
Submitted:
18 June 2024
Posted:
19 June 2024
You are already at the latest version
Abstract
The demand for a more sustainable agriculture, coupled with the need for optimized crop productivity, has driven the use of microorganisms for the biocontrol of diseases and pests, as well as for growth promotion. The use of plant growth-promoting rhizobacteria (PGPR) and plant growth-promoting fungi (PGPF) in Brazilian production systems has become increasingly common, with the number of microbial inoculant registrations in the country growing every year. PGPR and PGPF occupy various niches in the rhizosphere, playing a crucial role in soil nutrient cycling, as well as influencing various plant physiological processes. In this review, we address the main mechanisms used by these microbial agents in growth promotion and the use of strain co-inoculation as a strategy to enhance the efficacy of these products. Subsequently, we conduct an analysis of the available inoculants in Brazil, detailing the microorganisms available for the main Brazilian crops, and the prospects of this market regarding research and development of new products for the coming years based on the current challenges faced.
Keywords:
Beneficial microorganisms
; Co-inoculation
; Soil microbiome
1. Introduction
The need for more sustainable agriculture with less environmental impact has transformed the way we use chemical fertilizers, once seen as a solution for maintaining productivity. Today, we understand the impact of these inputs on soil, the environment, and human health. Additionally, these inputs are expensive and inefficient and a large portion of nutriends ends up getting retained in the soil in insoluble forms [1].
In this context, the use of growth-promoting microorganisms is increasing, which is a more sustainable practice that can help better utilize nutrients in the soil and reduce chemical fertilization. The interaction between plants and microorganisms is essential for plant development. Microorganisms contribute to the chemical composition of the rhizospheric environment, influencing growth, morphology, and root permeability through the release of metabolites [2].
Several processes are regulated by plant growth-promoting rhizobacteria (PGPR) and plant growth-promoting fungi (PGPF), such as nutrient mineralization and solubilization, production of growth-regulating phytohormones, induction of systemic resistance, increased resistance to abiotic stresses, inhibition of plant diseases through antibiotic production, reduction of ethylene in roots, production of siderophores, and iron chelation [3].
The use of growth-promoting microorganisms has become a trend in Brazilian agriculture. According to the National Association of Inoculant Producers and Importers, in 2022, 134.9 million doses were delivered by associated companies, which represents an increase of 30 million doses in comparison to 2021. In this panorama, there is growing interest in research and development of new inoculants with more effective, multifunctional strains and formulations that increase the shelf life of these products. Co-inoculation of microbial agents with different mechanisms of action and synergistic interactions is one of the strategies used, with an increase in registrations of these products in recent years. This practice was used in 35% of the soybeans grown in the country in the 22/23 harvest [4].
Based on this, this article aims to provide a review of the main mechanisms used by PGPR and PGPF in promoting growth and thus discuss the state of the Brazilian inoculant market (Figure 1.). Furthermore, it aims to describe the crops with the most registered microbial inoculants and their prevalent mechanisms of action. In conclusion, it will also discuss market trends based on challenges in Brazilian agriculture.
2. Plant Growth-Promoting Rhizobacteria (PGPR)
The beneficial interactions between plants and PGPR have emerged as a promising research area for the development of sustainable agriculture and the pursuit of more efficient cultivation methods. These bacteria can positively influence plant development through several mechanisms (Table 1), which will be discussed further on.
2.1. Biofertilization: A Sustainable Approach to Enhance Soil Fertility
Nitrogen stands as a critical limiting factor for plant growth; its availability in the soil directly influences agricultural production. Biological nitrogen fixation (BNF) has proven to be an effective strategy to reduce reliance on nitrogen-based fertilizers. BNF is a complex biological phenomenon in which certain bacteria convert atmospheric gaseous nitrogen into plant-assimilable nitrogenous compounds. This process not only provides nitrogen to plants but also enriches the soil with essential nutrients, boosting its fertility and structure [17]. Several genera of rhizobacteria are capable of fixing nitrogen, e.g., Rhizobium, Bradyrhizobium, Azospirillum, and Azotobacter, which are the most commonly used as biofertilizers [18].
Rhizobium and Bradyrhizobium establish nodules on the roots of leguminous plants, where biological nitrogen fixation takes place. These bacteria possess an enzyme called nitrogenase, which breaks the triple bond of gaseous nitrogen (N₂) and transforms it into ammonia (NH₃), a compound assimilable by plants. The ammonia is then incorporated into organic molecules, such as amino acids and proteins, within the plant [19].
In turn, Azospirillum and Azotobacter are free-living bacteria that also possess the nitrogenase enzyme, although they do not form nodules on roots. The N₂ molecules are converted into NH₃ inside the bacterial cells. The ammonia produced in the reaction can be released into the surrounding environment, increasing soil fertility [20]. BFN by rhizobacteria enhances soil quality, especially by providing a constant source of nitrogen [21].
Another essential element for the growth and development of plants is phosphorus (P). When phosphate-based fertilizers are applied to the soil, they are not always fully utilized by plants, and the excess phosphorus tends to bind to surrounding metallic cations, such as calcium (Ca²⁺), iron (Fe³⁺), and aluminum (Al³⁺), forming insoluble phosphates. These phosphates (calcium phosphate, iron phosphate, and aluminum phosphate) have low solubility in water, limiting P availability to plants [22].
Bacillus, Pseudomonas, Acinetobacter, and Pantoea species play an important role in improving P availability in the soil, especially in its insoluble forms. These bacteria are referred to as PSBs (phosphate-solubilizing bacteria) and produce and release several organic acids (formic, acetic, propionic, lactic, and succinic acids). The hydroxyl and carboxyl groups of the acids released by these bacteria react with phosphate minerals, dissolving and converting them into assimilable forms by the plants [23]. From the production of these organic acids, these bacteria also enhance the availability of other nutrients, such as potassium (K), which is one of the most crucial nutrients for plants. Potassium is typically applied in soluble forms, but it undergoes rapid fixation in the soil [24].
The combination of nitrogen-fixing and phosphorus and potassium-solubilizing bacteria presents an alternative for reducing the application of chemical inputs in agricultural fertilization, thereby rendering agriculture more sustainable and cost-effective [25].
2.2. Protection Against Oxidative Stress in Adverse Environmental Conditions
Rhizobacteria also play a crucial role in the mitigation of the oxidative stress effects on plants that occur due to an imbalance between the production of reactive oxygen species (ROS) and the inherent capacity of plants to neutralize them through antioxidant systems [26]. The accumulation of ROS, highly reactive molecules, results in oxidative damage to vital cellular components, including lipids, proteins, and nucleic acids in plant cells. Oxidative stress caused by ROS can lead to cellular deterioration, compromised photosynthetic efficiency, and inhibition of plant growth.
Various abiotic factors contribute to the establishment of this imbalance, such as high soil salinity, environmental pollutants, water deficiency, toxic substances, and even inherent characteristics of the environment [27].
Antioxidant enzymes such as glutathione reductase (GR), ascorbate peroxidase (APX), dehydroascorbate reductase (DHAR), monodehydroascorbate reductase (MDHAR), catalase (CAT), peroxidase (POX), and superoxide dismutase (SOD) neutralize ROS through oxidation, reduction, and dismutation processes of O2 into H2O2. Rhizobacteria Bacillus subtilis, Pseudomonas fluorescens, Pseudomonas putida, among others, help modulate the plant’s antioxidant defense system and enhance the activity of antioxidant enzymes [28]. Thus, plant tolerance to adverse conditions, e.g., drought, high temperatures, and pollution is enhanced, and plants that are more resilient when facing oxidative stress tend to maintain optimized nutrient absorption, reducing the need for excessive amounts of fertilizers [29].
2.3. Production of Phytohormones by Rhizobacteria
Phytohormones control plant growth, development, and responses to environmental stimuli. Some beneficial rhizobacteria possess the remarkable ability to synthesize and release phytohormones, which will positively influence plant growth and health [30].
Some examples are auxins and cytokinins, two phytohormones that work in a coordinated manner to stimulate the growth of both above-ground parts and roots. The increase in root development, for instance, leads to an expansion of the absorption area, enhancing the efficiency in resource uptake from the soil, and consequently promoting a more robust and healthier development of plants [31]. Gibberellins, another group of phytohormones of great importance, are responsible for stem elongation, leaf expansion, floral development, seed germination, and fruit development [32].
Other phytohormones produced by rhizobacteria, such as abscisic acid, prove to enhance plant tolerance to stress conditions like drought and high temperatures. Abscisic acid regulates stomatal movements, contributing to the containment of water loss [30]. Furthermore, salicylic acid triggers the plant’s defense system, providing increased resistance to pathogens and pests [33].
