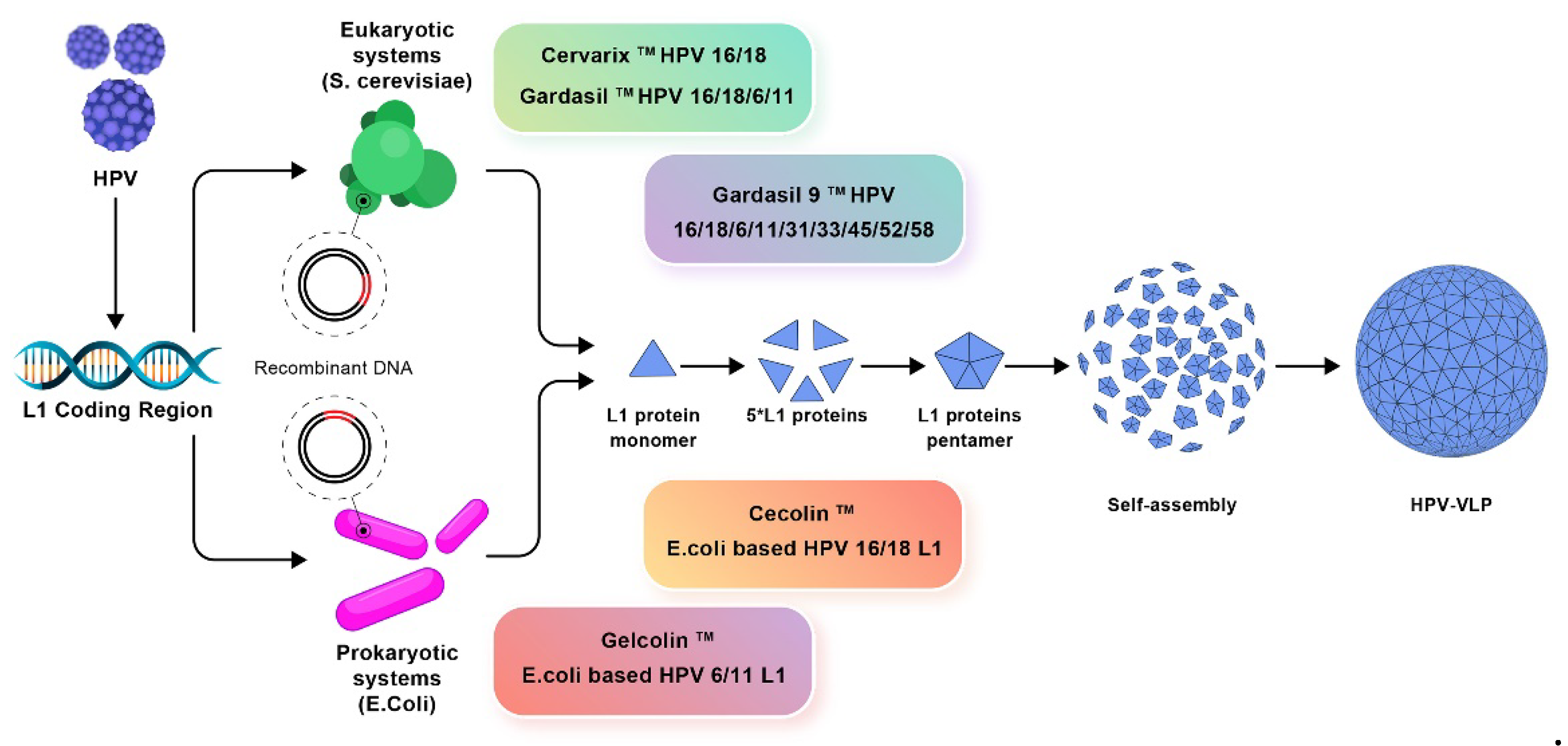
The technique of DNA cloning signaled the birth of genetic engineering, which allowed the facile transfer of genes among different biological species [123]. Thereby, genes encoding important recombinant proteins such as insulin and growth hormone were cloned and expressed in E. coli or Saccharomyces cerevisae; hundreds of pharmaceuticals such as therapeutic proteins and antibodies including the monoclonal ones became available in the market produced by the biotechnology industry [124]. Recombinant DNA technology indeed represents a valuable tool also in vaccines development which contributed to the fast design of COVID-19 vaccines applied during the recent pandemics.
Cationic NPs played a fundamental role in the delivery of mRNA vaccines, as highlighted by their successful use in COVID-19 vaccination [125,126]. Recently, the mRNA vaccines developed by Moderna and BioNTech/Pfizer against COVID-19, using lipid NPs (LNP) and mRNA technology have been approved for emergency use and are in clinical trials. The fast development of several COVID-19 vaccines was never seen before for any other vaccine in history [127]. Encapsulation not only shields the mRNA against premature degradation but also enhances its uptake into cells, primarily through endocytosis facilitated by the positive charge of the cationic NPs [127,128]. Cationic lipid nanoparticles (LNPs) have been used to encapsulate the mRNA molecules and protect them from degradation by nucleases before reaching their target cells. This strategy has been applied to other vaccines against viral infectious besides the COVID-19; for example, a vaccine against influenza (in phase 1 clinical trial) or HIV (in preclinical trial) [127].
The corona virus spike protein (S) has been widely used for vaccine development; prospective vaccines used strategies such as virus inactivation, live-vectoring, nucleic acid and recombinant antigen vaccines. Recombinant vaccine strategies were recently reviewed and discussed quoting designs such as cloning SARS-CoV-2 S gene for a DNA vaccine; obtaining a plasmid with SARS-CoV-2 S gene; obtaining S protein mRNA; using recombinant S-protein mimicking SARS-Cov-2 S protein; using non-replicating vectors carrying the SARS-CoV-2 S protein gene; using Virus-Like Particle without the virus genetic material [129].
Tuberculosis (TB) remains a formidable global health challenge, ranking as one of the top ten causes of death worldwide and the leading cause from a single infectious agent, surpassing even HIV/AIDS; in 2022, TB was responsible for 1.3 million deaths globally, including 167,000 people with HIV [130,131]. Caused by the bacterium Mycobacterium tuberculosis, TB primarily affects the lungs but can invade other parts of the body [131]. The disease is particularly prevalent in low-income countries, exacerbating public health and economic disparities [130]. Currently, the only widely used vaccine, Bacillus Calmette-Guérin (BCG), has been in use since 1921; although BCG is effective in preventing severe forms of TB in children, its efficacy in adults, particularly against pulmonary TB, which is the most common form and the main source of transmission, is variable and generally poor [132,133]. In this context, cationic adjuvants present a promising avenue for enhancing TB vaccine efficacy.
The efficacy of cationic liposomes, with or without cholesterol, was tested in various compositions as delivery systems for TB antigens; the goal was to identify cationic lipids with strong immunogenic properties and understand how cholesterol modifies the formulation's efficacy; cationic liposomes were prepared using the thin-film hydration method followed by sonication and designed to carry the FP antigen composed of Ag85B-ESAT6-Rv2034 and sized between 80-120 nm; cationic liposomes containing cholesterol increased liposome stability, cellular uptake, and DCs activation; cationic lipids improved interactions with DCs and enhanced the Th1-type immune response in a lipid-dependent manner; specifically, 1,2-dioleoyl-sn-glycero-3-ethylphosphocholine (EPC) demonstrated the highest immunogenicity, robust DCs activation, and a strong Th1 response measured as the number of CD4+ T/IFNγ+ especially when combined with cholesterol, despite maintaining moderate cytotoxicity; in contrast, DODAB exhibited higher cytotoxicity but was effective in dendritic cell activation, whereas DOTAP showed moderate uptake and activation, which were enhanced by cholesterol with minimal increase in cytotoxicity [134].
In our lab, we have been developing strategies to modulate DODAB dose-dependent cytotoxicity over the last decades [16,61,112,135,136]; for doing so we obtained combinations of DODAB with biocompatible polymers such as carboxymethylcellulose (CMC)[136] or poly (methyl methacrylate) (PMMA)[16] which indeed diminished substantially DODAB cytotoxicity against cells in culture. In addition, in other formulations, just a thin DODAB bilayer shell surrounded polymeric or inorganic NPs thereby reducing total DODAB dose in comparison to doses employed in formulations with DODAB large vesicles (LV) [137] or liposomes [30]. Importantly, in vaccine design with DODAB, its cytotoxicity against macrophages in culture could be reduced substantially by synthesizing hybrid PMMA/DODAB cationic NPs with excellent immunoadjuvant activity eliciting balanced Th1 and Th2 responses against ovalbumin as the model antigen [16]. Figure 5 illustrates the cytotoxicity of DODAB as large vesicles (LV), bilayer fragments (BF) or PMMA/DODAB NPs against macrophages and fibroblasts in culture [16].
The enhancement of the BCG vaccine using cationic adjuvants presents a promising strategy to overcome its limited efficacy and provide more robust and long-lasting protection against tuberculosis [138,139,140,141]. Cationic liposomes of DODAB/TDB/cholesterol were used to formulate the fusion protein (FP) HspX-PPE44-EsxV to enhance the efficacy of the BCG vaccine; the addition of cholesterol to these structures contributed to reduce liposomes’ sizes from 960 nm to 205 nm; the adjuvant effectively stimulated a robust Th1 immune response, evidenced by high levels of IFN-γ and IL-12, which are critical for protective immunity against Mycobacterium tuberculosis; additionally, the presence of IL-17 indicated a Th17 response, important for granuloma formation and neutrophil migration; mice primed with BCG and boosted with the DODAB/TDB/cholesterol/FP formulation exhibited the strongest immune responses, demonstrating that this strategy significantly enhanced BCG's efficacy; the high IgG2a/IgG1 ratio further confirmed the induction of a cellular immune response; there was superior protection against tuberculosis compared to BCG alone [138].
In another attempt to improve the efficacy of BCG vaccine, DODAB liposomes containing imiquimod (IMQ) a TLR7/8 agonist were employed to carry the FP DR2 composed of two antigens, Rv0572c and Rv3621c that targeted different stages of TB infection; DODAB/IMQ/DR2 vaccine induced strong CD4+ Th1 cell responses and robust IFN-γ+ CD4+ effector memory T cells (TEM) in mice; the vaccine also resulted in high levels of serum antibodies and cytokines, particularly IL-2+, CD4+, and CD8+ central memory T cells, indicating sustained immunity over 18 weeks; in vitro challenge experiments showed that splenocytes from vaccinated mice exhibited significant protection against M. tuberculosis, with low bacterial loads compared to control groups [140]. Usually, DODAB LV and liposomes are unilamellar and large varying in size from 500-1500 nm [30,137,142,143,144], therefore, they will not be directly drained to the lymph nodes and rather stimulate macrophages in situ at the injection site; they are micrometric in size.
Cationic liposomes of DODAB (480 nm in mean size) combined with two pattern recognition receptor agonists: monophosphoryl lipid A (MPLA) and trehalose 6,6'-dibehenate (TDB) were used to deliver a recombinant DNA antigen expressing the fusion protein CMFO, a recombinant fusion protein composed of four multistage antigens from Mycobacterium tuberculosis: CFP10, MPT64, Ag85B, and HspX; the inclusion of MPLA and TDB in the DODAB liposomes decreased the zeta potential but increased storage stability, allowing a slow and long-lasting release of DNA compared to DODAB liposomes alone; mice vaccinated with DODAB/MPLA/TBD/CMFO showed significantly higher levels of antigen-specific T cells and Th1-biased responses, including high levels of IFN-γ and TNF-α, compared to DODAB/CMFO and CMFO alone; DODAB/MPLA/TBD/CMFO formulation provided enhanced and persistent protection against Mycobacterium tuberculosis aerosol infection, comparable to that of the BCG vaccine [144].
The IC31 adjuvant, composed of the cationic peptide KLKL5KLK and the immunostimulatory oligodeoxynucleotide ODN1a, were employed to formulate the Ag85B-ESAT-6 fused antigen; the formulation Ag85B-ESAT-6/IC31 showed significant protection in mouse and guinea pig in an models of pulmonary tuberculosis; Ag85B-ESAT-6/IC31 induced strong antibody production in both BALB/C and C57BL/6 mice; it was also confirmed that the formulation elicited high levels of IFN-γ and low amounts of IL-4, IL-5, and IL-10, further supporting a Th1-biased response. Mice vaccinated with Ag85B-ESAT-6/IC31 showed a significant reduction in bacterial load in the lungs compared to unvaccinated mice and those vaccinated with Ag85B-ESAT-6 in Alum [145]. Against tuberculosis, it is essential to elicit a strong Th1 response because the infective agents are located inside the host cells.
The pressing challenge of developing a universal influenza vaccine lies in overcoming the limitations of current vaccines, which often fail to provide broad-spectrum protection due to the high antigenic variability of influenza viruses; in order to address this issue, it was developed an influenza universal vaccine based on the fused antigen NM2e; to this end, the adjuvant used was the cationic formulation composed of DODAB and poly(lactic acid) (PLA); the DODAB/PLA NPs were synthesized through a nanoprecipitation method, resulting in uniformly sized particles of 151 nm with a zeta potential of +30 mV; the NM2e@DODAB/PLA cationic nanovaccine exhibited a fourfold increase in antigen uptake by bone marrow-derived DC compared to the antigen alone accompanied by robust activation and maturation of DC, characterized by elevated expression of markers such as CD40, CD86, and MHC II, as well as increased production of cytokines including IFN-γ, TNF-α, and IL-6; in vivo studies demonstrated the prolonged presence of the antigen in lymph nodes for over 14 days post-immunization; the formulation induced high levels of specific IgG antibodies and strong cytotoxic T lymphocyte activity as indicated by increased secretion of granzyme B and IFN-γ; most notably, the NM2e@DDAB/PLA nanovaccine achieved over 90% cross-protection efficiency in mice challenged with both homosubtypic (H3N2) and heterosubtypic (H1N1) influenza viruses; NM2e@DDAB/PLA nanovaccine showed no significant toxicity from serum biochemical analyses and histopathological examinations [146]. This formulation for the influenza nanovaccine was based on the concept of combining a biocompatible polymer and a cationic compound as developed earlier in our group for PMMA/ DODAB hybrid NPs which yielded also similar results presenting the model antigen OVA [15,61,112,147]
In another attempt to produce more effective vaccine strategies to induce broad and protective immunity against influenza, the cationic lipid DOTAP was used to prepare DOTAP NPs to carry the recombinant influenza hemagglutinin (HA) antigen; DOTAP NPs significantly enhanced the recruitment of CD4 T cells producing IL-2 and IFN-γ compared to the squalene-based adjuvant AddaVax, even when AddaVax was combined with the TLR9 agonist CpG; DOTAP-adjuvanted vaccine elicited high frequencies of cytokine-producing CD4 T cells in both draining lymph nodes and spleens, with an approximately 9 to 12-fold increase in IL-2-producing cells and a 5.6 to 6.5-fold increase in IFN-γ-producing cells; the response to HA was broadly distributed across three identified CD4 T cell epitopes, with DOTAP NPs inducing a balanced and robust response to each epitope suggesting that DOTAP NPs generated a comprehensive immune response that is less susceptible to viral antigenic variation [148].
Malaria is one of several infectious diseases against which a new generation of CD8+ T cell-inducing vaccines are required; a number of heterologous vectors have been tested using DNA priming and boosting with vaccinia Ankara resulting in high levels of T cell response [149]. The infection is caused by Plasmodium parasites and transmitted by female Anopheles mosquitoes, remaining a major global health threat, particularly affecting children in tropical and subtropical regions [150]. The Mosquirix (RTS,S/AS01) vaccine, the most advanced against malaria, was approved by the World Health Organization for children in areas of moderate to high transmission but shows variable efficacy of 30-50% and requires multiple doses [151,152]. A self-amplifying RNA (samRNA) encoding Plasmodium falciparum reticulocyte binding protein homologue (5PfRH5) was used since this protein is crucial for the parasite's invasion of erythrocytes; the samRNA was stabilized by DODAB and DC-Chol cationic liposomes, which enhanced delivery and improved the immune response; liposomes were prepared by forming a dry lipid film, rehydrating it in HEPES buffer, and applying sonication to achieve sizes under 100 nm; the liposomes demonstrated high transfection efficiency and low cytotoxicity in vitro using Vero and HEK293T cells with GFP and luciferase reporters; in vivo, intradermal tattooing in mice resulted in significant GFP expression, and immunized mice produced antibodies that recognized native PfRH5 and inhibited P. falciparum growth; the use of cationic liposome-stabilized samRNA and tattooing for intradermal delivery are promising strategies in malaria vaccine development [153,154].
To improve malaria vaccine efficacy, a promising study explored the use of the cationic liposomal adjuvant CAF09, combined with full-length recombinant P. falciparum circumsporozoite protein (Pf rCSP); CAF09 consists of DODAB, monomycolyl glycerol (MMG), and polyinosinic:polycytidylic acid (Poly I:C), diameter around 400-500 nm [155]; the formulation induced robust antibody and CD8+ T-cell responses, providing comprehensive immunity; mice immunized with Pf rCSP-CAF09 developed high antibody titers and significant CD8+ T-cell responses, displaying durable sterilizing immunity against transgenic P. berghei expressing Pf CSP; the study highlighted the adjuvant's ability to elicit strong and long-lasting humoral and cellular immunity, addressing limitations of current vaccines primarily inducing antibody and CD4+ T-cell responses [156].