2.4. Production of ACC Deaminase
Ethylene is a gaseous hormone produced by plants and plays a vital role in their development and adaptation to adverse conditions. During biotic stresses, such as pathogen attacks, the increase in ethylene levels triggers a series of defense responses, including the production of phytoalexins and the activation of resistance genes [34]. Similarly, in abiotic stresses such as drought and salinity, ethylene assists plants in survival by adjusting growth, closing stomata to reduce water loss, and promoting the synthesis of antioxidant enzymes to combat oxidative stress. However, when ethylene levels exceed what is needed, undesirable effects can occur, such as inhibition of root and stem growth, reduction in photosynthetic rate, and even induction of premature senescence. Additionally, excess ethylene can lead to premature leaf and flower drop, compromising the plant’s ability to photosynthesize and reproduce. Therefore, precise regulation of ethylene levels is crucial to ensure optimal growth and development of plants [35].
Some PGPR produce ACC deaminase, an enzyme that reduces ethylene concentrations in the plant by cleaving the precursor aminocyclopropane carboxylic acid (ACC) into ammonia and α-ketobutyrate. This mechanism helps reduce damage caused by various stresses, particularly environmental ones, such as in saline soils [36], heavy metals [37], and water stress [38].
2.5. Chemical Signals: Volatile Organic Compounds (VOCs)
Volatile organic compounds (VOCs) are chemical substances widely released by most living organisms. Rhizobacteria can produce certain VOCs that promote growth promotion and the induction of systemic resistance [39].
These substances are released by bacteria into the surrounding environment and encompass a variety of categories, including alcohols, benzenoids, aldehydes, alkenes, acids, esters, terpenoids, and ketones. These compounds act as chemical signals, enabling bidirectional communication (between plants and microorganisms in the rhizosphere). Among the genera of rhizobacteria identified as VOC producers are species of Bacillus, Serratia, Enterobacter, and Pseudomonas [40].
The multiple categories of compounds and the intricate biochemical pathways involved in their signaling make the complete elucidation of these mechanisms a challenge. However, studies indicate that microbial VOCs are associated with promoting plant growth, possibly by modulating the synthesis and/or metabolism of phytohormones produced by rhizobacteria or by the plants themselves [30].
2.6. Production of Siderophores
Siderophores are secreted molecules that can be used both as biofertilizers and/or biocontrollers. As biofertilizers, they optimize the availability of iron in the root zone, and at the same time, their biocontroller activity relies on competing with phytopathogens for access to iron [41].
From a structural perspective, a typical siderophore presents one or more well-known functional groups such as hydroxamate, catecholate, or carboxylate. These functional groups establish bonds with iron ions, forming stable complexes. Although siderophores are widely recognized for their affinity with iron, it is important to emphasize that some of these organic molecules can also form complexes with other metals, including zinc, copper, manganese, and other metals [42].
There are two theories regarding how plants absorb iron from microbial siderophores. According to the first theory, siderophores with high redox potential donate iron (Fe²⁺) to the plant’s transport system after being reduced. This happens when siderophores with Fe³⁺ are transported to the roots, where reduction occurs in the apoplast, retaining Fe²⁺ and increasing iron concentrations in the roots. Following the second theory, microbial siderophores can bind to iron in the soil and exchange ligands with plant phytosiderophores, a process dependent on various factors such as complex stability, concentrations, and root environment conditions [43].
In addition, the high affinity of siderophores for iron results in limiting the access of this element by other microorganisms, suppressing the population of phytopathogenic agents in the rhizosphere. The reduction in disease incidence strengthens plant health and reduces the need for high doses of chemical control agents [44].
3. Plant Growth-Promoting Fungi (PGPF)
Some genera of saprophytic fungi present in the rhizosphere are capable of promoting growth by colonizing the plant’s roots, primarily through increased nutrient absorption, phytohormone production, and induction of systemic resistance. Among the most common are Aspergillus, Fusarium, Penicillium, Piriformospora, Phoma, and Trichoderma [45]. Here, we focus on fungi from Trichoderma genus and arbuscular mycorrhizal fungi (AMF). The latter are obligatory symbionts of plant roots and have a notable effect on nutrient acquisition and resistance to abiotic stress [46] (Table 2).
3.1. Trichoderma spp. as Biocontrol Agents and Fertilizers
Fungi of the genus Trichoderma have remarkable abilities in the biological control of pathogenic fungal agents that harm plant growth. This activity in controlling fungi is well-documented in the literature and various mechanisms of action are described.[57].
This effect can occur through direct or indirect mechanisms. Direct mechanisms rely on competition for resources, such as space and nutrients. Trichoderma spp. colonize surfaces and substrates, occupying spaces and consuming nutrients.
It limits the growth of pathogens, restricting their establishment in plants. Chitinases and Beta-1,3-glucanases produced by Trichoderma spp. are vital keys to its action against other fungi. The production of these enzymes is part of the process known as mycoparasitism, where, upon recognizing chemical signals from the cell wall of the pathogen, Trichoderma hyphae grow towards and wrap around the hyphae of the pathogenic fungus. They then initiate the secretion of enzymes that degrade and create pores in the cell wall, through which Trichoderma obtains nutrients [58,59].
Regarding indirect mechanisms, the activation of the plant’s defense system is observed. This is because Trichoderma spp. emit chemical signals that stimulate the production of defensive compounds in plants, such as phytoalexins and enzymes [60,61].
Organic compounds, such as VOCs, are also produced by Trichoderma spp. and influence the germination of fungal spores, inhibiting their development. VOCs produced by Trichoderma koningiopsis T-51 inhibited the mycelial growth of Botrytis cinerea by 73.78% and of Fusarium oxysporum by 43.68%. Additionally, there was a reduction in conidial germination and a delay in germ tube elongation [49].
It is important to highlight that the production of metabolites can vary between different species and strains of Trichoderma, as well as the environmental conditions. These metabolites play critical roles in the ecology of Trichoderma and their interactions with plants and other microorganisms, consolidating their role in controlling plant diseases and promoting soil health. Additionally, these fungi also play a vital role in improving nutrient and water uptake [62,63,64].
Out of 251 Trichoderma isolates from the Amazon rainforest soil, 49 demonstrated phosphate solubilization capacity. It was observed the production of organic acids, such as lactic acid, fumaric acid, ascorbic acid, malic acid, gluconic acid, D-isocitric acid, phytic acid, and citric acid. Additionally, two strains showed growth-promoting activity in soybean plants in a greenhouse setting [47]. High solubilization activity and growth promotion were also confirmed for T. asperellum UFT 201 in soybean cultivations in a greenhouse, and these results were later replicated in a field experiment [65,66].
The combination of these action mechanisms confers significant benefits to the plant. After selecting four strains with positive inhibitory activity against F. oxysporum, phosphate solubilization, and IAA production, an evaluation of tomato seed inoculation confirmed an increase in chlorophyll levels, aerial part length, fresh and dry weight of both the aerial part and roots, and a reduction in wilt disease caused by F. oxysporum ranging from 10 to 30% [67].
3.2. Arbuscular Mycorrhizal Fungi: Extensions of Roots in Soil
AMF have a symbiotic relationship with plant roots [62]. Intracellular hyphae form arbuscules, specialized structures that penetrate the root epidermis to exchange nutrients with hosts [68]. AMF provide many benefits to plants, such as increased photosynthetic rates, improved soil quality, influence on atmospheric CO2 fixation [69], enhanced nutrient absorption (particularly P), increased water uptake, protection against pathogens, assistance in obtaining micro and macronutrients [70].
The benefits arise from AMF functioning as extensions of roots, penetrating other parts of the rhizosphere. Additionally, their hyphae are thinner compared to roots, allowing them to access a greater soil volume. For protection against water deficit, AMF hyphae enhance access to small soil pockets [71]. Also, the presence of phytohormones like abscisic acid modulates plant mechanisms related to water deficit, such as aquaporins and transpiration [72].
AMF can also increase flood tolerance, improving growth and assisting in phosphorus absorption through osmotic adjustment performed by the fungi [73].
Heat stress is also attenuated by AMF, which increases nutrient and water absorption, along with improved photosynthetic rates. Other effects are the accumulation of proline and sugars, sodium reduction, increased carbon and homeostasis [74]. These mechanisms, aiding in extreme temperature tolerance, also contribute to salinity tolerance [75]. Therefore, AMF plays a pivotal role in terrestrial ecosystems’ microbiomes, helping host plants and ecosystem maintenance [76].
Among its roles in plant health regarding enhanced nutrient obtention, P absorption is the most thoroughly described and recognized as the primary regulator of the association. In conditions of high available concentrations of this mineral in the soil, the plant tends to reduce its investment in the association, whereas, under conditions of low availability, plant investment is higher. This enhanced P absorption occurs directly, through the absorption and transport of P via the hyphae to the arbuscules, and indirectly, by stimulating phosphate-solubilizing bacteria in the soil [77].