Cationic polymers are another important class of cationic materials used for preparing cationic nanoparticles, though their application is often limited by dose-dependent cytotoxicity, which necessitates careful dose minimization and optimization [20,88,135,157,158]. These polymers readily interact with oppositely charged proteins, facilitating their effective combination and delivery. Figure 8 illustrates the dose-dependent cytotoxicity of the cationic polymer PDDA against fibroblasts in culture and the use of biocompatible PMMA NPs hybridized with PDDA to circumvent its toxicity as reproduced from references [15,20,65].
Aiming to improve the critical issue of vaccine thermal instability, which hinders effective vaccination programs in resource-limited regions, a vaccine formulation used layer-by-layer coating of the Japanese encephalitis virus (JEV); polyethyleneimine (PEI) and silica layers improved the vaccine's thermostability as illustrated by Figure 7; the vaccine retained its potency at elevated temperatures (up to 42°C) for extended periods of time; the NPs, averaging 100 nm in diameter, demonstrated a robust thermal protective effect by forming a hydration layer that prevented thermal degradation, elicited strong immune responses (high serum IgG and neutralization antibody levels) and sustained cellular immunity depicted from IFN-γ secreting splenocytes; this innovative approach not only enhances the stability and efficacy of vaccines but also facilitates their distribution without the need for a cold chain, making it a promising solution for global vaccination efforts [159].
Glycoproteins derived from influenza A virus, herpes simplex virus type-2, and HIV-1 were combined with polyethyleneimine (PEI) to form dispersed nanoparticles with 800 nm as average size; when administered as a single intranasal dose, these PEI/antigen NPs induced robust protection against subsequent lethal viral infections, outperforming cholera toxin, a common experimental mucosal adjuvant; these NPs were efficiently taken up by antigen-presenting cells (APCs) in vitro and, in vivo, enhanced the trafficking of dendritic cells (DCs) to draining lymph nodes; nasal immunization with the recombinant envelope glycoprotein gp140 from HIV-1 carried by PEI resulted in high titers of antigen-specific IgA in vaginal lavages, demonstrating that nasal immunization can induce a systemic immune response; in Nlrp3-knockout mice, the PEI/gp140 complexes elicited a Th1-biased immune response, indicating that in normal mice, the NPs activated the Nlrp3 inflammasome towards a Th2-type response [83] . Additionally, four different PEI polymers—linear PEI (40 and 160 kDa) and branched PEI (25 and 750 kDa)—were combined with gp140 from HIV-1; post-immunization, all combinations elicited similar responses characterized by a moderate Th1 bias; when compared to alum, PEI showed significantly improved performance; the PEI-induced immune response featured an intermediate IgG1/IgG2a endpoint titer ratio, indicating a mixed Th1/Th2 immune response, corroborated by the cytokine profile analysis of antigen-restimulated splenocytes from mice immunized with gp140 and PEI; significant amounts of Th1 cytokines IL-2, TNF-α, and GM-CSF, as well as the Th2-associated cytokine IL-5, were detected [160].
Approaching the challenge of improving the delivery and immunogenicity of DNA vaccines, specifically against COVID-19, PEI covalently bound to cholesterol (PEI-CHOL) and mannose (PEI-CHOL-MAN) improved cellular uptake, reduced cytotoxicity, and target mannose receptors on APCs; the synthesis involved conjugating PEI with cholesteryl chloroformate, followed by reacting PEI-CHOL with D-mannose in the presence of sodium cyanoborohydride and acetic acid, resulting in NPs approximately 170 nm and 204 nm in size, with zeta potentials around +15 mV; the PEI-CHOL-MAN NPs demonstrated enhanced cellular uptake and significant stimulation of pro-inflammatory cytokine production, indicating improved dendritic cell maturation and a stronger immune response; the NPs exhibited improved biocompatibility with reduced cytotoxicity and demonstrated stability in various media, maintaining their integrity and protecting the encapsulated DNA from degradation; the incorporation of mannose and cholesterol enhanced targeted delivery, transfection efficiency, and safety, making this multifunctional PEI-based NPs formulation a promising approach for enhancing the efficacy of DNA vaccines against COVID-19 and potentially other infectious diseases [161].
PDDA was employed to enhance the delivery and immunogenicity of HIV-1 DNA vaccines using surface-engineered gold nanorods (Au NRs) as adjuvants; cationic molecules such as CTAB, PDDA and PEI modified the surface of gold nanorods, creating three distinct formulations: CTAB-Au NRs, PDDA-Au NRs, and PEI-Au NRs; the NPs had similar dimensions, approximately 15 nm × 60 nm, but differed in surface charge and stability, PDDA- and PEI-modified nanorods showed superior DNA adsorption capabilities and reduced cytotoxicity; PDDA- and PEI-Au NRs significantly improved both cellular and humoral immune responses in vivo compared to naked HIV-1 Env plasmid DNA; these formulations promoted dendritic cell maturation and enhanced T cell proliferation, indicating a strong immune activation; PDDAC-Au NRs induced a Th2-biased response, characterized by higher IgG1 production, while PEI-Au NRs facilitated efficient DNA release from endosomes due to the proton sponge effect; the strategic modification of gold nanorods not only enhanced the transfection efficiency but also provided a stable and biocompatible platform for DNA vaccine delivery; this approach demonstrated that surface-engineered gold nanorods can serve as promising DNA vaccine adjuvants, offering a versatile and effective means to improve the immunogenicity and efficacy of vaccines against HIV-1 [162].
Quaternized polysaccharides have emerged as a promising class of cationic polymers used as adjuvants in vaccine development, enhancing immunogenicity and offering robust protection against various pathogens [163,164]. In an effort to improve the efficacy of the Anthrax Vaccine Adsorbed (AVA), a study explored the use of fucoidan-quaternary chitosan NPs (FUC-HTCC NPs) as adjuvants; NPs, prepared via polyelectrolyte complexation, had sizes ranging from 100-300 nm and zeta potentials of ±25 mV; the FUC-HTCC NPs significantly enhanced both humoral and cellular immune responses, leading to higher IgG-anti-protective antigen titers and superior survival rates in mice challenged with anthrax toxin compared to traditional CpG oligodeoxynucleotides and AVA alone; NPs demonstrated low cytotoxicity, and excellent uptake by DCs [165].
Curdlan-chitosan conjugate NPs (C–O NPs) were used as mucosal adjuvants; NPs were synthesized via sonication with average size of 190 nm with a zeta potential of 16 mV; the C–O NPs demonstrated excellent biocompatibility and boosted macrophage phagocytosis and DCs activation in vitro; in vivo experiments showed that mice immunized with the H1N1 subunit antigen and C–O NPs produced high titers of HA-specific IgG and sIgA, indicating enhanced systemic and mucosal immune responses; this immune activation included the recruitment and activation of key immune cells such as macrophages and T cells and promoted a balanced Th1/Th2 response, essential for comprehensive protection; the study highlighted the potential of C–O NPs to improve vaccine stability and induce robust immunity and addressed the shortcomings of traditional influenza vaccines that often fail to elicit strong mucosal responses [166].
Subunit vaccines are commonly associated with low immunogenicity, necessitating innovative strategies to enhance their effectiveness; addressing this issue, researchers developed a novel carrier system using a complex of 2-hydroxypropyl-trimethylammonium chloride chitosan (HTCC) and amylose to entangle gold NPs (AuNPs) for improving the delivery and immunogenicity of SARS-CoV-2 subunit vaccines; the cationic adjuvants used were HTCC and amylose, which were combined with AuNPs to create positively charged nanocarriers; the NPs were prepared using a modified wet chemical method, resulting in particles with sizes ranging from 33 to 44 nm and a zeta potential of +21.1 mV; these HTCC/amylose/AuNPs demonstrated a high S protein loading capacity, low cytotoxicity, and excellent cellular uptake by RAW 264.7 cells; the immune response was significantly improved, eliciting high levels of specific IgG antibodies and enhancing T- and B-cell responses in mice compared to the free S protein; this new system offered a stable, efficient delivery method that significantly boosted the immunogenicity of subunit vaccines [167].
Taking advantage of chitosan's excellent mucoadhesive properties, hybrid DOTAP/chitosan NPs were developed to enhance the effectiveness of intranasal mRNA COVID-19 vaccines; DOTAP formed liposomes via the thin film hydration method, while chitosan was added to improve mucoadhesion by coating the liposomal surface; the optimized NPs, with a size of 360-590 nm and a positive zeta potential of +17 to +24 mV, demonstrated high mRNA encapsulation efficiency (~80.2%) and excellent biocompatibility; the formulation significantly enhanced the immune response by effectively delivering mRNA to the nasal mucosa, leading to high expression levels of the SARS-CoV-2 spike protein in target cells; in mice, this approach elicited strong local immune responses, with high levels of HA-specific IgG and secretory IgA antibodies in nasal and bronchoalveolar lavage fluids; the chitosan coating ensured prolonged retention and enhanced interaction with the nasal mucosa [168].
In a further effort to enhance the effectiveness of intranasal SARS-CoV-2 vaccines, the receptor-binding domain (RBD) of the SARS-CoV-2 spike protein was chemically conjugated to mannan; RBD-mannan conjugate was then assembled to polyarginine and 2',3'-cGAMP, a potent stimulator of the STING pathway, through electrostatic interactions; this process resulted in slightly cationic NPs approximately 74 nm in diameter; NPs demonstrated high mRNA encapsulation efficiency and excellent biocompatibility; intranasal administration in mice elicited robust local immune responses, with high levels of mucosal IgA and IgG in nasal and bronchoalveolar lavage fluids, and strong systemic immune responses, including elevated levels of RBD-specific neutralizing antibodies; the formulation also promoted a balanced Th1/Th2/Th17 cytokine response, crucial for effective immune protection [169].
4. Cationic Nanostructures in Vaccine Design against Cancer
Cancer nanovaccines should be efficiently delivered to the lymphatic system for induction of anti-tumor immunity; optimization of size, charge, colloidal stability and attached surface ligands should be performed beforehand aiming at the nanovaccine accumulation in lymph nodes and spleen [177]. Nanovaccines administered by subcutaneous or intramuscular routes can form depots at the injection site and need transportation by APCs to reach the LNs; using the peritumoral, intratumoral or intravenous routes allows capture of tumor antigens and antigen presentation in situ by local APCs or their delivery to the tumor-associated LNs [178]; chemo-, photothermal/photodynamic or radio-therapy destroys tumor cells releasing tumor antigens that are captured by the nanoadjuvants and delivered to tdLNs triggering personalized anti-tumor immunity [179].
The spleen, as the largest secondary lymphatic organ can also elicit a rapid and potent anti-tumor immunity; targeting the spleen with NPs could increase the efficacy of vaccines and cancer immunotherapy and treat intracellular infections including leishmaniasis, trypanosomiasis, splenic TB, AIDS, malaria, and hematological disorders; however, there is a macrophage barrier located in the red pulp of the spleen as an obstacle preventing the nanovaccine from reaching T or B lymphocytes; nanovaccines require also size optimization and albumin coating to achieve spleen accumulation. To achieve efficient spleen accumulation post i.v. administration, the size of nanovaccines has been optimized and surface ligands such as albumin were used to enhance their permanence in the circulation [180]. For example, the efficient delivery of clay-based nanovaccines to the mouse spleen promoted potent anti-tumor immunity for both prevention and treatment of lymphoma; the conventional subcutaneous (SC) vaccination normally induces limited anti-tumor immune responses with low therapeutic efficacy; clay-based nanovaccines directly delivered to the spleen intravenously (IV) induced strong anti-tumor immunity with high efficacy for tumor prevention and treatment; layered double hydroxide clay (LDH) sized from 77 to 285 nm and co-loaded with the model antigen OVA and CpG formed the nanovaccines where 215 nm for the size of LDH promoted antigen presentation by DCs and induced high spleen enrichment; in vivo, CO-LDH-215 induced potent anti-tumor immune responses and completely prevented the growth of E.G7-OVA tumor in the mouse model; IV injected CO-LDH-215 nanovaccine effectively delayed tumor growth in comparison to SC injected one, largely due to the direct and quick delivery of more nanovaccines to the spleen [180].
An obstacle preventing nanovaccines from reaching T or B lymphocytes in spleen or lymph nodes (LN) is a macrophage barrier located in the subcapsular sinus of the LN or the red pulp of the spleen [181]. Nanovaccines have to bypass the macrophage barrier and interact with B/T cell zones; the maximum achievable delivery efficiency was reported to be only 2% [182]. Improving the delivery efficiency of NPs to solid tumors will possibly be achievable through the engineering of multifunctional nanosystems or through manipulation of biological barriers. In this context, biomimetic NPs disguising their cargo might become very important.
Effective cancer immunotherapy was achieved from nanovaccines based on an injectable hydrogel formed with graphene oxide (GO) and polyethylenimine (PEI); mRNA of ovalbumin as the model antigen and adjuvants (R848)-laden nanovaccines were protected from degradation and directly delivered to lymph nodes; there was significant increase in antigen-specific CD8+ T cells and inhibition of tumor growth with only one treatment plus production of antigen specific antibody in the serum preventing the occurrence of metastasis [183].
Cancer immunotherapies seldom produce favorable outcomes in patients since most types of cancer can suppress antigen presentation, dendritic cell maturation, activation and infiltration of T lymphocytes in the tumor [184,185].
Nanomaterials have been reported to trigger immunogenic cell death (ICD) of cancer cells; dying cells then release tumor-associated antigens, pro-inflammatory cytokines (e.g., interleukin-1β (IL-1β) and IL-18) and damage-associated molecular patterns such as calreticulin, heat-shock proteins, adenosine triphosphate, and other biomarkers, all of them leading to APCs maturation plus activation and infiltration of T cells in tumor. Several nanomedicines with ICD-induced anticancer immunity have also major disadvantages associated to toxicity in vivo; the nanomaterial has to be biodegradable and toxicity in vivo has to be reduced [186].
Biodegradable, biocompatible, cancer-targeting, and ICD triggering nanomedicines are warranted. Alternatively, lysosomes can be targeted for triggering ICD; damaging lysosomes not only interrupts intracellular macromolecule digestion but also release hydrolytic enzymes such as cathepsins triggering ICD; this disrupts intracellular clearance; in addition, the leakage of inner lysosomal contents triggers cell death pathways such as apoptosis, necroptosis, and others [187].
Polycations of high-molecular-weight are cytotoxic due to lysosomal rupture often explained by the proton sponge effect; for example, protonation of PEI imine groups inside acidic intra-lysosomal environment would lead to lysosomal rupture [188]. Using a NPs pH sensor that was developed for pH measurements in the endosomal/lysosomal pathway, quantitative measurements of lysosomal pH as a function of PEI content showed that PEI did not induce change in lysosomal pH as previously suggested and quantification of PEI concentrations in lysosomes made it uncertain that the "proton sponge " effect would indeed be the dominant mechanism of PEI endosomal escape [189].
In our experience with PDDA, during dialysis of PDDA samples, we observed an influx of water to the dialysis bag revealing that the polymer induced an increase in osmotic pressure inside the bag (indeed PDDA is a very hydrophilic polymer). The increased influx of water in accordance with increased osmotic pressure due to the polymer enclosed in the lysosome would be a simple mechanism to explain lysosomal swelling and rupture culminating in cross- presentation of antigen carried by the polymer due to its presence in the cytosol. Anyway, PEI-mediated cytosolic delivery of oppositely charged antigens occurs by endosomal bursting as clearly shown for HeLa cells incubated with polyplexes carrying fluorescein isothiocyanate labeled oligo-deoxy nucleotides (FITC-ODNs) monitored by time-lapse microscopy; following internalization of the poly (ethylene imine) (PEI) polyplexes in endosomes, endosomal bursting took place releasing their ODNs cargo into the cytosol; after an initial appearance throughout the cytosol, the ODNs rapidly accumulated in the nucleus [69]. In order to activate potent cellular immune responses, cationic cancer nanovaccines should not only promote improved cellular uptake by DC but also lysosome escape, triggering cross-presentation to activate potent cellular immune response Th 1. Recalling Figure 1, DODAB BF carrying OVA induced strong Th1 response.
Using low molecular weight PEI (lwPEI), when lwPEI underwent self-assembly to form “high molecular weight” NPs, there was a dramatic enhancement in lysosome rupture, cell cytotoxicity, and immunogenic cancer cell death [188].
Similarly to cationic lipids, cationic polymers taken up by lysosomes such as PEI or PDDA display a dose-dependent cytotoxicity [16,20,135]. Interestingly, against tumors that do not respond to tumor antigens, an emerging approach has been turning a nonresponsive tumor into a responsive one by using nano-assemblies with PEI for rupturing lysosomes [190]. These positively charged nano-assemblies were constructed from low-molecular-weight branched PEI covalently bound to self-assembled peptides so that their uptake by cancer cells elicited immunogenic cells death (ICD) and anticancer immunity [190].
Immunotherapeutic agents commonly employed for cancer treatment are either antibodies, polypeptides or nucleic acids [191]; liposomes can protect both their hydrophobic or their hydrophilic cargos due to their two versatile compartments, the inner aqueous core and the hydrophobic microenvironment in the closed bilayer core [192].
Although excellent reviews on requirements for successful nanovaccines for cancer immunotherapy are available [193,194], just a few high-light emerging cationic nanovaccines [14,78,185,192,195]. During the past decades, cationic lipids or polymers in NPs have shown superior immunostimulatory properties as compared to their neutral and anionic analogues and recognized as potent inducers of antigen-specific T-cells; cationic NPs have offered clinically applicable vaccine formulation platforms [78]. For example, some cationic lipid-assisted nanoparticles (CLAN) fabricated with biocompatible and biodegradable block copolymer poly (ethylene glycol)-block-poly (lactic-co-glycolic acid) (PEG-b-PLGA) and cationic lipid packaged mRNA molecules, induced efficient internalization and translation in DC, yielded potent CD8+ T-cell activation and induced significant antitumor effect on the aggressive E·G7-OVA lymphoma model [196].