This stimulation occurs in the region known as the mycorrhizosphere, which is the zone around roots colonized by mycorrhizal fungi, directly influenced by hyphal and root exudation [78]. Composed of hyphae with a diameter of approximately 0.2 μm, much finer than roots but significantly denser, the mycorrhizosphere is broader than the rhizosphere and acts as a matrix, facilitating interactions between bacteria and fungi. Extraradical fungal hyphae form the hyphosphere, a region influenced by AMF hyphae, that bring about physical, chemical, and biological alterations. Examples of these alterations include soil particle aggregation and the direct delivery of water to the host plant [79,80].
Fungal hyphae release carbon-rich compounds promoting bacterial growth, and, on the other hand, hyphae can also produce compounds that signal or inhibit other microorganisms’ growth [81], affect soil pH, maintain a liquid film, and serve as bacterial concentration nodes, creating a microbiome that supports soil maintenance [82].
Through hyphal exudation of signaling molecules and compounds, AMF can influence the surrounding soil, modifying the local microbiota, referred to as the hyphosphere effect. It can also impact weathering, where exudates act as chelators, destabilizing mineral surfaces [83,84]. Thus, the hyphosphere provides an excellent habitat for other microorganisms, supporting the soil microbiome and influencing various soil processes [85].
4. Co-Inoculation of Beneficial Microorganisms
In recent years, there has been increased interest in combining microorganisms with similar activities but different mechanisms of action. This management strategy has been primarily studied in the control of diseases caused by nematodes, fungi, and insects, resulting in a reduction in the use of chemical products and a lower selection of resistant pathogens [86,87].
The interaction between growth-promoting microorganisms does not always result in the improvement of plant development or the biocontrol of pathogens; it is subject to several factors, such as the genetic variability of native bacteria, hosts, and environmental factors, such as light, temperature, and the organic matter of the soil [88].
These interactions are classified as synergistic, antagonistic, or non-interactive/additive, according to the observed effect. Synergy occurs when a combined effect is observed, meaning that co-inoculation provides a greater effect compared to the application of these agents individually. On the other hand, in antagonism, this effect is negative, whereas in non-interactive interactions, there is no impact compared to the isolated application of each microorganism [89].
An example of a synergistic interaction is the co-inoculation between AMF and PGPR, which has been examined by several studies, especially nitrogen-fixing bacteria (NFB). In general, gains in biomass and minerals were achieved, particularly in nitrogen and phosphorus rates [73,90].
The activity of AMF and NFB in the rhizosphere is essential for plant nutrition, and various factors contribute to a tripartite symbiosis. These microorganisms do not compete for the same colonization sites, indicating coexistence and possibly functional interactions. Furthermore, the inoculation of AMF has already been shown to be important in establishing NFB and improving nodulation, as nodulation is dependent on high levels of P, which can be increased by AMF colonization (Meena et al., 2018). It is also worth noting the importance of the correct combination of strains to achieve synergy in the co-inoculation of these microorganisms [91] (Table 3).
5. Biological Inoculants Registered in Brazil
According to the Brazilian Ministry of Agriculture (MAPA), there are 636 inoculants with 713 registrations (some inoculants are registered for more than one crop) in Brazil, covering 37 crops, ranging from vegetables such as lettuce, cabbage, and potatoes, but primarily grains [98]. Approximately 50% of these registrations are designated for soybean cultivation, the most widely planted crop in the country, covering an area of about 45 million hectares [99]. Bean and maize, also among the most planted crops in the country, have 73 and 70 registrations respectively. Jack bean (35), peanut (36), and wheat (23) close the list of crops with the most inoculant registrations. The remaining registrations are divided among the remaining 31 crops, with lettuce standing out with 8 (Figure 2).
Among these registrations, 622 consist of formulations with a single microorganism, with a significant portion being NFB such as Bradyrhizobium spp., Azospirillum brasilense, and Rhizobium spp. For soybeans, for example, there are 189 registrations of products with Bradyrhizobium japonicum, 57 for Bradyrhizobium Elkani, and 52 with the co-inoculation of B. japonicum and B. elkani. For beans, 65 out of 73 registrations are for Rhizobium tropici and 6 for Azospirillum brasilense. In maize cultivation, 45 out of 70 registrations contain A. brasilense [98]. The introduction of these inoculants, which began in 1964, played a pivotal role in rendering Brazil competitive in grain production and reducing the necessity for nitrogen fertilization. In recent years, formulations with co-inoculation of two NFB strains have also been implemented, resulting in increased yields compared to individual inoculation. This co-inoculation strategy is anticipated to gain further prominence in the coming years [100]. According to Telles et al. (2023), during the 2019/2020 harvest season, the co-inoculation technology of Bradyrhizobium spp. and A. brasilense was employed in 25% of all soybean-cultivated areas in Brazil, resulting in a cost-saving of 15.2 billion dollars through the replacement of urea application with biological nitrogen fixation. Additionally, it generated an estimated profit of 914 million dollars [101].
Analyzing a subset of the most important crops (Table 4), in addition to NFB, there are also registrations of plant growth-promoting rhizobacteria (PGPR) from the genus Bacillus, and Pseudomonas fluorescens. In certain cases, co-inoculation between two strains is implemented, such as B. subtilis and B. megaterium for soybean, maize, and bean crops, B. megaterium and Lysinobacillus sp for maize, strains that promote growth by providing phosphorus availability, and B. licheniformis and B. aryabhatai act on increasing stress tolerance in maize. In addition to P. fluorescens and Azospirillum brasilense for maize and soybean crops, there are also strains of B. subtilis and B. elkani for soybean. In these latter two cases, there are co-inoculation of strains with different activities, biological nitrogen fixation, and phosphorus solubilization. Co-inoculation with three strains is also observed, such as B. subtilis, B. amyloliquefaciens, and B. pumilus for promoting growth in soybean and maize, and P. fluorescense, B. amyloliquefaciens, and Priestia megaterium; B. subtilis, B. elkani, and Parabhurkodelia nodosa for soybean. This last species, introduced in the year 2024 to the market, isolated only in Brazilian soils so far, exhibits nitrogen fixation activity, among other beneficial interactions such as phosphate solubilization, hormone production, siderophore synthesis, and ACC deaminase activity [102].
In terms of fungi, the number of registrations is reduced to 5 registrations of Trichoderma for soybean and 5 registrations of AMF for soybean and maize. Among these, one formulation contains eight strains (4 T. harzianum; 3 T. asperelloides, and 1 T. koningiopsis), which are registered for soybean, alongside the other Trichoderma products. As for AMF, the registrations are for soybean and maize, containing a strain of Rhizophagus intraradices or a double inoculation with Rhizoglomus intraradices and Claroideoglomus claroideum (Table 4).
Brazil’s biological product market is experiencing substantial growth, NFB based inoculants well-established in the production of various crops, most notably maize and soybeans. Nonetheless, the adoption of inoculants with alternative growth-promoting mechanisms remains in its early stages.
An illustrative instance of this emerging trend is the utilization of AMF and PSB. Analogous to NFB in the context of nitrogen fertilization, AMF and PSB have the potential to revolutionize phosphate fertilization practices in Brazil. More than 50% of phosphorus fertilizers applied in Brazilian soils are imported, generating a variable cost according to the dollar exchange rate and are poorly utilized. It is believed that only 30% of this phosphorus is absorbed by plants, with a significant portion of the surplus accumulating in the soil. Brazilian soils contain elevated concentrations of P in insoluble forms, a consequence of decades of fertilization in iron (Fe) and aluminum (Al)-rich soils, culminating in the rapid immobilization of this vital nutrient. It is estimated that by 2018, Brazil has accumulated a staggering 33.4 teragrams (Tg) of phosphorus in its soil. This estimation is derived from an analysis of the ratio between phosphorus inputs from organic manure and mineral fertilizers and the phosphorus harvested by crops annually, dating back to 1967—a seminal year that marks the commencement of intensive phosphate fertilizer application. This accumulated phosphorus reservoir possesses an estimated economic value of US$ 22 billion [103].
The inoculation of AMF and PSB is an important alternative to enhance the efficiency of phosphorus (P) absorption, reducing the need for application and utilizing the soil’s legacy P [103,104]. However, the application of these microorganisms is still not widely disseminated in Brazil. For example, AMF products available are imported and do not utilize native strains from the country, highlighting a gap in investment in technologies for strain research and the development of production systems for these microorganisms.
Like phosphate fertilizers, potassium fertilizers are largely imported, around 97%, a percentage that has been increasing in recent years due to the accelerated growth of agriculture [99]. It is estimated that the efficiency of applied potassium (K) in the soil is 66%, with a considerable portion remaining stocked up in the soil and 13% lost through erosion and leaching processes [105]. Brazil is a country with extensive areas of mineral resources; rocks containing potassium have the potential as a source of this nutrient for agriculture. However, strategies are needed for the rapid release of this nutrient for plant uptake [106,107].