In another instance, the self-assembly of fluoroalkane PEI derivative (F-PEI) and mRNA encoding the model antigen ovalbumin (mRNA OVA), without additional adjuvants, could induce the maturation of DCs and trigger efficient antigen presentation, thereby eliciting anti-tumor immune responses that delayed the growth of established B16-OVA melanoma; this same approach to deliver MC38 neoantigen mRNA in combination with immune checkpoint blockade therapy, suppressed MC38 colon cancer and prevented tumor reoccurrence [197].
An anticancer nanovaccine improved cellular immune response by simultaneously delivering to the cytosol antigen and the immunostimulator polyIC RNA; the design involved the electrostatic attraction between PEI cationic moiety in a diblock co-polymer with an appropriate design and polyIC polyinosinic-polycytidylic acid (poly-IC); both antigen and polyIC underwent endosomal escape to the cytosol, synergistically enhancing antigen cross-presentation, co-stimulatory molecule expression, and cytokine production by dendritic cells; this approach enhanced CD8+ T cell responses relative to a mixture of antigen and poly-IC; hence inhibition of tumor growth in a mouse tumor model followed [198]. These cationic NPs allowed facile co-loading and dual-delivery of antigens and nucleic acid adjuvants; the di-block copolymers comprised a poly(ethylene glycol)-rich first block functionalized with reactive moieties for covalent conjugation of antigen via disulfide linkages, and a PEI moiety pH-responsive second block for electrostatic packaging of nucleic acids that also facilitated endosomal escape of associated vaccine cargo to the cytosol; endosomolytic NPs promoted the cytosolic co-delivery of poly-IC and protein antigen. Furthermore, poly-IC is an important co-stimulator used in the clinic to treat cancer; it is a double stranded RNA, which mimics viral genetic material and functions as a potent pattern recognition receptor ligand [199].
Figure 8 illustrates the co-loading of antigen and PolyIC using the di-block copolymer for obtaining NPs able to be taken up by the DCs by endocytosis and to escape the endosome for cross-presentation [198].
Figure 8.
Covalent combination of PEI moiety, antigen and poly-IC in an efficient cancer nanovaccine based on a di-block co-polymer able to elicit cross-presentation and efficient CD8+ T cell proliferation [198]. One should notice the disulfide binding of the antigen to the co-polymer. Reproduced with permission from Elsevier, Copyright 2022 [198].
Figure 8.
Covalent combination of PEI moiety, antigen and poly-IC in an efficient cancer nanovaccine based on a di-block co-polymer able to elicit cross-presentation and efficient CD8+ T cell proliferation [198]. One should notice the disulfide binding of the antigen to the co-polymer. Reproduced with permission from Elsevier, Copyright 2022 [198].
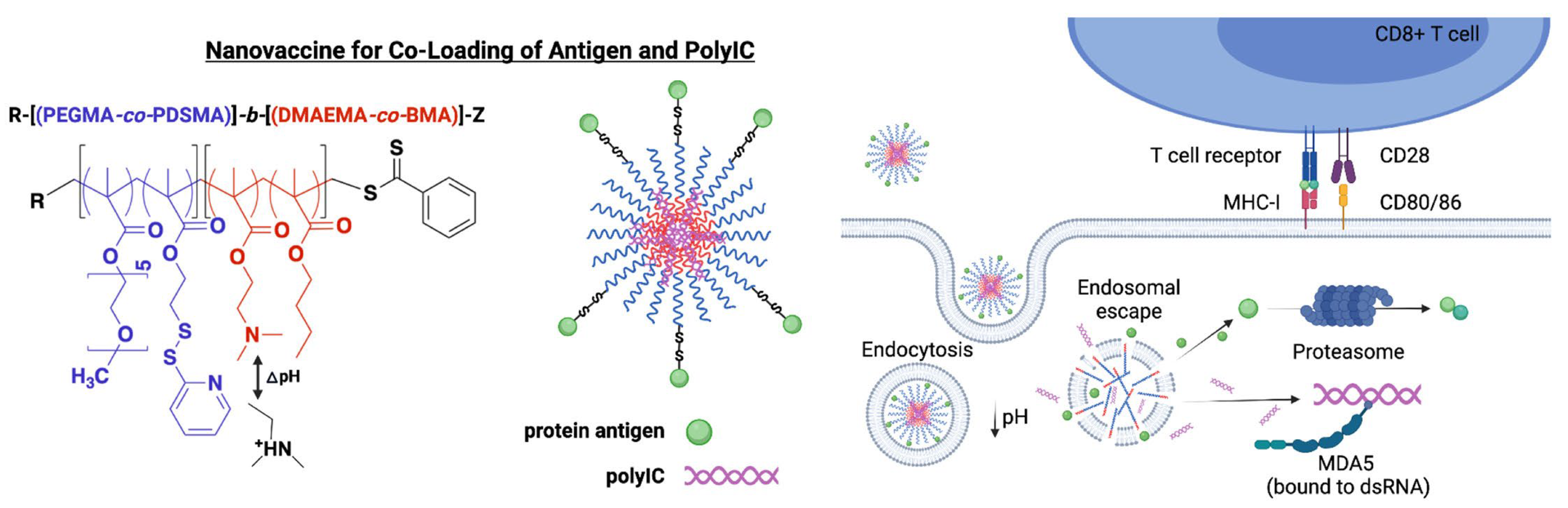
PolyIC is an agonist of Toll-like receptor 3 in the endosomal membrane and melanoma differentiation-associated protein 5 (MDA5) in the cytosol; polyIC protection from RNAase degradation could be achieved by electrostatic complexation with cationic polymers, such as (PEI) or poly-l-lysine (PLL); poly-IC complexed with PLL and stabilized with carboxymethylcellulose (poly-ICLC; trade mark Hiltonol®) has been evaluated in clinical trials and has been reported as stimulator of IFN-I production and/or enhanced T cell responses in humans; poly-ICLC activated MDA-5 more effectively than poly-IC since PLL promoted endosomal escape of poly-IC via the “proton sponge effect”[199].
PEI covalently bound to guanidine groups changed PEI’s cytotoxicity to innate immune activation; nanovaccines based on poly (phenyl biguanidine) stimulated dendritic cells, promoted their maturation via the TLR4 and NLRP3 pathways, and displayed robust in vivo immune activity significantly inhibiting tumor growth and extending survival in mice [200].
Antigens directly derived from cancer cell lysates or membranes have been used for cancer vaccines; their combination with cationic NPs of biosafe polyphenol molecules linked to mannose and metal ions such as Fe3+ or Cu2+ allowed further electrostatic attachment of CpG stimulator; the nanovaccine targeted DCs thereby eliciting a robust antitumor immune response [201]. Recently advances in cancer vaccines based on whole-tumor antigens were reviewed [202].
This year, a genetically engineered cancer cytomembrane nanovaccine simultaneously overexpressed co-stimulatory molecule CD40L and immune checkpoint inhibitor PD1 to elicit robust antitumor immunity for cancer immunotherapy; the CD40L and tumor antigens from cancer cytomembranes effectively stimulated dendritic cell (DC)-mediated immune activation of cytotoxic T cells, while PD1 on cancer cytomembranes significantly blocked PD1/PD-L1 signaling pathway, synergistically stimulating antitumor immune responses; there was satisfactory prevention in a breast tumor mouse model [203]. One should notice that inhibiting PD1/PD-L1 pathway means allowing T-cells to overcome the immunosuppression usual in the tumor microenvironment [185].
Lipid NPs (LNP) similar to HDL (high-density lipoprotein) carrying tumor-associated antigens from a specific cancer cell membrane were optimized regarding efficiency of lymphatic targeting and dendritic cell uptake; there was suppression of tumor formation and growth and augmented therapeutic efficacy of checkpoint inhibitors notably on the high-stemness melanoma in the mouse models; the effect of NPs charge was evaluated from three LNPs 40 nm in size and different surface charges (−30, −5, and +20 mV) obtained by incorporation of cetyltrimethylammonium bromide at different amounts; the negatively charged ones exhibited superior LN-targeted capacity than the positively charged ones but positively charged ones promoted the DCs uptake of LNPs [204].
Combinatory therapy against cancer often represents a promising strategy. For example, photothermal therapy (PTT) and immunotherapy were combined in the design of a nanovaccine in situ; NPs were assembled from Fe2+, CpG agonist of toll-like receptor 9, cationic lipid and polydopamine (PTT agent); after intratumoral injection, the cationic NPs were exposed to laser light irradiation capturing the released antigens and generating the nanovaccines in situ; there was cross-presentation of tumoral antigens, increased adaptive immune responses, and change of the immunosuppressive tumor environment to immunoresponsive one;hence, in situ nanovaccines highly improved survival rates and elicited a durable immune memory that prevented tumor metastasis, illustrating the synergism for PTT combined with immunotherapy [205].
Interestingly, the self-assembly of the antimicrobial peptide α-melittin yielding NPs evolved as a simple and effective approach against tumor cells; tumor cells were sensitive to α-melittin-NPs, which killed them and promoted the release of whole tumor-cell antigens in situ; furthermore, α-melittin-NPs drained into LNs and activated macrophages and DC after subcutaneous injection; thereafter, the activated effector T cells went to the circulation reaching the tumor and killing their target cancer cells; these NPs also induced the infiltration of innate immune cells such as NK cells and monocytes [206]. Melittin, a main component of bee venom, is a cationic amphiphilic peptide with a linear α-helix structure with antitumor, antiviral and anti-inflammatory effects in vitro and in vivo as comprehensively reviewed recently [207]. In fact, the potential of antimicrobial peptides and their assemblies shaped as NPs requires further investigations for the benefit of cancer immunotherapy [208,209].
Some probiotics such as Lactobacillus rhamnosus and Bifidobacterium longum coated with lipid membrane could be protected against loss of activity and thereby achieve better colonization in colon; subcutaneous transplant of colon cancer in mice was studied in the presence of this probiotic formulation; there were preventive and therapeutic effects that could be further improved by adding cancer nanovaccines; probiotics formulation alone efficiently reduced tumor numbers in colon whereas in combination with cancer nanovaccines improved immune responses were obtained [210].
Cancer nanovaccines have been reviewed from several points of view recently, emphasizing types of nanovaccines and carriers, ideal immune responses, factors determining their efficiency, appropriate antigens and many other important characteristics [202,211,212]. In this review, we presented and discussed main trends regarding cationic nanovaccines and their potential against infectious diseases and cancer.
5. Conclusions
Cationic compounds such as lipids, surfactants, polymers, peptides, metal ions and metal oxides all have been tried as adjuvants for vaccines either alone or in interesting hybrid assemblies with other materials, some of them biocompatible. Among all possibilities, the simpler ones avoiding covalent binding and depending only on interaction forces between antigen, immunostimulator and adjuvant seem to be more feasible and straightforward for scaling up in the biotechnology industry. Among the requirements for applications of cationic adjuvants in vaccinology is the systematic study of cytotoxicity, though in certain instances, precisely this cytotoxicity can be used to disrupt cancer cells in situ releasing multiple antigens in situ prone to local and systemic presentation to APCs thereby eliciting a strong antigen-specific cellular immune response as it should be against cancer and intracellular infective pathogens.
Author Contributions
Conceptualization, A.M.C.-R. and Y.P.B.; methodology, A.M.C.-R. and Y.P.B.; software, A.M.C.-R. and Y.P.B.; validation, A.M.C.-R. and Y.P.B.; formal analysis, A.M.C.-R. and Y.P.B.; investigation, A.M.C.-R. and Y.P.B.; resources, A.M.C.-R.; data curation, A.M.C.-R. and Y.P.B.; writing—original draft preparation, A.M.C.-R. and Y.P.B.; writing—review and editing, A.M.C.-R. and Y.P.B.; visualization, A.M.C.-R. and Y.P.B.; project administration, A.M.C.-R.; funding acquisition, A.M.C.-R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Conselho Nacional de Desenvolvimento Científico e Tecnológico, grants number 302758/2019-4 and 304091/2023-5.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Cellular and Molecular Immunology, 9th ed.; Elsevier Ltd: Philadelphia, PA, USA, 2018; p. 608. ISBN 978-0-323-47978-3. [Google Scholar]
- Manolova, V.; Flace, A.; Bauer, M.; Schwarz, K.; Saudan, P.; Bachmann, M.F. Nanoparticles Target Distinct Dendritic Cell Populations According to Their Size. Eur J Immunol 2008, 38, 1404–1413. [Google Scholar] [CrossRef] [PubMed]
- Fifis, T.; Mottram, P.; Bogdanoska, V.; Hanley, J.; Plebanski, M. Short Peptide Sequences Containing MHC Class I and/or Class II Epitopes Linked to Nano-Beads Induce Strong Immunity and Inhibition of Growth of Antigen-Specific Tumour Challenge in Mice. Vaccine 2004, 23, 258–266. [Google Scholar] [CrossRef] [PubMed]
- Fifis, T.; Gamvrellis, A.; Crimeen-Irwin, B.; Pietersz, G.A.; Li, J.; Mottram, P.L.; McKenzie, I.F.C.; Plebanski, M. Size-Dependent Immunogenicity: Therapeutic and Protective Properties of Nano-Vaccines against Tumors. The Journal of Immunology 2004, 173, 3148–3154. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.D.; Scholzen, A.; Minigo, G.; David, C.; Apostolopoulos, V.; Mottram, P.L.; Plebanski, M. Pathogen Recognition and Development of Particulate Vaccines: Does Size Matter? Methods 2006, 40, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.D.; Wilson, K.L.; Goubier, A.; Heyerick, A.; Plebanski, M. Design of Peptide-Based Nanovaccines Targeting Leading Antigens From Gynecological Cancers to Induce HLA-A2.1 Restricted CD8(+) T Cell Responses. Frontiers in immunology 2018, 9, 2968. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Ugozzoli, M.; Briones, M.; Kazzaz, J.; Soenawan, E.; O’Hagan, D.T. The Effect of CTAB Concentration in Cationic PLG Microparticles on DNA Adsorption and in Vivo Performance. Pharm Res 2003, 20, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Briones, M.; Ott, G.; O’Hagan, D. Cationic Microparticles: A Potent Delivery System for DNA Vaccines. Proc. Natl. Acad. Sci. U.S.A. 2000, 97, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Denis-Mize, K.S.; Dupuis, M.; MacKichan, M.L.; Singh, M.; Doe, B.; O’Hagan, D.; Ulmer, J.B.; Donnelly, J.J.; McDonald, D.M.; Ott, G. Plasmid DNA Adsorbed onto Cationic Microparticles Mediates Target Gene Expression and Antigen Presentation by Dendritic Cells. Gene Therapy 2000, 7, 2105–2112. [Google Scholar] [CrossRef]
- Xiang, S.D.; Selomulya, C.; Ho, J.; Apostolopoulos, V.; Plebanski, M. Delivery of DNA Vaccines: An Overview on the Use of Biodegradable Polymeric and Magnetic Nanoparticles. WIREs Nanomed Nanobiotechnol 2010, 2, 205–218. [Google Scholar] [CrossRef]
- Nazarizadeh, A.; Staudacher, A.H.; Wittwer, N.L.; Turnbull, T.; Brown, M.P.; Kempson, I. Aluminium Nanoparticles as Efficient Adjuvants Compared to Their Microparticle Counterparts: Current Progress and Perspectives. IJMS 2022, 23, 4707. [Google Scholar] [CrossRef]
- Stillman, Z.S.; Decker, G.E.; Dworzak, M.R.; Bloch, E.D.; Fromen, C.A. Aluminum-Based Metal–Organic Framework Nanoparticles as Pulmonary Vaccine Adjuvants. J Nanobiotechnol 2023, 21, 39. [Google Scholar] [CrossRef] [PubMed]
- Raponi, A.; Brewer, J.M.; Garside, P.; Laera, D. Nanoalum Adjuvanted Vaccines: Small Details Make a Big Difference. Seminars in Immunology 2021, 56, 101544. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ribeiro, A.M. Supramolecular Nanostructures for Vaccines. Biomimetics (Basel) 2021, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Betancourt, Y.; Távora, B. de C.L.F.; Faquim-Mauro, E.L.; Carmona-Ribeiro, A.M. Biocompatible Lipid Polymer Cationic Nanoparticles for Antigen Presentation. Polymers 2021, 13, 185. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Betancourt, Y.; Araujo, P.M.; Távora, B. de C.L.F.; Pereira, D.R.; Faquim-Mauro, E.L.; Carmona-Ribeiro, A.M. Cationic and Biocompatible Polymer/Lipid Nanoparticles as Immunoadjuvants. Pharmaceutics 2021, 13, 1859. [Google Scholar] [CrossRef]
- Lincopan, N.; Carmona-Ribeiro, A.M. Protein Assembly onto Cationic Supported Bilayers. Journal of Nanoscience and Nanotechnology 2009, 9, 3578–3586. [Google Scholar] [CrossRef] [PubMed]
- Lincopan, N.; Espíndola, N.M.; Vaz, A.J.; Costa, M.H.B. da; Faquim-Mauro, E.; Carmona-Ribeiro, A.M. Novel Immunoadjuvants Based on Cationic Lipid: Preparation, Characterization and Activity in Vivo. Vaccine 2009, 27, 5760–5771. [Google Scholar] [CrossRef] [PubMed]
- Lincopan, N.; Espíndola, N.; Vaz, A.; Carmonaribeiro, A. Cationic Supported Lipid Bilayers for Antigen Presentation. International journal of pharmaceutics 2007, 340, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Betancourt, Y.; Távora, B. de C.L.F.; Colombini, M.; Faquim-Mauro, E.L.; Carmona-Ribeiro, A.M. Simple Nanoparticles from the Assembly of Cationic Polymer and Antigen as Immunoadjuvants. Vaccines 2020, 8, 105. [Google Scholar] [CrossRef]
- Rozenfeld, J.H.K.; Silva, S.R.; Ranéia, P.A.; Faquim-Mauro, E.; Carmona-Ribeiro, A.M. Stable Assemblies of Cationic Bilayer Fragments and CpG Oligonucleotide with Enhanced Immunoadjuvant Activity in Vivo. J Control Release 2012, 160, 367–373. [Google Scholar] [CrossRef]
- Lincopan, N.; Santana, M.R.; Faquim-Mauro, E.; da Costa, M.H.B.; Carmona-Ribeiro, A.M. Silica-Based Cationic Bilayers as Immunoadjuvants. BMC Biotechnol 2009, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ribeiro, A.M. Biomimetic Systems in Nanomedicine. In Handbook of Nanobiomedical Research: Fundamentals, Applications and Recent Developments; 2014; pp. 401–456 ISBN 978-981-4520-64-5.
- Carmona-Ribeiro, A.M. The Versatile Dioctadecyldimethylammonium Bromide. In Application and Characterization of Surfactants; Najjar, R., Ed.; IntechOpen, 2017; pp. 157–182 ISBN 978-953-51-3326-1.
- Agger, E.M.; Rosenkrands, I.; Hansen, J.; Brahimi, K.; Vandahl, B.S.; Aagaard, C.; Werninghaus, K.; Kirschning, C.; Lang, R.; Christensen, D.; et al. Cationic Liposomes Formulated with Synthetic Mycobacterial Cordfactor (CAF01): A Versatile Adjuvant for Vaccines with Different Immunological Requirements. PloS one 2008, 3, e3116. [Google Scholar] [CrossRef] [PubMed]
- Tsuruta, L.R.; Quintilio, W.; Costa, M.H.; Carmona-Ribeiro, A.M. Interactions between Cationic Liposomes and an Antigenic Protein: The Physical Chemistry of the Immunoadjuvant Action. Journal of Lipid Research 1997, 38, 2003–2011. [Google Scholar] [CrossRef] [PubMed]
- Hilgers, L.A.; Snippe, H. DDA as an Immunological Adjuvant. Res Immunol 1992, 143, 494–503. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Santos, F.A.; Lincopan, N.; De Gaspari, E. Evaluation of Intranasal and Subcutaneous Route of Immunization in Neonatal Mice Using DODAB-BF as Adjuvant with Outer Membrane Vesicles of Neisseria Meningitis B. Immunobiology 2018, 223, 750–760. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, A.F.; De Gaspari, E. Dioctadecyldimethylammonium Bromide (DODAB-BF) as a New Adjuvant for Maternal-Fetal Immunization in Mice against Neisseria Meningitidis: Evaluation of Humoral Response. Pathogens and disease 2018, 76. [Google Scholar] [CrossRef] [PubMed]
- Davidsen, J.; Rosenkrands, I.; Christensen, D.; Vangala, A.; Kirby, D.; Perrie, Y.; Agger, E.M.; Andersen, P. Characterization of Cationic Liposomes Based on Dimethyldioctadecylammonium and Synthetic Cord Factor from M. Tuberculosis (Trehalose 6,6’-Dibehenate)-a Novel Adjuvant Inducing Both Strong CMI and Antibody Responses. Biochimica et biophysica acta 2005, 1718, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Baranov, M.V.; Kumar, M.; Sacanna, S.; Thutupalli, S.; Van Den Bogaart, G. Modulation of Immune Responses by Particle Size and Shape. Front. Immunol. 2021, 11, 607945. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, E.B.; Rosetti, A.S.; Lincopan, N.; De Gaspari, E. Neisseria Lactamica Antigens Complexed with a Novel Cationic Adjuvant. Human Vaccines & Immunotherapeutics 2013, 9, 572–581. [Google Scholar] [CrossRef]
- Chono, S.; Tanino, T.; Seki, T.; Morimoto, K. Influence of Particle Size on Drug Delivery to Rat Alveolar Macrophages Following Pulmonary Administration of Ciprofloxacin Incorporated into Liposomes. Journal of Drug Targeting 2006, 14, 557–566. [Google Scholar] [CrossRef]
- Epstein-Barash, H.; Gutman, D.; Markovsky, E.; Mishan-Eisenberg, G.; Koroukhov, N.; Szebeni, J.; Golomb, G. Physicochemical Parameters Affecting Liposomal Bisphosphonates Bioactivity for Restenosis Therapy: Internalization, Cell Inhibition, Activation of Cytokines and Complement, and Mechanism of Cell Death. Journal of Controlled Release 2010, 146, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Ramirez-Ortiz, Z.G.; Means, T.K. The Role of Dendritic Cells in the Innate Recognition of Pathogenic Fungi ( A. Fumigatus, C. Neoformans and C. Albicans ). Virulence 2012, 3, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Benne, N.; Van Duijn, J.; Kuiper, J.; Jiskoot, W.; Slütter, B. Orchestrating Immune Responses: How Size, Shape and Rigidity Affect the Immunogenicity of Particulate Vaccines. Journal of Controlled Release 2016, 234, 124–134. [Google Scholar] [CrossRef]
- Mottram, P.L.; Leong, D.; Crimeen-Irwin, B.; Gloster, S.; Xiang, S.D.; Meanger, J.; Ghildyal, R.; Vardaxis, N.; Plebanski, M. Type 1 and 2 Immunity Following Vaccination Is Influenced by Nanoparticle Size: Formulation of a Model Vaccine for Respiratory Syncytial Virus. Molecular pharmaceutics 2007, 4, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Kanchan, V.; Panda, A.K. Interactions of Antigen-Loaded Polylactide Particles with Macrophages and Their Correlation with the Immune Response. Biomaterials 2007, 28, 5344–5357. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sloat, B.R.; Yanasarn, N.; Cui, Z. Relationship between the Size of Nanoparticles and Their Adjuvant Activity: Data from a Study with an Improved Experimental Design. European Journal of Pharmaceutics and Biopharmaceutics 2011, 78, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Heung, L.J.; Wiesner, D.L.; Wang, K.; Rivera, A.; Hohl, T.M. Immunity to Fungi in the Lung. Seminars in Immunology 2023, 66, 101728. [Google Scholar] [CrossRef] [PubMed]
- Heung, L.J. Monocytes and the Host Response to Fungal Pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 34. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, H.-J.; Hang, T.; Yu, Y.; Liu, G.; He, G.; Xiao, S.; Yang, B.; Yang, C.; Liu, F.; et al. Physical Activation of Innate Immunity by Spiky Particles. Nature Nanotech 2018, 13, 1078–1086. [Google Scholar] [CrossRef]
- Cong, V.T.; Houng, J.L.; Kavallaris, M.; Chen, X.; Tilley, R.D.; Gooding, J.J. How Can We Use the Endocytosis Pathways to Design Nanoparticle Drug-Delivery Vehicles to Target Cancer Cells over Healthy Cells? Chem. Soc. Rev. 2022, 51, 7531–7559. [Google Scholar] [CrossRef]
- Sabourian, P.; Yazdani, G.; Ashraf, S.S.; Frounchi, M.; Mashayekhan, S.; Kiani, S.; Kakkar, A. Effect of Physico-Chemical Properties of Nanoparticles on Their Intracellular Uptake. IJMS 2020, 21, 8019. [Google Scholar] [CrossRef] [PubMed]
- Sahay, G.; Alakhova, D.Y.; Kabanov, A.V. Endocytosis of Nanomedicines. Journal of Controlled Release 2010, 145, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Behzadi, S.; Serpooshan, V.; Tao, W.; Hamaly, M.A.; Alkawareek, M.Y.; Dreaden, E.C.; Brown, D.; Alkilany, A.M.; Farokhzad, O.C.; Mahmoudi, M. Cellular Uptake of Nanoparticles: Journey inside the Cell. Chem Soc Rev 2017, 46, 4218–4244. [Google Scholar] [CrossRef] [PubMed]
- Sousa De Almeida, M.; Susnik, E.; Drasler, B.; Taladriz-Blanco, P.; Petri-Fink, A.; Rothen-Rutishauser, B. Understanding Nanoparticle Endocytosis to Improve Targeting Strategies in Nanomedicine. Chem. Soc. Rev. 2021, 2021 50, 5397–5434. [Google Scholar] [CrossRef]
- Zhu, G.H.; Gray, A.B.C.; Patra, H.K. Nanomedicine: Controlling Nanoparticle Clearance for Translational Success. Trends in Pharmacological Sciences 2022, 43, 709–711. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, X.; Ito, A. Tailoring Inorganic Nanoadjuvants towards Next-Generation Vaccines. Chem. Soc. Rev. 2018, 47, 4954–4980. [Google Scholar] [CrossRef] [PubMed]
- Du, G.; Sun, X. Engineering Nanoparticulate Vaccines for Enhancing Antigen Cross-Presentation. Current Opinion in Biotechnology 2020, 66, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Song, T.; Liao, Y.; Zuo, Q.; Liu, N.; Liu, Z. MnO 2 Nanoparticles as a Minimalist Multimode Vaccine Adjuvant/Delivery System to Regulate Antigen Presenting Cells for Tumor Immunotherapy. J. Mater. Chem. B 2022, 10, 3474–3490. [Google Scholar] [CrossRef] [PubMed]
- Breslin, J.W.; Yang, Y.; Scallan, J.P.; Sweat, R.S.; Adderley, S.P.; Murfee, W.L. Lymphatic Vessel Network Structure and Physiology. In Comprehensive Physiology; Prakash, Y.S., Ed.; Wiley, 2018; pp. 207–299 ISBN 978-0-470-65071-4.
- Tang, Y.; Liu, B.; Zhang, Y.; Liu, Y.; Huang, Y.; Fan, W. Interactions between Nanoparticles and Lymphatic Systems: Mechanisms and Applications in Drug Delivery. Advanced Drug Delivery Reviews 2024, 209, 115304. [Google Scholar] [CrossRef]
- Zhang, W.; Lu, L.; Zhu, Z.; Deng, F.; Zhang, W.; Wang, F.; Zeng, P.; Shi, H.; Wang, T.; Chen, Y.; et al. A Manganese-Based Nanodriver Coordinates Tumor Prevention and Suppression through STING Activation in Glioblastoma. Adv Healthcare Materials 2024, 2400421. [Google Scholar] [CrossRef]
- Chen, J.; Ye, Z.; Huang, C.; Qiu, M.; Song, D.; Li, Y.; Xu, Q. Lipid Nanoparticle-Mediated Lymph Node–Targeting Delivery of mRNA Cancer Vaccine Elicits Robust CD8 + T Cell Response. Proc. Natl. Acad. Sci. U.S.A. 2022, 119, e2207841119. [Google Scholar] [CrossRef] [PubMed]
- Pizzuto, M.; Bigey, P.; Lachages, A.-M.; Hoffmann, C.; Ruysschaert, J.-M.; Escriou, V.; Lonez, C. Cationic Lipids as One-Component Vaccine Adjuvants: A Promising Alternative to Alum. Journal of controlled release : official journal of the Controlled Release Society 2018, 287, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Wilmar, A.; Lonez, C.; Vermeersch, M.; Andrianne, M.; Perez-Morga, D.; Ruysschaert, J.-M.; Vandenbranden, M.; Leo, O.; Temmerman, S.T. The Cationic Lipid, diC14 Amidine, Extends the Adjuvant Properties of Aluminum Salts through a TLR-4- and Caspase-1-Independent Mechanism. Vaccine 2012, 30, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Lonez, C.; Bessodes, M.; Scherman, D.; Vandenbranden, M.; Escriou, V.; Ruysschaert, J.-M. Cationic Lipid Nanocarriers Activate Toll-like Receptor 2 and NLRP3 Inflammasome Pathways. Nanomedicine: Nanotechnology, Biology and Medicine 2014, 10, 775–782. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M. Lipid Bilayer Fragments and Disks in Drug Delivery. Curr Med Chem 2006, 13, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ribeiro, A.M. Synthetic Amphiphile Vesicles. Chem. Soc. Rev. 1992, 21, 209–214. [Google Scholar] [CrossRef]
- Naves, A.F.; Palombo, R.R.; Carrasco, L.D.M.; Carmona-Ribeiro, A.M. Antimicrobial Particles from Emulsion Polymerization of Methyl Methacrylate in the Presence of Quaternary Ammonium Surfactants. Langmuir 2013, 29, 9677–9684. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, L.D. de M.; Bertolucci, R.J.; Ribeiro, R.T.; Sampaio, J.L.M.; Carmona-Ribeiro, A.M. Cationic Nanostructures against Foodborne Pathogens. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ribeiro, A.M. Bilayer Vesicles and Liposomes as Interface Agents. Chem. Soc. Rev. 2001, 30, 241–247. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M. Biomimetic Nanomaterials from the Assembly of Polymers, Lipids, and Surfactants. Surfactants and Detergents 2019. [Google Scholar] [CrossRef]
- Ribeiro, R.T.; Galvão, C.N.; Betancourt, Y.P.; Mathiazzi, B.I.; Carmona-Ribeiro, A.M. Microbicidal Dispersions and Coatings from Hybrid Nanoparticles of Poly (Methyl Methacrylate), Poly (Diallyl Dimethyl Ammonium) Chloride, Lipids, and Surfactants. International journal of molecular sciences 2019, 20, 6150. [Google Scholar] [CrossRef] [PubMed]
- Thangsunan, P.; Kitiyodom, S.; Srisapoome, P.; Pirarat, N.; Yata, T.; Thangsunan, P.; Boonrungsiman, S.; Bunnoy, A.; Rodkhum, C. Novel Development of Cationic Surfactant-Based Mucoadhesive Nanovaccine for Direct Immersion Vaccination against Francisella Noatunensis Subsp. Orientalis in Red Tilapia (Oreochromis Sp.). Fish & Shellfish Immunology 2022, 127, 1051–1060. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M. Lipid-Based Biomimetics in Drug and Vaccine Delivery. In Biomimetics Learning from Nature; IntechOpen, 2010 ISBN 978-953-307-025-4.