The exploration of strains with high K solubilization activity and the development of biological products focusing on the availability of this nutrient is a gap to be filled in the Brazilian market. Obviously, Brazil’s solution to its dependency on the importation of chemical inputs is not solely the use of biological products, but also increased input production, as envisioned in the National Fertilizer Plan, aiming to reduce imports from 85% to 45% by 2050 [108].
Other inoculants that are expected to see increased registrations in the coming years are stress mitigators. The effects of adverse weather conditions are already being felt in Brazilian agriculture. According to the National Supply Company (Conab), the estimate for the harvest in the 2023/24 season is 8% lower compared to the previous season, a reduction of around 25.7 million tons. This loss of productivity is directly related to the delayed onset of rains in the Midwest, Southeast, and Matopiba regions, high temperatures, irregular and poorly distributed rainfall, and periods of drought lasting more than 20 days. In addition to the impacts on productivity, these effects may, in the long term, affect the cultivable regions for Brazil’s main crops, reducing areas suitable for soybean and maize cultivation [109]. Currently, there are already efficient strains on the Brazilian market for reducing water stress, registered for soybean and maize crops. However, given the magnitude of the economic impact that these climate changes can cause, there is a trend towards increased investment in research and development of biological products to mitigate the effects of climate change on agriculture and a greater variety of strains and formulations covering more cultures.
6. Conclusion
The use of PGPR and PGPF as biological inoculants in Brazilian agriculture has proven to be efficient and capable of generating cost savings for the production system, as evidenced by the individual or co-inoculated use of Bradyrhizobium spp. and Azospirillum brasilense. Given the current challenges faced in agricultural production in Brazil, such as costs and impacts related to the application of chemical fertilizers and losses due to climate change, there is a trend towards increased registration and application of inoculants that are effective in reducing fertilization and increasing plant tolerance to environmental stresses. In this scenario, AMF have great potential, as they not only play an important role in nutrient acquisition but also can help reduce the impact of abiotic stress.
Author Contributions
Conceptualization and project administration, M.F.L.A.; G.A.; validation, G.A. and L.A.; supervision, G.A.; data curation, M.F.L.A.; formal investigation and writing—original draft preparation, E.T.G.N; S.F.A; M.E.A.P.; K.G.G.; and M.F.L.A.
Funding
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001
Data Availability Statement
Data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Baweja P, Kumar S, Kumar G. Fertilizers and pesticides: Their impact on soil health and environment. Soil Biology, Cham: Springer International Publishing; 2020, p. 265–85.
- Olaetxea M, De Hita D, Garcia CA, Fuentes M, Baigorri R, Mora V, et al. Hypothetical framework integrating the main mechanisms involved in the promoting action of rhizospheric humic substances on plant root- and shoot- growth. Appl Soil Ecol 2018, 123, 521–537. [Google Scholar] [CrossRef]
- Etesami H, Adl SM. Plant growth-promoting rhizobacteria (PGPR) and their action mechanisms in availability of nutrients to plants. Environmental and Microbial Biotechnology, Singapore: Springer Singapore; 2020, p. 147–203.
- ANPII. Painel Interno ANPII; 2022. [cited 2024 may 20]. Available from: https://www.anpii.org.br/estatisticas/.
- Zeffa DM, Perini LJ, Silva MB, de Sousa NV, Scapim CA, Oliveira ALM de, et al. Azospirillum brasilense promotes increases in growth and nitrogen use efficiency of maize genotypes. PLoS One 2019, 14, e0215332. [Google Scholar] [CrossRef]
- Zilli JÉ, Pacheco RS, Gianluppi V, Smiderle OJ, Urquiaga S, Hungria M. Biological N2 fixation and yield performance of soybean inoculated with Bradyrhizobium. Nutr Cycling Agroecosyst 2021, 119, 323–336. [Google Scholar] [CrossRef]
- Oteino, N. Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Frontiers in microbiology. 2015. [CrossRef]
- afza MR, Aliasgharzad N, Khoshru B. P solubilizing potential of some plant growth promoting bacteria used as ingredient in phosphatic biofertilizers with emphasis on growth promotion of Zea mays L. Geomicrobiol J 2020, 37, 327–335. [Google Scholar] [CrossRef]
- Sun Y, Wu J, Shang X, Xue L, Ji G, Chang S, et al. Screening of siderophore-producing bacteria and their effects on promoting the growth of plants. Curr Microbiol 2022, 79, 150. [Google Scholar] [CrossRef]
- Uzma M, Iqbal A, Hasnain S. Drought tolerance induction and growth promotion by indole acetic acid producing Pseudomonas aeruginosa in Vigna radiate. PLoS ONE n.d.; 2022.
- Sandhya V, Ali SZ, Grover M, Reddy G, Venkateswarlu B. Effect of plant growth promoting Pseudomonas spp. on compatible solutes, antioxidant status and plant growth of maize under drought stress. Plant Growth Regul 2010, 62, 21–30. [Google Scholar] [CrossRef]
- Afzal, A. Rock phosphate solubilization by plant growth-promoting Bacillus velezensis and its impact on wheat growth and yield. Geomicrobiology Journal 2023, 40, 131–142. [Google Scholar] [CrossRef]
- Mosela M, Andrade G, Massucato LR, de Araújo Almeida SR, Nogueira AF, de Lima Filho RB, et al. Bacillus velezensis strain Ag75 as a new multifunctional agent for biocontrol, phosphate solubilization and growth promotion in maize and soybean crops. Sci Rep 2022, 12, 15284. [Google Scholar] [CrossRef]
- Rath M, Mitchell TR, Gold SE. Volatiles produced by Bacillus mojavensis RRC101 act as plant growth modulators and are strongly culture-dependent. Microbiol Res 2018, 208, 76–84. [CrossRef]
- Gowtham, HG. Induction of drought tolerance in tomato upon the application of ACC deaminase producing plant growth promoting rhizobacterium Bacillus subtilis Rhizo SF 48. Microbiological Research 2020, 234. [Google Scholar]
- 16. Carlos M-HJ, Stefani P-VY, Janette A-M, Melani M-SS, Gabriela P-O. Assessing the effects of heavy metals in ACC deaminase and IAA production on plant growth-promoting bacteria. Microbiol Res, 2016; 188–189, 53–61. [CrossRef]
- Zeng Q, Ding X, Wang J, Han X, Iqbal HMN, Bilal M. Insight into soil nitrogen and phosphorus availability and agricultural sustainability by plant growth-promoting rhizobacteria. Environ Sci Pollut Res Int 2022, 29, 45089–45106. [Google Scholar] [CrossRef] [PubMed]
- Shamseldin, A. Future outlook of transferring biological nitrogen fixation (BNF) to cereals and challenges to retard achieving this dream. Curr Microbiol 2022, 79. [Google Scholar] [CrossRef]
- Maitra S, Praharaj S, Brestic M, Sahoo RK, Sagar L, Shankar T, et al. Rhizobium as biotechnological tools for green solutions: An environment-friendly approach for sustainable crop production in the modern era of climate change. Curr Microbiol 2023, 80, 219. [Google Scholar] [CrossRef]
- 20. Zhang W, Chen Y, Huang K, Wang F, Mei Z. Molecular mechanism and agricultural application of the NifA-NifL system for nitrogen fixation. Int J Mol Sci. [CrossRef]
- Aasfar A, Bargaz A, Yaakoubi K, Hilali A, Bennis I, Zeroual Y, et al. Nitrogen fixing Azotobacter species as potential soil biological enhancers for crop nutrition and yield stability. Front Microbiol 2021, 12, 628379. [Google Scholar] [CrossRef]
- Timofeeva A, Galyamova M, Sedykh S. Prospects for using phosphate-solubilizing microorganisms as natural fertilizers in agriculture. Plants 2022, 11, 2119. [Google Scholar] [CrossRef] [PubMed]
- Sarmah R, Sarma AK. Phosphate solubilizing microorganisms: A review. Commun Soil Sci Plant Anal 2023, 54, 1306–1315. [Google Scholar] [CrossRef]
- Pandey D, Kehri HK, Zoomi I, Singh U, Chaudhri KL, Akhtar O. Potassium solubilizing microbes: Diversity, ecological significances and biotechnological applications. Sustainable Development and Biodiversity, Cham: Springer International Publishing; 2020, p. 263–86.