- Carmona-Ribeiro, A.M.; de Melo Carrasco, L.D. Cationic Antimicrobial Polymers and Their Assemblies. Int J Mol Sci 2013, 14, 9906–9946. [Google Scholar] [CrossRef]
- Rehman, Z.U.; Hoekstra, D.; Zuhorn, I.S. Mechanism of Polyplex- and Lipoplex-Mediated Delivery of Nucleic Acids: Real-Time Visualization of Transient Membrane Destabilization without Endosomal Lysis. ACS Nano 2013, 7, 3767–3777. [Google Scholar] [CrossRef]
- Simón-Vázquez, R.; Peleteiro, M.; González-Fernández, Á. Polymeric Nanostructure Vaccines: Applications and Challenges. Expert Opinion on Drug Delivery 2020, 17, 1007–1023. [Google Scholar] [CrossRef]
- Thompson, K.D.; Rodkhum, C.; Bunnoy, A.; Thangsunan, P.; Kitiyodom, S.; Sukkarun, P.; Yostawornkul, J.; Yata, T.; Pirarat, N. Addressing Nanovaccine Strategies for Tilapia. Vaccines 2023, 11, 1356. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M.; Araújo, P.M. Antimicrobial Polymer−Based Assemblies: A Review. International Journal of Molecular Sciences 2021, 22, 5424. [Google Scholar] [CrossRef] [PubMed]
- Rosa, H.; Petri, D.F.S.; Carmona-Ribeiro, A.M. Interactions between Bacteriophage DNA and Cationic Biomimetic Particles. J. Phys. Chem. B 2008, 112, 16422–16430. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ribeiro, A.M.; de Moraes Lessa, M. Interactions between Bilayer Membranes and Latex. Colloids and Surfaces A: Physicochemical and Engineering Aspects 1999, 153, 355–361. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M.; Pérez-Betancourt, Y. Cationic Nanostructures for Vaccines Design. Biomimetics (Basel, Switzerland) 2020, 5. [Google Scholar] [CrossRef]
- Choi, K.C.; Lee, D.H.; Lee, J.W.; Lee, J.S.; Lee, Y.K.; Choi, M.J.; Jeong, H.Y.; Kim, M.W.; Lee, C.-G.; Park, Y.S. Novel Lipid Nanoparticles Stable and Efficient for mRNA Transfection to Antigen-Presenting Cells. IJMS 2024, 25, 1388. [Google Scholar] [CrossRef] [PubMed]
- Anderluzzi, G.; Schmidt, S.T.; Cunliffe, R.; Woods, S.; Roberts, C.W.; Veggi, D.; Ferlenghi, I.; O’Hagan, D.T.; Baudner, B.C.; Perrie, Y. Rational Design of Adjuvants for Subunit Vaccines: The Format of Cationic Adjuvants Affects the Induction of Antigen-Specific Antibody Responses. Journal of Controlled Release 2021, 330, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Heuts, J.; Jiskoot, W.; Ossendorp, F.; Van Der Maaden, K. Cationic Nanoparticle-Based Cancer Vaccines. Pharmaceutics 2021, 13, 596. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ribeiro, A.M.; Midmore, B.R. Synthetic Bilayer Adsorption onto Polystyrene Microspheres. Langmuir 1992, 8, 801–806. [Google Scholar] [CrossRef]
- Liu, L.; Cao, F.; Liu, X.; Wang, H.; Zhang, C.; Sun, H.; Wang, C.; Leng, X.; Song, C.; Kong, D.; et al. Hyaluronic Acid-Modified Cationic Lipid–PLGA Hybrid Nanoparticles as a Nanovaccine Induce Robust Humoral and Cellular Immune Responses. ACS Appl. Mater. Interfaces 2016, 8, 11969–11979. [Google Scholar] [CrossRef] [PubMed]
- Galvão, C.N.; Sanches, L.M.; Mathiazzi, B.I.; Ribeiro, R.T.; Petri, D.F.S.; Carmona-Ribeiro, A.M. Antimicrobial Coatings from Hybrid Nanoparticles of Biocompatible and Antimicrobial Polymers. International Journal of Molecular Sciences 2018, 19, 2965. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Alexander-Katz, A. Cell Membranes Open “Doors” for Cationic Nanoparticles/Biomolecules: Insights into Uptake Kinetics. ACS Nano 2013, 7, 10799–10808. [Google Scholar] [CrossRef] [PubMed]
- Wegmann, F.; Gartlan, K.H.; Harandi, A.M.; Brinckmann, S.A.; Coccia, M.; Hillson, W.R.; Kok, W.L.; Cole, S.; Ho, L.-P.; Lambe, T.; et al. Polyethyleneimine Is a Potent Mucosal Adjuvant for Viral Glycoprotein Antigens. Nature Biotechnology 2012, 30, 883–888. [Google Scholar] [CrossRef]
- Kedmi, R.; Ben-Arie, N.; Peer, D. The Systemic Toxicity of Positively Charged Lipid Nanoparticles and the Role of Toll-like Receptor 4 in Immune Activation. Biomaterials 2010, 31, 6867–6875. [Google Scholar] [CrossRef]
- Liao, X.; Liu, Y.; Zheng, J.; Zhao, X.; Cui, L.; Hu, S.; Xia, T.; Si, S. Diverse Pathways of Engineered Nanoparticle-Induced NLRP3 Inflammasome Activation. Nanomaterials 2022, 12, 3908. [Google Scholar] [CrossRef]
- Li, T.; He, J.; Horvath, G.; Próchnicki, T.; Latz, E.; Takeoka, S. Lysine-Containing Cationic Liposomes Activate the NLRP3 Inflammasome: Effect of a Spacer between the Head Group and the Hydrophobic Moieties of the Lipids. Nanomedicine: Nanotechnology, Biology and Medicine 2018, 14, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Tracey, S.R.; Smyth, P.; Herron, U.M.; Burrows, J.F.; Porter, A.J.; Barelle, C.J.; Scott, C.J. Development of a Cationic Polyethyleneimine-Poly(Lactic- Co -Glycolic Acid) Nanoparticle System for Enhanced Intracellular Delivery of Biologics. RSC Adv. 2023, 13, 33721–33735. [Google Scholar] [CrossRef] [PubMed]
- Fischer, D.; Li, Y.; Ahlemeyer, B.; Krieglstein, J.; Kissel, T. In Vitro Cytotoxicity Testing of Polycations: Influence of Polymer Structure on Cell Viability and Hemolysis. Biomaterials 2003, 24, 1121–1131. [Google Scholar] [CrossRef] [PubMed]
- Pizzuto, M.; Gangloff, M.; Scherman, D.; Gay, N.J.; Escriou, V.; Ruysschaert, J.-M.; Lonez, C. Toll-like Receptor 2 Promiscuity Is Responsible for the Immunostimulatory Activity of Nucleic Acid Nanocarriers. Journal of Controlled Release 2017, 247, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ribeiro, A.M. Cationic Nanostructures for Vaccines. In Immune Response Activation; Duc, G.H.T., Ed.; InTech, 2014 ISBN 978-953-51-1374-4.
- Yew, N.S.; Wang, K.X.; Przybylska, M.; Bagley, R.G.; Stedman, M.; Marshall, J.; Scheule, R.K.; Cheng, S.H. Contribution of Plasmid DNA to Inflammation in the Lung after Administration of Cationic Lipid:pDNA Complexes. Human Gene Therapy 1999, 10, 223–234. [Google Scholar] [CrossRef]
- Carvalho, L.A.; Carmona-Ribeiro, A.M. Interactions between Cationic Vesicles and Serum Proteins. Langmuir 1998, 14, 6077–6081. [Google Scholar] [CrossRef]
- Kayitmazer, A.B.; Strand, S.P.; Tribet, C.; Jaeger, W.; Dubin, P.L. Effect of Polyelectrolyte Structure on Protein-Polyelectrolyte Coacervates: Coacervates of Bovine Serum Albumin with Poly(Diallyldimethylammonium Chloride) versus Chitosan. Biomacromolecules 2007, 8, 3568–3577. [Google Scholar] [CrossRef]
- Li, S.; Tseng, W.-C.; Stolz, D.B.; Wu, S.-P.; Watkins, S.C.; Huang, L. Dynamic Changes in the Characteristics of Cationic Lipidic Vectors after Exposure to Mouse Serum: Implications for Intravenous Lipofection. Gene Ther 1999, 6, 585–594. [Google Scholar] [CrossRef]
- Li, W.; Szoka, F.C. Lipid-Based Nanoparticles for Nucleic Acid Delivery. Pharm Res 2007, 24, 438–449. [Google Scholar] [CrossRef]
- G. Lima, E.; R. Gomes, L.; M. Carmona-Ribeiro, A. Stable Indomethacin Dispersions in Water from Drug, Ethanol, Cationic Lipid and Carboxymethyl-Cellulose. Pharmaceutical Nanotechnology 2016, 4, 126–135. [Google Scholar] [CrossRef]
- Barbassa, L.; Mamizuka, E.M.; Carmona-Ribeiro, A.M. Supramolecular Assemblies of Rifampicin and Cationic Bilayers: Preparation, Characterization and Micobactericidal Activity. BMC Biotechnol 2011, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- Gujrati, M.; Malamas, A.; Shin, T.; Jin, E.; Sun, Y.; Lu, Z.-R. Multifunctional Cationic Lipid-Based Nanoparticles Facilitate Endosomal Escape and Reduction-Triggered Cytosolic siRNA Release. Mol. Pharmaceutics 2014, 11, 2734–2744. [Google Scholar] [CrossRef] [PubMed]
- Rayamajhi, S.; Marchitto, J.; Nguyen, T.D.T.; Marasini, R.; Celia, C.; Aryal, S. pH-Responsive Cationic Liposome for Endosomal Escape Mediated Drug Delivery. Colloids and Surfaces B: Biointerfaces 2020, 188, 110804. [Google Scholar] [CrossRef] [PubMed]
- Ho, N.I.; in ’t Veld, L.G.M.; Raaijmakers, T.K.; Adema, G.J.; Huis In ’t Veld, L.G.M.; Raaijmakers, T.K.; Adema, G.J. Adjuvants Enhancing Cross-Presentation by Dendritic Cells: The Key to More Effective Vaccines? Frontiers in Immunology 2018, 9, 2874. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Zhang, Y.; Hou, L.; Qiao, X.; Zhang, Y.; Cheng, H.; Lu, H.; Chen, J.; Du, L.; Zheng, Q.; et al. Increases in Cellular Immune Responses Due to Positive Effect of CVC1302-Induced Lysosomal Escape in Mice. Vaccines 2023, 11, 1718. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zhao, C.; Wang, W.; Liu, X.; Deng, H. Biomimetic Noncationic Lipid Nanoparticles for mRNA Delivery. Proc. Natl. Acad. Sci. U.S.A. 2023, 120, e2311276120. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.; Wafa, E.I.; Geary, S.M.; Ebeid, K.; Alhaj-Suliman, S.O.; Salem, A.K. Cationic Nanoparticles Enhance T Cell Tumor Infiltration and Antitumor Immune Responses to a Melanoma Vaccine. Sci. Adv. 2022, 8, eabk3150. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, X.; Qi, Y.; Wang, M.; Yu, N.; Nan, F.; Zhang, H.; Tian, M.; Li, C.; Lu, H.; Jin, N. mRNA Vaccines Encoding the HA Protein of Influenza A H1N1 Virus Delivered by Cationic Lipid Nanoparticles Induce Protective Immune Responses in Mice. Vaccines 2020, 8. [Google Scholar] [CrossRef]
- Wusiman, A.; Gu, P.; Liu, Z.; Xu, S.; Zhang, Y.; Hu, Y.; Liu, J.; Wang, D.; Huang, X. Cationic Polymer Modified PLGA Nanoparticles Encapsulating Alhagi Honey Polysaccharides as a Vaccine Delivery System for Ovalbumin to Improve Immune Responses. International journal of nanomedicine 2019, 14, 3221–3234. [Google Scholar] [CrossRef]
- Ebrahimian, M.; Hashemi, M.; Maleki, M.; Hashemitabar, G.; Abnous, K.; Ramezani, M.; Haghparast, A. Co-Delivery of Dual Toll-like Receptor Agonists and Antigen in Poly(Lactic-Co-Glycolic) Acid/Polyethylenimine Cationic Hybrid Nanoparticles Promote Efficient in Vivo Immune Responses. Frontiers in immunology 2017, 8, 1077–1077. [Google Scholar] [CrossRef]
- Rapuano, R.; Carmona-Ribeiro, A.M. Physical Adsorption of Bilayer Membranes on Silica. Journal of Colloid and Interface Science 1997, 193, 104–111. [Google Scholar] [CrossRef]
- Moura, S.P.; Carmona-Ribeiro, A.M. Cationic Bilayer Fragments on Silica at Low Ionic Strength: Competitive Adsorption and Colloid Stability. Langmuir 2003, 19, 6664–6667. [Google Scholar] [CrossRef]
- Ribeiro, R.T.; Braga, V.H.A.; Carmona-Ribeiro, A.M. Biomimetic Cationic Nanoparticles Based on Silica: Optimizing Bilayer Deposition from Lipid Films. Biomimetics 2017, 2, 20. [Google Scholar] [CrossRef] [PubMed]
- Melo, L.D.; Mamizuka, E.M.; Carmona-Ribeiro, A.M. Antimicrobial Particles from Cationic Lipid and Polyelectrolytes. Langmuir 2010, 26, 12300–12306. [Google Scholar] [CrossRef] [PubMed]
- de Melo Carrasco, L.D.; Sampaio, J.L.M.; Carmona-Ribeiro, A.M. Supramolecular Cationic Assemblies against Multidrug-Resistant Microorganisms: Activity and Mechanism of Action. Int J Mol Sci 2015, 16, 6337–6352. [Google Scholar] [CrossRef]
- Pereira, E.M.A.; Kosaka, P.M.; Rosa, H.; Vieira, D.B.; Kawano, Y.; Petri, D.F.S.; Carmona-Ribeiro, A.M. Hybrid Materials from Intermolecular Associations between Cationic Lipid and Polymers. J. Phys. Chem. B 2008, 112, 9301–9310. [Google Scholar] [CrossRef]
- Sanches, L.M.; Petri, D.F.S.; de Melo Carrasco, L.D.; Carmona-Ribeiro, A.M. The Antimicrobial Activity of Free and Immobilized Poly (Diallyldimethylammonium) Chloride in Nanoparticles of Poly (Methylmethacrylate). Journal of Nanobiotechnology 2015, 13, 58. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ribeiro, A.M.; Castuma, C.E.; Sesso, A.; Schreier, S. Bilayer Structure and Stability in Dihexadecyl Phosphate Dispersions. J. Phys. Chem. 1991, 95, 5361–5366. [Google Scholar] [CrossRef]
- Andersson, M.; Hammarstroem, L.; Edwards, K. Effect of Bilayer Phase Transitions on Vesicle Structure, and Its Influence on the Kinetics of Viologen Reduction. J. Phys. Chem. 1995, 99, 14531–14538. [Google Scholar] [CrossRef]
- Vieira, D.B.; Carmona-Ribeiro, A.M. Synthetic Bilayer Fragments for Solubilization of Amphotericin B. Journal of Colloid and Interface Science 2001, 244, 427–431. [Google Scholar] [CrossRef]
- Vieira, D.B.; Carmona-Ribeiro, A.M. Cationic Nanoparticles for Delivery of Amphotericin B: Preparation, Characterization and Activity in Vitro. Journal of Nanobiotechnology 2008, 6, 6. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M. Biomimetic Nanoparticles: Preparation, Characterization and Biomedical Applications. Int J Nanomedicine 2010, 5, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ribeiro, A.M. Biomimetic Particles in Drug and Vaccine Delivery. Journal of Liposome Research 2007, 17, 165–172. [Google Scholar] [CrossRef]
- Yong, C.Y.; Liew, W.P.P.; Ong, H.K.; Poh, C.L. Development of Virus-like Particles-based Vaccines against Coronaviruses. Biotechnology Progress 2022, 38, e3292. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.A.J. Recombinant DNA Technology and DNA Sequencing. Essays in Biochemistry 2019, 63, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, Z.; Aria, H.; Ghaedrahmati, F.; Bakhtiari, T.; Azizi, M.; Bastan, R.; Hosseini, R.; Eskandari, N. An Update on Human Papilloma Virus Vaccines: History, Types, Protection, and Efficacy. Front. Immunol. 2022, 12, 805695. [Google Scholar] [CrossRef]
- Cohen, S.N.; Chang, A.C.Y.; Boyer, H.W.; Helling, R.B. Construction of Biologically Functional Bacterial Plasmids In Vitro. Proc. Natl. Acad. Sci. U.S.A. 1973, 70, 3240–3244. [Google Scholar] [CrossRef]
- Nielsen, J. Production of Biopharmaceutical Proteins by Yeast: Advances through Metabolic Engineering. Bioengineered 2013, 4, 207–211. [Google Scholar] [CrossRef]
- Schoenmaker, L.; Witzigmann, D.; Kulkarni, J.A.; Verbeke, R.; Kersten, G.; Jiskoot, W.; Crommelin, D.J.A. mRNA-Lipid Nanoparticle COVID-19 Vaccines: Structure and Stability. International Journal of Pharmaceutics 2021, 601, 120586. [Google Scholar] [CrossRef]
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the Clinic: An Update Post COVID -19 Vaccines. Bioengineering & Transla Med 2021, 6, e10246. [Google Scholar] [CrossRef]
- Pilkington, E.H.; Suys, E.J.A.; Trevaskis, N.L.; Wheatley, A.K.; Zukancic, D.; Algarni, A.; Al-Wassiti, H.; Davis, T.P.; Pouton, C.W.; Kent, S.J.; et al. From Influenza to COVID-19: Lipid Nanoparticle mRNA Vaccines at the Frontiers of Infectious Diseases. Acta Biomaterialia 2021, 131, 16–40. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, X.-Q.; Ho, W.; Li, F.; Gao, M.; Bai, X.; Xu, X. Enzyme-Catalyzed One-Step Synthesis of Ionizable Cationic Lipids for Lipid Nanoparticle-Based mRNA COVID-19 Vaccines. ACS Nano 2022, 16, 18936–18950. [Google Scholar] [CrossRef] [PubMed]
- Yadav, T.; Srivastava, N.; Mishra, G.; Dhama, K.; Kumar, S.; Puri, B.; Saxena, S.K. Recombinant Vaccines for COVID-19. Human Vaccines & Immunotherapeutics 2020, 16, 2905–2912. [Google Scholar] [CrossRef]
- Global Tuberculosis Report 2023 Available online:. Available online: https://www.who.int/teams/global-tuberculosis-programme/tb-reports/global-tuberculosis-report-2023 (accessed on 16 May 2024).
- Philips, J.A.; Ernst, J.D. Tuberculosis Pathogenesis and Immunity. Annual Review of Pathology: Mechanisms of Disease 2012, 7, 353–384. [Google Scholar] [CrossRef] [PubMed]
- Brewer, T.F. Preventing Tuberculosis with Bacillus Calmette-Guérin Vaccine: A Meta-Analysis of the Literature. Clinical Infectious Diseases 2000, 31, S64–S67. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, A. Bacillus Calmette-Guérin (BCG) Revaccination and Protection Against Tuberculosis: A Systematic Review. Cureus 2024, 16. [Google Scholar] [CrossRef] [PubMed]
- Szachniewicz, M.M.; Neustrup, M.A.; van Meijgaarden, K.E.; Jiskoot, W.; Bouwstra, J.A.; Haks, M.C.; Geluk, A.; Ottenhoff, T.H.M. Intrinsic Immunogenicity of Liposomes for Tuberculosis Vaccines: Effect of Cationic Lipid and Cholesterol. European Journal of Pharmaceutical Sciences 2024, 195, 106730. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ribeiro, A.M.; Ortis, F.; Schumacher, R.I.; Armelin, M.C.S. Interactions between Cationic Vesicles and Cultured Mammalian Cells. Langmuir 1997, 13, 2215–2218. [Google Scholar] [CrossRef]
- Carrasco, L.D.; Sampaio, J.L.M.; Santos, Hadassa; Carmona-Ribeiro, A. Self-Assembled Antibiotic Nanoparticles Against Intracellular Bacteria. DDL 2017, 7, 39–47. [Google Scholar] [CrossRef]
- Carmona Ribeiro, A.M.; Chaimovich, H. Preparation and Characterization of Large Dioctadecyldimethylammonium Chloride Liposomes and Comparison with Small Sonicated Vesicles. Biochimica et Biophysica Acta (BBA) - Biomembranes 1983, 733, 172–179. [Google Scholar] [CrossRef]
- Mansury, D.; Ghazvini, K.; Amel Jamehdar, S.; Badiee, A.; Tafaghodi, M.; Nikpoor, A.R.; Amini, Y.; Jaafari, M.R. Enhancement of the Effect of BCG Vaccine against Tuberculosis Using DDA/TDB Liposomes Containing a Fusion Protein of HspX, PPE44, and EsxV. Artificial Cells, Nanomedicine, and Biotechnology 2019, 47, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, J.; Billeskov, R.; Doherty, T.M.; Andersen, P. Synergistic Effect of Bacillus Calmette Guerin and a Tuberculosis Subunit Vaccine in Cationic Liposomes: Increased Immunogenicity and Protection1. The Journal of Immunology 2007, 178, 3721–3730. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Du, J.; Wang, X.; Xu, L.; Zhang, Y.; Sun, Q.; Shi, Z.; Xing, Y.; Su, Y.; Wang, S.; et al. Long-Term Immunogenicity and In Vitro Prophylactic Protective Efficacy of M. Tuberculosis Fusion Protein DR2 Combined with Liposomal Adjuvant DIMQ as a Boosting Vaccine for BCG. ACS Infect. Dis. 2023, 9, 593–608. [Google Scholar] [CrossRef] [PubMed]
- Derrick, S.C.; Yang, A.; Parra, M.; Kolibab, K.; Morris, S.L. Effect of Cationic Liposomes on BCG Trafficking and Vaccine-Induced Immune Responses Following a Subcutaneous Immunization in Mice. Vaccine 2015, 33, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ribeiro, A.M.; Chaimovich, H. Salt-Induced Aggregation and Fusion of Dioctadecyldimethylammonium Chloride and Sodium Dihexadecylphosphate Vesicles. Biophys J 1986, 50, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Sobral, C.N.C.; Soto, M.A.; Carmona-Ribeiro, A.M. Characterization of DODAB/DPPC Vesicles. Chemistry and Physics of Lipids 2008, 152, 38–45. [Google Scholar] [CrossRef]
- Tian, M.; Zhou, Z.; Tan, S.; Fan, X.; Li, L.; Ullah, N. Formulation in DDA-MPLA-TDB Liposome Enhances the Immunogenicity and Protective Efficacy of a DNA Vaccine against Mycobacterium Tuberculosis Infection. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Agger, E.M.; Rosenkrands, I.; Olsen, A.W.; Hatch, G.; Williams, A.; Kritsch, C.; Lingnau, K.; von Gabain, A.; Andersen, C.S.; Korsholm, K.S.; et al. Protective Immunity to Tuberculosis with Ag85B-ESAT-6 in a Synthetic Cationic Adjuvant System IC31. Vaccine 2006, 24, 5452–5460. [Google Scholar] [CrossRef]
- Gao, Y.; Han, S.; Lu, F.; Liu, Q.; Yang, J.; Wang, W.; Wang, Y.; Zhang, J.; Ju, R.; Shen, X.; et al. Dimethyl-Dioctadecyl-Ammonium Bromide/Poly(Lactic Acid) Nanoadjuvant Enhances the Immunity and Cross-Protection of an NM2e-Based Universal Influenza Vaccine. ACS Nano 2024, 18, 12905–12916. [Google Scholar] [CrossRef] [PubMed]
- Mathiazzi, B.I.; Carmona-Ribeiro, A.M. Hybrid Nanoparticles of Poly (Methyl Methacrylate) and Antimicrobial Quaternary Ammonium Surfactants. Pharmaceutics 2020, 12, 340. [Google Scholar] [CrossRef]
- Henson, T.R.; Richards, K.A.; Gandhapudi, S.K.; Woodward, J.G.; Sant, A.J. R-DOTAP Cationic Lipid Nanoparticles Outperform Squalene-Based Adjuvant Systems in Elicitation of CD4 T Cells after Recombinant Influenza Hemagglutinin Vaccination. Viruses 2023, 15, 538. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.V.; Reece, W.; Gothard, P.; Moorthy, V.; Roberts, M.; Flanagan, K.; Plebanski, M.; Hannan, C.; Hu, J.T.; Anderson, R.; et al. DNA-Based Vaccines for Malaria: A Heterologous Prime-Boost Immunisation Strategy. Developments in biologicals 2000, 104, 171–179. [Google Scholar] [PubMed]
- González-Sanz, M.; Berzosa, P.; Norman, F.F. Updates on Malaria Epidemiology and Prevention Strategies. Curr Infect Dis Rep 2023, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Laurens, M.B. RTS,S/AS01 Vaccine (MosquirixTM): An Overview. Hum Vaccin Immunother 2019, 16, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, A.Y.; Shehzad, A.; Islam, S.U.; Al-Suhaimi, E.A.; Lee, Y.S. MosquirixTM RTS, S/AS01 Vaccine Development, Immunogenicity, and Efficacy. Vaccines 2022, 10, 713. [Google Scholar] [CrossRef] [PubMed]
- Fotoran, W.L.; Silva, J.R. da; Glitz, C.; Ferreira, L.C. de S.; Wunderlich, G. Establishment of an Antiplasmodial Vaccine Based on PfRH5-Encoding RNA Replicons Stabilized by Cationic Liposomes. Pharmaceutics 2023, 15, 1223. [Google Scholar] [CrossRef] [PubMed]
- Fotoran, W.L.; Kleiber, N.; Glitz, C.; Wunderlich, G. A DNA Vaccine Encoding Plasmodium Falciparum PfRH5 in Cationic Liposomes for Dermal Tattooing Immunization. Vaccines 2020, 8, 619. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.T.; Khadke, S.; Korsholm, K.S.; Perrie, Y.; Rades, T.; Andersen, P.; Foged, C.; Christensen, D. The Administration Route Is Decisive for the Ability of the Vaccine Adjuvant CAF09 to Induce Antigen-Specific CD8(+) T-Cell Responses: The Immunological Consequences of the Biodistribution Profile. Journal of controlled release : official journal of the Controlled Release Society 2016, 239, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, D.A.; Christensen, D.; Muñoz, C.; Singh, S.; Locke, E.; Andersen, P.; Zavala, F. Robust Antibody and CD8+ T-Cell Responses Induced by P. Falciparum CSP Adsorbed to Cationic Liposomal Adjuvant CAF09 Confer Sterilizing Immunity against Experimental Rodent Malaria Infection. npj Vaccines 2017, 2, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Monnery, B.D.; Wright, M.; Cavill, R.; Hoogenboom, R.; Shaunak, S.; Steinke, J.H.G.; Thanou, M. Cytotoxicity of Polycations: Relationship of Molecular Weight and the Hydrolytic Theory of the Mechanism of Toxicity. International journal of pharmaceutics 2017, 521, 249–258. [Google Scholar] [CrossRef]
- Naumenko, E.; Akhatova, F.; Rozhina, E.; Fakhrullin, R. Revisiting the Cytotoxicity of Cationic Polyelectrolytes as a Principal Component in Layer-by-Layer Assembly Fabrication. Pharmaceutics 2021, 13, 1230. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Zhou, H.; Nian, Q.-G.; Yang, Y.; Qin, C.-F.; Tang, R. Robust Vaccine Formulation Produced by Assembling a Hybrid Coating of Polyethyleneimine–Silica. Chemical Science 2016, 7, 1753–1759. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, N.C.; Brinckmann, S.A.; Gartlan, K.H.; Puthia, M.; Svanborg, C.; Krashias, G.; Eisenbarth, S.C.; Flavell, R.A.; Sattentau, Q.J.; Wegmann, F. Polyethyleneimine Is a Potent Systemic Adjuvant for Glycoprotein Antigens. International Immunology 2014, 26, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Eusébio, D.; Paul, M.; Biswas, S.; Cui, Z.; Costa, D.; Sousa, Â. Mannosylated Polyethylenimine-Cholesterol-Based Nanoparticles for Targeted Delivery of Minicircle DNA Vaccine against COVID-19 to Antigen-Presenting Cells. International Journal of Pharmaceutics 2024, 654, 123959. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Liu, Y.; Chen, Z.; Li, W.; Liu, Y.; Wang, L.; Liu, Y.; Wu, X.; Ji, Y.; Zhao, Y.; et al. Surface-Engineered Gold Nanorods: Promising DNA Vaccine Adjuvant for HIV-1 Treatment. Nano letters 2012, 12, 2003–2012. [Google Scholar] [CrossRef]
- Zhao, D.; Yu, S.; Sun, B.; Gao, S.; Guo, S.; Zhao, K. Biomedical Applications of Chitosan and Its Derivative Nanoparticles. Polymers 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Pathak, K.; Misra, S.K.; Sehgal, A.; Singh, S.; Bungau, S.; Najda, A.; Gruszecki, R.; Behl, T. Biomedical Applications of Quaternized Chitosan. Polymers 2021, 13, 2514. [Google Scholar] [CrossRef] [PubMed]
- Chuang, C.-C.; Tsai, M.-H.; Yen, H.; Shyu, H.-F.; Cheng, K.; Chen, X.; Chen, C.; Young, J.; Kau, J.-H. A Fucoidan-Quaternary Chitosan Nanoparticle Adjuvant for Anthrax Vaccine as an Alternative to CpG Oligodeoxynucleotides. Carbohydrate Polymers 2020, 229, 115403. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Zhang, S.; Chen, Y.; Wang, F.; Jiang, W. Preparation and Characterization of Curdlan-Chitosan Conjugate Nanoparticles as Mucosal Adjuvants for Intranasal Influenza H1N1 Subunit Vaccine. International Journal of Biological Macromolecules 2024, 266, 131289. [Google Scholar] [CrossRef]
- Fan, B.; Gu, J.; Deng, B.; Guo, W.; Zhang, S.; Li, L.; Li, B. Positively Charged-Amylose-Entangled Au-Nanoparticles Acting as Protein Carriers and Potential Adjuvants to SARS-CoV-2 Subunit Vaccines. ACS Appl. Mater. Interfaces 2023, 15, 29982–29997. [Google Scholar] [CrossRef]
- Altay Benetti, A.; Tan, E.Y.Z.; Chang, Z.W.; Bae, K.H.; Thwin, M.T.; Muthuramalingam, R.P.K.; Liao, K.-C.; Wan, Y.; Ng, L.F.P.; Renia, L.; et al. Design and Characterization of a New Formulation for the Delivery of COVID-19-mRNA Vaccine to the Nasal Mucosa. Vaccines (Basel) 2024, 12, 409. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Yu, W.; Shen, L.; Yan, W.; Qi, J.; Hu, T. Mucosal SARS-CoV-2 Nanoparticle Vaccine Based on Mucosal Adjuvants and Its Immune Effectiveness by Intranasal Administration. ACS Appl. Mater. Interfaces 2023, 15, 35895–35905. [Google Scholar] [CrossRef] [PubMed]
- Pati, R.; Shevtsov, M.; Sonawane, A. Nanoparticle Vaccines Against Infectious Diseases. Front Immunol 2018, 9, 2224–2224. [Google Scholar] [CrossRef] [PubMed]
- Heng, W.T.; Yew, J.S.; Poh, C.L. Nanovaccines against Viral Infectious Diseases. Pharmaceutics 2022, 14, 2554. [Google Scholar] [CrossRef] [PubMed]
- Priyanka; Abusalah, M. A.H.; Chopra, H.; Sharma, A.; Mustafa, S.A.; Choudhary, O.P.; Sharma, M.; Dhawan, M.; Khosla, R.; Loshali, A.; et al. Nanovaccines: A Game Changing Approach in the Fight against Infectious Diseases. Biomedicine & Pharmacotherapy 2023, 167, 115597. [Google Scholar] [CrossRef]
- Singh, A. Eliciting B Cell Immunity against Infectious Diseases Using Nanovaccines. Nat. Nanotechnol. 2021, 16, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Torres-Sangiao, E.; Holban, A.M.; Gestal, M.C. Advanced Nanobiomaterials: Vaccines, Diagnosis and Treatment of Infectious Diseases. Molecules 2016, 21, 867. [Google Scholar] [CrossRef] [PubMed]
- Look, M.; Bandyopadhyay, A.; Blum, J.S.; Fahmy, T.M. Application of Nanotechnologies for Improved Immune Response against Infectious Diseases in the Developing World. Advanced Drug Delivery Reviews 2010, 62, 378–393. [Google Scholar] [CrossRef] [PubMed]
- Bo, Y.; Wang, H. Materials-Based Vaccines for Infectious Diseases. WIREs Nanomedicine and Nanobiotechnology 2022, 14, e1824. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.; Liu, H.; Zhang, S.; Hu, J.; Zhang, L. Delivery of Nanovaccine towards Lymphoid Organs: Recent Strategies in Enhancing Cancer Immunotherapy. J Nanobiotechnol 2021, 19, 389. [Google Scholar] [CrossRef]
- Mao, C.; Gorbet, M.-J.; Singh, A.; Ranjan, A.; Fiering, S. In Situ Vaccination with Nanoparticles for Cancer Immunotherapy: Understanding the Immunology. International Journal of Hyperthermia 2020, 37, 4–17. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Mooney, D.J. Biomaterial-Assisted Targeted Modulation of Immune Cells in Cancer Treatment. Nature Mater 2018, 17, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-X.; Jia, Y.-B.; Huang, Y.-R.; Liu, H.-N.; Sun, X.-M.; Cai, T.; Liu, R.-T.; Xu, Z.P. Efficient Delivery of Clay-Based Nanovaccines to the Mouse Spleen Promotes Potent Anti-Tumor Immunity for Both Prevention and Treatment of Lymphoma. Nano Res. 2021, 14, 1326–1334. [Google Scholar] [CrossRef]
- Den Haan, J.M.M.; Martinez-Pomares, L. Macrophage Heterogeneity in Lymphoid Tissues. Semin Immunopathol 2013, 35, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Tavares, A.J.; Poon, W.; Zhang, Y.-N.; Dai, Q.; Besla, R.; Ding, D.; Ouyang, B.; Li, A.; Chen, J.; Zheng, G.; et al. Effect of Removing Kupffer Cells on Nanoparticle Tumor Delivery. Proc. Natl. Acad. Sci. U.S.A. 2017, 114. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Li, X.; Ma, H.; Zhang, J.; Yu, D.; Zhao, R.; Yu, S.; Nie, G.; Wang, H. In Situ Transforming RNA Nanovaccines from Polyethylenimine Functionalized Graphene Oxide Hydrogel for Durable Cancer Immunotherapy. Nano Lett. 2021, 21, 2224–2231. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Huang, D.; Saw, P.E.; Song, E. Turning Cold Tumors Hot: From Molecular Mechanisms to Clinical Applications. Trends in Immunology 2022, 43, 523–545. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ribeiro, A.M. Chapter 1 - Immunoadjuvants for Cancer Immunotherapy. In Nanomedicine in Cancer Immunotherapy; Kesharwani, P., Ed.; Academic Press, 2024; pp. 1–36 ISBN 978-0-443-18770-4.