- Wang J, Li R, Zhang H, Wei G, Li Z. Beneficial bacteria activate nutrients and promote wheat growth under conditions of reduced fertilizer application. BMC Microbiol 2020, 20, 38. [Google Scholar] [CrossRef]
- 26. Dumanović J, Nepovimova E, Natić M, Kuča K, Jaćević V. The significance of reactive oxygen species and antioxidant defense system in plants: A concise overview. Front Plant Sci, 2021; 11. [CrossRef]
- Devireddy AR, Zandalinas SI, Fichman Y, Mittler R. Integration of reactive oxygen species and hormone signaling during abiotic stress. Plant J 2021, 105, 459–476. [Google Scholar] [CrossRef]
- Gowtham HG, Singh SB, Shilpa N, Aiyaz M, Nataraj K, Udayashankar AC, et al. Insight into recent progress and perspectives in improvement of antioxidant machinery upon PGPR augmentation in plants under drought stress: A review. Antioxidants (Basel) 2022, 11, 1763. [Google Scholar] [CrossRef]
- Batool T, Ali S, Seleiman MF, Naveed NH, Ali A, Ahmed K, et al. Plant growth promoting rhizobacteria alleviates drought stress in potato in response to suppressive oxidative stress and antioxidant enzymes activities. Sci Rep 2020, 10, 16975. [Google Scholar] [CrossRef] [PubMed]
- Orozco-Mosqueda MDC, Santoyo G, Glick BR. Recent advances in the bacterial phytohormone modulation of plant growth. Plants 2023, 12, 606. [Google Scholar] [CrossRef] [PubMed]
- Andrade LA, Santos CHB, Frezarin ET, Sales LR, Rigobelo EC. Plant Growth-Promoting Rhizobacteria for sustainable agricultural production. Microorganisms 2023, 11. [Google Scholar] [CrossRef] [PubMed]
- Hedden P, Sponsel V. A century of gibberellin research. J Plant Growth Regul 2015, 34, 740–760. [Google Scholar] [CrossRef]
- Urban L, Lauri F, Ben Hdech D, Aarrouf J. Prospects for increasing the efficacy of plant resistance inducers stimulating salicylic acid. Agronomy (Basel) 2022, 12, 3151. [Google Scholar] [CrossRef]
- Jasrotia S, Jasrotia R. Role of ethylene in combating biotic stress. Ethylene in Plant Biology, 2022; 388–397. [CrossRef]
- Chen H, Bullock DA Jr, Alonso JM, Stepanova AN. To fight or to grow: The balancing role of ethylene in plant abiotic stress responses. Plants 2021, 11, 33. [Google Scholar] [CrossRef] [PubMed]
- Choudhury AR, Trivedi P, Madhaiyan M, Choi J, Choi W, Park J-H, et al. ACC deaminase producing endophytic bacteria enhances cell viability of rice (Oryza sativa L.) under salt stress by regulating ethylene emission pathway. Environ Exp Bot 2023, 213, 105411. [Google Scholar] [CrossRef]
- Andy AK, Rajput VD, Burachevskaya M, Gour VS. Exploring the identity and properties of two bacilli strains and their potential to alleviate drought and heavy metal stress. Horticulturae 2023, 9, 46. [Google Scholar] [CrossRef]
- 38. Ojuederie OB, Babalola OO. Growth enhancement and extenuation of drought stress in maize inoculated with multifaceted ACC deaminase producing rhizobacteria. Front Sustain Food Syst. [CrossRef]
- Brilli F, Loreto F, Baccelli I. Exploiting plant volatile organic compounds (VOCs) in agriculture to improve sustainable defense strategies and productivity of crops. Front Plant Sci 2019, 10, 264. [Google Scholar] [CrossRef]
- Russo A, Pollastri S, Ruocco M, Monti MM, Loreto F. Volatile organic compounds in the interaction between plants and beneficial microorganisms. J Plant Interact 2022, 17, 840–852. [Google Scholar] [CrossRef]
- Hyder S, Rizvi ZF, los Santos-Villalobos S de, Santoyo G, Gondal A, Khalid N, et al. Applications of plant growth-promoting rhizobacteria for increasing crop production and resilience. J Plant Nutr 2023, 46, 2551–2580. [Google Scholar] [CrossRef]
- Saha M, Sarkar S, Sarkar B, Sharma BK, Bhattacharjee S, Tribedi P. Microbial siderophores and their potential applications: a review. Environ Sci Pollut Res Int 2016, 23, 3984–3999. [Google Scholar] [CrossRef] [PubMed]
- Ahmed E, Holmström SJM. Siderophores in environmental research: roles and applications: Siderophores in environmental research. Microb Biotechnol 2014, 7, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Gowtham HG, Singh SB, Shilpa N, Aiyaz M, Nataraj K, Udayashankar AC, et al. Insight into recent progress and perspectives in improvement of antioxidant machinery upon PGPR augmentation in plants under drought stress: A review. Antioxidants (Basel) 2022, 11, 1763. [Google Scholar] [CrossRef] [PubMed]
- Hossain MM, Sultana F, Islam S. Plant growth-promoting fungi (PGPF): Phytostimulation and induced systemic resistance. Plant-Microbe Interactions in Agro-Ecological Perspectives, Singapore: Springer Singapore; 2017, p. 135–91.
- Igiehon NO, Babalola OO. Biofertilizers and sustainable agriculture: exploring arbuscular mycorrhizal fungi. Appl Microbiol Biotechnol 2017, 101, 4871–4881. [Google Scholar] [CrossRef] [PubMed]
- Bononi L, Chiaramonte JB, Pansa CC, Moitinho MA, Melo IS. Phosphorus-solubilizing Trichoderma spp. from Amazon soils improve soybean plant growth. Sci Rep 2020, 10, 2858. [Google Scholar]
- Nieto-Jacobo, MF. Environmental growth conditions of Trichoderma spp. affects indole acetic acid derivatives, volatile organic compounds, and plant growth promotion. Frontiers in plant science. 2017. [Google Scholar]
- You J, Li G, Li C, Zhu L, Yang H, Song R, et al. Biological control and plant growth promotion by volatile organic compounds of Trichoderma koningiopsis T-51. J Fungi (Basel), 2022; 8. [CrossRef]
- Lee S, Yap M, Behringer G, Hung R, Bennett JW. Volatile organic compounds emitted by Trichoderma species mediate plant growth. Fungal Biol Biotechnol 2016, 3, 7. [Google Scholar] [CrossRef]
- Lei ZHAO, Zhang Y. Effects of phosphate solubilization and phytohormone production of Trichoderma asperellum Q1 on promoting cucumber growth under salt stress. Effects of phosphate solubilization and phytohormone production of Trichoderma asperellum Q1 on promoting cucumber growth under salt stress. Journal of Integrative Agriculture 2015, 1588–1597. [Google Scholar]
- Cely MVT, de Oliveira AG, de Freitas VF, de Luca MB, Barazetti AR, Dos Santos IMO, et al. Inoculant of arbuscular mycorrhizal fungi (Rhizophagus clarus) increase yield of soybean and cotton under field conditions. Front Microbiol 2016, 7, 720. [Google Scholar] [CrossRef]
- 53. Fayaz F, Zahedi M. Beneficial effects of arbuscular mycorrhizal fungi on wheat (Triticum aestivum L.) nutritional status and tolerance indices under soil salinity stress. Journal of Plant Nutrition, 2021; 185–201.
- 54. Hu Y, Xie W, Chen B. Arbuscular mycorrhiza improved drought tolerance of maize seedlings by altering photosystem II efficiency and the levels of key metabolites. Chem Biol Technol Agric, 2020; 7. [CrossRef]
- Al-Karaki GN, Williams M. Mycorrhizal mixtures affect the growth, nutrition, and physiological responses of soybean to water deficit. Acta Physiologiae Plantarum, 2021.
- Jumrani K, Bhatia VS, Kataria S, Alamri SA, Siddiqui MH, Rastogi A. Inoculation with arbuscular mycorrhizal fungi alleviates the adverse effects of high temperature in soybean. Plants 2022, 11, 2210. [Google Scholar] [CrossRef]
- Guzmán-Guzmán P, Kumar A, de Los Santos-Villalobos S, Parra-Cota FI, Orozco-Mosqueda MDC, Fadiji AE, et al. Trichoderma species: Our best fungal allies in the biocontrol of plant diseases-A review. Plants 2023, 12, 432. [Google Scholar] [CrossRef] [PubMed]
- Naher, L. , Yusuf, U. K., Ismail, A., & Hossain, K. Trichoderma spp.: a biocontrol agent for sustainable management of plant diseases. Pak J Bot 2014, 46, 1489–1493. 46,.
- Asad, SA. Mechanisms of action and biocontrol potential of Trichoderma against fungal plant diseases - A review. Ecol Complex 2022, 49, 100978. [Google Scholar] [CrossRef]
- Cumagun CJR. Advances in Formulation of Trichoderma for Biocontrol. Biotechnology and Biology of Trichoderma, Elsevier; 2014, p. 527–31.
- Mukhopadhyay R, Kumar D. Trichoderma: a beneficial antifungal agent and insights into its mechanism of biocontrol potential. Egypt J Biol Pest Contr 2020, 30. [Google Scholar] [CrossRef]
- Hermosa R, Cardoza RE, Rubio MB, Gutiérrez S, Monte E. Secondary metabolism and antimicrobial metabolites of Trichoderma. Biotechnology and Biology of Trichoderma, Elsevier; 2014, p. 125–37.
- Stewart A, Hill R. Applications of Trichoderma in Plant Growth Promotion. Biotechnology and Biology of Trichoderma, Elsevier; 2014, p. 415–28.
- Zeilinger, S. , Gruber, S. , Bansal, R., & Mukherjee, P. K. Secondary metabolism in Trichoderma–chemistry meets genomics. Fungal biology reviews 2016, 30, 74–90. [Google Scholar]
- Junior AFC, Chagas LFB, Colonia BSO, Miller LDO, De Oliveira JC. Trichoderma asperellum (UFT201) functions as a growth promoter for soybean plant. African Journal of Agricultural Research 2019, 14, 1772–1777. [Google Scholar] [CrossRef]
- Junior AFC, Chagas LFB, Colonia BSO, Miller LDO, De Oliveira JC. Efficiency of Trichoderma asperellum UFT 201 as plant growth promoter in soybean. Afr J Agric Res 2019, 14, 263–271. [Google Scholar] [CrossRef]
- Bader AN, Salerno GL, Covacevich F, Consolo VF. Native Trichoderma harzianum strains from Argentina produce indole-3 acetic acid and phosphorus solubilization, promote growth and control wilt disease on tomato (Solanum lycopersicum L.). J King Saud Univ Sci 2020, 32, 867–873. [Google Scholar] [CrossRef]
- Riaz M, Kamran M, Fang Y, Wang Q, Cao H, Yang G, et al. Arbuscular mycorrhizal fungi-induced mitigation of heavy metal phytotoxicity in metal contaminated soils: A critical review. J Hazard Mater 2021, 402, 123919. [Google Scholar] [CrossRef] [PubMed]
- Begum N, Qin C, Ahanger MA, Raza S, Khan MI, Ashraf M, et al. Role of arbuscular mycorrhizal fungi in plant growth regulation: Implications in abiotic stress tolerance. Front Plant Sci 2019, 10, 1068. [Google Scholar] [CrossRef]
- Etesami H, Jeong BR, Glick BR. Contribution of arbuscular mycorrhizal fungi, phosphate-solubilizing bacteria, and silicon to P uptake by plant. Front Plant Sci 2021, 12, 699618. [Google Scholar] [CrossRef]
- Chitarra W, Maserti B, Gambino G, Guerrieri E, Balestrini R. Arbuscular mycorrhizal symbiosis-mediated tomato tolerance to drought. Plant Signal Behav 2016, 11, e1197468. [Google Scholar] [CrossRef] [PubMed]
- Ouledali S, Ennajeh M, Ferrandino A, Khemira H, Schubert A, Secchi F. Influence of arbuscular mycorrhizal fungi inoculation on the control of stomata functioning by abscisic acid (ABA) in drought-stressed olive plants. S Afr J Bot 2019, 121, 152–158. [Google Scholar] [CrossRef]
- Wang Y, Qiu Q, Yang Z, Hu Z, Tam NF-Y, Xin G. Arbuscular mycorrhizal fungi in two mangroves in South China. Plant Soil 2010, 331, 181–191. [Google Scholar] [CrossRef]
- Diagne N, Ngom M, Djighaly PI, Fall D, Hocher V, Svistoonoff S. Roles of arbuscular mycorrhizal fungi on plant growth and performance: Importance in biotic and abiotic stressed regulation. Diversity (Basel) 2020, 12, 370. [Google Scholar] [CrossRef]
- Djighaly PI, Diagne N, Ngom M, Ngom D, Hocher V, Fall D, et al. Selection of arbuscular mycorrhizal fungal strains to improve Casuarina equisetifolia L. and Casuarina glauca Sieb. tolerance to salinity. Ann For Sci 2018, 75. [CrossRef]
- Qin Y, Zhang W, Feng Z, Feng G, Zhu H, Yao Q. Arbuscular mycorrhizal fungus differentially regulates P mobilizing bacterial community and abundance in rhizosphere and hyphosphere. Appl Soil Ecol 2022, 170, 104294. [Google Scholar] [CrossRef]
- Kobae, Y. Dynamic phosphate uptake in arbuscular mycorrhizal roots under field conditions. Front Environ Sci, 2019; 6. [Google Scholar] [CrossRef]
- Wang F, Zhang L, Zhou J, Rengel Z, George TS, Feng G. Exploring the secrets of hyphosphere of arbuscular mycorrhizal fungi: processes and ecological functions. Plant Soil 2022, 481, 1–22. [Google Scholar] [CrossRef]
- He J-D, Chi G-G, Zou Y-N, Shu B, Wu Q-S, Srivastava AK, et al. Contribution of glomalin-related soil proteins to soil organic carbon in trifoliate orange. Appl Soil Ecol 2020, 154, 103592. [Google Scholar] [CrossRef]
- Wang X-Q, Wang Y-H, Song Y-B, Dong M. Formation and functions of arbuscular mycorrhizae in coastal wetland ecosystems: A review. Ecosyst Health Sustain 2022, 8. [Google Scholar] [CrossRef]
- Haq IU, Zwahlen RD, Yang P, van Elsas JD. The response of Paraburkholderia terrae strains to two soil fungi and the potential role of oxalate. Front Microbiol 2018, 9. [Google Scholar] [CrossRef]
- Jansa J, Hodge A. Swimming, gliding, or hyphal riding? On microbial migration along the arbuscular mycorrhizal hyphal highway and functional consequences thereof. New Phytol 2021, 230, 14–16. [Google Scholar] [CrossRef] [PubMed]
- Bakker PAHM, Berendsen RL, Doornbos RF, Wintermans PCA, Pieterse CMJ. The rhizosphere revisited: root microbiomics. Front Plant Sci 2013, 4, 165. [Google Scholar] [CrossRef]
- Verbruggen E, Sheldrake M, Bainard LD, Chen B, Ceulemans T, De Gruyter J, et al. Mycorrhizal fungi show regular community compositions in natural ecosystems. ISME J 2018, 12, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Faghihinia M, Jansa J, Halverson LJ, Staddon PL. Hyphosphere microbiome of arbuscular mycorrhizal fungi: a realm of unknowns. Biol Fertil Soils 2023, 59, 17–34. [Google Scholar] [CrossRef]
- Spescha A, Weibel J, Wyser L, Brunner M, Hess Hermida M, Moix A, et al. Combining entomopathogenic Pseudomonas bacteria, nematodes and fungi for biological control of a below-ground insect pest. Agric Ecosyst Environ 2023, 348, 108414. [Google Scholar] [CrossRef]
- El-Sharkawy EES, Abdelrazik E. Biocontrol of Fusarium root rot in squash using mycorrhizal fungi and antagonistic microorganisms. Egypt J Biol Pest Contr 2022, 32. [Google Scholar] [CrossRef]
- Meena RS, Vijayakumar V, Yadav GS, Mitran T. Response and interaction of Bradyrhizobium japonicum and arbuscular mycorrhizal fungi in the soybean rhizosphere. Plant Growth Regul 2018, 84, 207–223. [Google Scholar] [CrossRef]
- Anuar MSK, Hashim AM, Ho CL, Wong M-Y, Sundram S, Saidi NB, et al. Synergism: biocontrol agents and biostimulants in reducing abiotic and biotic stresses in crop. World J Microbiol Biotechnol 2023, 39, 123. [Google Scholar] [CrossRef]
- Musyoka DM, Njeru EM, Nyamwange MM, Maingi JM. Arbuscular mycorrhizal fungi and Bradyrhizobium co-inoculation enhances nitrogen fixation and growth of green grams (Vigna radiata L.) under water stress. J Plant Nutr 2020, 43, 1036–1047. [CrossRef]
- Kavadia A, Omirou M, Fasoula DA, Louka F, Ehaliotis C, Ioannides IM. Co-inoculations with rhizobia and arbuscular mycorrhizal fungi alters mycorrhizal composition and lead to synergistic growth effects in cowpea that are fungal combination-dependent. Appl Soil Ecol 2021, 167, 104013. [Google Scholar] [CrossRef]
- De Almeida Leite R, Martins LC, Ferreira LV dos SF, Barbosa ES, Alves BJR, Zilli JE, et al. Co-inoculation of Rhizobium and Bradyrhizobium promotes growth and yield of common beans. Appl Soil Ecol 2022, 172, 104356. [Google Scholar] [CrossRef]
- Queiroz Rego, CH. Co-inoculation with Bradyrhizobium and Azospirillum increases yield and quality of soybean seeds. Agronomy Journal 2018, 110, 2302–2309. [Google Scholar] [CrossRef]
- Pérez-Rodriguez MM, Pontin M, Lipinski V, Bottini R, Piccoli P, Cohen AC. Pseudomonas fluorescens and Azospirillum brasilense Increase Yield and Fruit Quality of Tomato Under Field Conditions. J Soil Sci Plant Nutr 2020, 20, 1614–1624. [Google Scholar] [CrossRef]
- Mendes JBS, da Costa Neto VP, de Sousa CDA, de Carvalho Filho MR, Rodrigues AC, Bonifacio A. Trichoderma and bradyrhizobia act synergistically and enhance the growth rate, biomass and photosynthetic pigments of cowpea (Vigna unguiculata) grown in controlled conditions. Symbiosis 2020, 80, 133–143. [Google Scholar] [CrossRef]
- Qi R, Lin W, Gong K, Han Z, Ma H, Zhang M, et al. Bacillus co-inoculation alleviated salt stress in seedlings cucumber. Agronomy (Basel) 2021, 11, 966. [Google Scholar] [CrossRef]
- 97. Leite R da C, Pereira YC, Oliveira-Paiva CA de, Moraes AJG de, Silva GB da. Increase in yield, leaf nutrient, and profitability of soybean co-inoculated with Bacillus strains and Arbuscular mycorrhizal fungi. Rev Bras Cienc Solo, 2022; 46. [CrossRef]
- Brasil. Inoculantes; 2024. [cited 2024 April 27]. Available from: https://www.atermaisdigital.cnptia.embrapa.br/web/inoculantes.