- Park, J.-H.; Gu, L.; Von Maltzahn, G.; Ruoslahti, E.; Bhatia, S.N.; Sailor, M.J. Biodegradable Luminescent Porous Silicon Nanoparticles for in Vivo Applications. Nature Mater 2009, 8, 331–336. [Google Scholar] [CrossRef]
- Zhu, S.; Yao, R.; Li, Y.; Zhao, P.; Ren, C.; Du, X.; Yao, Y. Lysosomal Quality Control of Cell Fate: A Novel Therapeutic Target for Human Diseases. Cell Death Dis 2020, 11, 817. [Google Scholar] [CrossRef]
- Wojnilowicz, M.; Glab, A.; Bertucci, A.; Caruso, F.; Cavalieri, F. Super-Resolution Imaging of Proton Sponge-Triggered Rupture of Endosomes and Cytosolic Release of Small Interfering RNA. ACS Nano 2019, 13, 187–202. [Google Scholar] [CrossRef]
- Benjaminsen, R.V.; Mattebjerg, M.A.; Henriksen, J.R.; Moghimi, S.M.; Andresen, T.L. The Possible “Proton Sponge ” Effect of Polyethylenimine (PEI) Does Not Include Change in Lysosomal pH. Molecular Therapy 2013, 21, 149–157. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Wen, J.; Wang, W.; Hu, Z.; Ling, C.; Zhao, Z.; Cheng, Y.; Chang, Y.; Xu, M.; Jin, Z.; et al. Peptide-Driven Proton Sponge Nano-Assembly for Imaging and Triggering Lysosome-Regulated Immunogenic Cancer Cell Death. Advanced Materials 2024, 36, 2307679. [Google Scholar] [CrossRef] [PubMed]
- Gao, A.; Hu, X.; Saeed, M.; Chen, B.; Li, Y.; Yu, H. Overview of Recent Advances in Liposomal Nanoparticle-Based Cancer Immunotherapy. Acta Pharmacol Sin 2019, 40, 1129–1137. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.Z.; Ahmad, J.; Alasmary, M.Y.; Abdel-Wahab, B.A.; Warsi, M.H.; Haque, A.; Chaubey, P. Emerging Advances in Cationic Liposomal Cancer Nanovaccines: Opportunities and Challenges. Immunotherapy 2021, 13, 491–507. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Liu, C.; Qi, Z.; Qiu, T.; Zhang, J.; Yang, H. Engineering Customized Nanovaccines for Enhanced Cancer Immunotherapy. Bioactive Materials 2024, 36, 330–357. [Google Scholar] [CrossRef] [PubMed]
- Alfagih, I.M.; Aldosari, B.; AlQuadeib, B.; Almurshedi, A.; Alfagih, M.M. Nanoparticles as Adjuvants and Nanodelivery Systems for mRNA-Based Vaccines. Pharmaceutics 2020, 13, 45. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ribeiro, A.M.; Mathiazzi, B.I.; Pérez-Betancourt, Y. Cationic Nanostructures as Adjuvants for Vaccines. In Vaccine Design: Methods and Protocols, Volume 3. Resources for Vaccine Development; Thomas, S., Ed.; Springer US: New York, NY, 2022; ISBN 978-1-07-161892-9. [Google Scholar]
- Fan, Y.-N.; Li, M.; Luo, Y.-L.; Chen, Q.; Wang, L.; Zhang, H.-B.; Shen, S.; Gu, Z.; Wang, J. Cationic Lipid-Assisted Nanoparticles for Delivery of mRNA Cancer Vaccine. Biomater. Sci. 2018, 6, 3009–3018. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wu, Y.; Xiang, J.; Wang, H.; Zhuang, Q.; Wei, T.; Cao, Z.; Gu, Q.; Liu, Z.; Peng, R. Fluoroalkane Modified Cationic Polymers for Personalized mRNA Cancer Vaccines. Chemical Engineering Journal 2023, 456, 140930. [Google Scholar] [CrossRef]
- Carson, C.S.; Becker, K.W.; Garland, K.M.; Pagendarm, H.M.; Stone, P.T.; Arora, K.; Wang-Bishop, L.; Baljon, J.J.; Cruz, L.D.; Joyce, S.; et al. A Nanovaccine for Enhancing Cellular Immunity via Cytosolic Co-Delivery of Antigen and polyIC RNA. Journal of Controlled Release 2022, 345, 354–370. [Google Scholar] [CrossRef]
- Sultan, H.; Salazar, A.M.; Celis, E. Poly-ICLC, a Multi-Functional Immune Modulator for Treating Cancer. Seminars in Immunology 2020, 49, 101414. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, K.; Zhao, Z.; Shan, X.; Wang, Y.; Feng, Z.; Li, B.; Luo, C.; Chen, X.; Sun, J. Self-Adjuvanting Polyguanidine Nanovaccines for Cancer Immunotherapy. ACS Nano 2024, 18, 7136–7147. [Google Scholar] [CrossRef]
- Luo, X.; Chen, X.; Ma, R.; Fu, Z.; Liu, Z.; Su, Q.; Fu, H.; Yang, Y.; Xue, W. Cancer Cell Membrane Proteins-Encapsulated Nanovaccine Enhances Cancer Immunotherapy and Prevention by Provoking Antigen-Specific Cellular Immunity via the Dendritic Cell-Targeted Delivery. Chemical Engineering Journal 2024, 481, 148611. [Google Scholar] [CrossRef]
- Diao, L.; Liu, M. Rethinking Antigen Source: Cancer Vaccines Based on Whole Tumor Cell/Tissue Lysate or Whole Tumor Cell. Advanced Science 2023, 10, 2300121. [Google Scholar] [CrossRef]
- Pan, Y.; Wu, X.; Liu, L.; Zhao, C.; Zhang, J.; Yang, S.; Pan, P.; Huang, Q.; Zhao, X.; Tian, R.; et al. Genetically Engineered Cytomembrane Nanovaccines for Cancer Immunotherapy. Adv Healthcare Materials 2024, 13, 2400068. [Google Scholar] [CrossRef]
- Liu, M.; Feng, Y.; Lu, Y.; Huang, R.; Zhang, Y.; Zhao, Y.; Mo, R. Lymph-Targeted High-Density Lipoprotein-Mimetic Nanovaccine for Multi-Antigenic Personalized Cancer Immunotherapy. Sci. Adv. 2024, 10, eadk2444. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, H.; Yang, X.; Yu, Q.; Wang, H.; Zhang, L.; Zhao, Y.; Zhu, D. Cascade Carrier-Free Nanoparticles Forming In Situ Nanovaccines for Synergistic Photothermal-Immunotherapy of Cancer. Adv Funct Materials 2024, 2401489. [Google Scholar] [CrossRef]
- Yu, X.; Dai, Y.; Zhao, Y.; Qi, S.; Liu, L.; Lu, L.; Luo, Q.; Zhang, Z. Melittin-Lipid Nanoparticles Target to Lymph Nodes and Elicit a Systemic Anti-Tumor Immune Response. Nat Commun 2020, 11, 1110. [Google Scholar] [CrossRef]
- Zhang, H.-Q.; Sun, C.; Xu, N.; Liu, W. The Current Landscape of the Antimicrobial Peptide Melittin and Its Therapeutic Potential. Front. Immunol. 2024, 15, 1326033. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ribeiro, A.M. Antimicrobial Peptides and Their Assemblies. Future Pharmacology 2023, 3, 763–788. [Google Scholar] [CrossRef]
- Pérez-Betancourt, Y.; Zaia, R.; Evangelista, M.F.; Ribeiro, R.T.; Roncoleta, B.M.; Mathiazzi, B.I.; Carmona-Ribeiro, A.M. Characterization and Differential Cytotoxicity of Gramicidin Nanoparticles Combined with Cationic Polymer or Lipid Bilayer. Pharmaceutics 2022, 14, 2053. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, M.; Liu, X.; Chai, M.; Diao, L.; Ma, L.; Nie, S.; Xu, M.; Wang, Y.; Mo, F.; et al. Probiotics Formulation and Cancer Nanovaccines Show Synergistic Effect in Immunotherapy and Prevention of Colon Cancer. iScience 2023, 26, 107167. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-H.; Su, J.-Y.; Li, Y.-M. Rational Design of T-Cell- and B-Cell-Based Therapeutic Cancer Vaccines. Acc. Chem. Res. 2022, 55, 2660–2671. [Google Scholar] [CrossRef] [PubMed]
- Gurunathan, S.; Thangaraj, P.; Wang, L.; Cao, Q.; Kim, J.-H. Nanovaccines: An Effective Therapeutic Approach for Cancer Therapy. Biomedicine & Pharmacotherapy 2024, 170, 115992. [Google Scholar] [CrossRef]
Figure 1.
Immune responses Th1 and/or Th2 elicited by cationic nanostructures carrying ovalbumin (OVA) depend on affinity between OVA antigen and adjuvant. Molecules used to build the nanostructures were: the cationic lipid dioctadecyldimethyl ammonium bromide (DODAB), the cationic polymer poly (diallyldimethyl ammonium chloride) (PDDA) and the biocompatible polymer poly (methyl methacrylate) (PMMA). Reproduced from reference [16].
Figure 1.
Immune responses Th1 and/or Th2 elicited by cationic nanostructures carrying ovalbumin (OVA) depend on affinity between OVA antigen and adjuvant. Molecules used to build the nanostructures were: the cationic lipid dioctadecyldimethyl ammonium bromide (DODAB), the cationic polymer poly (diallyldimethyl ammonium chloride) (PDDA) and the biocompatible polymer poly (methyl methacrylate) (PMMA). Reproduced from reference [16].
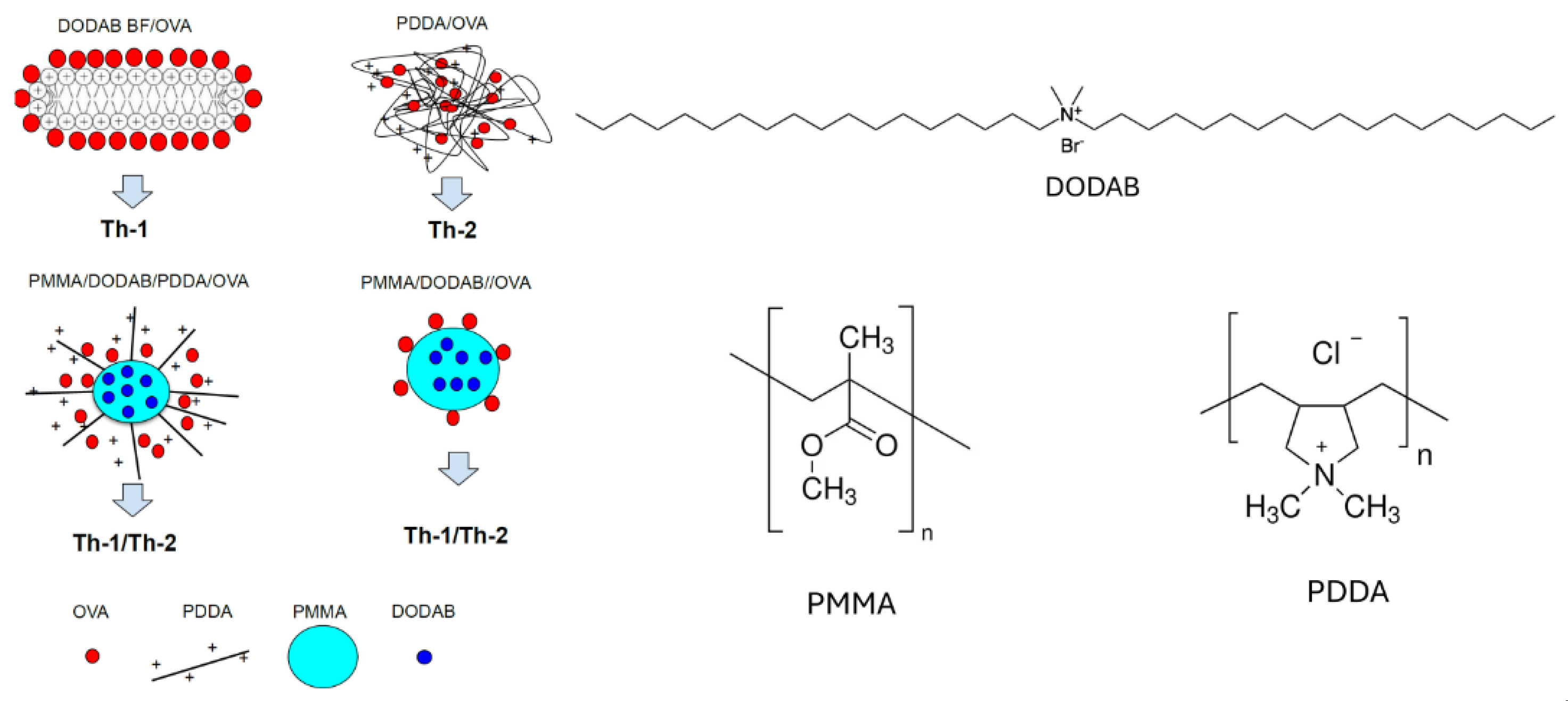
Figure 2.
Routes followed by cationic nanoparticles (NPs) acting as immunoadjuvants. NPs uptake induces reactive oxygen species (ROS) and receptor activation, such as the TLR-4 leading to the transcription of pro-inflammatory genes, translated as proinflammatory cytokines and co-stimulatory molecules. The proton–sponge effect delivers antigen to the cytosol where the antigen starts to be processed to yield smaller peptides to be presented as such by MHC molecules which will be transported to the cell surface. Improved cytosolic antigen delivery and the immune-stimulating properties of cationic NPs result in efficient differentiation of antigen-specific T-cells sub-sets. Reproduced from [78].
Figure 2.
Routes followed by cationic nanoparticles (NPs) acting as immunoadjuvants. NPs uptake induces reactive oxygen species (ROS) and receptor activation, such as the TLR-4 leading to the transcription of pro-inflammatory genes, translated as proinflammatory cytokines and co-stimulatory molecules. The proton–sponge effect delivers antigen to the cytosol where the antigen starts to be processed to yield smaller peptides to be presented as such by MHC molecules which will be transported to the cell surface. Improved cytosolic antigen delivery and the immune-stimulating properties of cationic NPs result in efficient differentiation of antigen-specific T-cells sub-sets. Reproduced from [78].
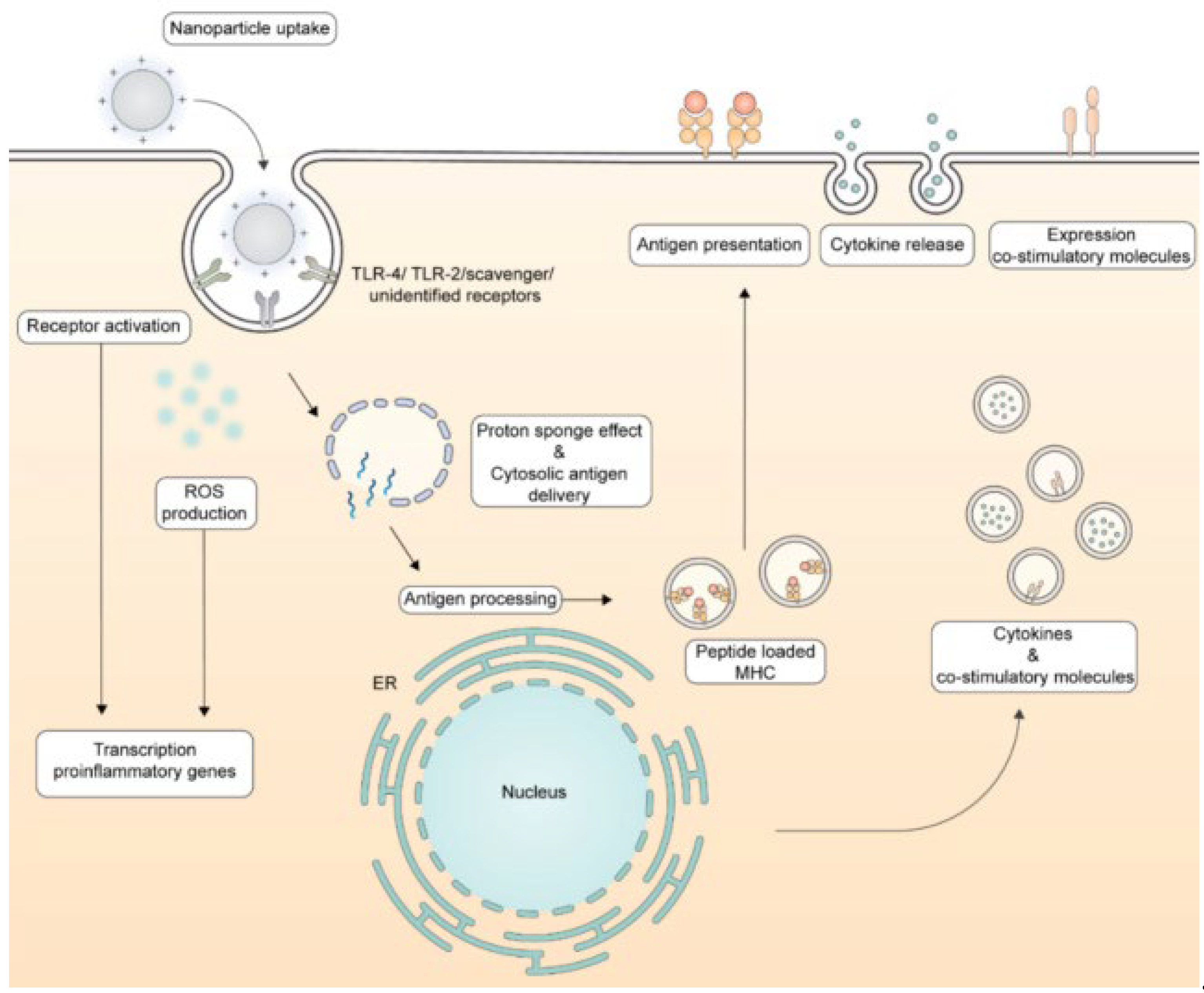
Figure 3.
Virus-like particles (VLPs) and the immune system. On step 1, VLP interacts with pattern recognition receptors (PRRs) on the dendritic cell (DC), such as a toll-like receptor (TLR), so that VLP is taken up by the DC. On step 2, occurs maturation of DC and secretion of pro-inflammatory cytokines (TNF-α and IL-1β) with recruitment of additional antigen presenting cells (APCs), including DCs and macrophages. On step 3, VLP taken up by the DC are then enzymatically digested into short peptides, which binds to major histocompatibility complex class I (MHC I) and class II (MHC II) and are transported onto surface of the DC. On step 4, short peptide displayed on MHC II, together with CD40 and CD80/86, interacts with T cell receptor (TCR), CD40L, and CD28 present on naïve helper T cell (Th), respectively, followed by proliferation and differentiation of Th cells into type 1 (Th1) and type 2 (Th2) Th cells. On step 5, peptides displayed on MHC I of the DC, along with CD40 and CD80/86 also interact with TCR, CD40L, and CD28 present on the naïve cytotoxic T lymphocyte (CTL). Aided by Th1, naïve CTL proliferates and differentiates into effector and memory CTLs, providing immediate and long-lasting cellular immunity, respectively. On step 6: Naïve B cell interacts with intact VLP carried over by blood stream or DC via B cell receptor (BCR). Aided by Th2, the B cell differentiates into plasma B cells which actively secrete antibodies, and memory B cells which provide long-lasting humoral immunity against the antigen present on the VLP. Reproduced from reference [120] with permission from John Wiley and Sons, Copyright 2022.
Figure 3.