- Conab. Boletim de safra de grãos - 7o Levantamento - Safra 2023/24 2024. Available from: URL: https://www.conab.gov.br/info-agro/safras/graos/boletim-da-safra-de-graos.
- Jones, F. Os primeiros inoculantes. revistapesquisa.fapesp.br. 2019. [cited 2023 September 24]. https://revistapesquisa.fapesp.br/os-primeiros-inoculantes/.
- Telles TS, Nogueira MA, Hungria M. Economic value of biological nitrogen fixation in soybean crops in Brazil. Environ Technol Innov 2023, 31, 103158. [Google Scholar] [CrossRef]
- Paulitsch F, dos Reis FB Jr, Hungria M. Twenty years of paradigm-breaking studies of taxonomy and symbiotic nitrogen fixation by beta-rhizobia, and indication of Brazil as a hotspot of Paraburkholderia diversity. Arch Microbiol 2021, 203, 4785–4803. [Google Scholar] [CrossRef]
- Pavinato PS, Cherubin MR, Soltangheisi A, Rocha GC, Chadwick DR, Jones DL. Revealing soil legacy phosphorus to promote sustainable agriculture in Brazil. Sci Rep 2020, 10, 15615. [Google Scholar] [CrossRef]
- Kalayu, G. Phosphate solubilizing microorganisms: Promising approach as biofertilizers. Int J Agron 2019, 2019, 1–7. [Google Scholar] [CrossRef]
- Sipert S, Cohim E, do Nascimento FRA. Identification and quantification of main anthropogenic stocks and flows of potassium in Brazil. Environ Sci Pollut Res Int 2020, 27, 32579–32593. [Google Scholar] [CrossRef]
- Schueler TA, Dourado ML, Videira SS, da Cunha CD, Rizzo ACL. Biosolubilization of verdete: An alternative potassium source for agriculture fertilizer. Biocatal Agric Biotechnol 2021, 34, 102031. [Google Scholar] [CrossRef]
- 107. Costa AD, Santos SR dos, Pereira GL, Santos WO. Glauconite as a potential source of potassium in Brazilian agriculture - a review. Cienc Agron, 2024; 55. [CrossRef]
- Brasil. Plano Nacional de Fertilizantes 2050; 2022. [cited 2024 April 27]. Available from: https://www.gov.br/planalto/pt-br/assuntos-estrategicos/documentos/planos/plano-nacional-fertilizantes/view.
- Zilli M, Scarabello M, Soterroni AC, Valin H, Mosnier A, Leclère D, et al. The impact of climate change on Brazil’s agriculture. Sci Total Environ 2020, 740, 139384. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Direct and indirect mechanisms of PGPR and PGPF for plant growth promotion in the soil.
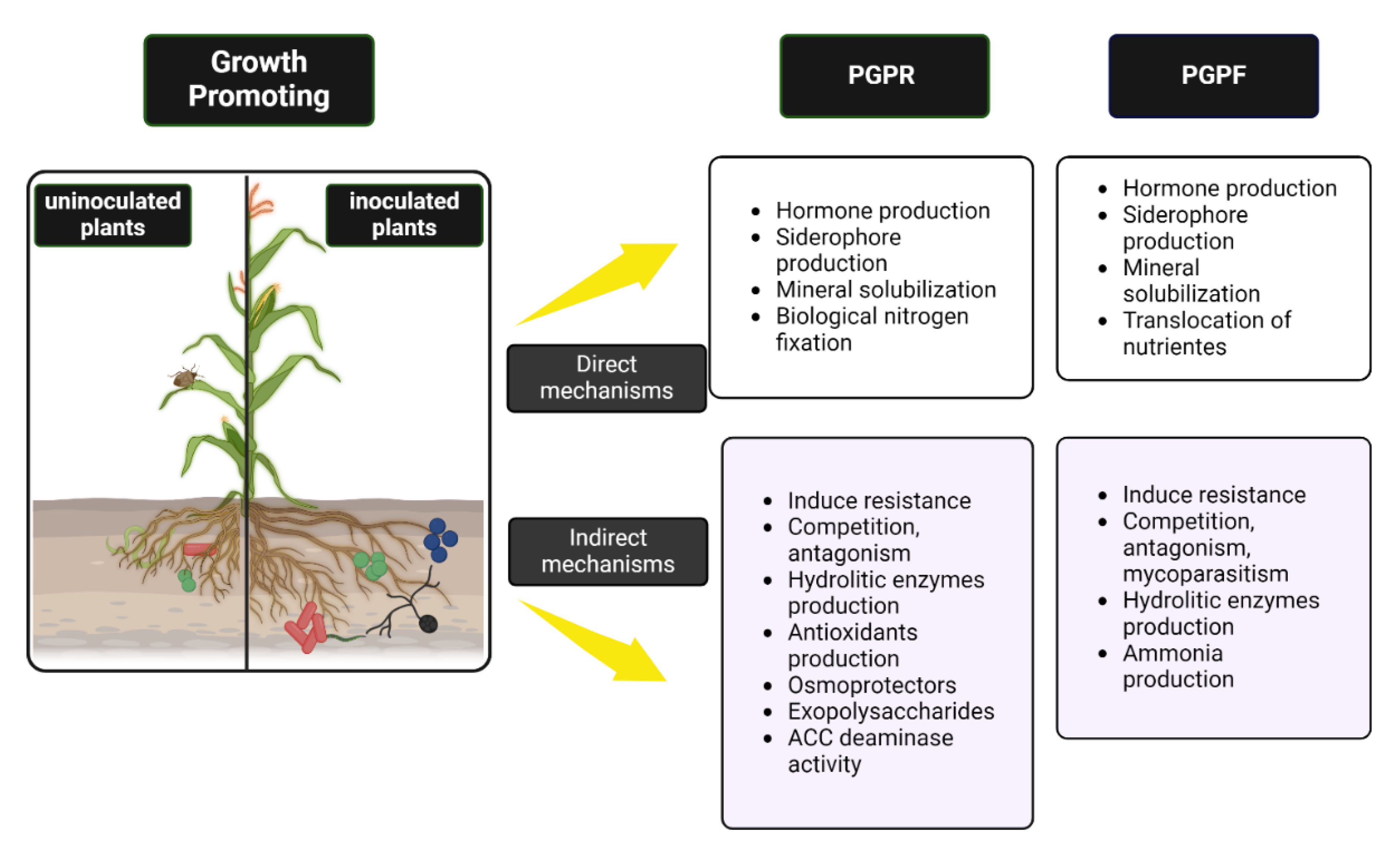
Figure 2.
– Number of inoculants registered for the main crops in Brazil.