Virus-like particles (VLPs) and the immune system. On step 1, VLP interacts with pattern recognition receptors (PRRs) on the dendritic cell (DC), such as a toll-like receptor (TLR), so that VLP is taken up by the DC. On step 2, occurs maturation of DC and secretion of pro-inflammatory cytokines (TNF-α and IL-1β) with recruitment of additional antigen presenting cells (APCs), including DCs and macrophages. On step 3, VLP taken up by the DC are then enzymatically digested into short peptides, which binds to major histocompatibility complex class I (MHC I) and class II (MHC II) and are transported onto surface of the DC. On step 4, short peptide displayed on MHC II, together with CD40 and CD80/86, interacts with T cell receptor (TCR), CD40L, and CD28 present on naïve helper T cell (Th), respectively, followed by proliferation and differentiation of Th cells into type 1 (Th1) and type 2 (Th2) Th cells. On step 5, peptides displayed on MHC I of the DC, along with CD40 and CD80/86 also interact with TCR, CD40L, and CD28 present on the naïve cytotoxic T lymphocyte (CTL). Aided by Th1, naïve CTL proliferates and differentiates into effector and memory CTLs, providing immediate and long-lasting cellular immunity, respectively. On step 6: Naïve B cell interacts with intact VLP carried over by blood stream or DC via B cell receptor (BCR). Aided by Th2, the B cell differentiates into plasma B cells which actively secrete antibodies, and memory B cells which provide long-lasting humoral immunity against the antigen present on the VLP. Reproduced from reference [120] with permission from John Wiley and Sons, Copyright 2022.
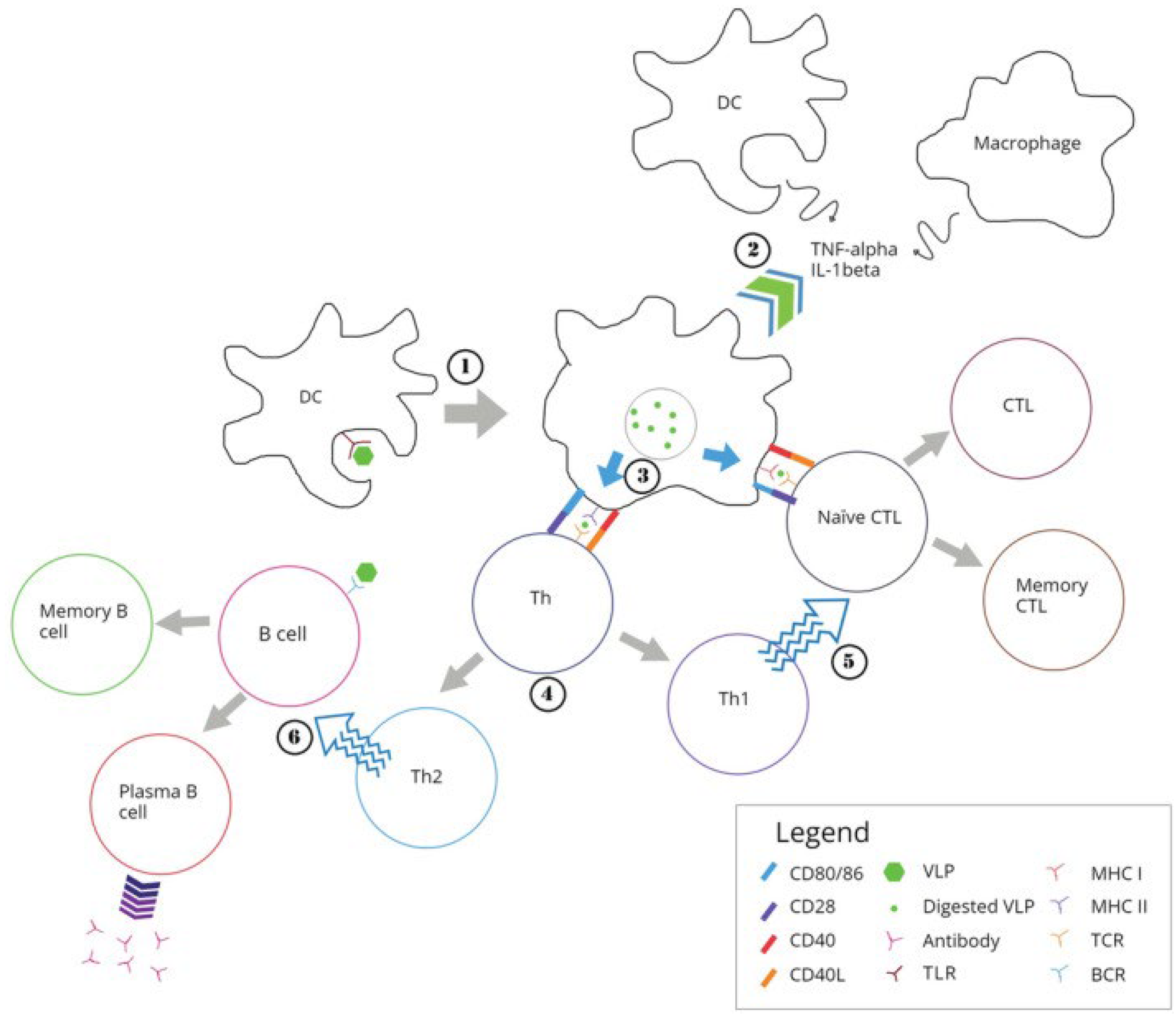
Figure 5.
Cell viability of macrophages (J774A.1) or fibroblasts (L929) as affected by three different adjuvants containing dioctadecyl dimethyl ammonium bromide (DODAB) at 3 (a,c) and 24 h interaction time (b,d). The adjuvants were DODAB bilayer fragments (DODAB BF) (in yellow), DODAB bilayer vesicles (DODAB LV) (in blue) and poly (methyl methacrylate) (PMMA)/DODAB hybrid NPs (in gray). The DODAB concentration used for the three different adjuvants (PMMA/DODAB NPs, DODAB BF and DODAB LV) was indicated by the red arrow and corresponded to 0.08 mM DODAB. Reproduced from reference [16].
Figure 5.
Cell viability of macrophages (J774A.1) or fibroblasts (L929) as affected by three different adjuvants containing dioctadecyl dimethyl ammonium bromide (DODAB) at 3 (a,c) and 24 h interaction time (b,d). The adjuvants were DODAB bilayer fragments (DODAB BF) (in yellow), DODAB bilayer vesicles (DODAB LV) (in blue) and poly (methyl methacrylate) (PMMA)/DODAB hybrid NPs (in gray). The DODAB concentration used for the three different adjuvants (PMMA/DODAB NPs, DODAB BF and DODAB LV) was indicated by the red arrow and corresponded to 0.08 mM DODAB. Reproduced from reference [16].
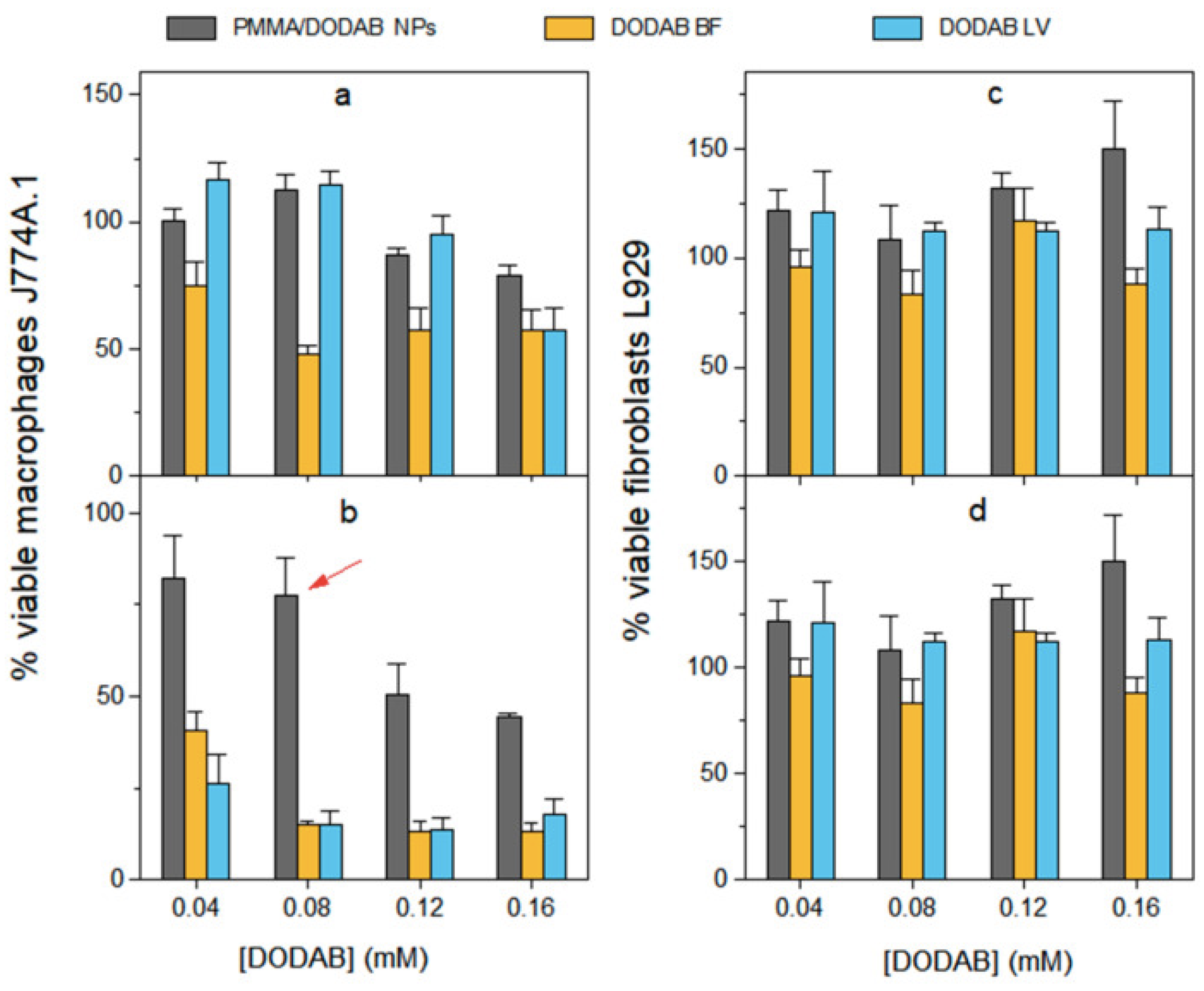
Figure 6.
Dose-dependent cytotoxicity of cationic hybrid NPs made of biocompatible polymer and traces of cationic polymer and cationic lipid. (A) PMMA/PDDA/DODAB NPs visualized by scanning electron microscopy (SEM) and synthesized at 0.56 M methyl methacrylate (MMA), 5 mg·mL−1 PDDA and 2 mM DODAB and exhaustively dialyzed before use; the bar represents 1000 nm. Reproduced from reference [65]. (B) Effect of poly diallyl dimethylammonium chloride (PDDA) concentration in hybrid NPs of biocompatible poly methyl methacrylate (PMMA) on L929 fibroblasts (a) or J774A.1 macrophages viability (%) after 3 or 24 h interaction (b). Reproduced from reference [15]. (C) PDDA dose-dependent lysis of fibroblasts in culture; the cells were submitted to increasing PDDA concentrations from (a) to (d) and are visualized by scanning electron microscopy (SEM) of fibroblasts L929 incubated for 3 h with: (a) culture medium; (b) 0.01; (c) 0.1; (d) 1 mg·mL−1 PDDA. The red arrows indicate holes on the cells at 0.1 mg·mL−1 PDDA [20].
Figure 6.
Dose-dependent cytotoxicity of cationic hybrid NPs made of biocompatible polymer and traces of cationic polymer and cationic lipid. (A) PMMA/PDDA/DODAB NPs visualized by scanning electron microscopy (SEM) and synthesized at 0.56 M methyl methacrylate (MMA), 5 mg·mL−1 PDDA and 2 mM DODAB and exhaustively dialyzed before use; the bar represents 1000 nm. Reproduced from reference [65]. (B) Effect of poly diallyl dimethylammonium chloride (PDDA) concentration in hybrid NPs of biocompatible poly methyl methacrylate (PMMA) on L929 fibroblasts (a) or J774A.1 macrophages viability (%) after 3 or 24 h interaction (b). Reproduced from reference [15]. (C) PDDA dose-dependent lysis of fibroblasts in culture; the cells were submitted to increasing PDDA concentrations from (a) to (d) and are visualized by scanning electron microscopy (SEM) of fibroblasts L929 incubated for 3 h with: (a) culture medium; (b) 0.01; (c) 0.1; (d) 1 mg·mL−1 PDDA. The red arrows indicate holes on the cells at 0.1 mg·mL−1 PDDA [20].
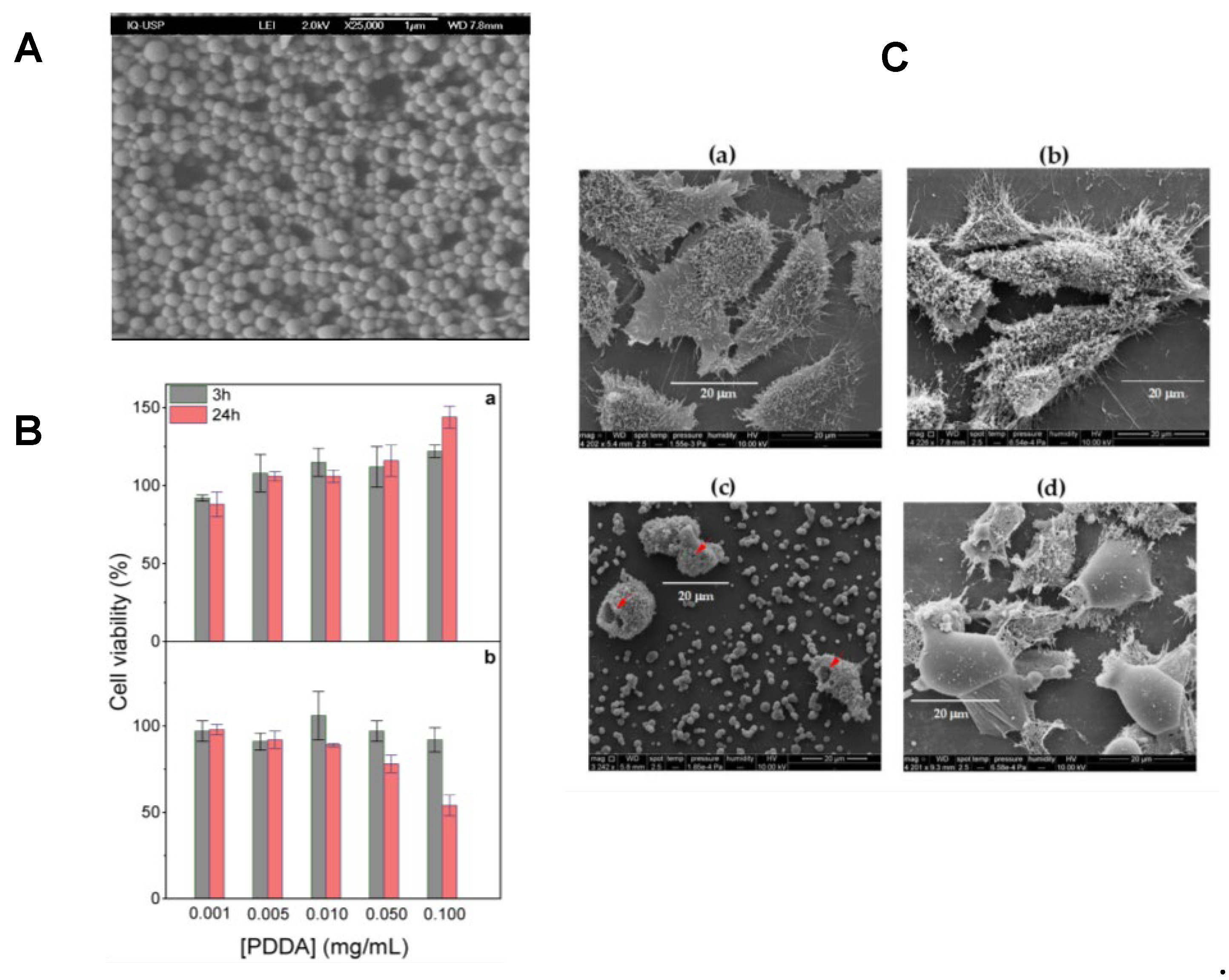
Figure 7.
Vaccine thermostability imparted by the layer-by-layer cationic assembly PEI-SiO2-PEI on JEV. (A) Schematic illustration of the assembly of PEI–silica hybrid nanocoatings on the vaccine. (B) Zeta-potential of JEV, and JEV coated with PEI, PEI–SiO2 and PEI–SiO2–PEI layers. (C) The vaccine protection against storage granted by silicification of JEV at pH 7.0. The modified vaccine could be used efficiently after long-term exposure at room temperature, which would facilitate vaccine transport and storage without a cold chain. Reproduced from reference [159].
Figure 7.
Vaccine thermostability imparted by the layer-by-layer cationic assembly PEI-SiO2-PEI on JEV. (A) Schematic illustration of the assembly of PEI–silica hybrid nanocoatings on the vaccine. (B) Zeta-potential of JEV, and JEV coated with PEI, PEI–SiO2 and PEI–SiO2–PEI layers. (C) The vaccine protection against storage granted by silicification of JEV at pH 7.0. The modified vaccine could be used efficiently after long-term exposure at room temperature, which would facilitate vaccine transport and storage without a cold chain. Reproduced from reference [159].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