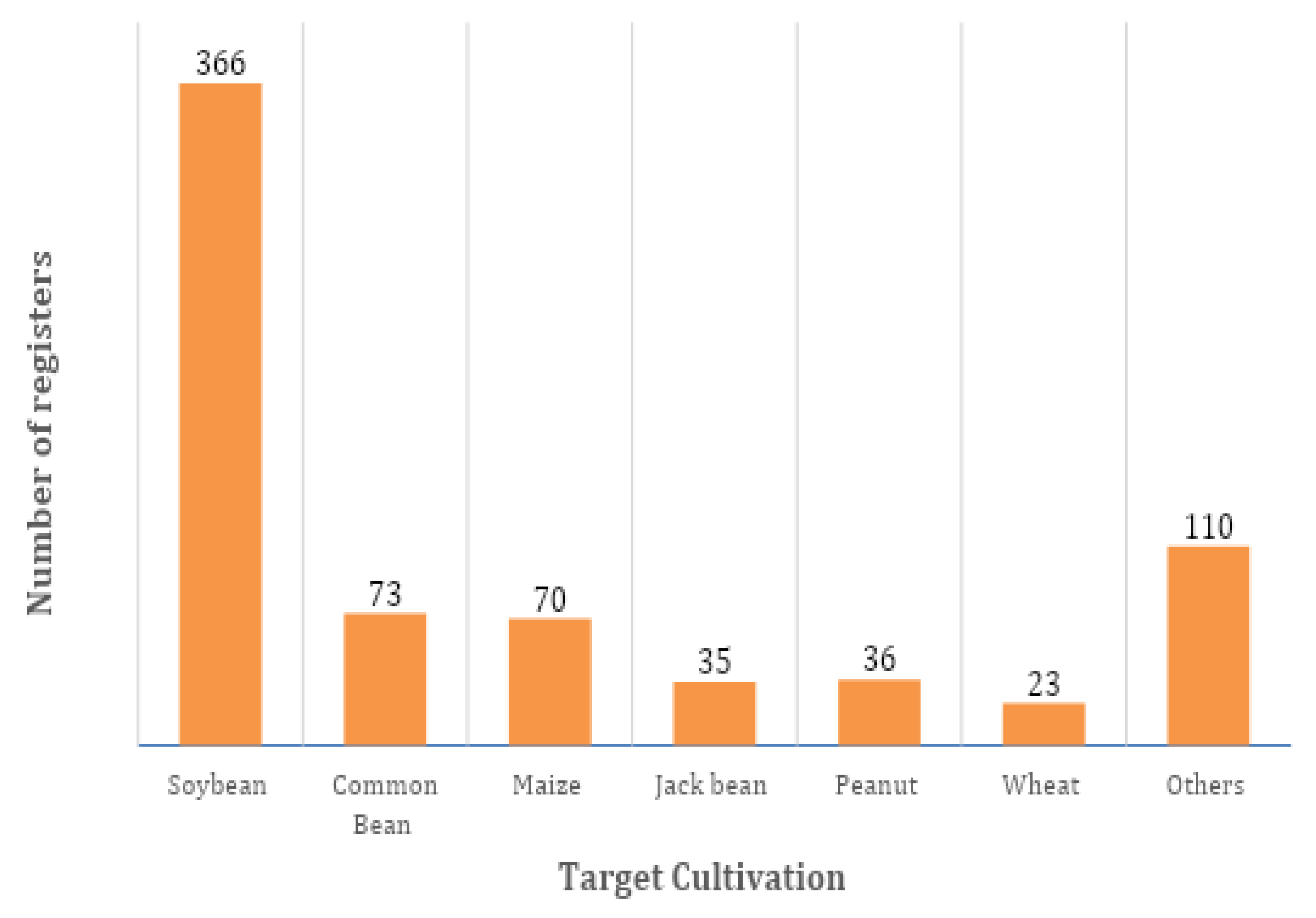

Table 1.
Some examples of PGPR growth promotion mechanisms.
| Specie | Mechanism of growth promotion | Culture | Reference |
|---|---|---|---|
| Azospirillum brasilense | Nitrogen fixation | Maize (Zea mays) | [5] |
| Bradyrhizobium sp. | Nitrogen fixation | Soybean (Glycine max) | Zilli et al., 2021[6] |
| Pseudomonas sp. | Phosphate solubilization | Pea (Pisum sativum) | Oteino et al. 2015[7] |
| Pseudomonas sp. | Phosphate solubilization | Maize (Zea mays) | Sarikhani et al., 2020[8] |
| Pseudomonas brassicae | Siderophore production | Mung bean (Vigna radiata) | Sun et al. 2022[9] |
| Pseudomonas aeruginosa | Phytohormone production | Mung bean (Vigna radiata) | Uzma et al., 2022[10] |
| Pseudomonas putida | Antioxidant activity | Maize (Zea mays L.) | Sandhya et al., 2010[11] |
| Bacillus velezensis | Phosphate solubilization and phytohormone production | Wheat (Triticum aestivum) | Afzal et al.2023[12] |
| Bacillus velezensis | Phosphate solubilization | Soybean (Glycine max) and Maize (Zea mays) | Mosela et al. 2022[13] |
| Bacillus mojavensis | VOCs production | Arabidopsis thaliana | Rath et al., 2018[14] |
| Bacillus subtilis | ACC deaminase activity | Tomato (Solanum lycopersicum) | Gowtham et al., 2020[15] |
| Serratia sp. | ACC deaminase activity and phytohormone production | Sunflower (Helianthus annuus) | Carlos et al., 2026[16] |
Table 2.
Some examples of PGPF growth promotion mechanisms.
| Specie | Mechanism of growth promotion | Culture | Reference |
|---|---|---|---|
| Trichoderma sp. | Phosphate solubilization | Soybean (Glycine max) | [47] |
| Trichoderma sp. | VOCs production | Arabidopsis thaliana | [48] |
| T. koningiopsis | VOCs production | Arabidopsis thaliana | [49] |
| T. viride | VOCs production | Tomato (Solanum lycopersicum) | [50] |
| T. asperellum | Phosphate solubilization and phytohormone production | Cucumber (Cucumber sativus) | [51] |
| Rhizophagus clarus | Increase of P and N content | Soybean (Glycine max) and Cotton (Gossypium hirsutum) | [52] |
| Glomus. intraradices | Salt tolerance | Wheat (Triticum aestivum) | [53] |
| Rhizophagus irregularis | Drought tolerance | Maize (Zea mays) | [54] |
| Mix of Rhizophagus clarus, R. intraradices, Septoglomus deserticola, Funneliformis mosseae | Water déficit tolerance | Soybean (Glycine max) | [55] |
| Rhizophagus irregularis , Funneliformis mosseae, and Funneliformis geosporum | High temperature tolerance | Soybean (Glycine max) | [56] |
Table 3.
Benefits of inoculating microorganisms.
| Species | Culture | Benefits | References |
| Bradyrhizobium diazoefficiens and Rhizobium tropici | Commom beans (Phaseolus vulgaris) | Growth promotion and grain yield | [92] |
| Bradyrhizobium japonicum and Azospirillum brasilense | Soybean (Glycine max) | Increased yield components, grain yield and seed quality | [93] |
| Pseudomonas fluorescens and Azospirillum brasilense | Tomato (Solanum lycopersicum) | Increased yield and fruit quality | [94] |
| Bradyrhizobium sp. and Trichoderma sp. | Cowpea (Vigna unguiculata) | Increased the growth rate, biomass and photosynthetic pigments | [95] |
| B. licheniformis and B. subtilis | Cucumber (Cucumber sativus) | Alleviated Salt Stress | [96] |
| B. subitilis, B. megaterium and Rhizophagus intraradices | Soybean (Glycine max) | Increase leaf nutrient and in yield | [97] |
| Rhizophagus irregulares and Bradyrhizobium sp. | Mung bean (Vigna radiata) | Growth promotion and alleviated water stress | [90] |
Table 4.
Genus of microorganisms registered for the main crops in Brazil, organized in single inoculation or co-inoculation.
Table 4.
Genus of microorganisms registered for the main crops in Brazil, organized in single inoculation or co-inoculation.
| Cultura | Gênero | Single inoculation | Co-inoculation |
|---|---|---|---|
| Soybean (Glycine max) | Bradyrhizobium sp. | 246 |
B. Japonicum + B. Elkani (52) B. Japonicum + A. brasilense (4) A. brasilense + P. fluorescense (2) B. megaterium + B. subtilis (2) B. subtilis + B. elkani (3) R. intraradices + Claroideoglomus claroideum (2) B. subtilis + B. elkani +Parabhurkodelia nodosa (3) B. subtilis + B. amyloliquefacens +B. pumilus (1) P. fluorescense + B. amyloliquefacens + Priestia megaterium (1) |
| Azospirillum sp. | 28 | ||
| Bacillus sp. | 8 | ||
| Pseudomonas sp. | 6 | ||
| Trichoderma sp. | 5 | ||
| Rhizophagus sp. (Rhizoglomus) | 5 | ||
| Commom beans (Phaseolus vulgaris) | Rhizobium sp. | 65 | Bacillus megaterium + Bacillus subtillis (1) |
| Azospirilum sp. | 6 | ||
| Bacillus sp. | 1 | ||
| Maize (Zea mays) | Azospirillum sp. | 41 |
B. megaterium + B. subtilis (2) A. brasilense + P. fluorescense (2) B. megaterium + Lysinobacillus sp. (1) B. licheniformis + B. aryabhatai (2) B. Japonicum + A. brasilense (2) R. intraradices + Claroideoglomus claroideum (2) B. subtilis + B. amyloliquefacens +B. pumilus (1) |
| Bacillus sp. | 8 | ||
| Rhizophagus sp. (Rhizoglomus) | 5 | ||
| Pseudomonas sp. | 3 | ||
| Methylobacterium sp. | 1 | ||
| Peanut (Arachis hypogaea) | Bradyrhizobium sp. | 36 | - |
| Jack bean (Canavalia ensiformis) | Bradyrhizobium sp. | 35 | - |
| Wheat (Triticum aestivum) | Azospirilum sp. | 23 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



